
Protivgeni i protivtela Molekulite koi go obele`uvaat steknatiot imun ...
33
Protivgeni i protivtela Molekulite koi go obele`uvaat steknatiot imun sistem se protivteloto i T-kleto~niot receptor. Dodeka delovite od vrodeniot imunitet se programirani za prepoznavawe obrasci i na toj na~in prepoznavaat osobini spodeleni od grupi na nadvore{ni molekuli, protivteloto i T-kleto~niot receptor prika`ivaat povisok stepen na specifi~nost, prepoznavaj}i odredeni protivgeni odrednici (determinanti) ili epitopi. Epitopite se imuni aktivni podra~ja od imunogenot koi se vrzuvaat za protivgen specifi~ni membranski receptori vo limfocitite ili izla~enite protivtela. Protivtelata se epitop vrzuva~ki proteini koi se sretnuvaat vo dve formi, ili kako membranski vrzani soedinenija vrz B kletkite ili kako rastvorlivi molekuli izla~eni od plazma kletkite. Membranski vrzanite protivtela poseduvaat protivgena specifi~nost za B kletkite. Razmno`uvaweto na protivgen-specifi~nite B kleto~ni klonovi e pottiknato od me|udejstvoto na membranskoto protivtelo so protivgenot. Izla~enite protivtela cirkuliraat vo krvta, kade slu`at kako efektori na humoralniot imunitet so prebaruvawe i ozna~uvawe protivgenite za otstranuvawe. Site protivtela delat strukturni osobini, se vrzuvaat za protivgen i u~estvuvaat vo ograni~en broj efektorni funkcii. T-kleto~niot receptor koj se prika`uva vrz povr{inata od membranata na T kletkata prepoznava samo preraboteni protivgeni fragmenti koi prika`uva vrz povr{inata od membranata na T kletkata prepoznava samo preraboteni protivgeni fragmenti koi se spoeni so GTK molekuli. Iako ova poglavje gi naglasuva protivteloto i prirodata na B-kleto~noto prepoznavawe protivgen, }e se napravi sporedba so T-kleto~noto prepoznavawe protivgen za da se razvie op{tata tema na protivgenost i da se prika`e kontrastot pome|u T i B kleto~noto me|udejstvo so protivgenite. Molekulskite osobini na GTK molekulite, na~inot na koj protivgenite se obrabotuvaat i vrzuvaat so GTK molekulite i prirodata na T-kleto~nite receptori koi prepoznavaat kompleksi protivgen-GTK }e se pokrie vo poglavja 8 i 9. Vo osnova, populacijata na protivtelata koi se sozdavaat kako odgovor na odredena protivgena drazba e heterogena. Pove}eto protivgeni se strukturni kompleksi, koi sodr`at mnogu razli~ni epitopi, a imuniot sistem obi~no reagira so sozdavawe protivtela na del od epitopite vo protivgenot. So drugi zborovi, nekolku razli~ni B-kleto~ni klonovi se stimuluraat i se razmno`uvaat. Rezultatot od plazma kletkite vo eden B- kleto~en klon e monoklonsko protivtelo koe specifi~no se vrzuva za edine~na protivgena odrednica. Izla~enite produkti od site stimulirani B-kleto~ni klonovi i grupata na monoklonskite protivtela gi so~inuvaat poliklonskite i heterogenite serumski protivtela kako odgovor na imunizira~kiot protivgen. ^lenovi od proteinskata familija protivtela imaat zaedni~ki strukturni osobini i op{to se poznati kako imunoglobulini (Ig). No i pokraj nivnite sli~nosti, ~lenovite od ovaa familija izvr{uvaat izvonredno razli~ni funkcii pri vrzuvawe isto kako i nekolku razli~ni efektorski funkcii koi sledat po vrzuvaweto na protivgenot. Pred da ja istra`ime strukturata i kompleksnosta na familijata protivtelata, ova poglavje }e ja opi{e prirodata na protivgenite i konceptite i osobinite na imunogenite supstancii. Protivgenite se definirani specifi~no kako molekulite koi stapuvaat vo me|udejstvo so imunoglobulinskiot receptor od B kletkite (ili so T-kleto~niot receptor koga e vo kompleks so GTK). Molekulskite svojstva na protivgenite i kako ovie svojstva pridonesuvaat za aktivirawe imunitetot se centralni za na{eto razbirawe na imuniot sistem. Ova poglavje gi opi{uva molekulskite osobini na protivgenite prepoznaeni so protivtelata i go istra`uva pridonesot napraven za imunogenosta od strana na biolo{kiot sistem na doma}inot. 1
Transcript of Protivgeni i protivtela Molekulite koi go obele`uvaat steknatiot imun ...

Protivgeni i protivtela
Molekulite koi go obele`uvaat steknatiot imun sistem se protivteloto i T-kleto~niot receptor. Dodekadelovite od vrodeniot imunitet se programirani za prepoznavawe obrasci i na toj na~in prepoznavaat osobinispodeleni od grupi na nadvore{ni molekuli, protivteloto i T-kleto~niot receptor prika`ivaat povisok stepenna specifi~nost, prepoznavaj}i odredeni protivgeni odrednici (determinanti) ili epitopi. Epitopite se imuniaktivni podra~ja od imunogenot koi se vrzuvaat za protivgen specifi~ni membranski receptori vo limfocititeili izla~enite protivtela. Protivtelata se epitop vrzuva~ki proteini koi se sretnuvaat vo dve formi, ili kakomembranski vrzani soedinenija vrz B kletkite ili kako rastvorlivi molekuli izla~eni od plazma kletkite.Membranski vrzanite protivtela poseduvaat protivgena specifi~nost za B kletkite. Razmno`uvaweto naprotivgen-specifi~nite B kleto~ni klonovi e pottiknato od me|udejstvoto na membranskoto protivtelo soprotivgenot. Izla~enite protivtela cirkuliraat vo krvta, kade slu`at kako efektori na humoralniot imunitetso prebaruvawe i ozna~uvawe protivgenite za otstranuvawe. Site protivtela delat strukturni osobini, sevrzuvaat za protivgen i u~estvuvaat vo ograni~en broj efektorni funkcii. T-kleto~niot receptor koj seprika`uva vrz povr{inata od membranata na T kletkata prepoznava samo preraboteni protivgeni fragmenti koiprika`uva vrz povr{inata od membranata na T kletkata prepoznava samo preraboteni protivgeni fragmenti koise spoeni so GTK molekuli. Iako ova poglavje gi naglasuva protivteloto i prirodata na B-kleto~notoprepoznavawe protivgen, }e se napravi sporedba so T-kleto~noto prepoznavawe protivgen za da se razvie op{tatatema na protivgenost i da se prika`e kontrastot pome|u T i B kleto~noto me|udejstvo so protivgenite.Molekulskite osobini na GTK molekulite, na~inot na koj protivgenite se obrabotuvaat i vrzuvaat so GTKmolekulite i prirodata na T-kleto~nite receptori koi prepoznavaat kompleksi protivgen-GTK }e se pokrie vopoglavja 8 i 9.Vo osnova, populacijata na protivtelata koi se sozdavaat kako odgovor na odredena protivgena drazba eheterogena. Pove}eto protivgeni se strukturni kompleksi, koi sodr`at mnogu razli~ni epitopi, a imuniotsistem obi~no reagira so sozdavawe protivtela na del od epitopite vo protivgenot. So drugi zborovi, nekolkurazli~ni B-kleto~ni klonovi se stimuluraat i se razmno`uvaat. Rezultatot od plazma kletkite vo eden B-kleto~en klon e monoklonsko protivtelo koe specifi~no se vrzuva za edine~na protivgena odrednica.Izla~enite produkti od site stimulirani B-kleto~ni klonovi i grupata na monoklonskite protivtela giso~inuvaat poliklonskite i heterogenite serumski protivtela kako odgovor na imunizira~kiot protivgen.
^lenovi od proteinskata familija protivtela imaat zaedni~ki strukturni osobini i op{to se poznati kakoimunoglobulini (Ig). No i pokraj nivnite sli~nosti, ~lenovite od ovaa familija izvr{uvaat izvonrednorazli~ni funkcii pri vrzuvawe isto kako i nekolku razli~ni efektorski funkcii koi sledat po vrzuvaweto naprotivgenot. Pred da ja istra`ime strukturata i kompleksnosta na familijata protivtelata, ova poglavje }e jaopi{e prirodata na protivgenite i konceptite i osobinite na imunogenite supstancii.Protivgenite se definirani specifi~no kako molekulite koi stapuvaat vo me|udejstvo so imunoglobulinskiotreceptor od B kletkite (ili so T-kleto~niot receptor koga e vo kompleks so GTK). Molekulskite svojstva naprotivgenite i kako ovie svojstva pridonesuvaat za aktivirawe imunitetot se centralni za na{eto razbirawe naimuniot sistem. Ova poglavje gi opi{uva molekulskite osobini na protivgenite prepoznaeni so protivtelata igo istra`uva pridonesot napraven za imunogenosta od strana na biolo{kiot sistem na doma}inot.
1

Imunogenost nasproti protivgenost
Imunogenosta i protivgenosta se povrzani so razli~ni imuni osobini koiponekoga{ se me{aat. Imunogenosta e sposobnosta da se pottikne humoraleni/ili kleto~no posreduvan imun odgovor:
Iako substancijata koja pottiknuva specifi~en imun odgovor obi~no seu c j j u c f u d rnarekuva protivgen, posodvetno e da se narekuva imunogen.
2

Protivgenosta e sposobnosta za da se kombinira specifi~nosta so krajniteprodukti od gornite odgovori (t.e. izla~enoto protivtelo i/ilipovr{inskite receptori vo T kletkite). Iako site molekuli koi imaatsvojstvoto na imunogenost isto taka imaat i svojstvoto na protivgenost, nemo`e obratno. Odredeni mali molekuli, nare~eni hapteni, se protivgeni none se sposobni samite da pottiknuvaat specifi~en imun odgovor. So drugizborovi, nemaat imunogenost.
3

Haptenite se korisni istra`uva~ki i dijagnosti~ki alatki
Hemiskoto vrzuvawe na heptenot so golem imunogen protein, nare~en nosa~,dava imunogen hapten-nosa~ kowugat. @ivotnite imunizirani so takovkowugat sozdavaat protivtela specifi~ni za tri vida protivgeniodrednici: (1) haptenska odrednica, (2) nepromeneti epitopi od nose~kiotprotein i (3) novi epitopi sozdadeni so kombinacija od podra~ja zaedni~kiza molekulata hapten i molekulata nosa~ (slika 4 1) Samiot hapten ne mo`eza molekulata hapten i molekulata nosa~ (slika 4-1). Samiot hapten ne mo`eda funkcionira kako imunogen epitop. No koga pove}e molekuli od edenhapten se kombiniraat so proteinski nosa~ (ili duri i vo neimunogenhomopolimer), haptenot stanuva pristapliv za imuniot sistem i mo`e dafunkcionira kako imunogen.
(slika 4-1 Na ovaa slika kowugatot hapten-nosa~ e imunogen, a hapten e( u u ,protivgen koj samiot od sebe ne e imunogen. Imunogenot sodr`i pove}e kopiod hapten - mal neimunogeno organsko soedinenie, vo ovoj slu~aj,dinitrofenol (DNF) - hemiski povrzan za golem proteinski nosa~ kako {toe govedski serum albumin (GSA). Imunizacijata so DNF-GSA sozdava trivida protivtela. Od niv, protivteloto protiv-DNF e dominantno,uka`uvaj}i deka vo ovoj slu~aj haptenot e imunodominanten epitop iliprotivgena odrednica, kako {to e ~esto pri takvite kowugati.r dr d c , r u
4

