
regions Specific a-chain B-cell - pnas.org · 3810 Immunology: Blackman and Koshland P. Blair,...
5
Proc. Nati. Acad. Sci. USA Vol. 82, pp. 3809-3813, June 1985 Immunology Specific 5' and 3' regions of the a-chain gene are undermethylated at distinct stages of B-cell differentiation (immunoglobulin p-chain gene regulation/DNA methylation/enhancer activation/shift of membrane to secreted forms of p chain/ chromatin structure) MARCIA A. BLACKMAN AND MARIAN ELLIOTT KOSHLAND Department of Microbiology and Immunology, University of California, Berkeley, CA 94720 Contributed by Marian Elliott Koshland, February 1, 1985 ABSTRACT The p-chain gene is expressed differently in successive stages of B-lymphocyte development. The heavy chain product appears as a cytoplasmic constituent in pre-B- cells, as part of the IgM receptor in maturing B cells, and as a component in the pentamer IgM antibody synthesized and secreted by the antigen-stimulated cell. We have used the methylation of CpG sequences as an assay system to define the chromatin changes associated with different expression of the p-chain. The methylation status of eight index sites was followed by restriction enzyme analysis of murine cell lines representing the major stages in the developmental pathway. The analyses showed that a single Msp I/Hpa II site 5' to the immunoglobulin enhancer becomes undermethylated with the onset of p-chain gene transcription. Four midgene Msp I/Hpa II sites exhibit a progressive loss of methyl groups unrelated to changes in p-chain gene expression, whereas a Msp I/Hpa II site and two Hha I sites surrounding the exon encoding the carboxyl terminus of the secreted form of pL chain (ps) become undermethylated during the transition to IgM secretion. These results indicate that structural changes in local regions of the p-chain gene correlate with specific developmental events. Although many of the complex events associated with immu- noglobulin gene expression have been defined, little is known of the mechanisms that regulate these events. The mechanisms operating on the pL heavy chain gene are of particular interest because the gene is expressed through several stages of B-cell differentiation, and at each stage the heavy chain product plays a critical role in the differentiative process. Thus, in the precursor B cell, the p-chain gene locus is the first to undergo rearrangement of the variable (V), diversity (D), and joining (J) gene segments, and the synthesis of a cytoplasmic form of p chain is a signal for rearrangement to proceed at a light chain locus. In the maturing B cell, synthesis of the membrane form of p chain (Am) allows monomeric IgM to be inserted in the lipid bilayer and serve as an antigen receptor. Finally, in the anti- gen-stimulated B blast, the switch in synthesis from the mem- brane (,,m) to the secreted (p,) form of pt chain permits the assembly of pentamer IgM and the secretion of the pentamer antibody from the cell (1). Analyses of p-chain gene expression have identified two structures involved in transcriptional control: a promoter sequence located 5' to the V region (2) and an enhancer sequence located within the JH-CM intron (JH, heavy chain gene J regions; CM, constant regions of the p-chain gene) (3, 4). The promoter and its flanking V gene are transcriptionally silent in the germ-line configuration and become activated only when they are brought within the proximity of the enhancer by a productive V-D-J rearrangement (5). The activity of the enhancer, on the other hand, appears to be independent of productive V-D-J rearrangement because sterile transcripts of unrearranged or incompletely rear- ranged g-chain genes are found not only in pre-B-cells (6) but also in T cells (7). Analyses of g-chain gene expression have also identified the switch from the gm to the Au form of A chain as a post-transcriptional event. The two forms have been shown to differ only in their carboxyl termini (8), which are encoded in separate exons located 3' to C. (9). Both exons are cotranscribed with the gene, and separate Am- and A,-chain mRNAs are then generated from the primary transcript by differential 3' cleavage and processing (10, 11). These data suggest that the form of A chain expressed is regulated through changes in the higher-order structure of the primary transcript that determine the relative accessibility of the cleavage and polyadenylylation sites 3' to the A. and Pm exons. In the studies reported here, the mechanisms of p-chain gene regulation were pursued by examining the methylation of the p-chain gene during B-cell differentiation. Most of the methyla- tion in mammalian DNA occurs at position 5 of cytosine in the symmetrical dinucleotide CpG. Considerable evidence has accumulated that the loss of methyl groups from these sites is an index of changes in DNA conformation. Hypomethylation has been shown to destabilize Z-DNA (12) and to facilitate the denaturation of double-stranded DNA (13). In many eukaryotic systems, hypomethylation has been found to correlate with the activation of gene expression (14-16), and it frequently ac- companies the development of DNaseI-hypersensitive sites (17-19). Thus, the loss of methyl groups is thought to define areas of altered, more open chromatin structure associated with transcriptional competence. The strategy used in these studies was to compare the methylation status of the 5' flanking and coding sequence of the C. gene in tissues and cell lines representative of successive stages in B-cell differentiation. The comparison was made by digesting high molecular weight DNA from the various cells with EcoRI and then with enzymes that are inhibited by the presence of methylcytosine within their restriction sites. The cleavage patterns obtained show that specific developmental events at the C. locus correlate with a stepwise loss of methyl groups, first in the 5' sequence and then in the 3' sequence. MATERIALS AND METHODS The B-cell lines used in these studies are listed in Table 1 and have been described (20). The T-cell lines WEHI 7.1 and WEHI 22.1 were obtained from the American Type Culture Collection. YAC-1, EL-4, and BW5147 were obtained from Abbreviations: uPm and As membrane and secreted forms of p chains; A, and um exons, exons encoding the different carboxyl termini of the fIL,. and Am chains; C, constant; V, variable; D, diversity; J, joining; kb, kilobase(s). 3809 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of regions Specific a-chain B-cell - pnas.org · 3810 Immunology: Blackman and Koshland P. Blair,...

