
Reginald H. Garrett Charles M. Grisham Chapter 30 Protein Synthesis.
Upload
osborn-davidsonCategory
view
252download
4
Reginald H. GarrettCharles M. Grisham
Chapter 3Thermodynamics of Biological
Systems

Outline
• What are the basic concepts of thermodynamics ?• What can thermodynamic parameters tell us about
biochemical events ?• What is the effect of concentration on net free
energy changes ?• What is the effect of pH on standard-state free
energies ?• Why are coupled reactions important to living
things ?• What are the characteristics of high-energy
biomolecules ? • What are the complex equilibria involved in ATP
hydrolysis ?

3.1 What Are the Basic Concepts of Thermodynamics?
• Definitions for thermodynamics:• The system: the portion of the universe with
which we are concerned.• The surroundings: everything else.• Isolated system cannot exchange matter or
energy.• Closed system can exchange energy but not
matter.• Open system can exchange either or both.

3.1 What Are the Basic Concepts of Thermodynamics?
Figure 3.1 The characteristics of isolated, closed, and open systems. Isolated systems exchange neither matter nor energy with their surroundings. Closed systems may exchange energy, but not matter, with their surroundings. Open systems may exchange either matter or energy with the surroundings.

Laws of Thermodynamics
First Law:• Conservation of energy – Energy can neither be
created nor destroyed.
Second Law:• All processes proceed toward equilibrium.
Third Law:• The entropy of a pure crystalline solid at 0oK is 0.

State functions
• Free energy, ΔG, is an indication of the chemical energy available from a reaction.
• Enthalpy, ΔH, is the heat of a reaction (bond energy). • Entropy, ΔS = S is a measure of disorder.
an ordered state = low entropya disordered state = high entropy
G = H - TS
• If G = 0, the reaction is at equilibrium.• If G < 0 (is negative), the reaction proceeds as written.• If G > 0 (is positive), the reaction proceeds in the reverse
of the direction as written.

Standard Conditions
• G of reaction depends on conditions.• Standard State (Go) has set reference
conditions:• Temperature = 298 oK• Pressure = 1 atmosphere• Concentration = 1 Molar
• Biological Standard State (Go') uses includes a standard H+ concentration of 10-7 M (pH 7)rather than 1 M.

The Effect of Concentration on ΔG
• How can we calculate the free energy change for reactions not at standard state ?
• Consider a reaction: A + B C + D
• Then: [C] [D]ΔG = ΔG° + RT ln --------
[A] [B]
• Thus concentrations at other than 1 M will change the value of G.
• This is Equation 3.13. It is used frequently throughout this text.

Evaluation of Standard State Free Energy
• Remember, at equilibrium ΔG = 0.• Substituting ΔG = 0 into the previous equation gives
the relationship between standard state free energy, ΔG°, and the equilibrium constant, Keq.
ΔG° = - RT ln Keq
• Since in biological systems the predominant pH is 7, it makes more sense to adopt a modified standard state – i.e., 1 M for all constituents except protons, for which the standard state is pH 7.
ΔG°' = - RT ln Keq'

3.3 What is the Effect of pH on Standard State Free Energies?
• For those reactions in which H+ is not a reactant or a product, the values of ΔG°´and ΔG° are about equal.
• Where a proton is involved, ΔG°´and ΔG° are as shown below:
• For reactions in which H+ is produced:
ΔG°´= ΔG° + RT ln [H+]
• And for reactions in which H+ is consumed:
ΔG°´ = ΔG° - RT ln [H+]

Oxidation- Reduction
• In redox reactions, the free energy is related to electron flow.
ΔG° = - nE°F
• Where n is the number of electrons.• Eo is the cell potential at standard state.• F is Faraday’s constant, 96480 J/mol-V
• This can also be evaluated at pH 7:
ΔG°' = -nE°'F

Example Calculation
Sample calculation of Go' and G.
3-phosphoglycerate <===> 2-phosphoglycerate
Given Keq: 2-PGKeq = ------- = 0.178 at 37oC
3-PG
Calculate Go'
Go' = -2.3 RT log 0.178
= -2.3(8.314)(310)(-0.75)
= + 4446 J/mol or 4.446 kJ/mol

