
Reaction of 7S and 19S Components of Immune Rabbit Antisera ...
13
Immunology, 1963, 6, 421. Reaction of 7S and 19S Components of Immune Rabbit Antisera with Human Group A and AB Red Cells C. L. GREENBURY, D. H. MOORE AND L. A. C. NUNN Department of Pathology, Stoke Mandeville Hospital, Aylesbury, Bucks. (Received 31st December 1962) Summary. 7S and 19S components of rabbit anti-A antibody, labelled with 1311 have been separated by chromatography on DEAE-cellulose. The reactions of these components with human group A1, A2 and AB red cells have been in- vestigated. At saturation point A1 cells combine with 0-22 p-g. of 7S antibody per million cells, this corresponds to a minimum of 8-3 x 105 A-antigen sites per cell. A2 cells can take up only one quarter as much antibody, and combine much less avidly with it than do A1 cells. Cells can take up only about one-fifth as many 19S as 7S antibody molecules. 19S antibody is more avid and, on a molecular basis, is 750 times more efficient at agglutinating red cells than 7S antibody. The sig- nificance of these findings is discussed with regard to the differences between A1 and A2 antigens, and the valency and mode of attachment to the red cells of the antibodies. INTRODUCTION For the purpose of studying the reaction of Rheumatoid Factor with antibody-coated red cells, we have been using as substrate human group A red cells and rabbit anti-A serum. As a basis for further progress it has been necessary to investigate in some detail the A cell/anti-A system. This study has yielded information of some general interest, and the findings are presented here. We have immunized rabbits with human group A red cells. The globulin from the immune rabbits has been labelled with 131I, and the 7S and 19S fractions have been separated by chromatography on DEAE-cellulose. By measuring the uptake by A1 and A2 cells of these antibodies in varying experimental conditions we have attempted to measure the number of A-antigen sites on the red cells and have compared the avidity, combining ratios and agglutinating properties of the 7S and 19S antibodies. MATERIALS AND METHODS Red Cells Except where otherwise stated, all the work was done with red cells from two donors, belonging respectively to groups A1 and A2. Blood was taken into A.C.D. and stored at 40 until used; the period of storage was usually less than 1 week and never more than 14 days. Before use, cells were washed three times in isotonic saline. A Coulter counter was used for counting red cells. 421
Transcript of Reaction of 7S and 19S Components of Immune Rabbit Antisera ...

Immunology, 1963, 6, 421.
Reaction of 7S and 19S Components of Immune RabbitAntisera with Human Group A and AB Red Cells
C. L. GREENBURY, D. H. MOORE AND L. A. C. NUNN
Department of Pathology, Stoke Mandeville Hospital, Aylesbury, Bucks.
(Received 31st December 1962)
Summary. 7S and 19S components of rabbit anti-A antibody, labelled with 1311have been separated by chromatography on DEAE-cellulose. The reactions ofthese components with human group A1, A2 and AB red cells have been in-vestigated. At saturation point A1 cells combine with 0-22 p-g. of 7S antibodyper million cells, this corresponds to a minimum of 8-3 x 105 A-antigen sites percell. A2 cells can take up only one quarter as much antibody, and combine much lessavidly with it than do A1 cells. Cells can take up only about one-fifth as many 19Sas 7S antibody molecules. 19S antibody is more avid and, on a molecular basis,is 750 times more efficient at agglutinating red cells than 7S antibody. The sig-nificance of these findings is discussed with regard to the differences between A1and A2 antigens, and the valency and mode of attachment to the red cells of theantibodies.
INTRODUCTION
For the purpose of studying the reaction of Rheumatoid Factor with antibody-coatedred cells, we have been using as substrate human group A red cells and rabbit anti-Aserum. As a basis for further progress it has been necessary to investigate in some detailthe A cell/anti-A system. This study has yielded information of some general interest,and the findings are presented here.We have immunized rabbits with human group A red cells. The globulin from the
immune rabbits has been labelled with 131I, and the 7S and 19S fractions have beenseparated by chromatography on DEAE-cellulose. By measuring the uptake by A1 andA2 cells of these antibodies in varying experimental conditions we have attempted tomeasure the number of A-antigen sites on the red cells and have compared the avidity,combining ratios and agglutinating properties of the 7S and 19S antibodies.
MATERIALS AND METHODSRed Cells
Except where otherwise stated, all the work was done with red cells from two donors,belonging respectively to groups A1 and A2. Blood was taken into A.C.D. and stored at 40until used; the period of storage was usually less than 1 week and never more than 14days. Before use, cells were washed three times in isotonic saline. A Coulter counter wasused for counting red cells.
421

422 C. L. Greenbury, D. H. Moore and L. A. C. N%unnImmunization of Rabbits
Rabbits were immunized with five intravenous injections, each of 5 ml. of a 5 per centsuspension of red cells, given at 5 day intervals. They were bled a week after the last injec-tion and on the following 2 days, 40-50 ml. of blood being taken at each bleeding. Serawere stored at -200 until used.
Preparation of Antibody for IodinationA crude y-globulin fraction was precipitated from the antisera by adding one volume of
serum to two volumes of 27 per cent w/v Na2SO4 and leaving for 3 hours at 370. Theprecipitate was washed in 18 per cent w/v Na2SO4, dissolved in saline and then dialysedagainst saline until sulphate-free. The protein was then iodinated.
