
Rapid Publications Growth Factor/Matrix-Induced...
3
Rapid Publications Growth Factor/Matrix-Induced Proliferation of Human Adult 0-Cells Alberto Hayek, Gillian M. Beattie, Vincenzo Cirulli, Ana D. Lopez, Camillo Ricordi, and Jeffrey S. Rubin Proliferation of human (J-cells in vitro is desirable for both transplantation and biological studies. In this study, human pancreatic islets obtained from cadavers were kept in tissue culture plates that favored cell attachment. When the cells attached to the matrix produced by the rat-bladder carcinoma cell line 804G, 5'-bromo-2'-de- oxyuridine (BrdU) labeling increased from 4.7 ± 2.5 to 13.2 ± 2.2%, while cells simultaneously labeled for insulin and BrdU increased from 0 to 32%. Addition of the growth factor hepatocyte growth factor/scatter factor (HGF/SF) increased BrdU labeling to 17.5 ± 1.8 and the percentage of double positive (BrdU + insulin) cells to 69%. This is the first in vitro demonstration that human P-cells grown in monolayer culture are able to replicate when exposed to selected matrices and growth factors. These experi- ments add further evidence that HGF/SF is an important mitogenic agent for human p-cells. Diabetes 44:1458- 1460, 1995 S ignificant in vitro proliferation of (3-cells obtained from the human adult pancreas is important for both transplantation and biological studies. In con- trast to the extensive literature available on rodent fetal and adult islet growth and the growth factors involved in such processes (1,2), information concerning human adult islets' growth potential is scant. We have previously shown that insulin content and secretion are enhanced during short-term culture of human adult islets by using biocompat- ible extracellular matrices favoring cell attachment (3). However, under the conditions described, we were unable to induce proliferation of P-cells. More recently, we found that the matrix secreted by the rat bladder carcinoma cell line 804G (4) is superior to other matrices in supporting the growth of islet-like cell clusters (ICCs) derived from mid- gestation human fetal pancreases (unpublished data). In related experiments, we screened a variety of peptide growth From the Whittier Institute (A.H., G.M.B., V.C., A.D.L.), Department of Pediatrics, University of California San Diego, La Jolla, California; the Diabetes Research Institute (C.R.), University of Miami, Miami, Florida; and the Laboratory of Cellular and Molecular Biology (J.S.R.), National Cancer Institute, Bethesda, Maryland. Address correspondence and reprint requests to Dr. Alberto Hayek, Whittier Institute, Dept. of Pediatrics, UCSD School of Medicine, 9894 Genesee Ave., La Jolla, CA 92037. Received for publication 18 September 1995 and accepted 28 September 1995. A.H. is a paid consultant of Desmos, Inc., San Diego, CA. BrdU, 5'-bromo-2'-deoxyuridine; HGF/SF, hepatocyte growth factor/scatter fac- tor; ICC, islet-like cell cluster; NIDDM, non-insulin-dependent diabetes mellitus; PBS, phosphate-buffered saline. factors for mitogenic, morphogenic, and insulinotropic ef- fects on cultured ICCs, finding that hepatocyte growth fac- tor/scatter factor (HGF/SF) had a marked mitogenic effect on the human fetal (3-cells contained within ICCs (5). In addi- tion, insulin content in ICCs was significantly increased in the presence of the growth factor. In the present study, we report that the combination of the 804G matrix and HGF/SF causes (3-cell proliferation in monolayer cultures of human adult islets. RESEARCH DESIGN AND METHODS Tissue culture experiments. Human adult islets were isolated with an automated method as described previously (6) and were further purified by handpicking single islets 50-100 |xm in diameter after dithizone staining (7). Measurements of cell numbers were obtained with repli- cates of 50 islets grown in monolayers for 5 days in 35-mm tissue culture dishes coated with 804G matrix before removal with nonenzymatic cell dissociation solution (Sigma, St. Louis, MO) for counting with a hema- cytometer. During incubation at 37°C in a humidified atmosphere with 5% CO 2 , the islets were kept in RPMI-1640 medium (Gibco, Grand Island, NY) containing 10% pooled normal human serum and antibiotics (100 U/ml penicillin, 0.1 mg/ml streptomycin, and 1 |xg/ml amphotericin B; Sigma). Glucose was added to the medium at 5.5 mmol/1 because of the finding that prolonged exposure to higher levels impairs function (8). Fresh medium and HGF/SF at a concentration of 25 ng/ml, previously