
Relation entre les deux souches bactériennes principales d ...composant de fertilisants agricoles...
92
i Université du Québec INRS-Institut Armand-Frappier Relation entre les deux souches bactériennes principales d’un biofilm dénitrifiant Par Jonathan Cloutier Mémoire présenté pour l’obtention du grade Maître des sciences (M. Sc.) en microbiologie appliquée Jury d’évaluation Président du jury et examinateur interne Examinateur externe Directeur de recherce Éric Déziel INRS-Institut Armand-Frappier France Daigle Université de Montréal Richard Villemur INRS-Institut Armand-Frappier
Transcript of Relation entre les deux souches bactériennes principales d ...composant de fertilisants agricoles...

i
UniversitéduQuébec
INRS-InstitutArmand-Frappier
Relationentrelesdeuxsouchesbactériennesprincipales
d’unbiofilmdénitrifiant
Par
JonathanCloutier
Mémoireprésentépourl’obtentiondugrade
Maîtredessciences(M.Sc.)enmicrobiologieappliquée
Juryd’évaluation
Présidentdujuryetexaminateurinterne
Examinateurexterne
Directeurderecherce
ÉricDéziel
INRS-InstitutArmand-Frappier
FranceDaigleUniversitédeMontréal
RichardVillemurINRS-InstitutArmand-Frappier

ii
Résumé
Lorsd’étudesportantsurunbiofilmprovenantd'unbioprocédéde
dénitrification,ilaétédéterminéqu’ilyavaitdeuxsouchesbactériennesquiétaient
lesplusabondantesentermedeproportiondelabiomasse.Cesdeuxsouches,NL23
etJAM1,ontéventuellementétédésignéescommenouvellesespècesbactériennes
HyphomicrobiumnitrativoransNL23etMethylophaganitratireduscenticrescens
JAM1.TandisquelasoucheNL23estenmesured'effectuertouteslesétapesde
dénitrification,lasoucheJAM1estincapablederéduirelenitriteetdoncnepeutpas
réduirecomplètementlenitrateenazotegazeux.Plusieursfacteursdansleur
relationsuggèrentunecertainecoopération.Cemémoireprésentelestravaux
effectuésafind’enapprendredavantagesurl’interactionentrecesdeuxbactéries
Dansunpremiertemps,desconditionsoptimalesdeco-culturesentrelesdeux
souchesontétédéveloppées.Lacaractérisationdeladynamiquedeconsommation
dunitrateetdunitriteadémontréquelesco-culturesdessouchesNL23etJAM1
n’étaientpasnécessairementplusrapidespoureffectuerladénitrificationquela
soucheNL23enculturepure.Parcontre,desco-culturesdelasoucheNL23avecun
mutantdelasoucheJAM1(inactifpourréduirelenitrate)ontdémontréune
dénitrificationplusrapidequelasoucheNL23enculturepure.
LarechercheaaussipermisdedémontrerquelasoucheJAM1apporteune
aidedecroissanceàlasoucheNL23enconditionsaérobiesetuneaideenréduction
denitrateenconditionsanaérobies.Parcontre,iln’apasétédémontréquela
soucheNL23procureunavantageàlasoucheJAM1.

iii
Remerciements
J’aimeraisremerciermondirecteurderecherche,RichardVillemur,de
m’avoirpermisdecomplétermamaîtrisedanssonlaboratoire,luiquiatoujoursété
disponiblepouroffrirsonaideetsesconseils.Sanssapatience,sacompréhension
etsagénérositéjen’auraispaspufinirleprojet.
J’aimeraisaussiremerciertouslesmembresdulaboratoiredeRichard
Villemurpourleuraide,enparticulierChristineMartineau,poursonaide
indispensabledansmesdébutsnerveux;KarlaVasquezpoursonsoutien,son
expérienceetsaluttepourl’entretienduIC;etFlorianMauffrey,poursesconseils
(etsesmutants).MerciaussiàGeneviève,Marc-AndréetValérie,ainsiqu’aux
membresduGRMEquiontrendul’expérienceencoreplusagréable.
Sanslesoutienmoraldemafamilleetdemesamis,cetteentrepriseaurait
étéimpossible.Merciàmesparents,àmasœuretàtouslesgensquiontfaitpartie
demaviedurantcettepériode.

iv
Tabledesmatières
1. Introduction..............................................................................................12. Revuedelittérature...............................................................................42.1. Dénitrification..........................................................................................................................................42.2. Biofilms.......................................................................................................................................................72.3. Osmoprotection.......................................................................................................................................82.4. Bioprocédés...............................................................................................................................................92.5. BiodômedeMontréal..........................................................................................................................102.6. Methylophaganitratireduscenticrescens.....................................................................................132.7. Hyphomicrobiumnitrativorans.......................................................................................................142.8. Co-cultures,partagederessources...............................................................................................152.9. Hypothèseetobjectifs........................................................................................................................17
3. Matérieletméthodes...........................................................................183.1. Souchesbactériennes..........................................................................................................................183.1.1. Mutants.............................................................................................................................................18
3.2. Culturedesbactéries...........................................................................................................................193.2.1. MilieuoptimaldelasoucheJAM1.........................................................................................193.2.2. MilieuoptimaldelasoucheNL23.........................................................................................213.2.3. Milieudecroissanceadaptéàlaco-culture......................................................................223.2.4. Milieuxsousformedegéloses................................................................................................23
3.3. Préparationdescultures...................................................................................................................233.3.1. Ensemencement............................................................................................................................243.3.2. Conditionsdeculture..................................................................................................................24
3.4. Récoltededonnées..............................................................................................................................253.4.1. Spectrophotométrie....................................................................................................................263.4.2. Chromatographieionique.........................................................................................................263.4.3. Extractiond’ADN..........................................................................................................................263.4.4. qPCR...................................................................................................................................................27
4. Résultats...................................................................................................304.1. Déterminationdesconditionsoptimalesdeculture.............................................................304.1.1. Suividecroissancepréliminaireenconditionsaérobies...........................................304.1.2. Suividecroissanceenconditionsanaérobies..................................................................314.1.3. Variationsproblématiques.......................................................................................................404.1.4. Réductiondelaquantitéinitialedenitrate......................................................................46
4.2. CaractérisationdelarelationentrelessouchesNL23etJAM1........................................494.2.1. Croissancedeculturespuresenprésencedesurnageant..........................................494.2.2. Recherched’unosmoprotecteur...........................................................................................534.2.3. Caractérisationapprofondiedelaconsommationdunitrate/nitriteàl’aidedemutantsdelasoucheJAM1......................................................................................................................57
5. Discussion................................................................................................645.1. Paramètresoptimauxpourlacroissancedesco-culturesdessouchesNL23etJAM1 645.2. Profildeconsommationdenitrateetdenitritelorsdeladénitrification...................665.3. LessouchesNL23etJAM1enbiofilm..........................................................................................70

v
5.4. Échangederessources.......................................................................................................................73
6. Conclusion................................................................................................75

vi
Listedestableaux
Tableau3.1:compositiondumélangedanschaqueréactiondeqPCR.28Tableau3.2:détailssurlatechniqueduqPCR.................................................29Tableau4.1:concentrationdeNaClpartubedanslarecherched’un
osmoprotecteur..............................................................................................................54

vii
ListedesfiguresFigure2.1:Représentationschématiquesimplifiéeducycledel’azote..............................................................4
Figure2.2:Voiemétaboliquedeladénitrification........................................................................................................6
Figure2.3:BioréacteurdubiodômedeMontréal.......................................................................................................11
Figure3.1:LessouchesNL23etJAM1.............................................................................................................................15
Figure3.2:Exempledecultureenanaérobiose...........................................................................................................25
Figure4.1:Croissanceenconditionsaérobies(InstantOcean15g/L).............................................................31
Figure4.2:Lacroissancedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies...........33
Figure4.3:Consommationdenitratedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................35
Figure4.4:concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................36
Figure4.5:Concentrationd’azotetotaledanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobie........................................................................................................................................................................................38
Figure4.6:Consommationdenitratepourchaqueréplicatdesco-cultures1:5........................................40
Figure4.7:Concentrationdenitritepourchaqueréplicatdesco-cultures1:5...........................................41
Figure4.8:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:5.....................................41
Figure4.9:Consommationdenitratepourchaqueréplicatdesco-cultures1:10......................................42
Figure4.10:Concentrationdenitritepourchaqueréplicatdesco-cultures1:10......................................43
Figure4.11:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:10...............................43
Figure4.12:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:5..................................44
Figure4.13:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:10................................45
Figure4.14:Consommationdenitratedesco-culturesdessoucheNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................47
Figure4.15:Concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enanaérobie....47
Figure4.16:Concentrationd’azotetotaldanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobies.....................................................................................................................................................................................48
Figure4.17:CroissanceenconditionsaérobiesdelasoucheNL23enprésencedesurnageant...........50
Figure4.18:CroissanceenconditionsaérobiesdelasoucheJAM1enprésencedesurnageant...........52
Figure4.19:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(0%deNaCl)................................................................................................................................................................................................55
Figure4.20:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1%deNaCl)................................................................................................................................................................................................55
Figure4.21:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1,5%deNaCl)................................................................................................................................................................................................56
Figure4.22:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(2%deNaCl)................................................................................................................................................................................................56
Figure4.23:Consommationdunitrateparlesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................59

viii
Figure4.24:Concentrationdenitritedanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................60
Figure4.25:Concentrationd’azotetotaldanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies)..........................................................................................61
Figure5.1:ProfildeconsommationdunitrateetdunitriteparlasoucheNL23lorsdeladénitrification..............................................................................................................................................................................66
Figure5.2:ProfildeconsommationdunitrateetdunitriteparlasoucheJAM1lorsdeladénitrification..............................................................................................................................................................................67
Figure5.3:Profildeconsommationdunitrateetdunitriteparuneco-culturedessouchesNL23etJAM1lorsdeladénitrification.............................................................................................................................................67
Figure5.4:Profildeconsommationdunitrateetdunitriteparuneco-culturedesespècesBacillussubtilisetPseudomonasdenitrificanslorsdeladénitrification.............................................................................68
Figure5.5:Supportenplastiquegarnid’unbiofilm...................................................................................................71
Figure7.1:Schémadelaméthodedecréationd’unbiofilm...................................................................................77

ix
Listedessiglesetdesabbréviations˚C:degrésCelsius
ADN:acidedésoxyribonucléique
DO600:densitéoptiqueàunelongueurd’ondede600nm
EPS:extracellularpolymericsubstances
FISH:fluorescentin-situhybridization
TSA:trypticasesoyagar
R2A:Reasoner’s2Aagar
ATCC:americantypeculturecollection
Pb:pairesdebases
FAM:carboxylfluorescein
BHQ:blackholequencher
rpm:rotationsparminute

x

1
1. Introduction
Lenitratereprésenteunpolluantpotentieldanslecontexte
d’environnementsaquatiquesnaturelsousimulés.Lamoléculeesttoxiquepourles
animauxàdesconcentrationsexcédantenviron30ppm(0,49mMd’azote).La
limiteimposéeàl’eaupotableauCanadaetauxÉtats-Unisestde10mg/Ld’azote-
nitrate(45mg/Ldenitrate),concentrationàlaquellel’eauestconsidérée
définitivementsécuritairepourleshumainslesplusvulnérables.Lenitriteest
encoreplustoxique:quelquesmilligrammesparlitresontsuffisantspourla
manifestationdeproblèmeschezlespoissonsetlalimiteacceptabledansl’eau
potableestde1mg/Ld’azotenitrite(3mg/Ldenitrite).Lenitrateestsouventun
composantdefertilisantsagricolesets’accumuledoncàhautesconcentrationsà
l’embouchuredecoursd’eau.Outreleseffetsdirectssurlasantédespopulations
humainesetanimales,ilpeutyavoirproliférationd’algues,hypoxieetautreseffets
reliéscausantdeschangementsnocifsàl’écosystèmedumilieu(Camargoetal.,
2005).Dansunsystèmeaquatiquefermé,commeunaquarium,l’accumulationdu
nitratepeutseproduiredemanièreinexorablepuisquel’eaudumilieuestrecyclée
etquelesorganismesvivantsensontdessourcescontinuelles.Unesolutionnon-
biologiqueestleremplacementtotaldel’eau,cequipeutserévélerêtredispendieux
etpeupratique.Unesolutionpotentielleestladénitrificationbiologiquepardes
organismesbactériens.
UndesécosystèmesenexhibitionauBiodômedeMontréalestlemésocosme
dufleuveSaint-Laurent,quicomporteungrandaquariummarindanslequelvivent
plusieursespècesanimalesoriginairedel’estuairedufleuveSaint-Laurent.
L’accumulationdunitrateétantproblématique,unbioréacteurfutinstallé,dans
l’espoirquedesespècesbactériennesdénitrifiantespuissentréduirelamolécule
toxiqueenazotegazeux.Cebioréacteurétaitcomposéd’unmécanismesupportant
lacroissanced’unbiofilmexhibantdel’activitédénitrifiante;cebiofilmestétudié
pardesmembresdulaboratoireduPr.RichardVillemurdepuis2000.Plusieurs

2
souchesbactériennesdistinctesenontétéisolées,etplusieursorganismesnon-
bactérienssontnotésd’ycohabiter.Deuxsouchesbactériennes,cependant,
composentlagrandemajoritédelacommunautébiologiquedubiofilm,entermes
debiomasseetd’activitédénitrifiante:lessouchesJAM1etNL23,touteslesdeux
méthylotrophesetanaérobesfacultatives.Depuisleurdécouverteetleur
caractérisationcommedesespècesdistinctes,ellessontdénomméesMethylophaga
nitratireducenticrescensetHyphomicrobiumnitrativorans,respectivement.
Ladénitrificationestunprocessusderéductiondunitrateetdecomposés
intermédiairesenquatreétapesprincipales.Lenitrateestd’abordréduitennitrite,
quiestréduitenacidenitrique,etensuiteenacidenitreuxpourfinalementproduire
del’azotegazeux.Àchacunedesétapesestassociéuneenzyme.LasoucheJAM1
n’effectuequetroisdesquatreétapesdedénitrification,nepossédantpasdegène
codantpourlanitriteréductase.Elleréduitlenitratedefaçonefficacegrâceàdeux
nitrateréductasesmembranaires,Nar1etNar2.Lasoucheestaussitoléranteaux
conditionssalées,exhibantunecroissancenormaledansunmilieucontenant
jusqu’à8%deNaCl.LasoucheNL23,quantàelle,effectuetouteslesétapesdela
dénitrification.Toutefois,sanitrateréductasepériplasmiqueexécutelaréduction
dunitratebeaucoupmoinsrapidementqueleduod’enzymesdelasoucheJAM1.
Contrairementàcettedernière,lasoucheNL23netolèrepasleseldanssonmilieu,
etuneconcentrationdeplusde1%NaClinhibesacroissance.Notamment,ceci
sembleparticuliercarlemilieudanslequelétaitcultivélebiofilmétaitmarinet
incluaitprèsde3%desel.
Puisquecesdeuxsouchesprincipalesétaientdeloinlesplusabondanteset
activesdubiofilm,ilestnaturellementintéressantdelesétudierenplusdedétails
defaçonindividuelle.Parcontre,unbiofilmestunecommunautécomplexede
microorganismes,etlessouchesNL23etJAM1interagissentdoncpresque
certainementàuncertainniveau.Ilestdoncégalementintéressantd’étudierla
relationentrelesdeuxsouches,spécialementlorsdeladénitrification.Ceprojetde
recherchetentededéterminers’ilexisteunecoopérationentrelesdeuxespèces

3
principalesdubiofilm.Cetterechercheserasurtoutaxéesurlepartagedesoxydes
d’azotemaisviseraégalementdetrouverd’autresélémentspouvantcontribuerà
unesynergie,telsquedesfacteursdecroissance,desmoléculesessentielles(ex:
vitamineB12),etc.

4
2. Revuedelittérature
2.1. Dénitrification
Leprocessusdedénitrificationestunedescomposantesmajeuresducycle
del’azote,oùl’azotecirculedansl’atmosphère,lesoletl’eausousdiversesformes
moléculaires.L’importancedecettecirculationprovientengrandepartiedela
dépendanced’organismesvivantsàl’azotepourlastructureetlaformationde
moléculesprotéiquesetnucléiques.Eneffet,aucunevieterrestrenepeutexister
sanscetélément,etlesorganismesexistantauseindemilieuxpauvresenazote
doiventdévelopperdesstratégiesspécialesdesurvie.
Figure2.1:Représentationschématiquesimplifiéeducycledel’azote Lecycledel’azoteestunsystèmecomplexederéactionschimiquesengrandepartiecatalyséespardesorganismesmicrobiens.(1)Fixationd’azote;(2)nitrification;(3)dénitrification;(4)co-dénitrification;(5)anammox;(6)l’oxydationd’ammoniacdurantlanitrification.(Hayatsuetal.,2008)

5
Ladénitrificationréfèrespécifiquementàtouteslesétapesdelaréductiondu
nitrateenazote(Fig.2.2).Danslaplupartdescas,cetteréductionestexécutéepar
desespècesbactériennesprofitantdupotentielénergétiquedesréactionschimiques
impliquées;leprocessusréduitséquentiellementlenitrateennitrite,enoxyde
nitrique,enoxydenitreuxetfinalementenazotegazeux,chaqueétapeoffrantun
avantageénergétiquedifférent.Chacunedesréactionsestcatalyséeparuneenzyme
spécifique:
- Lanitrateréductasecatalyselaréductiondunitrateennitrite,cequioffrele
plusd’énergiepotentielleparmilesétapesdeladénitrification.Deux
versionsdissimilatoiresdecetteenzymeexistent:uneversionmembranaire,
codéeparlesgènesnar,etuneversionpériplasmique,codéeparlesgènes
nap.Ilexisteégalementdesvariétésdenitrateréductasesassimilatoires.
(Zumft,1997)
- Lanitriteréductaseàsontourpermetlaréductiondunitriteenoxyde
nitrique,lapremièreformegazeusedesmoléculesazotéesdela
dénitrification.Ilyaaussideuxversionsdecetteenzyme:Cu-Nir,qui
contientunatomedecuivre,estcodéparlegènenirK;cd1-Nir,quicontient
uncytochromecd1,estcodéparlegènenirS.Cesdeuxvariétésd’enzymes
n’ontjamaisétéretrouvéesensembledanslamêmeespèceetaucuneétude
n’apurévélerunedifférencefonctionnelleentreelles.(Priemeetal.,2002,
Zumft,1997)
- L’oxydenitriqueréductasecatalyselaréductiondel’oxydenitriqueenoxyde
nitreux.Laversiondel’enzymequiestprésentedanslesbactéries
dénitrifiantesestcNor,quicontientuncytochromecetestcodéparlesgènes
nor(unitécatalytiquecodéeparnorB).Ilexisteégalementd’autresversions
del’oxydenitriqueréductasedanscertainesespècesbactériennesquifont
partiedusystèmedeleursdéfensescontrelatoxicitédel’oxydenitrique.
(Hinoetal.,2012)

6
- L’oxydenitreuxréductasecatalyselaréductiondel’oxydenitreuxenazote
gazeux.Cetteenzymecontientunatomedecuivreetestcodéeparlesgènes
nos(unitécatalytiquecodéeparlegènenosZ).(Pauletaetal.,2013)
Figure2.2:Voiemétaboliquedeladénitrification Lavoiecomplètededénitrificationtransformelenitrateenazotegazeuxparunesériederéactionschimiquescatalyséespardesenzymes(surlignées).Lesmoléculessontlenitrate(NO3),lenitrite(NO2),l’oxydenitrique(NO),l’oxydenitreux(N2O)etl’azotegazeux(N2).
Cesenzymespeuventsemanifestersousdesformesdifférentesetavoirdes
activitésvariablesselonl’espèce.Puisqu’elleoffreunavantageénergétique
inférieuràlarespirationaérobie,ladénitrificationestprincipalementutilisépardes
espècesstrictementanaérobiesoucommealternativepardesbactériesanaérobies
facultatives.Seuleunefractiond’espècesbactériennespossèdelacapacité
d’effectuerladénitrificationetcelles-cin’eneffectuentpasnécessairementtoutes
lesétapeselles-mêmes(Zumft,1997).Plusieursespècesbactériennessecontentent
d’effectuerseulementlarespirationdunitrate,tellequeSerratiamarcescens,quine
peutproduiredel’oxydenitriquequeparlaréductionnon-enzymatiquedunitrite
(Andersonetal.,1986).Celles-ciconstituentleplusimportantsous-groupede
bactériesprofitantdelaréductiond’oxydesd’azote(Gregoryetal.,2003).D’autres
espècesseserventplutôtd’enzymesderéductioncommemécanismededéfense
contrelesmoléculestoxiques.L’enzymeqNorestutilisépardesbactériescomme
Neisseriagonorrhoeaepoursedébarrasserd’oxydenitrique(Hinoetal.,2012).De

7
plus,certainesbactériesréduisentdesmoléculesintermédiairesdeladénitrification
enconditionsaérobies.Parexemple,l’espèceParacoccuspantatrophus
(précédemmentnomméeThisphaerapantatropha)estenmesuredeperformerune
dénitrificationcomplèteenprésenced’oxygène.Ilestpossiblequecetteactivité
permetteàlabactériededissiperlesmoléculesréductricesetretrouverun
équilibreredox.(Berksetal.,1995)
2.2. BiofilmsDanslanature,lavastemajoritédelaviemicrobienneexistesousformede
biofilm,etnondiffuseenmilieuliquide.Unbiofilmestcomposéd’organismesde
classificationstaxonomiquesvariéesainsiqued’unematricecomplexede
polymèresbiologiquesgénéralementconnussouslenom«substances
polymériquesextracellulaires»(enAnglais,EPS).Celles-cisontproduites
directementparlesorganismesvivantdanslebiofilmousontmodifiéespardes
enzymesproduitesparcesorganismes.Cependant,lacompositionexactedela
structured’unbiofilmestdifficileàdéterminer.LesEPSsontintimement
impliquéesdanslaviemicrobienned’unbiofilm,etleursfonctionssontmultiples.
LesEPSpeuventagircommesourcedenourriture,faciliterl’échangegénétique
entrebactéries,faciliterlarétentiond’eau,fourniruneprotectioncontredes
facteursantibactériensetbiend’autreschoses.L’architectured’unbiofilmestaussi
hautementvariéeetdépenddel’organisationdesEPS.Ilpeutyavoirdescanauxet
descompartiments,etlaformedubiofilmpeutvarierselonlasituation.Unedes
composantesessentiellesàlaformationd’unbiofilmestlagammedemolécules
(protéines,ADN,polysaccaridesetmoléculesamphiphiles)quiserventàl’adhésion
debactériescolonisatricesàunesurface.Eneffet,lapremièreétapedansla
formationd’unbiofilmestl’attachementàunesurface,suividelaformationd’une
microcoloniepardivisioncellulaire.Cen’estqu’aprèsl’adhésionoriginalequeles
autrescomposantesdelamatricedubiofilmsontproduitesetqued’autresespèces
bactériennesysontrecrutées.Sanslaprésenced’unesurfacepropice,lesbactéries

8
nepeuventpasformerdebiofilm,etrestentenphaseplanctoniqueous’agrègenten
d’autresformes(ex:floc).(Flemmingetal.,2010)
Ilestaussipossiblequ’unbiofilmsoitnécessairepourlafonctionde
certainessubstancesrelâchéespardesbactéries.Ilaétédocumentéquecertaines
souchesbactérienneshalophilespouvaientrelâcherdesparticulesosmoprotectrices
(ectoïne,glutamate)dansleurmilieupouraideràlasurviedebactériesdansleur
entourage.(Plakunovetal.,2008)
2.3. OsmoprotectionLorsqu’ellessontconfrontéesàdesfluctuationsdel’osmolaritédedeleur
milieu(comme,parexemple,uneaugmentationensel),lesbactériesdoivent
équilibrerlesgradientsosmotiquescrééspoursurvivre.Pouratteindrecetobjectif,
ellespeuventproduiredesosmoprotecteurs(ousolutéscompatibles)oulesobtenir
dansleurenvironnement.Unosmoprotecteurestunemoléculeavecunpotentiel
osmotiqueélevéquin’interfèrepasaveclesprocessusmétaboliquesd’unecellule,
mêmeenhautesconcentrations.Enl’accumulantdanssoncytoplasme,unebactérie
peutréduirelapressionosmotiqueentrelesdeuxcôtésdesamembrane.(Kempfet
al.,1998)
Ilexisteunevariétéd’osmoprotecteurs.Ceux-ciincluent:desdiholosides
commelesaccharoseetletréhalose;despolyolscommeleglycérol;ainsiquedes
acidesaminés(etdérivés)commelaproline,lesbétaïnes,lacarnitineetl’ectoïne
(Kempfetal.,1998).Plusieursbactériesontdessystèmescomplexesenplacepour
lerèglementfindeleurréponseàunstressosmotique.L’espècebactérienne
Bacillussubtilis,parexemple,faitlasynthèsedelaprolinepourseprotéger,mais
ellepossèdeaussiuntransporteurspécifiqueàlacapturedeprolinedans
l’environnement.Ellepeutaussicapturerdespeptidesetenextrairedelaproline
avecdesprotéases,unestratégiequiestadoptéeparplusieursautresespèces
également(ex:Listeriamonocytogenes,Lactobacilluscasei,Oenococcus

