
Relationship of Chromosome Changes to Neoplastic Cell
30
Relationship of Chromosome Changes to Neoplastic Cell Transformation Joseph A. DiPaolo, PhD, and Nicolae C. Popescu, PhD Chromosomal abnormalities are a frequent concomitant of neoplasia, and although it is tempting to relate these mutations and alterations in chromatin (DNA) function to cancer, their relationship to the initiation or progression of carcinogenesis is unknown. Mammalian cells in culture, after interacting with chemical carcinogens, often exhibit chromosome damage consisting of breaks and exchanges of chromatid material. The pattern of damage of banded metaphases indicates that negative bands are especially vulnerable to the action of chemical carcinogens, probably because of differential chromatin condensation. Damage to individual chromosomes may be random or non- random, depending on the species. Cell death can be correlated with chromatid altera- tions that occur shortly after treatment with chemical carcinogens. There is also a correlation between mutagenic and carcinogenic activity of some chemical carcinogens and the frequency of sister chromatid exchanges. The question of whether specific chromosome changes are absolutely required for neoplastic transformation cannot be answered because of conflicting data and diverse results from studies even with known carcinogens. Cell transformation may occur without any visible chromosome changes. A universal specific numerical or visible structural chromosomal alteration is not necessarily associated with chemical or viral transformation. Chromosome changes are independent of the etiologic agents: different carcinogens may produce transforma- tion associated with the same abnormal chromosomes, but not all transformed lines insvariably exhibit the same abnormality, even with the same chemical. In some species, chromosome having nucleolar organizer regions may be more frequently involved in numerical or structural deviations. Progressively growing tumors also mav occur as a result of the proliferation of transformed cells without detectable chromosome changes, indicating that tumorigenicity need not be related to an imbalance of chromosome number or structure. Our studies indicate that chromosome changes are not essential for establishment of neoplasms but that karyotypic instability may result in response to selective growth pressures. (Am J Pathol 85:709-738, 1976) THE STU DY OF THE TRANSFORMIATION in vitro of normal cells into neoplastic cells -hich results from exposure to a variety of chemical or xiral carcinogens presents an opportunity for identifying environmental agents that max be responsible for causing cancer in humans and for studying their mechanism of action. A number of experimental models for studying transformation, as verified by subsequent tumor formation, exist. Many- studies followved the original observations of Berwald and Sachs," From the Somatic Cell Genetics Section. Biology Branch. Carcinogenesis Program. Div ision of Cancer Cause and Prevention. National Cancer Institute. Bethesda. Mlarvland Presented at the Sixtieth Annual Mleeting of the Federation of Xmerican Societies of Experimental Biolog-. Anaheim. Calif. April 13. 1976 Xddress reprint requests to Dr J X DiPaolo. Building 37. Room 2A13. National Cancer Institute. Bethesda. MID 20014 709
Transcript of Relationship of Chromosome Changes to Neoplastic Cell

Relationship of Chromosome Changes to NeoplasticCell Transformation
Joseph A. DiPaolo, PhD, and Nicolae C. Popescu, PhD
Chromosomal abnormalities are a frequent concomitant of neoplasia, and although it istempting to relate these mutations and alterations in chromatin (DNA) function tocancer, their relationship to the initiation or progression of carcinogenesis is unknown.Mammalian cells in culture, after interacting with chemical carcinogens, often exhibitchromosome damage consisting of breaks and exchanges of chromatid material. Thepattern of damage of banded metaphases indicates that negative bands are especiallyvulnerable to the action of chemical carcinogens, probably because of differentialchromatin condensation. Damage to individual chromosomes may be random or non-random, depending on the species. Cell death can be correlated with chromatid altera-tions that occur shortly after treatment with chemical carcinogens. There is also acorrelation between mutagenic and carcinogenic activity of some chemical carcinogensand the frequency of sister chromatid exchanges. The question of whether specificchromosome changes are absolutely required for neoplastic transformation cannot beanswered because of conflicting data and diverse results from studies even with knowncarcinogens. Cell transformation may occur without any visible chromosome changes.A universal specific numerical or visible structural chromosomal alteration is notnecessarily associated with chemical or viral transformation. Chromosome changes areindependent of the etiologic agents: different carcinogens may produce transforma-tion associated with the same abnormal chromosomes, but not all transformed linesinsvariably exhibit the same abnormality, even with the same chemical. In some species,chromosome having nucleolar organizer regions may be more frequently involved innumerical or structural deviations. Progressively growing tumors also mav occur as aresult of the proliferation of transformed cells without detectable chromosome changes,indicating that tumorigenicity need not be related to an imbalance of chromosomenumber or structure. Our studies indicate that chromosome changes are not essentialfor establishment of neoplasms but that karyotypic instability may result in response toselective growth pressures. (Am J Pathol 85:709-738, 1976)
THE STUDY OF THE TRANSFORMIATION in vitro of normal cells intoneoplastic cells -hich results from exposure to a variety of chemical orxiral carcinogens presents an opportunity for identifying environmentalagents that max be responsible for causing cancer in humans and forstudying their mechanism of action. A number of experimental models forstudying transformation, as verified by subsequent tumor formation, exist.Many- studies followved the original observations of Berwald and Sachs,"
From the Somatic Cell Genetics Section. Biology Branch. Carcinogenesis Program. Divisionof Cancer Cause and Prevention. National Cancer Institute. Bethesda. Mlarvland
Presented at the Sixtieth Annual Mleeting of the Federation of Xmerican Societies of ExperimentalBiolog-. Anaheim. Calif. April 13. 1976
Xddress reprint requests to Dr J X DiPaolo. Building 37. Room 2A13. National Cancer Institute.Bethesda. MID 20014
709

710 DIPAOLO AND POPESCU American Journalof Pathology
who demonstrated that it was possible to transform Syrian hamster cellstrains. The subject of carcinogenesis and, in particular, in vitro carcin-ogenesis has been reviewed recently.2-4 Our own studies have utilizedseveral mammalian cell systems for transformation: newly establishedcultures derived from Syrian hamster,5'6 guinea pigs,7 or rat embryo;8 andestablished cell lines derived from the mouse BALB/3T3.9 Human fibro-blasts and human peripheral lymphocytes or cord blood have also beenused in conjunction with known carcinogens.
In some systems, quantitative analyses of the frequency of transforma-tion has been possible, indicating a linear relationship with dose.10" Thetransformation of Syrian hamster cells by a chemical carcinogen demon-strates a Poisson distribution and thus is consistent with a one-hit hypoth-esis.ll. 2The lack of transformation in the controls is further evidence thatthis transformation system results from an induction of the transformedstate rather than from selection of preexisting transformed cells. Theposition that transformation by chemical carcinogens is or may be a directprocess and need not involve viral activation is based upon experimentswhich demonstrate that the transformed cells are negative for virusesknown to transform hamster cells, fail to demonstrate activation of extra-cellular-type particles after BudR induction, lack reverse transcriptase anddetectable Syrian hamster complement-fixing antigen, and fail to expresshamster-type viruses following hybridization techniques using cRNAfrom hamster-type C virus."3 The critical site of action of a chemicalcarcinogen is unknown. Most importantly, whether the phenomenon oftransformation is due to a somatic genetic change or to an epigeneticalteration is unknown. Cell culture systems will be useful in this regardbecause they permit the study of the nature of the target-cell insult as wellas the modifying or modulating factors that influence transformation.Studies involving transformation with a specific etiologic agent such as achemical carcinogen should provide evidence of the importance of therole of chromosomes in terms of changes in structure or in number.
Malignant neoplasms are often characterized by gross cytogenetic alter-ations. Numerical and structural deviations have been reported sinceMakino 14 first observed a marker chromosome and have been an impor-tant criterion for distinguishing neoplastic cells from normal and quiteoften for distinguishing one neoplasm from another even though theymay be caused by the same agent. It is known that normal diploidkaryotypes may be present in malignant neoplasms and that chromosomeimbalance need not lead to malignancy.15l'6 On the other hand, chromo-somal instability associated with certain inherited syndromes or inducedby mutagenic agents may predispose normal somatic cells to malig-

