
Reisión de la seguridad amiental de la proteína Cry1F -...
15
Revisión de la seguridad ambiental de la proteína Cry1F Center for Environmental Risk Assessment, ILSI Research Foundation 1156 Fifteenth Street N.W., Washington D.C. 20005-1743 EE. UU. 24 de mayo de 2013 Palabras clave Cry1F, proteína insecticida cris- talina, toxina binaria, Bacillus thuringiensis, resistencia a los insectos, genéticamente mo- dificado, evaluación de riesgo ambiental Copyright © ILSI Research Foundation 2013 El presente trabajo está autorizado en virtud de la licencia Creative Commons Attribution 3.0 Unported License. Para ver una copia de esa licencia, visite http:// creativecommons.org/licenses/ by/3.0/ o envíe una carta a Creative Commons, 171 Second Street, Suite 300, San Francisco, California, 94105, EE. UU. INTRODUCCIÓN Este documento provee un análisis exhaustivo de la información y los datos relevantes para la evaluación del riesgo ambiental de Cry1F, una familia de proteínas codificadas por genes aislados deBacillus thuringiensis (Bt), y presenta un informe de síntesis de la seguridad ambiental de esas proteínas cuando se las produce en plantas de algodón (Gossypium hirsutum) y maíz (Zea mays) genéticamente modificadas (GM). Todas las fuentes de información aquí revisadas están disponibles públicamente e incluyen expedientes presentados a las autoridades regulatorias; documentos de decisiones preparados por las autoridades regulatorias; descripciones del producto preparadas por los desarrolladores de productos y bibliografía revisada por pares. Las evaluaciones de riesgo ambiental relacionadas con la siembra de cultivos GM se realizan caso por caso y consideran la biología de la planta, las características de los transgenes y las proteínas codificadas, el fenotipo que conceden los transgenes, los usos previstos del cultivo y la naturaleza del ambiente en que se introducirá la planta. Dichas evaluaciones, que consideran tanto los posibles riesgos como los niveles de exposición, por lo general implican comparaciones con una línea progenitora sin transformar o con una isolínea muy relacionada (Craig, Tepfer, Degrassi y Ripandelli, 2008; OCDE, 2007). El propósito de estas comparaciones es identificar los posibles riesgos que la planta GM podría presentar más allá de los ya aceptados, al cultivarse plantas similares no GM en el ambiente. Se evalúan las consecuencias de esos riesgos, si los hay. Diversas autoridades regulatorias han efectuado evaluaciones de riesgo ambiental en variedades de cultivos GM que producen Cry1F. La Tabla 1 muestra el estado 1 actual de las aprobaciones regulatorias para la liberación al ambiente del evento de algodón DAS- 1 Es posible que las regulaciones requieran la renovación periódica de los registros de pesticidas. Por ejemplo, el estado actual de los registros de USEPA se puede consultar en http://www.epa.gov/oppbppd1/biopesticides/pips/pip_list.htm. 24236-5 que contiene Cry1F y eventos de maíz DAS- 01507-1 y DAS-06275-8 que contienen Cry1F. 2 En algunos países, se otorga una aprobación regulatoria por separado cuando un evento ya aprobado se combina con otros eventos GM en conjunto (Que et al., 2010). La tabla muestra la fecha de la primera aprobación otorgada para el evento. Tabla 1. Aprobaciones regulatorias para la liberación al ambiente de variedades de algodón y maíz GM que contienen Cry1F (al 28 de febrero de 2013). País Algodón DAS- 24236-5 Maíz DAS- 01507-1 Maíz DAS- 06275-8 Argentina 2005 Brasil 2009 2008 Canadá 2002 2006 Colombia 2007 Estados Unidos 2004 2001 2004 Honduras 2009 Japón 2002 2008 Paraguay 2012 Uruguay 2011 2 Muchas otras autoridades regulatorias también han aprobado el uso de algodón y maíz Cry1F para alimentos destinados a seres humanos y animales. Para obtener más información, consulte http://cera-gmc.org/index. php?action=gm_crop_database.
Transcript of Reisión de la seguridad amiental de la proteína Cry1F -...

Revisión de la seguridad ambiental de la proteína Cry1FCenter for Environmental Risk Assessment, ILSI Research Foundation1156 Fifteenth Street N.W., Washington D.C. 20005-1743 EE. UU.
24 de mayo de 2013
Palabras clave
Cry1F, proteína insecticida cris-talina, toxina binaria, Bacillus thuringiensis, resistencia a los insectos, genéticamente mo-dificado, evaluación de riesgo ambiental
Copyright © ILSI Research Foundation 2013El presente trabajo está autorizado en virtud de la licencia Creative Commons Attribution 3.0 Unported License. Para ver una copia de esa licencia, visite http://creativecommons.org/l icenses/by/3.0/ o envíe una carta a Creative Commons, 171 Second Street, Suite 300, San Francisco, California, 94105, EE. UU.
INTRODUCCIÓN
Este documento provee un análisis exhaustivo de la información y los datos relevantes para la evaluación del riesgo ambiental de Cry1F, una familia de proteínas codificadas por genes aislados deBacillus thuringiensis (Bt), y presenta un informe de síntesis de la seguridad ambiental de esas proteínas cuando se las produce en plantas de algodón (Gossypium hirsutum) y maíz (Zea mays) genéticamente modificadas (GM). Todas las fuentes de información aquí revisadas están disponibles públicamente e incluyen expedientes presentados a las autoridades regulatorias; documentos de decisiones preparados por las autoridades regulatorias; descripciones del producto preparadas por los desarrolladores de productos y bibliografía revisada por pares.
Las evaluaciones de riesgo ambiental relacionadas con la siembra de cultivos GM se realizan caso por caso y consideran la biología de la planta, las características de los transgenes y las proteínas codificadas, el fenotipo que conceden los transgenes, los usos previstos del cultivo y la naturaleza del ambiente en que se introducirá la planta. Dichas evaluaciones, que consideran tanto los posibles riesgos como los niveles de exposición, por lo general implican comparaciones con una línea progenitora sin transformar o con una isolínea muy relacionada (Craig, Tepfer, Degrassi y Ripandelli, 2008; OCDE, 2007). El propósito de estas comparaciones es identificar los posibles riesgos que la planta GM podría presentar más allá de los ya aceptados, al cultivarse plantas similares no GM en el ambiente. Se evalúan las consecuencias de esos riesgos, si los hay.
Diversas autoridades regulatorias han efectuado evaluaciones de riesgo ambiental en variedades de cultivos GM que producen Cry1F. La Tabla 1 muestra el estado1 actual de las aprobaciones regulatorias para la liberación al ambiente del evento de algodón DAS-
1 Es posible que las regulaciones requieran la renovación periódica de los registros de pesticidas. Por ejemplo, el estado actual de los registros de USEPA se puede consultar en http://www.epa.gov/oppbppd1/biopesticides/pips/pip_list.htm.
24236-5 que contiene Cry1F y eventos de maíz DAS-01507-1 y DAS-06275-8 que contienen Cry1F.2 En algunos países, se otorga una aprobación regulatoria por separado cuando un evento ya aprobado se combina con otros eventos GM en conjunto (Que et al., 2010). La tabla muestra la fecha de la primera aprobación otorgada para el evento.
Tabla 1. Aprobaciones regulatorias para la liberación al ambiente de variedades de algodón y maíz GM que contienen Cry1F (al 28 de febrero de 2013).
País AlgodónDAS-
24236-5
MaízDAS-
01507-1
MaízDAS-
06275-8
Argentina 2005
Brasil 2009 2008
Canadá 2002 2006
Colombia 2007
Estados Unidos 2004 2001 2004
Honduras 2009
Japón 2002 2008
Paraguay 2012
Uruguay 2011
2 Muchas otras autoridades regulatorias también han aprobado el uso de algodón y maíz Cry1F para alimentos destinados a seres humanos y animales. Para obtener más información, consulte http://cera-gmc.org/index.php?action=gm_crop_database.

2 3
Investigaciones adicionales acerca de la base para la susceptibilidad de la toxina Bt indican que la unión de Cry1F a las membranas celulares del intestino medio puede ser necesaria, pero no suficiente para que un insecto sea susceptible a Cry1F (Ballester et al., 1999; Coates et al., 2011). Otros mecanismos de resistencia podrían involucrar el proceso de activación de la protoxina, degradación de la toxina activa, o un mecanismo indeterminado (Jurat-Fuentes y Adang, 2006; Eliseu J G Pereira et al., 2010).
Numerosos estudios han ayudado a establecer el espectro de actividad de Cry1F. Aparentemente, es tóxica para las especies lepidópteras pero no para las coleópteras (Balog, Szenasi, Szekeres y Palinkas, 2011; Oppert et al., 2010). Las especies susceptibles incluyen el gusano bellotero del tabaco (Heliothis virescens), gardama (Spodoptera exigua), falsa oruga medidora (Pseudoplusia includens), gusano del algodón (Helicoverpa zea), gusano cogollero (Spodoptera frugiperda), barrenador menor del maíz (Elasmopalpus lignosellus), polilla de la cera (Galleria mellonella), y taladro del maíz (Ostrinia nubilalis) (Adamczyk y Gore, 2004; Adamczyk et al., 2008; Ballester et al., 1999; Blanco et al., 2008, 2010; Buntin, 2008; Chambers et al., 1991; Hanley, Huang y Pett, 2003; Iracheta et al., 2000; Siebert, Babcock et al., 2008; Tindall, Siebert, Leonard, All y Haile, 2009; USEPA, 2001).
Modificaciones a los genes que codifican Cry1F en algodón y maíz GM
Algodón: La secuencia cry1F utilizada para producir el evento de algodón DAS-24236-5 se obtuvo de Cry1F de B. thuringiensis subsp. aizawai cepa PS811. Los nucleótidos que codifican la parte C-terminal de Cry1F, que normalmente se extrae durante la activación de la protoxina, se reemplazaron por nucleótidos 1811-1917 de cry1C y nucleótidos 1918-3447 de cry1Ab. Los aminoácidos que codifican estos nucleótidos se extraen durante la activación de la protoxina. Se preparó una versión sintética de esta secuencia utilizando codones óptimos para la expresión de proteínas en plantas (Murray, Lotzer y Eberle, 1989). Estos reemplazos de codones no alteraron la secuencia final de aminoácidos de la proteína activa Cry1F.6
Maíz: La secuencia cry1F utilizada para producir los eventos de maíz DAS-01507-1 y DAS-06275-8 se obtuvieron de cry1F de B. thuringiensis subsp. aizawai cepa PS811. Se preparó una versión sintética de esta
6 La secuencia de ADN que se utilizó en el proceso de transformación original, que resultó en el aislamiento del evento DAS-24236-5, también contenía el gen pat, que confiere tolerancia a los herbicidas de glufosinato de amonio. Para obtener un análisis completo de la seguridad ambiental de la proteína PAT, consulte “A Review of the Environmental Safety of the PAT Protein” (CERA, 2011).
ORIGEN Y FUNCIÓN DE LA PROTEÍNA Cry1F
Bacillus thuringiensis y la proteína insecticida de Cry1F
Bacillus thuringiensis es una bacteria en forma de bastón, gram positiva, capaz de formar endosporas de larga vida. A menudo se la llama bacteria del suelo, si bien está omnipresente en el ambiente (ver, por ejemplo, Apaydin, Çınar, Turanli, Harsa y Güneş, 2008; Martínez y Caballero, 2002; Seifinejad, Jouazni, Hosseinzadeh y Abdmishani, 2008). Se ha estudiado la especie con profundidad y se la ha utilizado comercialmente durante varios años debido a su capacidad de sintetizar proteínas con propiedades pesticidas (Hofte y Whiteley, 1989; OCDE, 2007; Schnepf et al., 1998; van Frankenhuyzen, 2009). Los preparados de aislados naturales de B. thuringiensis se usaron por primera vez como insecticida comercial en Francia en 1938 y la subespecie kurstaki de B. thuringiensis ha estado registrada en USEPA desde 1961 (USEPA, 1998). Los preparados microbianos de B. thuringiensis actualmente están aprobados para ser utilizados en todo el mundo, incluso en Australia, Canadá, la Unión Europea y los Estados Unidos (APVMA, 2013; DGSANCO, 2013; Health Canada, 2008; Kumar, Sharma y Malik, 1996; Schnepf et al., 1998). Debido a que esos preparados derivan de células cultivadas de bacterias Bt, contienen una mezcla compleja de las sustancias pesticidas producidas por la cepa específica de Bt usada.
Varios cientos de sustancias pesticidas fueron aisladas de cultivos de Bt (Crickmore et al., 2012) y estas sustancias muestran una formidable diversidad en su estructura química, modo de acción y especificidad en el blanco (Hofte y Whiteley, 1989; OCDE, 2007; Pigott y Ellar, 2007; Schnepf et al., 1998; Vachon, Laprade y Schwartz, 2012; van Frankenhuyzen, 2009). Incluyen compuestos fungicidas, ß-exotoxinas, proteínas Cyt (citolíticas), proteínas insecticidas vegetativas (Vip) y las ß-endotoxinas,3 un grupo que incluye las proteínas insecticidas Cry (cristalinas) (Hofte y Whiteley, 1989; OCDE, 2007; Schnepf et al., 1998). Esas sustancias pueden interactuar entre sí para influenciar el espectro de actividad y toxicidad de los preparados de bacterias individuales (OCDE, 2007; Schnepf et al., 1998).4 Las proteínas Cry han sido estudiadas con profundidad y utilizadas ampliamente en agricultura por su capacidad para causar la rotura celular en los tractos digestivos de las plagas de insectos, lo cual causa la interrupción de la alimentación y finalmente la muerte de los insectos (Mendelsohn, Kough, Vaituzis y Matthews, 2003; OCDE, 2007).
Un informe, publicado en 1991, describe el descubrimiento de una nueva proteína 133.6 kDa, en B. thuringiensis subsp. aizawai. Esta proteína es altamente tóxica para los lepidópteros Heliothis virescens y Ostrinia nubilalis, y moderadamente tóxica paraSpodoptera exigua (Chambers et al., 1991; Gaspers et al., 2011), pero no es tóxica para las especies de coleópteros (Oppert, Ellis y Babcock, 2010). La proteína es termolábil, altamente digestible por pepsina, y no glicosilada (Ladics, Bardina, Cressman, Mattsson y Sampson, 2006). La secuencia proteica la ubicó
3 También denominado thurigiensin (Liu et al., 2010; OCDE, 2007).
4 La actividad insecticida de los preparados de bacterias Bt se debe a una combinación de múltiples toxinas y calidades de las esporas bacterianas, que puede tener un impacto en la selectividad y el rango del receptor (Schnepf et al., 2005; Tabashnik, 1992). Por lo tanto, el espectro de actividad de los rociadores hechos de cultivos bacterianos Bt puede diferir del espectro de actividad de las proteínas Bt individuales producidas por una planta GM (OCDE 2007).
en el grupo Cry1 de proteínas Bt, pero poseía, en el mejor de los casos, solo 72 % de similitud de secuencia con las proteínas de los subgrupos existentes: Cry1A, Cry1B, Cry1C, Cry1D y Cry1E (Chambers et al., 1991; Crickmore et al., 2012) Asimismo, la región N-terminal de la nueva proteína Cry1, que típicamente codifica el componente insecticida de otras proteínas del grupo Cry1, era como máximo 52 % similar a las secuencias N-terminal de otras proteínas Cry1. Se estableció que la proteína no era miembro de ninguno de los subgrupos existentes, y se designó un nuevo subgrupo, Cry1F, para esta proteína. A la fecha, se han designado 11 proteínas pertenecientes al subgrupo Cry1F (Crickmore et al., 2012).
Mecanismo de la actividad insecticida de Cry1F
Al igual que muchas otras proteínas Cry, el mecanismo de actividad de Cry1F comienza con una escisión enzimática de la protoxina 130 kDa para liberar una toxina clave 65 kDa (Gao et al., 2006) En este proceso, se escinde un péptido corto del extremo N-terminal y se extrae casi la mitad del péptido del extremo C-terminal. Se cree que estos terminales participan en la formación de cuerpos de inclusión cristalinos (Gao et al., 2006). La proteína restante posee dos regiones fuertemente hidrofóbicas que, según se cree, participan en la unión con las membranas del intestino medio (Chambers et al., 1991; Jurat-Fuentes y Adang, 2001). Una vez unidas, las moléculas de las toxinas forman oligómeros, que crean poros en la membrana y provocan desestabilización osmótica y muerte celular (Jurat-Fuentes y Adang, 2001).
Se ha puesto mucho énfasis en comprender la naturaleza de la unión de membranas en el grupo de proteínas Cry15 para establecer si múltiples toxinas comparten el mismo sitio de unión (un factor que podría afectar el desarrollo de resistencia cruzada a las proteínas Bt en una población de insectos). Por ejemplo, Cry1F y Cry1A se unen al mismo receptor del intestino medio en Plutella xylostella (Ballester, Granero, Tabashnik, Malvar y Ferré, 1999), lo cual posiblemente contribuya a la resistencia cruzada observada en las dos toxinas (González-Cabrera, Herrero y Ferré, 2001). Del mismo modo, existe un sitio de unión compartido por Cry1A y Cry1F en Heliothis virescens (Blanco et al., 2008; Jurat-Fuentes y Adang, 2001), en Trichoplusia ni (Iracheta, Pereyra-Alférez, Galán-Wong y Ferré, 2000), y en Spodoptera frugiperda (Sena, Hernández-Rodríguez y Ferré, 2009). Sin embargo, en algunas especies Cry1F no interactúa con el receptor conocido de Cry1Ac en Heliothis virescens (Jurat-Fuentes y Adang, 2001, 2006), y la resistencia cruzada entre las proteínas Cry1F y Cry1A es baja en algunas especies de insectos (Pereira, Lang, Storer y Siegfried, 2008; Storer et al., 2010). Aparentemente, la presencia y la naturaleza de sitios de unión compartidos para las proteínas Cry1 difiere entre las especies de insectos (Blanco et al., 2010; Ferré y Rie, 2002; Pereira, Siqueira, Zhuang, Storer y Siegfried, 2010; Tabashnik et al., 2003).
5 Debido a las mayores diferencias en la secuencia de la proteína, existe una menor probabilidad de sitios de unión compartidos entre las proteínas Cry1F y Cry en otros grupos (Gouffon, Van Vliet, Van Rie, Jansens y Jurat-Fuentes, 2011; comparación de la unión de Cry1F y Cry2Ae).
secuencia borrando los codones que codifican los 569 aminoácidos de la parte C-terminal, que normalmente se escindirían durante la activación de la proteína. Asimismo, la secuencia empleó codones óptimos para la expresión de proteínas en plantas de maíz (Murray et al., 1989). Estos reemplazos de codones no alteraron la secuencia final de aminoácidos de la proteína activa Cry1F. Se agregó un residuo de leucina en la posición 604 de la proteína, lo cual creó un sitio de restricción XhoI para facilitar la clonación genética.7
La Tabla 2 brinda descripciones de los elementos genéticos utilizados en la producción de eventos de algodón y maíz Cry1F.