9
oeni)(Zaprasisetal.,2013).Deplus,certainesbactériespeuventfaireusagede
plusieursosmoprotecteursdifférents.Escherichiacoliestcapabledesynthétiserle
tréhaloseetlaglycinebétaïne,ainsiqued’utiliserlaprolineetl’ectoïneretrouvées
danssonenvironnement(Jebbaretal.,1992).L’ectoïne,dérivédela
tetrahydropyrimidine,estl’osmoprotecteurquelasoucheJAM1semblepouvoir
produire(voirsection2.6).Cetosmoprotecteurestparticulièrementintéressant
danslecadredescommunautésbactériennestolérantesauselcarilaétédémontré
quedesbactériesnon-halophilespeuventenfaireusagepourobtenirunetolérance
aupressionsosmotiques(Jebbaretal.,1992).
2.4. BioprocédésUnbioprocédéestuneapplicationdescapacitésdemicroorganismespourla
productiondebiensoupourd’autresprocessusutiles.Cesapplications
biotechnologiquessontcourammentutiliséesdansunepanopliededomaines,
incluantlaproductionalimentaire,laproductiondemoléculespourutilisation
pharmaceutique,etc.
Lapurificationd’eauuséeimpliquesouventdesbioprocédés,dontles
systèmesdeboueactivée,quidépendentdel’activitédebiomasseenflocdansdes
réservoirsaéréspourpurifierl’eauquis’yretrouve(Roediger,1983).Les
bioréacteursmembranaires(«Membranebioreactor»enAnglais,ouMBR)
combinentunsystèmedeboueactivéeavecuneultrafiltrationpourpurifierdel’eau.
Cettetechnologiereprésenteunmarchéévaluéàdescentainesdemillionsde
dollarsetresteunsujetderechercheactif(Le-Clechetal.,2006,Wangetal.,2016).
Labiomasseutiliséedansunbioprocédépeutaussiêtrecultivéesousformede
biofilmfixéàunsupportàl’aided’unbioréacteuràlitfluidisé,commedanslecas
d’untraitementdelixiviatdansunedécharged’ordures(Hajipouretal.,2011)oude
dégradationdesubstanceshuileusesparunbiofilmfongique(Pakulaetal.,1996).
Cependant,lesbiofilmspeuventaussiêtreproblématiquesdanslesbioprocédés.
DansunMBR,laformationd’unbiofilmsurlamembranedefiltrationmèneàla

10
dégradationdecelle-ci.Ceciestconsidérécommeundesproblèmesprincipaux
dansledéveloppementdesMBR(Le-Clechetal.,2006).
Ladénitrificationpeutelleaussiêtreexploitéecommebioprocédé.Lenitrate
etlesautrescomposésintermédiairesdeladénitrificationreprésententtousdes
dangerspotentielspourl’environnementainsiquepourlasantéd’organismes
vivants,incluantleshumains.Pourcetteraison,ledéveloppementdeméthodesde
dénitrificationenbioréacteuresttrèsactif.Undomained’attentionparticulièreest
letraitementd’eaupolluéeparl’industrieagroalimentaire.Plusieursétudes
récentesontinvestiguél’efficacitédebioréacteursavecdifférentessourcesde
carbonesetunevariétédeparamètres.Lesrésultatsmontrentengénéralune
améliorationpotentielleaveccestechniquesparrapportsauxtechniquesde
traitementtraditionnelles(créationdezoneshumides,constructiondefossés
écologiques,etc)(Liangetal.,2015).
2.5. BiodômedeMontréalLeBiodômedeMontréalainstalléen1998unbioréacteuràlitfluidisépour
remédieràunniveaudangereusementélevédenitrate(environ15mM)dansl’eau
deleuraquariumd’eaumarine(Fig.2.3).Àl’intérieurduréacteurseretrouvaitun
biofilmdénitrifiantnourrideméthanoletformédebactériespréalablement
présentesdanslemilieu.L’espoirétaitquelebioréacteurpourraitgarderla
concentrationdunitratedissousàdesvaleursplussécuritaires,danslebutd’éviter
dessolutionsplusdrastiques,commeleremplacementd’eau.Lesystèmede
dénitrificaitonn’apuatteindrecesattentes,etdoncdeseffortsd’optimisationdela
performancedubioréacteurontétéentreprises.(Labbé,2007)
Ilaétédémontréqu’ilétaitpossibledegénérerunbiofilmcapablederéduire
uneconcentrationde50mgd’azoteparlitreàprèsde1mgd’azoteparlitre.Le
biofilmétaitformédebactériesprésentesdansl’eaumarinedumésocosmedu
FleuveSt-LaurentduBiodômeetétaitcultivéenmodecontinudansunréacteuràlit

11
fluidisé,maisàuneéchellepilote,beaucoupmoinsvolumineusequedans
l’aquariumoriginal.Ilaétéconcluquelebioréacteurdénitrifiantpourraitêtre
efficace,maisqueplusieursfacteurs(épaisseurdubiofilm,dégradationdesupports,
zonesmortes)quiétaientfacilesàcontrôlerenlaboratoiredevenaientdesobstacles
importantàl’applicationduprocédéàgrandeéchelle.(Labelleetal.,2005)
Figure2.3:BioréacteurdubiodômedeMontréal Bioréacteurdedésoxygénation(A)etdedénitrification(B)pourlemésocosmeduFleuveSt-LaurentauBiodômedeMontréal.Installéen1998.(Labbé,2007)
Parlasuite,lacompositionmicrobiologiquedubiofilmdubioréacteuraété
investiguée.Delabiomasseissuedubiofilmaétécultivéesurtroistypesd’agar
(TSA,R2AetWinogradsky).TroisisolatsontétéobtenusetleurADNribosomal16S
analysés,cequiapermisdedéterminerqu’ilsétaientdesα-protéobactériesdes
genresHyphomicrobium,Paracoccus,etd’unnouveaugenrebactérien.Lasouchedu

12
genreHyphomicrobiumaéténomméeNL23etaétécaractériséecommeundes
acteursprincipauxdel’activitédénitrifiante(Labbéetal.,2003).L’espècedu
nouveaugenreasubséquemmentétécaractériséecommenouvelleespèce
dénomméeNitratireductoraquibiodomus(Labbéetal.,2004).Lesbactériesnon-
cultivablesdubiofilmontétéétudiéesàl’aided’unegénothèqued’ADNribosomal
16Stotaldubiofilm.Ilaétédéterminéque70%desclonesgénérésavaientla
séquenced’uneγ-protéobactériedugenreMethylophaga.Lesautresprofils
indiquaientlaprésenced’unevariétédeséquencesreliéesàdesespèces
bactériennesdénitrifiantes.DesétudessubséquentesparFISH(«Fluorescentin
situhybridization»)ontconfirmél’abondancedominante(jusqu’à79%)desouches
appartenantaugenreMethylophagaetl’importancedugenreHyphomicrobium(7-
8%).Ilaaussiétédéterminéquelebiofilmincluaitenviron15espèces
bactériennes.(Labbéetal.,2003,Labbéetal.,2007)
Ilétaitsurprenantàl’époquequedesespècesdugenreMethylophaga
puissentcomposeruneaussigrandeproportiond’unbiofilmenconditions
anaérobies;legenreétaitjusque-làcaractérisépardesespècesstrictementaérobes.
Àl’aidedeméthanolmarquépardesisotopesdecarbone(14C)etd’analysedes
séquences16Sdanslesbactériesl’ayantassimilé,ilaétédémontréqu’uneespèce
dugenreMethylophagaétaithautementimpliquéedansleprocessusde
dénitrification.Unefoisisolée,cettesoucheaéténomméeJAM1;celle-ciaété
désignéecommeespècedistincteaprèsdesétudesdecaractérisationapprofondies
(voirsection2.6).LesgènesnarG1etnarG2ontaussiétécorrélésavecl’activitéde
réductiondenitratedelasoucheJAM1.Lesrésultatsd’analysesbioinformatiques
ontsuggéréuneoriginedetransferthorizontalpourlegènenarG1.(Auclairetal.,
2010)

13
2.6. MethylophaganitratireduscenticrescensLorsdesexpériencesexplorantladiversitémicrobienned’unbiofilm
provenantdel’aquariumdumésocosmeduFleuveSt-LaurentauBiodômede
montréal,ilfutdéterminéqu’unedessouchesbactériennesreprésentaitenviron
79%detoutelabiomasseprésente.L’identitédecettesouchen’étaitpasconnueà
l’époque,maisilavaitétédéterminéqu’elleétaitétroitementapparentéeaux
espècesdugenreMethylophaga.(Labbéetal.,2007)Suivantdesrecherchesplus
poussées,cettesouche,initialementnomméeJAM1,futdésignéecommenouvelle
espèceMethylophaganitratireduscenticrescens.(C.Villeneuveetal.,2013)
LegenreMethylophagaestunedivisiondesγ-protéobactériescaractérisée
pardesespècesbactériennesdeformebacille,typiquementlégèrementhalophiles.
Lesespècesmembresdugenresontaussidécritescommeconsommatricesde
composésàuncarbone(saufleméthane)dontleméthanol,qu’ellesassimilentà
l’aidedelavoiemétaboliqueduribulosemonophosphate.(Janvieretal.,1995)
Typiquement,lesespècesdugenreMethylophagasontdesespèceshabitantdes
milieuxmarinsoùellespeuventconsommerleméthanolabondammentproduitpar
desmicroorganismes(phytoplancton)(Minceretal.,2016).Plusieursespècesdu
genresontaussireportéesd’êtrecapabledelaproductiond’osmoprotecteur,
notammentl’ectoïne.CesespècesincluentM.thalassica,M.alcalica,M.murataet
autres(Doroninaetal.,2005,Mustakhimovetal.,2012,Mustakhimovetal.,2009).
LasoucheJAM1esttoléranteaumilieuxsalésjusqu’à8%deNaCl,malgréque
sacroissanceoptimaleestobtenuedansdesmilieuxd’environ3%deNaCl.Elleest
capabledecroissanceentredestempératuresde15˚Cà37˚Cmaispréfèreune
températureautourde30˚C.UnpHentre6et11permetlacroissance,maislepH
optimumestde8.ElleestauxotrophepourlavitamineB12,etrequiertdonc
l’inclusiondecettemoléculedanslemilieupourlacroissance.Surgélosedumilieu
1403(milieuadaptéauxsouchesbactériennesmembresdugenreMethylophaga),
sescoloniessontrondesetontenviron2mmdediamètre.Lasoucheestleseul

14
membredugenreMethylophagaàêtreanaérobefacultative,c’est-à-dired’être
capabledecroissanceenutilisantlenitrate(Auclairetal.,2010,C.Villeneuveetal.,
2013)etl’oxydenitreux(Mauffrey,résultatsnon-publiés)commeaccepteurs
d’électron,maispréférantl’oxygènedanscerôles’ilestdisponible.Ellepeut
égalementréduirel’oxydenitriteenoxydenitreux(Mauffrey,résultatsnon-publiés)
maispaslenitrite(Auclairetal.,2010).
2.7. HyphomicrobiumnitrativoransOutrelasoucheJAM1,uneautrebactériecomposaitunpourcentage
considérable(7-8%)delabiomassetotaledubiofilm.(Labbéetal.,2007)Celle-ci
futdésignéeNL23etparlasuitefutelleaussidécritecommeunenouvelleespèce,
Hyphomicrobiumnitrativorans.(Martineauetal.,2013)
LegenreHyphomicrobiumfaitpartiedesα-protéobactéries.Laparticularité
desespècesmembresestleurproductiond’hypheslorsdelareproduction.Les
bactériessontaussidécritescommebacillesauxboutspointus.Aussi,étantdonné
deleurfaibletoléranceauxenvironnementssalés,ellessontrarementretrouvées
dansdesmilieuxmarins.Plusieursmembresdugenreontétéimpliquésdansla
dénitrification,dontH.zavarziniietH.denitrificans,entreautres(Fesefeldtetal.,
1998,Martineauetal.,2015).Onretrouvecourammentcesbactériesdansles
bioprocédésdetraitementd’eauxusées.(Gliescheetal.,2005)
LasoucheNL23netolèrepasunmilieusalin,cequiestsurprenanten
considérantlecontextedesadécouverteetdesonactivité;lacroissancemaximale
estobtenueenconditionsde0-0,5%deNaCl.Presqueaucunecroissancen’est
possibleenmilieuxavecplusde1%deNaCl.LasoucheNL23peutcroîtreàdes
températuresentre15˚Cet35˚C,maispréfèreunetempératurede30˚à35˚C.LepH
optimalestentre7,5et8,5,maisunpHentre7et9,5estsuffisantpoursupporter
unecertainecroissance.CommelasoucheJAM1,lasoucheNL23estanaérobe
facultative,préférantlesenvironnementsoxygénésmaisétantcapabledecroissance
15
efficaceenutilisantlenitratecommeaccepteurd’électronspourgénérerdel’azote
gazeux.Surgélose,lescoloniesforméessontdepetitdiamètre(~1mm)etblanches.
LasoucheNL23peuttirersoncarboneduméthanol,delaméthylamineoudu
formate.(Martineauetal.,2013)
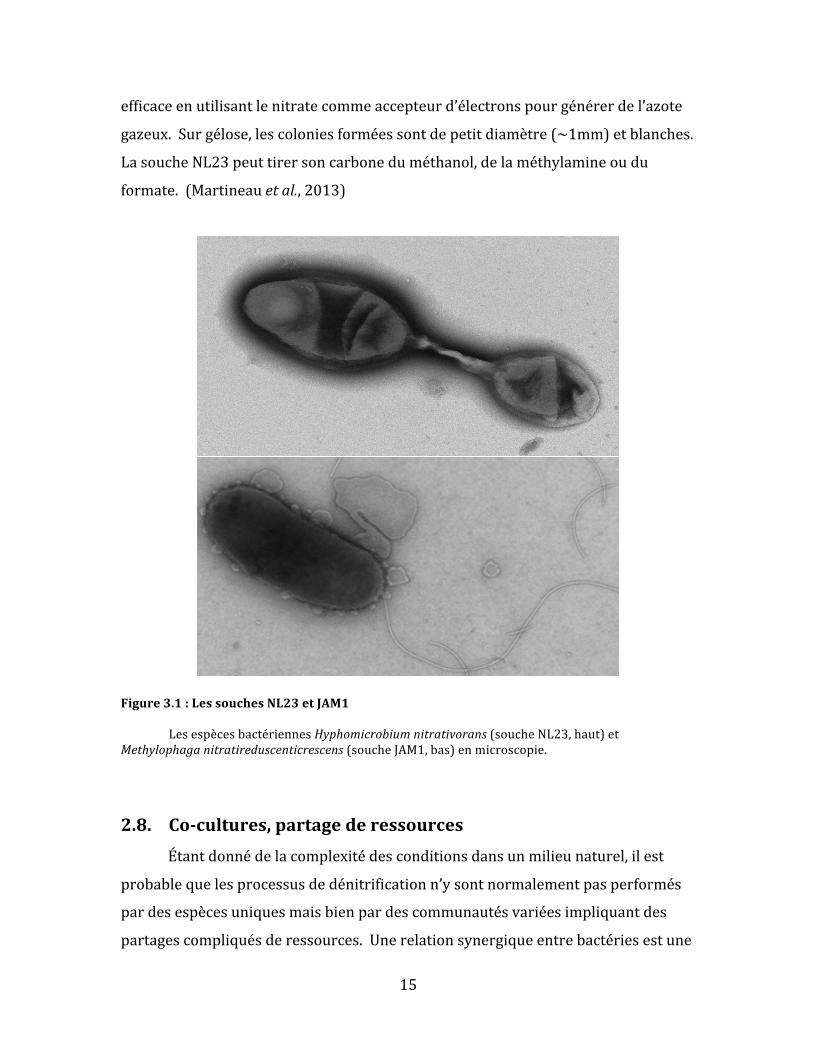
Figure3.1:LessouchesNL23etJAM1
LesespècesbactériennesHyphomicrobiumnitrativorans(soucheNL23,haut)etMethylophaganitratireduscenticrescens(soucheJAM1,bas)enmicroscopie.
2.8. Co-cultures,partagederessourcesÉtantdonnédelacomplexitédesconditionsdansunmilieunaturel,ilest
probablequelesprocessusdedénitrificationn’ysontnormalementpasperformés
pardesespècesuniquesmaisbienpardescommunautésvariéesimpliquantdes
partagescompliquésderessources.Unerelationsynergiqueentrebactériesestune

16
coopérationquiestbénéfiquepourtouteslesbactériesimpliquées.Ilexistedéjàdes
casdetellesinteractionsenco-culturesdénitrifiantesdocumentéesdansla
littératurescientifique.Parexemple,ilaétédémontréquel’espècebactérienne
Ralstoniapickettiauneactivitédénitrifiantede40à100foissupérieurelorsqu’elle
estcultivéeenprésencedel’espèceStreptomycesgriseus.L’aideétaitfourniepar
cettedernièresousformedeprotéasessecrétéesquiaugmentaientlaconcentration
d’histidinedanslemilieu.Cettehistidineaensuiteétéenmesured’augmenterle
tauxdedénitrificationdeR.pickettipourdesraisonsinconnues.(Takakietal.,2008)
Unautreexempledesynergieestlaco-culturedel’espèceAlcaligenesfaecalis
aveclasouchePseudomonasG9.Lapremièreestunebactériedénitrifiantequiest
incapablederéduirelenitrateennitrite,maisquiperformelesautresétapesdela
dénitrification.Àl’inverse,lasoucheG9exprimesanitrateréductase
constitutivement.Celle-ciperformeaussilesautresétapesdeladénitrificationmais
souffred’uneaccumulationtoxiquedenitriteenculturepure.Deplus,ellenepeut
pascroîtrelorsqu’elleestinoculéeenconditionsanaérobies.Enconditions
dénitrifiantes(anaérobies)avecseullenitratecommeaccepteurd’électronfourni,
chaquebactérieestincapablededémontrerunecroissanceenculturepure.Ila
pourtantétédémontréqu’uneco-culturedesdeuxbactériesaétécapablede
croissanceetd’activitédénitrifianteefficace.(VandePas-Schoonenetal.,2005)

17
2.9. HypothèseetobjectifsCertainesdescaractéristiquesuniquesàchaquesouche(NL23etJAM1)
laissentimaginerqu’ilpuisseexisteruncertainéchangederessources.Lasouche
JAM1nepeutpasréduirelenitrite,tandisquecettecapacitéestdisponibleàla
soucheNL23.Demême,lasoucheNL23nepeutexisterenprésencedehautes
concentrationsdesel;sepourrait-ilquelasoucheJAM1puissepartagerune
certaineosmoprotectiondelaquelleelledispose?Leportraitfinalseraitun
mutualismequiamélioreraitsubséquemmentl’efficacitédeladénitrification.
Donc:
Hypothèse:ilexisteunecoopérationentrelesNL23etJAM1enco-culturesous
formedepartagederessources(nitrate,nitrite,vitamineB12)oud’autresfacteurs
(ex:osmoprotection/ectoïne).
Objectifs:
• l’optimisationdesconditionsdelaco-culturedessouchesNL23etJAM1danslebutd’enfaciliterl’étude;
• lacaractérisationdesinteractionsentrelesdeuxsouches.

18
3. Matérieletméthodes
3.1. SouchesbactériennesLessouchesbactériennesdeceprojetontétéisoléesàpartird’unbiofilm
dontl’origineestlebioréacteurdénitrifiantdumésocosmeduFleuveSt-Laurentdu
BiodômedeMontréal.Celles-cisont:Methylophaganitratireducenticrescens
(soucheJAM1),Hyphomicrobiumnitrativorans(soucheNL23)(Fig.3.1).Plusieurs
échantillonsdechaqueespèceontétéconservéesà-80˚Cdansleursmilieuxde
culturerespectifscomplémentésde25%deglycérol.
3.1.1. Mutants
Enplusdesdeuxsouchesbactériennesci-dessus,troismutantsdetype
«knockout»delasoucheJAM1ontétéutilisés.Ceux-ciexhibenttousdes
déficiencesencequiconcerneleursenzymesdelanitrateréductase:JAM1ΔnarG1
estunmutantsanscopiedugènenarG1;JAM1ΔnarG2estunautresanscopiedu
gènenarG2;etJAM1ΔnarG1narG2estundouble-mutantnepossédantaucunecopie
desgènesnarG1etnarG2.Cessouchesontétécrééesparlaméthodedeladouble
recombinaisonparFlorianMauffrey,étudiantdedoctoratdanslelaboratoireduPr.
Villemur(Mauffreyetal.,2015).

19
3.2. Culturedesbactéries
3.2.1. MilieuoptimaldelasoucheJAM1LemilieuoptimalpourlacroissancedelasoucheJAM1telquedéterminé
danslestestsd’optimisationpréalables(Villeneuveetal.,2013)estlemilieuATCC
1403pourespècesdugenreMethylophaga.Unlitredumilieuliquidecontient:
NaCl----------------------------------------------24g
MgCl2·6H2O--------------------------------------3g
MgSO4·7H2O------------------------------------2g
KCl------------------------------------------------0,5g
CaCl2·2H2O--------------------------------------1g
Bis-Tris------------------------------------------0,5g
SolutionT-------------------------------------20ml
SolutiondeWolf-----------------------------10ml
Eaudistillée---------------------------------970ml
LasolutionTestunesolutionminéraledontlacompositionestlasuivante(pour1L
desolution):
KH2PO4--------------------------------------------7g
NH4Cl------------------------------------------100g
Bis-Tris----------------------------------------100g
(NH4)5Fe(C6H4O7)2------------------------------3g
Eaudistillée-------------------------------1000ml

20
LasolutiondeWolfestuneautresolutionminéraleenutilisationcourante.Unlitre
delasolutioncontient:
N(CH2CO2H)3-----------------------------------1,5g
MgSO4·7H2O------------------------------------3g
MnSO4·H2O------------------------------------0,5g
NaCl------------------------------------------------1g
FeSO4·7H2O-----------------------------------0,1g
CoCl2·6H2O------------------------------------0,1g
CaCl2---------------------------------------------0,1g
ZnSO4·7H2O-----------------------------------0,1g
CuSO4·5H2O---------------------------------0,01g
AlK(SO4)2·12H2O---------------------------0,01g
H3BO3------------------------------------------0,01g
Na2MoO4·2H2O-----------------------------0,01g
Eaudistillée-------------------------------1000ml
Suiteàl’ajoutdesminérauxlemilieuaétéajustéàunpHde7,3etautoclavéà121˚C
pendantaumoins15minutes.Avantutilisation,etseulementlorsquelasolutionest
àtempératuredelapièce,l’ajoutdevitamineB12etdeméthanolaétéeffectué.Pour
100mldemilieu,0,3mldeméthanolstériliséparfiltrationet0,1mldevitamineB12
(0,1mg/ml)(stérilisédelamêmefaçon)ontétéajoutés.

21
3.2.2. MilieuoptimaldelasoucheNL23
PourlasoucheNL23,lemilieudecroissanceoptimalestuneversion
modifiéedumilieuATCC337a,dontl’efficacitéaététestéelorsd’études
précédentes(Martineauetal.,2013).Danschaquelitredumilieuonretrouve:
KH2PO4------------------------------------------1,3g
Na2HPO4--------------------------------------1,13g
(NH4)2SO4---------------------------------------0,5g
MgSO4SO4·7H2O------------------------------0,2g
Solutiond’élémentstraces------------------5ml
Eaudistillée-------------------------------1000ml
Lasolutiond’élémentstracesestspécifiquementadaptéeaumilieu337a;unlitrede
lasolutionestcomposéde:
CaCl2·2H2O--------------------------------0,618g
FeSO4·7H2O-----------------------------------0,4g
Na2MoO4·H2O---------------------------------0,2g
MnSO4·4H2O------------------------------0,176g
Eaudistillé---------------------------------1000ml
Lasolutiond’élémentstracespouvaitcontenirunecertainequantitédeprécipité
suivantuncycleàl’autoclave,maiscecin’apasproduitd’effetsnotablessurla
croissancedesbactéries.Suiteàlacombinaisondesingrédients,lasolutionaété
ajustéeàunpHentre7,5et8etautoclavéeà121˚Cpendantaumoins15minutes.
DelavitamineB12(0,1mlà0,1mg/ml)etduméthanol(0,3ml)ontétéajoutés
avantl’utilisation.

22
3.2.3. Milieudecroissanceadaptéàlaco-culture
Avantledébutduprojet,ilétaitdéjàconnuquelessouchesNL23etJAM1,
ainsiqu’uneco-culturedesdeux,pouvaientsimplementcroitredansuneversion
diluée(15g/Laulieude30g/L)dumilieumarinartificiel«InstantOcean»
retrouvéauBiodôme.Lacompositionexacted’unlitredumilieu«InstantOcean»
pourco-cultureest:
Mélange«InstantOcean»------------------15g
NaNO3---------------------------------------1,214g
KH2PO4(ensolution32,67g/L)----------2,5ml
Solutiond’élémentstraces------------------1ml
Eaudistillée-------------------------------1000ml
Lemélanged’eausaléepouraquarium«InstantOcean»estacheté
commercialementetsacompositionexacteestdoncinconnue(«InstantOcean»est
lenomdufournisseur).
Lasolutiond’élémentstracesestspécifiqueaumilieu«InstantOcean»;unlitrede
lasolutionestcomposéde:
FeSO4·7H2O-----------------------------------0,9g
CuSO4·5H2O-------------------------------0,035g
MnSO4·H2O--------------------------------0,202g
Eaudistillée-------------------------------1000ml
Unefoisquelemilieuestcomplet,lepHaétéajustéentre7,8et8etautoclavéà
121˚Cpendantaumoins15minutes.Avantl’utilisation,0,1mldevitamineB12(0,1
mg/ml)et0,3mldeméthanolontétéajoutés.Laquantitédenitrateajoutéeau
milieuvarieselonlesbesoinsdel’expérienceetlesconnaissancesacquiseslorsdela
recherche.Cesdifférencessontindiquéesoùelless’appliquent.