Pokraj {irokiot opseg na epitopi koi se sretnuvaat kaj eden imunogen, sistemot hapten-nosa~ naimunolozite im ozozmo`uva alatka za istra`uvawe efektite od malite varijaci vo hemiskatau u r u f d r j cstruktura vrz imunolo{kata specifi~nost. Haptenot mo`e da bide hemiski definirana odrednicakoja potoa diskretno se modificira so hemiski sredstva za da se odredi dali ova }e vlijae vrzprepoznavaweto od protivtelata. Klasi~en prikaz na ovaa strategija se sretnuva vo pionerskatarabota na Karl Land{tajner, koj vo 1920-tite i 1930-tite godini sozdade ednostaven, hemiskidefiniran sistem za prou~uvawe povrzuvaweto na poeidine~no protivtelo. Land{tajner rabotel sohapteni so mali organski molekuli koi se protivgeni no ne se imunogeni. Vo svoite prou~uvawa,Land{tajner imuniziral zajaci so hapten-nosa~ kowugat a potoa ja testiral reaktivnosta na zaja~kiotimun serum so istiot hapten i so tesno povrzani hapteni vrzani za razli~en proteinski nosa~. Na tojna~in toj mo`el precizno da go izmeri odgovorot na protivhaptenskite protivtela vo imuniot serumno ne i toj od protivtelata od izvornite epitopi od nosa~ot. Land{tajner napravil testovi za toadali protivhaptenskoto protivtelo mo`e da se vrze za drugi hapteni so nezna~itelno razli~nihemiski strukturi Ako dojde{e do vrzuvawe toa be{e nare~eno -vkrstena reakcija Prekuhemiski strukturi. Ako dojde{e do vrzuvawe, toa be{e nare~eno -vkrstena reakcija. Prekuistra`uvawe koi modifikacii od haptenot spre~ile ili dozvolile vkrsteni-reakcii, Land{tajnermo`el datalno da ja razgleda specifi~nosta vo me|udejstvota na protivgenot so protivteloto.So upotreba na razli~ni derivati od aminobenzen kako hapteni, Land{tajner otkril deka celiotizled na haptenot igra golema uloga pri odreduvaweto dali toj mo`e da reagira so dadeno protivtelo.Na primer, protivserumot od zajacite imunizirani so aminobenzen ili eden od negovite karboksilniderivati (orto-aminobenzoi~na kiselina, meta-aminobenzoi~na kiselina ili para-aminobenzoi~nakiselina) vo kombinacija so protein nosa~ reagiral samo so izvorniot imunizira~ki hapten a nereagiral vkrsteno so koj bilo od drugite hapteni (tabela 4-1). Nasproti toa, ako celiot izgled nahaptenot se zadr`i ist a haptenot se modificira vo para pozicija so razli~ni nejonski derivati,toga{ protivserumot poka`uval razli~ni stepeni na vkrstena reakcijaa. Rabotata na Land{tajner japrika`a i specifi~nosta na imuniot sistem za malite strukturni varijacii vo haptenite ir c f u ru ur r j cogromnata raznovidnost na epitopite koi imuniot sistem mo`e da gi prepoznae.Mnogu biolo{ko zna~ajni supstancii, vklu~uvaj}i gi lekovite, peptidnite hormoni i steroidnitehormoni mo`at da funkcioniraat kako hapteni. Kowugatite od ovie hapteni so golemi proteinskinosa~i mo`e da se koristi za sozdavawe hapten-specifi~ni protivtela. Ovie protivtela se korisniza merewe prisustvoto na razli~ni supstancii vo teloto. Na primer, obi~nata alatka za doma{noispituvawe bremenost osloboduva protivhaptenski protivtela za odreduvawe dali urinata na `enatasodr`i human horionski gonadotropin (HHG), {to pretstavuva znak za bremenost.Iako prou~uvawata so hapten-nosa~ kowugati davaat jasno razgrani~uvawe pome|u konceptite naprotivgenosta i imunogenosta, vo realnosta, za odreduvawe dali substancijata so koja se sretnalimuniot sistem }e predizvika odgovor treba da se razgledaat pove}e faktori. Imunogenosta ne zavisisamo od vnatre{nite osobini na protivgenot tuku isto taka i od odreden broj osobini od konkretniotsamo od vnatre{nite osobini na protivgenot tuku isto taka i od odreden broj osobini od konkretniotbiolo{ki sistem so koj se sretnal protivgenot i na~inot na koj e pretstaven imunogenot. Slednivedelovi gi objasnuvaat osobinite koi gi delat pove}eto imunogeni i pridonesot na biolo{kiot sistemkoj sozdava prika`uvawe imunogenost.
5

Osobinite na imunogenot pridonesuvaat za imunogenosta
b |Imunogenosta delumno se utvrduva preku ~etiri osobini na imunogenot: negovata tu|ost, molekulskata golemina, hemiskiot sostav i slo`enosta isposobnosta za obrabotka i prika`uvawe so GTK molekulata vrz povr{inata od protivgen prika`uva~kata kletka ili o{tetenata sopstvena kletka.
Tu|ostZa pottiknuvawe imun odgovor, molekulata mora da bide prepoznaena kako tu|a od strana na biolo{kiot sistem. Nasproti kapacitetot za prepoznavawetu|o se nao|a podnesuvaweto na svoe, specifi~na podnoslivost (tolerancija) na svoi protivgeni (vidi poglavje 16). Pogolemiot del od sposobnosta zapodnesuvawe svoi protivgeni proizleguva pri razvojot na limfocitot, koga nezrelite limfociti se izlo`eni na svoite delovi. Kletkite koi pri ovojproces prepoznavaat svoi delovi se neaktivni. Tie koi }e go pre`iveat procesot se oslobodeni. Protivgenite koi ne bile prika`ani vrz nezrelitelimfociti vo ovoj kriti~en period podocna od strana na imuniot sistem mo`e da bidat prepoznati kako tu|i. Koga protivgenot ke se vnese voorganizam, stepenot na negovata imunogenost zavisi od stepenot na negovata tu|ost. Vo osnova, kolku e pogolemo filogenetskoto rastojanie pome|u dvavida, tolku e pogolema strukturnata (a so toa i protivgenata) razli~nost pome|u nivnite strukturni molekuli. Na primer, voobi~aenioteksperimetalen protivgen govedski serum albumin (GSA) ne e imunogen koga se vnesuva vo kravata no e mo{ne imunogen koga }e se vnese vo zajak. Osventoa, se o~ekuva deka GSA }e izvr{uva pogolema imunogenost kaj koko{kata otkolku kaj kozata, koja e potesno povrzana so govedata. Postojat isklu~ociod ova pravilo. Odredeni makromolekuli (na primer, kolagenot i citohromot c) mo{ne se zadr`ale pri evoluacijata i zatoa prika`uvaat mnogu malkuimunogenost niz lozi od razli~ni vidovi. Nasproti toa, odredeni vnatre{ni delovi (na pr. tkivoto od ro`nicata i spermata) efikasno se izoliraat odimuniot sistem, taka {to ako ovie tkiva se vnesat duri i vo `ivotnoto od koe poteknuvaat, }e funkcioniraat kako imunogeni.
Molekulska goleminaPostoi povrzanost pome|u goleminata na makromolekulata i negovata imunogenost. Najaktivnite imunogeni te`neat kon molekulska masa od > 1000,000daltoni (Da). Vo osnova, supstanciite so molekulska masa pomala od 5000 do 10000 Da se slabi imunogeni, iako za odredeni supstancii so molekulskamasa pomala od 1000 Da e doka`ano deka se imunogeni.
Hemiski sostav i heterogenostGoleminata i nadvore{nosta ne se dovolni, sami po sebe, za molekulata da ja napravat imunogeni~na. Isto taka potrebni se i drugi osobini. Naprimer, sinteti~kite homopolimeri (polimeri sostaveni od pove}e kopii od edna aminokiselina ili {e}er) te`neat da nemaat imunogenost bezrazlika na nivnata golemina. Heteropolimerite obi~no se po imunogeni od homopolimerite. Ovie prou~uvawa poka`uvaat deka hemiskatakompleksnost pridonesuva za imunogenosta. Se zabele`uva deka site ~etiri nivoa od organizacija kaj proteinot-prvi~na, vtori~na, treti~na i~etvrti~na-pridonesuvaat za strukturnata kompleksnot na proteinot i na toj na~in vlijaat vrz negovata imunogenost (slika 4-2).Soodvetno pretstavenite lipidni protivgeni mo`at da pottiknat B kleto~ni reakcii. Na primer, lipidite mo`e da slu`at kako hapteni prika~eni zasoodvetni molekuli nosa~i, kako {to se proteinite hemocijanin od morski pol`av (HMP) (keyhole limpet hemocyanin = HMP) ili govedskiot serumalbumin (BSA). Pri imunizirawe so ovie lipidno-proteinski kowugati, mo`no e da se dobijat protivtela koi se visoko specifi~ni za celnite lipidi.S b j j}So upotreba na ovoj pristap, nau~nicite sozdadoa protivtela kon mnogu razli~ni lipidni molekuli, vklu~uvaj}i steroidi, kompleks masno-kiselinskiprodukti i masno-rastvorlivi vitamin kako {to e vitaminot E. Takvite protivtela se od zna~itelna prakti~na vrednost - mnogu klini~ki analizi zaprisustvoto i koli~estvata na medicinski zna~ajnite lipidi se baziraat na protivtela. Na primer, odreduvaweto nivoata na kompleksnata grupalipidi poznati kako leukotrieni mo`e da bide korisno pri ispituvawe pacientite so astma (vidi poglavje 15). Analizite koi se baziraat naupotrebata na protivlipidni protivtela ovozmo`uvaat otkrivawe pikogramski koli~ini na leukotrien C4. Bidej}i protivleukotrienot C4 malkureagira ili ne reagira so sli~ni soedinenija, kako {to e leukotrienot D4 ili E4, mo`e da se koristi za merewe leukotrienot C4 kaj primeroci koi gosodr`at ova soedinenie i mnogu drugi strukturno povrzani lipidi. Druga medicinsko zna~ajna upotreba vklu~uva otkrivawe prednizon,imunosupresiven steroid koj ~esto se vnesuva kako del od programata za supresija na otfrlaweto presaden organ. Odr`uvaweto soodvetni nivoa vo krvna ovie i drugi imunosupresivni lekovi e zna~ajno za uspe{en rezultat pri presaduvaweto, a imunoanalizite koi se baziraat na protivtelata sekoristat rutinski za da gi napravat ovie proceni.
Peifatlivost za obrabotka i prika`uvawe protivgenRazvojot na humoralnite (protivtelo-posreduvani) i T-kleto~no posreduvanite imuni reakcii baraat me|udejstvo na T kletkite so protivgenot koj eobraboten i pretstaven zaedno so GTK molekulite. Golemite, nerastvorlivi ili nasobranite makromolekuli vo osnova se po imunogeni od malite,rastvorlivi bidej}i pogolemite molekuli polesno se fagocitiraat i obrabotuvaat. Makromolekulite koi ne mo`e da se razgradat i pretstavat so GTKmolekulite se slabi imunogeni. Ova mo`e da se prika`e so polimeri od D-amino kiselini, {to pretstavuvaat stereoizomeri na L-amino kiselinitekoi prirodno se pojavuvaat Bidej}i razgraduva~kite enzimi vo protivgen prika`uva~kite kletki mo`e da razgraduvaat samo proteini koi sodr`at L-koi prirodno se pojavuvaat. Bidej}i razgraduva~kite enzimi vo protivgen prika`uva~kite kletki mo`e da razgraduvaat samo proteini koi sodr`at Lamino kiselini, polimerite so D-amino kiselini ne mo`e da se obrabotat i zatoa se slabi imunogeni.
6