Expresión de Cry1F en algodón y maíz GM resistentes a insectos
Los niveles de expresión transgénica en una planta GM modificada pueden ser influenciados por muchos factores relacionados con el proceso de transformación genética, entre ellos el tipo de secuencias de promotor y terminador utilizadas, también la ubicación cromosómica donde el transgen se incorporó al genoma. Los niveles de expresión también pueden estar modificados por el tipo de tejido que se tome de la muestra, la edad de la planta al momento en que se tomó la muestra y las condiciones ambientales en que crecía la planta (Siebert et al., 2009).
7 La secuencia de ADN que se utilizó en el proceso de transformación original, que resultó en el aislamiento del evento DAS-01507-1, también contenía el gen pat, que confiere tolerancia a los herbicidas de glufosinato de amonio. La secuencia de ADN que se utilizó en el proceso de transformación original, que resultó en el aislamiento del evento DAS-06275-8, también contenía el gen bar, que confiere tolerancia a los herbicidas de glufosinato de amonio. Para obtener un análisis completo de la seguridad ambiental de las proteínas PAT y BAR, consulte “A Review of the Environmental Safety of the PAT Protein” (CERA, 2011).
Tabla 2. Elementos genéticos utilizados en la producción de variedades de algodón y maíz GM resistentes a insectos (USDA, 2000, 2003, 2004a)
Elemento genético
AlgodónDAS-24236-5
MaízDAS-01507-1
MaízDAS-06275-8
Promotor Promotor de manopina sintasa a partir de Agrobacterium tumefaciens, cepa LBA 4404 pTi15955, que icluye copias del potenciador de octipina sintasa a partir de pTiAch5
Promotor ubicuo (más intrón y secuencia no traducida de 5’) a partir de Zea mays
Promotor ubicuo (más intrón y secuencia no traducida de 5’) a partir de Zea mays
Gen Versión sintética, optimizada para plantas, en versión de longitud completa de Cry1F a partir de B.t. subsp. aizawai. Los nucleótidos 1-1810 de la secuencia de codificación codifican la porción tóxica de Cry1Fa2. Los nucleótidos 1811-1917 codifican una porción de la protoxina Cry1C. Los nucleótidos 1918-3447 codifican una porción de la protoxina Cry1Ab.
Versión optimizada para plantas de Cry1F truncada a partir de B.t. subsp. aizawai
Versión de maíz optimizada para plantas de Cry1F truncada a partir de B.t. subsp. aizawai
Terminador Terminador a partir de Agrobacterium tumefaciens cepa LBA 4404 pTi15955
Terminador a partir de Agrobacterium tumefaciens cepa LBA 4404 pTi15955
Secuencia de terminador a partir del inhibidor de proteinasa II Solanum tuberosum
4 5
Los datos obtenidos de los ensayos por inmunoabsorción ligados a enzimas (ELISA), que muestran niveles de expresión de la proteína Cry1F en eventos de algodón y maíz GM, están disponibles en presentaciones a las autoridades regulatorias de acceso público y documentos de decisión asociados a procesos de autorización de las autoridades regulatorias. Se recolectaron muestras de diferentes tipos de tejidos y en estadios de crecimiento múltiples de plantas cultivadas en diferentes ubicaciones para obtener datos representativos del rango típico de la expresión de la proteína. Las Tablas 3 y 4 presentan los máximos valores informados de la expresión de Cry1F en plantas de algodón y maíz GM, respectivamente. Los datos de expresión de proteínas pueden utilizarse para calcular la exposición potencial de varios organismos del ambiente a Cry1F cuando se cultivan plantas de algodón y maíz que producen Cry1F. Los datos de expresión de proteínas disponibles actualmente para Cry1F por el evento de algodón DAS-24236-5 y por los eventos de maíz DAS-01507-1 y DAS-06275-8 utilizados en forma individual y apilados con otros eventos GM se presentan en el Anexo I.
Tabla 3. Máximas concentraciones de proteína informadas de Cry1F en varios tejidos de plantas del evento de algodón GM DAS-24236-5 (USDA, 2003).
Tejido Cry1F ng/mg de peso seco de tejido1
Hoja joven (3-6 semanas) 16,8
Hoja terminal 20,7
Flor 12,3
Botón floral 9,4
Cápsula (temprano) 9,2
Planta entera (plántula) 23,3
Planta entera (polinización) 38,4
Planta entera (defoliación) 37,6
Raíz (plántula) 2,3
Raíz (polinización) 0,62
Raíz (defoliación) 1,7
Polen 1,1
Néctar No detectado
Semilla 8,2
1 Resultados basados en el peso de tejido fresco de polen, néctar y semilla.
Tabla 4 Máximas concentraciones informadas de Cry1F en varios tejidos de plantas de los eventos de maíz GM DAS-06275-8 y DAS-01507-1 (USDA, 2004a).
Tejido Etapa de crecimiento 1
Línea 6275ng/mg peso seco
de tejido
Línea 1507ng/mg peso seco
de tejido
Hoja V9 23,8 24
Planta entera V9 7,87 6,8
Planta entera R1 9,57 4,7
Polen R1 4,6 27,2
Tallo R1 16,4 10,3
Forraje R4 7,77 3,2
Planta entera Senescencia 3,07 2,4
1 V9 – el collar de la 9.a hoja es visible R1 - Emergencia de estigmas, panícula emitiendo polen R4 - Masa blanda, mayoría de granos pastosos con semisólidos Madurez - Humedad del grano de 25 a 35 %
PRUEBA EN ORGANISMOS NO BLANCO E IMPACTOS DE LA EXPOSICIÓN A LA PROTEÍNA Cry1F
La toxina Cry1F posee propiedades insecticidas contra ciertas especies de insectos lepidópteros al expresarse en plantas de algodón y maíz. La toxina ataca a plagas de insectos lepidópteros, lo cual reduce el daño alimenticio (Adamczyk y Gore, 2004; Adamczyk et al., 2008; Buntin, 2008; CFIA, 2002, 2005, 2006; EFSA, 2009a, 2009b; FSANZ, 2003, 2004; JBCH, 2004, 2006a, 2006b, 2006c, 2006d, 2006e, 2009, 2010; PDOA, 2006; Siebert, Babcock, et al., 2008; Siebert, Nolting, et al., 2008; USDA, 2000, 2001, 2003, 2004a, 2004b, 2004c; USEPA, 2001, 2005). Se denominan organismos no blanco (non-target organisms, NTO) los organismos del ambiente que no son plagas de maíz pero que están directa o indirectamente expuestos a Cry1F.
Las evaluaciones de impactos sobre los NTO incluyen la revisión de los datos entregados por el desarrollador del producto a las agencias- regulatorias para demostrar que los NTO expuestos a Cry1F, directa o indirectamente, no están dañados. La evaluación de riesgos de los NTO comienza normalmente con una determinación de los organismos que tienen posibilidad de estar directa o indirectamente expuestos a Cry1F. A menudo, se otorga una consideración particular a los NTO que tienen funciones beneficiosas para el ambiente, como los polinizadores o los enemigos naturales de las plagas agrícolas. Las autoridades regulatorias pueden prestarle atención especial a los NTO que se designaron como especies amenazadas o en peligro o especies con valor cultural reconocido. Dichas especies, o las especies representativas de grupos taxonómicos o funcionales a proteger, se evalúan para determinar si la exposición a Cry1F puede tener impactos adversos importantes.
Las evaluaciones de los posibles impactos sobre los NTO y las decisiones de las autoridades regulatorias informadas por la evaluación están fundamentadas en la bien documentada y larga historia de la evaluación de formulaciones de insecticidas químicos, incluyendo las formulaciones microbianas de B. thuringiensis (Carstens et al., 2012; Romeis et al., 2008, 2013; Sanvido et al., 2012; USEPA, 2007). El abordaje “por
niveles” para evaluar los impactos de los pesticidas químicos sobre los NTO se usa efectivamente desde hace muchos años; científicos y reguladores también determinaron que los estudios por niveles son apropiados para la evaluación de impactos posibles de los cultivos GM sobre los NTO (Duan, Lundgren, Naranjo y Marvier, 2010; Dutton, Romeis y Bigler, 2003; EFSA, 2006a; Garcia-Alonso et al., 2006; Raybould, 2006; Romeis et al., 2008, 2013; USEPA, 2007, 2011). Normalmente, los primeros estudios por niveles involucran la exposición de NTO o especies sustitutas a grandes concentraciones de pesticidas, en condiciones controladas de laboratorio. Esos estudios identifican a las especies que se ven muy afectadas por los pesticidas. Dichos efectos, cuando se encuentran, pueden requerir de un análisis mayor a un nivel superior. Las primeras pruebas por niveles también identifican aquellos NTO que no son afectados por la proteína pesticida y para los cuales no es necesario realizar pruebas en niveles superiores. Las pruebas en niveles superiores también pueden ser apropiadas cuando los resultados de las pruebas anteriores son inconclusos. Las pruebas en niveles superiores involucran mayores niveles de complejidad y de condiciones reales de evaluación (EFSA, 2006a; Garcia-Alonso et al., 2006; Romeis et al., 2008; USEPA, 2007, 2011).
La posibilidad de daño a los NTO a partir de la exposición a Cry1F fue considerada en las evaluaciones de riesgo realizadas por diversas autoridades regulatorias (CFIA, 2002, 2005, 2006; EFSA, 2009a, 2009b; FSANZ, 2003, 2004; JBCH, 2006c, 2006d, 2006e, 2009, 2010, 2004, 2006a, 2006b; PDOA, 2006; USDA, 2000, 2001, 2003, 2004a, 2004b, 2004c; USEPA, 2001, 2005). Los datos recabados de ensayos de laboratorio y de campo de algodón y maíz GM que producen Cry1F y presentados a las autoridades regulatorias establecieron que la proteína Cry1F es activa específicamente contra el subconjunto de plagas lepidópteras que se alimentan de las partes aéreas de las plantas de algodón y maíz y es inocua para las especies vertebradas y otros NTO (CFIA, 2002, 2005, 2006; EFSA, 2009a, 2009b; FSANZ, 2003, 2004; JBCH, 2006c, 2006d, 2006e, 2009, 2010, 2004, 2006a, 2006b; PDOA, 2006; USDA, 2000, 2001, 2003, 2004a, 2004b, 2004c; USEPA, 2001, 2005).
Vías de exposición ambiental
La exposición directa ocurre cuando los NTO se alimentan de tejidos de cultivos vivos que expresan Cry1F o de desechos de cultivos que se encuentran arriba o debajo del suelo. La exposición indirecta resulta de la depredación de un organismo a otro que estuvo directamente expuesto a Cry1F (J.-C. Tian et al., 2012). Además del consumo directo de las plantas de algodón o maíz GM, las autoridades regulatorias pueden considerar otras vías posibles de exposición indirecta a la toxina Cry1F: exposición a la toxina en el polen, exposición a la toxina depositada en el suelo por el material de plantas en descomposición y exposición a especies depredadoras que consumen herbívoros que se alimentaron de las plantas de maíz GM (CFIA, 2002, 2005, 2006; EFSA, 2009a, 2009b; FSANZ, 2003, 2004; JBCH, 2006c, 2006d, 2006e, 2009, 2010, 2004, 2006a, 2006b; PDOA, 2006; USDA, 2000, 2001, 2003, 2004a, 2004b, 2004c; USEPA, 2001, 2005). Las autoridades regulatorias pueden considerar datos de expresión de la proteína para establecer las posibles vías y los posibles niveles de exposición. Por ejemplo, los tejidos de plantas que producen poca Cry1F, o nada de ella, son poco proclives a plantear un riesgo a los NTO. (Consulte en las Tablas 3 y 4 y en el Anexo I los datos de los niveles de expresión en los tejidos de las variedades aprobadas de algodón y maíz.) Los datos presentados a las autoridades regulatorias
indican que Cry1F se degrada con rapidez una vez liberada del tejido de una planta en descomposición y no es probable que persista o se acumule en el ambiente del suelo (Herman, Wolt y Halliday, 2002; Shan, Embrey, Herman y McCormick, 2008; USDA, 2003).
Pruebas ecotoxicológicas de Cry1F en organismos no blanco
Se efectuaron pruebas ecotoxicológicas de Cry1F en NTO, en una variedad de organismos de prueba bien caracterizados que se utilizan típicamente para la prueba ecotoxicológica de pesticidas químicos, y los datos de estas pruebas fueron evaluados por las autoridades regulatorias en el transcurso de las evaluaciones del riesgo de la liberación al ambiente de variedades de algodón y maíz GM (Dutton et al., 2003; Raybould, 2007; Romeis et al., 2008; USEPA, 2007; Wolt et al., 2010). Debido a que Cry1F es tóxica para varias especies lepidópteras, las autoridades regulatorias en general han solicitado datos del impacto de Cry1F en los lepidópteros no blanco, tales como la mariposa monarca u otras especies lepidópteras de importancia local (Wolt, Conlan, and Majima, 2005). Las autoridades regulatorias también pueden solicitar datos sobre las especies polinizadoras representativas, es decir, las abejas melíferas; las especies representativas de artrópodos del suelo, tales como las lombrices de tierra, para demostrar que no existe un impacto significativo sobre estas especies de la exposición a Cry1F. Entre los organismos de prueba se incluyeron Apis mellifera (abeja melífera); Hippodamia convergens (catarina o mariquita) Chrysoperla carnea (crisopa); Danaus plexippus (mariposa monarca); Nasonia vitripennis (avispa parasitaria); Folsomia candida (colémbolo); Daphnia magna (crustáceo); y Eisenia foetida (lombriz de tierra). Los organismos de prueba fueron expuestos a niveles de Cry1F muchas veces superiores a los máximos niveles de exposición previstos de las concentraciones de tejido observadas de Cry1F en plantas de algodón y maíz GM (Ver Tablas 3 y 4). Ninguno de los organismos de prueba mostró una respuesta significativa a Cry1F (Hanley et al., 2003; Hellmich et al., 2001; J.-C. Tian et al., 2012, 2013; X.C. Tian et al., 2005; USDA, 2000, 2003; USEPA, 2001; Wolt et al., 2005) (Véase el Anexo II). Asimismo, se han efectuado pruebas toxicológicas en vertebrados y pruebas de equivalencia nutricional en Mus musculus (ratón); Oncorhynchus mykiss (trucha arcoiris); Gallus domesticus (gallina); Rattus norvegicus (rata); Sus domestica (cerdo); y Colinus virginianus (codorniz cotuí norteña) (Appenzeller, Malley, Mackenzie, Hoban y Delaney, 2009; Dryzga, Yano, Andrus y Mattsson, 2007; FSANZ, 2004; MacKenzie et al., 2007; Scheideler, Rice, Smith, Dana y Sauber, 2008; Stein et al., 2009; USDA, 2000, 2003; USEPA, 2001) (Véase el Anexo II).
Los resultados de las pruebas de Nivel 1 analizados más arriba indican que no sería necesario efectuar pruebas en niveles superiores desde el punto de vista de la regulación, ya que no se observaron efectos adversos;8 sin embargo, se realizaron estudios acerca de los efectos de Cry1F sobre las poblaciones naturales de NTO (Balog et al., 2011; Higgins et al., 2009; USDA, 2004c; USEPA, 2001). En estos estudios de campo no se encontraron diferencias significativas entre los artrópodos NTO en campos en los que se cultivaba maíz Cry1F y en campos en los que se cultivaba una variedad de maíz no GM.
8 La realización de estudios de campo se considera caso por caso, según el nivel de daño y exposición potenciales, y los objetivos se pueden modificar a medida que se acumulan información y experiencia (USEPA, 2007).

6 7
Las autoridades regulatorias han considerado el posible impacto de Cry1F en poblaciones naturales de NTO y determinaron que los efectos adversos sobre los NTO son poco probables por diversas razones. Primero, Cry1F tiene un espectro reducido de actividad pesticida. Segundo, en ensayos de laboratorio de Nivel I, que emplearon un rango de especies invertebradas presentes en los ecosistemas agrícolas de algodón y de maíz, o sustitutos para esas especies, se demostró que Cry1F no causa efectos observables significativos en esas especies. Tercero, en estudios de Nivel I también se demostró que Cry1F no tiene ningún efecto observable en especies vertebradas ni acuáticas representativas. Cuarto, los niveles de Cry1F utilizados en estos ensayos de Nivel I fueron mucho más altos que aquellos medidos en tejidos de algodón y maíz GM cultivados en el campo. Quinto, los estudios de campo de las variedades de maíz que producen Cry1F no muestran efectos adversos significativos sobre los escarabajos vagabundo, un atrópodo beneficioso no blanco (Balog et al., 2011). Sexto, al compararlo con el control de insectos mediante Cry1F, el control de insectos habitual, que utiliza pesticidas químicos, altera la diversidad de las especies de manera significativa y daña las especies no blanco (Higgins et al., 2009; USDA, 2003).9 En su conjunto, estos hallazgos indican que es poco probable que Cry1F tenga efectos adversos en las poblaciones naturales de organismos, excepto las plagas lepidópteras que son blanco de los cultivos que se propone controlar (Balog et al., 2011; CFIA, 2005, 2006; CTNBio, 2008, 2009a; EFSA, 2009a; Higgins et al., 2009; JBCH, 2006e, 2006c, 2006d; PDOA, 2006; J.-C. Tian et al., 2012; USDA, 2001, 2004b, 2004c; USEPA, 2005; Wolt et al., 2005).
ESTABLECIMIENTO Y PERSISTENCIA EN EL AMBIENTE DE PLANTAS DE ALGODÓN Y MAÍZ QUE EXPRESAN Cry1F
Biología de la especie de planta
La biología de la especie de planta no GM dentro del ambiente que la recibe es, por lo general, el punto de partida de las evaluaciones de riesgo ambiental de plantas GM (OCDE, 2003, 2007, 2008). Se puede utilizar información sobre la biología de la planta no GM para evaluar si una variedad GM de la planta puede convertirse en maleza, invasiva o dañina de cualquier otra forma para el ambiente. También puede brindar detalles acerca de las interacciones importantes entre la planta y otros organismos, que pueden ser importantes para considerar los posibles perjuicios. Al considerar la biología de la planta huésped, un evaluador de riesgo puede identificar los peligros posibles que pueden estar asociados con la expresión de la nueva proteína (por ejemplo, Cry1F) y luego puede evaluar la probabilidad de que dichos peligros se hagan realidad. Por ejemplo, el hecho de si una planta es una especie anual o perenne o si se autopoliniza o si la poliniza el viento, es relevante para evaluar la probabilidad de que la planta GM se establezca y persista fuera del cultivo (EFSA, 2006a; OCDE, 1992, 2003, 2007, 2008).
9 En este estudio también se encontró una dinámica similar para los atrópodos no blanco en campos de maíz Cry1F y maíz no Bt, tanto a nivel comunitario como de taxones individuales (Higgins et al., 2009).
