Provision of carbon skeletons for amide synthesis in non-nodulated soybean and pea roots in response...
6
Soil Science and Plant Nutrition (2008) 54, 732–737 doi: 10.1111/j.1747-0765.2008.00293.x © 2008 Japanese Society of Soil Science and Plant Nutrition Blackwell Publishing Ltd Carbon skeletons of amides in legume roots S. Ueda et al. ORIGINAL ARTICLE/SHORT PAPER Provision of carbon skeletons for amide synthesis in non-nodulated soybean and pea roots in response to the source of nitrogen supply Satoshi UEDA 1 , Motoki IKEDA 2 and Takeo YAMAKAWA 2 1 Department of Plant Resources, Graduate School of Bioresource and Bioenvironmental Sciences and 2 Department of Plant Resources, Faculty of Agriculture, Kyushu University, Fukuoka 812-8581, Japan Abstract Soluble amino acids in roots and primary amino acids, which were involved in primary ammonium assimi- lation, in the metabolites of 14 C-glucose fed to roots for 3 h in the dark were analyzed in the roots of non-nodulated soybean and pea plants grown in ammonium, nitrate or nitrogen-free media for 1 day. Compared with the effect of nitrate, ammonium supply strongly affected the content and synthesis of the amino acids in the roots. In both soybean and pea roots, the supply of ammonium increased considerably the concentrations of the primary amino acids, and asparagine was the most predominant amide, followed by glutamine. In nitrate-supplied soybean roots, the concentrations of asparagine, aspartate and alanine increased, but the concentration of glutamine was low. In the roots of pea plants grown in nitrate media, asparagine was the predominant amino acid, although the composition of the primary amino acids was little affected by nitrate supply. The proportion of amino acids synthesized from 14 C-glucose increased and asparagine rather than glutamine was predominantly synthesized in ammonium-supplied soybean and pea roots, whereas in nitrate-supplied roots asparagine was more actively synthesized than glutamine, although asparagine was not predominant. The ratio of C4 (asparagine + aspartate) to C5 (glutamine + glutamate) amino acids was twofold higher in ammonium-supplied and nitrate-supplied soybean roots than in roots receiving no nitrogen. In contrast, in pea roots, the C4/C5 ratio was twofold higher only in ammonium nutrition. The results obtained suggest that the roots of leguminous plants might possess an indigenous ability to provide a carbon skeleton for preferential synthesis of asparagine rather than glutamine with a high intensity of ammonium supply. Key words: asparagine synthesis, carbon skeleton, glutamine synthesis, leguminous plant, nitrogen nutrition. INTRODUCTION When an excess of ammonium nitrogen is exposed to roots, the synthesis of amides, such as glutamine (Gln) and asparagine (Asn), is enhanced considerably (Givan 1979). Continuous synthesis of both amides requires the provision of carbon skeletons, oxaloacetate for Asn and 2-oxoglutarate for Gln, which are produced in the roots from sugars transported from the shoots (Corzzi and Last 2000). The diminishing pools of these oxo-acids accompanied by ammonium assimilation in roots have to be replenished by the products of glycolysis, stimulated dark carbon fixation and the TCA cycle (Givan 1979; Ikeda et al. 1992). After ammonium (4 mmol L –1 ) was supplied to various plants for 24 h, Asn was the most predominant among the amino acids synthesized from 14 C-glucose in wheat and rice roots, whereas in tomato and cucumber roots it was Gln (Ikeda et al. 2004, 2005; Koga and Ikeda 2000). These results suggest that there may be a dif- ference among plant species in the predominance of amide synthesis in roots in response to ammonium supply. Several non-nodulated leguminous plants, even ureide exporters, transport Asn increasingly as assimilates of combined nitrogen in ammonium or nitrate nutrition (Atkins et al. 1980; McClure and Israel 1979; Oaks 1992; Correspondence: S. UEDA, Department of Plant Resources, Graduate School of Bioresource and Bioenvironmental Sciences, Kyushu University, Fukuoka 812-8581, Japan. Email: [email protected] Received 1 July 2007. Accepted for publication 21 April 2008.