Iodination ofy GlobulinThe method of McFarlane (1958) was used (without pre-oxidation of sulphydryl
groups). The y-globulin solution, buffered to pH 9 with glycine buffer, was added rapidlyto a solution of iodine monochloride in equilibrium with 131J* also at pH 9 in quantitiescalculated to introduce one atom of iodine per protein molecule of mol. wt. 160,000.The radioactivity used varied, but did not exceed 70 pc./mg. protein. Most of the non-protein-bound iodine was removed by passing the mixture through a column ofAmberliteIRA-400, any residual free iodine being removed by dialysis against the starting bufferused for DEAE-cellulose chromatography.During the earlier work upon which the present investigation was based, globulin was
iodinated according to J. S. Glover's (personal communication) method. 131It was addedto non-radioactive iodide, the mixture was oxidized to hypoiodite with chloramine-T, andthe protein at pH 9*5 was added rapidly. Free iodine was removed with Amberlite. Thismethod was abandoned because the iodinated globulin gave erratic chromatographicresults.
Separation of 7S and 19S Fractionzs by ChromatographyThe method of Sober, Gutter, Wyckoff and Peterson (1956) was employed, using
DEAE-cellulose (Whatman DE 50 powder) and eluting with five separate sodium phos-phate buffers of increasing molarity and decreasing pH: 0'01 M, pH 8; 0'025 M, pH 7;0'05 M, pH 6; 0'1 M, pH 5'5; 0'15 M, pH 4'6. The iodinated globulin was dialysed over-night at 40 against the starting buffer before application to the column. Of the first fraction20-40 per cent and up to 20 per cent of the last fraction consisted of antibody. Thefractions eluted by the three intermediate buffers contained much less or no antibody.The main protein-containing fractions eluted by the first buffer were pooled and are
referred to as 7S antibody; the main protein-containing fractions eluted by the last bufferwere similarly pooled and are referred to as 19S antibody. The specific radioactivity of the19S antibody was about 1-2 relative to the 7S antibody. Density gradient ultracentrifuga-tion showed that the first fraction from the DEAE-cellulose column contained only 7Sprotein, that is protein whose sedimentation coefficient was similar to that of a known 7Santibody; the last fraction, however, contained both 7S and 19S components of which allthe antibody sedimented with a known 19S protein (cf. Rockey and Kunkel, 1961).
After setting aside from the 7S and 19S fractions a small portion for the determination of* Preparation IBS-3 from Radiochemical Centre, Amersham.t Preparation IBS-1 from Radiochemical Centre, Amersham.

Reaction of Rabbit Antisera with Human Red Cells
protein and specific radioactivity, bovine albumin was added to the remainder in aconcentration fifty or more times that of the rabbit protein. This was done to minimizenon-specific uptake of the labelled rabbit protein, and also to protect the rabbit proteinto some extent from radiation damage.
Since it was necessary for saturation experiments to have a high concentration of anti-body, the antibody-bovine albumin mixtures were concentrated by dialysis againstpolyethylene glycol dissolved in saline to produce a final concentration of rabbit protein of1-2 mg./ml. in the case of 7S antibody, and about 0*6 mg./ml. in the case of 19S antibody.The antibodies were then stored at 40 until used.
Protein DeterminationAll protein determinations were made by the micro-Kjeldahl method, the ammonia
liberated being determined colorimetrically after nesslerization.
Bovine AlbuminOx serum obtained from the slaughterhouse was mixed with anhydrous Na2SO4 to a
final concentration of 18 per cent w/v, to precipitate globulins. The supernatant, afterdialysis against saline, was used as carrier for antibody.
Measurement of RadioactivityAll measurements were made in a well-type scintillation counter.
Haemagglutination TitrationsTwo-fold serial dilutions of the antibody were made in saline in Perspex agglutination
trays, and an equal volume (0.4 ml.) of a 2 per cent red cell suspension was added to eachdilution. After 1-2 hours at room temperature the end-point ('50 per cent agglutination')was estimated by observation of the pattern of the sedimented cells.
Experimental ProcedureSince we have been interested only in anti-A antibody, all antibody preparations were
absorbed with a large excess ofhuman group 0 cells immediately before use. Group 0 cellswere still able to take up a small quantity of protein from the absorbed preparation, butthe amount was very small and, except where stated below, did not form a significantsource of error. In all experiments in which it was relevant to do so, group 0 cells, treatedin the same way as the group A cells, were included as a control.The pattern of absorption experiments was constant, although the concentrations of
antibody and red cells varied according to experimental requirements. Red cells in dilutesuspension of known cell-count were dispensed into 4 x a in. test tubes which were thencentrifuged, and the supernatant fluid was removed. To the cell deposit was added aknown volume, usually 0-2-2 ml. of labelled antibody in bovine albumin and the cellswere resuspended by means of a vortex shaker. After 30-40 minutes at bench temperaturethe tubes were centrifuged and the cells were freed from unbound antibody by washingthree times. A dilute solution of bovine albumin in saline was used as washing fluid, sincethis prevented cells from sticking to the side and bottom of the test tube. After the finalwashing the original tube containing the red cells was counted in the scintillation counter;the cells were then washed out of the tube and the tube alone counted, the differencebetween the two counts being the radioactivity adhering to the cells. That the cells were
423

C. L. Greenbury, D. H. Moore and L. A. C. N&unntightly agglutinated in saturation experiments and over part of the range in avidity experi-ments was found to be an advantage rather than a handicap to accuracy, since agglutina-tion virtually prevented elution of antibody during washing. We found, as did Costea,Schwartz, Constantoulakis and Dameshek (1962), that the error due to free antibodytrapped between the agglutinated cells was negligible.