Datos del fenotipo
La información acerca del fenotipo de las plantas GM que expresan Cry1F se recaba del laboratorio, el invernadero y los estudios de campo, y se presenta en las presentaciones a las autoridades regulatorias para: (1) identificar todo cambio intencional al fenotipo que pueda afectar la seguridad ambiental de la planta; y (2) para identificar todo cambio no intencional causado a la biología de la planta que pueda afectar la seguridad ambiental. En las presentaciones a las autoridades regulatorias y en las publicaciones de pares, los datos del fenotipo se han concentrado en las características de la planta que podrían contribuir a su supervivencia o persistencia (es decir, el potencial para convertirse en maleza), o que podrían afectar negativamente el rendimiento agrícola (por ejemplo, la susceptibilidad a enfermedades y los datos de rendimiento). Las observaciones del fenotipo tuvieron en cuenta el fenotipo resultante del carácter transgénico deseado, en este caso, la resistencia a la depredación de insectos mediada por Cry1F. Algunos de los datos recolectados son cuantitativos (por ejemplo, altura de la planta o porcentaje de germinación de la semilla), mientras que otros datos son cualitativos y de observación (por ejemplo, síntomas de susceptibilidad a enfermedades). Se observaron diferencias estadísticamente significativas entre las plantas de algodón (CTNBio, 2009a; JBCH, 2006b, 2006d; USDA, 2004b) o maíz GM (EFSA, 2009a; JBCH, 2004, 2006e; USDA, 2000, 2001, 2004a, 2004c) que expresan Cry1F y los controles, pero estas diferencias no fueron consistentes entre los sitios de la prueba de campo y estuvieron dentro del rango informado para las variedades de algodón y maíz no GM. Conjuntamente, los reguladores determinaron que los datos del fenotipo no respaldan la hipótesis de que la expresión de Cry1F haya tenido algún impacto no intencional en la morfología general o las características del fenotipo de las plantas de algodón y maíz, además de conferir resistencia a las plagas de insectos lepidópteros.
Potencial para convertirse en maleza en ambientes agrícolas
Algodón: El algodón cultivado carece de características agresivas o que supongan un crecimiento como de maleza, y por lo general, no se lo considera una maleza agrícola económicamente importante, a pesar de que puede crecer como perenne en áreas que no tienen una temporada fría. Investigadores y reguladores han evaluado el potencial para que las variedades de algodón GM resistentes a los insectos se conviertan en maleza, incluido el algodón que produce la proteína Cry1F, y han descubierto que no hay características del algodón resistente a los insectos que puedan aumentar su potencial para convertirse en una maleza agrícola, debido a que las plantas voluntarias o de regeneración natural de algodón podrían controlarse fácilmente mediante técnicas convencionales de manejo de malezas (Eastick y Hearnden, 2006; Eastick, 2002; USDA, 2003; 2004b; USEPA, 2005).
Maíz: El maíz por lo general no es considerado una maleza, ya que posee pocas de las características que aumentan la probabilidad de que una planta se transforme en maleza, tales como dormancia, desgrane y competitividad (Baker, 1965, 1974). No existen datos que indiquen que la expresión de Cry1F altere la dormancia de la semilla, la capacidad de sobrevivir el invierno, u otras características que podrían alterar la prevalencia del maíz de regeneración natural en temporadas de crecimiento subsiguientes. El maíz de regeneración natural en temporada siguiente que produzca Cry1F no debería presentar ningún desafío inusual en el
manejo de malezas, y podrá manejarse de la misma manera que el maíz de regeneración natural convencional (Carpenter et al., 2002; Raybould et al., 2011; USDA, 2000, 2001, 2004a, 2004c; USEPA, 2001, 2005).
Potencial para convertirse en maleza en ambientes no agrícolas
El principal mecanismo por el cual Cry1F puede introducirse en un ambiente no agrícola es el movimiento de propágulos fuera de las áreas cultivadas (Lee y Natesan, 2006), y los reguladores evalúan cómo tales introducciones pueden dar como resultado una planta GM que se vuelva maleza o invasiva.
Algodón: Mientras que todas las plantas pueden exhibir características de crecimiento como maleza en determinadas condiciones, no se considera que las variedades comerciales de algodón representen un riesgo de conversión en maleza significativo en ambientes no agrícolas. El cruzamiento selectivo ha llevado a que las variedades modernas de algodón dependan de la intervención humana, y factores tales como el estrés hídrico y el frío limitan notablemente la capacidad de las variedades comerciales de sobrevivir en ambientes no agrícolas. A pesar de que la resistencia a los insectos mediada a través de la proteína Cry1F puede suministrar alguna ventaja inadvertida de aptitud biológica a una planta de algodón GM, investigadores y reguladores han determinado que esta ventaja no sería suficiente para permitir que el algodón GM que expresa Cry1F persista en un ambiente no agrícola (Carpenter et al., 2002; Eastick & Hearnden, 2006; Eastick, 2002; JBCH, 2006b, 2006c, 2006d; USDA, 2003, 2004b; USEPA, 2005).
Maíz: Como resultado del extenso cruzamiento selectivo, las variedades comerciales de maíz tienen reducida en gran medida su capacidad de persistir en ambientes no agrícolas sin la intervención del ser humano, y el maíz no se considera una maleza invasiva ni agresiva fuera de los sistemas agrícolas (Carpenter et al., 2002). Los datos agronómicos muestran que Cry1F no posee un impacto significativo sobre los caracteres asociados con el potencial para convertirse en maleza. A pesar que la liberación de factores de control natural (entre ellos los insectos herbívoros) es una explicación parcial del éxito de las especies invasivas (Blumenthal, 2005; Keane y Crawley, 2002; Mack, 1996; Mason, Braun, Warwick, Zhu y Stewart, 2004), las decisiones de las autoridades regulatorias han determinado que es poco probable que la resistencia a plagas de lepidópteros permita que el maíz productor de Cry1F se vuelva invasivo en ambientes no agrícolas (Carpenter et al., 2002; USDA, 2000, 2001, 2004a, 2004c; USEPA, 2001).
Movimiento del transgen a especies emparentadas sexualmente compatibles
El movimiento de transgenes de una planta GM a sus familiares silvestres se realiza mediante el polen, y la producción de híbridos viables en lo reproductivo depende de diversos factores: si el donante de polen se autopoliniza, la proximidad física y temporal de las plantas GM con las especies compatibles sexualmente, la movilidad y viabilidad del polen y la presencia de polinizadores correspondientes (Chandler y Dunwell, 2008).
Algodón: El genoma Gossypium es muy complejo y se organiza en ocho grupos de especies diploides y un grupo de especies tetraploides, que incluye a G. hirsutum. Pueden ocurrir cruces dentro de grupos, pero los cruces entre grupos son raros y la descendencia muestra anormalidades meióticas e infertilidad, incluidos los cruces entre G. hirsutum y miembros de las especies diploide. La hibridación entre G. hirsutum y las tres especies tetraploide silvestres (G. mustelinum, G. darwinii y G. tomentosum), así como también los cruces entre poblaciones asilvestradas de G. barbadense y G. hirsutum, pueden experimentarse fácilmente y resultar en descendencia fértil. Dentro de las condiciones favorables analizadas anteriormente, pueden ocurrir hibridaciones espontáneas cuando las variedades comerciales de G. hirsutum se cultivan cerca de poblaciones naturales de especies tetraploides (OCDE, 2008). No obstante, se considera que la frecuencia de estos cruces entre G. hirsutum transgénica y los familiares silvestres compatibles sexualmente no es mayor que los cruces entre las variedades producidas tradicionalmente de G. hirsutum y las especies silvestres (Carpenter et al., 2002; OCDE, 2008; USDA, 2003, 2004b; USEPA, 2005).
Maíz: Predominantemente, el maíz es polinizado por el viento y no tiene especies emparentadas sexualmente compatibles que se consideren invasivas (Carpenter et al., 2002; OCDE, 2003). El maíz se hibrida libremente con teosintes silvestres, pero se cree que la introgresión de genes es limitada (Baltazar, De Jesús Sánchez-Gonzalez, De la Cruz-Larios y Schoper, 2005; Castillo-Gonzalez y Goodman, 1997; OCDE, 2003). Las poblaciones de teosintes silvestres se limitan a la zona de México, Guatemala y una única población en Nicaragua; y si bien algunos agricultores mexicanos la consideran una maleza grave, otros la usan como forraje y la consideran una especie culturalmente significativa (González y Corral, 1997; Mondragon-Pichardo y Vibrans, 2005). No se espera que ocurran cruces entre teosintes y maíz GM que expresen Cry1F con más frecuencia que entre teosintes y variedades de maíz producidas tradicionalmente (Carpenter et al., 2002; USDA, 2000, 2001, 2003, 2004a; USEPA, 2001).
ANÁLISIS DE LA COMPOSICIÓN DE LAS PLANTAS DE ALGODÓN Y MAÍZ QUE EXPRESAN Cry1F
Muchos procesos de aprobación regulatoria requieren un análisis de la composición de las plantas GM que se utilizarán en alimentos para los seres humanos y los animales. Los datos de composición se pueden utilizar para identificar cambios no intencionales en el cultivo debido a la presencia del transgen. Por lo general, el análisis compara la planta GM con la línea parental no transformada o una isolínea estrechamente relacionada, y los analitos medidos dependen del cultivo y de sus usos planificados. El análisis puede utilizar plantas que se cultivan en diversas ubicaciones y puede incluir datos de varias temporadas de crecimiento, debido a que las condiciones ambientales locales pueden tener un impacto en la composición nutricional, incluso en variedades producidas en forma convencional.10 El objetivo del análisis es verificar que los valores obtenidos para la planta GM están dentro del rango observado en las variedades tradicionales cultivadas en condiciones comparables.
10 En algunos casos en los que las plantas de maíz GM contenían cry1F además del gen de tolerancia al herbicida (pat), se recabaron datos de composición de plantas que habían sido tratadas con glufosinato de amonio, y de plantas cultivadas en el mismo sitio pero no tratadas, para establecer si el herbicida tenía algún efecto.

8 9
La semilla y el forraje de maíz Cry1F y la semilla de algodón Cry1F se han sometido a un análisis proximal para determinar los niveles de proteína cruda, grasa cruda, fibra, humedad y ceniza. Además, se han determinado los niveles de minerales seleccionados, ácidos grasos, aminoácidos y antioxidantes. Algunas plantas del cultivo producen toxinas o compuestos antinutrientes, y también se miden los niveles de estos compuestos para determinar si la presencia de los transgenes ha resultado, de manera inadvertida, en una producción elevada de estas sustancias. Se sabe que el maíz produce los compuestos antinutrientes ácido fítico, rafinosa e inhibidor de la tripsina (OCDE, 2003), y que el algodón produce la toxina gosipol y los ácidos grasos ciclopropenoides (OCDE, 2008). Los niveles de estas sustancias producidos por las variedades de algodón y maíz que expresan Cry1F se midieron y compararon con los niveles de las variedades de algodón y maíz convencionales. En el Anexo III se sintetizan los datos de las fuentes disponibles al público. Todas las diferencias establecidas entre el evento de algodón GM DAS-23236-5 y las variedades del comparador estuvieron dentro del rango normal de variación para el algodón, o las diferencias fueron consideradas irrelevantes para la seguridad ambiental (CFIA, 2005; CTNBio, 2009a; FSANZ, 2004; Health Canada, 2006a; JBCH, 2006b, 2006c; UKACRE, 2004; UKDEFRA, 2005; USDA, 2003; USFDA, 2004a). Una comparación similar para los eventos de maíz DAS-01507-1 y DAS-06275-8 y las variedades del comparador no reveló diferencias relevantes para la seguridad ambiental (CFIA, 2006; COGEM, 2005; CTNBio, 2008, 2009b; EC, 2006; EFSA, 2005, 2006b, 2009a; FSANZ, 2003; Health Canada, 2006b, 2002; Herman et al., 2004; MSPS, 2012; PDOA, 2006; SAGPA, 2008; SFOPH/SFOA, 2001; USDA, 2001, 2004a, 2004c; USFDA, 2001, 2004b).
CONCLUSIÓN
La proteína Cry1F producida por plantas de algodón y maíz GM con resistencia a los insectos proviene de la bacteria común del suelo Bacillus thuringiensis y es específicamente tóxica para los lepidópteros. La prueba de toxicidad con un rango de organismos no blanco representativos demostró que Cry1F no producía efectos observables en concentraciones significativamente mayores que las concentraciones ambientales esperadas de Cry1F. Los datos de campo sugieren que el cultivo de plantas de maíz GM que expresan Cry1F no afecta la abundancia de artrópodos no blanco. Cry1F en plantas puede ser tóxica para los lepidópteros no blanco, pero las evaluaciones de riesgo de las autoridades regulatorias para los productos aprobados han concluido que el riesgo es bajo debido a la falta de exposición a la toxina en el ambiente, especialmente cuando se compara con otras prácticas de control de insectos. El peso de la evidencia de los análisis de datos de fenotipo y de composición demuestra que la expresión de Cry1F en variedades aprobadas de algodón y maíz no altera la fisiología general de las plantas del cultivo e indica que dichas plantas no tienen más probabilidades de convertirse en malezas ni en invasivas que las variedades convencionales de algodón y maíz.
REFERENCIASAdamczyk, J. J., y Gore, J. (2004). Laboratory and field performance of cotton containing Cry1Ac, Cry1F, and both Cry1Ac and Cry1F (Widestrike®) against beet armyworm and fall armyworm larvae (Lepidoptera: Noctuidae). Florida Entomologist, 87(4), 427–432.
Adamczyk, J. J., Greenberg, S., Armstrong, J. S., Mullins, W. J., Braxton, L. B., Lassiter, R. B. y Siebert, M. W. (2008). Evaluations of Bollgard®, Bollgard II®, and Widestrike® technologies against beet and fall armyworm larvae (Lepidoptera: Noctuidae). Florida Entomologist, 91(4), 531–536.
Apaydin, Ö., Çınar, Ç., Turanli, F., Harsa, Ş. y Güneş, H. (2008). Identification and bioactivity of native strains of Bacillus thuringiensis from grain-related habitats in Turkey. Biological Control, 45(1), 21–28. doi:10.1016/j.biocontrol.2008.01.011
Appenzeller, L. M., Malley, L., Mackenzie, S. A., Hoban, D. y Delaney, B. (2009). Subchronic feeding study with genetically modified stacked trait lepidopteran and coleopteran resistant (DAS-Ø15Ø7-1xDAS-59122-7) maize grain in Sprague-Dawley rats. Food and Chemical Toxicology, 47(7), 1512–20. Elsevier Ltd. doi:10.1016/j.fct.2009.03.041
APVMA. (2013). Record of Approved Active Constituents. Public Chemical Registration Information System - PUBCRIS. Obtenido el 24 de enero de 2013, de http://services.apvma.gov.au/PubcrisWebClient/welcome.do
Baker, H. (1965). Characteristics and modes of origins of weeds. En: H. G. B. y G. L. Stebbins (Ed.), (pp. 147–168). New York: Academic Press.
Baker, H. (1974). The evolution of weeds. Annual Review of Ecology and Systematics, 5, 1–24.
Ballester, V., Granero, F., Tabashnik, B. E., Malvar, T. y Ferré, J. (1999). Integrative model for binding of Bacillus thuringiensis toxins in susceptible and resistant larvae of the diamondback moth (Plutella xylostella). Applied and Environmental Microbiology, 65(4), 1413–1419. Obtenido de <Go to ISI>://000079530000008
Balog, A., Szenasi, A., Szekeres, D. y Palinkas, Z. (2011). Analysis of soil dwelling rove beetles (Coleoptera: Staphylinidae) in cultivated maize fields containing the Bt toxins, Cry34/35Ab1 and Cry1F x Cry34/35Ab1. Biocontrol Science & Technology, 21(3-4), 293–297.
Baltazar, B. M., De Jesús Sánchez-Gonzalez, J., De la Cruz-Larios, L. y Schoper, J. B. (2005). Pollination between maize and teosinte: an important determinant of gene flow in Mexico. Theoretical and Applied Genetics, 110(3), 519–526. doi:10.1007/s00122-004-1859-6
Blanco, C. A., Portilla, M., Jurat-Fuentes, J. L., Sánchez, J. F., Viteri, D., Vega-Aquino, P., Terán-Vargas, A. P., Azuara-Domínguez, A., López, J. D. Jr., Arias, R., Zhu, Y. -C., Lugo-Barrera, D. y Jackson, R. (2010). Susceptibility of isofamilies of Spodoptera frugiperda (Lepidoptera: Noctuidae) to Cry1Ac and Cry1F proteins of Bacillus thuringiensis. Southwestern Entomologist, 35(3), 409–415.
Blanco, C. A., Storer, N. P., Abel, C. A., Jackson, R., Leonard, R., Lopez, J. D., Payne, G., Siegfried, B. D., Spencer, T., Terán-Vargas, A. P. (2008). Baseline susceptibility of tobacco budworm (Lepidoptera: Noctuidae) to Cry1F toxin from Bacillus thuringiensis. Journal of Economic Entomology, 101(1), 168–173. Obtenido de http://www.ncbi.nlm.nih.gov/pubmed/18330132
Blumenthal, D. (2005). Interrelated causes of plant invasion. Science, 310(October), 243–244.
Buntin, G. D. (2008). Corn expressing Cry1Ab or Cry1F endotoxin for fall armyworm and corn earworm (Lepidoptera: Noctuidae) management in field corn for grain production. Florida Entomologist, 91(4), 523–530. Florida Entomological Society. doi:10.1653/0015-4040-91.4.523
Carpenter, J., Felsot, A., Goode, T., Hammig, M., Onstad, D. y Sankula, S. (2002). Comparative Environmental Impacts of Biotechnology-derived and Traditional Soybean, Corn, and Cotton Crops. Crops. Ames, IA: Council for Agricultural Science and Technology. Obtenido de http://oregonstate.edu/instruct/bi430-fs430/Documents-2004/7B-MIN%20TILL%20AG/CAST-ComparEnvImpactGMOCrops-2002.pdf
Carstens, K., Anderson, J., Bachman, P., De Schrijver, A., Dively, G., Federici, B., Hamer, M., Gielkens, M., Jensen, P., Lamp, W., Rauschen, S., Ridley, G., Romeis, J., Waggoner, A., (2012). Genetically modified crops and aquatic ecosystems: considerations for environmental risk assessment and non-target organism testing. Transgenic Research, 21(4), 813–842. doi:10.1007/s11248-011-9569-8
Castillo-Gonzalez, F. y Goodman, M. M. (1997). Research on gene flow between improved maize and landraces. En: J. A. Serratos, M. Willcox y F. Castillo (Eds.), Gene Flow Among Maize Landraces, Improved Maize Varieties, and Teosinte: Implications for Transgenic Maize. El Batan, México: CIMMYT.
CERA. (octubre de 2011). A review of the environmental safety of the PAT protein. doi:10.1051/ebr/2012004
CFIA. (2002). Determination of the Safety of Dow AgroSciences Canada Inc. and Pioneer Hi-Bred International’s Insect Resistant and Glufosinate-Ammonium Tolerant Corn (Zea mays L.) Line 1507 (Vol. 9). Ottawa, Ontario.
CFIA. (2005). Decision Document DD2005-52: Determination of the Safety of Dow AgroSciences Canada Inc. Insect Resistant (Gossypium hirsutum L.) Event 281-24-236. Ottawa, Canada.