-
Upload
satoshi-ueda -
Category
Documents
-
view
212 -
download
0
Transcript of Provision of carbon skeletons for amide synthesis in non-nodulated soybean and pea roots in response...

Soil Science and Plant Nutrition (2008) 54, 732–737 doi: 10.1111/j.1747-0765.2008.00293.x
© 2008 Japanese Society of Soil Science and Plant Nutrition
Blackwell Publishing LtdCarbon skeletons of amides in legume rootsS. Ueda et al.ORIGINAL ARTICLE/SHORT PAPER
Provision of carbon skeletons for amide synthesis in non-nodulated soybean and pea roots in response to the source of nitrogen supply
Satoshi UEDA1, Motoki IKEDA2 and Takeo YAMAKAWA2
1Department of Plant Resources, Graduate School of Bioresource and Bioenvironmental Sciences and 2Department of Plant Resources, Faculty of Agriculture, Kyushu University, Fukuoka 812-8581, Japan
Abstract
Soluble amino acids in roots and primary amino acids, which were involved in primary ammonium assimi-lation, in the metabolites of 14C-glucose fed to roots for 3 h in the dark were analyzed in the roots ofnon-nodulated soybean and pea plants grown in ammonium, nitrate or nitrogen-free media for 1 day.Compared with the effect of nitrate, ammonium supply strongly affected the content and synthesis of theamino acids in the roots. In both soybean and pea roots, the supply of ammonium increased considerablythe concentrations of the primary amino acids, and asparagine was the most predominant amide, followedby glutamine. In nitrate-supplied soybean roots, the concentrations of asparagine, aspartate and alanineincreased, but the concentration of glutamine was low. In the roots of pea plants grown in nitrate media,asparagine was the predominant amino acid, although the composition of the primary amino acids was littleaffected by nitrate supply. The proportion of amino acids synthesized from 14C-glucose increased andasparagine rather than glutamine was predominantly synthesized in ammonium-supplied soybean andpea roots, whereas in nitrate-supplied roots asparagine was more actively synthesized than glutamine,although asparagine was not predominant. The ratio of C4 (asparagine + aspartate) to C5 (glutamine+ glutamate) amino acids was twofold higher in ammonium-supplied and nitrate-supplied soybean rootsthan in roots receiving no nitrogen. In contrast, in pea roots, the C4/C5 ratio was twofold higher only inammonium nutrition. The results obtained suggest that the roots of leguminous plants might possess anindigenous ability to provide a carbon skeleton for preferential synthesis of asparagine rather than glutaminewith a high intensity of ammonium supply.
Key words: asparagine synthesis, carbon skeleton, glutamine synthesis, leguminous plant, nitrogen nutrition.
INTRODUCTION
When an excess of ammonium nitrogen is exposed toroots, the synthesis of amides, such as glutamine (Gln)and asparagine (Asn), is enhanced considerably (Givan1979). Continuous synthesis of both amides requiresthe provision of carbon skeletons, oxaloacetate for Asnand 2-oxoglutarate for Gln, which are produced in theroots from sugars transported from the shoots (Corzzi
and Last 2000). The diminishing pools of these oxo-acidsaccompanied by ammonium assimilation in roots haveto be replenished by the products of glycolysis, stimulateddark carbon fixation and the TCA cycle (Givan 1979;Ikeda et al. 1992).
After ammonium (4 mmol L–1) was supplied to variousplants for 24 h, Asn was the most predominant amongthe amino acids synthesized from 14C-glucose in wheatand rice roots, whereas in tomato and cucumber rootsit was Gln (Ikeda et al. 2004, 2005; Koga and Ikeda2000). These results suggest that there may be a dif-ference among plant species in the predominance ofamide synthesis in roots in response to ammonium supply.