Proc. Nati. Acad. Sci. USAVol. 82, pp. 3809-3813, June 1985Immunology
Specific 5' and 3' regions of the a-chain gene are undermethylatedat distinct stages of B-cell differentiation
(immunoglobulin p-chain gene regulation/DNA methylation/enhancer activation/shift of membrane to secreted forms of p chain/chromatin structure)
MARCIA A. BLACKMAN AND MARIAN ELLIOTT KOSHLANDDepartment of Microbiology and Immunology, University of California, Berkeley, CA 94720
Contributed by Marian Elliott Koshland, February 1, 1985
ABSTRACT The p-chain gene is expressed differently insuccessive stages of B-lymphocyte development. The heavychain product appears as a cytoplasmic constituent in pre-B-cells, as part of the IgM receptor in maturing B cells, and as acomponent in the pentamer IgM antibody synthesized andsecreted by the antigen-stimulated cell. We have used themethylation of CpG sequences as an assay system to define thechromatin changes associated with different expression of thep-chain. The methylation status of eight index sites wasfollowed by restriction enzyme analysis of murine cell linesrepresenting the major stages in the developmental pathway.The analyses showed that a single Msp I/Hpa II site 5' to theimmunoglobulin enhancer becomes undermethylated with theonset of p-chain gene transcription. Four midgene Msp I/HpaII sites exhibit a progressive loss of methyl groups unrelated tochanges in p-chain gene expression, whereas a Msp I/Hpa IIsite and two Hha I sites surrounding the exon encoding thecarboxyl terminus of the secreted form of pL chain (ps) becomeundermethylated during the transition to IgM secretion. Theseresults indicate that structural changes in local regions of thep-chain gene correlate with specific developmental events.
Although many of the complex events associated with immu-noglobulin gene expression have been defined, little is known ofthe mechanisms that regulate these events. The mechanismsoperating on the pL heavy chain gene are of particular interestbecause the gene is expressed through several stages of B-celldifferentiation, and at each stage the heavy chain product playsa critical role in the differentiative process. Thus, in theprecursor B cell, the p-chain gene locus is the first to undergorearrangement of the variable (V), diversity (D), and joining (J)gene segments, and the synthesis of a cytoplasmic form of pchain is a signal for rearrangement to proceed at a light chainlocus. In the maturing B cell, synthesis of the membrane formofp chain (Am) allows monomeric IgM to be inserted in the lipidbilayer and serve as an antigen receptor. Finally, in the anti-gen-stimulated B blast, the switch in synthesis from the mem-brane (,,m) to the secreted (p,) form of pt chain permits theassembly of pentamer IgM and the secretion of the pentamerantibody from the cell (1).Analyses of p-chain gene expression have identified two
structures involved in transcriptional control: a promotersequence located 5' to the V region (2) and an enhancersequence located within the JH-CM intron (JH, heavy chaingene J regions; CM, constant regions of the p-chain gene) (3,4). The promoter and its flanking V gene are transcriptionallysilent in the germ-line configuration and become activatedonly when they are brought within the proximity of theenhancer by a productive V-D-J rearrangement (5). Theactivity of the enhancer, on the other hand, appears to be
independent of productive V-D-J rearrangement becausesterile transcripts of unrearranged or incompletely rear-ranged g-chain genes are found not only in pre-B-cells (6) butalso in T cells (7).Analyses of g-chain gene expression have also identified
the switch from the gm to the Au form of A chain as apost-transcriptional event. The two forms have been shownto differ only in their carboxyl termini (8), which are encodedin separate exons located 3' to C. (9). Both exons arecotranscribed with the gene, and separate Am- and A,-chainmRNAs are then generated from the primary transcript bydifferential 3' cleavage and processing (10, 11). These datasuggest that the form of A chain expressed is regulatedthrough changes in the higher-order structure of the primarytranscript that determine the relative accessibility of thecleavage and polyadenylylation sites 3' to the A. and Pmexons.