23
3.2.4. Milieuxsousformedegéloses
Chaquevariétédemilieudécriteci-hautaétéprincipalementutiliséesous
formeliquidepoursoutenirdespopulationsdebactériesensuspensionoupour
permettrelaformationdebiofilmsursupportsadaptés.Cependant,cesmilieuxont
aussiétéutilessousformedegélosesenboîtesdePetri.Pourproduiredesgéloses,
lasolutionminérale(enconcentrationdouble)etlasolutiond’agar(3%)ontété
autoclavéesséparément,puiscombinéesavantd’êtrecouléesdanslesboîtesde
Petri.LavitamineB12etleméthanolontétéajoutésaprèsl’autoclavage.
3.3. Préparationdescultures
PuisquelessouchesJAM1etNL23ontdestempsdecroissancedifférents,la
culturedesbactériesadûsuivreunrégimerigoureux.Uneréserveàlongtermede
chaquesouchebactérienneaétéconservéeà-80˚C.Àpartirdecelle-ci,descolonies
distinctesontétéproduitessurgéloses;lacroissancesedéroulaitsurunepériode
d’environ10joursà30˚C,puiscesculturesétaientconservéesà4˚Cpourune
périoded’utilisationd’environunmois.Àpartirdecoloniesdistinctes,
préférablement,despetitsvolumes(environ15ml)decultureliquideontété
préparés,cequinécessitede2à3joursd’incubation.Cesculturesontàleurtour
étéconservéesà4˚Cpourunepérioded’utilisationde1-2semaines.Également,ces
culturesontétéutiliséespourcréerlesculturesenplusgrosvolumes(variables
selonlesbesoinsdel’expérience)quiservirontd’inoculumpourlesexpériences
elles-mêmes.Parexemple,ajouter0,6mldecultureliquideà10mldemilieu
optimalfraisapermislacroissancejusqu’àdensitémaximaledelasoucheJAM1en
24hetdelasoucheNL23en40h.Danslescasoùl’expérienceenvisagéeimplique
uneco-culture,lestempsd’ensemencementontétéplanifiéspourqueladensité
maximalesoitatteinteenmêmetempspourlesdeuxsouchesaudébutdeladite
expérience.

24
3.3.1. Ensemencement
Àl’ensemencement,ladensitéoptiquedesculturesbactériennesservant
d’inoculumaétémesuréeàunelongueurd’ondede600nm(DO600).Decette
façonlaconcentrationdelabiomasseapuêtreajustéeàunevaleurquiétait
toujourslamêmepourchaqueexpérience;danscecasleDO600viséétaitde0,1
danslacultureexpérimentale.Donc,danslecasd’uneco-culture,chaquesouche
individuelledevaitcontribuerunDO600de0,1danslaco-cultureexpérimentaleau
tempsinitial.Ceciapermisunecertaineconstancedansleprocessusexpérimental
quiaévitéquelesconcentrationsinitialesdebactériesaffectentlerésultatd’une
expérience.Deplus,danslescasoùunchangementdemilieuaeulieuaumoment
del’ensemencement(parexemple,uneco-cultureoùchaquebactériepassed’un
milieuoptimalàunmilieuaccommodantlesdeuxsouches),lesculturesde
l’inoculumontdûd’abordêtrecentrifugéespourendébarrasserlemilieuinitial
(8000rpm,10minutes,25˚C,deuxfois).
3.3.2. Conditionsdeculture
Lesbactériesontétécultivéesenconditionsaérobiesouanaérobies.Les
culturesaérobiesontétéfaitesenflaconsErlenmeyerouenplaquesmulti-puits,
selonlebesoin,etontrequisdel’agitationconstante(entre60et100rpm)pour
faciliterlacirculationd’oxygènedanslemilieu.Lesculturesanaérobiesontété
effectuéesdansdesfiolesdeborosilicatedanslesquellesl’oxygèneaétéévacuéà
l’aided’azotegazeux(Fig3.2).Danscesfioles,l’échantillonnageaétérendupossible
grâceàunbouchonenbutyledanslesquelspeuventêtreinsérésdesaiguilles.Dans
touslescas,lesculturesontétéincubéesàunetempératureconstantede30˚C.

25
Figure3.2:Exempledecultureenanaérobiose
Unefioleferméehermétiquementpourculturedebactériesenconditionsanaérobies.Uneculturecommecelle-cicontienttypiquement30mldemilieuliquideetestincubéeà30˚Cavecseringuedanslebouchonenbutyle.
3.4. Récoltededonnées
Pourprendreunéchantillon,lacultureaétéagitéepourassurerune
distributionégaledescellulesetdoncdelaconcentrationdansleliquide.En
anaérobie,uneseringueaétéinséréedanslebouchondebutylescellantlafiole,et
aumoins1mlaétéextrait.UnefoisquelamesuredeDO600aétépriseavecle
spectrophotomètre,l’échantillonaétéconservéà-20˚Cdanslebutd’effectuer
d’autrestests(principalementdelachromatographieionique).

26
3.4.1. Spectrophotométrie
Pourrécolterdesdonnéesutilessurcescultures,dessuivisdecroissances
ontétéeffectuéesavecplusieurscombinaisonsdeconditions.Enconditions
aérobies,lesuiviaétéeffectuéuniquementparmesuresdeDO600àl’aided’un
spectrophotomètre,ouparmesuresdeturbiditéàl’aided’unappareilautomatisé
d’analysedecroissance(«Bioscreen»).L’avantageprincipald’unappareil
automatiséaétélapossibilitédeprendredesmesuressimultanémentpourchaque
échantillondefaçonrapide,cequiapermisderécolterdesdonnéesàdesintervalles
detempsplusrapprochées.Enconditionsanaérobies,lesuiviaégalementété
effectuéparmesuresdeDO600(manuellesseulement).
3.4.2. Chromatographieionique
Lachromatographieioniqueaétéutiliséepourquantifierlaconsommation
denitrateetdenitritedanslemilieuparlesdeuxsouchesbactériennes.Les
échantillonsontétépréalablementcentrifugés(13000rpm,2min.)dansdes
microtubesdetypeEppendorfpourenretirerlesbactéries.Unefoisquelescellules
seretrouvaientaufonddutube,aumoins0,5mldusurnageantdechaque
échantillonaétéprélevépourêtreanalyséparchromatographieionique.
L’appareilutilisépourlaquantificationdunitrateetdunitriteétaitle850
ProfessionalICdeMetrohm,munid’unecolonneMetroseppAsupp5dedimensions
250X4mm.Lacolonneaétémaintenueà35˚Cdurantl’opération.Lesmesuresde
nitrateetdenitriteontétéeffectuéesparUV.L’éluantutiliséétaitcomposéde3.2
mmol/LdeNa2CO3etde1mmol/LdeNaHCO3.Ledébitétaitde0,7ml/min.
3.4.3. Extractiond’ADN
Labiomassed’intérêt(ensuspensionliquide)ad’abordétécentrifugéeafin
d’enretirerlemilieudeculture.Lescellulesrésultantesontensuiteétédispersées
dansunesolutiontamponTEN(10mMTris-Cl;1mMEDTA;1MNaCl)dansun
microtubedetypeEppendorfde1,5mLcontenant0,2mgdebillesdeverresde0,1

27
mm.Cemélangeasubséquemmentétéplacédansunhomogénéisateurpour
effectuerunbroyage:rapidementagiterlescellulesensuspensionpourqu’elles
éclatentcontrelesbillesdeverre.L’appareilutiliséétaitleFastPrep-24deMPBio.
L’ADNaétéextraitàl’aided’unmélangedephénol,dechloroformeetd’alcool
isoamylique(25:24:1).L’ADNapuensuiteêtreprécipitédansunemixture(9:1)
d’éthanolfroid(90%)etd’acétatedesodium.Letoutaétécentrifugé,etleprécipité
aétédissousdansl’eau.L’ADNaétéquantifiéparspectrophotométrie,cequia
égalementdonnéuneindicationdelapuretédelasolutionobtenue.Le
spectrophotomètreutilisélorsdecesétudesétaitleNanodrop2000deThermo
Scientific.
3.4.4. qPCR
Danslecadredecesétudes,l’applicationprincipalementintéressantedela
techniqueestlaquantificationdecopiesdegènesparticuliersprésentsdansuneco-
culture.Puisquelessouchesbactériennesétudiéesontchacuneleurpropreversion
uniquedelanitrateréductasedontlaséquencegénétiqueestconnue,ilaété
possibled’utiliserleqPCRpourmesurerlaquantitérelativedesbactériesdansle
mélange.Lesnitrateréductasesenquestionsontlanitrateréductasepériplasmique
delasoucheNL23(gènenapA)etunedesdeuxnitrateréductasesmembranairesde
lasoucheJAM1(gènenarG1).Donc,parexemple,siuneanalyseqPCRindiqueque
legènenarG1représente90%descopiesdegènesdenitrateréductase(narG1+
napA)dansl’échantillon,celaindiquequelasoucheJAM1comptaitneuffoisplusde
cellulesquelasoucheNL23danslaco-culture.
LatechniquedeqPCRutiliséelorsdecesétudesétaitleTaqMan.Danscette
variationduqPCR,deuxcourtesséquencesdenucléotidesserventd’amorces,tandis
qu’uneautrecourteséquencesertdesonde.Cettedernièreestconçuepour
s’apparieràunendroitentrelesdeuxamorces.Àlasondesontfixésun
fluorochromeetun«quencher».Lafluorescenceémiseparlefluorochromeest
dissimuléeparle«quencher»lorsquelesdeuxsontàproximité.Lapolymérase
Taqsemetalorsàajouterdesnucléotidesauxamorces,etquandellearriveàla

28
sonde,celle-ciestenlevéeestdégradée,libérantlefluorochromequiémetalorsune
fluorescencequantifiable.LeTaqManaétéutilisédansceprojetàcausedesoncoût
abordablecomparéauSYBRgreen,leprotocolequiétaitutiliséprécédemmentdans
lelaboratoire.Ilaaussiéténotéquedesproblèmesdedimèresd’amorcessont
aussiplusfréquentsavecleSYBRgreen.D’ailleurs,ilaétédéterminéqueleTaqMan
génèredesrésultatscomparablesauSYBRgreen(Blais,2014).
L’instrumentutilisépoureffectuerlesqPCRétaitleRotor-Gene6000de
CorbettLifeSciences(Qiagen).Chaquemicrotube(tubesspécialisés0,1mlavec
couverclesdeQiagen)contenait25µld’unmélangepréparéeenlaboratoire(voir
tableau3.1).L’ADNajoutéaumélangeaétéobtenugrâceàlaméthoded’extraction
décritedanslasection3.4.3.LeprogrammePCRétait:
i) 5minutesà95˚C
ii) 40cyclesdesétapessuivantes:
(1) 15secondesà95˚C
(2) 20secondesà55˚C
(3) 30secondesà65˚C
Tableau3.1:compositiondumélangedanschaqueréactiondeqPCR
Composante Volumeparréaction
H2O 14,88µl
Tampon10Xtaq«hotstart» 2,5µl
BSA(20µg/µl) 0,5µl
dNTP(10mM) 0,5µl
Amorce(f)(10mM) 0,5µl
Amorce(r)(10mM) 0,5µl
SondeTaqMan(5mM) 0,5µl
Taqpolymérase«hotstart»(5U/µl) 0,125µl
ADNgénomique(dilué1/5) 5µl
29
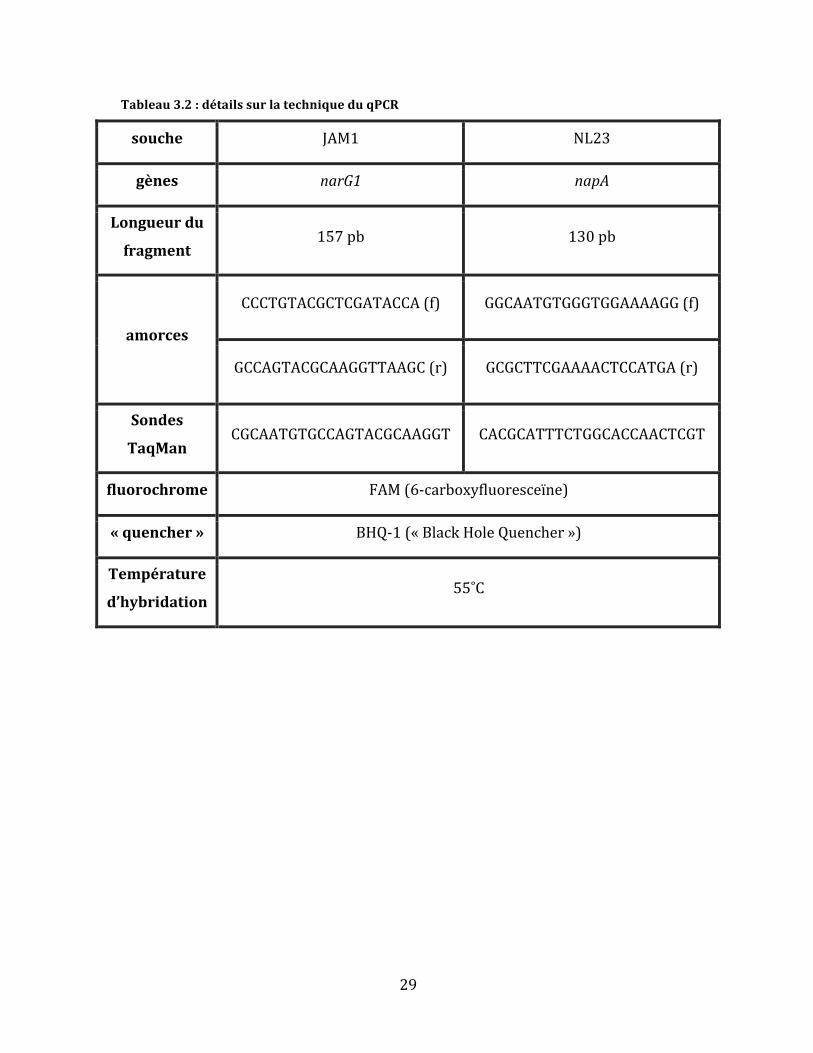
Tableau3.2:détailssurlatechniqueduqPCR
souche JAM1 NL23
gènes narG1 napA
Longueurdu
fragment157pb 130pb
amorces
CCCTGTACGCTCGATACCA(f) GGCAATGTGGGTGGAAAAGG(f)
GCCAGTACGCAAGGTTAAGC(r) GCGCTTCGAAAACTCCATGA(r)
Sondes
TaqManCGCAATGTGCCAGTACGCAAGGT CACGCATTTCTGGCACCAACTCGT
fluorochrome FAM(6-carboxyfluoresceïne)
«quencher» BHQ-1(«BlackHoleQuencher»)
Température
d’hybridation55˚C

30
4. Résultats4.1. Déterminationdesconditionsoptimalesdeculture
Lepremiervoletduprojetétaitconstituéd’expériencesvisantàdéterminer
quellesconditionsdeculturepermettraientlameilleurecroissancedessouches
JAM1etNL23.Unegrandepartiedutravailàcetégardétaitdéjàcomplétépardes
étudespréliminaires.Eneffet,laplupartdesparamètresdebase,incluantla
température,lepH,lasalinité,etlemilieudecroissanceavaientétéétablisavantle
débutdestravauxprésentésdanscemémoire.Lesconditionsoptimalesqu’ilrestait
àdéterminerimpliquaientprincipalementlesconcentrationsdenitrateetles
proportionsdesbactériesenco-cultures.
4.1.1. Suividecroissancepréliminaireenconditionsaérobies
Cespremierssuivisdecroissanceavaientpourbutdevaliderlechoixde
milieudecroissancepourlesco-culturesdessouchesJAM1etNL23ainsiquede
donnerunaperçudescourbesdecroissancetypiquesdecelles-ci.LessouchesJAM1
etNL23ontétécultivéesenconditionsaérobiesouanaérobies,soitsousformede
culturespuresouenco-cultures.Lemilieucommerciald’eaudemer«Instant
Ocean»aétéutilisépourcespremièresexpériencesenprésencedeprèsde40mM
d’azotesousformedenitrate.Ilaétédéterminéauparavantquelemilieu«Instant
Ocean»coupédemoitié(15g/L,aulieude30g/L)permettaitlacroissanceàlafois
dessouchesJAM1etNL23,lasoucheNL23étanttrèssensibleàlaconcentrationen
sel.
31
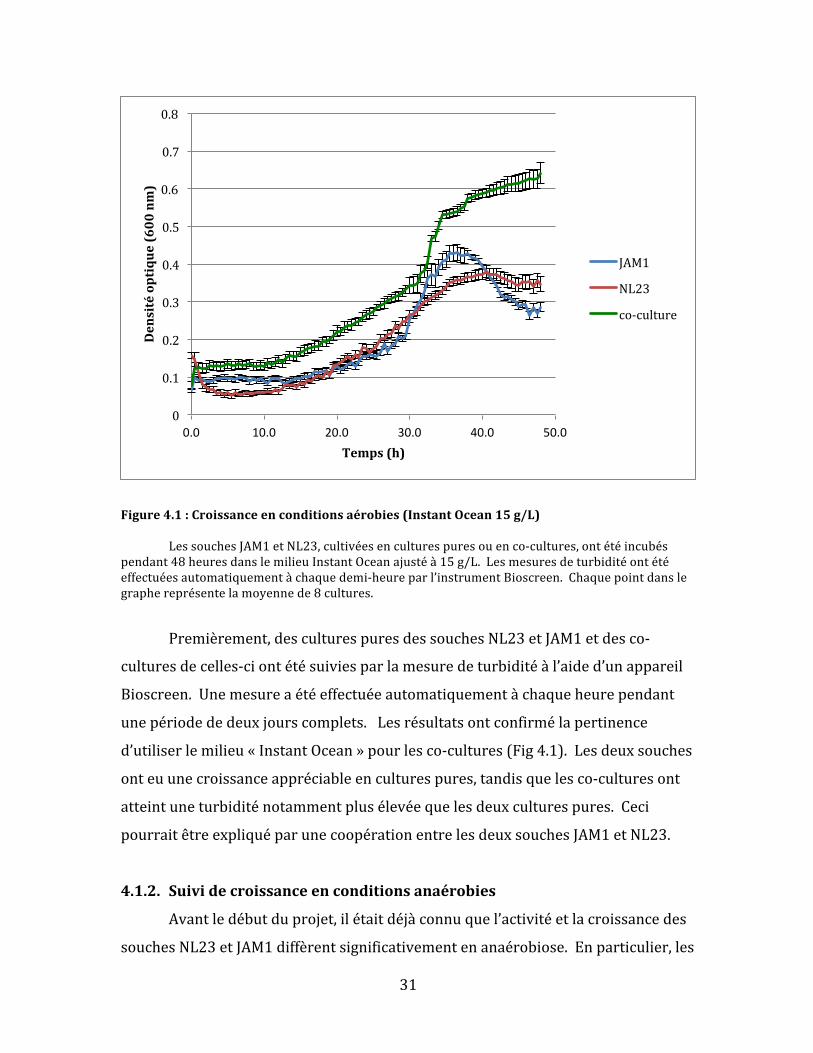
Figure4.1:Croissanceenconditionsaérobies(InstantOcean15g/L) LessouchesJAM1etNL23,cultivéesenculturespuresouenco-cultures,ontétéincubéspendant48heuresdanslemilieuInstantOceanajustéà15g/L.Lesmesuresdeturbiditéontétéeffectuéesautomatiquementàchaquedemi-heureparl’instrumentBioscreen.Chaquepointdanslegraphereprésentelamoyennede8cultures.
Premièrement,desculturespuresdessouchesNL23etJAM1etdesco-
culturesdecelles-ciontétésuiviesparlamesuredeturbiditéàl’aided’unappareil
Bioscreen.Unemesureaétéeffectuéeautomatiquementàchaqueheurependant
unepériodededeuxjourscomplets.Lesrésultatsontconfirmélapertinence
d’utiliserlemilieu«InstantOcean»pourlesco-cultures(Fig4.1).Lesdeuxsouches
onteuunecroissanceappréciableenculturespures,tandisquelesco-culturesont
atteintuneturbiditénotammentplusélevéequelesdeuxculturespures.Ceci
pourraitêtreexpliquéparunecoopérationentrelesdeuxsouchesJAM1etNL23.
4.1.2. Suividecroissanceenconditionsanaérobies
Avantledébutduprojet,ilétaitdéjàconnuquel’activitéetlacroissancedes
souchesNL23etJAM1diffèrentsignificativementenanaérobiose.Enparticulier,les
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
JAM1
NL23
co-culture

32
nitrateréductasesdelasoucheJAM1sontbeaucoupplusefficaces,etcelle-ci
démontreunecroissanceinitialeplusélevée(Martineauetal.,2015,Mauffreyetal.,
2015).Lespremierssuivisdecroissanceenconditionsanaérobiesontdoncétéfaits
avecdifférentsratiosd’inoculumpourdéterminerlesconditionsd’ensemencement
lesplusfavorables.LessouchesNL23etJAM1requièrentlaprésencedenitrateet
sesdérivéescommesourced’accepteursd’électronspourcroîtreenanaérobiose.
Dansleursmilieuxoptimauxrespectifs,unequantitéinitialedenitrateoptimaleest
d’environ40mM(CélineVilleneuve,2012)(donnéesnon-publiéespourlasouche
NL23),donccetteconcentrationinitialeaétéchoisie.
Lessuivisdecroissanceontétéeffectuéssurunesériedeco-culturesavec
desvariationsdansleratioentrelesdeuxsouchesbactériennes.L’expérience,qui
comptaitégalementcommetémoinslessouchesNL23etJAM1enculturespures,a
étéeffectuéeavecdesco-culturesderatios(JAM1:NL23)1:1,1:2,1:5,et1:10.
Cesratiosréfèrentuniquementauxquantitésrelativesdessouchesaumomentde
l’ensemencement,telsquemesurésparDO600.Lacroissanceétantbeaucoupplus
lenteencesconditions,etàcausedeconsidérationspratiques,desmesuresde
DO600ontétéprisesmanuellementunefoisparjourjusqu’àcomplétiondusuivi.
L’appareil«Bioscreen»nepermetpaslesculturesenconditionsanaérobies.Les
échantillonsainsirecueillisontégalementservipourlaquantificationdenitrateet
denitrite.Leséchantillonnagesjournaliersontétéeffectuéssurunepériodedeprès
de200heures,aprèsquoilesculturesontétéconservéespourdesétudesparqPCR.
33
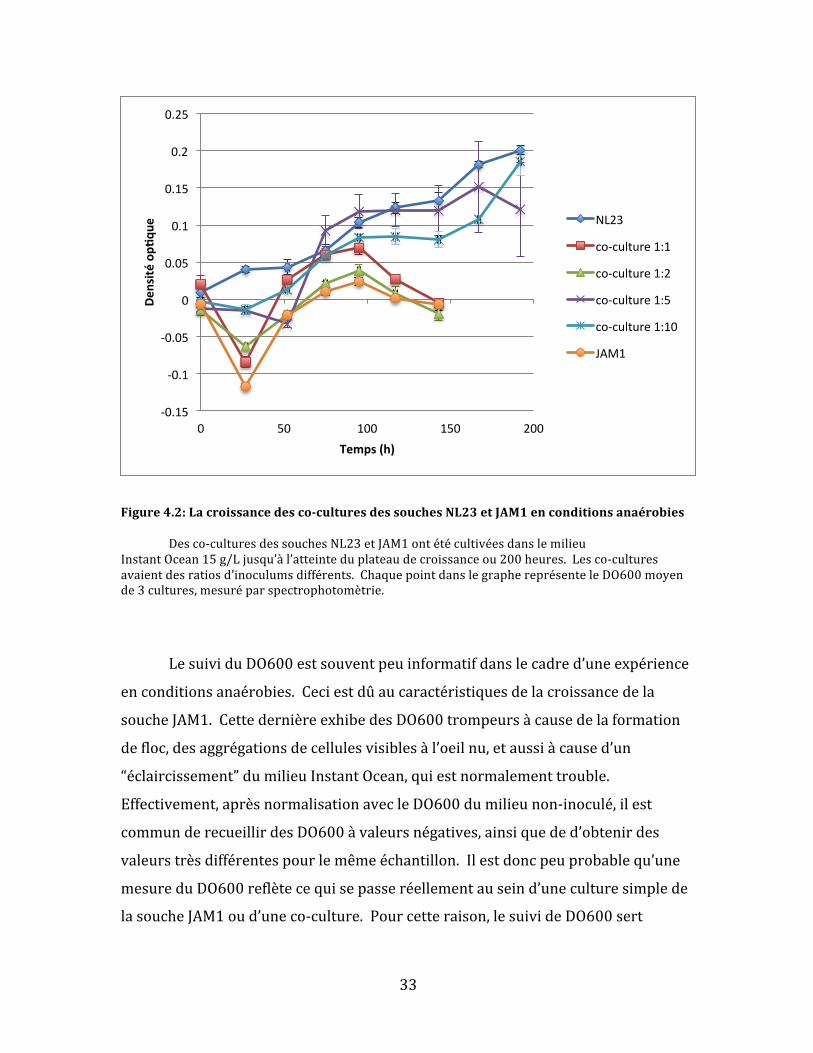
Figure4.2:Lacroissancedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.ChaquepointdanslegraphereprésenteleDO600moyende3cultures,mesuréparspectrophotomètrie.
LesuividuDO600estsouventpeuinformatifdanslecadred’uneexpérience
enconditionsanaérobies.Ceciestdûaucaractéristiquesdelacroissancedela
soucheJAM1.CettedernièreexhibedesDO600trompeursàcausedelaformation
defloc,desaggrégationsdecellulesvisiblesàl’oeilnu,etaussiàcaused’un
“éclaircissement”dumilieuInstantOcean,quiestnormalementtrouble.
Effectivement,aprèsnormalisationavecleDO600dumilieunon-inoculé,ilest
communderecueillirdesDO600àvaleursnégatives,ainsiqueded’obtenirdes
valeurstrèsdifférentespourlemêmeéchantillon.Ilestdoncpeuprobablequ’une
mesureduDO600reflètecequisepasseréellementauseind’uneculturesimplede
lasoucheJAM1oud’uneco-culture.Pourcetteraison,lesuivideDO600sert
-0.15
-0.1
-0.05
0
0.05
0.1
0.15
0.2
0.25
0 50 100 150 200
Densité
op+
que
Temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
JAM1