Several non-nodulated leguminous plants, even ureideexporters, transport Asn increasingly as assimilates ofcombined nitrogen in ammonium or nitrate nutrition(Atkins et al. 1980; McClure and Israel 1979; Oaks 1992;
Correspondence: S. UEDA, Department of Plant Resources,Graduate School of Bioresource and Bioenvironmental Sciences,Kyushu University, Fukuoka 812-8581, Japan. Email: [email protected] 1 July 2007.Accepted for publication 21 April 2008.

Carbon skeletons of amides in legume roots 733
© 2008 Japanese Society of Soil Science and Plant Nutrition
Ohyama et al. 1989; Pate 1980). Xylem sap fromnon-nodulated soybean roots grown in nitrate mediacontained averages of 1–8, 20–50 and 50–80% of totalN as ureide-N, amino acid-N and nitrate-N, respectively,with varying cultivars, and Asn was the most pre-dominant among the amino acids (Hunter et al. 1982;McClure and Israel 1979). In contrast, Asn and, to alesser extent, Gln are the predominant nitrogen com-pounds of the xylem exudates of temperate legumes,such as Pisum, Lupinus, Vicia, Trifolium, Medicago andLotus, regardless of nodulation (Pate 1980; Schubert1986). These findings suggest that carbon skeletons mightbe replenished for the synthesis of C4 amino acids(aspartate [Asp] and Asn) rather than C5 amino acids(glutamate [Glu] and Gln) in leguminous plants duringroot nitrogen assimilation. In the present study, we testedthe hypothesis that carbon skeletons are provided forthe synthesis of asparagine as a predominant amideamong amino acids in roots of legumes in response tonitrogen supply.
MATERIALS AND METHODS
Plant cultureSoybean (Glycine max L. Merill cv. Fukuyutaka) and pea(Pisum sativum L. cv. Hakuryuu) plants were grownin the same procedures as described previously (Ikedaet al. 2004). Soybean seedlings with fully expandedprimary leaves and pea seedlings with the first leaf weregrown in hydroponic culture in the phytotron of theBiotron Institute, Kyushu University (at 25°C for soybeanand 20°C for pea at 70% relative humidity under naturallight conditions). Quarter-strength modified Hoaglandnutrient solution (pH 6.5) containing 4 mmol L–1
NaNO3 as the nitrogen source was used as the culturesolution (Downs and Hellmers 1975; Ikeda et al. 1992).The nutrient solution was replaced every other day. Theseplants were used for the following two experiments,which were conducted in triplicate.
Determination of free amino acids in soybean and pea roots
Fourteen-day-old soybean and 22-day-old pea plantswere grown in nitrogen-free quarter-strength modifiedHoagland nutrient solution for 3 days, and then sub-jected to nitrogen treatment for 1 day. The nitrogentreatments included 2 mmol L–1 (NH4)2SO4 (+A),4 mmol L–1 NaNO3 (+N) and no nitrogen (−N) as thenitrogen source. After nitrogen treatment, the plantswere harvested and rinsed several times with deionizedwater, and then separated into roots and shoots. Theplant parts were frozen at –80°C and lyophilized. Freeamino acids were extracted and purified from dried
roots as previously described (Ikeda et al. 2004) andliquid-chromatographic determination of the aminoacids was carried out according to the method of Yamayaand Matsumoto (1988).
Feeding of 14C-glucose to soybean and pea roots
Fourteen-day-old soybean and 22-day-old pea plantswere grown in nitrogen-free quarter-strength modifiedHoagland nutrient solution for 3 days, and then sub-jected for 1 day to the nitrogen treatment describedabove. After treatment, each plant was acclimated for1 h at 25°C in the dark in respective nutrient solutionsin a growth chamber. These plants were placed in asealed bottle with the respective nutrient solution con-taining 0.1 mmol L–1 14C-glucose (400 kBq, D-[U-14C]glucose; Amersham Biosciences, Little Chalfont, UK)for 3 h with gentle shaking at 25°C in the dark in thegrowth chamber. A filter-paper segment moistened with10% (w/v) NaOH was placed beside the base of theroots in the bottle to recover 14CO2 released from theroots. After feeding of 14C-glucose, the roots of harvestedplants were rinsed several times with deionized water,and then separated into roots and shoots. These plantparts were frozen at –80°C and lyophilized. The filter-paper segment was transferred in a vial and dried at60°C. The radioactivity in this segment was regardedas the CO2 fraction.