RESULTS
UPTAKE OF 7S ANTIBODY
If the amount of antibody taken up by red cells at saturation point is known, it ispossible, making certain assumptions, to calculate the number of relevant antigen sites onthe cell. The simplest method of determining the uptake of antibody at saturation point isto add to respective aliquots of red cells increasing concentrations of antibody until theuptake reaches a maximum. The aim ofour saturation experiments has been to determinethis maximum. However, the highest concentration ofantibody attainable has been limitedby the amount of material available and by the necessity of maintaining a fifty-fold orgreater excess of unlabelled (bovine) protein, in order to reduce non-specific take-up.
Table 1 shows the results ofa typical experiment of this nature; other experiments in thisseries have conformed to the same pattern, differing only in the sera used.
TABLE 1
UPTAKE OF 7S ANTIBODY (RABBIT 88)
Antibody uptakeAntibody (pg. per 109 cells)dilution
A1 cells A2 cells 0 cells
Neat 265 62-4 141/2 252 58-4 7.71/4 237 493 5.91/8 219 44 0 2-71/16 195 37-2 0-61/32 170 30 7 0
Antibody: Concentrated 7S fraction from Rabbit 88.Rabbit protein content: 1-38 mg./ml.Absorbable antibody content: 0-482 mg./ml.0 5 ml. volumes of antibody dilutions added to centrifuged celldeposits containing the following number of cells:A1: lI-6x106;A2: 10-7x106;0: 11-6x106.
It will be seen that whereas saturation point has been approached, it has neverthelessnot been achieved. Moreover the curve of uptake versus antibody concentration is difficultto extrapolate. However, it is assumed that the antigen-antibody reaction obeys the lawof mass action; and that at equilibrium the relationships are as follows:
[Ab.Ag] =- K [Ag -Ab.Ag] [Ab -Ab.Ag] (1)where [Ab] and [Ag] are the initial concentrations of antibody and of antigen sites and[Ab.Ag] is the concentration of the antibody-antigen complex at equilibrium. In theregion ofgreat antibody excess, which prevailed in these experiments, the equation may bewritten:
[Ab] K [Ag] -K [Ab.Ag] (2)
424

Reaction of Rabbit Antisera with Human Red Cells
whence, where [Ag] (the concentration of red cells) is kept constant, as in these experi-ments, a plot of [Ab.Ag]/[Ab] versus [Ab.Ag] should give a straight line, and the error ofextrapolation is only that of the deviation from linearity. We have therefore expressed ourresults as curves of fixed antibody ([Ab.Ag]) versus fixed/total antibody ([Ab.Ag]/[Ab]).The intercept on the fixed antibody axis defines the amount of antibody that would betaken up by the red cells at infinite antibody concentration, i.e. at saturation.
0*3
0-25- VI
VI
A 1V
A
A0-15
0-i~~ ~ ~ ~ ~ -
1 2 3 4 5 6 7 X 10-3
FIG. 1. Uptake of 7S antibody by A1 cells. Abscissa: Antibody fixed by red blood cells (pg. per 106 cells). Antibody concentration (pg./ml.). Ordinate: Antibody fixed by red blood cells (pg. per 106 cells).v, Serum 88 preparation 1; o, Serum 88 preparation 2; A, Serum 86; a, Serum 62.
Fig. 1 shows the results obtained with A1 cells using the 7S fraction of sera from threerabbits. The points lie on curves rather than straight lines, and the possible reasons forthis are discussed later. However, it seems not unreasonable to extrapolate the curves asshown. One of the three sera (88) gives a significantly greater uptake than the other two,and this was repeated with a second preparation. The greater uptake from serum 88 mightbe accounted for by a degree of aggregation of the y-globulin molecules (suggestion byDr. R. A. Kekwick). This serum gave a similarly high result with A2 cells.The mean value for uptake at saturation with sera 62 and 86 is 0-22 ,ug. per 106 cells.
If the average molecular weight of 7S y globulin is assumed to be 1 6 x 105, each cell iscapable of absorbing 8-3 x 105 molecules of antibody.
Fig. 2 shows the results using the same antibody preparations as in Fig. 1 with A2 cells.The mean uptake at saturation from sera 62 and 86 is 0-051 ,ug. per 106 cells, a little undera quarter of the uptake by A1 cells. Serum 88 again gives a higher take-up than the othertwo sera, but the ratio of uptake by A1 and A2 cells is similar to that for the other two sera.In experiments with A2 cells, the small amount of protein which in spite of preabsorptionstill remains absorbable by 0 cells forms, with the higher concentrations of antibody, asignificant proportion of the protein taken up by the cells. It is not a constant quantity andhas not been allowed for, but it is likely that the actual amount of antibody which A2cells can absorb specifically is a little less than indicated.Where in Figs. 1 and 2 there are two curves for the same preparation, the curve giving
425

426 C. L. Greenbury, D. H. Moore and L. A. C. Nunn0 08
0*07
0.06 -7
005.