found to be effective (5), were provided every 48 h. DNA synthesis and insulin content and secretion. Monolayers derived from 50 islets per well were pulsed with 1.0 |xCi/ml of [methyl- 3 H]thymidine (specific activity 25 Ci/mmol; Amersham, Arlington, IL) in newly replenished medium. After 16 h, medium was collected to determine insulin secretion, while thymidine incorporation into DNA and insulin extractable from cells were quantified as before (3,9). Basal insulin release and insulin release in response to glucose stimulation were determined in 1-h static incubations in the presence of 1.6 and 16.7 mmol/1 glucose, respectively (3). For these experiments, HGF/SF was first removed by washing the monolayers twice with phosphate-buffered saline (PBS) and incubating for 1 h in the presence of 1.6 mmol/1 glucose. Insulin was measured with a solid-phase radioimmunoassay (DPC, Los Angeles, CA), and DNA content was measured fluorometri- cally (10). Incorporation of [ 3 H]thymidine was determined by liquid scintillation counting of trichloroacetic acid precipitates of the soni- cated cells. Immunocytochemistry. After 3 days in monolayer culture, the cells were incubated for 24 h with 0.1 mmol/1 5'-bromo-2'-deoxvuridine (BrdU) and fixed in 4% paraformaldehyde for detection of cells contain- ing insulin and/or BrdU (5). Double antigen-specific staining was visu- alized with the alkaline phosphatase (11) and immunoperoxidase (12) techniques. For morphometric analyses, the investigator was blinded to the identity of the samples. Insulin and BrdU immunoreactivity on islet cell monolayers was also identified by double immunofluorescence using confocal microscopy. After fixation, the cell monolayers were permeabilized in 0.1% saponin for 5 min and incubated in 50 mmol/1 glycine in PBS to saturate reactive groups generated by formaldehyde fixation. Nonspecific binding was blocked by incubation with 2% donkey normal serum and 1% bovine serum albumin before exposure to a 1458 DIABETES, VOL. 44, DECEMBER 1995
Transcript of Rapid Publications Growth Factor/Matrix-Induced...

Rapid PublicationsGrowth Factor/Matrix-Induced Proliferation of HumanAdult 0-CellsAlberto Hayek, Gillian M. Beattie, Vincenzo Cirulli, Ana D. Lopez, Camillo Ricordi,and Jeffrey S. Rubin
Proliferation of human (J-cells in vitro is desirable forboth transplantation and biological studies. In this study,human pancreatic islets obtained from cadavers werekept in tissue culture plates that favored cell attachment.When the cells attached to the matrix produced by therat-bladder carcinoma cell line 804G, 5'-bromo-2'-de-oxyuridine (BrdU) labeling increased from 4.7 ± 2.5 to13.2 ± 2.2%, while cells simultaneously labeled for insulinand BrdU increased from 0 to 32%. Addition of the growthfactor hepatocyte growth factor/scatter factor (HGF/SF)increased BrdU labeling to 17.5 ± 1.8 and the percentageof double positive (BrdU + insulin) cells to 69%. This isthe first in vitro demonstration that human P-cells grownin monolayer culture are able to replicate when exposedto selected matrices and growth factors. These experi-ments add further evidence that HGF/SF is an importantmitogenic agent for human p-cells. Diabetes 44:1458-1460, 1995
Significant in vitro proliferation of (3-cells obtainedfrom the human adult pancreas is important forboth transplantation and biological studies. In con-trast to the extensive literature available on rodent
fetal and adult islet growth and the growth factors involvedin such processes (1,2), information concerning human adultislets' growth potential is scant. We have previously shownthat insulin content and secretion are enhanced duringshort-term culture of human adult islets by using biocompat-ible extracellular matrices favoring cell attachment (3).However, under the conditions described, we were unable toinduce proliferation of P-cells. More recently, we found thatthe matrix secreted by the rat bladder carcinoma cell line804G (4) is superior to other matrices in supporting thegrowth of islet-like cell clusters (ICCs) derived from mid-gestation human fetal pancreases (unpublished data). Inrelated experiments, we screened a variety of peptide growth
From the Whittier Institute (A.H., G.M.B., V.C., A.D.L.), Department of Pediatrics,University of California San Diego, La Jolla, California; the Diabetes ResearchInstitute (C.R.), University of Miami, Miami, Florida; and the Laboratory of Cellularand Molecular Biology (J.S.R.), National Cancer Institute, Bethesda, Maryland.