CFIA. (2006). Decision Document DD2006-59: Determination of the Safety of Dow AgroSciences Canada Inc.’s Insect Resistant and Glufosinate - Ammonium Tolerant Corn (Zea mays L.) Event DAS-06275-8. Ottawa, Ontario.
Chambers, J. A., Jelen, A., Gilbert, M. P., Jany, C. S., Johnson, T. B. y Gawron-Burke, C. (1991). Isolation and characterization of a novel insecticidal crystal protein gene from Bacillus thuringiensis subsp. aizawai. Journal of Bacteriology, 173(13), 3966–3976. Obtenido de http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=208042&tool=pmcentrez&rendertype=abstract
Chandler, S. y Dunwell, J. M. (2008). Gene flow, risk assessment and the environmental release of transgenic plants. Critical Reviews in Plant Sciences, 27(1), 25–49. Taylor & Francis.
Coates, B. S., Sumerford, D. V, Lopez, M. D., Wang, H., Fraser, L. M., Kroemer, J. A., Spencer, T., Kim, K. S., Abel, C. A., Hellmich, R. L. y Siegfried, B. D. (2011). A single major QTL controls expression of larval Cry1F resistance trait in Ostrinia nubilalis (Lepidoptera: Crambidae) and is independent of midgut receptor genes. Genetica, 139(8), 961–72. doi:10.1007/s10709-011-9590-0
COGEM. (2005). Import and processing of insect resistant and herbicide tolerant maize 1507 x NK603. The Hague, Netherlands.
Craig, W., Tepfer, M., Degrassi, G. y Ripandelli, D. (2008). An overview of general features of risk assessments of genetically modified crops. Euphytica, 164(3), 853–880. Springer Netherlands. doi:10.1007/s10681-007-9643-8
Crickmore, N., Zeigler, D. R., Schnepf, E., Van Rie, J., Lerechus, D., Baum, J., Bravo, A., et al. (2012). Bacillus thuringiensis toxin nomenclature. Obtenido el 24 de enero de 2013 de http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/
CTNBio. (2008). Parecer Técnico No 1679/2008. Brasilia, Brazil.
CTNBio. (2009a). Parecer Técnico No 1757/2009. Brasilia, Brazil.
CTNBio. (2009b). Obtenido de Parecer Técnico No 2042/2009. Brasilia, Brazil.
DGSANCO. (2013). EU Pesticides Database. Bruselas, Bélgica. Obtenido el 24 de enero de 2013 de http://ec.europa.eu/sanco_pesticides/public/index.cfm?event=activesubstance.selection
Dryzga, M. D., Yano, B. L., Andrus, A. K. y Mattsson, J. L. (2007). Evaluation of the safety and nutritional equivalence of a genetically modified cottonseed meal in a 90-day dietary toxicity study in rats. Food and Chemical Toxicology, 45(10), 1994–2004. Obtenido de http://www.sciencedirect.com/science/article/B6T6P-4NNYJDK-1/2/b92516b58958f3d4944e0f5f323d19a1
Duan, J. J., Lundgren, J. G., Naranjo, S. y Marvier, M. (2010). Extrapolating non-target risk of Bt crops from laboratory to field. Biology Letters, 6(1), 74–77. doi:10.1098/rsbl.2009.0612
Dutton, A., Romeis, J. y Bigler, F. (2003). Assessing the risks of insect resistant transgenic plants on entomophagous arthropods: Bt-maize expressing Cry1Ab as a case study. BioControl, 48(6), 611–636. Springer Netherlands. doi:10.1023/A:1026313719424
Eastick, R. J. (2002). The Potential Weediness of Transgenic Cotton in Northern Australia. Technical Bulletin No. 305. Clayton South, Australia.
Eastick, R. J., y Hearnden, M. N. (2006). Potential for weediness of Bt cotton in northern Australia. Weed Science, 54(6), 1142–1151. doi:10.1614/WS-06-077R.1
EC. (2006). Commission decision of 3 March 2006 authorising the placing on the market of food containing, consisting of, or produced from genetically modified maize line 1507 (DAS-01507-1). Official Journal of the European Union, 70, 82–86.
EFSA. (2005). Application (reference EFSA-GMO-NL-2004-02) for the placing on the market of insect-tolerant genetically modified maize 1507, for food use. The EFSA Journal, 182, 1–22.
EFSA. (2006a). Guidance Document of the Scientific Panel on Genetically Modified Organisms for the Risk Assessment of Genetically Modified Plants and Derived Food and Feed. Parma, Italia. Obtenido de http://www.efsa.europa.eu/en/scdocs/doc/gmo_guidance_derived_feed_food.pdf
EFSA. (2006b). Application for the placing on the market of insect-protected, glufosinate and glyphosate-tolerant genetically modified maize 1507 x NK603 for food and feed uses from Pioneer Hi-Bred and Mycogen Seeds. Parma, Italia.
EFSA. (2009a). Scientific Opinion: Application (Reference EFSA-GMO-UK-2005-21) for the placing on the market of the insect-resistant and herbicide-tolerant genetically modified maize 59122 x 1507 x NK603 for food and feed uses, import and processing. The EFSA Journal, 1050, 1–32. Obtenido de http://www.efsa.europa.eu/en/efsajournal/doc/1050.pdf
EFSA. (2009b). Scientific Opinion: Application (Reference EFSA-GMO-NL-2005-15) for the placing on the market of the insect-resistant and herbicide-tolerant genetically modified maize 1507 x 59122, for food and feed uses, import and processing. The EFSA Journal, 1074, 1–28. Obtenido de http://www.efsa.europa.eu/en/efsajournal/pub/1074.htm
Ferré, J. y Rie, J. Van. (2002). Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annual Review of Entomology, 47, 501–533.
FSANZ. (2003). Final assessement report: Application A446 - Insect-protected and glufosinate ammonium-tolerant corn line 1507. Canberra, Australia.
FSANZ. (2004). Final Assessment Report: Application A518 - Food derived from insect-protected, herbicide-tolerant cotton line MXB-13. Canberra, Australia.
Gao, Y., Fencil, K. J., Xu, X., Schwedler, D. A., Gilbert, J. R. y Herman, R. A. (2006). Purification and characterization of a chimeric Cry1F δ-endotoxin expressed in transgenic cotton plants. Journal of Agricultural and Food Chemistry, 54(3), 829–835. American Chemical Society. doi:10.1021/jf0523869
Garcia-Alonso, M., Jacobs, E., Raybould, A., Nickson, T. E., Sowig, P., Willekens, H., Van Der Kouwe, P., Layton, R., Amijee, F., Fuentes, Angel M. y Tencalla, F. (2006). A tiered system for assessing the risk of genetically modified plants to non-target organisms. Environmental Biosafety Research, 5(02), 57–65. Obtenido de http://journals.cambridge.org/abstract_S1635792206000182

10 11
Gaspers, C., Siegfried, B. D., Spencer, T., Alves, A. P., Storer, N. P., Schuphan, I., & Eber, S. (2011). Susceptibility of European and North American populations of the European corn borer to the Cry1F insecticidal protein. Journal of Applied Entomology, 135(1-2), 7–16. doi:10.1111/j.1439-0418.2010.01541.x
González, J. de J. S. y Corral, J. A. R. (1997). Teosinte distribution in Mexico. En: J. A. Serratos, M. Wilcox, and F. Castillo (Eds.), Gene Flow Among Maize Landraces, Improved Maize Varieties, and Teosinte: Implications for Transgenic Maize. El Batan, México: CIMMYT. Obtenido de http://repository.cimmyt.org/xmlui/handle/10883/577
González-Cabrera, J., Herrero, S. y Ferré, J. (2001). High genetic variability for resistance to Bacillus thuringiensis toxins in a single population of diamondback moth. Applied and Environmental Microbiology, 67(11), 5043–5048. doi:10.1128/AEM.67.11.5043-5048.2001
Gouffon, C., Van Vliet, A., Van Rie, J., Jansens, S. y Jurat-Fuentes, J. L. (2011). Binding sites for Bacillus thuringiensis Cry2Ae toxin on heliothine brush border membrane vesicles are not shared with Cry1A, Cry1F, or Vip3A toxin. Applied and Environmental Microbiology, 77(10), 3182–3188. doi:10.1128/AEM.02791-10
Hanley, A. V., Huang, Z. Y. y Pett, W. L. (2003). Effects of dietary transgenic Bt corn pollen on larvae of Apis mellifera and Galleria mellonella. Journal of Apicultural Research, 42(4), 77–81.
Health Canada. (2002). Novel Food Information -- Food Biotechnology: Cry1F Insect-resistant/Glufosinate-Tolerant Maize Line 1507. Ottawa, Ontario.
Health Canada. (2006a). Novel Food Information -- Insect Resistant Cotton Event 281-24-236. Ottawa, Ontario.
Health Canada. (2006b). Novel Food Information: Cry1F Insect Resistant, Glufosinate Tolerant Corn Event TC6275. Ottawa, Ontario.
Health Canada. (2008). Bacillus thuringiensis: Re-evaluation Decision Document. Obtenido de http://www.hc-sc.gc.ca/cps-spc/pubs/pest/_decisions/rvd2008-18/index-eng.php
Hellmich, R. L., Siegfried, B. D., Sears, M. K., Stanley-Horn, D. E., Daniels, M. J., Mattila, H. R., Spencer, T., Bidne, K. G. y Lewis, L. C. (2001). Monarch larvae sensitivity to Bacillus thuringiensis-purified proteins and pollen. Proceedings of the National Academy of Sciences of the United States of America, 98(21), 11925–11930. doi:10.1073/pnas.211297698
Herman, R. A., Phillips, A. M., Collins, R. A., Tagliani, L. A., Claussen, F. A., Graham, C. D., Bickers, B. L., Harris, T.A. y Prochaska, L.M. (2004). Compositional equivalency of Cry1F corn event TC6275 and conventional corn (Zea mays L.). Journal of Agricultural and Food Chemistry, 52(9), 2726–2734. American Chemical Society. doi:10.1021/jf049969n
Herman, R. A., Wolt, J. D. y Halliday, W. R. (2002). Rapid degradation of the Cry1F insecticidal crystal protein in soil. Journal of Agricultural and Food Chemistry, 50(24), 7076–7078. American Chemical Society. doi:10.1021/jf025630u
Higgins, L. S., Babcock, J., Neese, P., Layton, R. J., Moellenbeck, D. J. y Storer, N. (2009). Three-year field monitoring of Cry1F, event DAS-Ø15Ø7-1, maize hybrids for nontarget arthropod effects. Environmental Entomology, 38(1), 281–292. Entomological Society of America. doi:10.1603/022.038.0135
Hofte, H., y Whiteley, H. R. (1989). Insecticidal crystal proteins of Bacillus thuringiensis. Microbiological Reviews, 53(2), 242–255. Obtenido de http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=372730&tool=pmcentrez&rendertype=abstract
Iracheta, M. M., Pereyra-Alférez, B., Galán-Wong, L. y Ferré, J. (2000). Screening for Bacillus thuringiensis crystal proteins active against the cabbage looper, Trichoplusia ni. Journal of Invertebrate Pathology, 76(1), 70–75. doi:10.1006/jipa.2000.4946
JBCH. (2004). Maize resistant to Lepidoptera and tolerant to glufosinate herbicide and glyphosate herbicide (cry1F, pat, cp4 epsps, Zea mays subsp. mays (L.) Iltis) (1507×NK603, OECD UI : DAS-Ø15Ø7-1×MON-ØØ6Ø3-6).
JBCH. (2006a). Maize resistant to Coleoptera and Lepidoptera, and tolerant to glufosinate herbicide and glyphosate herbicide (cry34Ab1, cry35Ab1, cry1F, pat, cp4 epeps, Zea mays subsp. mays (L.) Iltis) (58122 x 1507 x NK603, OECD UI: DAS-59122-7 x DAS-0150.
JBCH. (2006b). Cotton resistant to Lepidoptera, and tolerant to glufosinate herbicide and glyphosate herbicide (cry1F, cry1Ac, pat cp4 epsps, Gossypium hirsutum L.)(281 x 3006 x MON88913, OECD UI: DAS-24236-5 x DAS-21023-5 x MON-88913-8).
JBCH. (2006c). Cotton resistant to Lepidoptera and toleran to glufosinate herbicide (cry1F, cry1Ac, pat, Gossypium hirsutum L.)(281 x 3006, OECD UI: DAS-24236-5 x DAS-21023-5). Tokio.
JBCH. (2006d). Cotton resistant to Lepidoptera, and tolerant to glufosinate herbicide and glyphosate herbicide (cry1F, cry1Ac, pat, cp4 epsps, Gossypium hirsutum L.)(281 x 3006 x 1445, OECD UI: DAS-24236-5 x DAS-21023-5 x MON-1445-2). Tokio.
JBCH. (2006e). Maize resistant to Lepidoptera and tolerant to glufosinate herbicide (Modified cry1F, modified bar, Zea mays subsp. mays (L.) Iltis) (TC6275, OECD UI: DAS-06275-8) (pp. 1–14). Tokio.
JBCH. (2009). Maize resistant to Lepidoptera and Coleoptera and tolerant to glufosinate and glyphosate herbicides (cry1A.105, modified cry2Ab2, cry1F, pat, modified cp4 epsps, modified cry3Bb1, cry34Ab1, cry35Ab1, Zea mays subsp. mays (L.) Iltis). Tokio, Japón.
JBCH. (2010). Maize resistant to Lepidoptera, and tolerant to glufosinate and glyphosate herbicides (cry1A.105, modified cry2Ab2, cry1F, pat, modified cp4 epsps, Zea mays subsp. mays (L.) Iltis).
Jurat-Fuentes, J. L. y Adang, M. J. (2001). Importance of Cry1 δ-endotoxin domain II loops for binding specificity in Heliothis virescens (L.). Applied and Environmental Microbiology, 67(1), 323–329. doi:10.1128/AEM.67.1.323
Jurat-Fuentes, J. L. y Adang, M. J. (2006). The Heliothis virescens cadherin protein expressed in Drosophila S2 cells functions as a receptor for Bacillus thuringiensis Cry1A but not Cry1Fa toxins. Biochemistry, 45(32), 9688–9695. doi:10.1021/bi0606703
Keane, R. M., y Crawley, M. J. (2002). Exotic plant invasions and the enemy release hypothesis. Trends in Ecology & Evolution, 17(4), 164–170.
Kumar, A. P., Sharma, R. P., and Malik, V. S. (1996). The insecticidal proteins of Bacillus thuringiensis. Advances in Applied Microbiology, 42, 1–43.
Ladics, G. S., Bardina, L., Cressman, R. F., Mattsson, J. L. y Sampson, H. A. (2006). Lack of cross-reactivity between the Bacillus thuringiensis derived protein Cry1F in maize grain and dust mite Der p7 protein with human sera positive for Der p7-IgE. Regulatory Toxicology and Pharmacology, 44(2), 136–143. doi:10.1016/j.yrtph.2005.11.005
Lee, D. y Natesan, E. (2006). Evaluating genetic containment strategies for transgenic plants. Trends in Biotechnology, 24(3), 109. Obtenido de http://www.sciencedirect.com/science/article/B6TCW-4J6WNXK-4/2/f6bf81ff4bce3a0ae01d93e0566c5463
Liu, X.-Y., Ruan, L.-F., Hu, Z.-F., Peng, D.-H., Cao, S.-Y., Yu, Z.-N., Liu, Y., Zheng, J.-S., Sun, M. (2010). Genome-wide screening reveals the genetic determinants of an antibiotic insecticide in Bacillus thuringiensis. The Journal of Biological Chemistry, 285(50), 39191–39200. doi:10.1074/jbc.M110.148387
Mack, R. N. (1996). Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biological Conservation, 78, 107–121.
MacKenzie, S. A., Lamb, I., Schmidt, J., Deege, L., Morrisey, M. J., Harper, M., Layton, R. J., Prochaska, L. M., Sanders, C., Locke, M., Mattsson, J. L., Fuentes, A. y Delaney, B. (2007). Thirteen week feeding study with transgenic maize grain containing event DAS-Ø15Ø7-1 in Sprague-Dawley rats. Food and Chemical Toxicology, 45(4), 551–562. doi:10.1016/j.fct.2006.09.016
Martínez, C. y Caballero, P. (2002). Contents of cry genes and insecticidal toxicity of Bacillus thuringiensis strains from terrestrial and aquatic habitats. Journal of Applied Microbiology, 92(4), 745–52. Obtenido de http://www.ncbi.nlm.nih.gov/pubmed/11966916
Mason, P., Braun, L., Warwick, S. I., Zhu, B. y Stewart, C. N. (2004). Transgenic Bt-producing Brassica napus: Plutella xylostella selection pressure and fitness of weedy relatives. Environmental and Biosafety Research, 2, 263–276. doi:10.1051/ebr
Mendelsohn, M., Kough, J., Vaituzis, Z. y Matthews, K. (2003). Are Bt crops safe? Nature Biotechnology, 21(9), 1003–1009.
Mondragon-Pichardo, J., y Vibrans, H. (2005). Ethnobotany of the Balsas teosinte. Maydica, 50, 123–128.
MSPS. (2012). Resolución Número 0001488: Por la cual se autoriza el uso de granos de maíz, provenientes de híbridos de maíz genéticamente modificado, con la tecnología HX1 x MON810 x NK603 (DAS-01507-1 x MON-00810-6 x MON-00603-6), como alimento o materia prima para la. Bogotá, Colombia.
Murray, E. E., Lotzer, J. y Eberle, M. (1989). Codon usage in plant genes. Nucleic Acids Research, 17(2), 12–18.
OCDE. (1992). Recombinant DNA Safety Considerations. Organización para la Cooperación y el Desarrollo Económicos. París.
OCDE. (2003). Consensus Document on the Biology of Zea mays subsp. mays (Maize). Organización para la Cooperación y el Desarrollo Económicos. París.
OCDE. (2007). Consensus Document on Safety Information on Transgenic Plants Expressing Bacillus thuringiensis - Derived Insect Control Proteins. Organización para la Cooperación y el Desarrollo Económicos. Paris.
OCDE. (2008). Consensus Document on the Biology of Cotton (Gossypium spp.). Organización para la Cooperación y el Desarrollo Económicos. Paris.
Oppert, B., Ellis, R. T. y Babcock, J. (2010). Effects of Cry1F and Cry34Ab1/35Ab1 on storage pests. Journal of Stored Products Research, 46(3), 143–148. Elsevier Ltd. doi:10.1016/j.jspr.2010.01.003
PDOA. (2006). Determination of the safety of Pioneer’s and Dow Agro Science’s combined trait product corn: 59122 x 1507 x NK603 for direct use as food, feed and for processing. Quezon City, Filipinas.