Biolo{kiot sistem pridonesuva za imunogenosta
D j j b } b b jDuri ako makromolekulata ima svojstva koi pridonesuvaat za imunogenosta, nejzinata sposobnost za pottiknuvawe imun odgovor }e zavisi od odredeni osobini na biolo{kiot sistem so koj se sretnuvaprotivgenot. Faktorite koi pridonesuvaat za imunogenosta vklu~uvaat genetski sostav, na~in na koj e prika`anamaterijata i upotrebata na agensi (dodatoci) za zasiluvawe imunogenosta.
Genotip od `ivotnoto primatel
Glavniot faktor koj ja odreduva imunolo{kata reaktivnost mo`e da bide genotipot na primatelot. Genetskiot sostav (ili genotipot) od imuniziranoto `ivotno vlijae vrz tipot na imuniot odgovor koj`ivotnoto go manifestira, kako i stepenot na odgovorot. Na primer, Hju MekDevid poka`a deka dva razli~ni vidovi gluvci reagiraat dosta razli~no po izlo`uvaweto na sinteti~ki polipeptidenimunogen. Edniot vid sozdava visoki nivoa na serumsko protivtelo, drugiot vid sozdava niski nivoa. Koga e napraveno vkrstuvawe pome|u dvata vida, naslednicite od F1 generacijata poka`uvaat prose~enodgovor kon imunogenot. So nazadno vkrstuvawe, genot koj ja kontrolira imunolo{kata reaktivnost be{e ozna~en vo podra~jeto od grupata geni nare~eni glaven tkivnosovpadliv kompleks (GTK, vidipoglavje 8). Brojni eksperimenti so prosto definirani imunogeni poka`aa genetska kontrola na imunolo{kata reaktivnost, glavno posreduvana za genite vo GTK. Ovie podatoci uka`uvaat dekaproduktite od GTK genite, koi funkcioniraat vo prika`uvaweto na obraboteniot protibgen na T kletkite, igraat centralna uloga vo odreduvaweto stepenot do koj `ivotnoto reagira kon imunogenot.
Reakcijata na `ivotnoto kon protivgenot e pod vlijanie na genite koi gi kodiraat B- i T-kleto~nite receptori i od genite koi kodiraat razli~ni proteini vklu~eni vo imunite regulatorni mehanizmi.Genetskata promenlivost na site ovie geni vlijae vrz imunogenosta na dadena makromolekula kaj razli~ni `ivotni. Ovie genetski efekti vrz imunogenosta }e bidat podetalno objasneti vo natamo{nitepoglavja.
Imunogensko dozirawe i na~inot na vnesuvawe
Sekoj eksperimentalen imunogen sozdava odredena kriva na dozen odgovor, koja se odreduva so merewe imunot odgovor na razli~ni dozi i razli~ni na~ini na vnesuvawe. Rekacijata na protivteloto semeri so odreduvawe nivoto na protivteloto koe e prisutno vo serumot od imuniziranite `ivotni Procenuvaweto T kleto~nite reakcii e poslo`eno no mo`e da se odredi so procenuvawe porastot nameri so odreduvawe nivoto na protivteloto koe e prisutno vo serumot od imuniziranite `ivotni. Procenuvaweto T kleto~nite reakcii e poslo`eno no mo`e da se odredi so procenuvawe porastot nabrojot na T kletkite koi nosat T kleto~ni receptori koi go prepoznavaat imunogenot. Odredena kombinacija na optimalna doza i na~in na vnesuvawe }e pottikne najvisok imun odgovor kaj dadeno`ivotno.
Isto taka nedovolnata doza nema da stimulira imun odgovor bidej}i ne mo`e da aktivira dovolno limfociti ili bidej}i, vo nekoi slu~ai, odredeni niski dozi mo`e da pottiknat sostojba na imunolo{kanereaktivnost, ili podnoslivost. Nasproti toa, zna~itelno visokata doza isto taka mo`e da pottikne podnoslivost. Imunot odgovor kaj glu{ecot kon pro~isteniot pnevmokokalen kapsularenpolisaharid go prika`uva zna~eweto na dozata. Doza od 0,5 mg protivgen ne mo`e da pottikne imun odgovor kaj gluvcite, dodeka 1000 pati poniska doza od istiot protivgen (5 h 10-4 mg) pottiknuvahumoralen odgovor na protivtela. Edna doza od najmnogute eksperimaentalni imunogeni nema da pottikne silen odgovor, tuku, povtorno vnesuvawe vo period od nekolku nedeli dokolku ima potreba za toa.Takvite povtorni vnesuvawa, ili zasiluva~i, go zgolemuvaat klonskito razmno`uvawe na protivgen-specifi~nite B i T kletki i na toj na~in ja zgolemuvaat limfocitnata populacija koja e specifi~naza imunogenot.
Ekperimentalnite imunogeni obi~no se vnesuvaat parenteralno (para, okolu - enterik, utroba) - toa zna~i, na na~ini razli~ni od oralniot. Voobi~aeni se slednive na~ini na vnesuvawe:
vnatrevenozen (vv): vo venata; vnatreko`en (vk): vo ko`ata; podko`en (pk): pod ko`ata; vnatremuskulen (vm): vo muskulot; vnatreperitonealen (vp): vo peritonealnata praznina
Na~inot na vnesuvawe silno vlijae koi imuni organi i kleto~ni populaciite }e se vklu~at pri odgovorot. Protivgenot koj se vnesuva vnatrevenozno prvo se nosi do slezinata, dodeka protivgenot koj sevnesuva podko`no prvo odi vo lokalnite limfni jazli. Razlikite vo limfoidnite kletki koi gi naseluvaat ovie organimo`e da se vlijaat vrz natamo{niot imun odgovor.
P (A j )Pomaga~i (Adjuvansi)
Pomaga~i (adjuvansi od latinskiot izraz adjuvare, da pomogne) se supstancii koi, koga }e se izme{aat i se vnesat so protivgenot, ja zgolemuvaat imunogenosta na toj protivgen. Pomaga~ite ~esto sekoristat pri istra`uvawe i klini~ka podgotovka za zasiluvawe imunot odgovor koga protivgenot ima niska imunogenost ili koga protivgenot e dostapen vo mali koli~ini. Na primer, protivtelniotodgovor kaj glu{ec po imunizacija so BSA mo`e da se zgolemi do pet pati ili pove}e ako BSA se vnese zaedno so pomaga~. To~no kolku pomaga~ite go zgolemuvaat imunot odgovor ne e celosno jasno, nodel od poznatite pomaga~i (na primer, sintetskite poliribonukleotidi i bakteriskite lipopolisaharidi) sega se utvrdeni kako ligandi za tolovidnite receptori vosteblestite kletki i makrofagite (vidi poglavje 3) i na toj na~in stimuliraat imun odgovor preku aktivirawe vrodeniot iumunolo{ki sistem.
Vo osnova, pomaga~ite gi predizvikuvaat eden ili pove}e od slednive efekti:
Opstojuvaweto na protivgenot e prodol`eno; Kostimulatornite signali se zgolemeni; Lokalnoto vospalenie e zgolemeno; Stimulirano e nespecifi~noto razmno`uvawe na limfocitite.
Aluminium natrium sulfat (alum) e pomaga~ koj go prodol`uva opstojuvaweto na protivgenot i e edinstveniot pomaga~ koj e odobren za op{ta upotreba kaj lu|eto. Koga protivgenot }e se izme{a soalumot, solta go talo`i protivgenot. Vnesuvaweto na ovoj alumen talog doveduva do posporo osloboduvawe protivgenot od oblasta na vnesuvawe, taka {to efektivnoto vreme za izlo`uvawe protivgenotse zgolemuva od nekolku denovi bez pomaga~ do nekolku nedeli so pomaga~.Alumniot talog isto taka ja zgolemuva goleminata na protivgenot, so {to ja zgolemuvamo`nosta za fagocitoza.
Pomaga~ite voda vo maslo (water in oil) isto taka go prodol`uvaat opstojuvaweto na protivgenot. Preparatot poznat kako necelosen Frojndov pomaga~ sodr`i protivgen vo voden rastvor, rastitelnomaslo i emuluzivno soedinenie kako {to e manid monooleat, koi go rasprsnuva masloto vo mali kapki koi go opkru`uvaat protivgenot. Potoa protivgenot mnogu sporo se osloboduva od mestoto navnesuvawe. Ovoj preparat se bazira na necelosniot Frojndov pomaga~, prviot namerno formuliran visoko efektiven pomaga~, sozdaden od strana na Xul Frojnd pred mnogu godini i koj sodr`i toplotnoubieni tuberkulozni bakterii (Mycobacteria), kako dopolnitelna sostojka. Muramil dipeptidot, del od mikobakteriskiot kleto~en yid, gi aktivira steblestite kletki i makrofagite, pravej}i gocelosniot Frojndov pomaga~ daleku pomo}en od necelosniot oblik. Aktiviranite steblesti kletki i makrofagite se pofagocitni od nivnite neaktivirani sprotivni delovi i la~at povisoki nivoa nacelosniot Frojndov pomaga~ daleku pomo}en od necelosniot oblik. Aktiviranite steblesti kletki i makrofagite se pofagocitni od nivnite neaktivirani sprotivni delovi i la~at povisoki nivoa namolekuli koi pottiknuvaat kostimulacija i zasiluvawe na T-kleto~niot imun odgovor. Spored toa, prika`uvaweto na protivgenite i neophodniot kostimulatoren signal obi~no se zgolemeni voprisustvo na pomaga~.
Alum i Frojndovite pomaga~i isto taka stimuliraat lokalen, hroni~en vospalitelen odgovor koj gi privlekuva i fagocitite i limfocitite. Ova navleguvawe kletki vo mestoto na vnesuvaweto pomaga~~esto doveduva do sozdavawe gusta, makrofagno bogata masa kletki poznata kako granulom.
7

Epitopi
Kako {to spomnavme prethodno, imunite kletki ne stapuvaat vo me|udejstvo so, iliprepoznavaat, cela imunogena molekula. Namesto toa, limocitite prepoznavaatdiskretni oblasti na makromolekula nare~eni epitopi, ili protivgeni odrednici(determinanti). Prou~uvawata so mali protivgeni otkrija deka B i T kletkiteprepoznavaat razli~ni epitopi od istata protivgena molekula. Na primer, kogaglu{ecot se imunizira so glukagon, mal ~ove~ki hormon od 29 amino kiselini,b Tbe{e pottiknato protivtelo za epitopite vo amino terminalniot del, dodeka Tkletkite reagiraa samo na epitopi vo karboksilno terminalniot del.
Limfocitite mo`e da stapat vo me|udejstvo so kompleksen protivgen na nekokolkunivoa od strukturata od protivgenot. Epitop vo proteinskiot protivgen mo`e davklu~i elementi od prvi~nata, vtori~nata, treti~na pa duri i ~etvrti~nastruktura na proteinot (vidi slika 4-2). Vo polisaharidite, ~esto se prisutnirazgraneti verigi, a vrvovite od grankata mo`e da pridonesat za sozdavaweepitopi B kletkite prepoznavaat raatvorliv protivgen koga se vrzuva zaepitopi. B kletkite prepoznavaat raatvorliv protivgen koga se vrzuva zaprotivtelnata molekula vo membaranata od B kletkite. Bidej}i V kletkitevrzuvaat protivgenot koj e sloboden vo rastvor, epitopite koi tie gi prepoznavaatte`neat da bidat dosta pristapni oblasti vrz izlo`enata povr{ina od imunogenot.T-kleto~nite epitopi od proteinite se razlikuvaat po toa {to se peptidi koiobi~no proizleguvaat od enzimsko razgraduvawe na patogenite proteini i seprepoznavaat od T kleto~nite receptori samo koga se vo kombinacija so GTKprotivgen. Spored toa, nema uslov za pristapnost vo rastvor kako {to e so B-kleto~niot epitop. Tabela 4-2 gi naveduva golemite razliki pri protivgenosta naB- i T-kletkite.
8

B-kleto~nite epitopi imaat karakteristi~ni osobini
B-kleto~nite epitopi vo prirodnite proteini vo osnova se sostaveni od hidrofilni aminokiselini vrz proteinskata povr{ina koi se topografski pristapni za memebranski vrzanite iliza slobodnite protivtela. B-kleto~niot epitop mora da bide pristapliv za da mo`e da se vrze zaprotivtelo. Vo osnova, za ispaknatite delovi od povr{inata na proteinot poverojatno e da bidatprepoznati kako epitopi i ovie podra~ja obi~no se sostaveni od dominantno hidrofilni aminokiselini. Amino kiselinskite sekvenci koi se skrieni vo vnatre{nosta na proteinot ~esto sesostaveni od dominantno hidrofobni amino kiselini i ne mo`at da funkcioniraat kako B-kleto~niepitopi dokolku prvo proteinot ne se denaturira.
(pregled na slika 4-3 Proteinskite protivgeni obi~no sodr`at i posledovatelni i neposledovatelni( ) 5B-kleto~ni epitopi. (a) [ema na mioglobulin od sperma na kit gi poka`uva mestata na 5
posledovatelni B-kleto~ni epitopi (sino). (b) [ema vo vid na lenti od lizozim od belka na jajce odkoko{ka poka`uva ostatoci koi so~inuvaat eden neposledovatelen (konformaciski) epitop.Ostatocite koi se dopiraat so lesnite verigi, te{kite verigi, ili i so dvata se prika`ani vo crveno,sino i belo. Ovie ostatoci se mo{ne rasprostraneti vo sekvencata od amino kiselinite, no sepribli`uvaat so dipliweto proteinot. (referenca) strana 82)
B - kleto~nite epitopi mo`e da se sostaveni od sekventni posledovatelni ostatoci dol`polipeptidniot veriga ili od neposledovatelni ostatoci od delovi na verigata koi se spoeni sozdiplenata konformacija na protivgenot. Pove}eto protivtela koi nastanuvaat od globularniteproteini se vrzuvaat za proteinot samo koga toj e vo prirodna konformacija. Poradi toa {tor r u r j r r d f r c j r dtalo`eweto na vakvite protivgeni obi~no ja menuva strukturata na nivnite epitopi, protivtelatakon prirodniot protein ne se vrzuvaat za istalo`eniot protein.Pri edna serija eksperimenti, pet razli~ni posledovatelni (sekventni) epitopi, pri {to sekojsodr`e{e {est do osum posledovatelni amino kiselini, bea pronajdeni vo mioglobinot od sperma nakit. Sekoj od ovie epitopi se nao|a na povr{inata od molekulata pri vrskite pome|u alfa -spiralnite podra~ja(slika 4-3a). Mioglobinot od spermata na kitot isto taka sodr`i nekolkuneposledovatelni (nesekventi) epitopi, ili konformaciski odrednici. Ostatocite koi giso~inuvaat ovie epitopi se dosta oddale~eni vo prvi~nata amino-kiselinska sekvenca, no sezbli`uvaat vo treti~nata struktura od molekulata. Vakvite epitopi se prisutni samo koga proteinote vo negovata prirodna konformacija (priroden oblik). Dobro karakteriziran neposledovatelenepitop vo lizozimot od belkata od jajceto na koko{ka (LBK) (hen egg-white lysozyme = HEL) e
4 3b I j LKBprika`an na slika 4-3b. Iako ostatocite od amino kiselinite koi go so~inuvaat ovoj epitop na LKBse dosta oddale~eni vo prvi~nata sekvenca na amino kiselinite, se zbli`uvaat so treti~noto diplewena proteinot.
9