In the studies reported here, the mechanisms ofp-chain generegulation were pursued by examining the methylation of thep-chain gene during B-cell differentiation. Most ofthe methyla-tion in mammalian DNA occurs at position 5 of cytosine in thesymmetrical dinucleotide CpG. Considerable evidence hasaccumulated that the loss of methyl groups from these sites isan index of changes in DNA conformation. Hypomethylationhas been shown to destabilize Z-DNA (12) and to facilitate thedenaturation ofdouble-stranded DNA (13). In many eukaryoticsystems, hypomethylation has been found to correlate with theactivation of gene expression (14-16), and it frequently ac-companies the development of DNaseI-hypersensitive sites(17-19). Thus, the loss of methyl groups is thought to defineareas ofaltered, more open chromatin structure associated withtranscriptional competence.The strategy used in these studies was to compare the
methylation status of the 5' flanking and coding sequence ofthe C. gene in tissues and cell lines representative ofsuccessive stages in B-cell differentiation. The comparisonwas made by digesting high molecular weight DNA from thevarious cells with EcoRI and then with enzymes that areinhibited by the presence of methylcytosine within theirrestriction sites. The cleavage patterns obtained show thatspecific developmental events at the C. locus correlate witha stepwise loss of methyl groups, first in the 5' sequence andthen in the 3' sequence.
MATERIALS AND METHODSThe B-cell lines used in these studies are listed in Table 1 andhave been described (20). The T-cell lines WEHI 7.1 andWEHI 22.1 were obtained from the American Type CultureCollection. YAC-1, EL-4, and BW5147 were obtained from
Abbreviations: uPm and As membrane and secreted forms ofp chains;A, and um exons, exons encoding the different carboxyl termini of thefIL,. and Am chains; C, constant; V, variable; D, diversity; J, joining;kb, kilobase(s).
3809
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

3810 Immunology: Blackman and Koshland
P. Blair, R. Mishell, and H. Sakano, respectively (Universityof California, Berkeley). The T-cell lines have been described(7). Liver and 9-day embryo tissues were obtained fromBALB/c mice.
Standard methods were used for DNA analysis and aredescribed in the text or have been published (20). Cytoplas-mic RNA dot-hybridization was performed as described byWhite and Bancroft (21) with the exception that cells were
lysed with a final concentration of 0.1% Nonidet P-40. Thehybridization probes are described in Fig. 1.
EcoRI + Msp Ia b c d e
EcoRI + HpalIf a b c d e f
-3.0- _
RESULTS
The methylation pattern of the C,, gene and its 5' flankingsequence was analyzed first by use of the restrictionendonucleases Msp I and Hpa II. Both enzymes recognizethe same 5' C-C-G-G 3' sequence, but only Hpa II is sensitiveto the presence of a methyl group on the inner cytosine. Thus,digestion with Msp I provides a control for the location andaccessibility of the restriction site, whereas digestion withHpa II indicates the extent to which the site is methylated.Six C-C-G-G sequences are encoded between the JH4 geneand the ,Um exons of the C,, gene (Fig. 1); each was examinedfor changes in methylation by analyzing DNA from therepresentative tissues and cell lines listed in Table 1. Afterpretreatment with EcoRI, the DNA samples were digestedwith either Msp I or Hpa II, and the fragments generatedwere assayed by hybridization with a site-specific probe(described in Fig. 1). This system allowed the extent ofundermethylation at each site or pair of adjacent sites to bequantitated by comparing the amount of diagnostic fragmentreleased by Hpa II digestion to the amount released bycomplete digestion with Msp I.
Methylation of the Region 5' to the Enhancer. The analysesof the Msp I/Hpa II site 5' to the enhancer sequence are
shown in Fig. 2. Digestion with Msp I gave the patternexpected for quantitative cleavage of the site; in all of theDNA preparations examined, only a single band of 1.1kilobases (kb) hybridized to the Msp I-EcoRI fragment usedas probe (probe 1, Fig. 1). However, digestion with Hpa IIrevealed a clearcut difference in the restriction patterns. IntheDNA from nonlymphoid tissues (liver and 9-day embryo),the Msp I/Hpa II site nearest the enhancer was resistant todigestion. The blots showed a major band of 6.4 kb that wasgenerated by EcoRI cleavage, a second major band of 3.0 kbthat was generated by additional cleavage at the Msp I/HpaII site 5' to the JH gene segments, and a minor band of 1.1 kbthat represents 6% cleavage of the Msp I/Hpa II site 5' to theenhancer (Table 1). Conversely, in the DNA from lymphoidlines actively transcribing the A-chain gene, the Msp I/HpaII site nearest the enhancer was completely susceptible to
Ikb JH E switch C,, )is Pm
RI M M RI M M MM RI,,
Hh Hh Hh Hh Hh
i- EcoRI6.4 ECORI 12.5
2 3 4 5
FIG. 1. The murine C,. gene and its 5' flanking sequences. R,EcoRI; M, Msp I/Hpa II; Hh, Hha I [indicated only in the12.5-kilobase (kb) EcoRI fragment]; E, enhancer. Restriction siteswere determined from the nucleotide sequence (22-24) and fromrestriction mapping (Michael Tigges, personal communication).Probe 1 was subcloned from XMEP203, a JH genomic clone (25).Probes 2-5 were isolated or subcloned from pXH-5, a C,. genomicclone.