Address correspondence and reprint requests to Dr. Alberto Hayek, WhittierInstitute, Dept. of Pediatrics, UCSD School of Medicine, 9894 Genesee Ave., LaJolla, CA 92037.
Received for publication 18 September 1995 and accepted 28 September 1995.A.H. is a paid consultant of Desmos, Inc., San Diego, CA.BrdU, 5'-bromo-2'-deoxyuridine; HGF/SF, hepatocyte growth factor/scatter fac-
tor; ICC, islet-like cell cluster; NIDDM, non-insulin-dependent diabetes mellitus;PBS, phosphate-buffered saline.
factors for mitogenic, morphogenic, and insulinotropic ef-fects on cultured ICCs, finding that hepatocyte growth fac-tor/scatter factor (HGF/SF) had a marked mitogenic effect onthe human fetal (3-cells contained within ICCs (5). In addi-tion, insulin content in ICCs was significantly increased inthe presence of the growth factor. In the present study, wereport that the combination of the 804G matrix and HGF/SFcauses (3-cell proliferation in monolayer cultures of humanadult islets.
RESEARCH DESIGN AND METHODSTissue culture experiments. Human adult islets were isolated with anautomated method as described previously (6) and were further purifiedby handpicking single islets 50-100 |xm in diameter after dithizonestaining (7). Measurements of cell numbers were obtained with repli-cates of 50 islets grown in monolayers for 5 days in 35-mm tissue culturedishes coated with 804G matrix before removal with nonenzymatic celldissociation solution (Sigma, St. Louis, MO) for counting with a hema-cytometer. During incubation at 37°C in a humidified atmosphere with5% CO2, the islets were kept in RPMI-1640 medium (Gibco, Grand Island,NY) containing 10% pooled normal human serum and antibiotics (100U/ml penicillin, 0.1 mg/ml streptomycin, and 1 |xg/ml amphotericin B;Sigma). Glucose was added to the medium at 5.5 mmol/1 because of thefinding that prolonged exposure to higher levels impairs function (8).Fresh medium and HGF/SF at a concentration of 25 ng/ml, previouslyfound to be effective (5), were provided every 48 h.DNA synthesis and insulin content and secretion. Monolayersderived from 50 islets per well were pulsed with 1.0 |xCi/ml of [methyl-3H]thymidine (specific activity 25 Ci/mmol; Amersham, Arlington, IL) innewly replenished medium. After 16 h, medium was collected todetermine insulin secretion, while thymidine incorporation into DNAand insulin extractable from cells were quantified as before (3,9). Basalinsulin release and insulin release in response to glucose stimulationwere determined in 1-h static incubations in the presence of 1.6 and 16.7mmol/1 glucose, respectively (3). For these experiments, HGF/SF wasfirst removed by washing the monolayers twice with phosphate-bufferedsaline (PBS) and incubating for 1 h in the presence of 1.6 mmol/1glucose. Insulin was measured with a solid-phase radioimmunoassay(DPC, Los Angeles, CA), and DNA content was measured fluorometri-cally (10). Incorporation of [3H]thymidine was determined by liquidscintillation counting of trichloroacetic acid precipitates of the soni-cated cells.