Example Calculation
Calculate the overall free energy for this reaction, G, using physiological conditions in a red cell (erythrocyte) at 37oC:
3-PG = 62.1 mol/liter = 62.1 x 10-6 M
2-PG = 4.3 mol/liter = 4.3 x 10-6 M
4.3 x 10 -6G = Go' + 2.3 RT log ---------------
62.1 x 10-6

Example Calculation
G = 4446 + 5928 log 0.0692
= 4446 + 5928 (-1.16)
= 4446 - 6876
= - 2340 J/mol
The negative value of the overall G for this reaction indicates that in the red cell it is spontaneous toward 2-PG.

3.5 What are the Characteristics of High-Energy Biomolecules?
Energy Transfer - A Crucial Biological Need
• Energy acquired from sunlight or food must be used to drive endergonic (energy-requiring) processes in the organism.
• Basically, two classes of biomolecules do this:
• Reduced coenzymes (NADH, FADH2).
• High-energy phosphate compounds – those having a free energy of hydrolysis more negative than -25 kJ/mol.
• Note that thioesters are also high energy.

High-Energy Biomolecules
Table 3.3 is important
• Note what's really high:PEP and 1,3-BPG • Note what's low: sugar-P, etc. • Note what's in between: ATP • Note difference (Figure 3.6) between Go' -
noted in Table 3.3 - and the activation energy for phosphoryl-group transfer.
**Note: Some of the figure references in Table 3.3 in the text refer to the 3rd Edition of the text.

3.5 What are the Characteristics of High-Energy Biomolecules?
Structure
Figure 3.12
Figure 3.10
Figure 13.21
Figure 3.10
Figure 3.9
Figure 3.9
Figure 3.9
Figure 23.6
Figure 23.6
Figure 22.14
Page 570
Figure 25.28
Figure 15.12, 22.14
Page 558
Figure 26.8

3.5 What Are the Characteristics of High-Energy Biomolecules?
Figure 3.6 The activation energies for phosphoryl group transfer reactions are substantially larger than the free energy of hydrolysis of ATP.

Group Transfer Potentials Quantify the Reactivity of Functional Groups
Group transfer is analogous to ionization potential and reduction potential. All these are specific instances of free energy changes.

ATP
An Shuttle Device of Intermediate Energy
• PEP and 1,3-BPG are created in the course of glucose breakdown.
• Their energy (and phosphates) are transferred to ADP to form ATP.
• But ATP is only a transient energy carrier - it quickly passes its energy to a host of energy-requiring processes.
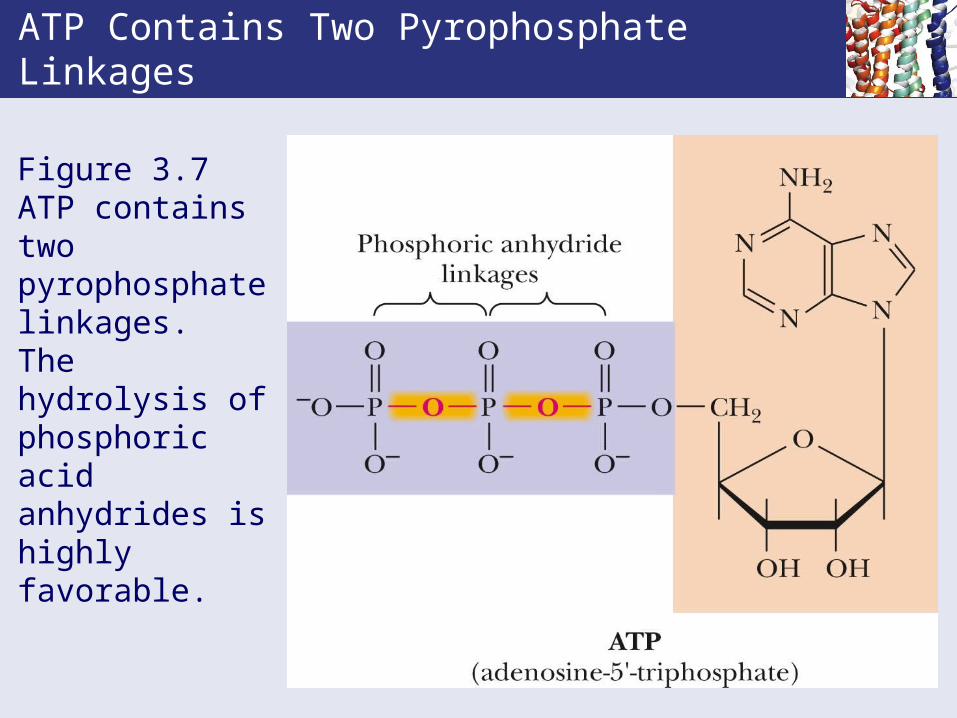
ATP Contains Two Pyrophosphate Linkages
Figure 3.7 ATP contains two pyrophosphate linkages. The hydrolysis of phosphoric acid anhydrides is highly favorable.