_ Im_ _ 00iS -1.1
M M.. . ..m
...4.. .N.O
E
probe
-3.0OL
6.4
FIG. 2. Methylation of the Msp I/Hpa II site 5' to the enhancerin B-cell lines. DNA was digested with EcoRI followed by either MspI or Hpa II. Samples were electrophoresed (10 ,ug per lane) on 0.8%agarose gels and hybridized with probe 1 (Fig. 1). Lanes: a, livercells; b, embryo; c, WEHI 231.2; d, K46R; e, MXW 231.1a.2; f,MOPC 104E.
Hpa II digestion, resulting in patterns similar to those of theMsp I controls.The finding that undermethylation in the JH4-enhancer
region correlated with p-chain gene transcription could beinterpreted in two ways. The loss of methyl groups couldreflect chromatin changes associated with V-D-J rearrange-ment, or, alternatively, chromatin changes associated withactivation of enhancer function. As an approach to resolvingthese alternatives, the methylation analyses were extended toT-cell lines. This system had the advantage that ,t-chain genetranscripts are often produced in the absence of gene re-arrangement, thus allowing the two events to be indepen-dently evaluated. Of the five representative T-cell linesexamined, the DNA from only two was detectably under-methylated at the Msp I/Hpa II site 5' to the enhancer (Fig.3A); in the EL-4 line (lane b) and the WEHI 22.1 line (lane e),13% and 5% of the sites, respectively, were cleaved by HpaII, whereas in the other DNA preparations no cleavage wasobserved. The undermethylation in the EL-4 and WEHI 22.1
Table 1. Densitometric quantitation of undermethylation atspecific C-C-G-G sites
% cleavage by Hpa II
Source of DNA Probe 1 Probe 2 Probe 3 Probe 4
NonlymphoidLiver 6 13 17 1Embryo 6 30 9 0
B-lymphoma lines70Z 100 54 ND NDWEHI 231.2 100 65 11 038C13 100 95 41 0K46R 100 100 60 2
IgM-secreting linesMxW 231.1a.2 100 100 100 92MOPC 104E 100 100 100 56
Proc. Natl. Acad. Sci. USA 82 (1985)
Eco RI ----A
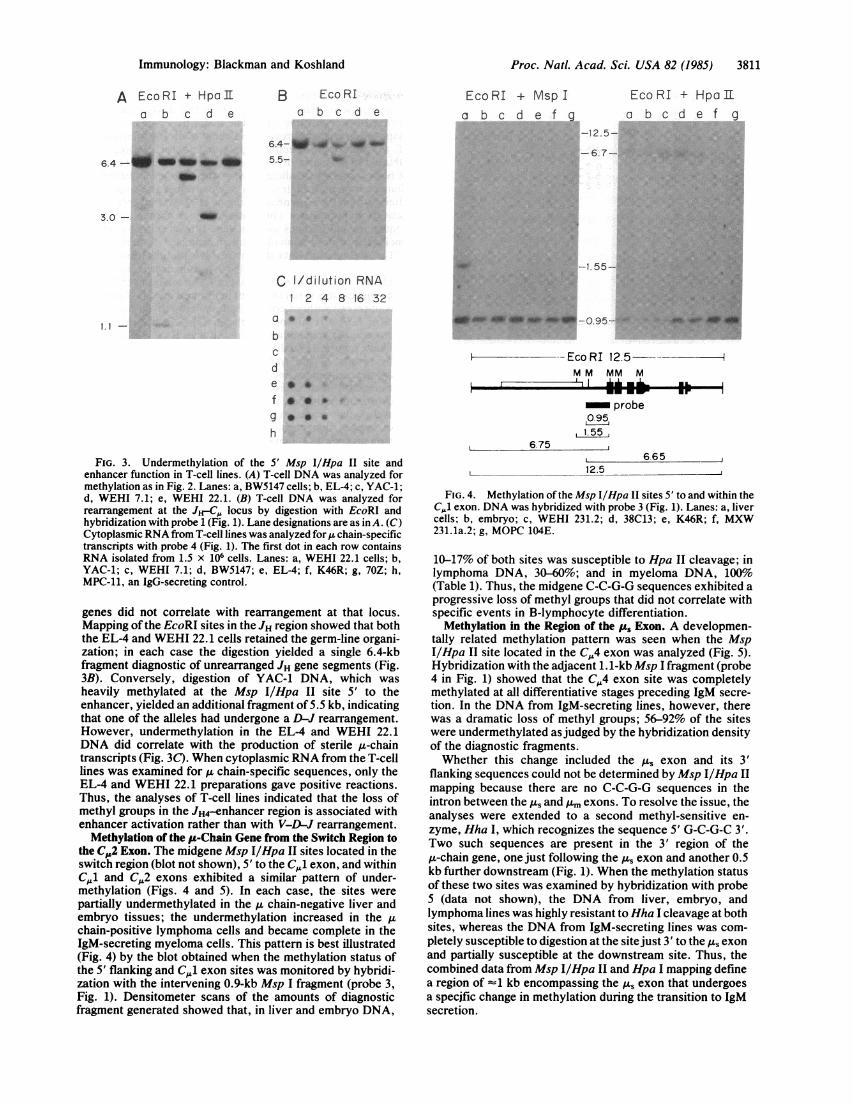
Proc. Natl. Acad. Sci. USA 82 (1985) 3811
B Eco RIa b c d e
6.4-UW _'le05.5- _
3.0 -
C 1/dilution RNA1 2 4 8 16 32
1. I -a *
bC
de
f vgee
h
FIG. 3. Undermethylation of the 5' Msp I/Hpa II site andenhancer function in T-cell lines. (A) T-cell DNA was analyzed formethylation as in Fig. 2. Lanes: a, BW5147 cells; b, EL-4; c, YAC-1;d, WEHI 7.1; e, WEHI 22.1. (B) T-cell DNA was analyzed forrearrangement at the JH-C,. locus by digestion with EcoRI andhybridization with probe 1 (Fig. 1). Lane designations are as in A. (C)Cytoplasmic RNA from T-cell lines was analyzed for , chain-specifictranscripts with probe 4 (Fig. 1). The first dot in each row containsRNA isolated from 1.5 x 106 cells. Lanes: a, WEHI 22.1 cells; b,YAC-1; c, WEHI 7.1; d, BW5147; e, EL-4; f, K46R; g, 70Z; h,MPC-11, an IgG-secreting control.