Immunocytochemistry. After 3 days in monolayer culture, the cellswere incubated for 24 h with 0.1 mmol/1 5'-bromo-2'-deoxvuridine(BrdU) and fixed in 4% paraformaldehyde for detection of cells contain-ing insulin and/or BrdU (5). Double antigen-specific staining was visu-alized with the alkaline phosphatase (11) and immunoperoxidase (12)techniques. For morphometric analyses, the investigator was blinded tothe identity of the samples. Insulin and BrdU immunoreactivity on isletcell monolayers was also identified by double immunofluorescenceusing confocal microscopy. After fixation, the cell monolayers werepermeabilized in 0.1% saponin for 5 min and incubated in 50 mmol/1glycine in PBS to saturate reactive groups generated by formaldehydefixation. Nonspecific binding was blocked by incubation with 2% donkeynormal serum and 1% bovine serum albumin before exposure to a
1458 DIABETES, VOL. 44, DECEMBER 1995
A. HAYEK AND ASSOCIATES
TABLE 1Effect of HGF/SF on replication and insulin content and releasefrom adult human islets grown in monolayer on 804G matrix
% of eachindividual control
Cell number16-h insulin secretion/cellInsulin release after glucose stimulation[3H]thymidine incorporation/|xg DNAInsulin content/(xg DNA
139 ± 9t54 ± 18*97 ±81:
272 ± 63t137 ± 24*
Data are means ± SE. Replicates (n = 6) of 50 islets per matrix-coated well were cultured for 5 days in RPMI medium containing 5.5mmol/1 glucose in the presence or absence of 25 ng/ml HGF/SF.*P < 0.05; tP < 0.001; $NS.
mixture of two primary antibodies: IgG fraction of a sheep anti-humaninsulin polyclonal antiserum (The Binding Site, Birmingham, U.K.) andbiotinylated mouse anti-BrdU monoclonal antibody (Caltag Laborato-ries, San Francisco, CA). After washing, the monolayers were incubatedwith lissamine/rhodamine-conjugated affinity-purified donkey anti-sheepIgG(H+L) preadsorbed on chicken, guinea pig, hamster, horse, human,mouse, rabbit, and rat serum proteins (Jackson Immunoresearch, WestGrove, PA). After several washes, fluorescein isothiocyanate-conju-gated streptavidin (Jackson Immunoresearch) was applied and themonolayers were again washed extensively before mounting in slow-fade medium (Molecular Probes, Eugene, OR). They were then viewedon a Zeiss Axiovert 35M microscope using a 40x 1.3 NA objective lensequipped with a laser scanning confocal attachment (MRC-1024, Bio-Rad, Cambridge, MA). Fluorescent images relative to each marker werecollected using an argon/krypton mixed-gas laser. Color compositeimages were printed on a Tektronix Phaser II-SDX.Statistical analysis. Except where indicated, data are presented aspercentage changes from control conditions (monolayers grown on804G matrix in the absence of HGF/SF; means ± SE). Significantdifferences of the relative (percentage) changes were evaluated withStudent's t test using 100 as the population mean. For the morphometricstudies, data were analyzed with software for the Macintosh (StatviewIV, Abacus Concepts, Berkeley, CA). Statistical significance of observeddifferences was tested using analysis of variance and Fisher's protectedleast significance difference test with 95% as the limit of significance.