Free Energy of Hydrolysis
Comparison of ΔGo' Values
• ATP ADP + Pi -30.5 kJ/mol
• ATP AMP + PPi -32.3 kJ/mol
• PPi 2 Pi -33.6 kJ/mol
• AMP Adenosine + Pi -9.2 kJ/mol
• Why is there a large difference in the last value ?

Phosphoric Acid Anhydrides
Accounting for the high Go' of ATP hydrolysis
• ADP and ATP are examples of phosphoric acid anhydrides (similar to acyl anhydrides).
• Factors in the large negative Go': • electrostatic repulsion. • stabilization of products by resonance. • stabilization of products by ionization (and
interaction with aqueous media). • entropy factors.
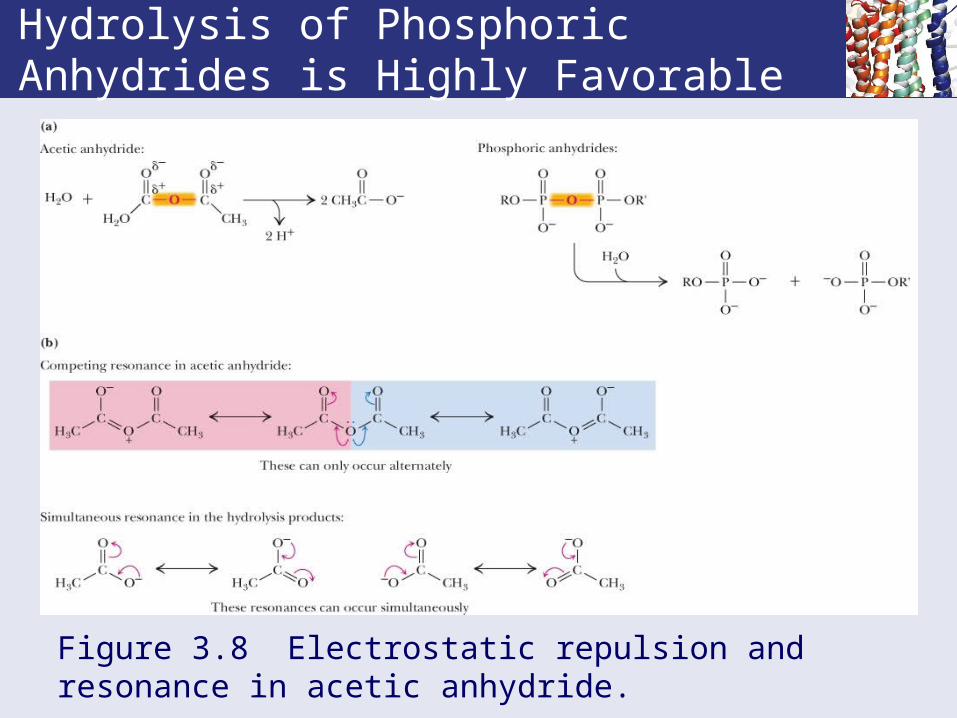
Hydrolysis of Phosphoric Anhydrides is Highly Favorable
Figure 3.8 Electrostatic repulsion and resonance in acetic anhydride.