genes did not correlate with rearrangement at that locus.Mapping of the EcoRI sites in the JH region showed that boththe EL-4 and WEHI 22.1 cells retained the germ-line organi-zation; in each case the digestion yielded a single 6.4-kbfragment diagnostic of unrearranged JH gene segments (Fig.3B). Conversely, digestion of YAC-1 DNA, which washeavily methylated at the Msp I/Hpa II site 5' to theenhancer, yielded an additional fragment of 5.5 kb, indicatingthat one of the alleles had undergone a D-J rearrangement.However, undermethylation in the EL-4 and WEHI 22.1DNA did correlate with the production of sterile ,u-chaintranscripts (Fig. 3C). When cytoplasmic RNA from the T-celllines was examined for A chain-specific sequences, only theEL-4 and WEHI 22.1 preparations gave positive reactions.Thus, the analyses of T-cell lines indicated that the loss ofmethyl groups in the JH4-enhancer region is associated withenhancer activation rather than with V-D-J rearrangement.
Methylation of the ,u-Chain Gene from the Switch Region tothe C,,2 Exon. The midgene Msp I/Hpa II sites located in theswitch region (blot not shown), 5' to the C,.,1 exon, and withinC,.1 and C.2 exons exhibited a similar pattern of under-methylation (Figs. 4 and 5). In each case, the sites werepartially undermethylated in the ,u chain-negative liver andembryo tissues; the undermethylation increased in the ,uchain-positive lymphoma cells and became complete in theIgM-secreting myeloma cells. This pattern is best illustrated(Fig. 4) by the blot obtained when the methylation status ofthe 5' flanking and Cj1 exon sites was monitored by hybridi-zation with the intervening 0.9-kb Msp I fragment (probe 3,Fig. 1). Densitometer scans of the amounts of diagnosticfragment generated showed that, in liver and embryo DNA,
EcoRI + Msp Ia b c d e f g
Eco RI + Hp Tr
a b c d e f 9
-12.5-
-6.7-
-1.55-
:~~~~~~~~~~~~~~~~~~j.. ...
*a...ai*.~~ -0.95.-.- - '*'4P4
*~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~;i:m0:.. :'
ON 0*1#O.95
-- Eco RI 12.5M M MM M
probe,0.951.55
6.75665
12.5
FIG. 4. Methylation of the Msp I/Hpa II sites 5' to and within theCJ exon. DNA was hybridized with probe 3 (Fig. 1). Lanes: a, livercells; b, embryo; c, WEHI 231.2; d, 38C13; e, K46R; f, MXW231.1a.2; g, MOPC 104E.