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 711December 1976
nancv."7 Chromosome breakage is also associated with the addition oftoxic materials to cells and may result in cell death.18
In other cancers, the alterations may occur during progression of thedisease. Nonrandom chromosomal changes have been observed especiallyin human neoplasms even prior to the development of the new chromo-somal banding techniques.19 Banding analysis shows that in chromosomesfrom cases of Burkitt's lymphoma most possess an extra band on onechromosome " and not all meningiomas lack a complete or part of anotherchromosome.21 Many cancers lack marker chromosomes. At least 50% ofpatients with acute leukemia did not have recognizable visible karyotypicchanges.2324 Thus, the cytogenetic changes may be secondary to theneoplastic state.
If the chromosomal changes reflect progression of the disease, theoriginal neoplasm may be due to an epigenetic influence or a mutation atthe gene level. Somatic cell hybridization is providing evidence that aspecific genetic or epigenetic function is the pivotal event for cancer,independent of the etiologic agent. Although a useful approach, puzzlingresults have appeared (reviewed Barski and Belehradek 25). Recently, theT antigen, the tumor-specific transplantation antigen, and the simianvirus 40 genome (SV40) have been assigned to human chromosome C-7.26Other data indicate that human cells transformed by SV40 fail to formprogressively growing tumors in nude mice and that the T antigen maybe assigned to a different chromosome.28
In general, data from the study of human neoplasia and from experi-ments, particularlv with chemical carcinogens," indicate that cytogeneticchanges in neoplastic states may be partially due to the genetic variabilityof the individuals involved or the experimental material used. Specifictypes of transformation generally do not have characteristic chromosomeabnormalities, but individual ones mav have chromosome alterationsindicating clonal patterns of growth.
Since alterations at the chromosomal level are an important aspect ofcell transformation by chemical carcinogens or viruses, our first objectivew%as to determine the chromosomal constitutions of the cells used. Gener-allv, the chromosomes were obtained from cell cultures in the logarithmicgrowth phase. In studies with human lymphocytes, the cells were culturedfor 72 hours in the presence of phytohemaglutinin, with Colcemid addedfor the final 3 to 4 hours.30 At a concentration of 0.04 to 0.08 jig Colce-mid/ml of medium, chromosome contraction of cells in suspension ormonolaver was optimal. However, no Colcemid treatment was requiredfor some transformed guinea pig cultures since these chromosomes tendedto be more contracted than those from other species. At the end of the

712 DIPAOLO AND POPESCU American Journalof Pathology
Colcemid treatment, the cells were detached from the petri dish with arubber policeman. In one experiment with hamster cells transformed bysimian adeno-7 (SA7) virus, a short treatment with trypsin was necessaryto disperse the cell foci typical of these cultures. Subsequently, the cellsuspension was concentrated by centrifuging 4 to 5 minutes at 80Q to 1000rpm in a table top centrifuge. The cells were then suspended in a hypo-tonic solution. For monolayer cultures, sodium citrate (0.75%) and potas-sium chloride KCI (0.5%) were found to be unsuitable, as each affectedthe chromosome morphology or, in some cases, disintegrated the meta-phases. Therefore, a solution of distilled water and culture medium (1: 3)was used. Cells were incubated for 15 to 30 minutes at room temperature.Human leukocyte cultures were incubated in the KCI solution for 10 to 12minutes at 37 C. All cells were fixed with Carnoy's fixative three times,with a 24-hour interval time between the second and the third fixation.After the third fixation, the chromosomes were prepared by pipettingthree or four drops of fixative containing the cells onto precleaned, dry orwet slides followed by manual drying.
Caspersson's discovery of differential affinity of metaphase chromo-somes for fluorescent dye 31 makes it possible to distinguish the chromo-somes longitudinally. Each chromosome is divisible into many bright anddark regions or Q bands. The banding pattern is unique for each individ-ual pair of chromosomes of the species. Subsequently, it was found thatvarious types of treatments 3-"- prior to staining with Giemsa also pro-duced chromosome cross-bands which corresponded to those revealedafter fluorescent dyes.-",-" The fluorescent technique, however, cannot betotally replaced by Giemsa banding method since certain rearrangementsin human karvotypes, as well as sequential examination of the samekarvotype, may require a fluorescent staining method. For example, thesilver technique for visualization of regions associated with the nucleolarorganizer will not be successful if applied to G-band preparations.
For G banding, several protocols were employed in which the pre-treatment prior to Giemsa staining differed. These consisted of trypsin 33or ureae treatment or a combination of the two as well as heat treatmentin alkali.32 Our standard procedure is a modification of one used byGallimore and Richardson for rat chromosomes.37 In addition to theprocedures for cross-bands, it is imperative to examine the chromosomesby procedures that identify constitutive heterochromatin, C bands, sincecertain species such as Syrian hamsters have large blocks of this material.In addition, this staining procedure often can be helpful for resolving theorigin of some rearrangements associated with neoplastic cells. C-bandtechnique also permits determination of the extent to which the relative

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 713December 1976
amount of constitutive heterochromatin in neoplastic cells is increased, ordecreased, and if a cytologically aneuploid cell is still geneticallv euploid.The original NaOH technique for C bands described by Arrighi and
Hsu ` was used to analyze the constitutive heterochromatin in mouse,"Svrian hamster," rat, and human chromosomes.4" More recentlv, partic-ularlv for species whose chromosomes contain heterochromatin associatedprimarilv with the centromeric region, a BaOH procedure was used.4
Recently, the new methods for demonstrating the nucleolar organizeror chromatid exchanges of chromosomes were used. The silver stainingtechnique advocated by Howell and Benton" and their associates forvisualization of the nucleolar organizers of human chromosomes wasfurther developed by Goodpasture and Bloom" and was successfullvapplied to different types of mammalian cells. Now sister chromatidexchanges (SCE) can be distinguished without the use of radioactivecompounds and autoradiographv; cells are first exposed to 5-bromode-oxvuridin (BrdU) for two rounds of replication. At the second mitosis, thechromosomes exhibit a specific staining pattern in which only onechromatid is differentially stained with fluorochrome (Hoechst 33258), asfirst described by Latt " or with Giemsa stain after exposure of the slidesto the Hoechst fluorochrome and light for BudR activation, as describedbv Perrv and Wolff." A simplified method described by Korenberg andFreedlender,47 used in our experiments, requires incubation in a solutionof sodium phosphate prior to Giemsa staining.
Norma G-Band Kyoty
The human chromosomes were arranged according to the nomencla-ture established at the Paris conference." For rat and mouse chromo-somes, the standard karyotvpe established by international com-mittees 49' was used. G-banded Syrian hamster chromosomes werearranged 51 following the original conventional karvotype of stainedchromosomes introduced by Lehman et al.52 Most pairs were arrangedaccording to their size; however, arm ratios were also considered (Figure1). For example, the pair A4 may be larger than pair A3, but its pro-nounced submetacentric morphology was decisive in placing it before thelargest metacentric pair B5. Of the acrocentric chromosome group, D, thefirst acrocentric pair D16, although not the largest pair, was placed firstbecause it is the only acrocentric pair without short arms.The guinea pig G-band karyotype consisting of 64 chromosomes was
arranged according to the banding pattern and chromosome size, regard-less of arm ratio. The autosome pairs were not divided into groups. By theG-banding method, however, the largest and medium size chromosomes