RESULTS AND DISCUSSIONAddition of HGF/SF to adult human islets grown in mono-layer culture on 804G matrix had a significant stimulatoryeffect as reflected by the 39% increase in cell number and172% increase in [3H]thymidine incorporation into DNA after5 days of culture. The level of extractable insulin, normalizedto DNA content, was increased 37%, while secretion into themedium was decreased by almost 50%. Replacement of themedium containing the HGF/SF with buffer containing lowand high glucose concentrations produced the same acute
TABLE 2Morphometric analysis of adult islet monolayers immunostainedfor BrdU and insulin
Condition
PlasticPlastic + HGF/SF804G804G + HGF/SF
BrdUlabeling
index (% oftotal cells)
4.75 ± 2.562.5 ± 0.87
13.25 ± 2.29*17.5 ± 1.85t
Insulin-containingcells (% oftotal cells)
88.5 ± 2.8487.75 ± 3.0482.5 ± 3.1290.5 ± 1.56
Double positivefor insulin +
BrdU (% of totalBrdU cells)
0403269
Data are means ± SE, or sum of all double stained cells counted infour dishes. Replicates (n = 4) of 50 islets per well (coated oruncoated) were cultured as in TABLE 1. BrdU (0.1 mmol/1) wasadded 24 h before fixation and double immunostaining. At least 100cells were counted in each replicate. *P < 0.01; tP < 0.001 com-pared with plastic alone.
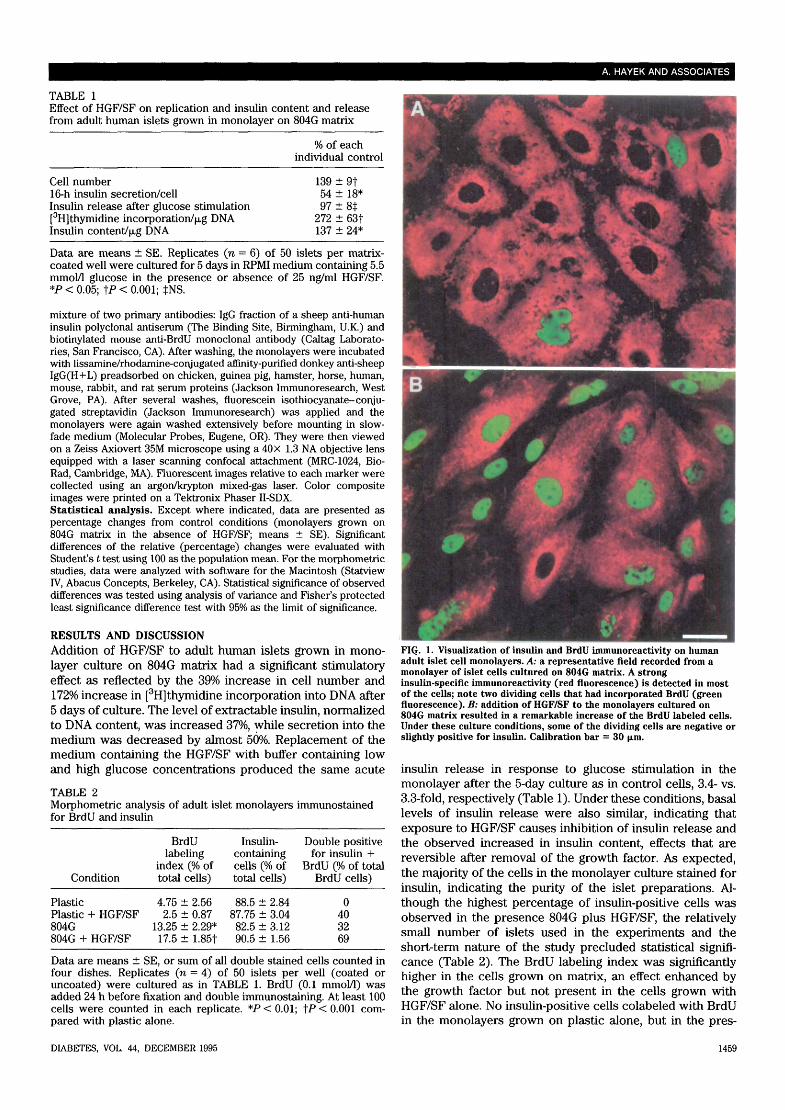
FIG. 1. Visualization of insulin and BrdU inununoreactivity on humanadult islet cell monolayers. A: a representative field recorded from amonolayer of islet cells cultured on 804G matrix. A stronginsulin-specific immunoreactivity (red fluorescence) is detected in mostof the cells; note two dividing cells that had incorporated BrdU (greenfluorescence). B: addition of HGF/SF to the monolayers cultured on804G matrix resulted in a remarkable increase of the BrdU labeled cells.Under these culture conditions, some of the dividing cells are negative orslightly positive for insulin. Calibration bar = 30 \im.