10-17% of both sites was susceptible to Hpa II cleavage; inlymphoma DNA, 30-60%; and in myeloma DNA, 100%(Table 1). Thus, the midgene C-C-G-G sequences exhibited aprogressive loss of methyl groups that did not correlate withspecific events in B-lymphocyte differentiation.
Methylation in the Region of the pu Exon. A developmen-tally related methylation pattern was seen when the MspI/Hpa II site located in the C,4 exon was analyzed (Fig. 5).Hybridization with the adjacent 1.1-kb Msp I fragment (probe4 in Fig. 1) showed that the C,4 exon site was completelymethylated at all differentiative stages preceding IgM secre-tion. In the DNA from IgM-secreting lines, however, therewas a dramatic loss of methyl groups; 56-92% of the siteswere undermethylated asjudged by the hybridization densityof the diagnostic fragments.Whether this change included the As exon and its 3'
flanking sequences could not be determined by Msp I/Hpa IImapping because there are no C-C-G-G sequences in theintron between the A, and lm exons. To resolve the issue, theanalyses were extended to a second methyl-sensitive en-zyme, Hha I, which recognizes the sequence 5' G-C-G-C 3'.Two such sequences are present in the 3' region of the,u-chain gene, one just following the Au exon and another 0.5kb further downstream (Fig. 1). When the methylation statusof these two sites was examined by hybridization with probe5 (data not shown), the DNA from liver, embryo, andlymphoma lines was highly resistant to Hha I cleavage at bothsites, whereas the DNA from IgM-secreting lines was com-pletely susceptible to digestion at the sitejust 3' to the As exonand partially susceptible at the downstream site. Thus, thecombined data from Msp I/Hpa II and Hpa I mapping definea region of -1 kb encompassing the p.s exon that undergoesa specific change in methylation during the transition to IgMsecretion.
A EcoRI + HpaI[a b c d e
Immunology: Blackman and Koshland

3812 Immunology: Blackman and Koshland
EcoRI + Msp I EcoRi + HpaI[a b c d e f g a b c d e f g
i~~~ . .gi. .... E,
-125gi-125 h 0~ ~ . ....A 2- ;.j... ..
ok- 121 - ...: ...' ''..'
MM MM M
.7 O'LI
probe
---
5.25.7
FIG. 5. Methylation of Msp I/Hpa II sites in the C,,2 and C,,4exons. DNA was hybridized with probe 4 (Fig. 1). Cell lines are thesame as in Fig. 4.
DISCUSSION
By following the methylation of C,. and its 5' flankingsequence as a function of B-cell differentiation, two patternsof change could be identified: (i) a stage-specific loss ofmethyl groups at sites in the 5' and 3' sequences, and (ii) a
progressive undermethylation of midgene sites over the lifehistory of the B cell. The finding of two distinct patternsexplains the contradictory results obtained in previous analy-ses of limited regions of the A-chain gene (26, 27). Moreover,the finding of stage-specific changes in methylation providesnew insight into the mechanisms that regulate p-chain gene
expression.The onset of I-chain gene transcription was found to
correlate with the loss of methyl groups from a single MspI/Hpa II site located in the 5' flanking sequence 150 basepairs downstream from the JH4 segment and 750 base pairsupstream from the enhancer region. This CpG sequence isheavily methylated in nonlymphoid and embryo DNA andbecomes completely undermethylated in B-cell lines express-
ing A chain, indicating that chromatin changes in the vicinityof the Msp I/Hpa II site are involved in the induction ofA-chain gene expression. The role of these changes wasdefined by extending the analyses to T-cell lines in which itis possible to distinguish the two requirements for transcrip-tion: enhancer activation and V-D-J recombination. Theanalyses showed that the chromatin changes in the region ofthe 5' Msp I/Hpa II site are directly related to enhancerfunction. The site was found to be detectably undermethyl-ated in two T-cell lines that retain the germ-line u-chain gene
structure. The same two lines were the only ones to express
sterile p-chain transcripts, and the level of transcription wasroughly proportional to the extent of undermethylation.