0-04
003 -I _
002
0*01-
0 1 2 3 4 5 6 7 8 9 10 11 12X104FIG. 2. Uptake of 7S antibody by A2 cells. Abscissa: Antibody fixed by red blood cells (pg. per 106 cells)Antibody concentration (pg./ml.). Ordinate: Antibody fixed by red blood cells (pg. per 106 cells).
V, Serum 88; *, Serum 86; o, Serum 62.
the lower uptake was obtained when the preparation was retested a day or two after thefirst experiment. This small decrease in uptake with time has been a constant finding.
INDIVIDUAL VARIATIONS IN THE UPTAKE OF ANTIBODY BY RED CELLS
The experiments illustrated in Figs. 1 and 2 were all done with the cells of two indivi-duals. It was thought desirable to have some idea of how the number of antigen sitesvaried from person to person. Accordingly cells from some seventy patients were examinedby measuring the uptake ofa known volume of cells when exposed to a large excess ofwholeunchromatographed y globulin. The cells from seventeen members of the laboratory staffwere examined in more detail, the uptake of 7S antibody being measured by means ofcurves of the type shown in Fig. 1. These results are illustrated in Fig. 3. The findingsamong the seventy subjects tested suggest that the smaller sample shown in Fig. 3 was afair one, although the scatter is probably rather larger than it shows.
It will be seen that there is quite a wide variation in the number of sites per red cell.A2 cells invariably have fewer antigen sites than A1 cells as measured by their antibodyuptake; A1B and A2B cells also tend to have fewer A-sites than the average for A1 and A2cells respectively. This was also apparent in the larger sample referred to above. It isinteresting to note that cells from the only known A1Al subject whom we were able to test(a child ofA1B parents) did not have very many more sites than the average A1 cells whichare presumably of genotype A10.
SPECIFICITY OF RABBIT ANTISERA FOR A1 AND A2 CELLS
An excess of A1 cells rapidly removed all antibody absorbable by either A1 or A2 cellsfrom the sera 62, 86 and 88, which were raised by immunization with A1 cells. Although,as will be seen below, A2 cells combine less avidly with the antibody, they are neverthelesscapable ofremoving not less than 90 per cent ofthe antibody in the course ofthree successiveabsorptions and it seems probable that there is no antibody which they would ultimately

Reaction of Rabbit Antisera with Human Red Cells
Cell types
AlAl Al A1B A2 A2B
E(a
02-
1.0 CD D w 0) (4 c oCD (
o o 0 ?0 7 - 7- ~(N '(N
6 8 o Oco t
oC C.,
aN C CD CD 0O C0 CD 0O N
Uptake of antibody at asturation (pg. per 106s cells)
FIG. 3. Comparison of the uptake of 7S antibody by red cells from seventeen individuals.
not absorb. Thus there does not appear to be any A-antigen recognizable by the rabbit onA1 cells that is not present on A2 cells.
Three attempts were made to raise an antibody to A2 cells, with the object of seeingwhether there was a determinant group on A2 cells not present on A1 cells or better able tocombine with an anti-A2 antibody. Two of the rabbits immunized with A2 cells failed toproduce any antibody not absorbable by 0 cells. The third rabbit produced a 7S antibodyof which the greater part was absorbable by 0 cells, although about 10 per cent was A-specific; this contrasted with the sera raised against A1 cells in which the greater part ofthe antibody was A-specific.The anti-A produced in response to the injection of A2 cells appeared to have two com-
ponents. The major component behaved with respect to A1 and A2 cells in the same wayas antibodies from rabbits injected with A1 cells; that is, it was more avidly absorbedby A1 than by A2 cells, and the maximum uptakes of antibody by A1 and A2 cells were ofthe same order as those obtained with anti-A1 sera. The second component, consisting ofperhaps rather less than half the antibody, had poor avidity for both A1 and A2 cells,and consequently it was difficult to estimate it quantitatively, to saturate cells with it,or to say whether it behaved differently towards A1 and A2 cells.
UPTAKE OF 19S ANTIBODY
Fig. 4 shows curves for the uptake of the 19S fraction from sera 86 and 88 by A1 cells.These two sera give results in close agreement: the mean value being 0-28 ,ug. per 106 redcells at saturation; this is of the same order as the uptake of 7S y globulin. If the molecularweight of the 1 9S globulin is taken as one million, this uptake corresponds to only I * 7 x 105molecules of antibody per red cell.
Earlier experiments with serum 62, in which the hypoiodite method was used foriodination and in which the specific activity was known only for the whole unchromato-graphed y-globulin fraction indicated that for this serum also, the uptake at saturation of19S antibody was similar to that of 7S antibody. The maximum uptake (by weight) byA2 cells appeared to be about a quarter that of A1 cells, as with 7S antibody.