34
principalementdeméthodepermettantd’observerqualitativementetrapidementle
progrèsdelacroissancedesbactéries.
Danslecasdecetteexpérience,cependant,ilestintéressantdenoterqueles
co-culturesavecunratiopluségal(lesco-cultures1:1et1:2)ontsuiviunecourbe
decroissancequiressemblebeaucoupauxculturespuresdelasoucheJAM1,tandis
quelesautresco-culturesontsuiviunecourbedecroissancequiressembleàcelle
desculturespuresdelasoucheNL23(Fig4.2).Cecipourraitréfletersimplement
quelecomportementtypedelasoucheJAM1devientmoinsvisibledanslesdonnées
lorsquecettedernièreestmoinsabondantedanslesco-cultures.Toutefois,ilest
aussipossiblequ’unedensitéplusimportantedesdeuxsouchesontpuse
développerenco-cultureslorsquel’inoculumdedépartdesoucheJAM1étaitmoins
important.Cetteexplicationseraitcompatibleavecl’hypothèsedudépart.Pour
approfondirlacompréhensiondelasituation,ilafalluinvestiguerauniveaudela
consommationdunitrateetdunitrite(Fig4.3et4.4).
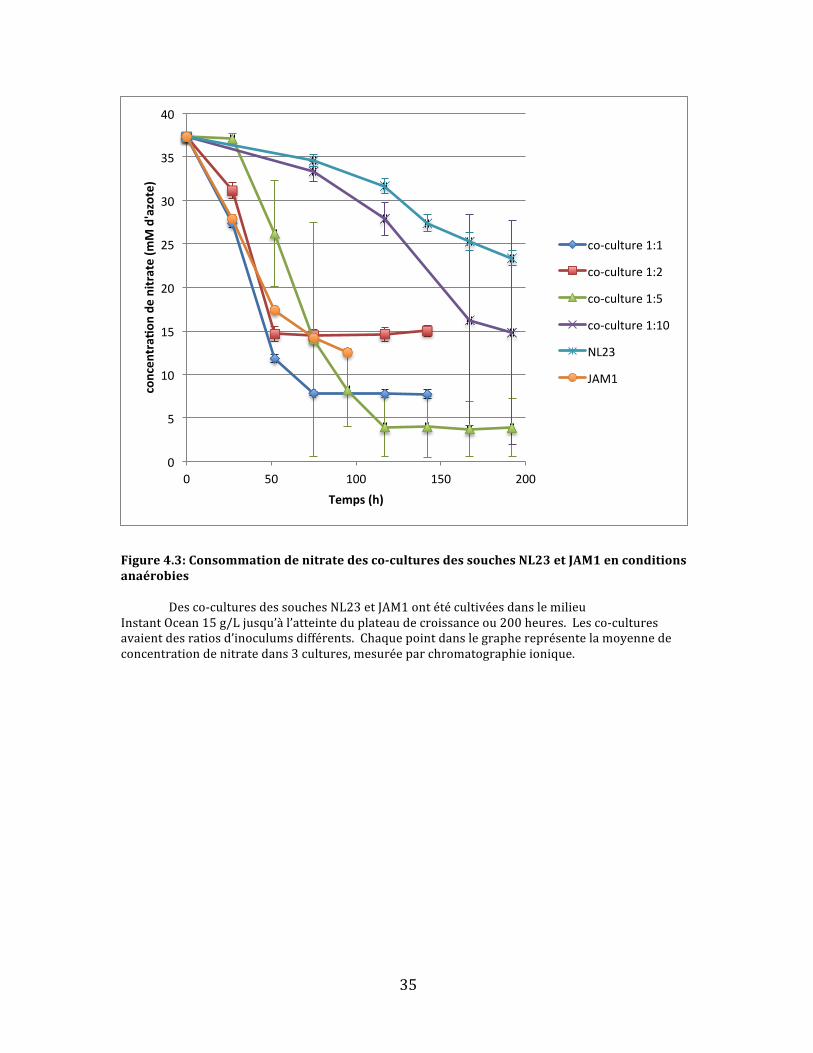
35
Figure4.3:Consommationdenitratedesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratedans3cultures,mesuréeparchromatographieionique.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concen
tra+
ondenitrate(m
Md'azote)
Temps(h)
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
NL23
JAM1
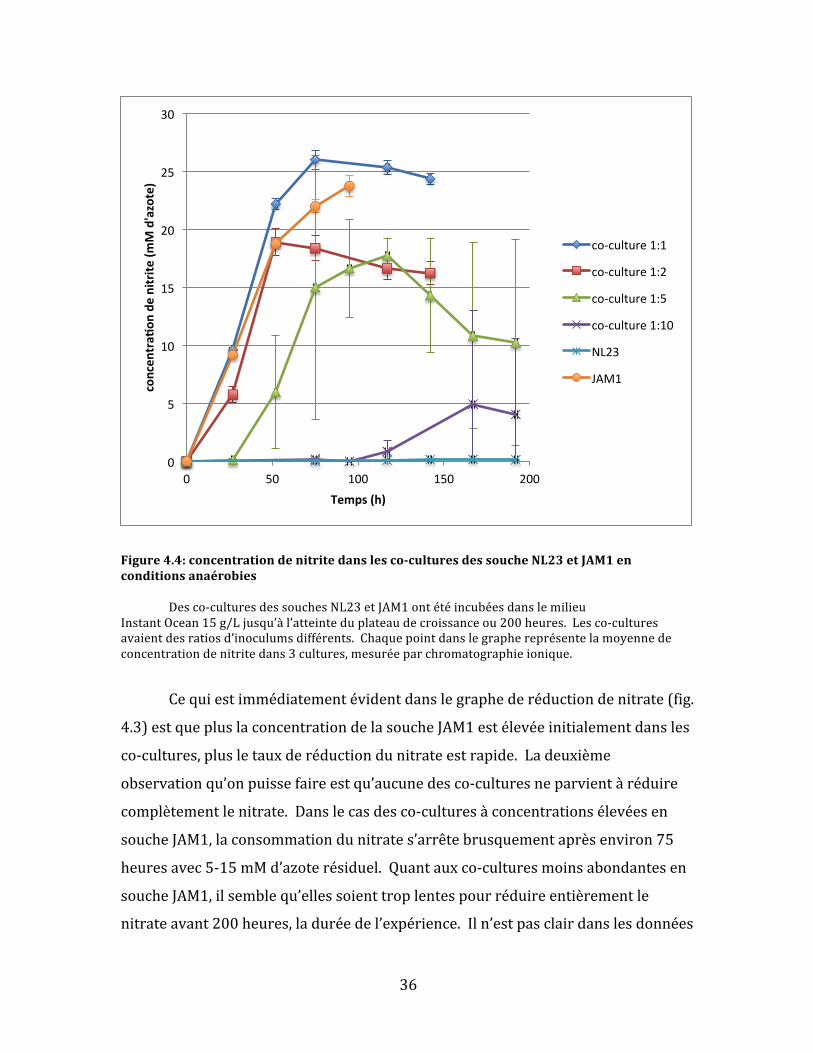
36
Figure4.4:concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétéincubéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritedans3cultures,mesuréeparchromatographieionique.
Cequiestimmédiatementévidentdanslegraphederéductiondenitrate(fig.
4.3)estquepluslaconcentrationdelasoucheJAM1estélevéeinitialementdansles
co-cultures,plusletauxderéductiondunitrateestrapide.Ladeuxième
observationqu’onpuissefaireestqu’aucunedesco-culturesneparvientàréduire
complètementlenitrate.Danslecasdesco-culturesàconcentrationsélevéesen
soucheJAM1,laconsommationdunitrates’arrêtebrusquementaprèsenviron75
heuresavec5-15mMd’azoterésiduel.Quantauxco-culturesmoinsabondantesen
soucheJAM1,ilsemblequ’ellessoienttroplentespourréduireentièrementle
nitrateavant200heures,laduréedel’expérience.Iln’estpasclairdanslesdonnées
0
5
10
15
20
25
30
0 50 100 150 200
concen
tra+
ondenitrite
(mMd'azote)
Temps(h)
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
NL23
JAM1

37
recueilliessilaréductionauraitpucontinuerjusqu’àépuisementdesréservesde
nitrate.
Lesuividelaconsommationdunitritecorroborel’hypothèse:Lesco-
cultures1:1et1:2ontaccumulélenitriterapidementpendant50-100heures
avantdeplafonner(entre18et26mMd’azote)indéfiniment.Lesco-cultures1:5
ontatteintunniveaudenitritemaximum(environ17mMd’azote)àenviron120
heurespourredescendreverslafindusuivi.Finalement,lesco-cultures1:10n’ont
démontréqu’unepetiteaccumulationdenitritetarddansl’expérience(170heures).
Lestempsoùlespointsmaximauxd’accumulationdenitritesontatteintssemblent
correspondreàpeuprèsàceuxd’arrêtd’activitéderéductiondenitrate.
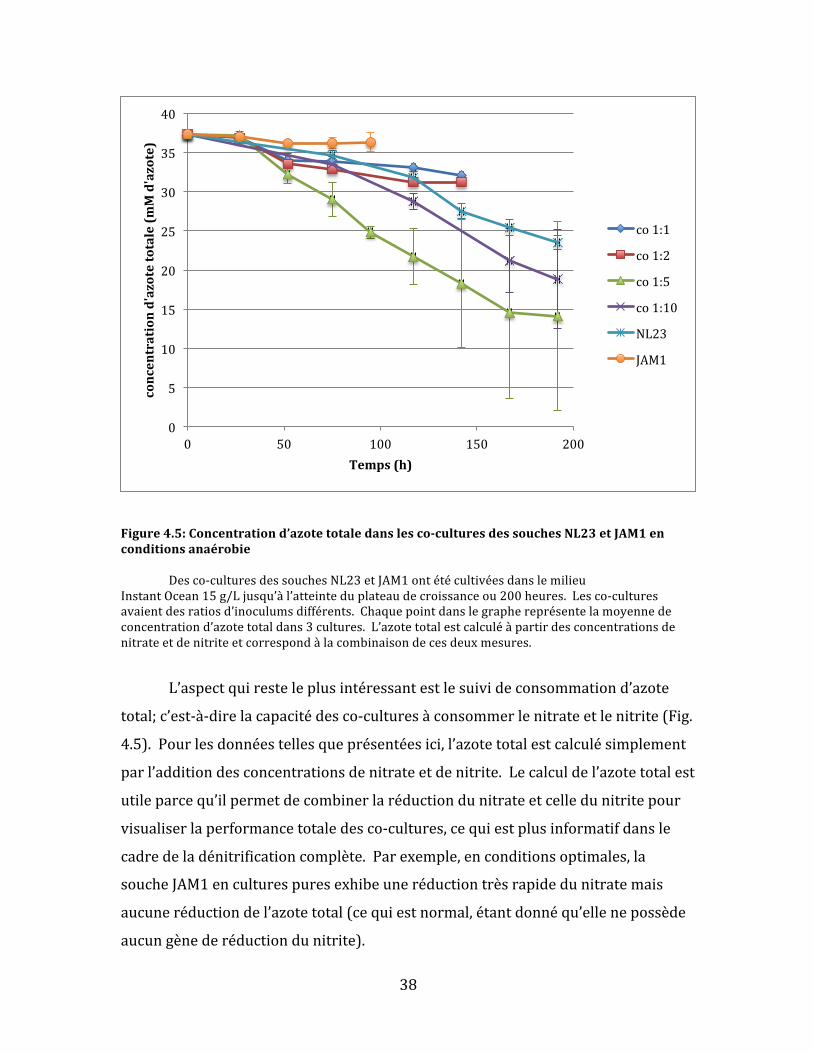
38
Figure4.5:Concentrationd’azotetotaledanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobie Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Ljusqu’àl’atteinteduplateaudecroissanceou200heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotaldans3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
L’aspectquiresteleplusintéressantestlesuivideconsommationd’azote
total;c’est-à-direlacapacitédesco-culturesàconsommerlenitrateetlenitrite(Fig.
4.5).Pourlesdonnéestellesqueprésentéesici,l’azotetotalestcalculésimplement
parl’additiondesconcentrationsdenitrateetdenitrite.Lecalculdel’azotetotalest
utileparcequ’ilpermetdecombinerlaréductiondunitrateetcelledunitritepour
visualiserlaperformancetotaledesco-cultures,cequiestplusinformatifdansle
cadredeladénitrificationcomplète.Parexemple,enconditionsoptimales,la
soucheJAM1enculturespuresexhibeuneréductiontrèsrapidedunitratemais
aucuneréductiondel’azotetotal(cequiestnormal,étantdonnéqu’ellenepossède
aucungènederéductiondunitrite).
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotale(m
Md'azote)
Temps(h)
co1:1
co1:2
co1:5
co1:10
NL23
JAM1

39
Pourlaprésenteexpérience,lesco-cultures1:5ontdémontrélaplusgrande
consomptiond’azote,suividesco-cultures1:10.Ilfautnoterquecesdeuxco-
culturesontconsomméplusrapidementlenitrateetlenitritequelesculturespures
delasoucheNL23,cequipourraitêtreunindicedelacoopérationbénéfiquedes
deuxsouchesdanscesco-cultures.Parcontre,lesco-cultures1:1et1:2ontété
moinsefficacesquelasoucheNL23enculturespures.Aucunedesco-culturesn’a
démontréuneréductioncomplètedel’azotetotaleavant200heuresdesuivide
croissance.
Malgrélacroissanceréussiedesdeuxsouchesenculturespuresetenco-
cultures,cessuivisontsoulevéuneproblématiqueimportante:lesco-culturesne
sontjamaisarrivéesàréduirecomplètementlenitrateleurétantfourniaudépart,ni
l’azotetotal.Eneffet,lesco-culturesàratiosplusinégauxontsimplementdémontré
uneréductiontrèslentedunitrate.Lesco-culturesàratiospluségauxsesont
comportéesdefaçonpresqueidentiqueàlasoucheJAM1enculturepure:une
réductionrapide(maisincomplète)dunitratesuivid’unestagnationd’activité
dénitrifiante,incluantuneaccumulationdenitrite,ceciendépitdelaprésencedela
soucheNL23,quialacapacitéderéduirecenitrite.Uneexplicationprobableadonc
étéformulée:étantdonnéquelenitriteestparticulièrementtoxiqueaux
organismesvivants(DeRoosetal.,2003),sonaccumulationenimportante
concentrationinhiberaitlacroissancedelasoucheNL23.CommelasoucheJAM1
estbeaucoupplusrapidepourréduirelenitratequelasoucheNL23,ilestprobable
quecelle-ciaitétéresponsabled’uneaccumulationdenitritesupérieureàcequela
soucheNL23peutsupporter.Enconséquence,sil’abondancedunitritepouvaitêtre
limitée,lasoucheNL23pourraitavoirlachancederéduirecenitriteàmesurequ’il
soitproduit.
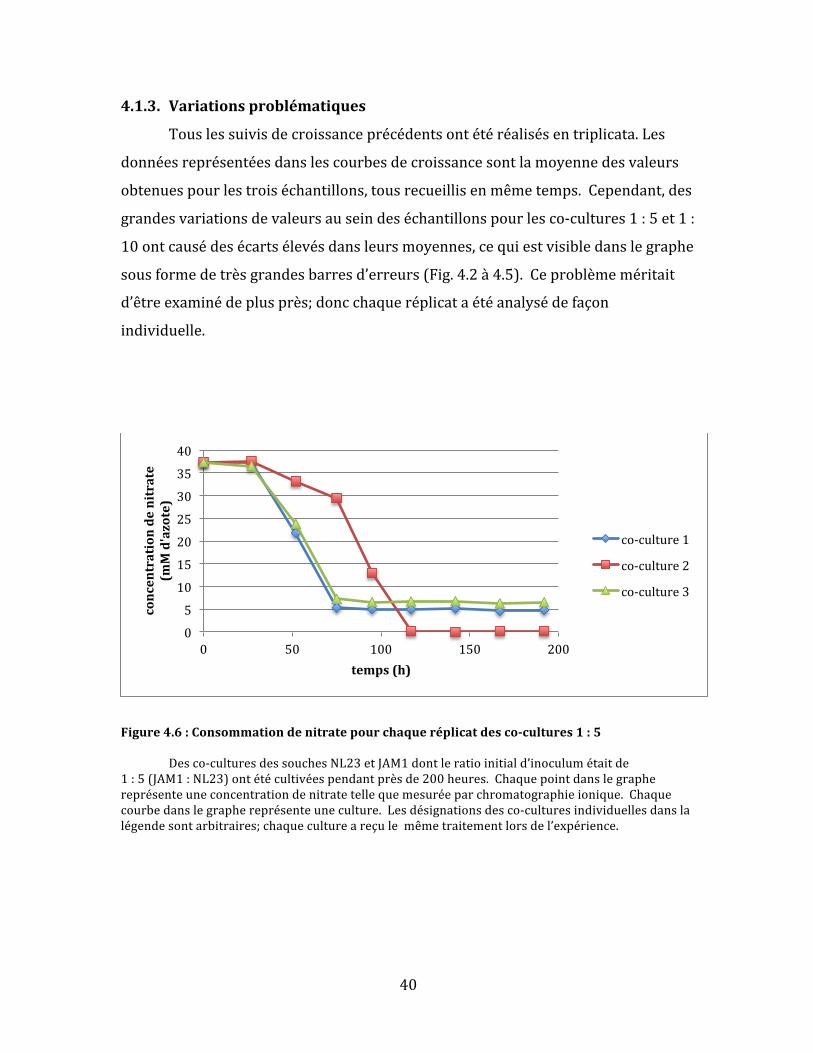
40
4.1.3. Variationsproblématiques
Touslessuivisdecroissanceprécédentsontétéréalisésentriplicata.Les
donnéesreprésentéesdanslescourbesdecroissancesontlamoyennedesvaleurs
obtenuespourlestroiséchantillons,tousrecueillisenmêmetemps.Cependant,des
grandesvariationsdevaleursauseindeséchantillonspourlesco-cultures1:5et1:
10ontcausédesécartsélevésdansleursmoyennes,cequiestvisibledanslegraphe
sousformedetrèsgrandesbarresd’erreurs(Fig.4.2à4.5).Ceproblèmeméritait
d’êtreexaminédeplusprès;doncchaqueréplicataétéanalysédefaçon
individuelle.
Figure4.6:Consommationdenitratepourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitratetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationdenitrate
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
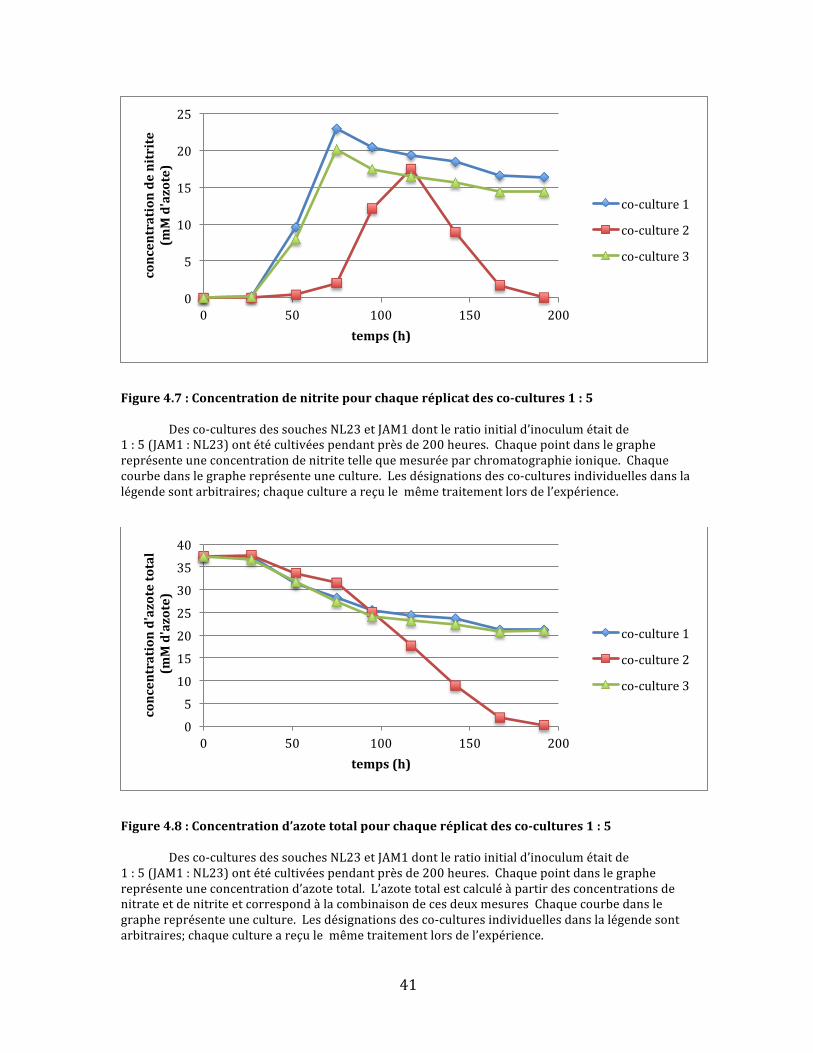
41
Figure4.7:Concentrationdenitritepourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitritetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
Figure4.8:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:5 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationd’azotetotal.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesuresChaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
0 50 100 150 200
concentrationdenitrite
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotal
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
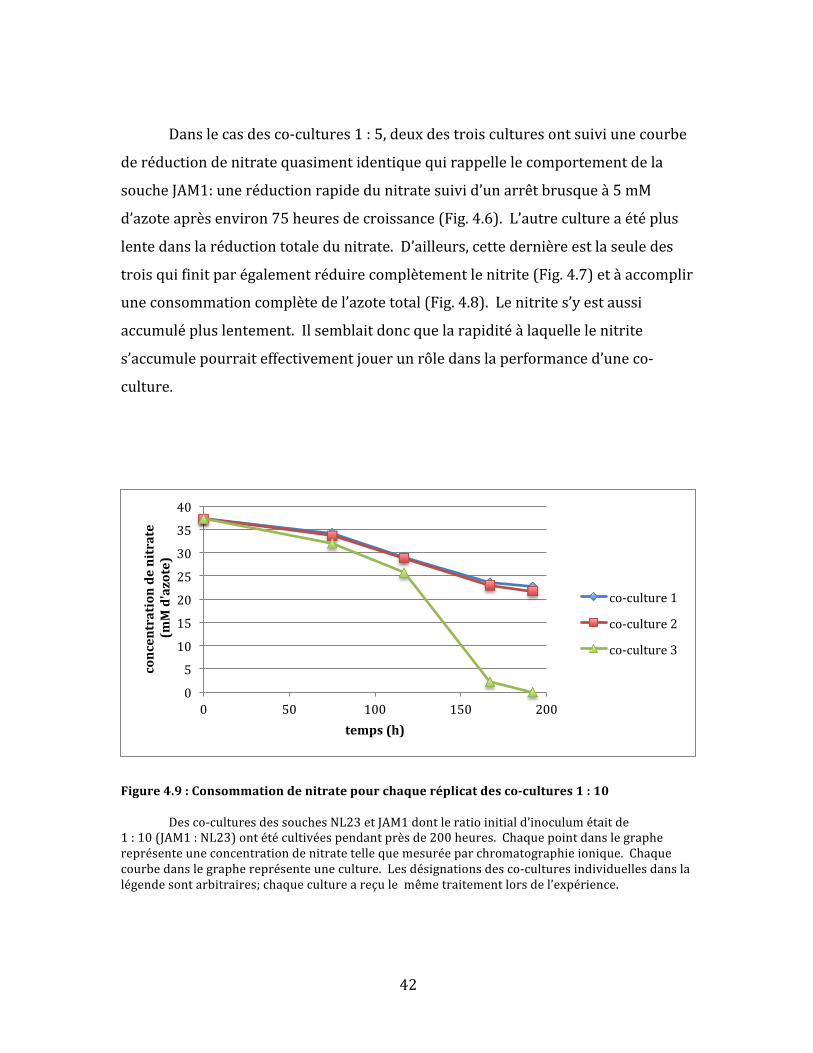
42
Danslecasdesco-cultures1:5,deuxdestroisculturesontsuiviunecourbe
deréductiondenitratequasimentidentiquequirappellelecomportementdela
soucheJAM1:uneréductionrapidedunitratesuivid’unarrêtbrusqueà5mM
d’azoteaprèsenviron75heuresdecroissance(Fig.4.6).L’autrecultureaétéplus
lentedanslaréductiontotaledunitrate.D’ailleurs,cettedernièreestlaseuledes
troisquifinitparégalementréduirecomplètementlenitrite(Fig.4.7)etàaccomplir
uneconsommationcomplètedel’azotetotal(Fig.4.8).Lenitrites’yestaussi
accumulépluslentement.Ilsemblaitdoncquelarapiditéàlaquellelenitrite
s’accumulepourraiteffectivementjouerunrôledanslaperformanced’uneco-
culture.
Figure4.9:Consommationdenitratepourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitratetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationdenitrate
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
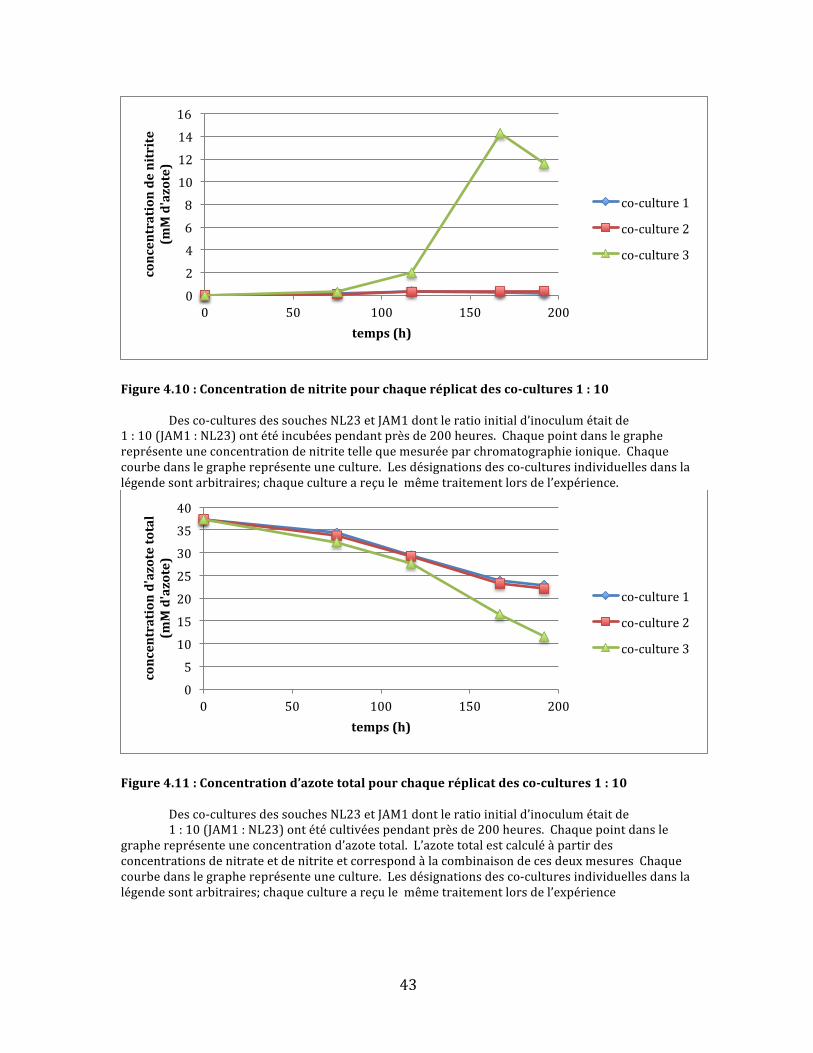
43
Figure4.10:Concentrationdenitritepourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétéincubéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationdenitritetellequemesuréeparchromatographieionique.Chaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience.
Figure4.11:Concentrationd’azotetotalpourchaqueréplicatdesco-cultures1:10 Desco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde
1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Chaquepointdanslegraphereprésenteuneconcentrationd’azotetotal.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesuresChaquecourbedanslegraphereprésenteuneculture.Lesdésignationsdesco-culturesindividuellesdanslalégendesontarbitraires;chaquecultureareçulemêmetraitementlorsdel’expérience
0
2
4
6
8
10
12
14
16
0 50 100 150 200
concentrationdenitrite
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
0
5
10
15
20
25
30
35
40
0 50 100 150 200
concentrationd'azotetotal
(mMd'azote)
temps(h)
co-culture1
co-culture2
co-culture3
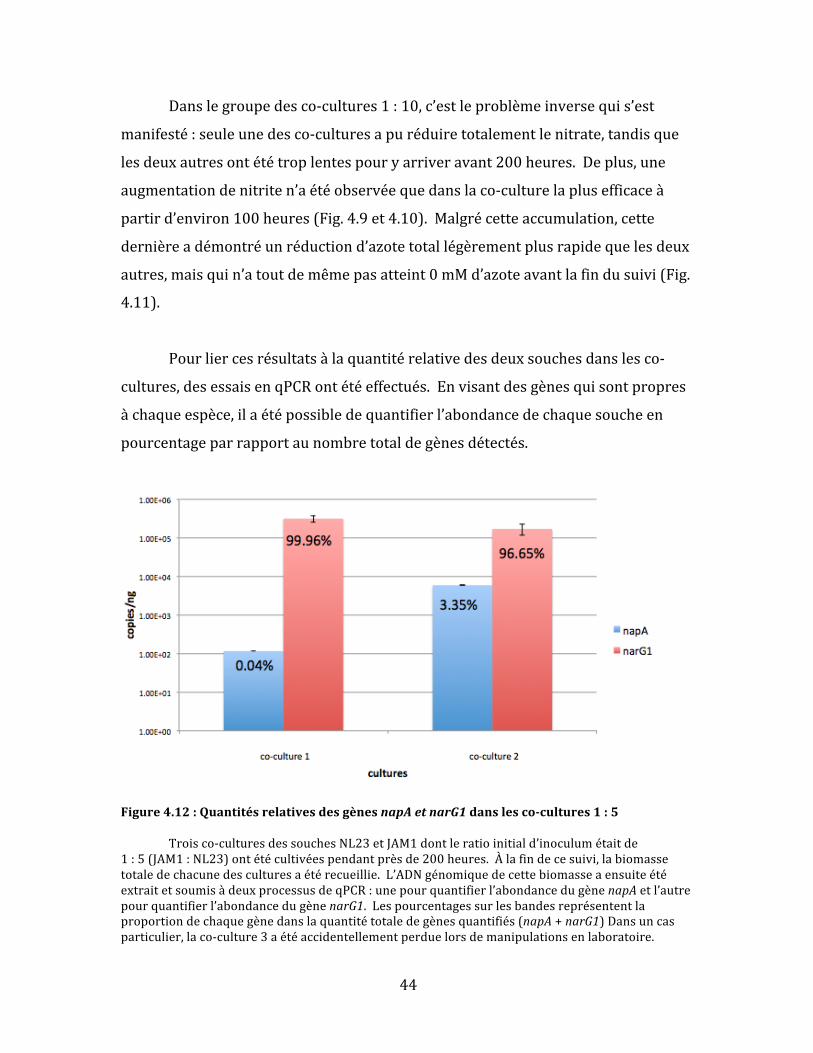
44
Danslegroupedesco-cultures1:10,c’estleproblèmeinversequis’est
manifesté:seuleunedesco-culturesapuréduiretotalementlenitrate,tandisque
lesdeuxautresontététroplentespouryarriveravant200heures.Deplus,une
augmentationdenitriten’aétéobservéequedanslaco-culturelaplusefficaceà
partird’environ100heures(Fig.4.9et4.10).Malgrécetteaccumulation,cette
dernièreadémontréunréductiond’azotetotallégèrementplusrapidequelesdeux
autres,maisquin’atoutdemêmepasatteint0mMd’azoteavantlafindusuivi(Fig.
4.11).
Pourliercesrésultatsàlaquantitérelativedesdeuxsouchesdanslesco-
cultures,desessaisenqPCRontétéeffectués.Envisantdesgènesquisontpropres
àchaqueespèce,ilaétépossibledequantifierl’abondancedechaquesoucheen
pourcentageparrapportaunombretotaldegènesdétectés.
Figure4.12:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:5 Troisco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:5(JAM1:NL23)ontétécultivéespendantprèsde200heures.Àlafindecesuivi,labiomassetotaledechacunedesculturesaétérecueillie.L’ADNgénomiquedecettebiomasseaensuiteétéextraitetsoumisàdeuxprocessusdeqPCR:unepourquantifierl’abondancedugènenapAetl’autrepourquantifierl’abondancedugènenarG1.Lespourcentagessurlesbandesreprésententlaproportiondechaquegènedanslaquantitétotaledegènesquantifiés(napA+narG1)Dansuncasparticulier,laco-culture3aétéaccidentellementperduelorsdemanipulationsenlaboratoire.
45
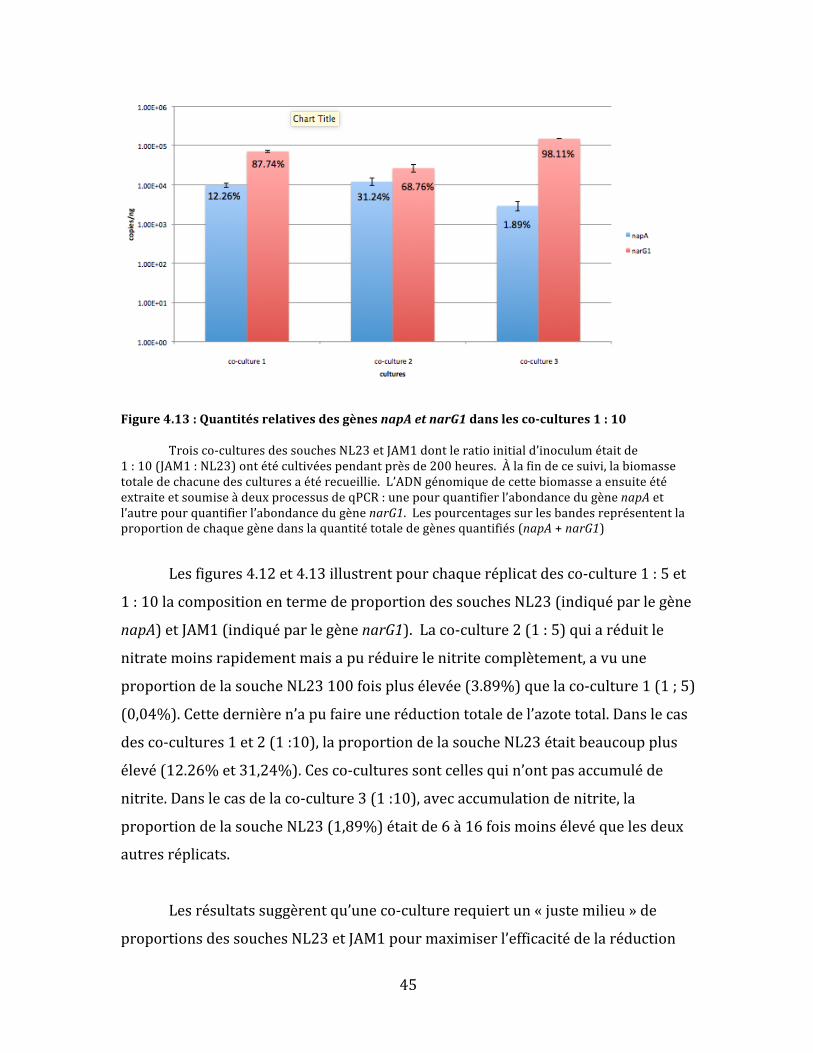
Figure4.13:QuantitésrelativesdesgènesnapAetnarG1danslesco-cultures1:10 Troisco-culturesdessouchesNL23etJAM1dontleratioinitiald’inoculumétaitde1:10(JAM1:NL23)ontétécultivéespendantprèsde200heures.Àlafindecesuivi,labiomassetotaledechacunedesculturesaétérecueillie.L’ADNgénomiquedecettebiomasseaensuiteétéextraiteetsoumiseàdeuxprocessusdeqPCR:unepourquantifierl’abondancedugènenapAetl’autrepourquantifierl’abondancedugènenarG1.Lespourcentagessurlesbandesreprésententlaproportiondechaquegènedanslaquantitétotaledegènesquantifiés(napA+narG1)
Lesfigures4.12et4.13illustrentpourchaqueréplicatdesco-culture1:5et
1:10lacompositionentermedeproportiondessouchesNL23(indiquéparlegène
napA)etJAM1(indiquéparlegènenarG1).Laco-culture2(1:5)quiaréduitle
nitratemoinsrapidementmaisapuréduirelenitritecomplètement,avuune
proportiondelasoucheNL23100foisplusélevée(3.89%)quelaco-culture1(1;5)
(0,04%).Cettedernièren’apufaireuneréductiontotaledel’azotetotal.Danslecas
desco-cultures1et2(1:10),laproportiondelasoucheNL23étaitbeaucoupplus
élevé(12.26%et31,24%).Cesco-culturessontcellesquin’ontpasaccumuléde
nitrite.Danslecasdelaco-culture3(1:10),avecaccumulationdenitrite,la
proportiondelasoucheNL23(1,89%)étaitde6à16foismoinsélevéquelesdeux
autresréplicats.
Lesrésultatssuggèrentqu’uneco-culturerequiertun«justemilieu»de
proportionsdessouchesNL23etJAM1pourmaximiserl’efficacitédelaréduction