714 DIPAOLO AND POPESCU American Journalof Pathology
could be accurately identified; the smaller ones (pairs 17-31), wheresometimes only one or two bands could be distinguished," are at timesdifficult to identify.The chromosomal constitution of the established cell lines used in our
experiments was also examined bv G-band karvotypes. A cloned deriva-tive of the BALB/3T3 mouse line had an aneuploid constitution with asubtetraploid stemline and three or four abnormal chromosomes.9 39 Clone11 derived from the V79 Chinese hamster line had a pseudodiploidkarvotvpe with a narrow distribution around the normal modal number.
Normal C-Band Pattern
The Svrian hamster is of particular interest due to its large amount ofconstitutive heterochromatin located on the sex chromosomes and on theshort arm of most autosomes. In an early study of the time course of DNAreplication in Syrian hamster chromosomes, Galton and Holt " found thathalf of the X chromosomes, the whole Y, and numerous short-arm seg-ments were late replicating and hypothesized that these regions are ofheterochromatic nature. This hvpothesis has been confirmed with C-bandstaining by Hsu and Arrighi 55 and in this laboratorv;" a half of the X, theentire Y, the short arms of several submetacentric pairs as well as the longarm of chromosomes E20 are heterochromatic. The short arm of theautosomes, as well as the long arm of the chromosome E20, did not alwaysstain darkly after C-band procedure. This was most evident on slidesstained more than 1 month after preparation. On such slides, only acentromeric darklv stained area occurred. This observation suggests thatthe short arm of the autosomes in Svrian hamster cells contains a specialclass of heterochromatin. Further details about the nature of hetero-chromatin in Syrian hamster cells will be presented elsewhere.
Guinea pig, mouse, rat, and human C-band patterns were also ana-lvzed. All have a relativelv small amount of constitutive heterochromatinmainly associated with the centromere. Guinea pig sex chromosomescould be recognized bv their increased amount of heterochromatin ascompared to the autosome.5
Nucleolar Organizer Regions On Syrian Hamster Chromosomn
For several mammalian species, in situ DNA/RNA hybridization pro-cedure has localized the ribosomal genes.-,57 With the new ammoniacalsilver stain (Ag-AS) Goodpasture and Bloom demonstrated that silver-stained nucleolar organizer regions (NOR) coincide to those visualizedbv in situ hybridization." NOR were identified on Syrian hamsterchromosomes. Svrian hamster complement contains 6 chromosomes bear-

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 715December 1976
ing NOR. Three are on the short arms of acrocentrics from the pairs D17-D19. One member of the pair E20 and a pair of medium size sub-metacentric chromosomes from group B have NOR located at the telo-mere (Figure 2).
Inital Ciromosom Change After Chenical InsultMost, if not all, carcinogens are known to produce chromosome dam-
age.w However, the clastogenic effect of chemical, physical, and biologicagents is not limited to those capable of inducing malignant transforma-tion. What is the relevance of insult to chromosomes? Is chromosomedamage a determinant of transformation and malignancy, or is it primar-ilv a reflection of toxicity? The majority of damaged cells are inviable. Wehave attempted to determine the effects of chemical carcinogens on thechromosomes of different species and to locate by banding resolution theposition at which individual breaks or exchanges occur. Chromosomebanding has been analyzed after x-ray-induced chromosomal damage,594'as well as with some chemical agents,62 but no study has involved theimmediate affects of chemical carcinogens.
Cultured Syrian hamster fetal cells w'" maintain their normal constitu-tion in terms of chromosome abnormalities or deviations from the normalmodal number for up to 20 subeultures. Chromatid aberrations such aschromatid gaps, or chromosome aberrations such as dicentrics., occur in1% or less of the metaphases. Occasionally, metaphases with one or twochromosomes missing were found, but this may be an artifact of chromo-some preparation. Secondarv cultures contain 1 to 2% tetraploid meta-phases.
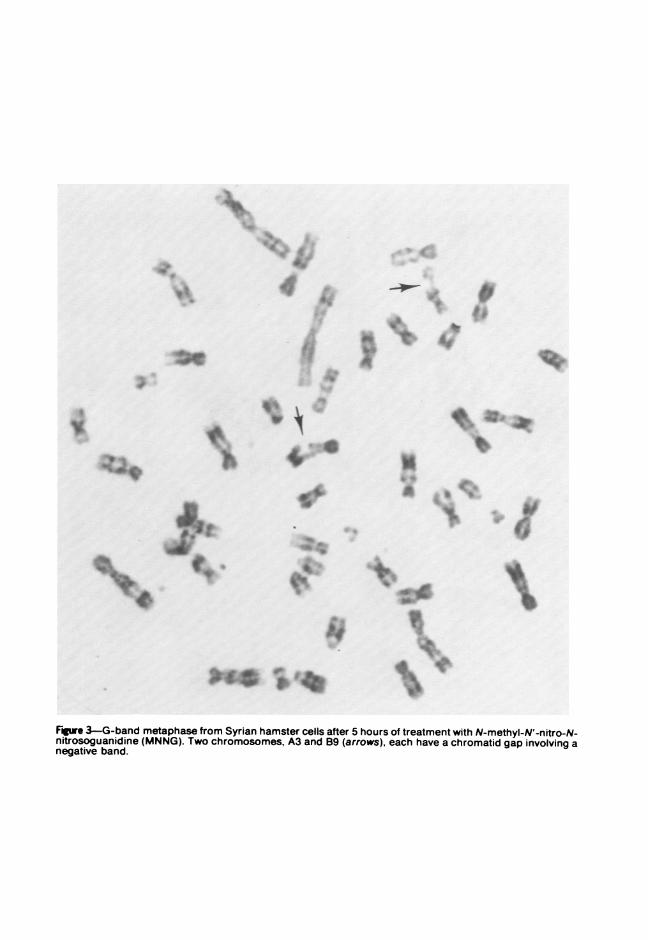
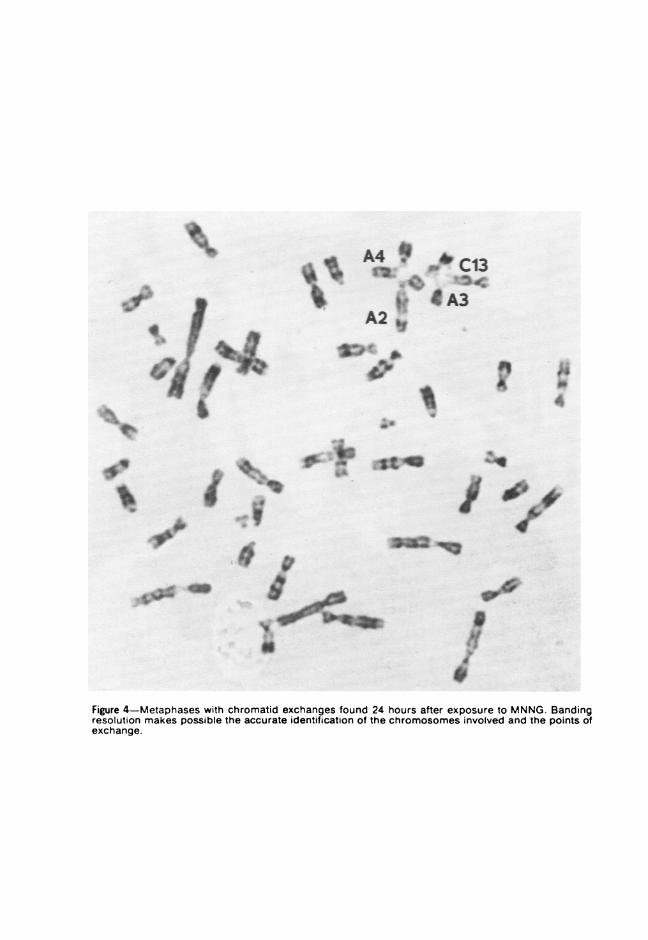
In a series of experiments, secondary Syrian hamster cells were treatedwith 0.5 ,g N-methvl-N'-nitro-N-nitrosoguanidine (MNNG)/ml mediumand chromosomes were prepared at 5, 13, or 24 hours thereafter. Thelength of exposure to MNNG did not appear to affect the frequency ofaberrations, which ranged from 20 to 25%. Whereas x-ray-induced dam-age to Syrian hamster chromosomes consisted of both chromatid andchromosome aberrations,' MNNG, an alkylating agent, produced mainlychromatid aberrations. G-band analysis of MNNG-treated cells revealedthat the breaks and gaps were randomly distributed among the chromo-somes of the Syrian hamster complement. In terms of banding locationswithin chromosomes, however, MNNG-induced breakage appeared toinvolve primarily the negative bands (Figure 3). C-band analysis alsoallowed the recognition of the chromosomes involved in different types ofrearrangements (Figure 4) and the identification of displaced portions ofanv chromosome. The longer autosomal chromosomes were most often