A*
427

C. L. Greenbury, D. H. Moore and L. A. C. JNunn032
028 ;8
024
020 --V
0-16
0-12
081 2 4 6 8 10 12 14 16 18 20 22 24X103FIG. 4. Uptake of 19S antibody by A1 cells. Abscissa: Antibody fixed by red blood cells (jig. per 106 cells)Antibody concentration (pig./ml.). Ordinate: Antibody fixed by red blood cells (pg. per 106 cells).
V, Serum 88; A, Serum 86.
It has not been shown conclusively that the 19S and 7S antibodies combine with thesame sites, but the likelihood that this is so, may be inferred from the fact that when wehave exposed red cells to a mixture containing a great excess of both types of antibody theuptake has not exceeded that absorbed in the presence of either antibody alone.
AVIDITY
Provided that the concentration of antibody is sufficiently low to leave the major part ofthe antigen uncombined, such that [Ag-Ab.Ag] [Ag], the law of mass action can berewritten:
K [Ab.Ag] 3[Ag] [Ab -Ab.Ag] (3)
which, at the point where half the antibody is combined with antigen, i.e. when[Ab-Ab.Ag] = [Ab.Ag], becomes:
1 (4)Kz[Ag]Therefore in these conditions the association constant can be defined as the reciprocal
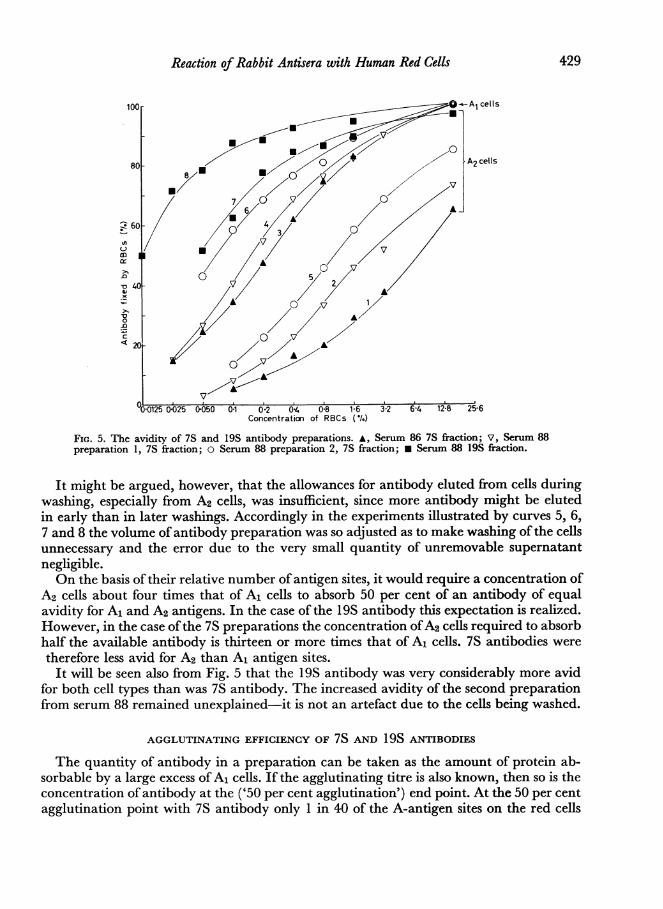
of the antigen concentration required to absorb half the available antibody. When con-structing Fig. 5 it has been found convenient to plot cell concentration, rather than antigenconcentration, and it must be remembered that A1 cells carry four times as many A-antigensites as A2 cells.
Fig. 5 shows a series of curves each ofwhich is derived from the addition of aliquots of asuitably dilute antibody preparation to a range of cell deposits to give the final cellconcentrations shown on the horizontal axis. The vertical axis shows the proportion of theavailable antibody which was taken up by the cells. The total quantity ofantibody availablehas been taken as the amount of antibody absorbed by 16 per cent A1 cells, since thisconcentration ofA1 cells absorbs not less than 95 per cent of antibody, and as stated above,A2 cells are capable of absorbing virtually all the antibody available to A1 cells. In theexperiments from which curves 1, 2, 3 and 4 were derived the cells were washed beforecounting and this process has eluted some antibody; allowance has been made for this bymeasturing the loss of radioactivity at each washing, and extrapolating back to zerowashings.
428
Reaction of Rabbit Antisera with Human Red Cells
Concentration of RBCs (*.)
FIG. 5. The avidity of 7S and 19S antibody preparations. A, Serum 86 7S fraction; V, Serum 88preparation 1, 7S fraction; o Serum 88 preparation 2, 7S fraction; * Serum 88 19S fraction.
It might be argued, however, that the allowances for antibody eluted from cells duringwashing, especially from A2 cells, was insufficient, since more antibody might be elutedin early than in later washings. Accordingly in the experiments illustrated by curves 5, 6,7 and 8 the volume of antibody preparation was so adjusted as to make washing of the cellsunnecessary and the error due to the very small quantity of unremovable supernatantnegligible.On the basis of their relative number of antigen sites, it would require a concentration of
A2 cells about four times that of A1 cells to absorb 50 per cent of an antibody of equalavidity for A1 and A2 antigens. In the case of the 19S antibody this expectation is realized.However, in the case ofthe 7S preparations the concentration ofA2 cells required to absorbhalf the available antibody is thirteen or more times that of A1 cells. 7S antibodies weretherefore less avid for A2 than A1 antigen sites.It will be seen also from Fig. 5 that the 19S antibody was very considerably more avid
for both cell types than was 7S antibody. The increased avidity of the second preparationfrom serum 88 remained unexplained-it is not an artefact due to the cells being washed.