Posledovatelnite i neposledovatelnite epitopi vo osnova se odnesuvaat razli~no koga proteinot e denaturiran, podelen ilinamalen. Na primer, soodvetnoto delewe mioglobinot od smermata na kitot mo`e da dade pet delovi, pri {to sekoj zadr`uvar r, d d d r d d d d , r j dr ueden posledovatelen epitop, kako {to e prika`ano od ispituvawata deka protivteloto mo`e da se vrze za sekoj del. Od drugastrana, neposledovatelnite epitopi vo osnova se gubat koga proteinot se deli ili koga se smaluvaat negovite disulfidnivrski. Na primer, LKB ima ~etiri me|uveri`ni disulfidni vrski, {to go odreduva kone~niot oblik na proteinot (slika 4-4a).Mnogu protivtela vo LKB prepoznavaat nekolku epitopi i sekoj od osumte razli~ni epitopi e prepoznat kako razli~noprotivtelo. Pove}eto od ovie epitopi se konformaciski odrednici zavisno od celosnata struktura na proteinot. Akome|uveri`nite disulfidni vrski na LKB se namalat so merkaptoetanol, neposledovatelnite epitopi se gubat. Poradi ovaapri~ina, protivteloto za prirodniot LKB ne se vrzuva za namaleniot LKB.
Eksperimentot so spre~uvawe prika`an na slika 4-4b i v ubavo ja prika`uvaa ovaa ideja. Protivteloto za konformaciskataodrednica, vo ovoj primer peptidna jamka prisutna vo prirodniot LKB, mo`e{e da se vrze za epitopot samo ako disulfidnatavrska koja ja odr`uva strukturata na jamkata be{e celosna. Informaciite za stukturnite uslovi na protivtelnotokombinacisko mesto se dobija so ispituvawe sposobnosta na strukturnite srodnosti na prirodniot protivgen da se vrze so toaprotivtelo. Ako strukturnata srodnost gi ima kriti~nite epitopi koi se prisutni vo prirodniot protivgen, }e se vrze zaprotivtelno kombinira~koto mesto, pri {to prirodniot protivgen }e go blokira nejziniot efekt. Pri ovoj metod naspre~uvawe, sposobnosta na zatvorenata jamka (slika 4-4b, desno) da spre~i povrzuvawe poka`uva deka zatvorenata jamka be{edovolno sli~na na LKB za da bide prepoznata od strana na protivteloto za prirodniot LKB. Iako otvorenata jamka (slika 4-4b, levo) ja ima{e istata amino kiselinska sekvenca kako i zatvorenata jamka, nema{e epitopi prepoznati od protivteloto izatoa ne mo`e{e da go blokira vrzuvaweto na LKB (slika 4-4v).
B-kleto~nite epitopi se stremat da bidat locirani vo podvi`nite oblasti na imunogenot i ~esto ja poka`uvaatpodvi`nata oblast. Xon A. Tajner i negovite kolegi gi analizirale epitopite na odreden broj proteinski protivgeni(miohemeriterin, insulin, citohrom c, mioglobin i hemoglobin) so sporeduvawe poziciite na poznatite B-kleto~ni epitopiso podvi`nosta na istite ostatoci. Nivnata analiza otkrila deka glavnite protivgeni odrednici kaj ovie proteini vo osnovabile locirani vo najpodvi`nite podra~ja. Ovie ispituva~i predlo`ija deka podvi`nata oblast na epitopite e maksimalnokomplementarna so oblasta za vrzuvawe protivtelo Se ispostavi deka pocvrstite epitopi se vrzuvaat poneefektivno Sepakkomplementarna so oblasta za vrzuvawe protivtelo. Se ispostavi deka pocvrstite epitopi se vrzuvaat poneefektivno. Sepak,poradi zagubata na entropija kako rezultat od vrzuvaweto za podvi`nata oblast, vrzuvaweto na protivteloto za podvi`niotepitop vo osnova e so pomal afinitet otkolku vrzuvaweto na protivteloto za cvrstiot epitop.
Kompleksnite proteini sodr`at pove}e preklopuva~ki B-kleto~ni epitopi, pri {to del od niv se imunodominantni. Votekot na mnogu godini, vo imunologijata postoe{e dogma deka sekoj globularen protein ima mal broj epitopi, sekoj namenet zavisoko pristapen podra~je i odreden po celosnoto sozdavawe proteinot. Sepak, neodamna se poka`a deka pove}eto odpovr{inata na globularniot protein e potencijalno imunogeni~na. Ova se poka`a so sporeduvawe protivgen-vrzuva~kiteoblici od razli~ni monoklonski protivtela za razli~ni globularni proteini. Na primer, koga 64 razli~ni monoklonskiprotivtela za GSA se sporeduvaat spored nivnata sposobnost da se vrzat za panel (kombinacija) od 10 razli~ni albumini odcica~i, se pojavija 25 razli~ni povtoruva~ki protivgen-vrzuva~kioblici, {to uka`uva na toa deka ovie 64 razli~niprotivtela prepoznale minimum 25 razli~ni epitopi za GSA.
Taka {to povr{inata na proteinot pretstavuva golem broj potencijalno protivgeni oblasti Podgrupata na protivgeniTaka {to, povr{inata na proteinot pretstavuva golem broj potencijalno protivgeni oblasti. Podgrupata na protivgenioblasti kaj daden protein koj e prepoznat od imuniot sistem kaj `ivotnite e dosta pomal od potencijalnata protivgena grupa ivarira od vid do vid, duri i kaj poedinci od dadeni vidovi. Kaj `ivotnite, odredeni epitopi od protivgenot se prepoznavaatkako imunogeni, drugi ne. Pokraj toa, odredeni epitopi, nare~eni imunodominantni, pottiknuvaat ponaglasen imun odgovor kajodredeni `ivotni okolku drugi epitopi. Mnogu e verojatno deka vnatre{nite topografski osobini na epitopot isto kako iregulatornite mehanizmi na `ivotnite vlijaat vrz imunodominacijata na epitopite.
10

Osnovna struktura na protivtelata
Prepoznavaweto na imunogenot od povr{inskite protivtela vo B kletkite pottiknuva razmno`uvawei diferencirawe vo pamte~kite B kletki i plazma kletki (vidi poglavje 11). Plazma kletkite la~atrastvorlivi protivtelo-molekuli so specifi~nost za protivgenot koj e identi~en so povr{inskiotreceptor od B kletkata roditel. Slednive delovi }e gi objasnat strukturnite krarakteristiki naprotivtelnite molekulite ovomo`uvaj}i im da gi ispolnat nivnite dve glavni funkcii:1. Vrzuvawe so nadvore{eni protivgeni koi se sretnale so doma}inot2. Posreduvawe efektorni funkcii za neutralizirawe ili otstranuvawe nadvore{nite napa|a~i.U{te od devetnaesetiot vek e poznato deka protivtelata se nao|aat vo krvniot serum. (vo sredniotdvaesetti vek, se poka`a deka protivtelata se prusutni vo drugi te~nosti koi gi la~i teloto: mleko,solzi plunkata `ol~kata itn) Krvta vo centrifuga mo`e da se podeli na te~nost i kleto~en delsolzi, plunkata, `ol~kata itn). Krvta vo centrifuga mo`e da se podeli na te~nost i kleto~en del.Te~niot del e plazmata, a kleto~niot del sodr`i crveni kletki, beli kletki i krvni plo~ki.Plazmata gi sodr`i site rastvorlivi mali molekuli i makromolekuli od krvta, vklu~uvaj}i gofibrinogenot i drugi proteini potrebni za sosiruvawe na krvta. Ako se dozvoli krvta ili plazmatada se sosirat (zgrut~at), te~nata faza koja ostanuva se narekuva serum. Prviot dokaz dekaprotivtelata se sodr`at vo odredeni delovi od serumski proteini dojde od klasi~en eksperiment naArni Tiselius i Elvin A. Kabat vo 1939 godina. Tie imuniziraa zajaci so proteinot jajcev albumin(albuminot od belkite vo jajcata), podgotvija serum i potoa go podelija serumot na dva dela.Elektroforetskoto dvoewe na eden del od serumot otkri ~etiri glavni delovi koi odgovoraat naserumskiot albumin i alfa, beta, i gama globulinite. Pred elektroforezata, drugiot del od serumotbil izme{an so imunizira~kiot protivgen (ovalbumin), ovozmo`uvaj}i sozdavawe imun precipitat(soedinenie od protivgen i protivtelo) koj bil otstranet, ostvaj}i gi ostanatite serum proteini, koine reagirale so protivgenot Sporedbata na elektroforetskite oblici na ovie dva dela serumi otkrine reagirale so protivgenot. Sporedbata na elektroforetskite oblici na ovie dva dela serumi otkrideka ima zna~itelen pad vo gama-globulinskiot vrv vo delot koj reagiral so protivgenot (slika 4-5).Taka {to, za gama-globulinskiot del se utvrdi deka sodr`i serumski protivtela, koi se nare~eniimunoglobulini za da se razlikuvaat od site drugi proteini koi bi mo`ele da se sodr`at vo gama-globulinskiot del. Prvi~nite eksperimenti na Kabat i Tiselius gi podelija serumskite proteini natri glavni nealbuminski delovi - alfa, beta i gama. Znaeme deka iako imunoglobulinot G (IgG),najobilnata klasa protivtelni molekuli, navistina najve}e se sretnuva kaj gama-globulinskiot del,zna~itelni koli~ini od nego i drugi klasi molekuli-protivtela se sretnuvaat vo alfa i betadelovite vo serumot.
11

Protivtelate se heterodimeri
Protivtelnite molekuli imaaat zaedni~ka struktura od ~etiri peptidni verigi (slika 4-6).Ovaa struktura se sostoi od dva identi~ni lesni (L) verigi, polipeptidi od okolu 22000 Dai dva identi~ni te{ki (T) verigi, pogolemi polipeptidi od okolu 55000 Da ili pove}e.Sekoj lesna veriga se vrzuva za te{ka veriga preku disulfidna vrska i so nekovalentnivrski kako {to se solnite vrski, vodorodnite vrski i hidrofobi~ni stapuva vo me|udejstvoza sozdavawe heterodimer (T-L). Sli~ni nekovalentni i disulfidni vrski me|usebno gipovrzuvaat dvete kombinacii od identi~ni te{ki i lesni (T-L) verigi za da ja formiraatosnovnata struktura na protivteloto od ~etiri verigi (T-L)2, dimer od dimeri. Kako {to }e
b j fvidime ponatamu, to~niot broj na precizni pozicii na disulfidnite vrski koi gipovrzuvaat dimerite se razlikuva pome|u klasite i potklasite na protivtelata.Prvite 110 ili pribli`no tolku amino kiselini od amino-krajnoto podra~je na lesni ite{ki verigi dosta varira pome|u protivtelata so razli~na protivgena specifi~nost.Ovie segmenti od visoko promenlivata sekvenca se nare~eni V podra~ja: VL kaj lesniteverigi i VT kaj te{kite verigi. Site razliki vo specifinosta prika`ani od razli~niprotivtela mo`e da se svedat na razliki vo sekvencite na amino kiselinite vo Vpodra~jata. Vsu{nost, pove}eto razliki razliki pome|u protivtelata vo oblasti vo Vpodra~jata nare~eni komplementarno odreduva~ko podra~je (KOP), a ova KOP, i kajlesnite i te{kite verigi, go so~inuvaat protivgeno vrzuva~koto mesto na protivtelnatamolekula. Nasproti toa, vo sekoja posebna klasa protivtela, se gledaat pomali razliki koga}e se sporedat sekvencite niz ostatokot od molekulata. Podra~jata so relativno postojanasekvenca pokraj promenlivite podra~ja se identi~ni so K podra~jata, KL kaj lesnite verigii KT kaj te{kite verigi. Protivtelata se glikoproteini, so odredeni isklu~oci, oblastitena prika~uvawe jaglehidratite se ograni~eni na postojanoto podra~je. Ne ja razbiramecelosno ulogata koja ja igra glikozilacijata na molekulite, no verojatno ja zgolemuvarastvorlivosta na molekulite. Nesoodvetnata glikozilacija ili nejzinoto otsustvo vlijaevrz stapkata so koja protivtelata se ~istat od serumot i ja namaluva efikasnosta name|udejstvota pome|u protivteloto i drugite proteini so koj stapuva vo soodnosme|udejstvota pome|u protivteloto i drugite proteini so koj stapuva vo soodnos.
12