Analysis of metabolites and amino acids derived from 14C-glucose in roots
Freeze-dried roots were extracted three times with80% (v/v) ethanol at 80°C. The remaining roots afterextraction were placed in a vial and freeze-dried. Theradioactivity in this sample was regarded as the insolublefraction. Purification and thin-layer chromatographicanalyses of labeled amino acids in the extracts werecarried out according to the methods of Ikeda et al.(2004).
All data were subject to statistical analysis usingDuncan’s multiple range test.
RESULTS
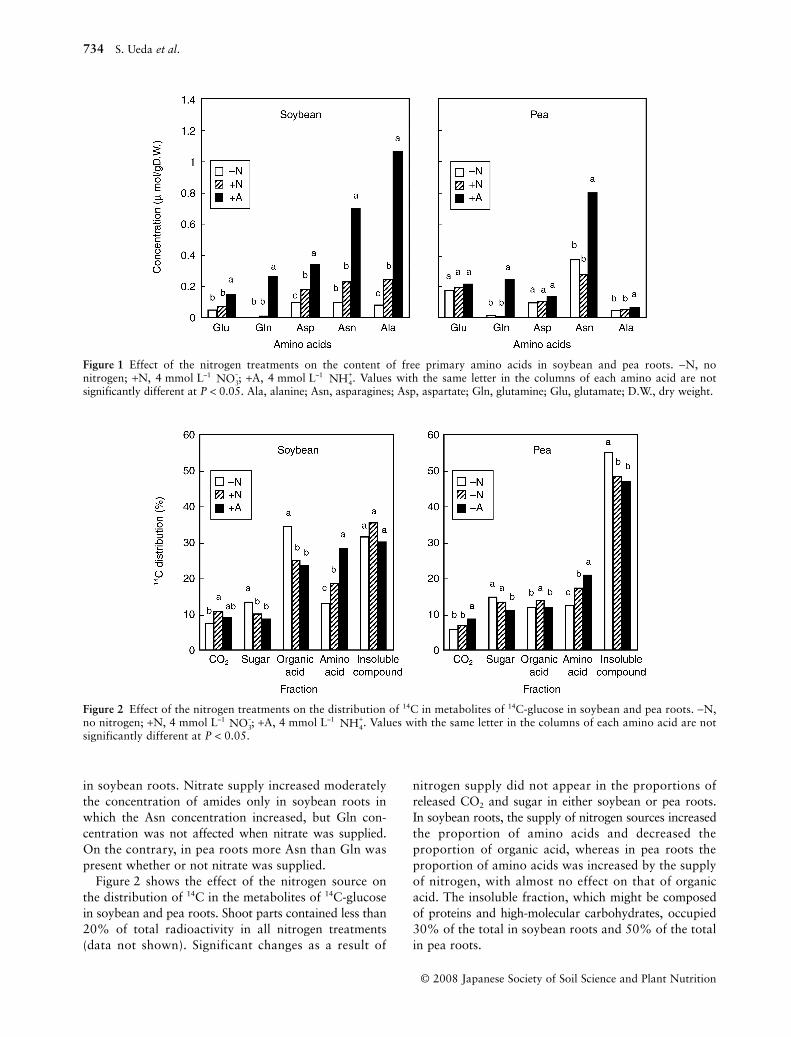
The effect of the nitrogen source on the concentrationsof the primary amino acids that are involved in theprimary ammonium assimilation (Glu, Gln, Asp andAsn) and Alanine (Ala) in the soybean and pea roots isshown in Fig. 1. In both soybean and pea roots, theammonium supply increased markedly the concentra-tions of the primary amino acids. The concentration ofAsn was twofold higher than that of Gln in soybeanand pea roots, although the concentration of Ala was high