46
dunitrateetdunitritetoutenévitantlatoxicitédunitrite.Malheureusement,il
s’estavérédifficiled’obtenirceparfaitmélangedefaçonassezconstantepour
produiredesrésultatsentriplicata,etlavariabilitéimprévisibledesrésultatsa
empêchéleprogrèsduprojetdanscettedirection.
4.1.4. Réductiondelaquantitéinitialedenitrate
Unenouvellestratégieaétéétabliepourcontournerleproblème
d’accumulationdenitrite.L’idéeétaitdediminuerlaconcentrationinitialede
nitratedanslemilieu«InstantOcean»pourréduirel’accumulationmaximalede
nitrite.Danslesessaisprécédents(Auclairetal.,2010),ilsemblaitêtrepossible
pourlesco-culturesdetoléreruncertainniveaudenitrite(jusqu’àenviron15mM
d’azote)avantd’éprouverdesdifficultés.L’hypothèseaétéémisequ’une
accumulationdenitritesousceniveauéviteraitleproblèmedetoxicité.
Pourvalidercettenouvelleapproche,l’expérienceprécédente(voirsection
4.1.2)aétérépétéeentriplicata,maisavecdumilieu«InstantOcean»contenant
environ15mMdenitrateaulieude40mM.Aveccettequantitéinitialedenitrate,
laconcentrationmaximalethéoriquepossibledenitriteestd’environ15mM.De
plus,cetteconcentrationcorrespondà-peu-prèsauniveaumaximalobservédansle
mésocosmedufleuveSaint-Laurentaubiodôme(Labbéetal.,2003).
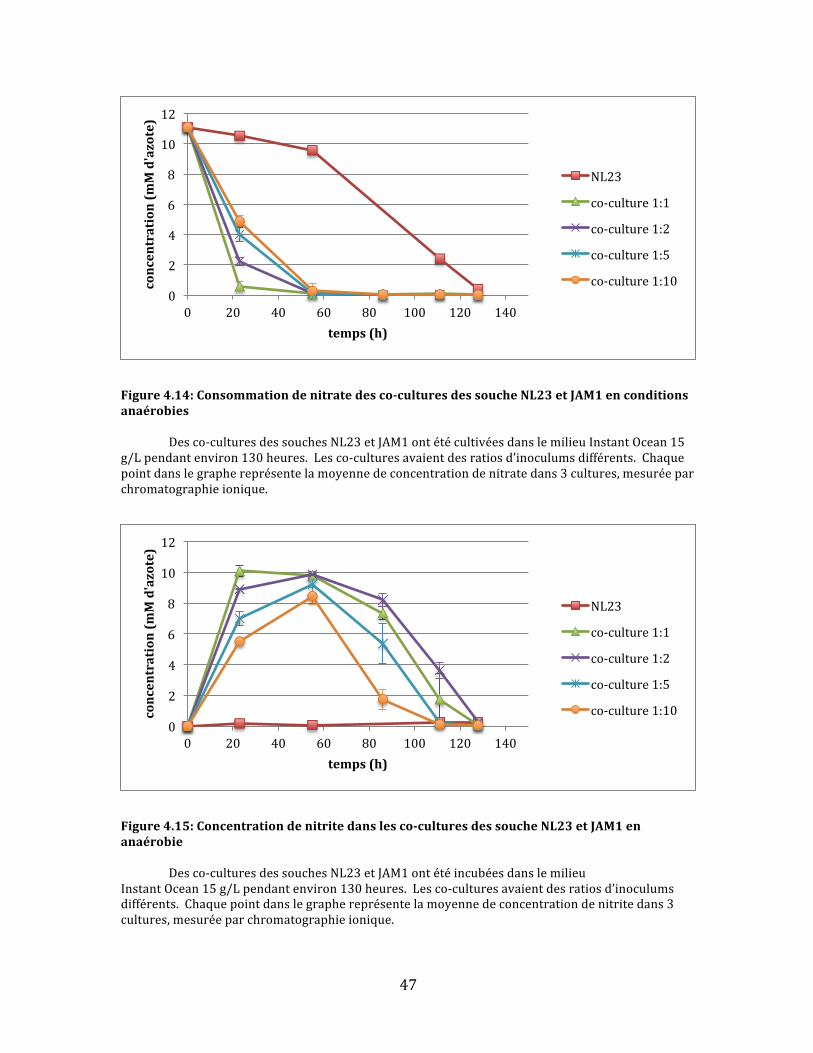
47
Figure4.14:Consommationdenitratedesco-culturesdessoucheNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratedans3cultures,mesuréeparchromatographieionique.
Figure4.15:Concentrationdenitritedanslesco-culturesdessoucheNL23etJAM1enanaérobie Desco-culturesdessouchesNL23etJAM1ontétéincubéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritedans3cultures,mesuréeparchromatographieionique.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10
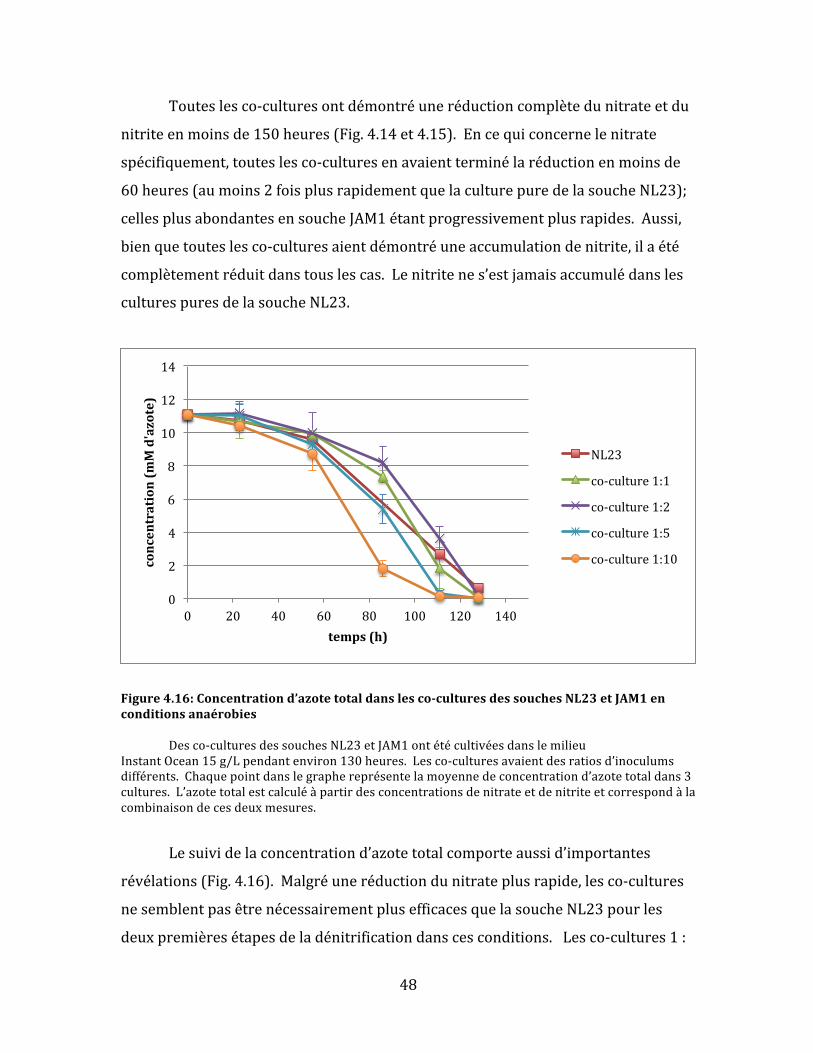
48
Touteslesco-culturesontdémontréuneréductioncomplètedunitrateetdu
nitriteenmoinsde150heures(Fig.4.14et4.15).Encequiconcernelenitrate
spécifiquement,touteslesco-culturesenavaientterminélaréductionenmoinsde
60heures(aumoins2foisplusrapidementquelaculturepuredelasoucheNL23);
cellesplusabondantesensoucheJAM1étantprogressivementplusrapides.Aussi,
bienquetouteslesco-culturesaientdémontréuneaccumulationdenitrite,ilaété
complètementréduitdanstouslescas.Lenitritenes’estjamaisaccumulédansles
culturespuresdelasoucheNL23.
Figure4.16:Concentrationd’azotetotaldanslesco-culturesdessouchesNL23etJAM1enconditionsanaérobies Desco-culturesdessouchesNL23etJAM1ontétécultivéesdanslemilieuInstantOcean15g/Lpendantenviron130heures.Lesco-culturesavaientdesratiosd’inoculumsdifférents.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotaldans3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
Lesuividelaconcentrationd’azotetotalcomporteaussid’importantes
révélations(Fig.4.16).Malgréuneréductiondunitrateplusrapide,lesco-cultures
nesemblentpasêtrenécessairementplusefficacesquelasoucheNL23pourles
deuxpremièresétapesdeladénitrificationdanscesconditions.Lesco-cultures1:
0
2
4
6
8
10
12
14
0 20 40 60 80 100 120 140
concentration(mMd'azote)
temps(h)
NL23
co-culture1:1
co-culture1:2
co-culture1:5
co-culture1:10

49
5et1:10ontdémontréiciuneréductionlégèrementplusrapide.L’étapelimitante
danscesystèmeestdonclaréductiondunitriteparlasoucheNL23.Ilsemble
probablequesontauxdeconsommationdunitriten’estpasouestpeuaccélérépar
laprésencedelasoucheJAM1ouuneréductionaccéléréedunitrate.
Pourtouteslesexpériencesdesuividecroissancesubséquentes,lesco-
culturesontétépréparéesavecunratioinitialde1:5(JAM1:NL23)etenviron15
mMdenitrateinitial.
4.2. CaractérisationdelarelationentrelessouchesNL23etJAM1
4.2.1. Croissancedeculturespuresenprésencedesurnageant
Enplusdupartagedunitrate/nitrite,ilaétéintéressantdechercherdes
indicesdecoopérationentrelessouchesNL23etJAM1.Lapremièretentativede
trouverunetelleinteractionavaitpourbutdedéterminers’ilyavaitexistencede
moléculessécrétéesdanslemilieuparchaquesouchequipouvaientaiderla
croissancedel’autre.L’expérienceaétéexécutéecommesuit:
- DesculturespuresdessouchesNL23etJAM1ainsiquedesco-culturesdes
deuxontétéeffectuéesenconditionsaérobiesdanslemilieu«Instant
Ocean»15g/L.
- Unefoisquelacroissancemaximaleaitétéatteinte,cesculturesontété
centrifugéespourrecueillirlesurnageant.Lescellulesrestantesdansce
surnageantontensuiteétéenlevéesàl’aided’uneseringuemunied’unfiltre.
- Chaquesoucheaétéinoculéedanslesurnageantrécoltédelasouche
opposée(soucheNL23danslesurnageantdelasoucheJAM1etvice-versa).
- Chaquesoucheaaussiétéinoculéedanssonpropresurnageant,dansle
surnageantdelaco-culturedesdeuxsouches(àproportionségales)etd’un
surnageanttémoinnégatif(milieunon-inoculé).
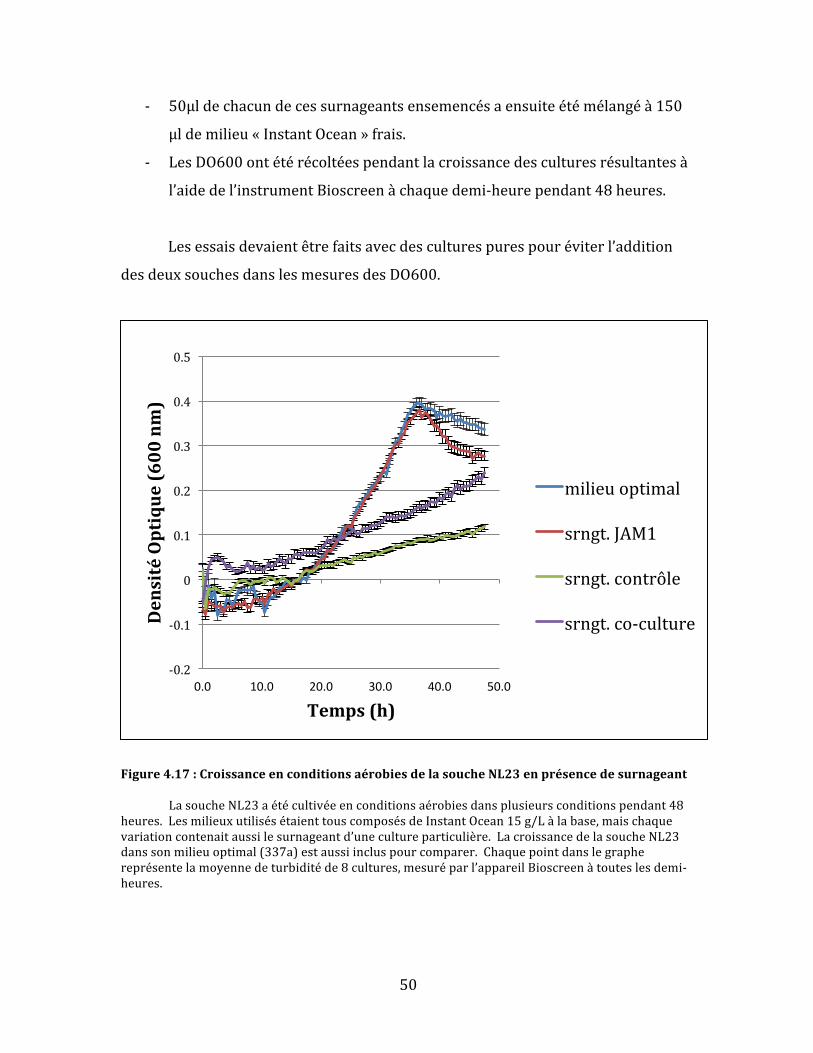
50
- 50µldechacundecessurnageantsensemencésaensuiteétémélangéà150
µldemilieu«InstantOcean»frais.
- LesDO600ontétérécoltéespendantlacroissancedesculturesrésultantesà
l’aidedel’instrumentBioscreenàchaquedemi-heurependant48heures.
Lesessaisdevaientêtrefaitsavecdesculturespurespouréviterl’addition
desdeuxsouchesdanslesmesuresdesDO600.
Figure4.17:CroissanceenconditionsaérobiesdelasoucheNL23enprésencedesurnageant LasoucheNL23aétécultivéeenconditionsaérobiesdansplusieursconditionspendant48heures.LesmilieuxutilisésétaienttouscomposésdeInstantOcean15g/Làlabase,maischaquevariationcontenaitaussilesurnageantd’unecultureparticulière.LacroissancedelasoucheNL23danssonmilieuoptimal(337a)estaussiincluspourcomparer.Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.0 10.0 20.0 30.0 40.0 50.0
DensitéOptique(600nm)
Temps(h)
milieuoptimal
srngt.JAM1
srngt.contrôle
srngt.co-culture