Pereira, Eliseu J G, Siqueira, H. A. A., Zhuang, M., Storer, N. P. y Siegfried, B. D. (2010). Measurements of Cry1F binding and activity of luminal gut proteases in susceptible and Cry1F resistant Ostrinia nubilalis larvae (Lepidoptera: Crambidae). Journal of Invertebrate Pathology, 103(1), 1–7. doi:10.1016/j.jip.2009.08.014
Pereira, Eliseu J.G., Lang, B. A., Storer, N. P.y Siegfried, B. D. (2008). Selection for Cry1F resistance in the European corn borer and cross-resistance to other Cry toxins. Entomologia Experimentalis et Applicata, 126(2), 115–121. doi:10.1111/j.1570-7458.2007.00642.x
Pigott, C. R. y Ellar, D. J. (2007). Role of receptors in Bacillus thuringiensis crystal toxin activity. Microbiology and Molecular Biology Reviews, 71(2), 255–81. doi:10.1128/MMBR.00034-06
Que, Q., Chilton, M.-D. M., de Fontes, He, M., Nuccio, C., Zhu, T., Wu, Y., Chen, J. S., Shi, L. (2010). Trait stacking in transgenic crops: challenges and opportunities. GM Crops, 1(4), 220–229. doi:10.4161/gmcr.1.4.13439
Raybould, A. (2006). Problem formulation and hypothesis testing for environmental risk assessment of genetically modified crops. Environmental Biosafety Research, 5, 119–126. doi:10.1051/ebr
Raybould, A. (2007). Ecological versus ecotoxicological methods for assessing the environmental risks of transgenic crops. Plant Science, 173(6), 589–602. Obtenido de http://www.sciencedirect.com/science/article/pii/S0168945207002488
Raybould, A., Higgins, L. S., Horak, M. J., Layton, R. J., Storer, N. P., De La Fuente, J. M. y Herman, R. A. (2011). Assessing the ecological risks from the persistence and spread of feral populations of insect-resistant transgenic maize. Transgenic Research, 21(3), 655–664. doi:10.1007/s11248-011-9560-4
Romeis, J., Bartsch, D., Bigler, F., Candolfi, M. P., Gielkens, M. M. C., Hartley, S. E., Hellmich, R. L., Huesing, J. E., Jepson, P.C., Layton, R., Quemada, H., Raybould, A., Rose, R. I., Schiemann, J., Sears, M. K., Shelton, A. M., Sweet, J., Vaituzis, Z. y Wolt, J. D. (2008). Assessment of risk of insect-resistant transgenic crops to nontarget arthropods. Nature Biotechnology, 26(2), 203–208. Nature Publishing Group. Obtenido de http://dx.doi.org/10.1038/nbt1381
Romeis, J., Raybould, A., Bigler, F., Candolfi, M. P., Hellmich, R. L., Huesing, J. E. y Shelton, A. M. (2013). Deriving criteria to select arthropod species for laboratory tests to assess the ecological risks from cultivating arthropod-resistant genetically engineered crops. Chemosphere, 90(3), 901–909. Elsevier Ltd. doi:10.1016/j.chemosphere.2012.09.035
SAGPA. (2008). Visto el Expediente N S01:0066649/2008 del Registro del Ministerio de Economia y Produccion. Buenos Aires, Argentina.
Sanvido, O., Romeis, J., Gathmann, A., Gielkens, M., Raybould, A. y Bigler, F. (2012). Evaluating environmental risks of genetically modified crops: ecological harm criteria for regulatory decision-making. Environmental Science & Policy, 15(1), 82–91. doi:10.1016/j.envsci.2011.08.006
Scheideler, S. E., Rice, D., Smith, B., Dana, G. y Sauber, T. (2008). Evaluation of nutritional equivalency of corn grain from DAS-Ø15Ø7-1 (Herculex I) in the diets of laying hens. Journal of Applied Poultry Research, 17(3), 383–389. doi:10.3382/japr.2007-00080
Schnepf, E., Crickmore, N., Van Rie, J., Lereclus, D., Baum, J., Feitelson, J., Zeigler, D. R. y Dean, D. H.(1998). Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Reviews, 62(3), 775–806. Obtenido de http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=98935&tool=pmcentrez&rendertype=abstract
Schnepf, H. E., Lee, S., Dojillo, J., Burmeister, P., Fencil, K., Morera, L., Nygaard, L., Narva, K. E. y Wolt, J. D. (2005). Characterization of Cry34/Cry35 binary insecticidal proteins from diverse Bacillus thuringiensis strain collections. Applied and Environmental Microbiology, 71(4), 1765–1774. American Society for Microbiology.
Seifinejad, A., Jouazni, G., Hosseinzadeh, A. y Abdmishani, C. (2008). Characterization of Lepidoptera-active cry and vip genes in Iranian Bacillus thuringiensis strain collection. Biological Control, 44(2), 216–226. doi:10.1016/j.biocontrol.2007.09.010
Sena, J. A. D., Hernández-Rodríguez, C. S., and Ferré, J. (2009). Interaction of Bacillus thuringiensis Cry1 and Vip3A proteins with Spodoptera frugiperda midgut binding sites. Applied and Environmental Microbiology, 75(7), 2236–2237. American Society for Microbiology (ASM). Obtenido de http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2663230&tool=pmcentrez&rendertype=abstract
SFOPH/SFOA. (2001). Information to the Swiss Authorities on the Potential Environmental Impact of Genetically Modified Plants in Accordance with Annexes IIB and III of Directive 90/220/EEC. Berna, Suiza.
Shan, G., Embrey, S. K., Herman, R. A. y McCormick, R. (2008). Cry1F protein not detected in soil after three years of transgenic Bt corn (1507 Corn) use. Environmental Entomology, 37(1), 255–262. Entomological Society of America. doi:10.1603/0046-225x(2008)37[255:cpndis]2.0.co;2
Siebert, M. W., Babcock, J. M., Nolting, S., Santos, A. C., Adamczyk, J. J., Jr, Neese, P. A., King, J. E., Jenkins, J. N., McCarty, J., Lorenz,G. M., Fromme, D. D. y Lassiter, R. B. (2008). Efficacy of Cry1F insecticidal protein in maize and cotton for control of fall armyworm (Lepidoptera: Noctuidae). Florida Entomologist, 91(4), 555–565.
Siebert, M. W., Nolting, S., Leonard, B. R., Braxton, L. B., All, J. N., Van Duyn, J. W., Bradley, J. R., Bacheler, J., Huckaba, R. M. (2008). Efficacy of transgenic cotton expressing Cry1Ac and Cry1F insecticidal protein against heliothines (Lepidoptera: Noctuidae). Journal of Economic Entomology, 101(6), 1950–1959. Entomological Society of America. doi:10.1603/0022-0493-101.6.1950

12 13
Siebert, M. W., Patterson, T. G., Gilles, G. J., Nolting, S. P., Braxton, L. B., Leonard, B. R., Van Duyn, J. W., Lassiter, R. B. (2009). Quantification of Cry1Ac and Cry1F Bacillus thuringiensis insecticidal proteins in selected transgenic cotton plant tissue types. Journal of Economic Entomology, 102(3), 1301–1308. Entomological Society of America. doi:10.1603/029.102.0357
Stein, H. H., Sauber, T. E., Rice, D. W.,Hinds, M. A., Smith, B. L., Dana, G., Peters, D. N. y Hunst, P. (2009). Growth performance and carcass composition of pigs fed corn grain from DAS-Ø15Ø7-1 (Herculex I) hybrids. Professional Animal Scientist, 25, 689–694.
Storer, N. P., Babcock, J. M., Schlenz, M., Meade, T., Thompson, G. D., Bing, J. W. y Huckaba, R. M. (2010). Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. Journal of Economic Entomology, 103(4), 1031–1038. doi:10.1603/EC10040
Tabashnik, B. E. (1992). Evaluation of synergism among Bacillus thuringiensis toxins. Applied and Environmental Microbiology, 58(10), 3343–3346. Obtenido de http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=183101&tool=pmcentrez&rendertype=abstract
Tabashnik, B. E., Carrière, Y., Dennehy, T. J., Morin, S., Sisterson, M. S., Roush, R. T., Shelton, A. M., Zhao, J. Z. (2003). Insect resistance to transgenic Bt crops: Lessons from the laboratory and field. Journal of Economic Entomology, 96(4), 1031–1038. Obtenido de http://www.ncbi.nlm.nih.gov/pubmed/14503572
Tian, J.-C., Collins, H. L., Romeis, J., Naranjo, S. E., Hellmich, R. L., & Shelton, A. M. (2012). Using field-evolved resistance to Cry1F maize in a lepidopteran pest to demonstrate no adverse effects of Cry1F on one of its major predators. Transgenic Research, 21(6), 1303–1310. doi:10.1007/s11248-012-9604-4
Tian, J.-C., Wang, X.-P., Long, L.-P., Romeis, J., Naranjo, S. E., Hellmich, R. L., Wang, P., Earle, E. D., Shelton, A. M. (2013). Bt crops producing Cry1Ac, Cry2Ab and Cry1F do not harm the green lacewing, Chrysoperla rufilabris. (J. L. Jurat-Fuentes, Ed.) PLoS ONE, 8(3), e60125. doi:10.1371/journal.pone.0060125
Tian, X. C., Kubota, C., Sakashita, K., Izaike, Y., Okano, R., Tabara, N., Curchoe, C., Jacob, L., Zhang, Y., Smith, S., Bormann, C., Xu, J., Sato, M., Andrew, S. y Yang, X. (2005). Meat and milk compositions of bovine clones. Proceedings of the National Academy of Sciences of the United States of America, 102(18), 6261–6266. doi:10.1073/pnas.0500140102
Tindall, K. V., Siebert, M. W., Leonard, B. R., All, J.y Haile, F. J. (2009). Efficacy of Cry1Ac:Cry1F proteins in cotton leaf tissue against fall armyworm, beet armyworm, and soybean looper (Lepidoptera: Noctuidae). Journal of Economic Entomology, 102(4), 1497–1505. Entomological Society of America. doi:10.1603/029.102.0414
UKACRE. (2004). Advice on a notification for marketing of insect resistant GM cotton. Londres, Reino Unido.
UKDEFRA. (2005). Comments or objections to the placing on the market of a GMO under Part C of Directive 2001/18/EC (Article 15). Londres, Reino Unido.
USDA. (2000). Petition for determination of nonregulated status: B.t. Cry1F insect resistant, glufosinate tolerant maize line. Washington, DC.
USDA. (2001). Approval of Mycogen Seeds c/o Dow AgroSciences LLC and Pioneer Hi-Bred International, Inc. Request 00-136-01P Seeking a Determination of Non-regulated Status for Bt Cry1F Insect Resistant, Glufosinate Tolerant Corn Line 1507. Washington, DC.
USDA. (2003). Petition for determination of non-regulated status: B.t. Cry1F insect-resistant cotton event 281-24-236. Washington, DC.
USDA. (2004a). Application for an Extension of the Determination of Nonregulated Status for B.t. Cry1F Insect-Resistant, Glufosinate-Tolerant Maize (00-136-01p): Maize Line 6275. Washington, DC.
USDA. (2004b). Approval of Mycogen/Dow Petitions 03-036l-01p and 03-036-02p seeking determination of nonregulated status for insect-resistant cotton events 281-24-236 and 3006-210-23 genetically engineered to express synthetic B.t. Cry1F and Cry1Ac, respectively. Washington, DC.
USDA. (2004c). Approval of Mycogen Seeds c/o Dow AgroSciences LLC Request (03-181-01p) Seeking Extension of Determination of Non-regulated Status for Bt Cry1F Insect Resistant, Glufosinate Tolerant Corn Line 6275. Washington, DC.
USEPA. (1998). Reregistration Eligibility Decision (RED) Bacillus thuringiensis. Washington, DC.
USEPA. (2001). Biopesticide Registration Action Document: Bacillus thuringiensis Cry1F Corn. Washington, DC.
USEPA. (2005). Bacillus thuringiensis Cry1F/Cry1Ac Construct 281/3006 Insecticidal Crystal Protein as expressed in cotton (006445 & 006481) Fact sheet. Washington, DC.
USEPA. (2007). White Paper on Tier-Based Testing for the Effects of Proteinaceous Insecticidal Plant-Incorporated Protectants on Non-Target Arthropods for Regulatory Risk Assessments. Environmental Protection (p. 36). Washington DC. Obtenido de http://www.epa.gov/pesticides/biopesticides/pips/non-target-arthropods.pdf
USEPA. (2011). Harmonized Test Guidelines. Obtenido de http://epa.gov/ocspp/pubs/frs/home/guidelin.htm
USFDA. (2001). Biotechnology Consultation: Note to the File, BNF No. 000073. Washington, DC.
USFDA. (2004a). Biotechnology Consultation Note to the File: BNF No. 000085. Washington, DC.
USFDA. (2004b). Biotechnology Consultation, Note to the File, BNF No. 000093. Washington, DC.
Vachon, V., Laprade, R. y Schwartz, J.-L. (2012). Current models of the mode of action of Bacillus thuringiensis insecticidal crystal proteins: A critical review. Journal of Invertebrate Pathology, 111(1), 1–12. Elsevier Inc. doi:10.1016/j.jip.2012.05.001
Van Frankenhuyzen, K. (2009). Insecticidal activity of Bacillus thuringiensis crystal proteins. Journal of Invertebrate Pathology, 101(1), 1–16. doi:10.1016/j.jip.2009.02.009
Wolt, J. D., Conlan, C. A. y Majima, K. (2005). An ecological risk assessment of Cry1F maize pollen impact to pale grass blue butterfly. Environmental and Biosafety Research, 4(2005), 243–251. doi:10.1051/ebr
Wolt, J. D., Keese, P., Raybould, A., Fitzpatrick, J. W., Burachik, M., Gray, A., Olin, S. S., Schiemann, J., Sears, M., Wu, F. (2010). Problem formulation in the environmental risk assessment for genetically modified plants. Transgenic Research, 19(3), 425–36. doi:10.1007/s11248-009-9321-9
ANEXO I:
Las tablas que aparecen a continuación presentan un resumen de datos de expedientes de solicitantes y de documentos de decisiones de las autoridades regulatorias. Siempre que es posible, los datos y las estadísticas que los acompañan se presentan como aparecen en el documento citado para facilitar las referencias cruzadas. En las fuentes citadas, se puede encontrar información adicional acerca de las metodologías de recolección y de toma de muestras.
Datos resumidos para algodón
Tabla I.1. Resumen de la expresión de la nueva proteína Cry1F en la línea de algodón MXB-13 (FSANZ, 2004)
Tejido de algodón Expresión promedio de la proteína (ng/mg peso seco)1
Hoja joven (3-6 semanas) 6,81
Hoja terminal 8,19
Flores 5,44
Botón floral 4,88
Cápsula (temprano) 3,52
Planta entera (plántula) 14,1
Planta entera (polinización) 25,3
Planta entera (defoliación) 22,0
Raíz (plántula) 0,88
Raíz (polinización) 0,54
Raíz (defoliación) 0,51
Polen 0,06
Néctar No detectado
Semilla 4,13
Fracción de algodón procesado
Semilla de algodón 3,1
Grano 3,9
Cascarilla 0,16
Harina tostada No detectado
Aceite refinado No detectado
1 Los resultados se informan en ng/ proteína/ mg muestra de peso seco, mientras que se usó peso fresco en semilla de algodón, polen, néctar y productos procesados.
2 La concentración calculada es menor que el LOQ del método.
Tabla I.2. Resumen de la expresión de la proteína Cry1F en el evento de algodón 281-24-236 determinado por ELISA (USDA, 2003)
Tejido Cry1F ng/mg de peso seco de tejido1
Promedio2 Desviación estándar
Rango
Hoja joven (3-6 semanas) 6,48 3,3 1,2–16,8
Hoja terminal 7,67 5,3 1,3–20,7
Flor 5,71 2,1 3,0–12,3
Botón floral 5,04 1,8 2,0–9,4
Cápsula (temprano) 4,02 2,0 1,2–9,2
Planta entera (plántula) 11,5 4,3 5,5–23,3
Planta entera (polinización) 22,8 7,2 12,1–38,4
Planta entera (defoliación) 21,1 9,9 8,4–37,6
Raíz (plántula) 0,72 0,6 0,21–2,3
Raíz (polinización) 0,36 0,1 0,10–0,62
Raíz (defoliación) 0,61 0,5 0,12–1,7
Polen (0,09)3 0,3 ND4–1,1
Néctar ND No aplicable ND–ND
Semilla 5,13 1,2 3,2–8,2
1 Resultados basados en el peso fresco de tejido de polen, néctar y semilla.2 Los promedios se calculan a partir de muestras tomadas en los seis sitios.3 La concentración calculada es menor que el LOQ del método.4 El valor de absorbancia de la muestra fue menor que la absorbancia estándar
más baja.