716 DIPAOLO AND POPESCU American Journalof Pathology
involved in different tvpes of exchanges. The sex chromosomes, X and Y,as well as the E20 chromosome, which is totally heterochromatic on thelong arm, did not participate in chromatid exchanges.The addition of chemical carcinogens to human leukocyte cultures
obtained from peripheral blood or fetal cells freshly derived from thera-peutic abortions has not resulted in transformation but sometimes pro-duced C-band type figures. The formation of G bands after exposure ofcells in vitro to different chemical compounds was first reported by Hsu etal." after treatment with actinomycin D or other compounds on G2 phasesof the cell cvcle. It has since been found that treatment with hydroxyureain the S phase of the cell cycle also induced G bands in living cells.41Carcinogenic polycyclic hydrocarbons, 4-nitroquinoline-1-oxide (4NQO),aflatoxin B,(AFT), and N-acetoxy-2-fluorenylacetamide (N-AcFAA), butnot urethane, caused toxicity as indicated by chromosomal damage. Aftera 24-hour treatment of human leukocyte cultures or Svrian hamsterembryo secondary cultures with chemical agents at concentrations knownto produce transformation of hamster cells in vitro, the chromosomesexhibited G bands without any further preparation; noncarcinogenicchemicals were ineffective in producing these bands."8
Although subtle variations may have occurred with different agents, thebands produced by each carcinogen appear to be similar to those observedfollowing orthodox G-band procedures on fixed cell preparations. Theability of these known carcinogens to produce bands is a transitory phe-nomenon. When the carcinogen was removed and the fetal cells culturedfor an additional 24 hours; the chromosomes were no longer banded.These events are considered reflections of a nonspecific tvpe of chemicaltoxicity because they may also be caused by other types of chemicals suchas hvdroxurea and azure B, which are not suspected to be in vivo carcino-gens, and because the rearrangements that occurred following carcinogentreatment did not lead to permanently altered cells.With human and hamster material, we conclude that toxic chemicals
such as a chemical carcinogen lead to nonspecific chromatid lesions,whereas the treatment of fetal rat cells with 7,12-dimethyl-benz[a]anthracene (DMBA) may induce a highly specific lesion on onechromosome. Either cold or 3H-DMBA was added to exponentially grow-ing secondarv rat fetal cells for 5, 9, or 24 hours, including a 4-hourexposure to Colcemid. Chromosome preparations from treated cells, whencompared to the other chromosomes of the rat complement, had a higherfrequency of gaps or breaks associated with the chromosome A2. With C-band analysis, four specific regions of the A2 chromosome exhibitedlesions. In the rat chromosomes, C-band nomenclature proposed by Le-

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 717December 1976
van,67 the segments associated with the breaks or gaps occur on negativebands and are located exactly in the Q2-2, Q24, Q2-6, and Q3-4 bands.Band Q24 of chromosome A2 (Figure 5) is the most vulnerable. Treat-ment with 3H-thymidine also induced damage on band Q24 in a smallpercentage of the cells. In an experiment in which radioactive carcinogenwas used, 1 to 2% of the metaphases showed nonrandom grain accumula-tion on chromosome A2 between bands Q2-2 and Q34 after a 5- or 9-hourexposure. After removal of emulsion, treatment with DNase and reproc-essing for autoradiography, no label occurred, indicating that the vis-ible grains represented binding of the radioactive material to the chromo-somal DNA. The accumulation of grains on the chromosome A2 was notnecessarily associated with a chromosome lesion. In addition, with theBrdU technique for SCE, some SCE were located in what must beconsidered the most vulnerable segment of chromosome A2. These studieson rat chromosomes demonstrate that a specific chromosome appears tobe involved with several nonspecific reactions: a high frequency of breakswith DMBA, a certain degree of susceptibility to 3H-thymidine, and asusceptibility to 3H-DMBA as well as to other chemicals. Of interest is therecent finding of Sugiyama with the food additive, 2-(2-furyl-3-(5-nitro-2-furyvlacrylamide in vivo which also caused breaks on A2 rat chromo-somes.w
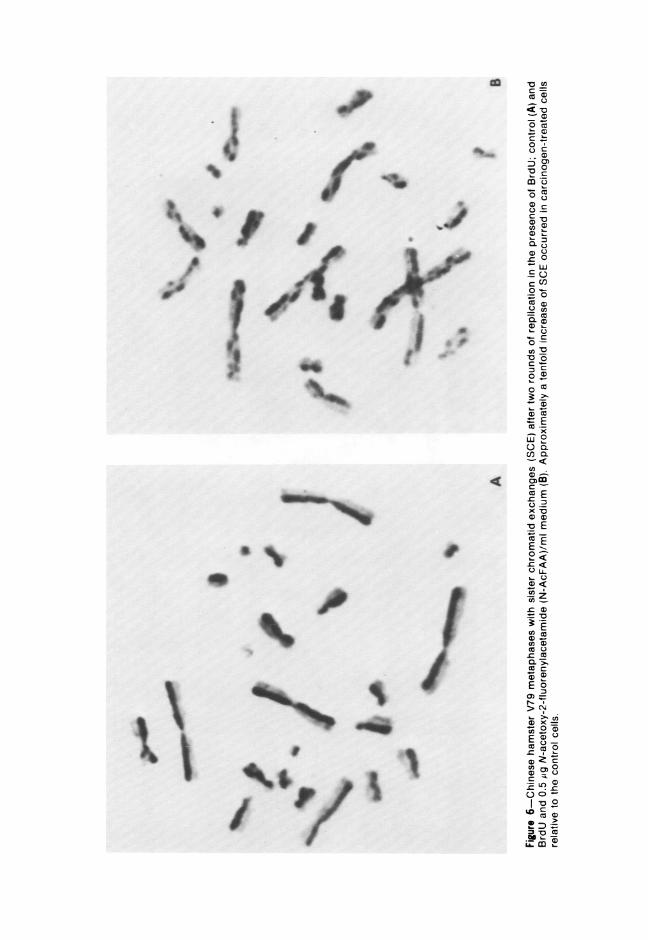
Studies are underway to determine the effects of various chemicals onthe frequency of SCE. This approach is particularly interesting since SCEfrequencies are far more sensitive indicies of chromosome damage thanthe frequencies of chromosome aberrations,' and some SCE may beassociated with mutational changes. Preliminary data indicates that chem-icals which are highly mutagenic and cause a high frequency of celltransformation per se may also induce a high frequency of SCE. Forexample, the number of SCE in V79 Chinese hamster cells after 0.5 ;g N-AcFAA/ml medium exhibit a tenfold increase in SCE relative to controls(Figure 6). Agents which are carcinogens in vivo but require metabolicactivation in vitro seldom induce SCE without such activation. Chemicalswhich show no mutagenic or carcinogenic activity in vitro or in vivo donot cause SCE.
Banding Analysis of Nepastic CellsFrom a total of nine different transformed cell lines obtained after
treatment with diverse chemical carcinogens, the number of markersranged from zero in four lines to as many as three in the remaining lines.70Each colonv transformed by a chemical carcinogen was derived from aculture that originated from a different hamster embryo culture. Transfor-

718 DIPAOLO AND POPESCU American Journalof Pathology
mation associated with the same specific marker was produced by differ-ent carcinogens, but not all the transformed lines had the same marker,even with the same chemical carcinogen. Some markers occurred intumor-derived cultures but not the transformed lines; others appeared inlater passages of both the transformed line and the tumor-derived culturesbut not in the early passage of these lines. The markers, M1-M7, found inthe various cell lines are shown in Figure 7. These markers demonstratethat marker formation may include different chromosomes of differentsizes belonging to different groups in the karyotype. The additionalmarkers Ma and Mb, after cell progression in vivo, demonstrate that newabnormal chromosomes may occur as a response to host pressure. C-bandanalvsis indicated no alteration in chromosome constitutive hetero-chromatin in chemically transformed lines, but in the tumor-derivedcultures, heterochromatic alteration was found in two different tumorcultures. In one of these, the heterochromatic alteration consisted of theaddition of a heterochromatic piece which was associated with the reac-tivation of an ordinarily inert short arm of chromosome number A4.40Nontranslocational numenrcal changes found in greater than 25% of thenear diploid line cells from G-band karyotype were summarized (Figure8). The changes in the number of chromosomes associated with eachidentifiable pair show that the increase and decrease in certain chromo-somes is random except for three lines in which trisomy for D17 waspresent as a result of transformation with BP, AFT, or 4NQO. Also, 4NQOmax cause transformation without involvement of D17 chromosomes.
These results are in contrast to those obtained by Sachs and associates 71and Benedict and associates,72 who reported that a complete or partialtrisomv or translocation of a piece of one chromosome was responsible forthe expression of malignancy of cells that had been exposed in vitro todimethvlnitrosamine or 1-al-D-arabinofuranosylcytosine. Whereas Sachsand associates propose that the expression and suppression of malignancyis controlled by the appearance or disappearance of a specific chromo-some, Benedict's group considers decreased malignancy to result frombalance between copies of chromosomes carrying genes for expression andsuppression of malignancy. In the final analysis, a discrepancy existsbetween these two groups on the chromosomal control of suppression ofmalignancv. These results are not substantiated by our analysis since theso-called expressor-bearing chromosome did not appear as part of a newchromosome nor was it present in the analysis of numerical changes ofspecific chromosome pairs. Interestingly, the chromosome that is in-criminated as the carrier of suppressor genes was frequently found inexcess.