3.5 What Are the Characteristics of High-Energy Biomolecules?
Figure 3.9 Hydrolysis of ATP to ADP (and/or of ADP to AMP) leads to relief of electrostatic repulsion.

Phosphoric-Carboxylic Anhydrides
• These mixed anhydrides - also called acyl phosphates - are very energy-rich.
• Acetyl-phosphate: G°´ = -43.3 kJ/mol.• 1,3-BPG: G°´ = -49.6 kJ/mol.• Bond strain, electrostatics, and resonance
are responsible.
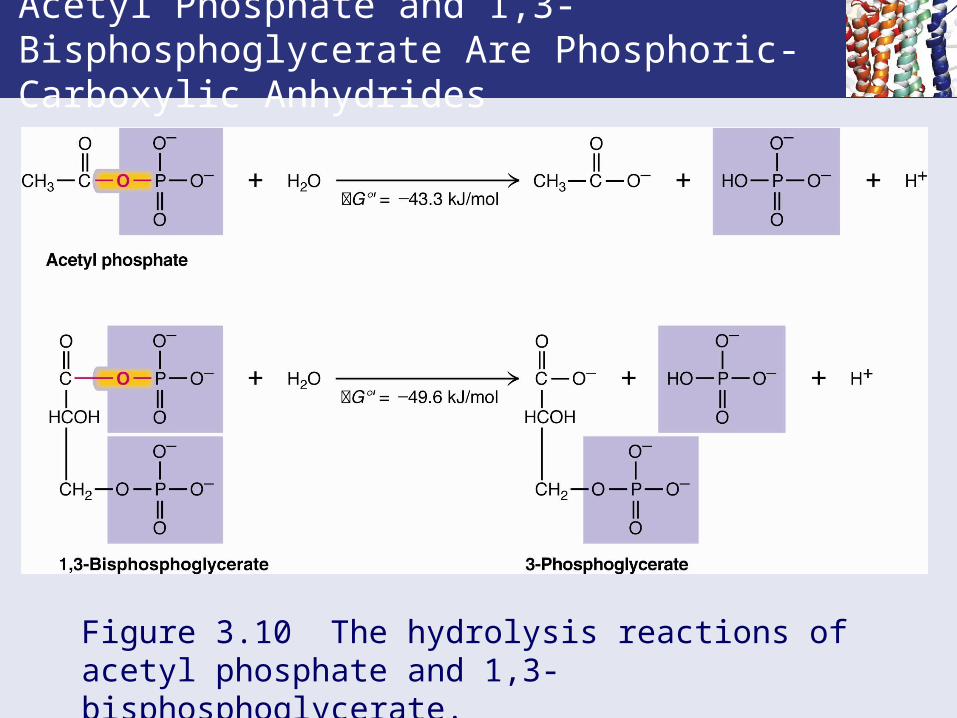
Acetyl Phosphate and 1,3-Bisphosphoglycerate Are Phosphoric-Carboxylic Anhydrides
Figure 3.10 The hydrolysis reactions of acetyl phosphate and 1,3-bisphosphoglycerate.

Enol Phosphates
• Phosphoenolpyruvate (PEP) has the largest free energy of hydrolysis of any biomolecule.
• Formed by dehydration of 2-phospho-glycerate.
• Hydrolysis of PEP yields the enol form of pyruvate - and tautomerization to the keto form is very favorable.

PEP Hydrolysis Yields -62.2 kJ/mol
Figure 3.11 PEP is produced by the enolase reaction and in turn drives the phosphorylation of ADP to form ATP in the pyruvate kinase reaction.

Enol Phosphates are Potent Phosphorylating Agents
Figure 3.12 Hydrolysis and subsequent tautomerization account for the very large ΔG° of PEP.