AGGLUTINATING EFFICIENCY OF 7S AND 19S ANTIBODIES
The quantity of antibody in a preparation can be taken as the amount of protein ab-sorbable by a large excess of A1 cells. If the agglutinating titre is also known, then so is theconcentration of antibody at the ('50 per cent agglutination') end point. At the 50 per centagglutination point with 7S antibody only 1 in 40 of the A-antigen sites on the red cells
429

C. L. Greenbury, D. H. Moore and L. A. C. Nunnare bound, and with 19S antibody only 1 in 7000; in these conditions 1 per cent A1 cellsabsorb practically all the antibody present.The initial concentration of 7S antibody necessary to cause 50 per cent agglutination of
1 per cent A1 cells has for sera 86 and 88 varied from 0 59 to 0 45 jig./ml., the mean ofwhichcorresponds to 1.9 x 104 molecules per red cell. For 19S antibody the 1 per cent AD50concentration has lain between 0 004 and 0 0045 pg./ml., corresponding to only twenty-five molecules per cell. Parallel titrations with cells from other donors have shown that theA1 cells used do not differ in agglutinability from the average.
Since so large a number of 7S molecules are required to produce agglutination, thequestion arises whether there is in these preparations a mixture of antibody molecules ofdifferent agglutinating capacity. If the agglutinating antibody was more avid than thenon-agglutinating, it would be possible, using very small quantities of cells to absorb theformer and leave the latter. When aliquots of 7S preparations were absorbed with arange of red cell concentrations, reduction in haemagglutinating titre paralleled theamount of antibody removed. Thus if there are two antibodies their mean avidity must besimilar.
DISCUSSIONThat antibody is heterogeneous and that it will appear in more than one fraction what-
ever means of fractionation is chosen is generally accepted, e.g. Askonas, Farthing andHumphrey (1960). We have found similarly, that although the major part of the 7Santibody appears in the peak eluted from DEAE-cellulose at pH 8 with 0-01 M buffer, thereis also some antibody eluted at pH 7 with 0X025 M buffer. We have used as 7S y globulin theantibody eluted in the first peak, but it cannot be supposed that this is a homogeneousentity and would not be subdivisible by other methods of separation. Nevertheless ifquantitative deductions concerning antibody are to be made from measurement of radio-activity, certain assumptions are necessary; notably that the antibody possesses the samespecific radioactivity as the average for other protein with which it elutes, that its molecularweight is of a known order and that the process of iodination has not significantly alteredits properties.We have no evidence ofour own on the first point, but Masouredis, Melcher and Koblick
(1951) showed that the specific activity of labelled rabbit antibody was the same as thatfor y globulin; also Pressman and Sternberger (1950) found that the specific activity ofantibody bore a relationship to whole y globulin (18 per cent Na2SO4 precipitate) similarto that of our 7S preparation to whole y globulin. We have taken 160,000 and 1,000,000as the molecular weights of the 7S and 19S antibodies respectively. Askonas et al. (1960)have shown that the method ofiodination we have used does not alter the chromatographicproperties of rabbit antibody.
If, in the absence of aggregation, only one molecule of antibody can attach itself to asingle antigenic site (determinant group), there must be at least as many sites on the cellas the number of molecules of antibody that the cell can carry. It then becomes necessaryto consider in what degree our findings define the number of antibody molecules absorb-able by the red cell, and thus the number of A-antigen sites which it possesses.Our absorption curves (Figs. 1 and 2) diverge considerably from linearity. The curvature
is probably due to heterogeneity of association constants within the antigen-antibodysystem; although non-specific absorption of labelled protein which, while remaining low,rises rather steeply in higher antibody concentrations (see Table 1) must also play a small
430

Reaction of Rabbit Antisera with Human Red Cells
part. In agreement with Hughes-Jones, Gardner and Telford (1962) we have found exten-sive heterogeneity within our antibody, but we think that there also exist considerable realor apparent differences among the antigen sites. Such differences could occur in at leastthree ways: (1) Interference between antibody molecules, whereby due to their crowdingit becomes increasingly difficult for late-comers to gain access to antigen sites. (2) Variationin accessibility of antigenic sites due to their topographical distribution; they may forinstance be situated at different distances from the cell surface. (3) Heterogeneity inphysico-chemical configuration among antigenic sites; it seems not unreasonable tosuppose that in view of the qualitative difference demonstrated between A1 and A2 sitesthat there could be a wide variety among the sites on an individual cell, ranging from agood to a very imperfect fit with the antibody. We have no evidence which favours anyone of these possibilities, and they may well all be operative, since they are not mutuallyexclusive. Moreover in so far as there is any merit in suggestions (2) and (3) it becomes ofdoubtful meaning to speak of the number of antigenic sites on the cell surface. In the lightof these arguments then, it must be said that the extrapolations we have made of oursaturation curves do no more than set a minimum for the number of antibody moleculesthat a red cell can take up, but they enable comparisons to be made between differentantibodies and red cells.