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 719December 1976
A number of transformations with near-diploid modes derived fromSA7-transformed foci and foci obtained by treatment with a chemicalcarcinogen and SA773 were analyzed for structural and numerical chromo-some deviations.74 The markers found following transformation by viruswere different from those described for chemical transformation (Figure9). Markers occurred as a result of deletions, translocations, and centro-meric fusions resulting in new chromosomes configurations; all of thesediffered from those obtained with chemical carcinogens. Among the sixvirus-transformed lines analyzed from four different cell pools, one wasdiploid in terms of G-band analysis, three had no marker chromosomes,and the remaining two each had unique markers. Analysis of numericaldeviations, increases, or decreases of chromosomes of the various hamsterlines transformed by SA7 alone or in combination with chemical carcino-gen failed to reveal a common or specific deviation of a specific chromo-some pair of the hamster karyotype. In some cases in which the variousfoci were derived from the same hamster pool, there was the same loss orgain of a chromosome in some lines derived from some of the foci. Thefoci indicated by the prefix 73 were all derived from the same pool. In thecase of A and B onlv, SA7 was used, whereas with 73C through E, the cellswere first treated with either 3-methylcholanthrene (3-MCA) or DMBAand then treated with SA7. Characteristic of the 73 pool was the loss ofone entire X chromosome or of a piece of one X chromosome (Figure 10).A similar observation was reported by Sachs and associates 75 in hamstercells transformed by polyoma; however, chromosomes identical to thosereferred to as responsible for expression of malignancy by polyoma werenot observed in cells transformed by SA7.
Copple and McDougall have characterized by G-band analysis a num-ber of clones derived from a Syrian hamster cell line (333-8-9) putativelytransformed bv herpes virus Type 2.7 The markers reported are morpho-logically indistinguishable from those markers described as distinctive forspecific, independently derived HSV-transformed cell lines.77 Some mark-ers Copple and McDougall pointed out are similar to those present herefor Svrian hamster cells transformed in vitro by chemical carcinogens orhamster tumors induced in vivo by 3-MCA.Th Consequently, they con-cluded that the presence of marker chromosomes is neither virus strainnor serotvpe specific nor even virus specific.
In some studies with hamster cells and in many other reports withcarcinogenic chemicals or viruses,71 there is an involvement of thechromosomes belonging to the acrocentric group or the smallest meta-centric pair, the same chromosomes that we have determined to beassociated with NOR. These chromosomes have an affinity for one another

720 DIPAOLO AND POPESCU American Journalof Pathology
(Figure 2B), and a carcinogenic insult may make them prone to in-volvement in nondisjunction. The importance of the numerical change inthis group of chromosomes remains to be investigated.
Viral transformation differs from the transformation by chemical car-cinogens in that a specific antigen(s) is associated with each virus thatcauses transformation. The in situ hybridization method with cRNA onfixed cells or chromosomes after treatment with SA7 virus has shown thatin some SA7-transformed cell lines the SA7 genetic material is integratedand randomly distributed over the nucleus and chromosomes. Adeno-virus-infected cells and adenovirus-transformed metaphases also did nothave anv chromosome or chromosome segment with specific grain local-ization;82- however, 3T6 mouse cells acutely infected with murine sar-coma leukemia virus were reported to have grain concentration atchromosome centromere regions of several chromosomes."
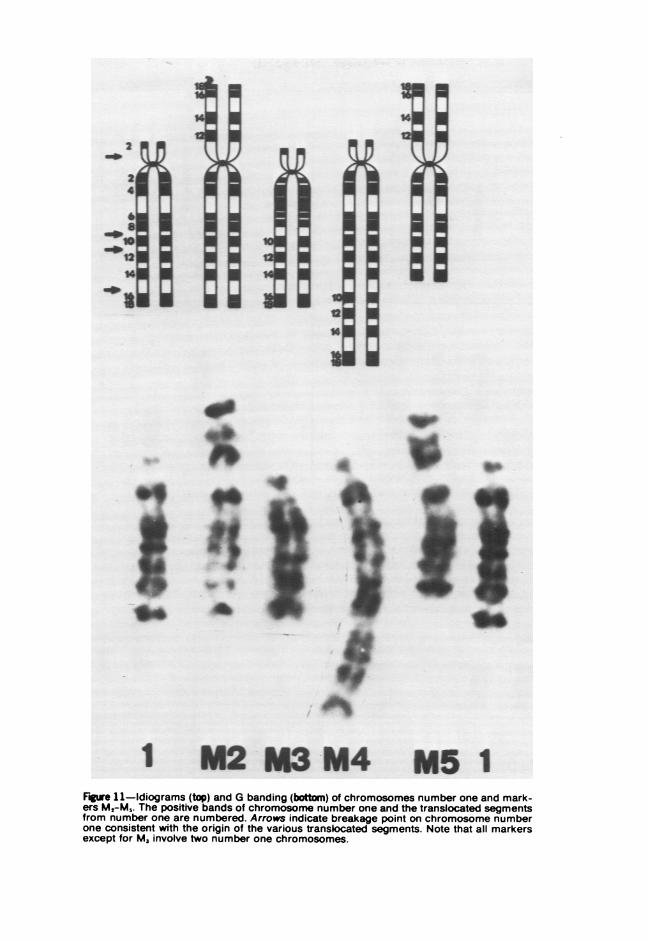
Guinea pig cells may also be transformed in vitro by chemical carcino-gens, but the process of transformation differs from that associated withtransformation of Svrian hamster cells; with guinea pig cells, the eventsleading to neoplasia are extended in time so that 4 to 18 months ofculturing mav pass before the cells produce progressively growing tu-mors.7 The chemical carcinogen-induced neoplastic transformation ofguinea pig cells need not be accompanied by numerical or morpholog-icallv visible chromosomal alterations.5" For example, it has been shownby G-band analvsis that cells transformed by a carcinogenic polycyclichvdrocarbon mav have a truly diploid constitution, or may be pseudo-diploid with an abnormal metacentric chromosome. Abnormal metacentricchromosomes may also arise in cultures transformed by carcinogens otherthan polyvyclic hvdrocarbons. Metacentrics need not be associated withtransformation since abnormal metacentric chromosomes also occurred incells from a morphologically nontransformed and non-tumor-producingcell line after extensive subculture. Some aneuploid cell lines may exhibita new submetacentric or subtelocentric chromosome originating fromtranslocation involving one or two number one chromosomes. Althoughchromosome number one was involved in three lines studied, the in-volvement is considered to be nonrandom because G-band analysisshowed that the new marker chromosomes involved different regions ofthe number one chromosomes (Figure 11). It has been suggested thatchromosome number one of the guinea pig is associated with nucleolarorganization.'M This has been confirmed using the AgAS technique.87 Thegreater frequencv of contact between such chromosomes creates an in-creased risk of chromatid exchange, possibly explaining their frequentparticipation in abnormal chromosome formation or nondisjunction.