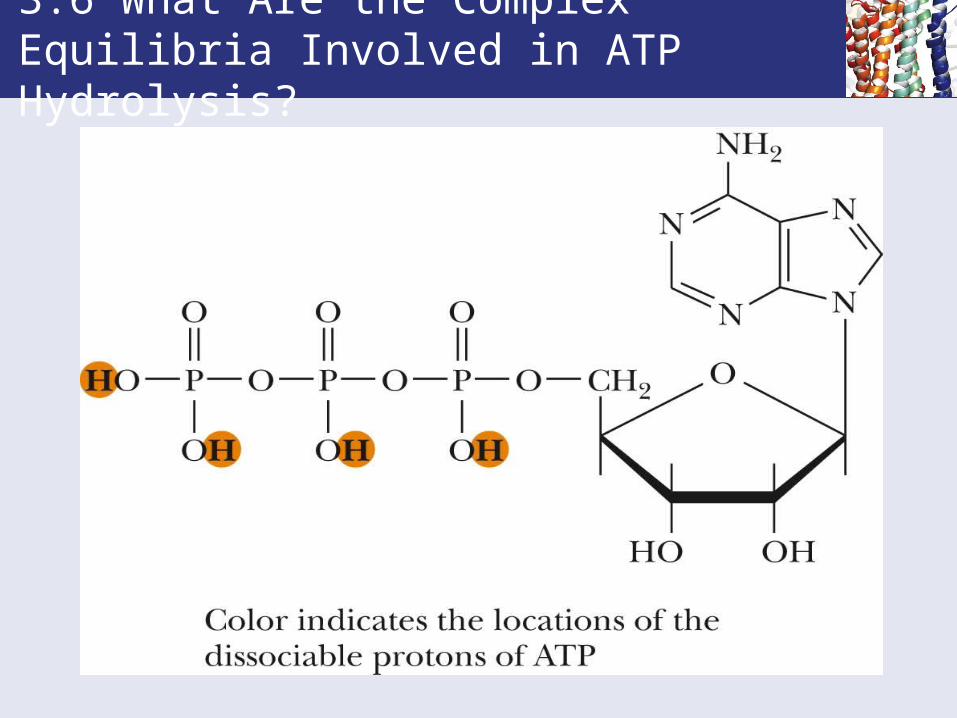
Ionization States of ATP
• ATP has four dissociable protons.
• pKa values range from 0-1 to 6.95.
• Free energy of hydrolysis of ATP is relatively constant from pH 1 to 6, but rises steeply at high pH.
• Since most biological reactions occur near pH 7, this variation is usually of little consequence.
3.6 What Are the Complex Equilibria Involved in ATP Hydrolysis?

The Free Energy of Hydrolysis for ATP is pH-Dependent
Figure 3.14 The pH dependence of the free energy of hydrolysis of ATP. Because pH varies only slightly in biological environments, the effect on ΔG is usually small.