This evidence is supported by the stage-specific changesthat have been observed in heavy chain gene enhancer
structure. Mills et al. (28) have shown that transcription ofhuman p-chain genes correlates with the development ofDNase I-hypersensitive sites. The major hypersensitive sitemaps to the-enhancer region and could mark the interactionof a lymphocyte-specific regulatory factor. Moreover,DNase I hypersensitivity is generated in the enhancer regionof both g-chain gene alleles, just as the changes in methyla-tion occur on both alleles. Thus, the pattern of stage-specificalterations in the JH4-enhancer region suggests that enhanceractivation propagates chromatin changes through the 5'sequences and initiates A-chain gene transcription by influ-encing the conformation of upstream promoter elements (29).The methylation data do not resolve the question as to
whether chromatin changes in the JH4-enhancer region playa role in D-J or V-D recombination. The analyses of T-celllines suggest this is not the case; in one cell line, D-J joiningwas observed without concomitant undermethylation of the5' Msp I/Hpa II site or expression of A-chain transcripts.However, the requirements may differ in B cells. Reth andAlt have reported (30) that D-J rearrangement in immature Bcells results in the transcription of D-J-C,. RNA frompromoter sequences located 5' to the D gene segments.Presumably, such transcription requires activation of thedownstream enhancer, if not as a precondition, at least as aclosely synchronized event. The expression of D-J-CL RNAalso implies that chromatin changes in the JH4-enhancerregion precede V-D joining and thus may serve a key role inthat process.A second stage-specific change in C,. gene methylation
occurs in the 3' sequences and correlates with the transitionof a resting B lymphocyte to an IgM-secreting blast cell.Restriction analyses of lymphoid cell lines showed that aMsp I/Hpa II site in C. exon 4 and a Hha I site just 3' to the,us exon remain heavily methylated through all of the devel-opmental stages preceding IgM secretion and then becomelargely undermethylated in IgM-secreting lines. A Hha I sitefurther downstream from the As exon exhibits a similar butless dramatic loss of methyl groups. These data indicate thata region of approximately 1 kb surrounding that exon under-goes a developmentally regulated change in chromatin struc-ture. A clue as to how this change may be initiated comesfrom the DNA sequence directly 3' to the ps exon. Richardset al. (24) have identified a region of potential Z-DNAstructures that, by flipping to B-DNA, could generate up-stream changes in chromatin conformation.The differentiation of a B cell to an IgM-secreting cell is
characterized by two changes in A-chain expression: a shiftin p-chain RNA synthesis from the gm to the Au form and anamplification in the levels of A chain-specific message (31,32). It is unlikely that chromatin changes in the region of the.As exon play a direct part in regulating the shift from g.m tops chains. The available evidence indicates that the ratio ofAs to Am chains is controlled by differential cleavage andpolyadenylylation of the primary RNA transcript and not byevents at the DNA level, such as early termination oftranscription (1, 31, 33-35). Moreover, significant amounts ofA.-chain RNA are produced by lymphoma cell lines in whichthe 3' Msp I/Hpa II and Hha I sites are completely methyl-ated; the secreted form constitutes 40-50% of the Au chain-specific RNA in these lines compared to 70% to 90% inIgM-secreting plasmacytomas (31, 32).On the other hand, it is likely that chromatin changes in the
region surrounding the A1s exon contribute to the markedamplification in p-chain message that accompanies the pen-tamer IgM response. Analyses of lymphoid lines have shownthat IgM-secreting plasmacytomas contain at least 10 timesmore functional g-chain mRNA than the lymphoma counter-parts of resting B cells (35, 36). This increase can beaccounted for by the increases that have been observed intranscriptional activity. In run-on assays of isolated nuclei,
Proc. Natl. Acad. Sci. USA 82 (1985)
_JI

Proc. Natl. Acad. Sci. USA 82 (1985) 3813
the frequency of transcription initiation was found to be anorder of magnitude higher in IgM-secreting lines than in Blymphomas (34, 35, 37). Thus, the evidence that a transcrip-tional mechanism governs the levels of g-chain messagesuggests that chromatin changes in the 3' sequences of thegene may be one ofthe controlling factors. Such a mechanismmight also contribute to the concurrent A,- to A,-chain shift.It is possible that a marked increase in transcription ratealters the folding of the RNA transcript and thereby makesthe cleavage and polyadenylylation sites 3' to the p, exonmore accessible to reaction (38).
In these studies the methylation of the C,. gene was usedas an assay system to define differentiation-related changesin chromatin structure. Because the analyses were restrictedto lymphoid cell lines arrested at particular differentiativestages, the data obtained do not provide any information onthe kinetics ofDNA hypomethylation and thus on its functionin the regulation of p-chain gene expression. However, thefinding that local changes in methylation correlate withspecific events at the heavy chain locus suggests experimen-tal approaches to answer this question. By following themodulation of specific CpG sequences in Abelson-trans-formed pre-B-cell lines that undergo gene recombination, orin normal B lymphocytes stimulated with mitogen, it shouldbe possible to evaluate whether hypomethylation serves toactivate the differentiative event, to stabilize chromatinchanges once they have been activated, or simply to transfermemory of the change in differentiated state to the progenycells.
The authors thank Dr. M. Tigges for his restriction maps andadvice on subcloning, Dr. E. Mather for helpful discussion, Drs. A.Bothwell and F. Alt for the C,, genomic clone, Dr. H. Sakano for theJH genomic clone. This research was supported by Grant A1-07079from the National Institutes of Allergy and Infectious Diseases andby Training Grant CA-09179 from the National Cancer Institute.