Vol. 85, No. 3 IKARYULUGY OF TRANSFORMED CELLS 721December 1976
A high frequency of trisomv of the A2 chromosome was found in rats asa result of treatment with DMBA regardless of whether a carcinoma,"sarcoma," or leukemia90 developed. Thus, the type of chromosome altera-tion has been theorized to be dependent upon the etiologic agent andindependent of the type of cancer in the rat. Because of this interestingconclusion, we compared the chromosome banding patterns of rat fibro-sarcomas induced bv in vitro transformation of rat embrvo cells with thoseinduced by in vivo injections of rats with DMBA and in untreated controlcell lines.8 The numerical and/or structural chromosonal aberrations preferen-tiallv involved chromosomes number one, two, or three, whether or not thecell lines were transformed (Figure 12). Because of the absence of specificalteration in chromosome distribution or any specific and constantchromosomal aberrations, one must conclude that fibrosarcomas as aresult of in vivo or in vitro malignant transformation are not associatedwith specific chromosome changes such as trisomv A2. The preferentialinvolvement of these three autosomes is nonspecific; in rat cells trans-formed with adenovirus, SV40, and herpes viruses (Gallimore et al.,quoted bv McDougall 91) the same three autosomes in marker formationare involved. The frequent involvement of these three autosomes invirally or chemically transformed cells appears independent of the acutechromosome damage which may be specific after chemical agents butnonspecific after adenovirus infection.'1
All of our studies discussed thus far have utilized freshly derived cellsthat are considered normal in several respects, including chromosomalconstitution. The BALB/3T3 cells originally obtained from Aaronson andTodaro"2 can be cloned, and sublines derived that are sensitive to avarietv of chemical carcinogens. These cells produced tumors only afterbeing transformed by chemical carcinogens. The sublines are aneuploidand exhibit a normal heterochromatin pattern before and after transfor-mation by chemical carcinogens. Although each transformed line ana-lvzed had a characteristic modal chromosome number, transformationwas not accompanied by a general chromosomal modification of thecontrol hypotetraploid chromosome number. When two transformed linesobtained with SV40 were examined, one hypotetraploid and the othersubdiploid, the former had the normal heterochromatin distribution andlatter had an area of interstitial heterochromatin in both arms. It waspostulated that the interstitial heterochromatin was the result of inducedhost DNA due to viral infection.' Variants obtained by treatment withFudR from twice recloned DMBA-transformed BALB/3T3 cells hadgrowth parameters resembling the nontransformed cells. Some of thevariant cell lines had an increased number and some had a decreased
LeTA %Vf-.. %-. -- -- A r-% %Iae -pn

722 DIPAOLO AND POPESCU American Journalof Pathology
number of chromosomes compared to the parental cell lines, but eachvariant had unique marker chromosomes not present in the parentalDMBA-transformed line.93 Although it is unknown whether the variantsare true revertants (genotypic change), the chromosome distributionsindicate that the loss of the neoplastic property is accompanied bychanges in a relatively large number of chromosomes.
References1. Berwald Y, Sachs L: In vitro transfonnation of normal cells to tumor cells by
carcinogenic hydrocarbons. J Natl Cancer Inst 35:641-{661, 19652. Heidelberger C: Chemical oncogenesis in culture. Adv Cancer Res 18:317-360,
19733. Heidelberger C: Chemical carcinogenesis. Ann Rev Biochem 44:79-121, 19754. Casto BC, DiPaolo JA: Virus, chemicals and cancer. Prog Med Virol 16:1-47, 19735. DiPaolo JA, Donovan P, Nelson RL: Quantitative studies of in vitro transformation
bv chemical carcinogens. J Natl Cancer Inst 42:867-874, 19696. DiPaolo JA: Quantitative aspects of in vitro chemical carcinogenesis. Chemical
Carcinogenesis. Edited by POP Ts'o, JA DiPaolo. New York, Marcel Dekker, Inc.,1974, pp 443-455
7. Evans CH, DiPaolo JA: Neoplastic tranformation of guinea pig fetal cells in cultureinduced by chemical carcinogens. Cancer Res 35:1035-1044, 1973
8. Olinici CD, DiPaolo JA: Chromosome banding patterns of rat fibrosarcomas in-duced by in vitro transformation of embrvo cells or in vivo injection of rats by 7,12-dimethvlbenz(a)anthracene. J Natl Cancer Inst 52:1627-1634, 1974
9. DiPaolo JA, Takano K, Popescu NC: Quantitation of chemicallv-induced neo-plastic transformation of BALB/3T3 cloned cell lines. Cancer Res 32:2686-2695,1972
10. Chen TT, Heidelberger C: Quantitative studies on the malignant transformation ofmouse prostate cells by carcinogenic hydrocarbons in vitro. Int J Cancer 4:166-178,1969
11. Hubennan E, Sachs L: Cell susceptibility to transformation and cytotoxicity by thecarcinogenic hydrocarbon benzo(a)pyrene. Proc Natl Acad Sci USA 56:1123-1129,1966
12. DiPaolo JA, Donovan Pj, Nelson RL: In vitro transformation of hamster cells bypolycvclic hvdrocarbon: Factors influencing the number of cells transformed. Nature[New Biol] 230:240-242, 1971
13. Reitz MS, Saxinger WC, Gallo RC, Ting RC, DiPaolo JA: Evidence for the lack ofexpression of hamster type-C virus in chemical carcinogen transformed Svrian ham-ster cells. Proc Am Assoc Cancer Res 17:176, 1976
14. Makino S: A cytological study of the Yoshida sarcoma, an ascites tumor of whiterats. Chromosoma 4:649-674, 1952
15. Sandberg AA, Hossfeld DK: Chromosomal changes in human tumors and leu-kemias. Handbuch der allgemeinen Pathologie. Edited bv W Altmann, et al. Berlin,Springer-Verlag, 1974, pp 141-158
16. M itelman F, Levan G, Brandt L: Highly malignant cells with normal karyotype inG-banding. Hereditas 80:291-293, 1975
17. German J: Genes which increase chromosomal instability in somatic cells andpredispose to cancer. Progress in Medical Genetics, Vol VIII. Edited bv AG Stein-berg, AG Bearn. New York, Grune & Stratton, 1972, pp 61-102
18. DiPaolo JA, Popescu NC: Chromosome bands induced in human and S-rian ham-ster cells by chemical carcinogens. Br J Cancer 30:103-108, 1974

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 723December 1976
19. Nowell PC, Hungerford DA: A minute chromosome in human chronic granulocyticleukemia Science 132:1497, 1960
20. Mlanolov G, Manolova Y: Marker band in one chromosome 14 from Burkitt lvnm-phomas. Nature (Lond) 237:33-34, 1972
21. Zankl H, Zang, KD: Cytological and cytogenetic studies on brain tumors: Identi-fication of the missing G chromosome in human meningiomas as no. 22 by fluoren-cence technique. Humangenetik 14:167-169, 1972
22. Mark J: Karvotvpe patterns in human meningiomas: A comparison between studieswith G- and Q-banding techniques. Hereditas 75:213-220, 1973
23. Sakurai M, Sandberg AA: Chromosomes and causation of human cancer andleukemia. XI. Correlation of karvotypes with clinical features of acute mveloblasticleukemia. Cancer 37:285-299, 1976
24. Rowley J, Potter D: Chromosomal banding pattems in acute nonlymphocyticleukemia. Blood 47:705-721, 1976
25. Barski G, Belehradek J: Expression of malignancy in interspecies cell hvbrids.Differentiation and Control of Malignancv of Tumor Cells. Edited by W Nakahara,et al. Tokvo, Universit, Park Press, 1974, pp 419-441
26. Croce CM: Genetics of cell transformation by SV40. Biology of Radiation Carci-nogenesis. Edited by JM Yuhas, RW Tennant, JD Regan. New York, Raven Press,1976, pp 207-217
27. Stiles CD, Desmond W, Sato G, Saier M: Failure of human cells transformed bvsimian virus 40 to form tumors in athvmic nude mice. Proc Natl Acad Sci USA72:4971-4975, 1975
28. McDougall JK: Unpublished data29. DiPaolo JA: Karvological instability of neoplastic somatic cells. In Vitro 11:89-96,
197530. Mtoorhead PS. Nowell PC. Mellman WJ, Battips DM, Hungerford DA: Chromo
some preparations of leukocytes cultured from human peripheral blood. Exp CellRes 20:61.3-616, 1960
31. Caspersson T, Zech L Johanson C, Modest E: Identification of human chromo-somes bv DNA-binding fluorescent agents. Chromosoma 30:215-227, 1970
32. Sumner AT, Evans HJ, Buckland RA: New technique for distinguishing betweenchromosomes. Nature [New Biol] 232:31-32, 1971
32. Seabright M: A rapid banding technique for human chromosomes. Lancet2:971-972, 1971
34. Shiraishi Y, Yosida TH: Banding pattern analysis of human chromosomes bv use ofa urea treatment technique. Chromosoma 37:75-83, 1972
35. Aula P, Saksela E: Comparison of areas of quinacrine mustard fluorescence andmodified Giemsa staining in human metaphase chromosomes. Exp Cell Res71:161-167, 1972
36. Dutrillaux B, Finaz C, DeGrouchy J, Lejeune J: Comparison of banding patterns ofhuman chromosomes obtained with heating, fluorescence and proteolvtic digestion.Cvtogenetics 11:113-116, 1972
37. Gallimore PH, Richardson CR: An improved banding technique exemplified in thekarvotype analysis of two strains of rats. Chromosoma 41:259-263, 1973
38. Arrighi FE, Hsu TC: Localization of heterochromatin in human chromosomes.Cvtogenetics 10:81-86, 1971
39. Popescu NC, DiPaolo JA: Heterochromatin, satellite DNA, and transformed neo-plastic cells. J Natl Cancer Inst 49:60-606, 1972
40. DiPaolo JA, Popescu NC: Distribution of chromosome constitutive hetero-chromatin of Syrian hamster cells transformed by chemical carcinogens. Cancer Res33:3259-3265, 1973