734 S. Ueda et al.
© 2008 Japanese Society of Soil Science and Plant Nutrition
in soybean roots. Nitrate supply increased moderatelythe concentration of amides only in soybean roots inwhich the Asn concentration increased, but Gln con-centration was not affected when nitrate was supplied.On the contrary, in pea roots more Asn than Gln waspresent whether or not nitrate was supplied.
Figure 2 shows the effect of the nitrogen source onthe distribution of 14C in the metabolites of 14C-glucosein soybean and pea roots. Shoot parts contained less than20% of total radioactivity in all nitrogen treatments(data not shown). Significant changes as a result of
nitrogen supply did not appear in the proportions ofreleased CO2 and sugar in either soybean or pea roots.In soybean roots, the supply of nitrogen sources increasedthe proportion of amino acids and decreased theproportion of organic acid, whereas in pea roots theproportion of amino acids was increased by the supplyof nitrogen, with almost no effect on that of organicacid. The insoluble fraction, which might be composedof proteins and high-molecular carbohydrates, occupied30% of the total in soybean roots and 50% of the totalin pea roots.
Figure 1 Effect of the nitrogen treatments on the content of free primary amino acids in soybean and pea roots. −N, nonitrogen; +N, 4 mmol L–1 ; +A, 4 mmol L–1 . Values with the same letter in the columns of each amino acid are notsignificantly different at P < 0.05. Ala, alanine; Asn, asparagines; Asp, aspartate; Gln, glutamine; Glu, glutamate; D.W., dry weight.
NO3− NH4
+
Figure 2 Effect of the nitrogen treatments on the distribution of 14C in metabolites of 14C-glucose in soybean and pea roots. −N,no nitrogen; +N, 4 mmol L–1 ; +A, 4 mmol L–1 . Values with the same letter in the columns of each amino acid are notsignificantly different at P < 0.05.
NO3− NH4
+

Carbon skeletons of amides in legume roots 735
© 2008 Japanese Society of Soil Science and Plant Nutrition
The effect of nitrogen supply on the composition ofprimary amino acids formed from 14C-glucose in soybeanand pea roots is shows in Fig. 3. In both soybean andpea roots grown in ammonium media, the primaryamino acids accounted for approximately 90% in theamino acid fraction, and Asn accounted for the highestproportion of labeled soluble amino acids (35 and 30%for soybean and pea roots, respectively). Glutaminealso increased, but not as much as Asn. In nitratenutrition, the primary amino acids accounted forapproximately 60% in the amino acid fraction in bothsoybean and pea roots. Glutamine did not increase ineither soybean or pea roots grown in nitrate media.Asparagine was the most abundant amino acid andaccounted for near 20% in soybean roots, whereas thesupply of nitrate had little effect on the proportion ofeither amide derived from glucose in pea roots. BecauseAsn is derived from Asp and Gln is derived from Glu,the ratio of C4 amino acids to C5 amino acids in thesoluble amino acids derived from 14C-glucose roots was
calculated (Table 1). The ratio of nitrogen-suppliedsoybean roots was twofold higher than that in roots withno supplied nitrogen. In pea roots, the ratio increasedonly with ammonium nutrition.
DISCUSSION
The glucose feeding experiment in soybean and pearoots was carried out under the same conditions ofnitrogen nutrition and light as that used in other plantroots in previous work (Ikeda et al. 2004, 2005; Kogaand Ikeda 2000) because growth conditions affect Glnand Asn biosynthesis even in roots (McGrath and Coruzzi1991). Although the growth temperatures were differentbetween the soybean and pea plants, the present resultsseem to support our hypothesis that Asn biosynthesisrather than Gln biosynthesis is predominant in leguminousplant roots that receive nitrogen. Unexpectedly, however,the pea plants responded weakly to nitrogen supplycompared with the soybean plants.