Since A1 cells can absorb 8f3 x 105 molecules per cell, they cannot possess fewer than thisnumber of antigenic sites. This figure is of the same order as that calculated by Filliti-Wurmser, Jacquot-Armand, Aubel-Lesure and Wurmser (1954) for the number of sites onB cells, using quite different experimental methods. If the red cell is regarded as having asmooth surface of 165 p2 (Ponder, 1948) then the mean distance between sites is 130 A.But it is probable that each molecule of A-substance possesses many A-sites, in which casemost of the sites will lie closer together than 130 A. Edsall and Foster (1948) and Oncley,Scatchard and Brown (1947) give the length of the human y-globulin molecule as 230 A.If this figure is valid for rabbit 7S y globulin and if the antibody is bivalent, it would atleast be possible, unless sterically prevented, for most antibody molecules to combine withtwo antigen sites; in which case our estimation of the number of sites would be con-siderably too low.The finding that A2 cells possess fewer A-sites than A1 cells is in agreement with recent
work by Goodman (1962) with human antibodies. It also accords well with the hypothesisadvanced by Watkins and Morgan (1957) that A and B specificities are due to the con-version of a common precursor, H-substance, under the influence of the genes concerned,and that the A2 gene is less efficient than the A1 gene in bringing about this conversion. ThatA1B and A2B cells tend to have fewer A-sites than A1 and A2 cells (presumably A10and A20) could then be explained by supposing that a single A-gene converts more thanhalf the H-substance to A-specificity; but when a B-gene is present competing for theavailable H-substance, the A-gene has to be content with a smaller share. This wouldalso account for the finding that A1Al cells do not carry twice as many sites as the averageA10 cells.We have found that the antigen sites on A2 cells, as well as being fewer in number,
differ qualitatively from those on A1 cells, combining less avidly with antibody raisedagainst A1 cells and being poorly antigenic in the rabbit. The presence of anti-Al in A2individuals also suggests that there is a qualitative difference. Nevertheless there appearto be no antibodies produced by the rabbit in response to A1 cells which cannot be ab-sorbed by A2 cells; and conversely the antibody produced against A2 cells was absorbed as
431

C. L. Greenbury, D. H. Moore and L. A. C. Nunnwell by A1 as by A2 cells. It seems then that the qualitative difference between these twoantigens is too subtle to confer different specificities as far as the rabbit is concerned. It maybe that if the A1 gene gives rise to a spectrum of sites which fit the antibody with varyingdegrees of perfection, that the A2 antigen sites, as well as being fewer in number, belong tothe less well fitting part of the spectrum.The findings that A1 cells can take up only a little more 19S than 7S antibody and only
about one-fifth of the number of molecules requires explanation. The three simplestpossibilities are: (a) that 19S molecules have the same valency as 7S molecules, but that amolecule once attached to a site, by virtue of its size, prevents access by other moleculesto five times as many sites as it actually occupies, (b) that 19S antibodies are specific forsites both different from and fewer than those occupied by 7S antibody, (c) that 19Smolecules possess the same number of effective valencies per unit weight as 7S moleculesand combine with five or six times as many sites.The first hypothesis requires the additional postulate of a better antigen-antibody fit in
order to account for the greater avidity of 19S antibody; it also requires the acceptance ofthe coincidence that the degree of occlusion by 19S molecules matches their weight ratherthan, as one might expect, their dimensions. The second hypothesis requires not only adifferent specificity for the 19S antibody, but a better fit with the antigen and, at thesame time, prevention of access to their own sites of 7S antibody; since we have shown thatcells take up no more protein in the presence of an excess of both 7S and 19S antibodiesthan in the presence of either alone. We therefore prefer for the moment the thirdhypothesis, that of multivalency of 19S antibody, which provides the simplest explanationof the experimental results, namely that saturated cells take up approximately the sameweight of 19S and 7S antibody and that the former is much the more avid; the increasedavidity being accounted for by the larger number ofcombining sites on the 19S molecule.It has been shown by Petermann and Pappenheimer (1941) that some 19S antibodiescan be split into smaller bivalent fragments, which retain their specificity; although it isnot clear that more than one such fragment can be derived from a single molecule. How-ever, according to Kunkel (1960) some large molecules lose their antibody activity on'depolymerization'.The 19S antibody has an extraordinary agglutinating efficiency, there being only twenty-
five molecules per cell available at the AD5o. If the cells remain as biconcave discs it isdifficult to conceive more than perhaps one or two of the fifty molecules available on anytwo cells being in a position to bind them together. In contrast at the AD5o of 7S antibody,red cells are each carrying 19,500 molecules of antibody.