insulin release in response to glucose stimulation in themonolayer after the 5-day culture as in control cells, 3.4- vs.3.3-fold, respectively (Table 1). Under these conditions, basallevels of insulin release were also similar, indicating thatexposure to HGF/SF causes inhibition of insulin release andthe observed increased in insulin content, effects that arereversible after removal of the growth factor. As expected,the majority of the cells in the monolayer culture stained forinsulin, indicating the purity of the islet preparations. Al-though the highest percentage of insulin-positive cells wasobserved in the presence 804G plus HGF/SF, the relativelysmall number of islets used in the experiments and theshort-term nature of the study precluded statistical signifi-cance (Table 2). The BrdU labeling index was significantlyhigher in the cells grown on matrix, an effect enhanced bythe growth factor but not present in the cells grown withHGF/SF alone. No insulin-positive cells colabeled with BrdUin the monolayers grown on plastic alone, but in the pres-
DIABETES, VOL. 44, DECEMBER 1995 1459

INDUCED PROLIFERATION OF (i-CELLS
ence of either HGF/SF or 804G matrix, 30-40% of theBrdU-labeled cells were insulin positive. The combination of804G matrix with HGF/SF was additive, as judged by thefrequency of cells colabeling for insulin and BrdU (Table 2,Fig. 1).
These results are the first to demonstrate a significanteffect of a growth factor, HGF/SF, in conjunction with matrixon p-cell replication in adult human islets in culture. Earlier,we had shown that islet cell function was influenced byextracellular matrices that facilitated attachment to theculture dish, but none of the matrices used showed asignificant effect on p-cell replication (3). In this study, weobserved that the matrix produced by 804G cells can itselfpromote (3-cell proliferation and that its effects were additiveto those of HGF/SF. This growth factor was chosen becausea previous survey of several growth factors indicated thatHGF/SF had a significant insulinotropic activity increasingthe proportion of insulin-positive cells and the number ofreplicating p-cells in human fetal ICCs (5). Other laboratorieshave reported increased insulin secretion from free-floatinghuman islets exposed to hormones such as placental lacto-gen, prolactin, and growth hormone, but the limited data onislet cell replication showed only an increase from 0.2 to 1%(assuming a cell content of ~ 1,000 cells per islet) (13). Incontrast, our results demonstrated an increase in BrdUlabeling index from 4.75% for cells plated on plastic to 17.5%for cells on 804G matrix treated with HGF/SF. Furthermore,while apparently none of the cells replicating on plastic were(3-cells, as judged by absence of insulin-staining, ~70% of thecells undergoing DNA synthesis on matrix plus HGF/SFcontained insulin and therefore were presumed to be p-cells.A recent Perspectives in Diabetes on the dynamics of (3-cellmass points out that the low rate of replication of (3-cells inthe rat pancreas has led some authors to suggest that one isborn with "all the (B-cells one will ever have" and to questionthe existence of a (3-cell population that retains the ability toproliferate (14). The information presented here shows thatmost (3-cells in the human pancreas retain the capacity toreplicate in vitro when exposed to mitogenic stimuli such asthose provided by HGF/SF and 804G matrix. Further re-search will be required to determine whether these replica-tive mechanisms also operate in vivo.