51
Lesrésultatsdémontrentclairementqu’en48heureslasoucheNL23a
atteintunDO600nettementplusélevéeenprésencedusurnageantdelasouche
JAM1qu’enprésencedusurnageantdutémoinnégatif(Fig.4.17).Lesurnageant
desco-culturessembleavoireuuneffetpotentielégalement,maismoinssignificatif.
Étantdonnéquelacroissancedesculturesdutémoinnégatifetdusurnageantdes
co-culturesn’avaitpasatteintsonplateau,ilestpossiblequecesdernièresauraient
éventuellementatteintunDO600semblableàcelledelacultureavecsurnageantde
lasoucheJAM1.LacroissancelasoucheNL23enprésencedesonpropre
surnageantadémontréàpeuprèslamêmecourbequeletémoinnégatifetn’estpas
inclusdanslegraphetelqueprésentéici.L’autreélémentdugrapheestlacourbe
decroissancedelasoucheNL23danssonmilieuoptimal(337a)sansmodifications.
Leprofildecroissanceenmilieu337as’alignepresqueparfaitementavecceluidela
culturedanslesurnageantdelasoucheJAM1.Ainsi,lasoucheNL23cultivéeen
milieu«InstantOcean»(quin’estpasunmilieuoptimal)enprésencedusurnageant
d’uneculturedelasoucheJAM1aétécapabled’atteindreunniveaudecroissance
quiressembleàceluidesacultureenmilieuoptimal.Puisquelemilieu337aestun
milieuminéralnon-salin,cecilaissepeut-êtreentrevoirlerôled’unosmoprotecteur.
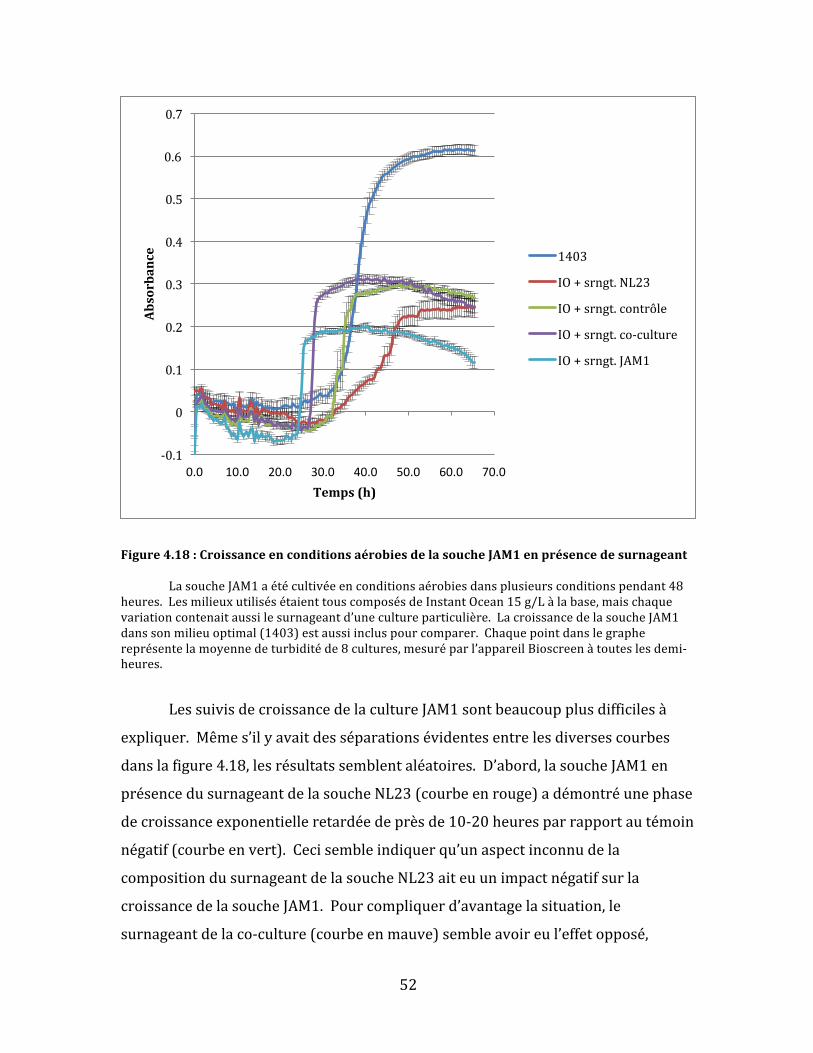
52
Figure4.18:CroissanceenconditionsaérobiesdelasoucheJAM1enprésencedesurnageant LasoucheJAM1aétécultivéeenconditionsaérobiesdansplusieursconditionspendant48heures.LesmilieuxutilisésétaienttouscomposésdeInstantOcean15g/Làlabase,maischaquevariationcontenaitaussilesurnageantd’unecultureparticulière.LacroissancedelasoucheJAM1danssonmilieuoptimal(1403)estaussiincluspourcomparer.Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
LessuivisdecroissancedelacultureJAM1sontbeaucoupplusdifficilesà
expliquer.Mêmes’ilyavaitdesséparationsévidentesentrelesdiversescourbes
danslafigure4.18,lesrésultatssemblentaléatoires.D’abord,lasoucheJAM1en
présencedusurnageantdelasoucheNL23(courbeenrouge)adémontréunephase
decroissanceexponentielleretardéedeprèsde10-20heuresparrapportautémoin
négatif(courbeenvert).Cecisembleindiquerqu’unaspectinconnudela
compositiondusurnageantdelasoucheNL23aiteuunimpactnégatifsurla
croissancedelasoucheJAM1.Pourcompliquerd’avantagelasituation,le
surnageantdelaco-culture(courbeenmauve)sembleavoireul’effetopposé,
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.0 10.0 20.0 30.0 40.0 50.0 60.0 70.0
Absorbance
Temps(h)
1403
IO+srngt.NL23
IO+srngt.contrôle
IO+srngt.co-culture
IO+srngt.JAM1

53
puisquelaphaseexponentielledeJAM1estarrivéelégèrement(environ5heures)
plustôtensaprésence.Bienquecerésultatsoitsurprenant,ilpourraitêtre
expliquéparlapossibilitéquelasoucheJAM1émetdesparticulesdurantsa
croissancequiluisontbénéfiques,tandisquelacroissancedelasoucheNL23
résulteenlasécrétiondeparticulesquisontnéfastesàsacroissance.Lacroissance
delasoucheJAM1enprésencedesonpropresurnageant(courbeenbleupâle)
semblaitcorroborer,enpartie,cettehypothèse,maisaatteintunedensitémaximale
notammentinférieureauxautrescultures.Lestroisautresculturesontatteintun
plateauàpeuprèségal,etlaculturedelasoucheJAM1enmilieuoptimal(1403)
(courbeenbleufoncé)aatteintunDO600quienestdeuxfoissupérieures.Malgré
lanatureinconnuedesdétails,unechoseétaitcertaine:lasoucheNL23nesemble
pasfournird’aideàlacroissancedelasoucheJAM1enconditionsaérobies.
4.2.2. Recherched’unosmoprotecteur
CommelasoucheJAM1estadaptéeàdesconditionsdemilieuxsalins,une
despossibilitésintéressantesdanssoncasseraitlaproductionpotentielled’un
osmoprotecteur,unemoléculecapabledeprotégerunebactériecontrelesstress
osmotiques(parexemple:unehauteconcentrationensel).Avantledébutdece
projet,desrecherchesbioinformatiquesontmontréquelegénomedelasouche
JAM1contientdesgènespermettantlaproductiond’ectoïne,unemolécule
osmoprotectrice.LasoucheNL23aunerésistancelimitéeauxenvironnements
salés,doncl’existencedecettemoléculedansuneco-cultureseraitd’une
importancesignificativepourlacompréhensiondelarelationentrelesdeux
espèces.L’ectoïnepourraitaussiexpliquerenpartiel’aidedecroissancequela
soucheJAM1fournitàlasoucheNL23,tellequ’observédansl’expériencedela
sectionprécédente.
Uneexpérienceadoncétédéveloppéepourvérifierl’existenced’une
protectionosmotiquedelapartdelasoucheJAM1enverslasoucheNL23.Le
protocoled’expérienceaétébasésurleprocessusde«BacterialMilking»(Saueret
al.,1998),quiajustementétédéveloppéepourlarécoltedesolutésincluant

54
l’ectoïne.LasoucheJAM1ad’abordétécultivéedansuneversionmodifiéedeson
milieuoptimal(1403),oùletauxdesalinitéaétéaugmentéà6%NaCl(aulieude
2,4%).Lesculturesontensuiteétécentrifugées,etlesculotsdispersésdansle
milieuoptimaldelasoucheNL23(337a).L’espoirétaitquelaproductionde
moléculesosmoprotectricesinduiteparlemilieuhautementsalinrésulteenla
présencedecesparticulesdanslenouveaumilieu.Aprèsenviron6heuresdansle
milieu337a,lescellulesontétécentrifugéesdenouveau,etcettefoislesurnageanta
étéconservé.Celui-ciaétéséparéenplusieurstubesdanslesquelsdesquantités
progressivementcroissantesdeselontétéajoutées.
Tableau4.1:concentrationdeNaClpartubedanslarecherched’unosmoprotecteur
tube PourcentageNaCL(masse)1 0%(0,00g)2 1%(0,05g)3 1,5%(0,075g)4 2,0%(0,10g)
LasoucheNL23aensuiteétéinoculéedanschacundesnouveauxmilieux,etles
culturesontétésuiviesàl’aidedel’appareilBioscreenpendant48heures.La
soucheNL23aégalementétécultivéedanslemilieu337anon-modifié(pasexposé
auxcellulesdelasoucheJAM1)àtitredecomparaison(Fig.4.19à4.22).
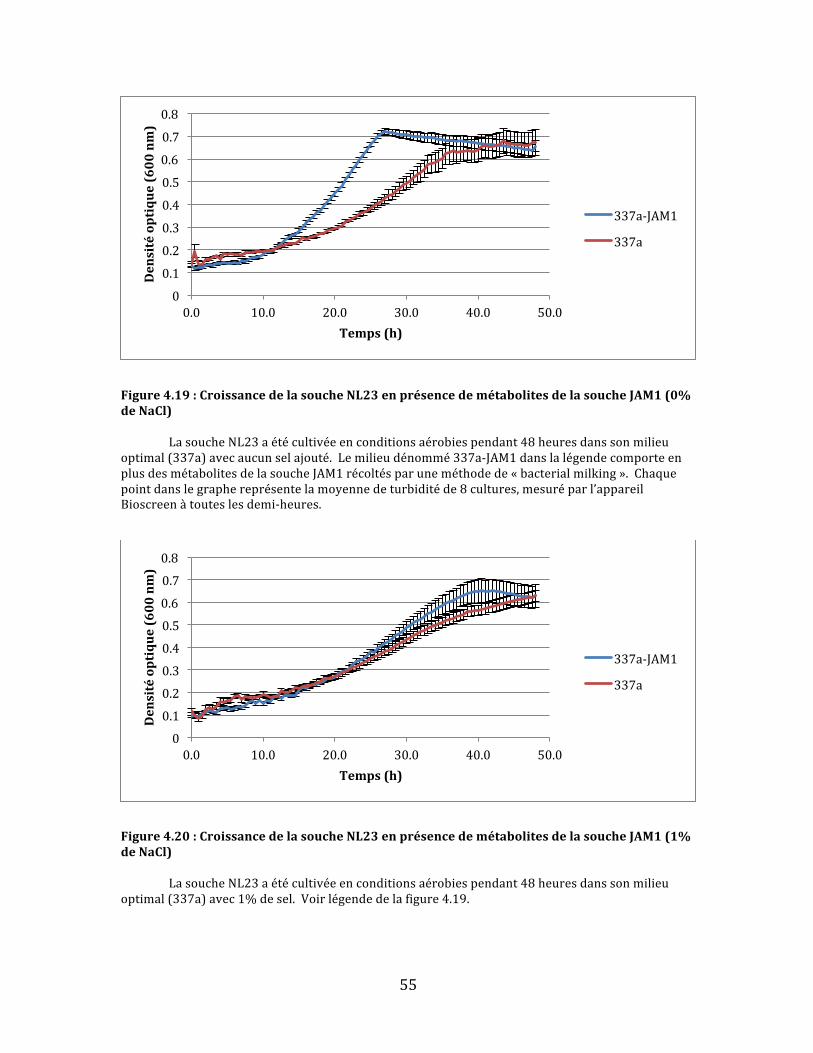
55
Figure4.19:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(0%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avecaucunselajouté.Lemilieudénommé337a-JAM1danslalégendecomporteenplusdesmétabolitesdelasoucheJAM1récoltésparuneméthodede«bacterialmilking».Chaquepointdanslegraphereprésentelamoyennedeturbiditéde8cultures,mesuréparl’appareilBioscreenàtouteslesdemi-heures.
Figure4.20:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec1%desel.Voirlégendedelafigure4.19.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
337a-JAM1
337a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
Densitéoptique(600nm)
Temps(h)
337a-JAM1
337a
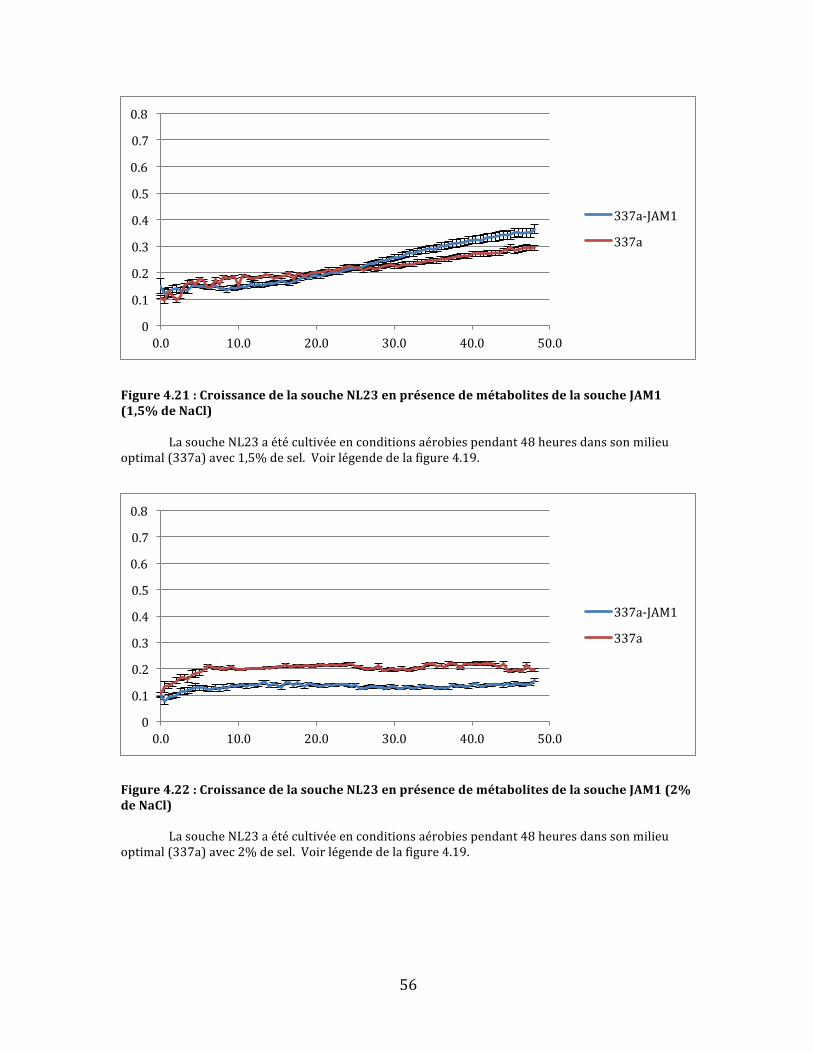
56
Figure4.21:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(1,5%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec1,5%desel.Voirlégendedelafigure4.19.
Figure4.22:CroissancedelasoucheNL23enprésencedemétabolitesdelasoucheJAM1(2%deNaCl) LasoucheNL23aétécultivéeenconditionsaérobiespendant48heuresdanssonmilieuoptimal(337a)avec2%desel.Voirlégendedelafigure4.19.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
337a-JAM1
337a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.0 10.0 20.0 30.0 40.0 50.0
337a-JAM1
337a

57
LesculturesdelasoucheNL23danslemilieu337amodifiéontmontréune
croissanceplusrapidequeletémoinauxconcentrations0%,1%et1,5%deNaCl.
Cetteobservationauraitpuporteràconfusion,maisuneexpérienceprécédentede
ceprojetavaitdéjàdémontréquelasoucheNL23pouvaitcroîtreplusrapidement
enprésenced’unsurnageantdelasoucheJAM1(voirsection4.2.1).Cetaspectdu
résultatn’estdoncpassurprenant.Cequiétaitréellementlepointmajeurdansles
donnéesrecueilliesétaitl’absencedecroissancedanslegraphepourla
concentration2%(Fig.4.22).Eneffet,cecomportementestexactementdequoion
s’attendaitsilasoucheJAM1n’offraitaucuneprotectionosmotique.Toutefois,ilest
possiblequeleprotocoleutilisénepermettaitpasderécolterdesmolécules
osmoprotectrices.Unerépétitiondel’expérienceaconfirmélesrésultatsjusqu’à
uneconcentrationdeselde2,5%.
4.2.3. Caractérisationapprofondiedelaconsommationdunitrate/nitriteà
l’aidedemutantsdelasoucheJAM1
Lorsd’étudesenparallèlesaveccelles-ci,desmutantsdelasoucheJAM1ont
étédéveloppéspouraideràlacaractérisationdesonprofildedénitrification
(Mauffreyetal.,2015).Lestroismutantsenquestionétaienttousrendusdéficients
dansleurhabiletéderéduirelenitrate:
- LasoucheJAM1ΔnarG1(AbsencedusystèmeNar1)
- LasoucheJAM1ΔnarG2(AbsencedusystèmeNar2)
- LasoucheJAM1ΔnarG1narG2(doublemutant)
Commelenitrateestprincipalementimportantdanslecontextedela
dénitrification,l’applicationexpérimentaledesmutantsétaitsurtoutintéressanteen
conditionsanaérobies.Leplanétaitdoncdefaireunsuividecroissancedesco-
culturesdelasoucheNL23avecchacundesmutantsetdelasoucheJAM1non
mutante(ainsiquedesculturespuresdechaquesoucheimpliquéecommetémoin).
Lesco-culturesdelasoucheNL23avecledoublemutantallaientêtre
particulièrementinformatif,puisquel’effetdelasoucheJAM1surlaperformancede

58
lasoucheNL23seraitvisibledanslesuividedénitrificationparchromatographie
ionique.Leratiod’inoculumdedépartpourtouteslesco-culturesétaitde1:5
(nitrate:nitrite),basésurdesrésultatsprécédents(voirsection4.1.2).Lesuivia
étéeffectuésurunepérioded’environ225heuresetleséchantillonsontété
mesurésenDO600etenconcentrationsdenitrateetdenitrite(Fig.4.21à4.23).
Cependant,seulslesrésultatsdelachromatographieioniquesontprésentés,carle
suiviparDO600nerévèlepasbeaucoupd’informationssurl’activitédescultures.
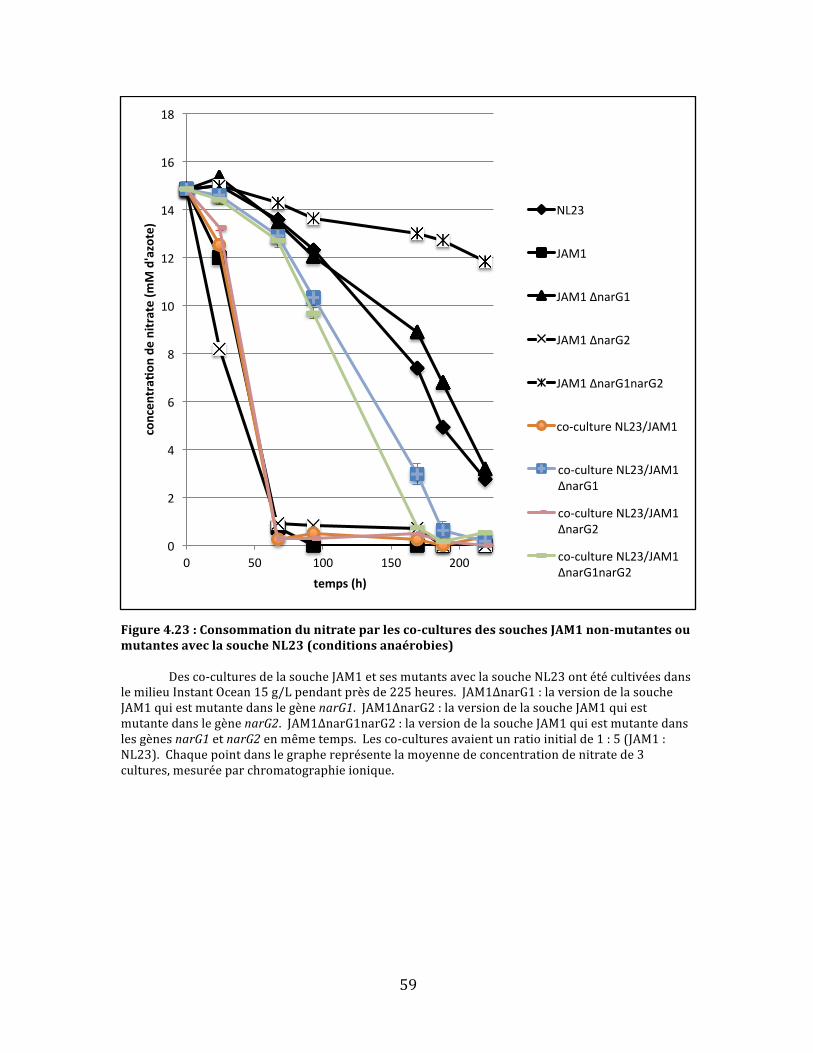
59
Figure4.23:Consommationdunitrateparlesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétécultivéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.JAM1ΔnarG1:laversiondelasoucheJAM1quiestmutantedanslegènenarG1.JAM1ΔnarG2:laversiondelasoucheJAM1quiestmutantedanslegènenarG2.JAM1ΔnarG1narG2:laversiondelasoucheJAM1quiestmutantedanslesgènesnarG1etnarG2enmêmetemps.Lesco-culturesavaientunratioinitialde1:5(JAM1:NL23).Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitratede3cultures,mesuréeparchromatographieionique.
0
2
4
6
8
10
12
14
16
18
0 50 100 150 200
concen
tra+
ondenitrate(m
Md'azote)
temps(h)
NL23
JAM1
JAM1∆narG1
JAM1∆narG2
JAM1∆narG1narG2
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1
co-cultureNL23/JAM1∆narG2
co-cultureNL23/JAM1∆narG1narG2
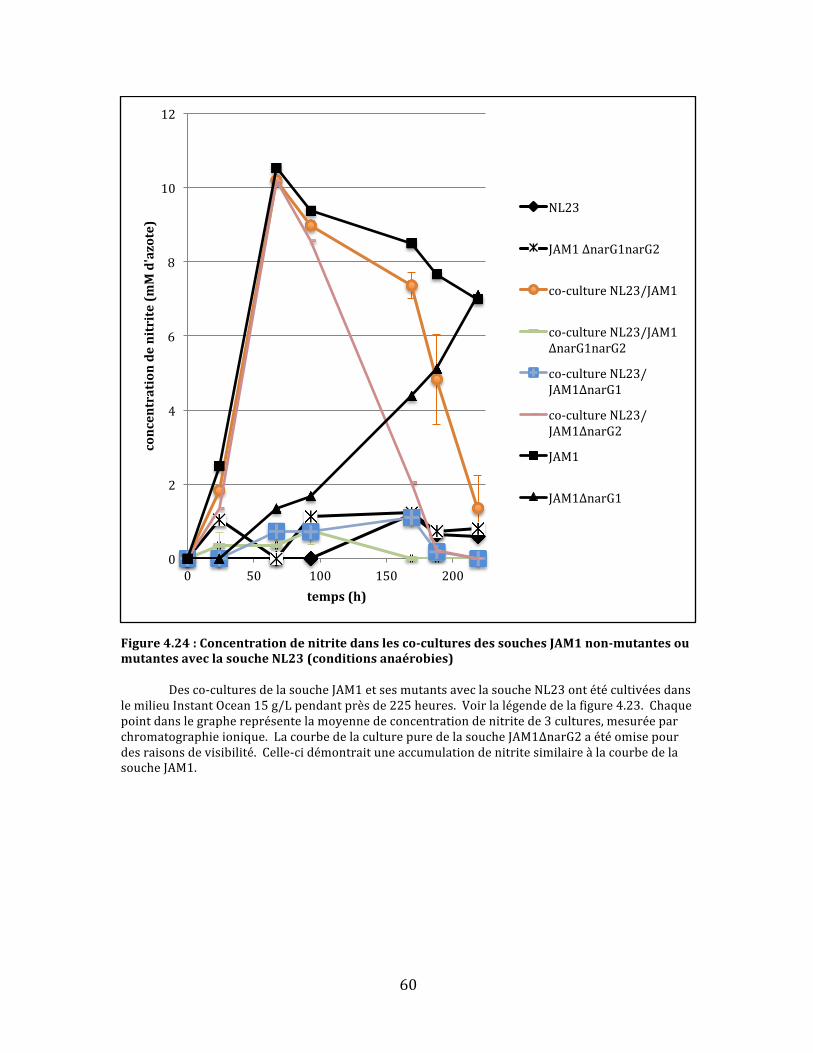
60
Figure4.24:Concentrationdenitritedanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétécultivéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.Voirlalégendedelafigure4.23.Chaquepointdanslegraphereprésentelamoyennedeconcentrationdenitritede3cultures,mesuréeparchromatographieionique.LacourbedelaculturepuredelasoucheJAM1ΔnarG2aétéomisepourdesraisonsdevisibilité.Celle-cidémontraituneaccumulationdenitritesimilaireàlacourbedelasoucheJAM1.
0
2
4
6
8
10
12
0 50 100 150 200
concentrationdenitrite(m
Md'azote)
temps(h)
NL23
JAM1∆narG1narG2
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1narG2
co-cultureNL23/JAM1ΔnarG1
co-cultureNL23/JAM1ΔnarG2
JAM1
JAM1ΔnarG1
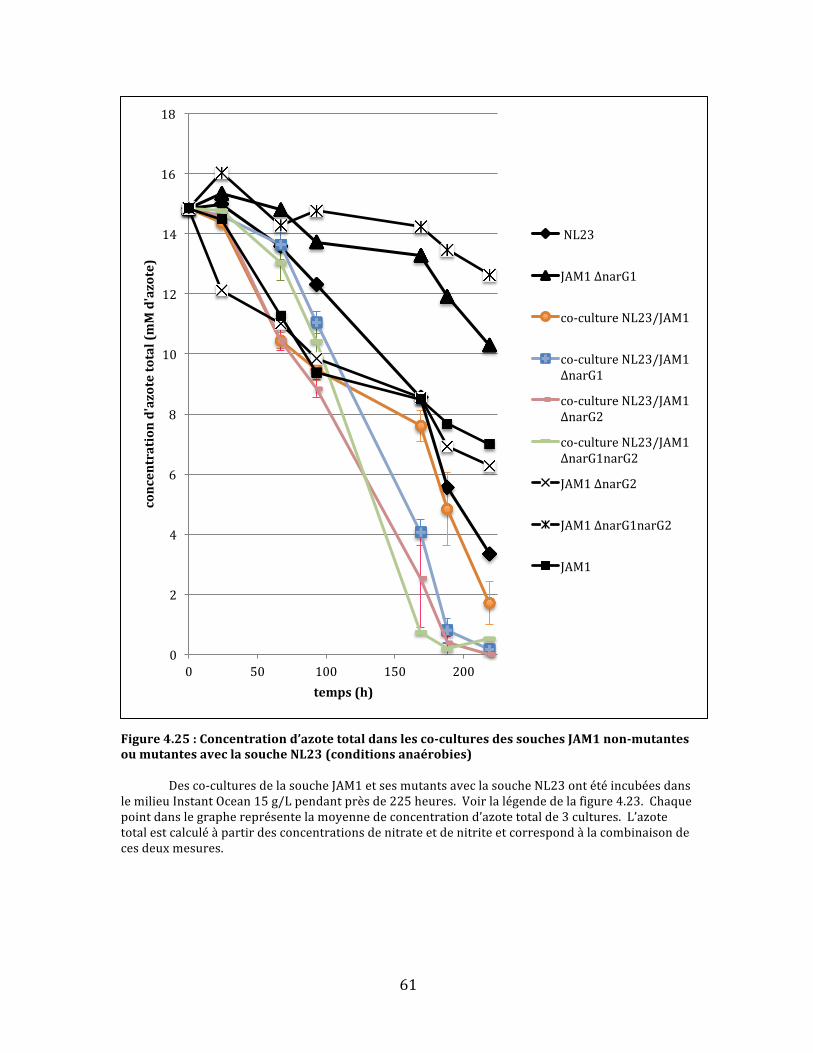
61
Figure4.25:Concentrationd’azotetotaldanslesco-culturesdessouchesJAM1non-mutantesoumutantesaveclasoucheNL23(conditionsanaérobies) Desco-culturesdelasoucheJAM1etsesmutantsaveclasoucheNL23ontétéincubéesdanslemilieuInstantOcean15g/Lpendantprèsde225heures.Voirlalégendedelafigure4.23.Chaquepointdanslegraphereprésentelamoyennedeconcentrationd’azotetotalde3cultures.L’azotetotalestcalculéàpartirdesconcentrationsdenitrateetdenitriteetcorrespondàlacombinaisondecesdeuxmesures.
0
2
4
6
8
10
12
14
16
18
0 50 100 150 200
concentrationd'azotetotal(mMd'azote)
temps(h)
NL23
JAM1∆narG1
co-cultureNL23/JAM1
co-cultureNL23/JAM1∆narG1
co-cultureNL23/JAM1∆narG2
co-cultureNL23/JAM1∆narG1narG2
JAM1∆narG2
JAM1∆narG1narG2
JAM1