Hemiskite i enzimskite metodi ja otkrija osnovanata struktura na protivteloto
Na{ite informacii za osnovanata struktura na antitelata proizleguva od razli~ni eskperimenti. Na{ite znaewe za osnovanata struktura naprotivtelata proizleguva od razli~ni eskperimenti. Koga gama-globulinskiot del od serumot }e se oddeli na delovi so golema i mala molekulskate`ina, protivtelata od okolu 150000 Da, nare~eni imunoglobulin G (IgG), se sretnuvaat kaj delot so mala molekulska te`ina. Pri klu~eneksperiment, brzoto razgraduvawe IgG so proteoliti~niot enzim papain sozdade tri delovi, od koi dva bea identi~ni, a tretiot be{e mo{nerazli~en (slika 4-7). Dvata identi~ni delovi (sekoj od 450000 Da) poseduvaa protivgen vrzuva~ka aktivnost i bea nare~eni Fpv fragmenti(üFragment, protivgen vrzuva~kiû ili na angliski "Fragment, antigen binding" = Fab). Drugiot del (50000 Da) voop{to ne poseduva{e aktivnostza vrzuvawe protivgeni. Bidej}i se otkri deka toj kristalizira pri ~uvawe vo ladno, be{e nare~en Fk fragment (üFragment, kristalizira~ki ilina angliski "Fragment, crystallizable = Fc). Razgraduvaweto na IgG so razli~en proteoliti~en enzim, pepsin, u{te edna{ poka`a dekaprotivgen-vrzuva~kite osobini na protivteloto mo`e da se oddelat od ostanatiot del od molekulata. Razgraduvaweto na pepesinot sozdade edenfragment od 100000 Da, sostaven od dve podeinici kako Fpv nare~eni F(pv’)2 fragment, koj se vrzuva za protivgen. Fk fragment ne se izvadi sotretmanot so pepsin bidej}i be{e razgraden na brojni mali peptidi.
Klu~en naod se napravi pri sveduvawe pove}everi`nata struktura na IgG koga molekulata be{e podlo`ena na smaluvawe merkaptoetanol ialkilacija, hemisko tretirawe koe nepovratno gi rase~uva disulfidnite vrski no ne i peptidnite vrski. Ako tretiraniot primerok sealkilacija, hemisko tretirawe koe nepovratno gi rase~uva disulfidnite vrski no ne i peptidnite vrski. Ako tretiraniot primerok sehromatografira na kolona koja gi oddeluva molekulite po golemina, stanuva jasno deka neo{tetenata molekula IgG od 150000 Da, vsu{nost, bilasostavena od podedinici. Sekoja IgG molekula sodr`i dva pogolemi pepetidni verigi od okolu 50000 Da, nare~eni te{ki (T) verigi i dva pomaliverigi od pribli`no 22000 Da, nare~eni lesni (L) verigi (slika 4-7).
Samite protivtela se koristat za odreduvawe kako enzimskite produkti pri digestija - Fpv, F(pv’)2 i Fk - se povrzani so redukciskite produktiod lesnite i od te{kite verigi. Na ova pra{awe se odovori so upotreba na protivserum od koza koja be{e imunizirana ili so Fpv fragmenti iliso Fk fragmenti od IgG na zajak. Protivteloto od Fpv fragmentot mo`e{e da reagira i so T i L verigite, dodeka protivteloto od Fk fragmentotreagira{e samo so T verigata. Ovie naodi dovedoa do zaklu~ok deka Fpv fragmentot sodr`i del od te{kiot plus nedoprena lesna veriga i deka Fksodr`i samo delovi so te{kata veriga. Od ovie rezultati i onie koi se spomeneti pogore, be{e sozdadena strukturata na IgG prika`ana na slika4-6. Spored ovoj model, IgG molekulata se sostoi od dve identi~ni T verigi i dve identi~ni L verigi, koi se povrzani so disulfidni vrski.
Verojatniot sleden ~ekor pri dobivaweto slika za strukturata na protivteloto ja odreduva{e nivnata amino kiselinska sekvenca, sepak nastanaproblem. Populacijata od protivtelata vo serum gama-globulinskiot del se sostoi od heterogen spektar na protivtela - mnogu razli~niprotivtela sekoj prisuten vo mali koli~ini Duri i da se napravi imunizacija so kowugat hapten nosa~ sozdadenata populacija protivtelataprotivtela sekoj prisuten vo mali koli~ini. Duri i da se napravi imunizacija so kowugat hapten-nosa~, sozdadenata populacija protivtelatanaso~ena protiv haptenot }e bide heterogena, bidej}i mnogu protivtela prepoznavaat razli~ni epitopi od haptenot. Sepak, analizirawesekvencata na amino kiselinite, bara ~ist primerok od molekulata koja se prou~uva. Ovaa pre~ka za analizirawe sekcencata se nadmina soupotreba na imunoglobulini od pacienti so multipen mielom, rak na plazma kletkite koi sozdavaat protivtela. Plazma kletkite kaj zdravo licese kletki vo krajna faza koi la~at molekulski vidovi na protivtela vo ograni~en period na vreme a potoa umiraat. Nasproti toa, klon od plazmakletkite kaj liceto so multipen mielom ispadanal od kontrola na `ivotniot ciklus i se razmno`uva na nereguliran na~in bez potreba za kakvobilo aktivirawe od strana na protivgen za pottiknuvawe razmno`uvawe. Iako takvata kancerozna plazma kletka, nare~ena mielomska kletka, etransformirana, nejzinata ma{inerija za sintetizirawe proteini i sekretornite funkcii ne se promeneti. Spored toa, kletkata prodol`uva dago la~i molekualskoto homogeno protivtelo. Ova protivtelo ne se razlikuva od obi~nite protivtelni molekuli no e nare~eno mielomski proteinza da go ozna~i negoviot izvor. Kaj pacient so mnogukraten mielom, na mielomskiot protein otpa|aat 95% od serumskite imunoloblini. Kaj mnogupacienti, mielomskite kletki isto taka la~at vi{ok koli~ini na lesni verigi. Ovoj vi{ok lesni verigi najprvo be{e otkrien vo urinata odpacientite so mielom i bea imenuvani kako Bens-Xonsonovi proteini spored nivnite otkriva~i.
Mnogukratniot mielom isto taka se pojavuva kaj drugi `ivotni. Kaj gluvcite mo`e da se javi spontano, kako {to se javuva kaj lu|eto, ili sovnesuvawe mineralno maslo vo peritonealnata praznina koe mo`e da se sozdade uslovi vo prilog na pottiknuvaweto mielom Klonovite odvnesuvawe mineralno maslo vo peritonealnata praznina koe mo`e da se sozdade uslovi vo prilog na pottiknuvaweto mielom. Klonovite odopasnite plazma kletki koi se razvivaat se nare~eni plazmacitomi, a mnogu od niv se nare~eni PKMM, {to zna~i pottiknuvawe plazmacitomskikletki so mineralno maslo. Golem broj vidovi na PKMM od glu{ec koi la~at razli~ni klasi imunoglobulini se vo upotreba od strana naAmerican Type-Culture Collection, neprofitno skladi{te na vidovi kletki koi ~esto se koristat vo istra`uvaweto. Metodot za sozdavawemonoklonski protivtela so sakanata specifi~nost koj e razvien od Gorges Kohler i Cezar Mil{tajn (vidi dolu) gi koristi plazmacitomskitelinii. Ovaa hibridomna tehnologija ovozmo`uva sozdavawe homogeni protivtela za strukturnite i funkcionalnite prou~uvawa i za brojniklini~ki upotrebi.
13
Sekvencata od lesnite verigi otkri postojani i promenlivi podra~ja
Koga sekvencite na amino kiselinite od nekolku Bens-Xonsonovi proteini (lesni verigi) odrazli~ni lica }e se sporedat, }e se dobijat izvonredni oblici. Za amino-krajnata polovina odverigata, koja se sostoi od 100 do 110 amino kiselini, se otkri deka varira pome|u razli~ni Bens-Xonsonovi proteini. Ovoj podra~je e nare~eno varijabilno (V) podra~je. Karboksil-krajnitepolovini od molekulite, nare~eni konstantni (K) podra~ja, gi imaat edna od dvete sekvenci na aminokiselini. Ova dovede do sva}awe deka postojat dva vida lesni verigi, kapa (κ) i lambda (λ). Kaj lu|eto,60% od lesnite verigi se kapa a 40% se lambda, dodeka kaj gluvcite 95% od lesnite verigi se kapa asamo 5% se lambda. Obi~na protivtelna molekula sodr`i samo eden vid lesna veriga, ili kapa ililambda, nikoga{ i dvete.Sekvencite od amino kiselinite na lambda konstantnoto podra~je na lesnata veriga poka`uvaat malirazliki koi se koristat za klasificirawe lambda lesnite verigi vo podgrupi Kaj gluvcite i lu|etorazliki koi se koristat za klasificirawe lambda lesnite verigi vo podgrupi. Kaj gluvcite i lu|eto,postojat ~etiri podgrupi: lambda1, lambda2, lambda3 i lambda4. Zamenite za amino kiselinite nasamo nekolku pozicii se odgovorni za razlikite vo podtipovite.
Postojat pet vidovi glavni klasi na te{ki verigi
Za analizaata na sekvencata na te{kite verigi, mielomskite proteini se reducirani somerkaptoetanol i alkilirani, a te{kite verigi se odvoeni so gel filtracija vo denaturira~kirastvor. Koga }e se sporedat sekvencatite na amino kiselinite od nekolku te{ki verigi namielomski proteini, }e se pojavi oblik sli~en kako onoj kaj lesnite verigi. Amino-krajniot del odverigata koj se sostoi od 100 do 110 amino kiselini poka`a golema varijacija vo sekvencata pome|uverigata, koj se sostoi od 100 do 110 amino kiselini, poka`a golema varijacija vo sekvencata pome|ute{kite verigi od mielomot i zatoa be{e nare~eno varijabilno (V) podra~je. Ostanatiot del odproteinot otkri pet osnovni modeli na sekvenci, {to odgovaraat na pet razli~ni konstantni (K)podra~ja so te{ki verigi: mi, delta, gama, epsilon i alfa. Sekoj od ovie pet razli~ni te{ki verigi enare~en izotip. Dol`inata na konstantnoto podra~je e pribli`no 330 amino kiselini za delta, gama,i alfa i 440 amino kiselini za mi i epsilon. Te{kite verigi kaj dadena protivtelna molekula jaodreduvaat klasata na toa protivtelo: IgM (mi), IgG (gama), IgA (alfa), IgD (delta) ili IgE(epsilon). T verigite od sekoja klasa mo`e da se spojat so kapa ili so lambda lesnite verigi. Ednaprotivtelna molekula ima dve identi~ni te{ki i dve identi~ni lesni verigi, T L , ili pove}e (T L )od ovaa osnovna struktura so ~etiri verigi (tabela 4-3).Malite razliki vo sekvencite na amino kiselinite so alfa i gama te{ki verigi dovede doponatamo{na klasifikacija na te{kite verigi vo podizotipovi koi ja odreduvaat potklasata naf c j r d j dr duprotivtelnite molekuli koi tie gi so~inuvaat. Kaj lu|eto, postojat dva vida podizotipovi na alfate{kite verigi - alfa1 i alfa2 (i dve potklasi, IgA1 i IgA2) i ~etiri podiztotipovi na gama te{kiverigi: gama1, gama2, gama3 i gama4 (i ~etiri potklasi), IgG1, IgG2, Ig3 i IgG4). Kaj gluvcite,postojat ~etiri podiztotipovi, gama1, gama2a, gama2b i gama3 i soodvetnite potklasi.
14

Imunoglobulinite poseduvaat mnogu oblasti vrz baza naimunoglobulinskoto diplewe
Celosnata struktura na imunoglobulinskata molekula se odreduva sporedprvi~nata, vtori~nata, treti~nata i ~etvrti~nata organizacija naproteinot. Prvi~nata struktura - sekvencata od amino kiselinite - gisodr`i promenlivite i postojanite podra~ja od te{kite i lesnite verigi.Vtori~anata struktura se sozdava so diplewe izdol`enata polipeptidnaVtori~anata struktura se sozdava so diplewe izdol`enata polipeptidnaveriga, prvo vo sebe, potoa vo serija protivparalelni beta zdipleni lenti(slika 4-8).
(slika 4-8 Strukturna formula na beta zdiplena plo~ka koja sodr`i dveprotivparalelni beta lenti. Strukturata e povrzana so vodorodni vrski odsosednite granki na polipeptidnite verigi. Strani~nite grupi na aminod r d r r rukiselini (R) se podredeni perpendikularno kon ramni{teto na plo~kata.
15

Potoa verigite se diplat vo treti~na struktura od kompaktni globularnidomeni, koi so sosednite domeni se povrzani polipeptidni lenti pome|upodra~jata od beta zdiplenata lenta. Na krajot, globularnite domeni odsosednite te{ki i lesni polipeptidni verigi stapuvaat vo me|udejstvo so~etvrti~nata struktura (slika 4-9), formiraj}i funkcionalni domeni koi $ovozmo`uvaat na molekulata specifi~no da se vrze za protivgenot i vo istovreme da izvr{i odreden broj biolo{ki efektorni funkcii.
(slika 4-9 Dva prikaza na neo{tetena protivtelna molekula. Me|udejstvopome|u domenite vo oddelnite verigi od imunoglobulinskata molekula sezna~ajni za ~etvrti~nata struktura. (a) Modelot na IgG molekulata, vrzbaza na rendgenskata kristolografska analiza, poka`uva povrzanostipome|u domenite vo oddelnite verigi na imunoglobulinskata molekula sekriti~ni za ~etvrti~nata struktura. Sekoja topka pretstavuva ostatok odaminokiselina Pogolemite temni topki se jaglehidrati Zabele`ete dekaaminokiselina. Pogolemite temni topki se jaglehidrati. Zabele`ete dekaKT2 domenite se izdadeni poradi vnatre{niot jaglehidrat, ova ovozmo`uvapristapnost do drugite molekuli kako {to se komplementnite delovi. Dvetelesni verigi se prika`ani vo nijansi od crvena boja. Dvete te{ki verigi, vonijansi od sina boja. (b) [ematskiot prikaz poka`uva soodnos pome|udomenite od te{kite i od lesnite verigi.
16

Detalnata analiza na amino-kiselinskite sekvenci od imunoglobulinskitete{ki i lesni verigi poka`a deka i dvete verigi sodr`at nekolkuhomologni edinici od okolu 110 amino kiselinski ostatoci. Vo sekojaedinica, nare~ena domen, me|uveri`nata disulfidna vrska formira jamka odokolu 60 amino kiselini. Lesnite verigi sodr`at eden promenliv domen,(VL) i eden postojan domen (KL). Te{kite verigi sodr`at eden promenlivdomen (VT) i ili tri ili ~etiri postojani domeni (KT1, KT2, KT3 i KT4), vozavisnost od klasata na protivteloto (slika 4-10).
(slika 4-10 (a) Te{kite i lesnite verigi se zdipleni vo domeni, pri {tosekoj sodr`i okolu 110 ostatoci od amino kiselini i me|uveri`nadisulfidna vrska koja formira jamka od 60 amino kiselini. Amino-krajnitedomeni, koi odgovoraat na V podra~ja, se vrzuvaat za protivgen.Efektornite funkcii se posredeni od drugite domeni. (b) Mi i epsilonte{kite verigi sodr`at dopolnitelen domen koj go zamenuva zglobnotote{kite verigi sodr`at dopolnitelen domen koj go zamenuva zglobnotopodra~je.
17