In summary, we present data documenting the ability ofthe extracellular matrix produced by 804G cells and HGF/SFto stimulate replication of human adult (3-cells in monolayercultures. These findings suggest a means of increasing the
number of cultured (3-cells for clinical research and a newmodel system for the study of processes that regulate thegrowth of fully differentiated pancreatic endocrine cells.
ACKNOWLEDGMENTSThis work was supported by the National Institute of Diabe-tes and Digestive and Kidney Diseases Grant RO1-DK-39087and The Herbert O. Perry Fund. V.C. is the recipient of afellowship from the Swiss National Science Fund.
We are grateful to Dr. Vito Quaranta at the ScrippsResearch Institute, La Jolla, CA, for providing material forthe matrix used in these studies and to Dr. Mark H. Ellismanfor his help in performing the confocal microscopy studies.
REFERENCES1. Bonner-Weir S, Smith FE: Islet cell growth and the growth factors
involved. Trends Endocrinol Metab 5:60-64, 19942. Hellerstrom C, Swenne I: Growth patterns of pancreatic islets. In The
Diabetic Pancreas. Volk BW, Wellman KF, Eds. New York, Plenum, 1985,p. 53-79
3. Beattie GM, Lappi DA, Baird A, Hayek A: Functional impact of attach-ment and purification in the short term culture of human pancreaticislets. J Clin Endocrinol Metab 73:93-98, 1991
4. Langhofer M, Hopkinson SB, Jones JCR: The matrix secreted by 804Gcells contains laminin-related components that participate in hemides-mosome assembly in vitro. J Cell Sci 105:753-764, 1993
5. Otonkoski T, Beattie GM, Rubin JS, Lopez AD, Baird A, Hayek A:Hepatocyte growth factor/scatter factor induced growth of human fetalpancreatic (3-cells in vitro. Diabetes 43:947-953, 1994
6. Ricordi C, Lacy PE, Finke EH, Olack BJ, Scharp DW: Automated methodfor isolation of human pancreatic islets. Diabetes 37:413-420, 1988
7. Latif ZA, Noel J, Alejandro R: A simple method of staining fresh andcultured islets. Transplantation 45:827-830, 1988
8. Eizirik DL, Korbutt GS, Hellerstrom C: Prolonged exposure of humanpancreatic islets to high glucose concentrations in vitro impairs the B-cellfunction. J Clin Invest 90:1263-1268, 1992
9. Otonkoski T, Beattie GM, Mally MI, Ricordi C, Hayek A: Nicotinamide isa potent inducer of endocrine differentiation in cultured human fetalpancreatic cells. J Clin Invest 92:1459-1466, 1993
10. Hinegardner RT: An improved fluorometric assay for DNA. Anal Biochem39:197-201, 1971
11. Erber WN, Mason DY: Immunoalkaline phosphatase labeling of terminaltransferase in hematologic samples. Am J Clin Pathol 88:43-50, 1987
12. Sternberger LA, Hardy PH, Cuculis JJ, Meyer HG: The unlabeled antibodymethod of immunohistochemistry: preparation and properties of solubleantigen-antibody complex (horseradish peroxidase-antiperoxidase) andits use in identification of spirochetes. J Histochem Cytochem 18:315—333, 1970
13. Brelje TC, Scharp DW, Lacy PE, Ogren L, Talamantes F, Robertson M,Friesen HG, Sorenson RL: Effect of homologous placental lactogens,prolactins, and growth hormones on islet B-cell division and insulinsecretion in rat, mouse, and human islets: implication for placentallactogen regulation of islet function during pregnancy. Endocrinology132:879-887, 1993
14. Finegood DT, Scaglia L, Bonner-Weir S: Dynamics of (3-cell mass in thegrowing rat pancreas: estimation with a simple mathematical model.Diabetes 44:249-256, 1995
1460 DIABETES, VOL. 44, DECEMBER 1995


