Metal Ions Affect the Free Energy of Hydrolysis of ATP
Figure 3.15 The free energy of hydrolysis of ATP as a function of total Mg2+ ion concentration at 38°C and pH 7.0.
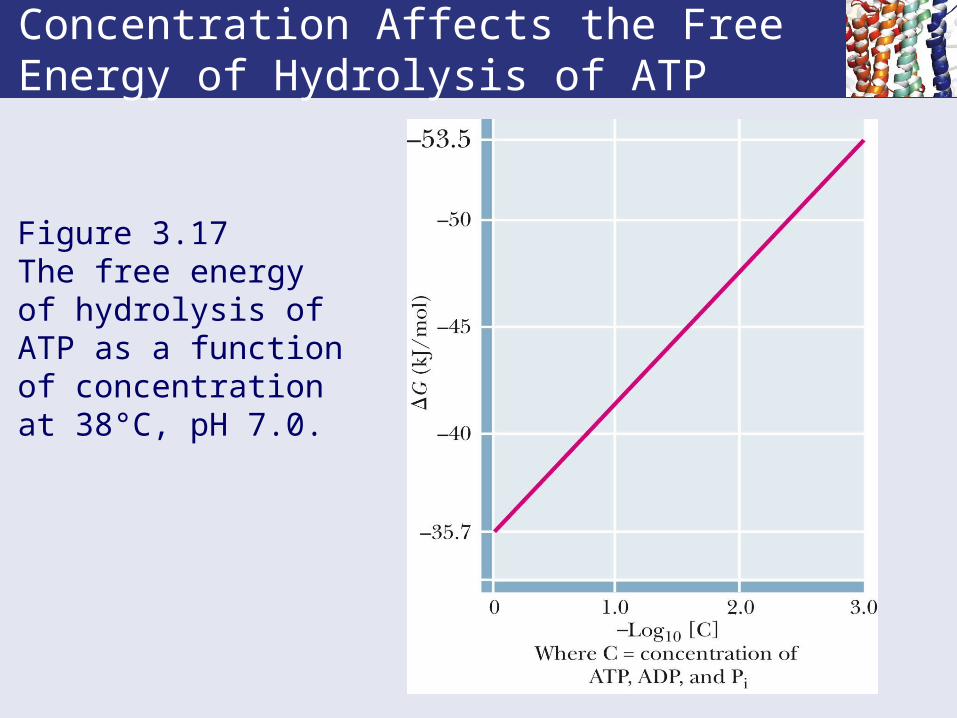
Concentration Affects the Free Energy of Hydrolysis of ATP
Figure 3.17 The free energy of hydrolysis of ATP as a function of concentration at 38°C, pH 7.0.

The Effect of Concentration
Recall that free energy changes are concentration dependent
So the free energy available from ATP hydrolysis depends on concentration
• We will use the value of -30.5 kJ/mol for the Go' (standard-state free energy) of hydrolysis of ATP.
• At non-standard-state conditions (in a cell, for example), the G is different.
• Equation 3.12 is crucial - be sure you can use it properly.
• In typical cells, the G for ATP hydrolysis is typically -50 kJ/mol.

3.7 Coupled MetabolicReactions/Processes
• Many reactions of cells and organisms run against their thermodynamic potential – that is, in the direction of positive ΔG.
• Examples – synthesis of ATP, creation of ion gradients.
• These processes are driven in the thermodynamically unfavorable direction via coupling with highly favorable processes.

Coupled Metabolic Reactions/Processes
Figure 3.18 The pyruvate kinase reaction. Hydrolysis of PEP is very favorable, and it is used to drive phosphorylation of ADP to form ATP, a process that is energetically unfavorable.

Successive Metabolic Reactions
• The Go' values for successive reactions are additive.
• Consider what this means in terms of equilibrium concentrations of reactants and products.
• Example: 2-P-glycerate pyruvate.2-P-glycerate PEP +1.8 kJ/molPEP Pyruvate -31.7 kJ/mol
Net: 2-P-glycerate pyruvate -29.9 kJ/mol• Here the first reaction (unfavorable Go') is
pulled forward by the highly exergonic second reaction.

3.8 What is the Daily Human Requirement for ATP?• The average adult human consumes approximately
11,700 kJ of food energy per day.• Assuming thermodynamic efficiency of 50%, about
5860 kJ of this energy ends up in form of ATP.• Assuming 50 kJ of energy required to synthesize
one mole of ATP, the body must cycle through 5860/50 or 117 moles of ATP per day.
• This is equivalent to 65 kg of ATP per day.• The typical adult human body contains 50 g of
ATP/ADP.• Thus each ATP molecule must be recycled nearly
1300 times per day.

ATP Changes Keq by 108
• Consider a process: A → B
• Compare this to: A + ATP → B + ADP + Pi
• Assume typical cellular concentrations of ATP, ADP and Pi, and using the standard state free energy of ATP hydrolysis, it can be shown that coupling of ATP hydrolysis to the reaction of A to B changes the equilibrium ratio of B/A by more than 200 million-fold.

3.4 What Can Thermodynamic Parameters Tell Us About Biochemical Events?
Figure 3.5 Unfolding of a soluble protein exposes significant numbers of nonpolar groups to water, forcing order on the solvent and resulting in a negative entropy change.

End Chapter 3Thermodynamics of Biological
Systems


