Kristalografskata analiza so Iks zraci poka`a deka imunoglobulinskite domeni se zdipleni vokarakteristi~ni kompaktni strukturi nare~eni imunoglobulinska dipla. Ovaa struktura se sostoi odr r ru ur r u u d ru ur d"sendvi~" od dve postaveni plo~i, od koi sekoja sodr`i protivparalelni beta lenti od aminokiselini, koi se spoeni so jamki so razli~na dol`inaRendgenskata kristolografska analiza otkrideka imunoglobulinskite domeni se obvitkani vo karaktersti~no mala struktura nare~enaimunoglobulinski nabor. Ovaa struktura sodr`i üsendvi~û od dve (slika 4-11). Beta lentite voplo~ata se stabiliziraat so vodorodni vrski pome|u amino (-NH) grupite od ednata plo~a sokarbonilnite grupi od sosednnata plo~a (vidi slika 4-8). Beta plo~kite se karakteriziraat sonaizmeni~ni hidrofobni i hidrofilni amino kiselini ~ii {to strani~ni verigi se organiziraniprependikularno na povr{inata na plo~kata, odnosno hidrofobnite amino kiselini se postaveni konvntare od sendvi~ot, a hidrofilnite amino kiselini gledaat kon nadvor. Dvete beta plo~ki voimunoglobulinskata dipla se stabilizirani preku hidrofobni soodnosi pome|u niv i prekukonzervirana disulfidna vrska.I j j |Iako promenlivite i postojanite domeni imaat sli~na struktura, postojat diskretni razliki pome|univ. Sekvencata na V domen e nezna~itelno podolga od onaa na K domen i sodr`i dopolnitelen parbeta lenti vo strukturata na beta plo~kata, isto kako i dopolnitelna sekvenca na jamka koja gopovrzuva ovoj par od beta lentite (vidi slika 4-11).Osnovnata struktura na imunoglobulinskata dipla pridonesuva za ~etvrtinata struktura naimunoglobulinite preku olesnuvawe nekovalentnite soodnosi pome|u domenite po povr{inite nabeta - plo~kite (vidi slika 4-8). Me|udejstvoto sozdava vrski pome|u identi~nite domeni (na pr. (KT2/KT2, KT3/ KT3 i KT4/ KT4) i pome|u neidenti~nite domeni (VT /VL i KT1/ KL). Strukturata naimunoglobulinskite dipli isto taka ovozmo`uva promenlivi dol`ini i sekvencini od aminokiselini koi ja sozdavaat jamkata koja gi povrzuva beta plo~kite. Kako {to objasnuva slednotopoglavje, del od povrzuva~kite sekvenci (loop sequences) na VT i VL domenite sodr`at promenliviamino kiselini i ja so~inuvaat protivgen vrzuva~koto mesto od molekulataamino kiselini i ja so~inuvaat protivgen vrzuva~koto mesto od molekulata.(slika 4-11 (a) Dijagram na edna imunoglobulinska lesna veriga {to prika`uva imunoglobulinzdiplena struktura na nejzinite varijabilni i konstantni domeni. Dvete beta zdipleni plo~i od sekojdomen se dr`at zaedno preku hidrofobni vrski i konzervirana disulfidna vrska. Beta plo~ite koi jaso~inuvaat sekoja lenta se prik`ani vo razli~ni boi. Amino kiselinskata sekvenca vo trite jamki odsekoj promenliv domen poka`uva zna~itelna varijabilnost, ovie hipervarijabilni podra~ja (sino) goso~inuvaat protivgen vrzuva~koto mesto. Hipervarijabilnite podra~ja obi~no se nare~eni KOP(komplementarno odreduva~ki podra~ja). Domenite od te{kite verigi ja imaat ista karakteristi~nastruktura. (b) beta zdiplenite plo~i se otvoreni za da ja otkrijat vrskata so oddelnite beta lenti ipovrzuva~kite jamki. Zabele`ete deka varijabilniot domen sodr`i 2 beta plo~i pove}e odkonstantniot domen. (referenca)
18
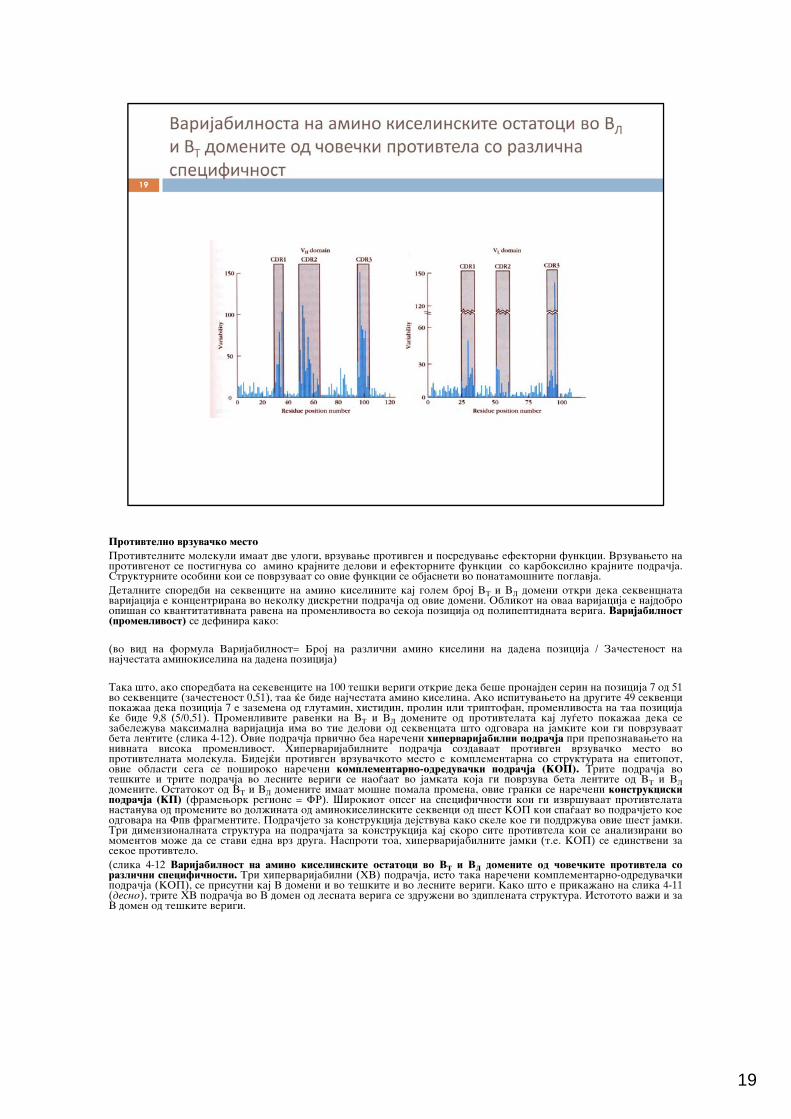
Protivtelno vrzuva~ko mestoProtivtelnite molekuli imaat dve ulogi, vrzuvawe protivgen i posreduvawe efektorni funkcii. Vrzuvaweto naProtivtelnite molekuli imaat dve ulogi, vrzuvawe protivgen i posreduvawe efektorni funkcii. Vrzuvaweto naprotivgenot se postignuva so amino krajnite delovi i efektornite funkcii so karboksilno krajnite podra~ja.Strukturnite osobini koi se povrzuvaat so ovie funkcii se objasneti vo ponatamo{nite poglavja.Detalnite sporedbi na sekvencite na amino kiselinite kaj golem broj VT i VL domeni otkri deka sekvencnatavarijacija e koncentrirana vo nekolku diskretni podra~ja od ovie domeni. Oblikot na ovaa varijacija e najdobroopi{an so kvantitativnata ravena na promenlivosta vo sekoja pozicija od polipeptidnata veriga. Varijabilnost(promenlivost) se definira kako:
(vo vid na formula Varijabilnost= Broj na razli~ni amino kiselini na dadena pozicija / Za~estenost nanaj~estata aminokiselina na dadena pozicija)
Taka {to, ako sporedbata na sekevencite na 100 te{ki verigi otkrie deka be{e pronajden serin na pozicija 7 od 51r r r r j r jvo sekvencite (za~estenost 0,51), taa }e bide naj~estata amino kiselina. Ako ispituvaweto na drugite 49 sekvencipoka`aa deka pozicija 7 e zazemena od glutamin, histidin, prolin ili triptofan, promenlivosta na taa pozicija}e bide 9,8 (5/0,51). Promenlivite ravenki na VT i VL domenite od protivtelata kaj lu|eto poka`aa deka sezabele`uva maksimalna varijacija ima vo tie delovi od sekvencata {to odgovara na jamkite koi gi povrzuvaatbeta lentite (slika 4-12). Ovie podra~ja prvi~no bea nare~eni hipervarijabilni podra~ja pri prepoznavaweto nanivnata visoka promenlivost. Hipervarijabilnite podra~ja sozdavaat protivgen vrzuva~ko mesto voprotivtelnata molekula. Bidej}i protivgen vrzuva~koto mesto e komplementarna so strukturata na epitopot,ovie oblasti sega se po{iroko nare~eni komplementarno-odreduva~ki podra~ja (KOP). Trite podra~ja vote{kite i trite podra~ja vo lesnite verigi se nao|aat vo jamkata koja gi povrzuva beta lentite od VT i VLdomenite. Ostatokot od VT i VL domenite imaat mo{ne pomala promena, ovie granki se nare~eni konstrukciskipodra~ja (KP) (framework regions = FR). [irokiot opseg na specifi~nosti koi gi izvr{uvaat protivtelatanastanuva od promenite vo dol`inata od aminokiselinskite sekvenci od {est KOP koi spa|aat vo podra~jeto koeodgovara na Fpv fragmentite. Podra~jeto za konstrukcija dejstvuva kako skele koe gi poddr`uva ovie {est jamki.T j j jTri dimenzionalnata struktura na podra~jata za konstrukcija kaj skoro site protivtela koi se analizirani vomomentov mo`e da se stavi edna vrz druga. Nasproti toa, hipervarijabilnite jamki (t.e. KOP) se edinstveni zasekoe protivtelo.(slika 4-12 Varijabilnost na amino kiselinskite ostatoci vo VT i VL domenite od ~ove~kite protivtela sorazli~ni specifi~nosti. Tri hipervarijabilni (HV) podra~ja, isto taka nare~eni komplementarno-odreduva~kipodra~ja (KOP), se prisutni kaj V domeni i vo te{kite i vo lesnite verigi. Kako {to e prika`ano na slika 4-11(desno), trite HV podra~ja vo V domen od lesnata veriga se zdru`eni vo zdiplenata struktura. Istototo va`i i zaV domen od te{kite verigi.
19

KOP vrzuva protivgen
Naodot deka KOP se protivgen vrzuva~ki podra~ja od protivtelata se potvrdi direktno so visokorazdelna kristalografija so Iks zraci na kompleksot protivgen-protivtelo. Kristalografskataanaliza e kompletirana za mnogu Fpv fragmenti od monoklonskite protivtela vo kompleks bilo sogolemi globularni proteinski protivegni ili so mali protivgeni, vklu~uvaj}i jaglehidrati,nukleinski kiselini, pepetidi i mali hapteni. Dopolnitelno, dobieni se celosnite strukturi zanekolku neo{teteni monoklonski protivtela. Rendgenskata difrakciska analiza na protivgen-protivtelnite kompleksi poka`a deka nekolku KOP kontaktiraat so protivgenot, a se vide i odredenbroj kompleksi kade site {est KOP kontaktiraa so protivgenot. Vo osnova, pove}e ostatoci od KOPna te{kite verigi kontaktiraat so protivgenot otkolku kaj KOP na lesnite verigi. So drugizborovi, VT domenot ~esto pridonesuva mnogu pove}e za vrzuvaweto protivgen otkolku VL domenot.Dominantnata uloga na te{kata veriga vo vrzuvaweto protivgen be{e prika`ana vo ispituvawe voDo a a a ulo a a e a a er a o rzu a e o ro e be e r a a a o s u a e okoe edna te{ka veriga specifi~na za glikoproteinsaki protivgen za ~ove~kiot imunonedostato~envirus (HIV) se kombinira{e so razli~ni lesni verigi so razli~na protivgna specifi~nost (vidipoglavje 20). Site hibridni protivtela se vrzuvaat za HIV glikoproteinskiot protivgen, {touka`uva na faktot deka samo te{kata veriga e dovolna za da pottikne specifi~nost. Sepak, ne trebada se zaklu~i deka lesnata veriga e nezna~itelna, vo nekoi protivgen-protivtelni reakcii, lesnataveriga ima pozna~aen pridones.(slika 4-13 Kompjuterska simulacija na soodnos pome|u protivteloto i protivgenot od virusotinfluenca, globularen protein. (a) Protivgenot (`olto) e prika`an kako stapuva vo soodnos soprotivtelnata molekulata, varijabilnoto podra~je od te{kata veriga e vo crveno, a promenlivotopodra~je na lesnata veriga e vo sino. (b) Komplementarnosta na dvete molekuli se otkriva prekuoddeluvawe protivgenot od protivteloto so 8 Angtremi.Prirodata na kontaknata povr{ina pome|u protivteloto i protivgenot e zna~itelno prou~ena sokompjuterski sozdadenata slika na soodnosot pome|u protivgenot i protivteloto od virusot nainfluenca (slika 4-13). Kontaktite pome|u golemiot globularen proteinski protivgen i protivtelose ostvaruvaat na {iroka, ~esto plitka, branovidna povr{ina. Vo oblasta na kontaktot,ispaknatinite ili vdlabnatinite na protivgenot verojatno se sovpa|aat so komplementnitevdlabnatini ili ispaknatini vo protivtelno vrzuva~koto mesto, sozdadeno od strana na KOP. Voslu~ajot na dobro proueniot lizozimski/protivlizozimski sistem, kristalografskite prou~uvawapoka`aa deka povr{inskite oblasti na soodnost se dosta golemi, i se protegaat od okolu 650Angstremi2 do pove}e od 900 Angstremi2. Vo ovaa oblast, ostatoci od 15 do 22 amino kiselini voprotivteloto kontaktiraat so sli~en broj ostatoci od proteinskiot protivgen.
20