Analyses of free amino acids in roots with highperformance liquid chromatography showed that ammo-nium supply increased the concentrations of Asn andGln considerably in both soybean and pea roots (Fig. 1).However, the concentration of Asn was much higherthan that of Gln. When nitrate was supplied, theconcentration of Asn increased in soybean roots, butchanged very little in pea roots (Fig. 1). The supply ofnitrate has no effect on the concentration of Gln in soy-bean and pea roots. Analyses of free amino acids inxylem sap and roots have been repeatedly carried out
Figure 3 Effect of the nitrogen treatments on the composition of the primary amino acids derived from 14C-glucose in soybeanand pea roots. −N, no nitrogen; +N, 4 mmol L–1 ; +A, 4 mmol L–1 . Values with the same letter in the columns of eachamino acid are not significantly different at P < 0.05. Ala, alanine; Asn, asparagines; Asp, aspartate; Gln, glutamine; Glu,glutamate.
NO3− NH4
+
Table 1 Ratios of C4 amino acids to C5 amino acids derivedfrom 14C-glucose in soybean and pea roots
Nitrogen source Soybean Pea
4 mmol L–1 2.03 a 1.47 a4 mmol L–1 2.25 a 0.73 bNo nitrogen 1.16 b 0.72 b
Values with the same letter are not significantly different at P < 0.05 within each plant.
NH4+
NO3−

736 S. Ueda et al.
© 2008 Japanese Society of Soil Science and Plant Nutrition
in non-nodulated soybean and pea roots (Chaillou et al.1991; Frechilla et al. 2002; Lasa et al. 2002; McClureand Israel 1979; Oaks 1992). In these studies, theproportion of Asn is generally larger than the proportionof Gln in nitrogen-supplied legume plants. Our resultssupport these previous studies, although nitrate effectswere not notable in the present experiment. In non-nodulated alfalfa roots, Asn increased markedly com-pared with Gln in ammonium nutrition (Pasqualiniet al. 2001). Hence, the predominant amino acid inroots grown in ammonium media seems to be Asnin leguminous plants, which is different from non-leguminous dicots (Bialczyk et al. 2004; Ikeda et al.2004, 2005).
The continuous synthesis of amides such as Asn andGln requires the provision of carbon skeletons oxaloac-etate and 2-oxoglutarate that are produced in the rootsfrom sugars transported from the shoots (Corzzi andLast 2000). Although a major sugar in photosynthatetranslocation is sucrose, glucose is produced by thehydrolysis of sucrose in the roots. Hence, 14C-glucoseas a source of carbon skeletons was fed to soybean andpea roots. The cycling of amino acids from shoots toroots plays an important role in the composition ofamino acids in the roots (Cooper and Clarkson 1989).It was, however, considered that the transported 14Cfrom the shoots to the roots had little influence on thedistribution of 14C in the roots because shoot partscontained less than 20% of the total radioactivity inall the nitrogen treatments in the present experiment(data not shown). In soybean and pea roots, nitrogensupply increased the proportion of the amino acidsfraction (Fig. 2). In fact, the ratio of amino acids toorganic acids was increased by nitrogen supply (fromFig. 2). The present results indicate that the supply ofnitrogen enhances the utilization of 14C-glucose as thesource of carbon skeletons for the synthesis of aminoacids in soybean and pea roots. At the present time, noconsistent difference in the release of CO2 derivedfrom glucose fed to roots can be explained in relationto the synthesis of amino acids because CO2 is producedin the oxidative degradation of glucose and at the sametime the produced CO2 is fixed as bicarbonate in darkcarbon fixation in the primary amino acid synthesisin roots.