Tabla I.3. Cálculos de exposición prolongada1 (HEEE, por sus siglas en inglés) para la expresión de la proteína Cry1F en 281-24-236 (USDA, 2003)
Tejido HEEE (ng/ mg tejido)
Hoja (terminal) 18,1
Planta entera (defoliación) 40,5
Raíz (defoliación) 1,6
Polen 0,7
Néctar <0,05 ng/µL
Semilla 7,5
1 HEEE se calculó como el Promedio + 1,96* Desviación Estándar de los valores de expresión informados en la Tabla I.2.
Tabla I.4. Resumen de la expresión de la proteína Cry1F en las fracciones de algodón procesado del evento 281-24-236 (USDA, 2003)
Fracción procesada ng Cry1F/ mg peso fresco de tejido
Semilla de algodón 3,3
Grano 3,0
Cascarilla 0,22
Harina tostada No detectado
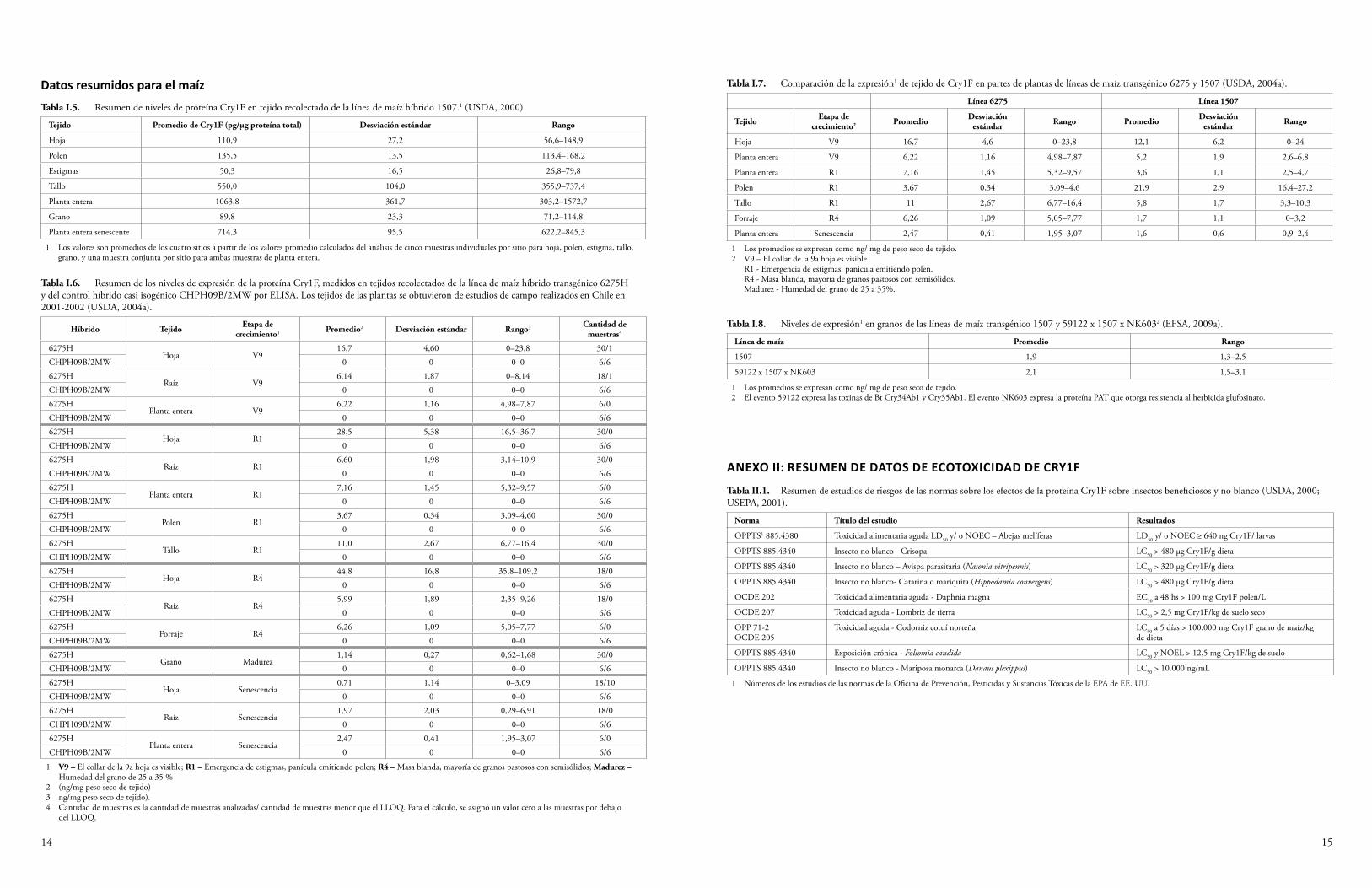
14 15
Datos resumidos para el maízTabla I.5. Resumen de niveles de proteína Cry1F en tejido recolectado de la línea de maíz híbrido 1507.1 (USDA, 2000)
Tejido Promedio de Cry1F (pg/µg proteína total) Desviación estándar Rango
Hoja 110,9 27,2 56,6–148,9
Polen 135,5 13,5 113,4–168,2
Estigmas 50,3 16,5 26,8–79,8
Tallo 550,0 104,0 355,9–737,4
Planta entera 1063,8 361,7 303,2–1572,7
Grano 89,8 23,3 71,2–114,8
Planta entera senescente 714,3 95,5 622,2–845,3
1 Los valores son promedios de los cuatro sitios a partir de los valores promedio calculados del análisis de cinco muestras individuales por sitio para hoja, polen, estigma, tallo, grano, y una muestra conjunta por sitio para ambas muestras de planta entera.
Tabla I.6. Resumen de los niveles de expresión de la proteína Cry1F, medidos en tejidos recolectados de la línea de maíz híbrido transgénico 6275H y del control híbrido casi isogénico CHPH09B/2MW por ELISA. Los tejidos de las plantas se obtuvieron de estudios de campo realizados en Chile en 2001-2002 (USDA, 2004a).
Híbrido Tejido Etapa de crecimiento1 Promedio2 Desviación estándar Rango3 Cantidad de
muestras4
6275HHoja V9
16,7 4,60 0–23,8 30/1
CHPH09B/2MW 0 0 0–0 6/6
6275HRaíz V9
6,14 1,87 0–8,14 18/1
CHPH09B/2MW 0 0 0–0 6/6
6275HPlanta entera V9
6,22 1,16 4,98–7,87 6/0
CHPH09B/2MW 0 0 0–0 6/6
6275HHoja R1
28,5 5,38 16,5–36,7 30/0
CHPH09B/2MW 0 0 0–0 6/6
6275HRaíz R1
6,60 1,98 3,14–10,9 30/0
CHPH09B/2MW 0 0 0–0 6/6
6275HPlanta entera R1
7,16 1,45 5,32–9,57 6/0
CHPH09B/2MW 0 0 0–0 6/6
6275HPolen R1
3,67 0,34 3,09–4,60 30/0
CHPH09B/2MW 0 0 0–0 6/6
6275HTallo R1
11,0 2,67 6,77–16,4 30/0
CHPH09B/2MW 0 0 0–0 6/6
6275HHoja R4
44,8 16,8 35,8–109,2 18/0
CHPH09B/2MW 0 0 0–0 6/6
6275HRaíz R4
5,99 1,89 2,35–9,26 18/0
CHPH09B/2MW 0 0 0–0 6/6
6275HForraje R4
6,26 1,09 5,05–7,77 6/0
CHPH09B/2MW 0 0 0–0 6/6
6275HGrano Madurez
1,14 0,27 0,62–1,68 30/0
CHPH09B/2MW 0 0 0–0 6/6
6275HHoja Senescencia
0,71 1,14 0–3,09 18/10
CHPH09B/2MW 0 0 0–0 6/6
6275HRaíz Senescencia
1,97 2,03 0,29–6,91 18/0
CHPH09B/2MW 0 0 0–0 6/6
6275HPlanta entera Senescencia
2,47 0,41 1,95–3,07 6/0
CHPH09B/2MW 0 0 0–0 6/6
1 V9 – El collar de la 9a hoja es visible; R1 – Emergencia de estigmas, panícula emitiendo polen; R4 – Masa blanda, mayoría de granos pastosos con semisólidos; Madurez – Humedad del grano de 25 a 35 %
2 (ng/mg peso seco de tejido)3 ng/mg peso seco de tejido).4 Cantidad de muestras es la cantidad de muestras analizadas/ cantidad de muestras menor que el LLOQ. Para el cálculo, se asignó un valor cero a las muestras por debajo
del LLOQ.
Tabla I.7. Comparación de la expresión1 de tejido de Cry1F en partes de plantas de líneas de maíz transgénico 6275 y 1507 (USDA, 2004a).
Línea 6275 Línea 1507
Tejido Etapa de crecimiento2 Promedio Desviación
estándar Rango Promedio Desviación estándar Rango
Hoja V9 16,7 4,6 0–23,8 12,1 6,2 0–24
Planta entera V9 6,22 1,16 4,98–7,87 5,2 1,9 2,6–6,8
Planta entera R1 7,16 1,45 5,32–9,57 3,6 1,1 2,5–4,7
Polen R1 3,67 0,34 3,09–4,6 21,9 2,9 16,4–27,2
Tallo R1 11 2,67 6,77–16,4 5,8 1,7 3,3–10,3
Forraje R4 6,26 1,09 5,05–7,77 1,7 1,1 0–3,2
Planta entera Senescencia 2,47 0,41 1,95–3,07 1,6 0,6 0,9–2,4
1 Los promedios se expresan como ng/ mg de peso seco de tejido.2 V9 – El collar de la 9a hoja es visible R1 - Emergencia de estigmas, panícula emitiendo polen. R4 - Masa blanda, mayoría de granos pastosos con semisólidos. Madurez - Humedad del grano de 25 a 35%.
Tabla I.8. Niveles de expresión1 en granos de las líneas de maíz transgénico 1507 y 59122 x 1507 x NK6032 (EFSA, 2009a).
Línea de maíz Promedio Rango
1507 1,9 1,3–2,5
59122 x 1507 x NK603 2,1 1,5–3,1
1 Los promedios se expresan como ng/ mg de peso seco de tejido.2 El evento 59122 expresa las toxinas de Bt Cry34Ab1 y Cry35Ab1. El evento NK603 expresa la proteína PAT que otorga resistencia al herbicida glufosinato.
Tabla II.1. Resumen de estudios de riesgos de las normas sobre los efectos de la proteína Cry1F sobre insectos beneficiosos y no blanco (USDA, 2000; USEPA, 2001).
Norma Título del estudio Resultados
OPPTS1 885.4380 Toxicidad alimentaria aguda LD50 y/ o NOEC – Abejas melíferas LD50 y/ o NOEC ≥ 640 ng Cry1F/ larvas
OPPTS 885.4340 Insecto no blanco - Crisopa LC50 > 480 µg Cry1F/g dieta
OPPTS 885.4340 Insecto no blanco – Avispa parasitaria (Nasonia vitripennis) LC50 > 320 µg Cry1F/g dieta
OPPTS 885.4340 Insecto no blanco- Catarina o mariquita (Hippodamia convergens) LC50 > 480 µg Cry1F/g dieta
OCDE 202 Toxicidad alimentaria aguda - Daphnia magna EC50 a 48 hs > 100 mg Cry1F polen/L
OCDE 207 Toxicidad aguda - Lombriz de tierra LC50 > 2,5 mg Cry1F/kg de suelo seco
OPP 71-2OCDE 205
Toxicidad aguda - Codorniz cotuí norteña LC50 a 5 días > 100.000 mg Cry1F grano de maíz/kg de dieta
OPPTS 885.4340 Exposición crónica - Folsomia candida LC50 y NOEL > 12,5 mg Cry1F/kg de suelo
OPPTS 885.4340 Insecto no blanco - Mariposa monarca (Danaus plexippus) LC50 > 10.000 ng/mL
1 Números de los estudios de las normas de la Oficina de Prevención, Pesticidas y Sustancias Tóxicas de la EPA de EE. UU.
ANEXO II: RESUMEN DE DATOS DE ECOTOXICIDAD DE CRY1F

16 17
Tabla II.2. Resumen de estudios de riesgos de las normas sobre los efectos de la proteína Cry1F (USDA, 2003).
Norma Título del estudio Fuente de la proteína Resultados
OCDE 401 Toxicidad aguda - Ratón Proteína Cry1F derivada de microbios LD50>600 mg Cry1F/kg
OPP B, 71-2 Toxicidad alimentaria aguda - Codorniz cotuí norteña
Harina de algodón preparada con semilla de algodón 3006-210/281-24-236
LC50 a 8 días >100.000 µg harina/ kg de dieta
OCDE 207 Toxicidad aguda - Lombriz de tierra Cry1F derivada de microbios, sola o combinada con proteína Cry1Ac derivada de microbios
LC50 a 14 días >247 mg Cry1F/kg de suelo762 x EEC en suelo
Propuesta por OCDE Toxicidad crónica - Colémbolos Cry1F derivada de microbios, sola o combinada con proteína Cry1Ac derivada de microbios
LC50>702 µg Cry1F/kg2167 x EEC en suelo
OCDE 202 Toxicidad alimentaria aguda - Daphnia magna Combinación de proteínas Cry1F y Cry1Ac derivadas de microbios
EC50 a 48 hs > 510 µg Cry1F/L395 x EEC en agua
OCDE 203 Toxicidad alimentaria aguda - Trucha Arco Iris Harina de algodón preparada con semilla de algodón 3006-210/281-24-236
LC50 a 8 días >0,209 µg harina/ kg de dieta162 x EEC en agua
OPPTS 885.4380 Toxicidad alimentaria aguda LD50 – Abejas melíferas
Combinación de proteínas Cry1F y Cry1Ac derivadas de microbios
LC50 > 1,98 µg Cry1F/g dieta2,8 x expresión prolongada en polen
OPPTS 885.4340 Insecto no blanco - Crisopa Combinación de proteínas Cry1F y Cry1Ac derivadas de microbios
LC50≥5,2µg Cry1F/g de dieta7 x expresión prolongada en polen104 x expresión prolongada en néctar
OPPTS 885.4340 Insecto no blanco - Avispa parasitaria Cry1F derivada de microbios, sola o combinada con proteína Cry1Ac derivada de microbios
LC50>5,2µg Cry1F/mL7 x expresión prolongada en polen104 x expresión prolongada en néctar
OPPTS 885.4340 Insecto no blanco - Catarina o mariquita Cry1F derivada de microbios, sola o combinada con proteína Cry1Ac derivada de microbios
LC50>300 µg Cry1F/mL428 x expresión prolongada en polen
Tabla II.3. Toxicidad potencial de Cry1F (FSANZ, 2004).
Norma Título del estudio Material de prueba/ control Resultados
OCDE 401 Límite de toxicidad oral aguda - Ratón5 machos y 5 hembras de ratón CD-12000 mg/kg peso corporal (600 mg Cry1F/kg peso corp.) administrados en 2 dosis de sonda gástrica, con 1 hora de diferencia
Proteína de Cry1F obtenida de pseudomonas fluorescens/ 0,5 % de metil celulosa
LD50>600 mg Cry1F/kg peso corporal
OCDE 401 Límite de toxicidad oral aguda - Ratón5 machos y 5 hembras de ratón CD-15000 mg/kg peso corporal (375 mg Cry1F/kg peso corp. y 350 mg Cry1Ac/kg peso corp.) administrados en 3 dosis de sonda gástrica, con 1 hora de diferencia
Mezcla 50:50 de Cry1F (15 % pura) y Cry1Ac (14 % pura) obtenida de Pseudomonas fluorescens /0,5 % metil celulosa
LD50>375 mg Cry1F/kg de peso corporal y LD50>350 mg Cry1Ac/kg de peso corporal
ANEXO III: RESUMEN DE ANÁLISIS DE COMPOSICIÓN DE PLANTAS GM QUE EXPRESAN CRY1F, INCLUYENDO ANÁLISIS DE TOXINAS Y ANTINUTRIENTES
Datos resumidos para algodón
Proximales
Tabla III.1. Resumen de niveles de expresión de la proteína Cry34Ab1 medida en tejidos recolectados de híbridos de maíz 59122 (evento DAS-59122-7) (USDA, 2004).
Componente1 MXB-132 Control Valor P de prueba t pareada Valor P ajustado por Dunnet Rango de bibliografía3
Ceniza3,9
[3,5–4,1]0,21
4,0[3,7–4,4]
0,280,238 0,489 4,1–4,9
Grasa22,9
[20,9–23,7]1,02
22,6[21,4–24,3]
1,150,657 0,941 16,1–26,7
Humedad3,5
[2,6–5,6]1,09
3,3[2,5–4,2]
0,650,659 0,943 5,4–15,9
Proteína27,9
[26,4–29,0]0,95
27,6[26,1–29,3]
1,190,717 0,966 12–32
Carbohidratos45,4
[43,5–47,2]1,34
45,8[42,1–48,1]
2,090,691 0,956 42,8–47,8
Calorías (Kcalorías/100 gm)
499[489–505]
5,32
497[491–504]
4,930,552 0,875 479–508
Fibra cruda15,9
[14,7–17,0]0,79
17,6[16,6–18,6]
0,690,003 0,009 17,2
Fibra detergente ácida25,2
[23,9–26,4]0,96
25,2[23,1–27,2]
0,960,989 1,0 26
Fibra detergente neutra34,1
[30,7–36,9]2,35
35,9[32,8–38,5]
1,920,316 0,613 37
1 Todos los valores (promedio y rango) expresados como % peso seco.2 Los valores exhibidos son el promedio (negrita), el rango (entre paréntesis) y la desviación estándar.3 Rango de bibliografía combinada.

18 19
Tabla III.2. Análisis proximal de fracciones procesadas de semilla de algodón (FSANZ, 2004).1
ComponenteGranos
MXB-13 Control Rango de bibliografía
Humedad 6,9 7,6 No aplicable
ComponenteCascarilla
MXB-13 Control Rango de bibliografía
Ceniza 2,8 3,0 2,39–3,97
Grasa 2,0 3,0 1,0–3,3
Humedad 10,6 10,3 8,5–12,3
Proteína 6,2 7,1 4,0–6,9
Carbohidratos 89,0 86,8 No aplicable
Calorías (Kcalorías/100 gm) 399 403 No aplicable
ComponenteHarina tostada
MXB-13 Control Rango de bibliografía
Ceniza 6,7 6,0 4,6–9,8
Grasa 2,0 4,6 0,6–4,7
Humedad 9,2 2,2 9–13,3
Proteína 51,3 47,2 43,0–52,4
Carbohidratos 40,0 42,1 No aplicable
Calorías (Kcalorías/100 gm) 383 399 No aplicable
Fibra cruda 9,3 12,4 8,4–15,3
Fibra detergente ácida 14,1 18,5 12,2–23,9
Fibra detergente neutra 20,2 24,2 15,8–32,4
ComponenteAceite refinado
MXB-13 Control Rango de bibliografía
Grasa 100,1 100,2 No aplicable
Humedad <0,1 <0,1 No aplicable
Proteína <0,1 <0,1 No aplicable
1 Todos los valores están expresados en % de peso seco excepto el aceite refinado, que está en % de peso fresco (FSANZ, 2004).
Tabla III.3. Resumen del análisis de minerales de MXB-13 y de la semilla de algodón de control de 6 sitios (FSANZ, 2004).
Componente(mg/100 gm peso seco) MXB-131 Control Valor P de prueba t
pareada Valor P ajustado por Dunnet Rango de bibliografía2
Calcio160
[140–190]18,25
151[129–185]
20,890,076 0,178 108–210
Cobre0,93
[0,79–1,11]0,11
0,91[0,83–1,03]
0,080,829 0,992 0,4–1,19
Hierro5,59
[4,76–6,67]0,71
6,17[4,95–7,65]
1,000,099 0,227 3,79–15,1
Magnesio417
[370–450]35,14
421[377–461]
31,680,799 0,988 305–460
Manganeso1,51
[1,35–1,66]0,14
1,42[1,27–1,68]
0,150,149 0,328 1,0–2,0
Molibdeno<0,2
[<0,2]-
<0,2[<0,2]
-- - 0,1–0,4
Fósforo687
[590–769]61,39
699[579–869]
107,720,763 0,980 447–750
Potasio1219
[1109–1324]70,87
1237[1065–1371]
102,260,406 0,731 990–1280
Sodio26,5
[<10–40]19,16
15,6[<10–24]
7,25- - 3–38
Zinc4,43
[4,09–4,82]0,31
4,23[3,61–5,38]
0,620,247 0,502 2,49–4,2
Azufre275
[226–315]35,26
276[248–293]
16,650,857 0,996 144–260
1 Los valores exhibidos son el promedio (negrita), el rango (entre paréntesis) y la desviación estándar.2 Rango de bibliografía combinada.