62
Lepremieraspectintéressantdesrésultatsestlecomportementdela
dénitrificationdessouchesmutantes.Ledoublemutant(JAM1ΔnarG1narG2)était
évidemmentlemoinsperformantenréductiondunitrate,puisquecelui-cine
possèdepasdenitrateréductasesfonctionnelles(Fig.4.23).Lalégèrebaissede
concentrationdenitrateetd’azotetotalepourraitêtredueàdesfluctuations
causéesparlechromatographe.LemutantJAM1ΔnarG1avaitunprofilderéduction
denitratetrèssimilaireàlasoucheNL23,tandisquelemutantJAM1ΔnarG2aun
profilderéductiondenitratequiressembleplutôtàceluidelasoucheJAM1.Ces
observationsétaientprévisibles;desexpériencesantérieuresavaientdéjà
déterminéquelegènenarG1estleplusactifdelapaire(Mauffreyetal.,2015).
Ilestintéressantdeconstaterquetouteslesco-culturesontréduitlenitrate
plusrapidementquelasoucheNL23enculturespures(Fig.4.23).Pourlesco-
culturesNL23/JAM1,cerésultatn’étaitpasnouveau,puisquecerésultatavaitdéjà
étéobtenu.ÉtantdonnéquelasoucheJAM1ΔnarG2performeexactementcomme
saversionnonmutante,lerésultatpourlesco-culturesNL23/JAM1ΔnarG2n’était
pasétonnantnonplus.Lesco-culturesNL23/JAM1ΔnarG1étaientplusrapidesdans
laréductiondunitratequelemêmemutantenculturespures.Cetauxderéduction
plusélevépourraitêtreattribuéàl’additiondel’activitédeNapAetdeNar2.
Toutefois,cecinepourraitpasexpliquerlemêmecomportementchezlesco-
culturesNL23/JAM1ΔnarG1narG2.Eneffet,ilsemblequelasimpleprésencedela
soucheJAM1double-mutanteaitpermisàlasoucheNL23d’augmentersonactivité
deréductiondenitrate.Ilexisteraitdoncunfacteurd’aideinconnufourniparla
soucheJAM1.Ilestimpossibleparcetteexpériencededéterminercequ’estla
naturedecefacteur.Celui-cipourraitêtrereliéàl’aidedecroissanceobservéeen
conditionsaérobiesdansuneexpérienceprécédente(voirsection4),uneprotection
osmotique,ouunautreélémentjusqu’iciinconnu.
Danslegraphedeconcentrationsdenitrite(Fig.4.24),onvoitquelesseules
co-culturesàavoiraccumulédunitriteétaientlesco-culturesNL23/JAM1et
NL23/JAM1ΔnarG2.Cecirenforcel’idéequelemutantJAM1ΔnarG2secomporte

63
largementcommelasoucheJAM1non-mutante.Aucunesdesautresco-cultures
n’ontaccumulédenitrite.
Commed’habitudelesplusgrandesdécouvertessetrouvaientdanslegraphe
dusuividelaréductiond’azotetotal(Fig.4.25).Premièrement,lesrésultats
corroborentlanotionquelesco-culturesNL23/JAM1nesontpasbeaucoupplus
efficacesàcetégardquelasoucheNL23enculturespures.Deuxièmement,toutes
lesco-culturesavecdesmutantsontréduitl’azotetotalplusrapidementquela
soucheNL23enculturespuresetlesco-culturesNL23/JAM1.Ceciétaitunrésultat
trèssurprenant,surtoutdanslecasdesco-culturesNL23/JAM1ΔnarG1narG2,qui
necomptaientabsolumentpassurlacapacitéderéductiondenitratedelasouche
JAM1.Pourtant,celles-ciontatteint0mMd’azotetotalplusde20heuresavantles
co-culturessansmutantoulasoucheNL23enculturespures.
UnedernièreobservationnotableestquelasoucheJAM1etlemutant
JAM1ΔnarG2ontsembléréduirel’azotetotal.Commeilyapeud’ammoniumdans
lemilieuInstantOcean,l’assimilationdel’azotedoitsefairevialaréductiondu
nitrateennitriteparlavoieassimilatriceprésentedanslasoucheJAM1(Mauffreyet
al.,2016).

64
5. Discussion
5.1. Paramètresoptimauxpourlacroissancedesco-culturesdes
souchesNL23etJAM1
Avantledébutduprojet,unepartiedelarecherchepréliminairesurles
conditionsdesco-culturesdessouchesNL23etJAM1avaitdéjàétéfaite.Lemilieu
InstantOcean15/Lavaitdéjàétédécouvertcommeétantapteàsupporterla
croissancedesdeuxbactériesenculturespuresouenco-cultures.Deplus,la
plupartdesautresparamètresimportants,dontlatempératured’incubation,lepH
dumilieuetlesélémentstracesajoutésétaientdéjàétabliscommeétantlesmêmes
quepourlaculturedubiofilmoriginal.Laproblématiquerestanteétaitsurtoutaxée
surlarecherched’unesolutionauproblèmed’accumulationtoxiquedenitrite.
Lapremièresolutiontentéeétaitd’ajusterlesratiosdebactériesdanslesco-
culturesaumomentdel’ensemencement.Cettesolutionétaitlogiquemaiselleavait
aussiétéutiliséeavecsuccèsensituationdeco-culturesdeClostridiumbutyricumet
Rhodobactersphaeroides.Eneffet,dépendantduratiodesdeuxbactériesdans
l’inoculum,ilpouvaityavoiraméliorationdel’efficacitédelaphotoproduction
d’hydrogènegazeuxparlesco-cultures.Enlimitantl’abondancedeC.butyricum,qui
démontreunecroissancenettementplusrapidequesonhomologue,unéquilibre
bénéfiqueaétéatteint.(Laurinavicheneetal.,2016).Ainsi,pluslaproportiondela
soucheJAM1dansl’inoculumdedépartétaitimportante,plusletauxderéduction
denitrateétaitrapide,étantdonnéquelasoucheJAM1estplusefficacepourréduire
lenitratequelasoucheNL23.Toutefois,enréduisantlaquantitérelativeinitialede
lasoucheJAM1parrapportàlasoucheNL23,ilaétépossibledanscertainscas
d’éviterleniveaufataldenitrite.Lesrésultatsdanscesessaisétaientsouvent
difficilesàreproduireetàanalyserparlasuiteàcausedevariabilitésconsidérables
etimprévisibles.Laplupartdesexpériencescomportaientdestriplicatsdans
lesquelslesculturesavaientuncomportementcomplètementdifférentmalgrédes

65
conditionsdecultureidentiques.Lacausedecettevariabilitédemeureinconnue,
maispourraisêtreattribuéàunparamètretropdifficileàcontrôler,cequiest
parfoislecasdansdessystèmesbiologiques.Desrésultatspréliminairesont
suggéréqueleratioentrelessouchesNL23etJAM1doittendreversuncertain
équilibrepoureffectuerunedénitrificationefficace.Évidemment,lacapacitéde
reproduiredesrésultatsavecconstanceestdésirable.Iladoncétéconcluque
l’ajustementduratioinitialdesbactériesdansl’inoculumnepouvaitêtrequ’une
partiedelasolutionauproblème.
Ladeuxièmesolutionauproblèmed’accumulationdenitriteétaitde
simplementbaisseràuntierslaconcentrationdenitrateinitialedanslemilieude
croissance.Lebutétaitdelimiterlaquantitémaximalepossibledenitriteaccumulé.
Deplus,lesconcentrationsdenitrateobservéesdanslanatureoudansles
aquariumsd’oùprovientlaproblématiquen’approchentjamais40mMd’azotetel
quedanscesexpériences,etleseffetstoxiquessurlesanimauxsontressentisàdes
concentrationsnettementinférieures(DeRoosetal.,2003,Labbé,2007).L’impact
decetteactionaétédécisif.Avecunequantitémoindredenitrateàsadisposition,
lesco-culturesn’ontjamaispuaccumulerassezdenitritepourquelaconcentration
deviennenocive.Touteslesexpériencessubséquentesavaientdoncunratioinitial
de1:5(JAM1:NL23)etenviron15mMdenitrateaulieude40mM,ainsi
combinantlesrésultatsdesdeuxsolutions.
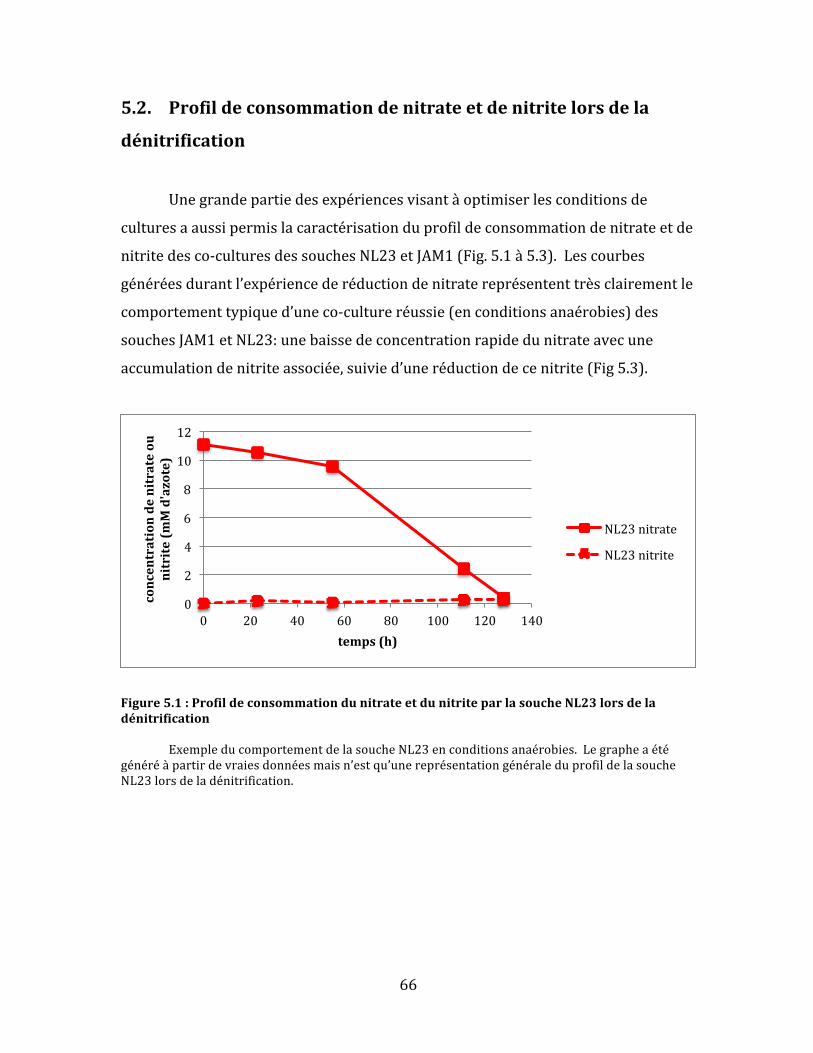
66
5.2. Profildeconsommationdenitrateetdenitritelorsdela
dénitrification
Unegrandepartiedesexpériencesvisantàoptimiserlesconditionsde
culturesaaussipermislacaractérisationduprofildeconsommationdenitrateetde
nitritedesco-culturesdessouchesNL23etJAM1(Fig.5.1à5.3).Lescourbes
généréesdurantl’expériencederéductiondenitratereprésententtrèsclairementle
comportementtypiqued’uneco-cultureréussie(enconditionsanaérobies)des
souchesJAM1etNL23:unebaissedeconcentrationrapidedunitrateavecune
accumulationdenitriteassociée,suivied’uneréductiondecenitrite(Fig5.3).
Figure5.1:ProfildeconsommationdunitrateetdunitriteparlasoucheNL23lorsdeladénitrification ExempleducomportementdelasoucheNL23enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofildelasoucheNL23lorsdeladénitrification.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
NL23nitrate
NL23nitrite
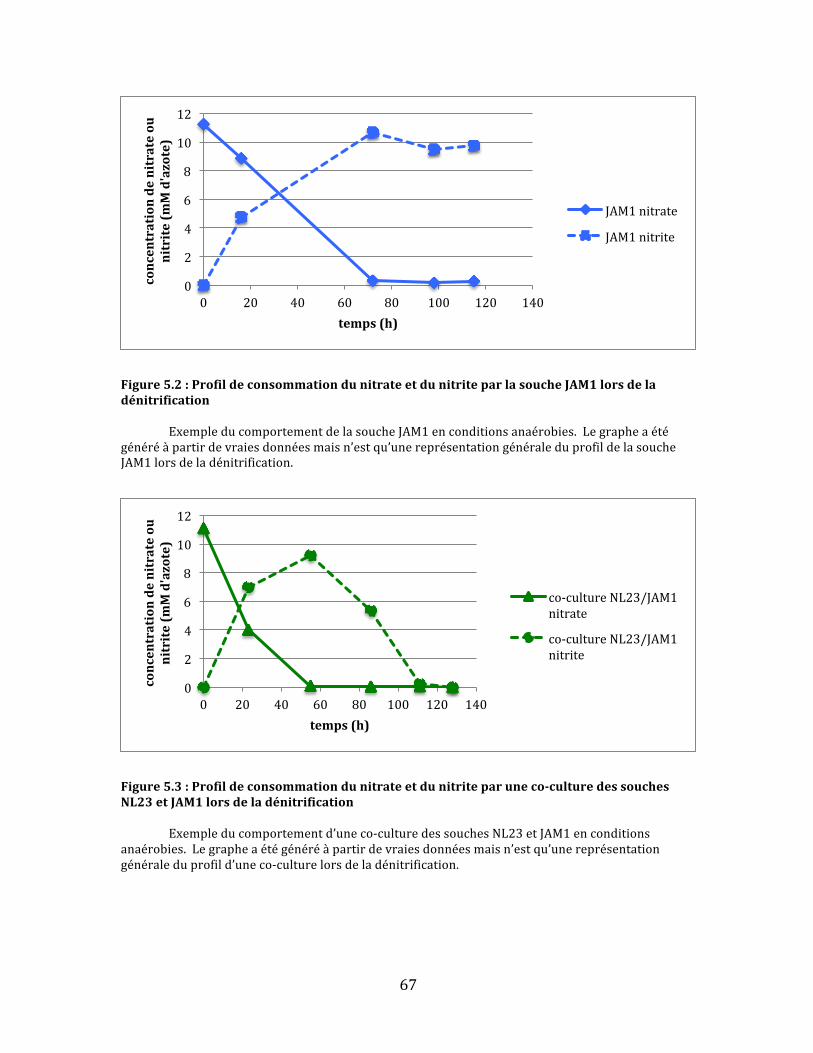
67
Figure5.2:ProfildeconsommationdunitrateetdunitriteparlasoucheJAM1lorsdeladénitrification ExempleducomportementdelasoucheJAM1enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofildelasoucheJAM1lorsdeladénitrification.
Figure5.3:Profildeconsommationdunitrateetdunitriteparuneco-culturedessouchesNL23etJAM1lorsdeladénitrification Exempleducomportementd’uneco-culturedessouchesNL23etJAM1enconditionsanaérobies.Legrapheaétégénéréàpartirdevraiesdonnéesmaisn’estqu’unereprésentationgénéraleduprofild’uneco-culturelorsdeladénitrification.
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
JAM1nitrate
JAM1nitrite
0
2
4
6
8
10
12
0 20 40 60 80 100 120 140
concentrationdenitrateou
nitrite(mMd'azote)
temps(h)
co-cultureNL23/JAM1nitrate
co-cultureNL23/JAM1nitrite
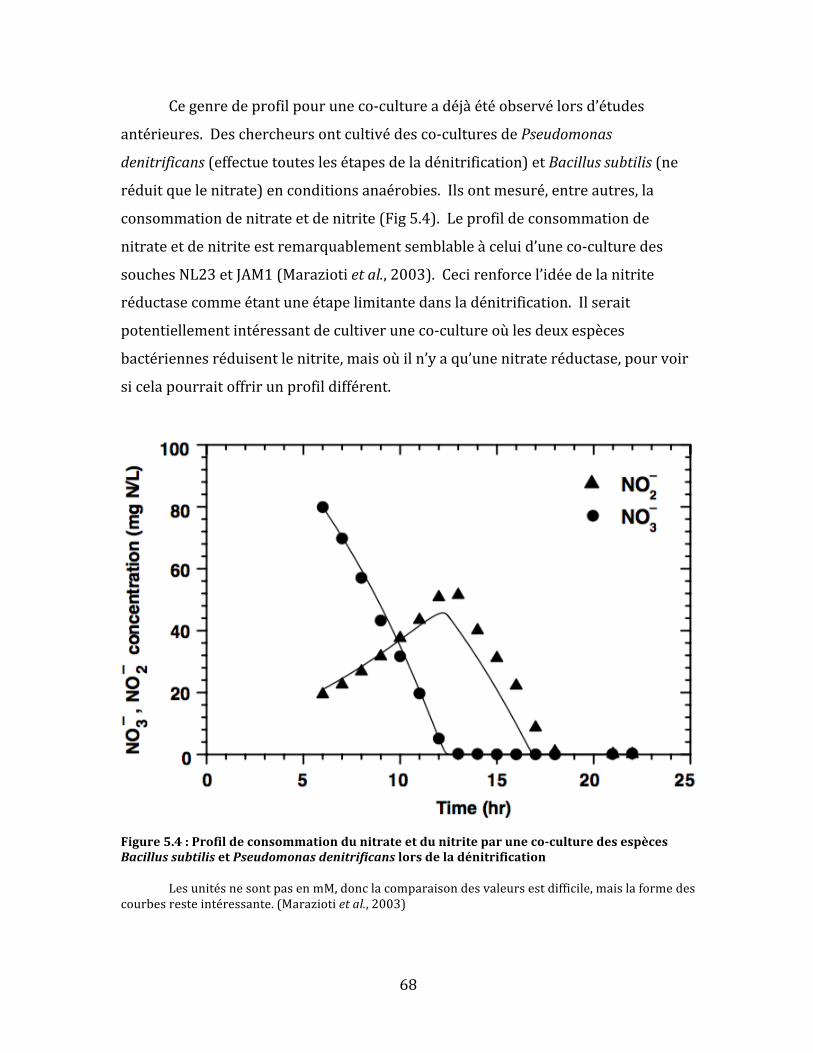
68
Cegenredeprofilpouruneco-cultureadéjàétéobservélorsd’études
antérieures.Deschercheursontcultivédesco-culturesdePseudomonas
denitrificans(effectuetouteslesétapesdeladénitrification)etBacillussubtilis(ne
réduitquelenitrate)enconditionsanaérobies.Ilsontmesuré,entreautres,la
consommationdenitrateetdenitrite(Fig5.4).Leprofildeconsommationde
nitrateetdenitriteestremarquablementsemblableàceluid’uneco-culturedes
souchesNL23etJAM1(Maraziotietal.,2003).Cecirenforcel’idéedelanitrite
réductasecommeétantuneétapelimitantedansladénitrification.Ilserait
potentiellementintéressantdecultiveruneco-cultureoùlesdeuxespèces
bactériennesréduisentlenitrite,maisoùiln’yaqu’unenitrateréductase,pourvoir
sicelapourraitoffrirunprofildifférent.
Figure5.4:Profildeconsommationdunitrateetdunitriteparuneco-culturedesespècesBacillussubtilisetPseudomonasdenitrificanslorsdeladénitrification LesunitésnesontpasenmM,donclacomparaisondesvaleursestdifficile,maislaformedescourbesresteintéressante.(Maraziotietal.,2003)

69
Lorsdel’expérienceaveclesmutantsdelasoucheJAM1,ilyavaitaussi
l’occasiondecorroborerdesnotionsreliéesauxdeuxnitrateréductases(Nar1et
Nar2)delasoucheJAM1.IlaétéconfirméquelanitrateréductaseNar1estla
principaleresponsabledelaréductiondunitrate.Eneffet,unesouchemutantede
lasoucheJAM1oùladeuxièmenitrateréductase(Nar2)aétédésactivée(doncNar1
actifseulement)reproduitpresqueexactementlemêmetauxderéductiondu
nitratequelasouchesauvage.DanslecasinverseoùNar1estinactivé,lesrésultats
ontdémontréquelasouchemutanteavecNar2actifseulementaunedynamiquede
réductiondunitrateéquivalenteàlasoucheNL23quipossèdeunenitrateréductase
detypeNap.
L’aspectleplusimportantengénéraldecetteoptiqueétaitl’efficacitéd’une
co-culturepourréduirel’azotetotal,c’estàdirelavitesseàlaquellelenitrateetle
nitritesontréduitsàzéro.Unedeshypothèsescentralesduprojetétaitquelesdeux
espècessontplusefficacesensemblesqu’individuellement.Incidemment,
l’améliorationdurendementmétaboliqueparl’applicationdeco-culturesestlebut
deplusieurseffortsderecherche(ex:laproductiond’hydrogèneparuneco-culture
deC.butyricumetR.sphaeroides,(Laurinavicheneetal.,2016).Lespremiers
résultatsàcetégardn’étaientpasencourageants:Lesco-culturesnesemblaientpas
êtrenécessairementplusefficacespourréduirel’azotetotal,prenantàpeuprès
autantdetempsquequelasoucheNL23enculturepurepourleréduireàzéro.Les
seulesco-culturesquiontdémontréuneaméliorationdecetaspectétaientcelles
aveclasoucheNL23avecuneversionmutantedeJAM1quiavaitNar1désactivé.
CombinéaveclanotionquelasoucheJAM1fournituneaidedecroissanceàla
soucheNL23(expliquéplusbas),cecipourraitindiquerquelasoucheNL23est
négativementaffectéeparl’activitédénitrifiantedelasoucheJAM1,mais
positivementaffectéeparsaprésence.