724 DIPAOLO AND POPESCU American Journalof Pathology
41. Popescu NC. DiPaolo JA: Sequential G and C chromosome banding. Lancet1:209-210. 1974
42. Sumner AT: A simple technique for demonstrating centromeric heterochromatin.Exp Cell Res 75:304-:306. 1972
43. Howell WM. Denton TE: An ammoniacal-silver stain technique specific for satel-lite III DNA regions on human chromosomes. Experientia 30:1.364-1:366, 1974
44. Goodpasture C. Bloom SE: 'isualization of nucleolar organizer regions in mamma-lian chromosomes using silver staining. Chromosoma 53:3 7-30. 1973
43. Latt SA: Microfluorometric detection of deoxyribonucleic acid replication in hu-man metaphase chromosomes. Proc Natl Acad Sci USA 70:3395-3399, 1973
46. WN'olff S. Pen-v P: Differential Giemsa staining of sister chromatids and the study ofsister chromatid exchanges without autoradiography. Chromosoma 48:341-&53. 1974
47. Korenberg JR. Freedlender EF: Giemsa technique for the detection of sister chroma-tid exchanges. Chromosoma 48:3-360. 1974
48. Paris Conference (1971): Standardization in human cytogenetics. Birth Defects8:1-46. 1972
49. Committee for a Standardized Karyotype of Rattus Norvegicus: Standard Karyo-type of the Norwvay rat. Rattus norvegicus. Cytogenetics 12:199-205. 1973
350. Committee on standardized genetic nomenclature for mice: Standard karyotype ofmouse. NMus musculus. J Hered 63:69-72. 1972
31. Popescu NC. DiPaolo JA: Identification of Syrian hamster chromosomes by acetic-saline-Giemsa (ASG) and trvpsin techniques. Cytogenetics 11:300-507. 1972
32. Lehman JM. MacPherson I. Moorhead PS: Karvotype of the Syrian hamster. J NatlCancer Inst 31:6:39-61. 196:3
5:3. Popescu NC. Evans CH. DiPaolo JA: Chromosome patterns (G and C bands) of invitro chemical carcinogen-transformed guinea pig cells. Cancer Res 36:1404-1413.1976
34. Galton NI. Holt SF: DNA replication pattems of the sex chromosomes in somaticcells of the Svrian hamster. Cytogenetics 3:97-111. 1964
35. Hsu TC. Arrighi F: Distribution of constitutive heterochromatin in mammalianchromosomes. Chromosoma 34:243-233. 1971
.56. Pardue NIL. Hsu TC: Locations of 18s and 28s ribosomal genes on the chromo-somes of Indian muntjac. J Cell Biol 64:231-254. 1975
37. Hsu TC. Spirito SE, Pardue ML: Distribution of 18 + 28S ribosomal genes inmammalian genomes. Chromosoma 33:23-36, 1973
38. Shaw MNW: Human chromosome damage by chemical agents. Ann Rev NIed21:409-4:32. 1970
39. Caspersson T. Haglund U. Lindell B. Zech L: Radiation induced nonrandomchromosome breakage. Exp Cell Res 73:341-543, 1972
60. Seabright N: High resolution studies on the pattern of induced exchanges in thehuman karvotype. Chromosoma 40:3.33-346. 1973
61. Holmberg NI. Jonasson J: Preferential location of X-ray induced chromosomebreakage in the R-bands of human chromosomes. Hereditas 74:37-68. 1973
62. Ayraud N. Cantrelle C. Llyod N: Distribution des points de cassure chromoso-mique induit par l'arabinosvl cytosine sur des lymphocytes humains in vitro. AnnGenet 19:29-35. 1976
6;3. DiPaolo JA. Nelson RL. Donovan PJ: Norphological, oncogenic, and karvologicalcharacteristics of Syrian hamster embr-o cells transformed in vitro by- carcinogenicpolycyclic hydrocarbons. Cancer Res 31:1118-1127, 1971
64. Popescu NC. Casto BC. DiPaolo JA: Infrequent chromosome aberrations in Syrianhamster cells following partial synchrony by amino-acid deprivation. J Cell Phys86:399-604. 1973
65. DiPaolo JA. Donovan PJ. Popescu NC: Kinetics of Syrian hamster cells during X-

Vol. 85, No. 3 KARYOLOGY OF TRANSFORMED CELLS 725December 1976
irradiation enhancement of transformation in vitro by chemical carcinogen. RadiatRes 66:310-325. 1976
66. Hsu TC, Pathak S. Shafer DA: Induction of chromosome cross-banding by treatingcells with chemical agents before fixation. Exp Cell Res 79484-487, 1973
67. Levan G: Nomenclature for G-bands in rat chromosomes. Hereditas 77:37-52.1974
68. Sugivrama T. Goto K. Uenaka H: Acute cytogenetic effect of 2(2-furvl )-3-(5-nitro-2-furvl)-acrvlamide (AF-2. a food preservative) on rat bone marrow cells in vivo. MutatRes 31:241-246, 1973
69. Perrv P. Evans HJ: Cytological detection of mutagen-carcinogen exposure by sisterchromatid exchange. N'ature (Lond) 258:121-125, 1973
70. DiPaolo JA. Popescu NC. Nelson RL: Chromosomal banding pattems and in ritrotransformation of Syrian hamster cells. Cancer Res 33:3250-3258, 1973
71. Yamamoto T. Rabinowitz Z, Sachs L: Identification of chromosomes that controlmalignancy. Nature 243:247-250. 1973
72. Benedict WF. Rucker N. Mark C. Kouri RE: Correlation betueen balance ofspecific chromosomes and expression of malignancy in hamster cells. J Natl CancerInst 54:1374-162, 1973
73. Casto BC. Pieczynski N'J, DiPaolo JA: Enhancement of adenovirus transformationbv treatment of hamster embryo cells with diverse chemical carcinogens. Cancer Res:34:72-78, 1974
74. Popescu NC. Olinici CD. Casto BC, DiPaolo JA: Random chromosome changesfollowing SA7 transformation of Syrian hamster cells. Int J Cancer 14:461-472. 197d4
73. Yamamoto T. Hayashi NI. Rabinowitz A, Sachs L: Chromosomal control of malig-nancy in tumors from cells transformed by polvoma virus. Int J Cancer 11:355-366.1973
76. Copple CD. McDougall JK: Clonal derivatives of a herpes type 2 transformedhamster cell line (.33.3-8-9): Cytogenetic analysis. tumorigenicity and virus se-quence detection. Int J Cancer 17:01-310. 1976
77. Nachtigal MI. Duff R, Rapp F: Chromosome aberrations in Syrian hamster embryocells transformed after exposure to ultraviolet irradiated herpes simplex virus type 1or 2. J Natl Cancer Inst 34:97-105, 1973
78. Popescu NC. Cioloca L, Liciu F. Encut I: Chromosomal analysis of some trans-planted tumors induced by 3-methylcholanthrene in golden hamsters. Int J Cancer4:785-792. 1969
79. Barski G. Cornefert F: Caracteristiques carvologiques des tumeurs pulmonaires dehamster produites par lad6novirus 12. Ann Inst Pasteur (Paris) 107:114-120. 1964
80. Nachtigal MI. Popescu NC, Nachtigal S: Chromosomes of tumors induced by 3-methylcholanthrene in the golden hamster. J Natl Cancer Inst 38:697-722, 1967
81. NMacKinnon E. Kalnins VI. Stich HF. Yohn DS: Viruses and mammalian chromo-somes. VI. Comparative karvologic and immunofluorescent studies on Syrian ham-ster and human amnion cells infected with human adenovirus type 12. Cancer Res26:612-618, 1966
82. NMcDougall JK, Dunn AR, Jones KE: In situ hybridization of adenovirus RNA andDNA. Nature (Lond) 236::346-348, 1972
83. Dunn AR, Gallimore PH, Jones KW, NMcDougall JK: In situ hybridization ofadenovirus RNA and DNA. II. Detection of adenovirus-specific DNA in transformedand tumor cells. Int J Cancer 11:628-636, 1973
84. NMcDougall JK. Vause KE. Gallimore PH, Dunn AR: Cytogenetic studies in per-missive and abortive infected by adenovirus type 12. Int J Cancer 14:236-243. 1974
85. Loni CNM Green NM: Detection and localization of virus-specific DNA by in situhybridization of cells during infection and rapid transformation by the murinesarcoma-leukemia virus. Proc Natl Acad Sci USA 71:3418-3422. 1974