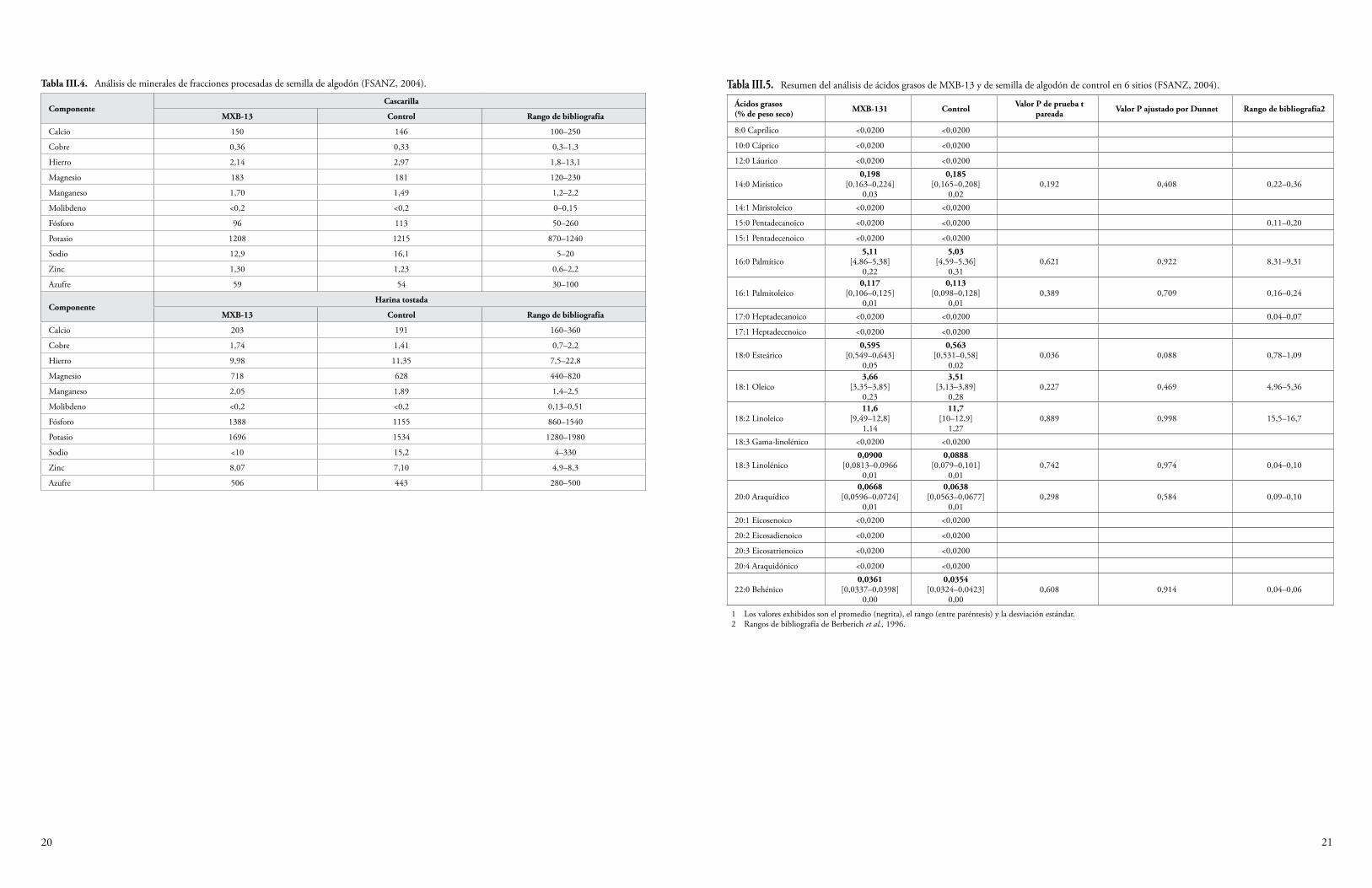
20 21
Tabla III.4. Análisis de minerales de fracciones procesadas de semilla de algodón (FSANZ, 2004).
ComponenteCascarilla
MXB-13 Control Rango de bibliografía
Calcio 150 146 100–250
Cobre 0,36 0,33 0,3–1,3
Hierro 2,14 2,97 1,8–13,1
Magnesio 183 181 120–230
Manganeso 1,70 1,49 1,2–2,2
Molibdeno <0,2 <0,2 0–0,15
Fósforo 96 113 50–260
Potasio 1208 1215 870–1240
Sodio 12,9 16,1 5–20
Zinc 1,30 1,23 0,6–2,2
Azufre 59 54 30–100
ComponenteHarina tostada
MXB-13 Control Rango de bibliografía
Calcio 203 191 160–360
Cobre 1,74 1,41 0,7–2,2
Hierro 9,98 11,35 7,5–22,8
Magnesio 718 628 440–820
Manganeso 2,05 1,89 1,4–2,5
Molibdeno <0,2 <0,2 0,13–0,51
Fósforo 1388 1155 860–1540
Potasio 1696 1534 1280–1980
Sodio <10 15,2 4–330
Zinc 8,07 7,10 4,9–8,3
Azufre 506 443 280–500
Tabla III.5. Resumen del análisis de ácidos grasos de MXB-13 y de semilla de algodón de control en 6 sitios (FSANZ, 2004).
Ácidos grasos(% de peso seco) MXB-131 Control Valor P de prueba t
pareada Valor P ajustado por Dunnet Rango de bibliografía2
8:0 Caprílico <0,0200 <0,0200
10:0 Cáprico <0,0200 <0,0200
12:0 Láurico <0,0200 <0,0200
14:0 Mirístico0,198
[0,163–0,224]0,03
0,185[0,165–0,208]
0,020,192 0,408 0,22–0,36
14:1 Miristoleico <0,0200 <0,0200
15:0 Pentadecanoico <0,0200 <0,0200 0,11–0,20
15:1 Pentadecenoico <0,0200 <0,0200
16:0 Palmítico5,11
[4,86–5,38]0,22
5,03[4,59–5,36]
0,310,621 0,922 8,31–9,31
16:1 Palmitoleico0,117
[0,106–0,125]0,01
0,113[0,098–0,128]
0,010,389 0,709 0,16–0,24
17:0 Heptadecanoico <0,0200 <0,0200 0,04–0,07
17:1 Heptadecenoico <0,0200 <0,0200
18:0 Esteárico0,595
[0,549–0,643]0,05
0,563[0,531–0,58]
0,020,036 0,088 0,78–1,09
18:1 Oleico3,66
[3,35–3,85]0,23
3,51[3,13–3,89]
0,280,227 0,469 4,96–5,36
18:2 Linoleico11,6
[9,49–12,8]1,14
11,7[10–12,9]
1,270,889 0,998 15,5–16,7
18:3 Gama-linolénico <0,0200 <0,0200
18:3 Linolénico0,0900
[0,0813–0,09660,01
0,0888[0,079–0,101]
0,010,742 0,974 0,04–0,10
20:0 Araquídico0,0668
[0,0596–0,0724]0,01
0,0638[0,0563–0,0677]
0,010,298 0,584 0,09–0,10
20:1 Eicosenoico <0,0200 <0,0200
20:2 Eicosadienoico <0,0200 <0,0200
20:3 Eicosatrienoico <0,0200 <0,0200
20:4 Araquidónico <0,0200 <0,0200
22:0 Behénico0,0361
[0,0337–0,0398]0,00
0,0354[0,0324–0,0423]
0,000,608 0,914 0,04–0,06
1 Los valores exhibidos son el promedio (negrita), el rango (entre paréntesis) y la desviación estándar.2 Rangos de bibliografía de Berberich et al., 1996.
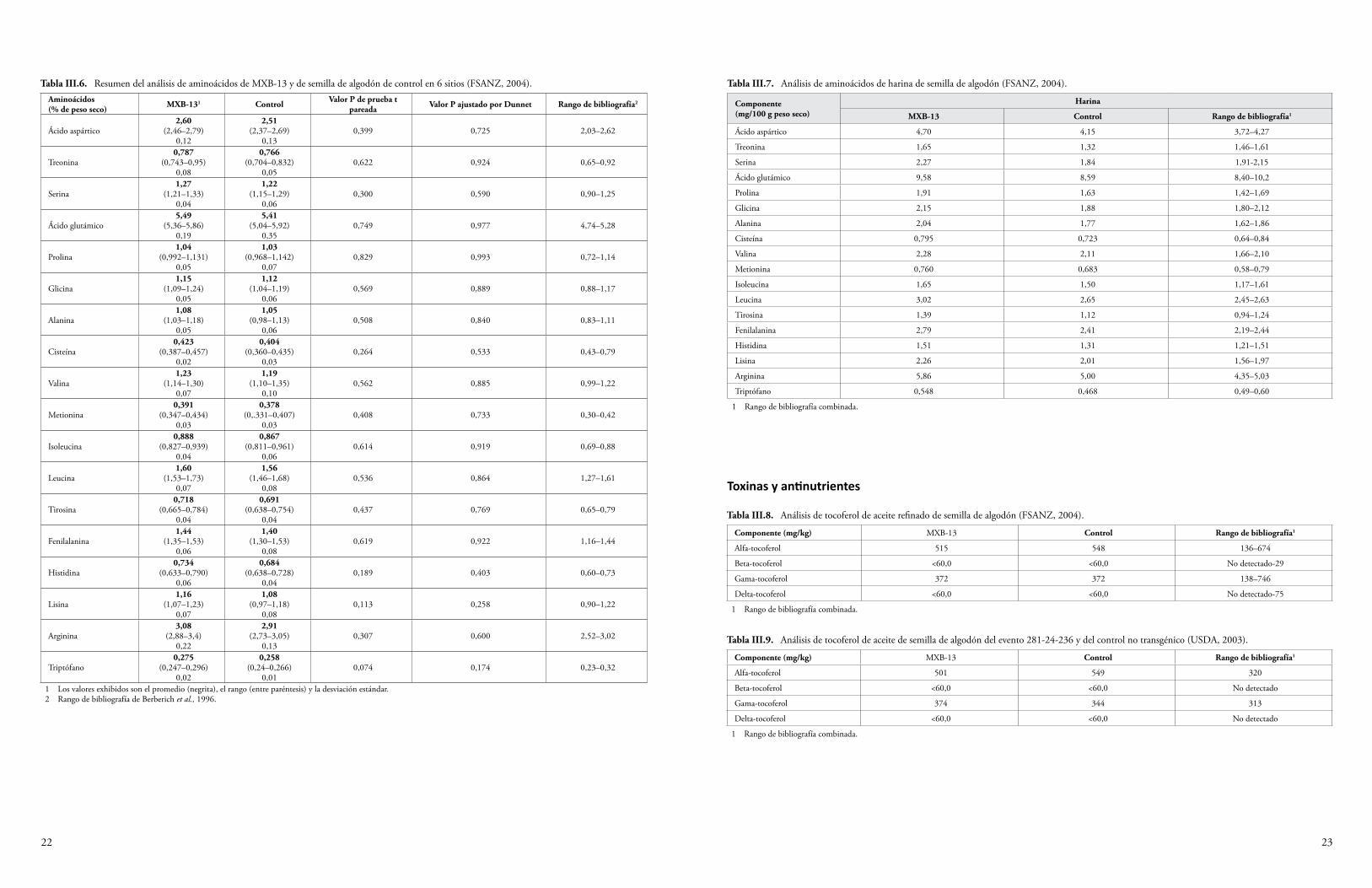
22 23
Tabla III.6. Resumen del análisis de aminoácidos de MXB-13 y de semilla de algodón de control en 6 sitios (FSANZ, 2004).Aminoácidos(% de peso seco) MXB-131 Control Valor P de prueba t
pareada Valor P ajustado por Dunnet Rango de bibliografía2
Ácido aspártico2,60
(2,46–2,79)0,12
2,51(2,37–2,69)
0,130,399 0,725 2,03–2,62
Treonina0,787
(0,743–0,95)0,08
0,766(0,704–0,832)
0,050,622 0,924 0,65–0,92
Serina1,27
(1,21–1,33)0,04
1,22(1,15–1,29)
0,060,300 0,590 0,90–1,25
Ácido glutámico5,49
(5,36–5,86)0,19
5,41(5,04–5,92)
0,350,749 0,977 4,74–5,28
Prolina1,04
(0,992–1,131)0,05
1,03(0,968–1,142)
0,070,829 0,993 0,72–1,14
Glicina1,15
(1,09–1,24)0,05
1,12(1,04–1,19)
0,060,569 0,889 0,88–1,17
Alanina1,08
(1,03–1,18)0,05
1,05(0,98–1,13)
0,060,508 0,840 0,83–1,11
Cisteína0,423
(0,387–0,457)0,02
0,404(0,360–0,435)
0,030,264 0,533 0,43–0,79
Valina1,23
(1,14–1,30)0,07
1,19(1,10–1,35)
0,100,562 0,885 0,99–1,22
Metionina0,391
(0,347–0,434)0,03
0,378(0,.331–0,407)
0,030,408 0,733 0,30–0,42
Isoleucina0,888
(0,827–0,939)0,04
0,867(0,811–0,961)
0,060,614 0,919 0,69–0,88
Leucina1,60
(1,53–1,73)0,07
1,56(1,46–1,68)
0,080,536 0,864 1,27–1,61
Tirosina0,718
(0,665–0,784)0,04
0,691(0,638–0,754)
0,040,437 0,769 0,65–0,79
Fenilalanina1,44
(1,35–1,53)0,06
1,40(1,30–1,53)
0,080,619 0,922 1,16–1,44
Histidina0,734
(0,633–0,790)0,06
0,684(0,638–0,728)
0,040,189 0,403 0,60–0,73
Lisina1,16
(1,07–1,23)0,07
1,08(0,97–1,18)
0,080,113 0,258 0,90–1,22
Arginina3,08
(2,88–3,4)0,22
2,91(2,73–3,05)
0,130,307 0,600 2,52–3,02
Triptófano0,275
(0,247–0,296)0,02
0,258(0,24–0,266)
0,010,074 0,174 0,23–0,32
1 Los valores exhibidos son el promedio (negrita), el rango (entre paréntesis) y la desviación estándar.2 Rango de bibliografía de Berberich et al., 1996.
Tabla III.7. Análisis de aminoácidos de harina de semilla de algodón (FSANZ, 2004).
Componente(mg/100 g peso seco)
Harina
MXB-13 Control Rango de bibliografía1
Ácido aspártico 4,70 4,15 3,72–4,27
Treonina 1,65 1,32 1,46–1,61
Serina 2,27 1,84 1,91-2,15
Ácido glutámico 9,58 8,59 8,40–10,2
Prolina 1,91 1,63 1,42–1,69
Glicina 2,15 1,88 1,80–2,12
Alanina 2,04 1,77 1,62–1,86
Cisteína 0,795 0,723 0,64–0,84
Valina 2,28 2,11 1,66–2,10
Metionina 0,760 0,683 0,58–0,79
Isoleucina 1,65 1,50 1,17–1,61
Leucina 3,02 2,65 2,45–2,63
Tirosina 1,39 1,12 0,94–1,24
Fenilalanina 2,79 2,41 2,19–2,44
Histidina 1,51 1,31 1,21–1,51
Lisina 2,26 2,01 1,56–1,97
Arginina 5,86 5,00 4,35–5,03
Triptófano 0,548 0,468 0,49–0,60
1 Rango de bibliografía combinada.
Toxinas y antinutrientes
Tabla III.8. Análisis de tocoferol de aceite refinado de semilla de algodón (FSANZ, 2004).
Componente (mg/kg) MXB-13 Control Rango de bibliografía1
Alfa-tocoferol 515 548 136–674
Beta-tocoferol <60,0 <60,0 No detectado-29
Gama-tocoferol 372 372 138–746
Delta-tocoferol <60,0 <60,0 No detectado-75
1 Rango de bibliografía combinada.
Tabla III.9. Análisis de tocoferol de aceite de semilla de algodón del evento 281-24-236 y del control no transgénico (USDA, 2003).
Componente (mg/kg) MXB-13 Control Rango de bibliografía1
Alfa-tocoferol 501 549 320
Beta-tocoferol <60,0 <60,0 No detectado
Gama-tocoferol 374 344 313
Delta-tocoferol <60,0 <60,0 No detectado
1 Rango de bibliografía combinada.
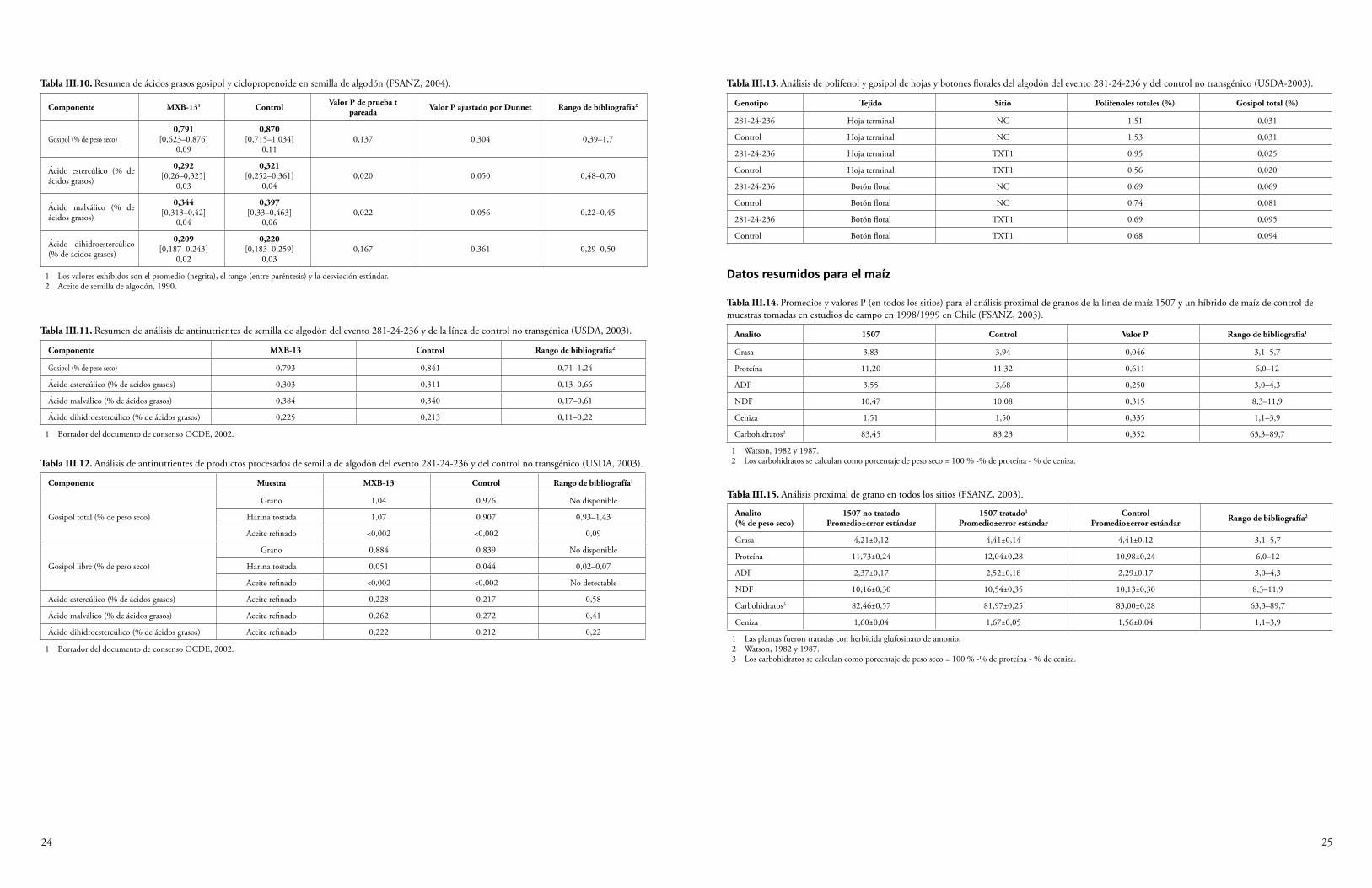
24 25
Tabla III.10. Resumen de ácidos grasos gosipol y ciclopropenoide en semilla de algodón (FSANZ, 2004).