70
5.3. LessouchesNL23etJAM1enbiofilm
Unedesgrandesquestionsaudébutduprojetétaitlerôledelastructuredu
biofilmdansl’interactiondesbactériesNL23etJAM1,surtoutdanslecasde
l’osmoprotectiondelasoucheNL23danslesmilieuxconcentrésenNaCl(plusque
1.5%).Ladécisionadoncétéprisedetenterlacréationd’unbiofilmcomposéde
chacunedessouchesNL23etJAM1ouen«co-biofilm».Lacréationetl’observation
d’unbiofilmconsistaientdéjàunprocessusconnudanslelaboratoire.Desétudes
précédentesportaientsouventsurlacompositionetlaperformancedubiofilm
originalrecueilliauBiodômedeMontréal.Lesméthodesdecréationdubiofilm
développéesdanscesétudesprécédentesonttoutdemêmeservid’exempledansla
méthodeemployéeici(unedescriptiondecetteméthodeestdisponibleenannexe).
Unbiofilmcommecelui-ciauraitpuservirdansplusieursexpériences
différentes,maisl’idéeinitialeétaitd’étudierunrôlepossibledelastructuredu
biofilmdanslaprotectiondelasoucheNL23enmilieusalé.Lebiofilmoriginal(au
biodômeetenlaboratoire)étaitcultivéenmilieuavec3%desel.LasoucheNL23
étantunedesespècesprincipalesdecesystème,elledevaitnécessairementrecevoir
uneaideextérieure.CetaidepourraitprovenirdelasoucheJAM1,parexemplepar
l’entremisedel’osmoprotecteurectoïne;ilaétédocumentépourd’autresco-
culturesquel’aided’unosmoprotecteurn’estefficacequ’enbiofilm.Dansunesérie
d’expériences,unesouchebactériennehalophiledugenreChromohalobacteraété
cultivéeenco-cultureavecuneespècehalotolérantedugenreDietza.L’espèce
halophilenedémontreaucunecroissanceenmilieuxcontenantmoinsde5%desel,
tandisquel’espècehalotolérantenedémontreaucunecroissanceenmilieux
contenantplusde12%desel.Enco-culturesplanctoniques(ensuspensionliquide),
iln’yavaitpasd’effetprotecteurd’uncôtéoudel’autre;seulesdescellulesde
l’espècehalotoléranteontétédétectéesdanslemilieusanssel,etseulesdescellules
del’espècehalophileontétédétectéesdanslemilieude15%desel.Quandles
espècesétaientcultivéesen«co-biofilm»,l’espècehalotoléranten’apportaitaucune

71
aideàl’autrebactérieenmilieusanssel.Cependant,unequantitéàpeuprèségale
desdeuxespècesontétédétectéedansle«co-biofilm»à15%desel.Les
chercheursontsuggérélerôledesosmoprotecteursectoïneetglutamatedansle
résultat(Plakunovetal.,2008).Malgrétout,ilneseraitpassurprenantquela
protectiondanslecasdessouchesNL23etJAM1provienneplutôtdirectementdela
structuredubiofilmetdesEPSquilecompose,étantdonnélagrandevariétéde
fonctionsquiluisontattribué.
L’expérienceplanifiéeétaitdecultiverlebiofilmenmilieuInstantOceanavec
1,5%deNaCl,puisdeletransférerdanslemêmemilieuà3%deNaCl,pourune
autresériedetransferts.Lasurvie(ounon)delasoucheNL23,encomparaison
avecuneco-cultureliquide,auraitalorsétéuneindicationduniveaudeprotection
offertparlebiofilm.Uneautreindicationdelaprotectionauraitétéletaux
dénitrification.SilacultureenbiofilmdanslemilieuInstantOcean(3%NaCl)était
arrivéeàréduirelenitrite,celaauraitétéunsigned’activitédelapartdelasouche
NL23.
Figure5.5:Supportenplastiquegarnid’unbiofilm Dessupportsenplastiquecommecelui-ciontétéutiliséspourfournirunesurfaced’accrochageausoucheNL23etJAM1pourlacréationd’unbiofilm.Lessupportssonassezpetitspourêtrefacilementinsérésetretirésdesfiolesdeculture.

72
Pourdesraisonsqu’onignore,nilessouchesindividuelles,nilaco-culture
dessouchesNL23etJAM1n’ontdémontrélacapacitédeformerunbiofilm.Les
raisonsexactesdecerésultatsontévidemmentinconnues,maisonpeutdévelopper
deshypothèses.L’échecpourraitêtredûàl’absenced’uneespècecolonisatrice
importante,puisquedescaractéristiquesspécifiquessontrequispourfaciliter
l’adhérencedecellulesàuneparoi.Lesfacteursassociésauxstagesinitiauxdela
formationd’unbiofilmpourE.coli,parexemple,incluentlamotilité(ex.:flagelles),
laprésencedefimbriae,laprésencedecurlietlaproductiondecertainsEPSclés,
dontlepolysaccaridePGA(β-1,6-N-acetyl-D-glucosamine)(VanHoudtetal.,2005).
Lacapacitédeformationdebiofilmpeutvarierénormémententreespècesetmême
entresouchesdelamêmeespèces,commeilaétédémontrépourCronobacter
sakazakii(Yeetal.,2015).Ilestaussidocumentéquelaformationdebiofilmpour
certainesespècesestfacilitéeparlacolonisationinitialed’unesurfaceparuneautre
espèce.L’espècebactérienneSphingomonasnatatoriaestcapabledeformerdes
biofilmsfragilesenculturepure,maiselleformedesbiofilmsplusrobustes
lorsqu’elleestexposéeàunesurfacerecouvertedebactériesdel’espèceMicrococcus
luteus.Ceciestdûàune«coagrégation»,unereconnaissanceetunattachement
spécifiqueentrelesdeuxbactéries(Minetal.,2009).
Lebiofilmoriginalestunecommunautécomplexedebactériesetd’autres
organismes;ilestpossiblequ’unouplusieursdeceux-ciagissentcomme
spécialistescolonisateursdesurfacesetsontdoncnécessairesàlaformation
efficaced’unbiofilm.Dansunedesanalysesdubiofilmoriginal,ilavaitéténoté
qu’unegrandevariétédecelluleseucaryotescomposaientuneproportion
importante(20%)delabiomasseaudébutdudéveloppementdubiofilm;ces
cellulesdevenaientmoinsabondantesparlasuite(5-6%)(Labbéetal.,2007).Ceci
pourraitêtreindicatifd’unrôledecolonisationounon,maislechangement
dramatiquedelacompositiondelabiomassedubiofilmestcertainementune
indicationquecertainesbactériessontresponsablesdedifférentsélémentsdelavie
microbienneenbiofilm,incluantlacolonisation.Ilestentièrementpossiblequeles
souchesNL23etJAM1nesoientpasaxéessurcettepartiedudéveloppementd’un

73
biofilm.Ilestaussipossiblequelesconditionsdanslesquellesl’expériences’est
dérouléen’étaientpasoptimales.Lasurfaceenplastiquedessupportsn’étaitpeut-
êtrepasapteaubiofilm;dessurfacesenverreontététestéeségalement,sansplus
derésultat.
Éventuellement,cetaspectduprojetaétéabandonnéavecleverdictquela
formationd’unbiofilmcomposédessouchesNL23etJAM1n’estpaspossibleavec
lesméthodesutilisées.
5.4. Échangederessources
Lesexpériencesontdémontréqu’ilyavaitaumoinsexistenced’uneaide
bénéfiquedecroissanceetd’activitédénitrifiantedanslesco-culturesdelapartde
lasoucheJAM1.Eneffet,ilsemblequ’ilexisteunfacteurinconnuquiaidelasouche
NL23àcroîtreplusrapidement(démontréenconditionsaérobies)etàréduirele
nitrateplusrapidement(démontréenconditionsanaérobies).L’identitédece
mécanismeresteindéterminée;Ilpourraits’agird’uneparticuleajoutéeou
transférée,oualorsl’enlèvementd’unélémentlimitant.Deplus,ilseraitpossible
queplusieursfacteurssoientprésents,etmêmequedifférentsfacteurssoient
responsablesdesaméliorationsobservéesenaérobioseetenanaérobiose.L’aspect
unidirectionneldecetaideaaussiétédémontré;lasoucheJAM1n’ayantjamais
montrélamoindreindicationdeprofiterdelaprésencedelasoucheNL23,voire
mêmesaprésencepouvantêtrenocive.Dansl’étudemenéeparTakakietal.(2007)
(voirsection2.8),l’espèceStreptomycesgriseusfournissaituneaidede
dénitrificationàRalstoniapickettiiparl’entremisedeprotéasesrelâchéesdans
l’environnement.Pourarriveràcetteconclusion,leschercheursontpremièrement
cultivéR.pickettiienprésencedesurnageantdeS.griseusenconditionsanaérobies.
Cetteastuceaétéutiliséeiciégalement,maisseulementenconditionsaérobies.
Pourconfirmerquec’estbienunemoléculesecrétéeoul’absenced’unélément
limitantquiestresponsabledel’améliorationdeladénitrificationdelasouche
NL23,ilfaudraitrépéterl’expérienceenconditionsanaérobies.Deuxièmement,le

74
surnageantdeS.griseusaétésoumisàuneultracentrifugationpourcréerune
séparationdescomposantesparmassemolaire.Entestantl’effetdechaquefraction
demoléculesséparément,ilestdevenupossibled’identifierlesplusmassives
commeétantresponsablesdutauxsupérieurdedénitrification,cequiamenéà
l’hypothèsedesprotéases.Ceciestunestratégiequipourraitêtreintéressantepour
identifierlanaturedufacteurd’aidedelasoucheJAM1.Deplus,cettemêmeétudea
concluquel’histidinejouaitpeut-êtreunrôledansl’améliorationdel’activité
dénitrifiante.CecipourraitêtrerapidementtestéaveclasoucheNL23également.
Uncandidatd’unfacteuraidantlasoucheNL23estl’ectoïne.Ilétait
connuquelasoucheJAM1possèdelesgènesnécessairesàlaproductiond’ectoïne,
uneparticuleosmoprotectrice.Ilaétérécemmentdéterminédanslelaboratoire
quelasoucheJAM1produisaitdel’ectoïnedanslemilieudeculture(donnéesnon-
publiées).Lorsdeceprojet,leseffortspourdémontrerl’effetd’osmoprotectionde
lasoucheJAM1ontéchoué(voirsection4.3.2),maiscelaneprouvepasquel’ectoïne
nejouepasderôle.Dessolutionspossiblesseraientdetrouverunmoyende
quantifierl’ectoïnedirectementoudeconcentrerl’ectoïneproduitparlasouche
JAM1pourinduiresoneffet.Ilseraitaussipossiblequel’ectoïnen’estpas
responsabledeseffetsobservésdanslessituationsplanctoniques.Dansuneétudeà
cesujet,ilaétédéterminéquel’ectoïneétaitsimplementtropdiluéedansune
cultureensuspensionliquidepouravoiruneffetnotable(Plakunovetal.,2008).
Danscecas,l’effetdel’ectoïnenepourraitêtreobservéquesiuneméthodeétait
trouvéepourproduireunbiofilmaveclessoucheNL23etJAM1.

75
6. Conclusion
Desconditionsoptimalesdeco-culturesdessouchesNL23etJAM1ontété
développées,cequiapermisd’optimiserlesétudessuivantes.Laproblématique
d’accumulationdenitriteparlaréductiontroprapidedunitrateaétécontournée
parlabaissedenitrateinitialedanslemilieuetl’ajustementduratiodessouchesau
momentdel’ensemencement.Lesmêmesexpériencesontaussidévoiléplusieurs
aspectsducomportementdesdeuxsouchesenconditionsaérobiesetanaérobies,
oùcelles-ciperformentladénitrification.
Ilaétédémontréquelesco-culturesdessouchesNL23etJAM1n’étaientpas
nécessairementplusrapidesendénitrificationquelasoucheNL23encultures
pures.Toutefois,ilaégalementétédémontréquelesco-culturesdelasoucheNL23
aveclessouchesmutantesdeJAM1quiontlanitrateréductaseNar2désactivée
étaientcapablesd’unedénitrificationplusrapidequelasoucheNL23encultures
pures.
Lestentativesdeformationsd’unbiofilmuniquementcomposédessouches
NL23etJAM1ontéchoué.Ilsemblequedanslesconditionsimposées,cesdeux
espècesbactériennesnepeuventpasdévelopperdebiofilm.
IlaétédémontréquelasoucheJAM1apporteuneaidedecroissanceàla
soucheNL23enconditionsaérobiesetuneaidederéductiondenitrateen
conditionsanaérobies.LasoucheNL23n’apasfaitdemêmepourlasoucheJAM1
danslesconditionsd’expérience.Cependant,iln’apuêtredémontréquelleestla
naturedecetaide.

76
Annexe
Protocoledecréationd’unbiofilm
Certainesexpériencesontrequislaformationd’unbiofilm.Pourcefaire,des
supportsenplastiquesontétéinsérésdanslesfiolesdecultureenprésencede
bactériesensuspension.Cesensemencementssontensuiteincubésenconditions
anaérobies.Puisqueledéveloppementd’unbiofilmapuprendreplusieurs
semaines,lemilieudanslequelsetrouvaitlacultureadûêtrerégulièrementchangé
pouryéviterladéplétiondesressources.Lessupportsontdoncététransférés
régulièrementdansunenouvellefiolecontenantdumilieufrais.Pourlesco-
culturesdessouchesJAM1etNL23,lapériodedetempsentrelestransfertsétait
d’environunesemaine.
Pourtransférerlessupportsenévitantlacontaminationexterneetentreles
échantillons,touslesoutilsutilisésontdûêtrestérilesetletravailadûêtrefaitdans
lahottebiologique.Lessupportsontétéenlevésdesfiolesàl’aidedepincespuis
insérésdanslesnouvellesfiolespréalablementautoclavées.Celles-ciontensuiteété
ferméeshermétiquementetl’oxygèneàl’intérieuraétéenlevépourcréer
l’environnementd’anaérobiose.Danslecasdeceprojet,leprocessusaétérépété
jusqu’àcequ’ilestdevenuévidentqu’iln’yauraitpasformationdebiofilm(4
semainesouplus).
77
Figure7.1:Schémadelaméthodedecréationd’unbiofilm Leprocessusdecréationdubiofilmimpliqueletransfertàrépétitiondesupportscoloniséspardesbactériesenanaérobies.

78
Listedesréférences
AndersonIC&LevineJS(1986)RelativeRatesofNitricOxideandNitrousOxideProductionbyNitrifiers,Denitrifiers,andNitrateRespirers.Appliedandenvironmentalmicrobiology51(5):938-945.
Doi:AuclairJ,LepineF,ParentS&VillemurR(2010)Dissimilatoryreductionofnitrate
inseawaterbyaMethylophagastraincontainingtwohighlydivergentnarGsequences.TheISMEjournal4(10):1302-1313.
Doi:10.1038/ismej.2010.47BerksBC,FergusonSJ,MoirJW&RichardsonDJ(1995)Enzymesandassociated
electrontransportsystemsthatcatalysetherespiratoryreductionofnitrogenoxidesandoxyanions.Biochimicaetbiophysicaacta1232(3):97-173.
Doi:BlaisM-A(2014)SuividelacontaminationfécaledanslagranderégiondeMontréal.
(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).CamargoJA,AlonsoA&SalamancaA(2005)Nitratetoxicitytoaquaticanimals:a
reviewwithnewdataforfreshwaterinvertebrates.Chemosphere58(9):1255-1267.
Doi:10.1016/j.chemosphere.2004.10.044DeRoosAJ,WardMH,LynchCF&CantorKP(2003)Nitrateinpublicwatersupplies
andtheriskofcolonandrectumcancers.Epidemiology14(6):640-649. Doi:DoroninaN,LeeTD,IvanovaE&TrotsenkoYA(2005)Methylophagamuratasp.
nov.:ahaloalkaliphilicaerobicmethylotrophfromdeterioratingmarble.Microbiology74(4):440-447.
Doi:FesefeldtA,KloosK,BotheH,LemmerH&GliescheC(1998)Distributionof
denitrificationandnitrogenfixationgenesinHyphomicrobiumspp.andotherbuddingbacteria.Canadianjournalofmicrobiology44(2):181-186.
Doi:FlemmingHC&WingenderJ(2010)Thebiofilmmatrix.Naturereviews.
Microbiology8(9):623-633. Doi:10.1038/nrmicro2415GliescheC,FesefeldtA&HirschP(2005)HyphomicrobiumStutzerandHartleb
1898,76AL.Bergey’sManual®ofSystematicBacteriology:476-494. Doi:GregoryLG,BondPL,RichardsonDJ&SpiroS(2003)Characterizationofanitrate-
respiringbacterialcommunityusingthenitratereductasegene(narG)asafunctionalmarker.Microbiology149(1):229-237.
Doi:

79
HajipourA,MoghadamN,NosratiM&ShojasadatiS(2011)Aerobicthermophilictreatmentoflandfillleachateinamoving-bedbiofilmbioreactor.JournalofEnvironmentalHealthScience&Engineering8(1):3-14.
Doi:HayatsuM,TagoK&SaitoM(2008)Variousplayersinthenitrogencycle:Diversity
andfunctionsofthemicroorganismsinvolvedinnitrificationanddenitrification.SoilScienceandPlantNutrition54(1):33-45.
Doi:10.1111/j.1747-0765.2007.00195.xHinoT,NaganoS,SugimotoH,ToshaT&ShiroY(2012)Molecularstructureand
functionofbacterialnitricoxidereductase.Biochimicaetbiophysicaacta1817(4):680-687.
Doi:10.1016/j.bbabio.2011.09.021JanvierM&GrimontP(1995)ThegenusMethylophaga,anewlineofdescentwithin
phylogeneticbranchγofProteobacteria.Researchinmicrobiology146(7):543-550.
Doi:JebbarM,TalibartR,GlouxK,BernardT&BlancoC(1992)Osmoprotectionof
Escherichiacolibyectoine:uptakeandaccumulationcharacteristics.JBacteriol174(15):5027-5035.
Doi:KempfB&BremerE(1998)Uptakeandsynthesisofcompatiblesolutesas
microbialstressresponsestohigh-osmolalityenvironments.Archivesofmicrobiology170(5):319-330.
Doi:LabbéN(2007)Caractérisationdudynamismemicrobiendudénitrificateurmarindu
BiodômedeMontréal.(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).
LabbéN,JuteauP,ParentS&VillemurR(2003)Bacterialdiversityinamarinemethanol-feddenitrificationreactoratthemontrealbiodome,Canada.Microbialecology46(1):12-21.
Doi:10.1007/s00248-002-1056-6LabbéN,LaurinV,JuteauP,ParentS&VillemurR(2007)Microbiological
communitystructureofthebiofilmofamethanol-fed,marinedenitrificationsystem,andidentificationofthemethanol-utilizingmicroorganisms.Microbialecology53(4):621-630.
Doi:10.1007/s00248-006-9168-zLabbéN,ParentS&VillemurR(2004)Nitratireductoraquibiodomusgen.nov.,sp.
nov.,anovelα-proteobacteriumfromthemarinedenitrificationsystemoftheMontrealBiodome(Canada).Internationaljournalofsystematicandevolutionarymicrobiology54(1):269-273.
Doi:LabelleMA,JuteauP,JolicoeurM,VillemurR,ParentS&ComeauY(2005)Seawater
denitrificationinaclosedmesocosmbyasubmergedmovingbedbiofilmreactor.Waterresearch39(14):3409-3417.
Doi:10.1016/j.watres.2005.06.001

80
LaurinavicheneT&TsygankovA(2016)DifferenttypesofH2photoproductionbystarch-utilizingco-culturesofClostridiumbutyricumandRhodobactersphaeroides.InternationalJournalofHydrogenEnergy41(31):13419-13425.
Doi:Le-ClechP,ChenV&FaneTA(2006)Foulinginmembranebioreactorsusedin
wastewatertreatment.Journalofmembranescience284(1):17-53. Doi:LiangX,LinL,YeY,GuJ,WangZ,XuL,JinY,RuQ&TianG(2015)Nutrientremoval
efficiencyinarice-strawdenitrifyingbioreactor.Bioresourcetechnology198:746-754.
Doi:http://dx.doi.org/10.1016/j.biortech.2015.09.083MaraziotiC,KornarosM&LyberatosG(2003)Kineticmodelingofamixedculture
ofPseudomonasdenitrificansandBacillussubtilisunderaerobicandanoxicoperatingconditions.Waterresearch37(6):1239-1251.
Doi:MartineauC,MauffreyF&VillemurR(2015)Comparativeanalysisofdenitrifying
activitiesofHyphomicrobiumnitrativorans,Hyphomicrobiumdenitrificans,andHyphomicrobiumzavarzinii.Appliedandenvironmentalmicrobiology81(15):5003-5014.
Doi:MartineauC,VilleneuveC,MauffreyF&VillemurR(2013)Hyphomicrobium
nitrativoranssp.nov.,isolatedfromthebiofilmofamethanol-feddenitrificationsystemtreatingseawaterattheMontrealBiodome.Internationaljournalofsystematicandevolutionarymicrobiology63(Pt10):3777-3781.
Doi:10.1099/ijs.0.048124-0MauffreyF,MartineauC&VillemurR(2015)ImportanceoftheTwoDissimilatory
(Nar)NitrateReductasesintheGrowthandNitrateReductionoftheMethylotrophicMarineBacteriumMethylophaganitratireducenticrescensJAM1.Frontiersinmicrobiology6.
Doi:MinK&RickardA(2009)Coaggregationbythefreshwaterbacterium
Sphingomonasnatatoriaaltersdual-speciesbiofilmformation.Appliedandenvironmentalmicrobiology75(12):3987-3997.
Doi:MincerTJ&AicherAC(2016)Methanolproductionbyabroadphylogeneticarrayof
marinephytoplankton.PloSone11(3):e0150820. Doi:MustakhimovI,ReshetnikovA,FedorovD,KhmeleninaV&TrotsenkoYA(2012)
RoleofEctRastranscriptionalregulatorofectoinebiosynthesisgenesinMethylophagathalassica.Biochemistry(Moscow)77(8):857-863.
Doi:MustakhimovI,ReshetnikovA,KhmeleninaV&TrotsenkoYA(2009)EctR—anovel
transcriptionalregulatorofectoinebiosynthesisgenesinthehaloalcaliphilicmethylotrophicbacteriumMethylophagaalcalica.DokladyBiochemistryandBiophysics.Springer,p305-308.

81
PakulaR&FreemanA(1996)Anewcontinuousbiofilmbioreactorforimmobilizedoil-degradingfilamentousfungi.Biotechnologyandbioengineering49(1):20-25.
Doi:PauletaSR,Dell’AcquaS&MouraI(2013)Nitrousoxidereductase.Coordination
ChemistryReviews257(2):332-349. Doi:http://dx.doi.org/10.1016/j.ccr.2012.05.026PlakunovVK,ZhurinaMV&BeliaevSS(2008)[Resistanceofthepetroleum-
oxidizingmicroorganismDietziasp.tohyperosmoticshockinreconstitutedbiofilms].Mikrobiologiia77(5):581-589.
Doi:PriemeA,BrakerG&TiedjeJM(2002)Diversityofnitritereductase(nirKandnirS)
genefragmentsinforesteduplandandwetlandsoils.Appliedandenvironmentalmicrobiology68(4):1893-1900.
Doi:RoedigerM(1983)Activatedsludgesystem.(GooglePatents).SauerT&GalinskiEA(1998)Bacterialmilking:Anovelbioprocessforproductionof
compatiblesolutes.Biotechnologyandbioengineering59(1):128. Doi:TakakiK,FushinobuS,KimS-W,MiyaharaM,WakagiT&ShounH(2008)
StreptomycesgriseusenhancesdenitrificationbyRalstoniapickettiiK50,whichispossiblymediatedbyhistidineproducedduringco-culture.Bioscience,biotechnology,andbiochemistry72(1):163-170.
Doi:VandePas-SchoonenK,Schalk-OtteS,HaaijerS,SchmidM,DenCampHO,StrousM,
KuenenJG&JettenM(2005)Completeconversionofnitrateintodinitrogengasinco-culturesofdenitrifyingbacteria.BiochemicalSocietytransactions33(1):205-209.
Doi:VanHoudtR&MichielsCW(2005)Roleofbacterialcellsurfacestructuresin
Escherichiacolibiofilmformation.Researchinmicrobiology156(5):626-633. Doi:VilleneuveC(2012)Caractérisationetclassificationdesnouvellesespèces
MethylophaganitratireducenticrescensetMethylophagafrappieri..(UniversitéduQuébec,InstitutNationaldelaRechercheScientifique).
VilleneuveC,MartineauC,MauffreyF&VillemurR(2013)Methylophaganitratireducenticrescenssp.nov.andMethylophagafrappierisp.nov.,isolatedfromthebiofilmofthemethanol-feddenitrificationsystemtreatingtheseawaterattheMontrealBiodome.Internationaljournalofsystematicandevolutionarymicrobiology63(Pt6):2216-2222.
Doi:10.1099/ijs.0.044545-0WangX,ChangVW&TangCY(2016)Osmoticmembranebioreactor(OMBR)
technologyforwastewatertreatmentandreclamation:Advances,challenges,andprospectsforthefuture.JournalofMembraneScience504:113-132.
Doi:

82
YeY,LingN,JiaoR,WuQ,HanY&GaoJ(2015)EffectsofcultureconditionsonthebiofilmformationofCronobactersakazakiistrainsanddistributionofgenesinvolvedinbiofilmformation.LWT-FoodScienceandTechnology62(1,Part1):1-6.
Doi:http://dx.doi.org/10.1016/j.lwt.2015.01.035ZaprasisA,BrillJ,ThüringM,WünscheG,HeunM,BarzantnyH,HoffmannT&
BremerE(2013)OsmoprotectionofBacillussubtilisthroughimportandproteolysisofproline-containingpeptides.Appliedandenvironmentalmicrobiology79(2):576-587.
Doi:ZumftWG(1997)Cellbiologyandmolecularbasisofdenitrification.Microbiology
andmolecularbiologyreviews:MMBR61(4):533-616. Doi:


