It is interesting to speculate on the reason for this very large difference. The first possi-bility to be considered is that the 7S antibody is 'incomplete' and the fraction gains itsagglutinating property from contamination with 19S antibody. This does not seemlikely in our preparations, since in the two '7S' fractions submitted to density gradientultracentrifugation, the agglutinating titre of the bottom fraction in the centrifuge tubewas considerably less (one-eighth) than in the fraction containing the bulk of the antibody.Moreover the 7S antibody is not 'incomplete' in the ordinary sense since trypsinizationof the cells only increases its titre by one or two tubes. A second possibility is that 7Smolecules differ among themselves in their agglutinating ability and that agglutination isdue only to a minor fraction.We have no evidence concerning such heterogeneity within the7S antibody other than our inability to separate fractions ofdifferent agglutinating capacityby differential absorption with small quantities of cells. It may be that the agglutinating
432

Reaction of Rabbit Antisera with Human Red Cells 433
ability of 7S antibody is solely a function of its molecular size. It would be expected from aconsideration of their respective avidities that it would require more 7S than 19S mole-cules to cause agglutination, but it would still be necessary to postulate that only one inmany hundreds of 7S molecules was able to attach itself to two cells simultaneously.There remains a further possibility: we have shown that A-sites are sufficiently close
together for two to lie within the reach of a single 7S molecule; if there is no steric impedi-ment it would seem that a molecule attached to an antigen site by one of its specific groupswould be more likely to find a site for its second specific group on its own rather than aneighbouring cell. Molecules attached to a single cell by both their 'valencies' would thenbe unable to take part in agglutination. Whether or not 7S molecules are in fact attachedby their two combining sites to a single cell might be resolved by measuring the uptake ofmonovalent molecular fragments such as those produced by Porter (1959) by papaindigestion. We are at present investigating this possibility.
ACKNOWLEDGMENTS
This work was undertaken with the assistance of a research grant from the Oxford RegionalHospital Board.We thank Dr. E. J. Holborow for ultracentrifuging our preparations, Dr. Muriel Seaman for
many donations of blood, Dr. T. Freeman for demonstrating McFarlane's method of iodination,Mr. P. S. Golding, who took part in the earlier experimental work and Mrs. Jeanne Smith forpreparing the diagrams.
REFERENCES
ASKONAS, B. A., FARTHING, C. P. and HUMPHREY, J. H.(1960). 'The significance of multiple antibodycomponents in serum of immunized rabbits.'Immunology, 3, 336.
COSTEA, N., SCHWARTZ, R., CONSTANTOULAKIS, M.and DAMESHEK, W. (1962). 'The use of radioactiveantiglobulin for the detection of erythrocyte sensi-tization.' Blood, 20, 214.
EDSALL, J. T. and FOSTER, J. F. (1948). 'Studies ondouble refraction of flow. IV. Human serumy-globulin and crystallized bovine serum albumin.'J. Amer. chem. Soc., 70, 1860.
FILIrrI-WuRMsER, S., JACQUOT-ARMAND, Y., AUBEL-LESURE, G. and WURMSER, R. (1954). 'A physico-chemical study of human isohaemagglutination.'Ann. Eugenics, 18, 183.
GOODMAN, H. S. (1962). 'Antigen and antibody-combining properties and their influence on theimmune reaction of red cells.' Nature (Lond.), 194,934.
HUGHES-JONES, N. C., GARDNER, B. and TELFORD, R.(1962). 'The kinetics of the reaction between theblood-group antibody anti-c and erythrocytes.'Biochem. j., 85, 466.
KABAT, E. A. (1956). Blood Group Substances, TheirChemistry and Immunochemistry, p. 290. AcademicPress, New York.
KUNKEL, H. G. (1960). 'Macroglobulins and highmolecular weight antibodies.' The Plasma Proteins(Ed. F. W. Putnam), vol. 1, p. 291. AcademicPress, New York.
MASOUREDIS, S., MELCHER, L. R. and KOBLICK, D. C.(1951). 'Specificity of radio-iodinated (1311.)
immune globulin as determined by quantitativeprecipitation reactions.'_J. Immunol., 66, 297.
McFARLANE, A. S. (1958). 'Efficient trace-labelling ofproteins with iodine.' Nature (Lond.), 182, 53.
ONCLEY, J. L., SCATCHARD, G. and BROWN, A. (1947).'Physico-chemical characteristics of certain of theproteins of normal human plasma.' J. Phys. ColloidChem., 51, 184.
PETERMANN, M. L. and PAPPENHEIMER, A. M., JUNR.(1941). 'The action of crystalline pepsin on horseanti-pneumococcus antibody.' Science, 93, 458.
PONDER, E. (1948). Hemolysis and Related Phenomena.Grune & Stratton, New York.
PORTER, R. R. (1959). 'The hydrolysis of rabbity-globulin and antibodies with crystalline papain.'Biochem. J., 73, 119.
PREssMAN, D. and STERNBERGER, L. A. (1950). 'Therelative rates of iodination ofserum components andthe effect of iodination on antibody activity.'J. Amer. chem. Soc., 72, 2226.
ROCKEY, J. H. and KUNKEL, H. G. (1961). 'Studies ofthe rabbit antibodies which sensitize red blood cellsfor agglutination by rheumatoid factors.' Arthr. andRheum., 4, 449.
SOBER, H. A., GUTTER, F. J., WYCKOFF, MI. W. andPETERSON, E. A. (1956). 'Chromatography ofproteins. II. Fractionation of serum protein onanion-exchange cellulose.' J. Amer. chem. Soc., 78,756.
WATKINS, W. M. and MORGAN, W. T. J. (1957).'The A and H character of the blood group sub-stances secreted by persons belonging to group A2.'Acta genet. statist. med., 6, 521.


