Componente MXB-131 Control Valor P de prueba t pareada Valor P ajustado por Dunnet Rango de bibliografía2
Gosipol (% de peso seco)0,791
[0,623–0,876]0,09
0,870[0,715–1,034]
0,110,137 0,304 0,39–1,7
Ácido estercúlico (% de ácidos grasos)
0,292[0,26–0,325]
0,03
0,321[0,252–0,361]
0,040,020 0,050 0,48–0,70
Ácido malválico (% de ácidos grasos)
0,344[0,313–0,42]
0,04
0,397[0,33–0,463]
0,060,022 0,056 0,22–0,45
Ácido dihidroestercúlico (% de ácidos grasos)
0,209[0,187–0,243]
0,02
0,220[0,183–0,259]
0,030,167 0,361 0,29–0,50
1 Los valores exhibidos son el promedio (negrita), el rango (entre paréntesis) y la desviación estándar.2 Aceite de semilla de algodón, 1990.
Tabla III.11. Resumen de análisis de antinutrientes de semilla de algodón del evento 281-24-236 y de la línea de control no transgénica (USDA, 2003).
Componente MXB-13 Control Rango de bibliografía2
Gosipol (% de peso seco) 0,793 0,841 0,71–1,24
Ácido estercúlico (% de ácidos grasos) 0,303 0,311 0,13–0,66
Ácido malválico (% de ácidos grasos) 0,384 0,340 0,17–0,61
Ácido dihidroestercúlico (% de ácidos grasos) 0,225 0,213 0,11–0,22
1 Borrador del documento de consenso OCDE, 2002.
Tabla III.12. Análisis de antinutrientes de productos procesados de semilla de algodón del evento 281-24-236 y del control no transgénico (USDA, 2003).
Componente Muestra MXB-13 Control Rango de bibliografía1
Gosipol total (% de peso seco)
Grano 1,04 0,976 No disponible
Harina tostada 1,07 0,907 0,93–1,43
Aceite refinado <0,002 <0,002 0,09
Gosipol libre (% de peso seco)
Grano 0,884 0,839 No disponible
Harina tostada 0,051 0,044 0,02–0,07
Aceite refinado <0,002 <0,002 No detectable
Ácido estercúlico (% de ácidos grasos) Aceite refinado 0,228 0,217 0,58
Ácido malválico (% de ácidos grasos) Aceite refinado 0,262 0,272 0,41
Ácido dihidroestercúlico (% de ácidos grasos) Aceite refinado 0,222 0,212 0,22
1 Borrador del documento de consenso OCDE, 2002.
Tabla III.13. Análisis de polifenol y gosipol de hojas y botones florales del algodón del evento 281-24-236 y del control no transgénico (USDA-2003).
Genotipo Tejido Sitio Polifenoles totales (%) Gosipol total (%)
281-24-236 Hoja terminal NC 1,51 0,031
Control Hoja terminal NC 1,53 0,031
281-24-236 Hoja terminal TXT1 0,95 0,025
Control Hoja terminal TXT1 0,56 0,020
281-24-236 Botón floral NC 0,69 0,069
Control Botón floral NC 0,74 0,081
281-24-236 Botón floral TXT1 0,69 0,095
Control Botón floral TXT1 0,68 0,094
Datos resumidos para el maíz
Tabla III.14. Promedios y valores P (en todos los sitios) para el análisis proximal de granos de la línea de maíz 1507 y un híbrido de maíz de control de muestras tomadas en estudios de campo en 1998/1999 en Chile (FSANZ, 2003).
Analito 1507 Control Valor P Rango de bibliografía1
Grasa 3,83 3,94 0,046 3,1–5,7
Proteína 11,20 11,32 0,611 6,0–12
ADF 3,55 3,68 0,250 3,0–4,3
NDF 10,47 10,08 0,315 8,3–11,9
Ceniza 1,51 1,50 0,335 1,1–3,9
Carbohidratos2 83,45 83,23 0,352 63,3–89,7
1 Watson, 1982 y 1987.2 Los carbohidratos se calculan como porcentaje de peso seco = 100 % -% de proteína - % de ceniza.
Tabla III.15. Análisis proximal de grano en todos los sitios (FSANZ, 2003).
Analito (% de peso seco)
1507 no tratadoPromedio±error estándar
1507 tratado1
Promedio±error estándarControl
Promedio±error estándar Rango de bibliografía2
Grasa 4,21±0,12 4,41±0,14 4,41±0,12 3,1–5,7
Proteína 11,73±0,24 12,04±0,28 10,98±0,24 6,0–12
ADF 2,37±0,17 2,52±0,18 2,29±0,17 3,0–4,3
NDF 10,16±0,30 10,54±0,35 10,13±0,30 8,3–11,9
Carbohidratos3 82,46±0,57 81,97±0,25 83,00±0,28 63,3–89,7
Ceniza 1,60±0,04 1,67±0,05 1,56±0,04 1,1–3,9
1 Las plantas fueron tratadas con herbicida glufosinato de amonio.2 Watson, 1982 y 1987.3 Los carbohidratos se calculan como porcentaje de peso seco = 100 % -% de proteína - % de ceniza.
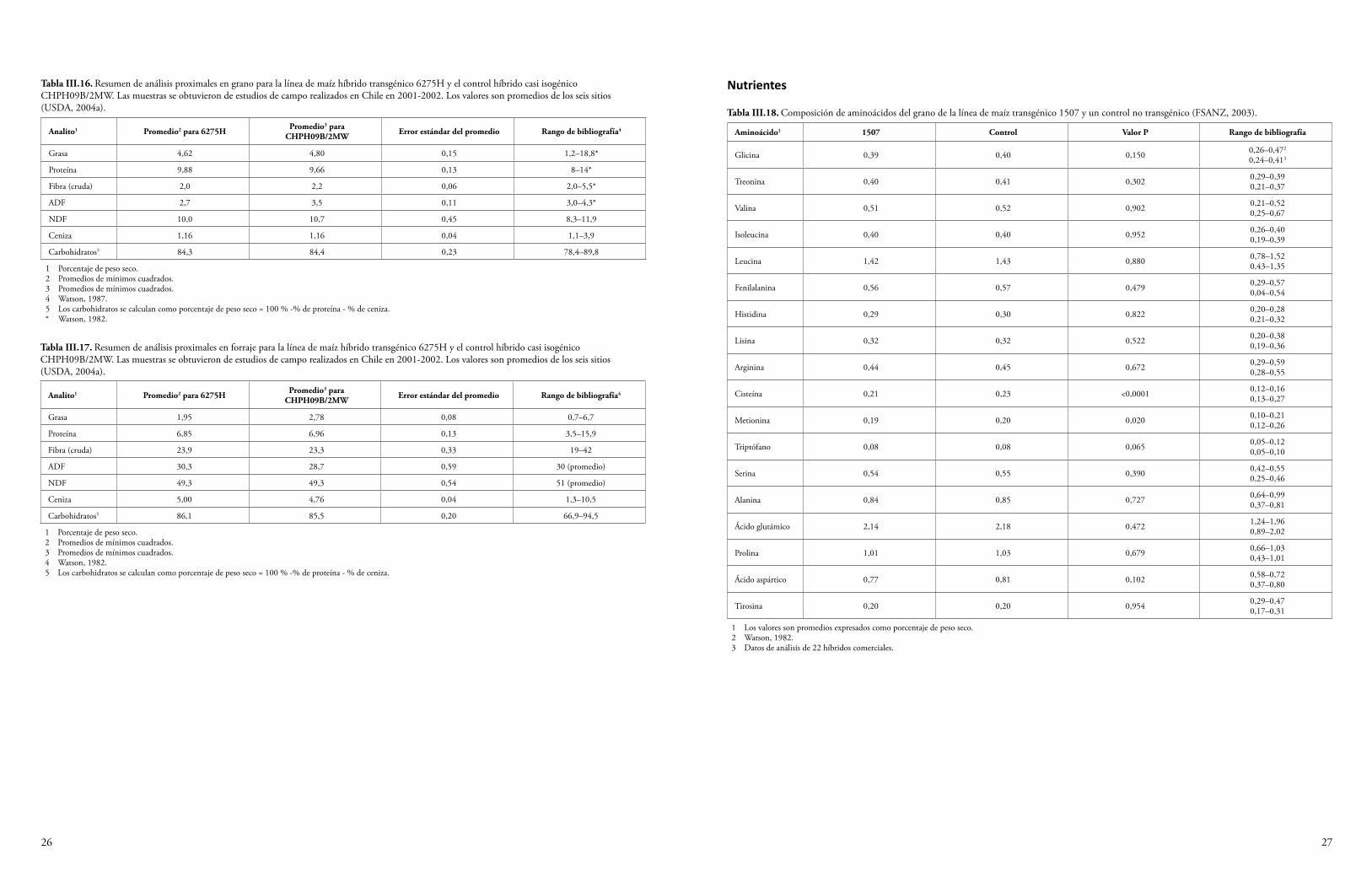
26 27
Tabla III.16. Resumen de análisis proximales en grano para la línea de maíz híbrido transgénico 6275H y el control híbrido casi isogénico CHPH09B/2MW. Las muestras se obtuvieron de estudios de campo realizados en Chile en 2001-2002. Los valores son promedios de los seis sitios (USDA, 2004a).
Analito1 Promedio2 para 6275H Promedio3 para CHPH09B/2MW Error estándar del promedio Rango de bibliografía4
Grasa 4,62 4,80 0,15 1,2–18,8*
Proteína 9,88 9,66 0,13 8–14*
Fibra (cruda) 2,0 2,2 0,06 2,0–5,5*
ADF 2,7 3,5 0,11 3,0–4,3*
NDF 10,0 10,7 0,45 8,3–11,9
Ceniza 1,16 1,16 0,04 1,1–3,9
Carbohidratos5 84,3 84,4 0,23 78,4–89,8
1 Porcentaje de peso seco.2 Promedios de mínimos cuadrados.3 Promedios de mínimos cuadrados.4 Watson, 1987.5 Los carbohidratos se calculan como porcentaje de peso seco = 100 % -% de proteína - % de ceniza.* Watson, 1982.
Tabla III.17. Resumen de análisis proximales en forraje para la línea de maíz híbrido transgénico 6275H y el control híbrido casi isogénico CHPH09B/2MW. Las muestras se obtuvieron de estudios de campo realizados en Chile en 2001-2002. Los valores son promedios de los seis sitios (USDA, 2004a).
Analito1 Promedio2 para 6275H Promedio3 para CHPH09B/2MW Error estándar del promedio Rango de bibliografía4
Grasa 1,95 2,78 0,08 0,7–6,7
Proteína 6,85 6,96 0,13 3,5–15,9
Fibra (cruda) 23,9 23,3 0,33 19–42
ADF 30,3 28,7 0,59 30 (promedio)
NDF 49,3 49,3 0,54 51 (promedio)
Ceniza 5,00 4,76 0,04 1,3–10,5
Carbohidratos5 86,1 85,5 0,20 66,9–94,5
1 Porcentaje de peso seco.2 Promedios de mínimos cuadrados.3 Promedios de mínimos cuadrados.4 Watson, 1982.5 Los carbohidratos se calculan como porcentaje de peso seco = 100 % -% de proteína - % de ceniza.
Tabla III.18. Composición de aminoácidos del grano de la línea de maíz transgénico 1507 y un control no transgénico (FSANZ, 2003).
Aminoácido1 1507 Control Valor P Rango de bibliografía
Glicina 0,39 0,40 0,150 0,26–0,472
0,24–0,413
Treonina 0,40 0,41 0,302 0,29–0,390,21–0,37
Valina 0,51 0,52 0,902 0,21–0,520,25–0,67
Isoleucina 0,40 0,40 0,952 0,26–0,400,19–0,39
Leucina 1,42 1,43 0,880 0,78–1,520,43–1,35
Fenilalanina 0,56 0,57 0,479 0,29–0,570,04–0,54
Histidina 0,29 0,30 0,822 0,20–0,280,21–0,32
Lisina 0,32 0,32 0,522 0,20–0,380,19–0,36
Arginina 0,44 0,45 0,672 0,29–0,590,28–0,55
Cisteína 0,21 0,23 <0,0001 0,12–0,160,13–0,27
Metionina 0,19 0,20 0,020 0,10–0,210,12–0,26
Triptófano 0,08 0,08 0,065 0,05–0,120,05–0,10
Serina 0,54 0,55 0,390 0,42–0,550,25–0,46
Alanina 0,84 0,85 0,727 0,64–0,990,37–0,81
Ácido glutámico 2,14 2,18 0,472 1,24–1,960,89–2,02
Prolina 1,01 1,03 0,679 0,66–1,030,43–1,01
Ácido aspártico 0,77 0,81 0,102 0,58–0,720,37–0,80
Tirosina 0,20 0,20 0,954 0,29–0,470,17–0,31
1 Los valores son promedios expresados como porcentaje de peso seco.2 Watson, 1982.3 Datos de análisis de 22 híbridos comerciales.
Nutrientes
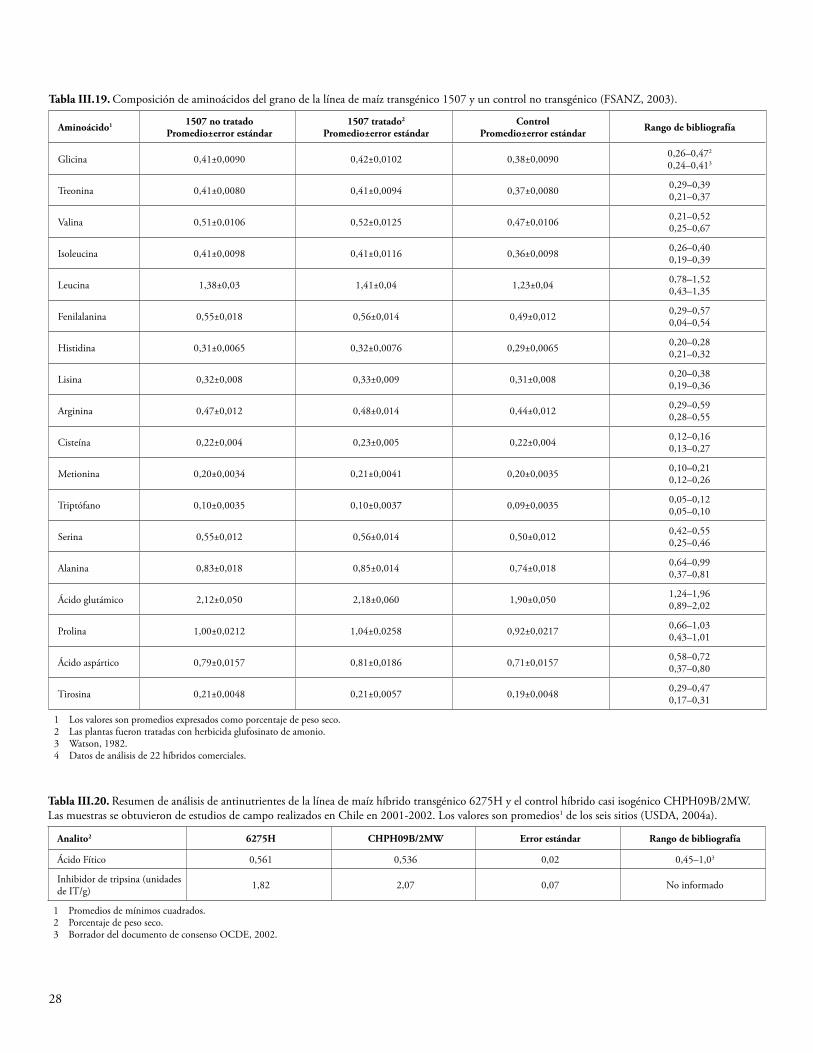
28
Tabla III.19. Composición de aminoácidos del grano de la línea de maíz transgénico 1507 y un control no transgénico (FSANZ, 2003).
Aminoácido1 1507 no tratadoPromedio±error estándar
1507 tratado2
Promedio±error estándarControl
Promedio±error estándar Rango de bibliografía
Glicina 0,41±0,0090 0,42±0,0102 0,38±0,0090 0,26–0,472
0,24–0,413
Treonina 0,41±0,0080 0,41±0,0094 0,37±0,0080 0,29–0,390,21–0,37
Valina 0,51±0,0106 0,52±0,0125 0,47±0,0106 0,21–0,520,25–0,67
Isoleucina 0,41±0,0098 0,41±0,0116 0,36±0,0098 0,26–0,400,19–0,39
Leucina 1,38±0,03 1,41±0,04 1,23±0,04 0,78–1,520,43–1,35
Fenilalanina 0,55±0,018 0,56±0,014 0,49±0,012 0,29–0,570,04–0,54
Histidina 0,31±0,0065 0,32±0,0076 0,29±0,0065 0,20–0,280,21–0,32
Lisina 0,32±0,008 0,33±0,009 0,31±0,008 0,20–0,380,19–0,36
Arginina 0,47±0,012 0,48±0,014 0,44±0,012 0,29–0,590,28–0,55
Cisteína 0,22±0,004 0,23±0,005 0,22±0,004 0,12–0,160,13–0,27
Metionina 0,20±0,0034 0,21±0,0041 0,20±0,0035 0,10–0,210,12–0,26
Triptófano 0,10±0,0035 0,10±0,0037 0,09±0,0035 0,05–0,120,05–0,10
Serina 0,55±0,012 0,56±0,014 0,50±0,012 0,42–0,550,25–0,46
Alanina 0,83±0,018 0,85±0,014 0,74±0,018 0,64–0,990,37–0,81
Ácido glutámico 2,12±0,050 2,18±0,060 1,90±0,050 1,24–1,960,89–2,02
Prolina 1,00±0,0212 1,04±0,0258 0,92±0,0217 0,66–1,030,43–1,01
Ácido aspártico 0,79±0,0157 0,81±0,0186 0,71±0,0157 0,58–0,720,37–0,80
Tirosina 0,21±0,0048 0,21±0,0057 0,19±0,0048 0,29–0,470,17–0,31
1 Los valores son promedios expresados como porcentaje de peso seco.2 Las plantas fueron tratadas con herbicida glufosinato de amonio.3 Watson, 1982.4 Datos de análisis de 22 híbridos comerciales.
Tabla III.20. Resumen de análisis de antinutrientes de la línea de maíz híbrido transgénico 6275H y el control híbrido casi isogénico CHPH09B/2MW. Las muestras se obtuvieron de estudios de campo realizados en Chile en 2001-2002. Los valores son promedios1 de los seis sitios (USDA, 2004a).
Analito2 6275H CHPH09B/2MW Error estándar Rango de bibliografía
Ácido Fítico 0,561 0,536 0,02 0,45–1,03
Inhibidor de tripsina (unidades de IT/g) 1,82 2,07 0,07 No informado
1 Promedios de mínimos cuadrados.2 Porcentaje de peso seco.3 Borrador del documento de consenso OCDE, 2002.


