In the amino acid fraction, Ala did not occupy thehighest proportion, although the carbon skeleton of Alais pyruvate, which is one of the first oxo-acids producedfrom glucose (Fig. 3). Ammonium supply markedlyincreased the proportion of Asn and Gln in soybeanand pea, and Asn was the most abundant amino acid.Moreover, the ratios of C4 amino acids to C5 amino acidsincreased twofold in ammonium-grown roots comparedwith roots grown in nitrogen-free media (Table 1).
The results suggest preferential use of the C4 skeleton forthe synthesis of Asn in soybean and pea roots grown inammonium media. In cucumber and tomato roots, theproportion of 14C-Gln was greater than that of 14C-Asnand the value of C4/C5 amino acids was smaller thanunity (Ikeda et al. 2004, 2005), whereas in rice andwheat roots, the proportion of 14C-Asn was greaterthan that of 14C-Gln and the value of C4/C5 aminoacids was larger than unity (Ikeda et al. 2004; Kogaand Ikeda 2000). Hence, leguminous plants are similarto monocots rather than non-leguminous dicots withrespect to amide synthesis in roots in response toammonium supply.
The supply of nitrate showed different trends in thecomposition of major amino acids derived from glucosebetween soybean and pea roots (Fig. 3). The ratio ofC4/C5 amino acids was twofold that of control plantsonly in soybean roots, however, the proportion of Asnin pea roots was also much greater than that of Gln,similar to the proportion in soybean roots. Nitrateneeds to be reduced to ammonia before its assimilationin roots (Oaks and Hirel 1985). It is understandablethat amide synthesis and the provision of carbon skele-tons for nitrate assimilation in roots might be regulatedby the intensity of the ammonium supply in the rootcells because of direct uptake or nitrate reduction.Although nitrate reduction in tropical legumes, suchas soybean, occurs mostly in shoots and in temperatelegumes, such as pea, it occurs predominantly in theroots (Andrews 1986), the present results showed thatthe nitrate effects after 1 day were more evident onsoybean roots than on pea roots. When pea plants aretreated with nitrate for longer periods or at higherconcentrations, the nitrate effect might be more evident.Asparagine synthesis is enabled by both the stimulationof dark carbon fixation (Ikeda et al. 1992) and theexpression of asparagines synthetase (Kawachi et al.2002) when ammonium concentrations are high in theroots of leguminous plants.
In conclusion, the results suggest that the roots ofleguminous plants might possess an indigenous abilityto provide a carbon skeleton for preferential synthesisof Asn rather than Gln in root cells with a high intensityof ammonium supply.
REFERENCES
Andrews M 1986: The partitioning of nitrate assimilationbetween root and shoot of higher plants. Plant CellEnviron., 9, 511–519.
Atkins CA, Pate JS, Griffiths GJ, White ST 1980: Economyof carbon and nitrogen in nodulated and non-nodulated(NO3-grown) cowpea (Vigna unguiculata L. Walp). PlantPhysiol., 66, 978–983.

Carbon skeletons of amides in legume roots 737
© 2008 Japanese Society of Soil Science and Plant Nutrition
Bialczyk J, Lechowski Z, Dziga D 2004: Composition of thexylem sap of tomato seedlings cultivated on media with
and nitrogen source as or . Plant Soil,263, 265–272.
Chaillou S, Vessey JK, Morot-Gaudry JF, Raper CD, HenryLT, Boutin JP 1991: Expression of characteristics ofammonium nutrition as affected by pH of the rootmedium. J. Exp. Bot., 42, 189–196.
Cooper HD, Clarkson DT 1989: Cycling of amino-nitrogenand other nutrients between shoots and roots in cereals– a possible mechanism integrating shoot and root inthe regulation of nutrient uptake. J. Exp Bot., 40, 753–762.
Corzzi G, Last R 2000: Assimilation of inorganic nitrogeninto N-transport amino acids. In Biochemistry andMolecular Biology of Plants. Ed. BB Buchanan, WCruissem and RL Jones, pp. 362–379, American Societyof Plant Physiologists, Rockville.