Na slika 4-14, gledame deka globularnite proteinski protivgeni i malipeptidni ili haptenski protivgeni stapuvaat vo soodnos so protivteloto narazli~ni na~ini. Obi~no, proteinskite protivgeni so golemi povr{ini sevo soodnos so golemi oblasti od protivtelo vrzuva~koto mesto (slika 4-14a).Pomalite protivgeni kako {to se peptidite zazemaat pomalku prostor imo`e da se smestat vo pregradite ili procepot od vrzuva~koto mesto (slika4-14b). Soodnosot na pepetidite dobieni so HIV proteaza i Fpv fragmentotod protivproteaznoto protivtelo detalno ni ja prika`uva bliskatakomplementarnost pome|u protivteloto i maliot protivgen (slika 4-14v).
(slika 4-14 Protivtelno vrzuva~ki mesta. Vo osnova, vrzuva~kite mesta zamalite molekuli pretstavuvaat dlaboki xepovi, dodeka vrzuva~kite mesta zagolemite molekuli se poplitki, pobranovidni povr{ini. (a, b) Modeli napromenlivite domeni na {est Fpv fragmenti so nivnite protivgenvrzuva~ki podra~ja prika`ani violetovo Trite protivtela na slika (a) sevrzuva~ki podra~ja prika`ani violetovo. Trite protivtela na slika (a) sespecifi~ni za lizozim, golem globularen protein. Trite protivtela naslika (b) se specifi~ni za pomali molekuli ili dosta pomali segmenti odmakromolekulite: McPC603 za fosfoholin, BV04 za mal segment odednoveri`na DNK i 17/9 za peptid od hemaglutininot, proteinska obvivkana influenca virusot. (v) Detalen prikaz na kompleksot pome|u mal peptiddobien so HIV proteaza i Fpv fragment od protivproteaznoto protivtelo.
21

(slika 4-15 Konformaciski promena pri vrzuvaweto na protivgen so protivtelo.Prika`an e istiot kompleks kako na slika 4-14v pome|u peptidot dobien od HIVPrika`an e istiot kompleks kako na slika 4-14v, pome|u peptidot dobien od HIVproteazata i Fpv fragmentot od protivproteaznoto protivtelo, pri {to se pravisporedba na strukturata na Fpv pred i po vrzuvaweto na peptidot. Crvenata linijaja prika`uva strukturata na na Fpv fragmentot pred se vrze so peptidot, a sinatalinija ja poka`uva negovata struktura po negovoto vrzuvawe. Doa|a do zna~ajnikonformaciski promeni pri sozdavaweto kaj KOP od Fpv pri vrzuvweto naprotivgenot. Tie se posebno naglaseni kaj KOP1 (L1) od lesnata veriga i kaj KOP3(T3) od te{kata veriga.
Konformaciskite promeni mo`e da bidat pottiknati od vrzuvaweto protivgenKonformaciskite promeni mo`e da bidat pottiknati od vrzuvaweto protivgen
Kako {to se kompletiraa pove}e rendgenski kristalografski analizi na Fpvfragmentite, stana jasno deka vo odredeni slu~ai, vrzuvaweto na protivgenotpottiknuva konformaciski promeni kaj protivteloto, protivgenot ili i dvete.Izvonreden primer za konformaciska promena se gleda kaj sozdavaweto kompleksipome|u Fpv fragmentite i niviot peptiden epitop prika`an na slika 4-15. Privrzuvaweto kaj Fpv se pojavuvaat zna~ajni strukturni promeni. Podra~jeto KOP1od lesnata veriga se dvi`i so 1 Angstrem, a KOP3 od te{kata veriga se dvi`i so2 7 A T2,7 Angstremi. Taka {to, dopolnitelni na promenata na dol`inata i na aminokiselinskiot sostav na KOP jamkite, sposobnosta na ovie jamki zna~ajno da jamenuvaat konformacijata pri vrzuvaweto protivgen im ovozmo`uva naprotivtelata da zazemat oblik koj e poefektivno komplementaren od onoj nanivnite epitopi.Kako {to ve}e poka`avme, konformaciskite promeni posle protivgenotovrzuvawe ne treba da se ograni~at na protivteloto. Iako ne e prika`ano na slika4-15, konformacijata na proteazniot pepetid vrzan za Fpv ne poka`uva strukturnasli~nost so soodvetniot epitop od prirodnata HIV proteaza. Se predlo`i deka
HIVpopre~uvaweto na proteaznata aktivnost od strana na protiv-HIV proteaznotoprotivtelo e rezultat na negovoto iskrivuvawe na enzimskata prirodnakonformacija.
22

Domeni vo konstantno podra~je
I b j b f jImunoglobulinskite domeni od konstantnoto podra~je u~estvuvaat vo razni biolo{ki funkcii koi se odredeni so amino kiselinskata sekvenca vo sekoj domen.
KT1 i KL domeni
Domenite KT1 i KL slu`at za da gi prodol`at grankite na protivtelnata molekula, pri toa olesnuvaj}i go me|udejstvoto so protivgenot i zgolemuvaj}i ja maksimalnata rotacija na grankite (racete) naFpv. Dopolnitelno, me|uveri`nata disulfidna vrska pome|u ovie domeni od konstantnoto podra~je pomaga da se dr`at zaedno VT i VL (vidi slika 4-10). Domenite KT1 i KL mo`e isto taka da pridonesat zaprotivtelnoto razli~ie ovozmo`uvaj}i poslu~ajni vrzuvawa pome|u domenite VT i VL otkolku ako se pojavi povrzanost samo od soodnosot pome|u VT i VL. Ovie zabele{ki imaat zna~ajno vlijanie vrzgradeweto {iroka paleta na protivtela (razli~ie ili repertoar). Kako {to }e ni poka`e poglavje 5, preureduvawata po slu~aen izbor na imnoglobulinskite geni sozdava unikatni VT i VL sekvenci zate{kite i za lesnite verigi prika`ani vrz sekoj B limfocit, potoa povrzuvaweto na sekvencite na VT i VL sozdava unikatno protivgen vrzuva~ko mesto. Prisustvoto na domenite KT1 i KL go zgolemuvabrojot na mo`ni stabilni interakcii pome|u VT i VL, na toj na~in pridonesuvaat za celosnata raznovidnost na protivtelnite molekuli koi se prika`ani kaj `ivotnite.
Zglobno podra~je
Gama, delta i alfa te{kite verigi sodr`at prodol`ena peptidna sekvenca pome|u domenite KT1 i KT2 koi nemaat homologija so drugite domeni (vidi slika 4-10). Ova podra~je, nare~eno zglobno podra~je,e bogato so ostatoci od prolin i e podvi`no, pri {to na IgG, IgD i IgA im dava segmentna fleksibilnost. Kako rezultat na toa, dvete race od Fpv mo`at da zazemat razli~ni agli eden sprema drug kogase vrzuva protivgenot. Ovaa podvi`nost na zglobnoto podra~je mo`e da se prika`e so elektronska mikrografija na kompleksite protivgen-protivtelo. Na primer, koga molekulata koja sodr`i dve grupidinitrofenol (DNF) reagira so protiv-DNF protivtelo a kompleksot e zadr`an vo mre`a, se boi negativno i se nabquduva so elektronski mirkoskop, se gledaat golemi kompleksi (na primer, dimeri,trimeri, tetrameri). Agolot pome|u grankite na protivtelnata molekula so oblik na U se razlikuva kaj razli~ni kompleksi, {to ja otslikuva podvi`nosta na zglobnoto podra~je (slika 4-16).r r , r r ) u r r u r u j r , j u d dr j ( )
(slika 4-16 Eksperimentalen prikaz na podvi`nosta na zglobnoto podra~je kaj protivtelna molekula. (a) Haptenot vo koj dve dinitrofenilni (DNF) grupi se povrzani so kratki povrzuva~ki prostornigrupi reagiraat so protiv-DNF protivtelata za da formiraat trimeri, tetrameri i drugi pogolemi kompleksi protivgen-protivtelo. (b) Pri eelektronska mikrografija na negativno oboeniot preparatod ovie soedinenija, jasno se gledat dve triagolni trimerni strukturi. Protivtelniot protein se gleda kako svetla struktura nasproti zadninata elektronski gustata podloga. Poradi podvi`nosta nazglobnoto podra~je, agolot pome|u grankite na protivtelnata molekulata se menuva. (referenca)
Dvete ispap~eni amino kiselini vo zglobnoto podra~je se prolin i cistin. Golemiot broj prolinski ostatoci vo zglobnoto podra~je mu dava prodol`ena polipeptidna konformacija, so {to go pravizna~itelno ranliv za razgraduvawe so proteoliti~ni enzimi, poradi toa ova podra~je se razgraduva so papain ili pepsin (slika 4-7). Cisteinskite ostatoci sozdavaat me|uveri`ni disulfidni vrski koigi dr`at spoeni dvete te{ki verigi. Brojot na me|uveri`ni disulfidni vrski vo zglobnoto podra~je zna~itelno varira pome|u razli~ni klasi protivtela i pome|u vidovi. Iako mi i epsilon verigitenemaat zglobno podra~je, tie imaat dopolnitelen domen od 110 amino kiselini (KT2/KT2) koj ima zglobnovidna osobina.
Drugi domeni vo konstantnoto podra~je
Kako {to vidovme prethodno, te{kite verigi IgA, IgD i IgG sodr`at tri domeni vo konstantnoti i vo zglobnoto podra~je. dodeka te{kite verigi od IgE i IgM sodr`at ~etiri konstantni domeni nonemaat zglobno podra~je. Soodvetnite domeni vo dvete grupi se slednive:
(tabela)( )
IgA, IgD, IgG IgE, IgM
KT1/KT1 KT1/KT1
Zglobno podra~je KT2/KT2
KT2/KT2 KT3/KT3
KT3/KT3 KT4/KT4
Iako domenite KT2/KT2 vo IgE i IgM ja zazemaat istata pozicija vo polipeptidnite verigi kako i zglobnoto podra~je kaj drugite imunoglobulinski klasi, funkcijata na ovoj dopolinitelen domen s#u{tene e utvrdena.
Rendgenskata kristalografska analiza otkri deka dvata domeni KT2 na IgA, IgD i IgG (i domenite KT3 vo IgE i IgM) se razdvoeni so oligosaharidni strani~ni verigi koi spre~uvaat kontakt pome|upritisnatite domeni (vidi slika 4-9b). Kako rezultat na toa, ovie dva globularni domeni se popristapni vo vodenata sredina od drugite. Ovaa pristapnost pretstavuva eden od elementite koipridonesuvaat za biolo{kata aktivnost na ovie domeni pri aktiviraweto komplementnite delovi od strana na IgG i IgM.
Karboksilno krajniot domen vo IgA, IgD i IgG e ozna~en KT3/KT3, a vo IgE i IgM e ozna~en KT4/KT4. Ovie pet klasi na protivtela i nivnite potklasi mo`e da se nare~at ili izla~en imunoglobin (iIg)ili membranski vrzan imunoglobulin (mIg). Karboksilno krajniot domen kaj izla~enite imunoglobilini se razlikuva vo strukturata i vo funkcijata od soodvetnite domeni vo membranski vrzaniteimunoglobulini. Izla~enite imunoglobulini imaat hidrofilna amino kiselinska sekvenca so razli~na dol`ina na kaboksilniot kraj. Funkciite na ovoj domen kaj razli~ni klasi izla~eno protivtelo }egi razgledame podocna. Vo membranski vrzaniot imunoglobulin, karboksilno krajniot domen sodr`i tri podra~ja:
Vonkleto~na hidrofilna prostornaû sekvenca sostavena od 26 amino kiselinski ostatociVonkleto~na hidrofilna üprostorna sekvenca sostavena od 26 amino kiselinski ostatoci
Hidrofobna transmembranska sekvenca
Kratka citoplazmatska opa{ka
Dol`inata na transmembranskata sekvenca e postojana me|u site imunoglobulinski izotipovi, dodeka dol`inite na vonkleto~nata prostorna sekvenca i citoplazmatskata opa{ka variraat.
B kletkite la~at razli~ni klasi na mIg pri razli~ni fazi vo razvojot. Nezrelite B kletki, nare~eni pred-B kletki, la~at samo mIgM. Podocna pri zreeweto, se pojavuva mIgD i zaedno so IgM seprika`uvaat vrz povr{inata od zrelite B kletki pred tie da se aktiviraat so protivgen. Pamte~kata B kletka mo`e da prika`e mIgG, mIgA ili mIgE. Duri koga istovremeno vrz edna kletka seprika`ani razli~ni klasi, protivgenata specifi~nost za site membranski protivtelni molekuli prika`ani vrz edna kletka e identi~na, taka {to sekoja protivtelna molekula se vrzuva za istiotepitop. Genetskiot mehanizam koj ovozmo`uva edna B kletka da prika`uva pove}e imunoglobulinski izotipovi pri {to site se so ista protivgena spencifi~nost e objasnet vo poglavje 5.
23