726 DIPAOLO AND POPESCU American Journalof Pathology
86. Ohno S. WNeiler C. Stenius C: A dormant nucleolus organizer in the guinea pig((aria cobaya. Exp Cell Res 25:498-503 1961
87. DiPaolo JA. Popescu NC: Unpublished data88. Ahlstrom U: Chromosomes of primary carcinomas induced b- 7. 12-dimethyl-
benz(a)anthracene in the rat. Hereditas 78:2:35-244. 197489. Levan G. Ahlstr8m U. \Mitelman F: The specificity of chromosome A2 involvement
in DMIBA-induced rat sarcomas. Hereditas 77:26:3-280. 197490. Kurita Y. Sugivama T. Nishizuka Y: Cx-togenetic studies on rat leukemia induced
bh- pulse doses ofT,. 12-dimethv-lbenz(a anthracene. Cancer Res 28:17:38-1752. 196891. McDougall JK: Adenoviruses: Interaction wvith the host cell genome. Prog Nied
Virol 21:118-1:32. 197592. Aaronson SA. Todaro GJ: Development of 3T;3-like lines from Balb c mouse em-
brvo cultures: Transformation susceptibility to SV40. J Cell Physiol 72:141-148. 19689:3. Olinici CD. Evans CH. DiPaolo JA: Chromosome patterns of nontransformed
variants from chemicallv transformed Balb 3T3 cells. J Cell Physiol 83:401-407.197,4

00o a
Co>E c>%E~eoX 0o
S~~~_qfp E _y0'flu
(..s_ e_* v
:Fsef
s
:::-
.St. n
.-L-4 v.- -
v
_.i " +e-X- -
:.-* <*ffi.Se t.. S- *1~~-jel.~~~
..^: Joh
mfe,4.: 7~ab *flPw=yZ~~~~~~~~~~~~~~~~~~~~~~~~~~I
in.t u
-
-lilk.. Us Mt
a t.a0 C.
* - Sow.- U
cLf* ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~..........ia. K
4.
C-.

cwz
: ; z EPD0o00
0?.
.. .. ...... ..
..... ... .....
N-
00
0
E0
Da-
CD 0
0 .0 0>) 00
O
0
.- OC-C
_000D. 0
0 0 0
o0 0000
Ex E-tr_ C1 0
e
- o%o0-
:;_ o _-
C D
'CLOc oZ< _Z>00
0. ZD 0
E0
ED 0
C00
E 00
E 00
= 0oE
cn o_coCE0C
coo
o 0
-00.C 0O1Cc Z-
_..::..., ..
I.
g,, _
z _.. ....... ... X.:r .... w.. :,.:. .) . * ....... t
..
Fgwe 3-G-band metaphase from Syrian hamster cells after 5 hours of treatment with N-methyl-N'-nitro-N-nitrosoguanidine (MNNG). Two chromosomes, A3 and B9 (arrows), each have a chromatid gap involving anegative band.
__._:* ..... ... - , :
......W
:,
....
Figure 4-Metaphases with chromatid exchanges found 24 hours after exposure to MNNG. Bandingresolution makes possible the accurate identification of the chromosomes involved and the points ofexchange.
..... ::.

.*
:*::
..... .._.... :_ . ......... ...
... ,....o:. ^ : w w..... . .. .. .. ... ..a:
E..... ... -
.:... ..^.. .... . . . ..... .*:. ^,
T] fulllv-*...... :w.,.f......... ..... w .. " =L ...* . ... -. .. ........ _
::::_r. _., ~~~~~~~~~~~~~~~~~~~~~~~~~....... ...._ .......~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~... .. _. .. . .Fgure 5-G-band metaphase of a rat cell with both A2 chromosomes having a gap after 5 hours oftreatment with 7,12-dimethylbenz[a]anthracene (DMBA). The gaps are at the Q2-4 band (arrow).
. . r cn
7E co5
0'
cma,C0-
C:a0o
CC)
0'
0j m
0moCo
D:C_
CI L
0
O Ca
C5D
En- C
LLJ t
(-) aC.)Q
U:
ccm. E
xa,aE
cc EEE0-CD <
cn
CD-',a
a, _
z a)
a)C
a)-
00> C>C: C~
Ea o
,Da
> vman °
r
<,2CZas )

-iX!0
,E

I I-I
S-0-
4c
0-
0 0
0 0
z zZ,I I
p:0
zq,
-i - I-
II
'U
-T-
]IU
E
- _
Si
', U
c
Co CO
_
CD
_ yEco
U
o
Er
_C-
0 c
-
cno
C)
0£
co.E
o0Eo
0t
_-_
0=
00
- C
E o
00
-o
0,
00
Co
Co
D <
or
Dz
C0
E 0
c E
EoC -
o =
CIO
_~~~~~~.oE
N
U)
_ >
I
M
U J
_-a
0s-.ia an
* 1
a6 a6 0a4
4
4,qa
q,a A
a
L-I aIN
I--IMEME9-1
F--
L._-JLL.=.L---mm.| -,
m
m
a w4c

0o
IC -I CLO
:E-'
E -cx
'° o cIC_X.(
."_"4.II"...
.......
.. -_ e _
-- a>S
A6
VW

.:-p..
U:
il::-jjkj- ::I . i _
J1 '
r-
I.
u,
..)
c
0C0
-
c*a)
*(D
0
: -
..Ii (1
m*20C_
C
'Ec C'J0D LA
-o
4:.:C.
L-C.. o.c..o
0-
U,
C) CN- 0
0Ecc.. --
cz
C oo o.. _
E °_
e mW uVat:>- ()C° _
*I.:
100L-CUa_
-0 'a( En
c U)
D E-
-o0En
0
CE
o0- 0
_c°* C3
I7-i
.4
..1.z'
ii j '-'ff% .......
.=
I
J.
I ..'i.
.it
il
J
i
ii
t
III
i.
I
.4ifi.
I*
.4.
:t.:
iV
4 -:4
'a ..fi on.
x.
.7
10
I-l"A'Au:..
WIWI%.7
%.
..
I9 I
!I,IIIpII
C _._I
I
II
I INaU
U
IIIII
Ipa
IIIIIIIII
IIIII
p
I.III
I
U0'
a
.
Figure 11-Idiograms (top) and G banding (botton) of chromosomes number one and mark-ers M2-Ms. The positive bands of chromosome number one and the translocated segmentsfrom number one are numbered. Arrows indicate breakage point on chromosome numberone consistent with the origin of the various translocated segments. Note that all markersexcept for M3 involve two number one chromosomes.
c AP
1 1
w
x
AI._

Ii..
a
I,
I.M.
Figure 12-Marker chromosomes and their derivation in nontransformed lines (a and b) isolated from DMBA-treated dishes; DMBA-transformed lines (c and d) and tumors induced in vivo (6 and fl.
.11
b
If'o
d
I
IMI 0


