Downs RJ, Hellmers H, eds 1975: Water and nutrition. InEnvironments and the Experimental Control of PlantGrowth. pp. 95–116, Academic Press, London.
Frechilla S, Lasa B, Aleu M, Juanarena N, Lamsfus C, Aparicio-Tejo PM 2002: Short-time ammonium supply stimulatesglutamate dehydrogenase activity and alternative pathwayrespiration in roots of pea plants. J. Plant Physiol., 159,811–818.
Givan CV 1979: Metabolic detoxification of ammonia intissues of higher plants. Phytochemistry, 18, 375–382.
Hunter WJ, Fahring CJ, Olsen SR, Porter LK 1982: Locationof nitrate reduction in different soybean cultivars. CropSci., 22, 944–948.
Ikeda M, Kusano T, Koga N 2004: Carbon skeletons foramide synthesis during ammonium nutrition in tomatoand wheat roots. Soil Sci. Plant Nutr., 50, 141–147.
Ikeda M, Mizoguchi K, Yamakawa T 1992: Stimulation ofdark carbon fixation in rice and tomato roots byapplication of ammonium nitrogen. Soil Sci. PlantNutr., 38, 315–322.
Ikeda M, Ueda S, Yamakawa T 2005: Preferential provision ofcarbon skeletons for amide synthesis during ammonium
nutrition in the roots of various plants. In Plant Nutritionfor Food Security, Human Health and EnvironmentalProtection. Ed. CJ Li et al., pp. 190–191, TsinghuaUniversity Press, Beijing.
Kawachi T, Sueyoshi K, Nakajima A, Yamagata H, SugimotoT, Oji Y 2002: Expression of asparagine synthetase inrice (Oryza sativa) roots in response to nitrogen. Physiol.Plant., 114, 41–46.
Koga N, Ikeda M 2000: Enhancement of glucose utilizationin provision of carbon skeletons for ammonium assimila-tion in wheat roots. J. Fac. Agr. Kyushu Univ., 45, 7–13.
Lasa B, Frechilla S, Aparicio-Tejo PM, Lamsfus C 2002: Roleof glutamate dehydrogenase and phosophoenolpyruvatecarboxylase activity in ammonium nutrition tolerance inroots. Plant Physiol. Biochem., 40, 969–976.
McClure PR, Israel DW 1979: Transport of nitrogen in thexylem of soybean plants. Plant Physiol., 64, 411–416.
McGrath RB, Coruzzi GM 1991: A gene network controllingglutamine and asparagines biosynthesis in plants. PlantJ., 1, 275–280.
Oaks A 1992: A re-evaluation of nitrogen assimilation inroots. Bioscience, 42, 105–111.
Oaks A, Hirel B 1985: Nitrogen metabolism in roots. Ann.Rev. Plant Physiol., 36, 345–365.
Ohyama T, Kato N, Saito K 1989: Nitrogen transport inxylem of soybean plant supplied with . Soil. Sci.Plant Nutr., 35, 131–137.
Pasqualini S, Ederli L, Piccioni C et al. 2001: Metabolic regu-lation and gene expression of root phosphoenolpyruvatecarboxylase by different nitrogen sources. Plant CellEnviron., 24, 439–447.
Pate JS 1980: Transport and partitioning of nitrogenous solutes.Ann. Rev. Plant Physiol., 31, 313–340.
Schubert KR 1986: Product of biological nitrogen fixationin higher plants: synthesis, transport, and metabolism.Ann. Rev. Plant Physiol., 37, 539–574.
Yamaya T, Matsumoto H 1988: Analysis of phenilisothiocar-bamyl-amino acids at pico-mole level by high performanceliquid chromatography and application to plant materials.Soil Sci. Plant Nutr., 34, 297–302.
HCO3− NO3
− NH4+
15NO3−