1. Wall, R. & Kuehl, M. (1983) Annu. Rev. Immunol. 1, 393-422.2. Bentley, D. L., Farrell, P. J. & Rabbitts, T. H. (1982) Nucleic
Acids Res. 10, 1841-1856.3. Banerji, J., Olson, L. & Schaffner, W. (1983) Cell 33, 729-740.4. Gillies, S. D., Morrison, S. L., Oi, V. T. & Tonegawa, S.
(1983) Cell 33, 717-728.5. Mather, E. L. & Perry, R. P. (1981) Nucleic Acids Res. 9,
6855-6866.6. Alt, F. W., Rosenberg, N., Enea, V., Siden, E. & Baltimore,
D. (1982) Mol. Cell. Biol. 2, 386-400.7. Kemp, D. J., Harris, A. W., Cory, S. & Adams, J. M. (1980)
Proc. NatI. Acad. Sci. USA 77, 2876-2880.8. Kehry, M., Ewald, S., Douglas, R., Sibley, C., Raschke, W.,
Fambrough, D. & Hood, L. (1980) Cell 21, 393406.
9. Early, P., Rogers, J., Davis, M., Calame, K., Bond, M., Wall,R. & Hood, L. (1980) Cell 20, 313-319.
10. Alt, F. W., Bothwell, A. L. M., Knapp, M., Siden, E., Mather,E., Koshland, M. & Baltimore, D. (1980) Cell 20, 293-301.
11. Rogers, J., Early, P., Carter, C., Calame, K., Bond, M.,Hood, L. & Wall, R. (1980) Cell 20, 303-312.
12. Behe, M. & Felsenfeld, G. (1981) Proc. Natl. Acad. Sci. USA78, 1619-1623.
13. Dawid, I. B., Brown, D. D. & Reeder, R. H. (1970) J. Mol.Biol. 51, 341-360.
14. Razin, A. & Riggs, A. D. (1980) Science 210, 604-610.15. Doerfler, W. (1983) Annu. Rev. Biochem. 52, 93-124.16. Bird, A. P. (1984) Nature (London) 307, 503-504.17. Groudine, M., Eisenman, R. & Weintraub, H. (1981) Nature
(London) 292, 311-317.18. McGhee, J. D., Wood, W. I., Dolan, M., Engel, J. D. &
Felsenfeld, G. (1981) Cell 27, 45-55.19. Mather, E. L. & Perry, R. P. (1983) Proc. Natl. Acad. Sci.
USA 80, 4689-4693.20. Yagi, M. & Koshland, M. E. (1981) Proc. Natl. Acad. Sci.
USA 78, 4907-4911.21. White, B. A. & Bancroft, F. C. (1982) J. Biol. Chem. 257,
8569-8572.22. Kawakami, T., Takahashi, N. & Honjo, T. (1980) Nucleic
Acids Res. 8, 3933-3945.23. Sakano, H., Maki, R., Kurosawa, Y., Roeder, W. &
Tonegawa, S. (1980) Nature (London) 286, 676-683.24. Richards, J. E., Gilliam, A. C., Shen, A., Tucker, P. W. &
Blattner, R. F. (1983) Nature (London) 306, 483-487.25. Maki, R. A., Traunecker, A., Sakano, H., Roeder, W. &
Tonegawa, S. (1980) Proc. Natl. Acad. Sci. USA 77,2138-2142.
26. Rogers, J. & Wall, R. (1981) Proc. Natl. Acad. Sci. USA 78,7497-7501.
27. Storb, U. & Arp, B. (1983) Proc. Natl. Acad. Sci. USA 80,6642-6646.
28. Mills, V. C., Fisher, L. M., Kuroda, R., Ford, A. M. & Gould,H. J. (1983) Nature (London) 306, 809-812.
29. Perry, R. P. (1984) Nature (London) 310, 14-15.30. Reth, M. G. & Alt, F. W. (1984) Nature (London) 312,
418-423.31. Nelson, K. J., Haimovich, J, & Perry, R. P. (1983) Mol. Cell.
Biol. 3, 1317-1332.32. Lamson, G. & Koshland, M. E. (1984) J. Exp. Med. 160,
877-892.33. Kemp, D. J., Morahan, G., Cowman, A. F. & Harris, A. W.
(1983) Nature (London) 301, 84-86.34. Yuan, D. & Tucker, P. W. (1984) J. Exp. Med. 160, 564-583.35. Tigges, M. & Koshland, M. E. (1984) Fed. Proc. Fed. Am.
Soc. Exp. Biol. 43, 1426 (abstr.).36. Perry, R. P., Kelley, D. E., Coleclough, C. & Kearney, J. F.
(1981) Proc. Nati. Acad. Sci. USA 78, 247-251.37. Mather, E. L., Nelson, K. J., Haimovich, J. & Perry, R. P.
(1984) Cell 36, 329-338.38. Koshland, M. E. (1983) J. Immunol. 131, i-ix.
Immunology: Blackman and Koshland


