Protivtelno ostvareni efektorni funkcii
Pokraj vrzuvaweto protivgen protivtelata u~estvuvaat vo {irok obseg drugi biolo{ki aktivnosti Koga se razgleduva ulogata na protivteloto vo odbrana protiv bolest mora da sePokraj vrzuvaweto protivgen, protivtelata u~estvuvaat vo {irok obseg drugi biolo{ki aktivnosti. Koga se razgleduva ulogata na protivteloto vo odbrana protiv bolest, mora da sezapomni deka protivtelata vo osnova ne ubivaat ili ostranuvaat patogeni samo so vrzuvawe za niv. Za da bidat efikasni protiv patogenite, protivtelata ne treba samo da prepoznavaatprotivgen, tuku isto taka i da pottiknuvaat reakcii - efektorni funkcii - koi doveduvaat do otstranuvawe protivgenot i smrt na patogenot. Iako varijabilnite podra~ja naprotivteloto se edinstvenite sredstva za vrzuvawe za protivgenot, konstatnoto podra~je od te{kite verigi (KT) e odgovorno za mno{tvo soodnosi so drugi proteini, kletki i tkiva{to rezultira so efektorni funkcii na humoralniot odgovor.Bidej}i ovie efektorni funkcii se javuvaat kako rezultat od soodnosot pome|u konstantnite podra~ja od te{kite verigi i drugi serumski proteini ili kleto~ni membranskireceptori, site klasi od imunoglobulinite nemaat isti funkcionalni osobini. Ovde e pretstaven pregled na ~etirite glavni efektorni funkcii ostvareni od strana na konstantnitedomenit. Petatta funkcija koja e edinstvena za mastocitite, ezinofilite i bazofilite, }e ja objasnime podocna.
Opsonizacijata e pottiknata so protivteloto
Opsonizacijata, pottiknuvaweto fagocitoza so protivgeni kaj makrofagite i neutrofilite, e zna~aen faktor za protivbakteriska odbrana. Proteinskite molekuli nare~eni Fkreceptori (FkR), koi mo`at da se vrzat za konstantnot podra~je od Ig molekuli, se prisutni vrz povr{inite od makrofagite i neutrofilite kako i kaj drugi kletki koi ne sevklu~eni vo fagocitozata. Vrzuvaweto na fagocitnite Fk receptori od strana na nekolku protivtelni molekuli vo kompleks so istiot protivgen, kako {to se bakteriskite yidovi,sozdava soodnos koj go obezbeduva patogenot vrz fagocitnata membrana. Ova vkrsteno povrzuvawe (cross-linking) na FkR preku vrzuvawe za niza od protivtelni Fk podra~japottiknuva signalno prenosna pateka {to rezultira so fagocitoza na kompleksot protivgen-protivtelo. Vo fagocitot, patogenot stanuva cel na razli~ni destruktivni procesi koivklu~uvaat enzimsko razgraduvawe, oksidacisko o{tetuvawe i membransko-naru{uva~ki efekti na protivbakteriskite peptidi.
Protivtelata aktiviraat komplementr r
IgM i, kaj lu|eto, pove}eto potklasi na IgG mo`at da aktiviraat kolekcija na serumski glikoproteini nare~eni komplementen sistem. Komplementot vklu~uva odredena grupaproteini koi mo`at da gi izdup~at kleto~nite membrani. Zna~aen strani~en produkt od komplementno aktivirana pateka e proteinskiot del nare~en K3b, koj se vrzuva nespecifi~noza kleto~nite i protivgen protivtelnite kompleksi blisku do mestoto kade bil aktiviran komplementot. Mnogu vidovi kletki - na primer, crvenite krvni kletki i makrofagite -imaat receptori za K3b i na toj na~in se vrzuvaat za kletki ili kompleksi kon koi e zalepen K3b. Vrzuvaweto na zalepeniot K3b od strana na makrofagi vodi kon fagocitoza nakletkite ili molekulskite soedinenija prika~eni za K3b. Vrzuvaweto na protivgen protivtelnite kompleksi za receptorite K3b od crevenite krvni zrnca mu ovozmo`uva naeritrocitot da gi prenese kompleksite vo crniot drob ili slezinata, kade makrofagite koi prestojuvaat tamu gi ostranuvaat bez uni{tuvawe crvenite krvni zrnca. Soodnosot pome|uprotivteloto i komplementniot sistem e zna~aen za deaktiviraweto i otstranuvaweto protivgenite i ubivaweto na patogenite. Procesot na aktivirawe komplement e detalnoobjasnet vo poglavje 7.
Protivtelno zavisna kleto~no posreduvana citotoksi~nost (PZKC) gi ubiva kletkite
Povrzuvaweto na protivteloto koi za celnite kletki (na pr. virusno zarazeni kletki od doma}inot) so Fk receptorite od odreden broj kletki, posebno kletkite prirodni ubijci(KPU), mo`e da gi naso~at citotoksi~nite aktivnosti od efektornata kletka protiv celnata kletka. Pri ovoj proces, nare~en Protivtelno zavisna kleto~no posreduvanacitotoksi~nost (PZKC), protivteloto se odnesuva kako novo steknat receptor koj $ ovozmo`uva na napa|a~kata kletka da ja prepoznae i ubie celnata kletka. Fenomenot PZKC eobjasnet vo poglavje 14.
Nekoi protivtela mo`at da go preminat epitelniot sloj so transcitozar d r j r c
Dostavuvawe protivtelo kon mukoznata povr{ina od respiratorniot, gastrointestinalniot i urogenitalniot trakt, isto kako negoviot prenos kon mlekoto vo gradite, bara dvi`eweimunoglobulinot niz epitelnite sloevi, proces nare~en transcitoza. Kapacitetot za transportirawe zavisi od osobinite na konstantnoto podra~je. Kaj lu|eto i gluvcite, IgA eglavnata protivtelna klasa koja pretrpuva takva transcitoza, iako i IgM mo`e da se transportira do mukoznata povr{ina. Odredeni vidovi cica~i, kako {to se lu|eto i gluvcite,isto taka prenesuvaat zna~itelni koli~ini na pove}eto potklasi na IgG od majakata do embrionot. Bidej}i maj~iniot i embrionskiot cirkulatoren sistem se oddelni, protivtelotomora da se prenese niz placentalnoto tkivo koe gi oddeluva majkata i embrionot. Kaj lu|eto, ovoj prenos se slu~uva vo tretiot trimester od bremenosta. Zna~ajna posledica e toa {torazvojot na fetusot prima primerok od kolekcijata protivtela od majkata kako za{titen dar protiv patogenite. [to se odnesuva do drugite efektorni funkcii objasneti ovde,kapacitetot za pretrpuvawe transplacentno prenesuvawe zavisi od svojstvata na konstantnoto podra~je na protivtelna molekulata.Prenesuvawe IgG od majkata vo embrionot e forma na pasivna imunizacija, {to pretstavuva dobivawe imunitet preku primawe preformirani protivtela namesto so aktivnosozdavawe protivtela po izlo`uvawe kon protivgenot. Sposobnosta da se prenesat protivtelata od edno lice na drugo so prenoe na protivtelata e osnova za protivtelnata terapija,zna~ajna i {iroko upotrebuvana medicinska postapka (vidi klini~ki fokus, strana 98).
24

Protivtelni klasi i biolo{ki aktivnosti
Razli~nite imunoglobulinski izotipovi i klasi prethodno bea kratko spomenati. Sekoja klasa se razlikuva po unikatna sekvencaod amino kiselini vo konstantnite podra~ja od te{kite verigi koi pottiknuvaat specifi~ni strukturni i funkcionalni osobini.Vo ovoj del, podetalno se objasneti strukturata i efektornite funkcii na sekoja klasa. Molekulskite osobini i biolo{kiteaktivnosti na imunoglobulinskite klasi se opi{ani vo tabela 4-4. Strukturite na pette glavni klasi {ematski se prika`ani naslika 4-17.Imunoglobulin M (IgM)
Na IgM otpa|a 5% do 10% od vkupniot serumski imunoglobulin, so prose~na serumska koncentracija od 1,5 mg/ml. MonomerniotIgM (180000 Da) vrz B kletkite se prika`uva kako membranski vrzano protivtelo. IgM go la~at plazma kletkite kako pentamer vokoi pet monomerni edinici se spoeni zaedno so disulfidni vrski koi gi povrzuvaat karboksilno krajnite domeni od te{kataveriga (Kmi4/Kmi4) i nivnite Kmi3/Kmi3 domeni (vidi slika 4-17e). Pette monometrni podedinici se naredeni so nivnite Fkpodra~ja vo centarot na pentamerot i 10 protivgen vrzuva~ki mesta na periferija od molekulata. Sekoj pentamer sodr`idopolnitelen polipeptid povrzan za Fk nare~en J (vrzuva~ka) veriga, koja preku disulfidna vrska e povrzana so karboksilnokrajniot ostatok od cisteinot na dve od desette mi verigi J verigata e potrebna za polimerizacija na monomerite pri sozdavawetokrajniot ostatok od cisteinot na dve od desette mi verigi. J verigata e potrebna za polimerizacija na monomerite pri sozdavawetopentametren IgM, koj se dodava neposredno pred la~eweto na pentamerot.IgM e prvata klasa koja se sozdava vo prvi~niot odgovor kon protivgenot i isto taka prviot imunoglobolin koj se sintetizira vonovoroden~eto. Poradi negovata pentameri~na struktura so 10 protivgen vrzni mesta, serumskiot IgM ima povisoka valentnost oddrugite izotipovi. IgM molekulata mo`e da vrze 10 mali haptenski molekuli. Sepak, poradi steri~nata hidrantnost(navodenetost), istovremeno mo`e da se vrzat samo pet ili pomalku molekuli od pogolemite protivgeni. Poradi negovata visokavalencija, pentamerniot IgM e poefikasen od drugite izotipovi pri vrzuvaweto protivgeni so mnogu povtoruva~ki izotipovikako {to se virusnite ~estici i crvenite krvni zrnca (CKZ). Na primer, koga CKZ se inkubirani so specifi~no protivtelo, tiese grupiraat zaedno vo golemi grupi pri proces nare~en aglutinacija. Za postignuvawe na istoto nivo na aglutinacija potrebni seod 100 do 1000 pati pove}e molekuli od IgG i IgM. Sli~en fenomen se pojavuva so virusnite ~estici: za neutralizirawe virusnataaktivnost potrebni se pomalku IgM i IgG. Isto taka IgM e poefikasen otkolku IgG pri aktivirawe komplementot.Aktiviraweto na komplementot bara najmalku dva bliski Fk podra~ja, a pentamernata struktura na edna molekula od IgM goispolnuva ova barawe.
(slika 4 17Osnovni strukturi na pette klasi izla~eni protivtela(slika 4-17Osnovni strukturi na pette klasi izla~eni protivtela.Lesnite verigi se prika`ani vo nijansi od temno-rozovo, disulfidnite vrski se prika`ani kako tenki crni linii. Zabele`etedeka IgG, IgA i IgD te{kite verigi (sino, portokalovo i zeleno) sodr`at ~etiri domeni i zglobno podra~je, dodeka IgM i IgEte{kite verigi (violeotovo i `olto) sodr`at pet domeni no nemaat zglobno podra~je. Polimernite formi na IgM i IgA sodr`atpolipeptid. nare~en J veriga, koj e povrzan so dve disulfidni vrski za Fk podra~ja od dva razli~ni monomeri. Serumskiot IgM esekoga{ pentamer. Naj~esto serum IgA postoi kako monomer, iako ponekoga{ se prisutni dimeri, trimeri i duri tetrameri. Naovie sliki ne se prika`ani me|uveri`nite disulfidni vrski koi gi povrzuvaat lesnite i te{kite verigi (vidi slika 4-6).
Poradi negovata golema, IgM ne difundira dobro i zatoa se sretnuva vo mnogu niski koncentracii vo me|ukleto~nite tkivnite~nosti. Prisustvoto na J verigata mu ovozmo`uva na IgM da se vrze za receptorite od sekretornite kletki, koi go prenesuvaatniz epitelnite obvivki za da izlezat vo nadvore{nite skreti koi gi mijat mukoznite membrani. Iako IgA e glavniot izotip koj sesretnuva pri ovie sekreti, IgM igra zna~ajna pomo{na uloga kako la~eben imunoglobulin.
25

Imunoglobulin G (IgG)
IgG, najobilnata klasa vo serumot, so~inuva okolu 80% od vkupnite serumskiimunoglobulin. Molekulata IgG sodr`i dve gama te{ki verigi i dve κ ili dvelambda lesni verigi (vidi slika 4-17a). Postojat ~etiri humani potklasi na IgG,koi se razlikuvaat spored sekvencata konstantnoto podra~je od gama-verigata i seobele`uvaat spored namaluvaweto na nivnata koncentracija vo serumot: IgG1,IgG2, IgG3 i IgG4 (vidi tabela 4-3).Sekvencite na amino kiselinite koi gi razlikuvaat ~etirite potklasi na IgG i sekodiraat so razli~ni zarodi{ni KT geni, ~ii sekvenci od DNK se 90% do 95%r r r T Dhomologni. Strukturnite osobini koi gi razlikuvaat ovie potklasi edna od drugase goleminata na zglobnoto podra~je i brojot i pozicii na me|uveri`nitedisulfidni vrski pome|u te{kite verigi (slika 4-18). Malite razliki vo aminokiselinite pome|u potklasite na IgG vlijaat vrz biolo{kata aktivnost namolekulata:
IgG1, IgG3 i IgG4 lesno minuvaat niz placentata i igraat zna~jana uloga voza{titata na embrionskiot razvoj.IgG3 e najefikasniot komplementen aktivator koj sledi po IgG1 IgG2 e pomalkuIgG3 e najefikasniot komplementen aktivator, koj sledi po IgG1. IgG2 e pomalkuefikasen, a IgG4 ne mo`e voop{to da aktivira komplement.IgG1 i IgG3 se vrzuvaat so visok afinitet za Fk receptorite na fagocitnitekletki i na toj na~in posreduvaat opsonizacija. IgG4 ima prose~en afinitet za Fkreceptorite a IgG2 ima zna~itelno mal afinitet.
(slika 4-18 Osnovna struktura na ~etirite potklasi IgG kaj lu|eto, koi serazlikuvaat po brojot i rasporedot na me|uveri`nite disulfidni vrski (tenkicrni linii) koi gi povrzuvaat te{kite verigi. Zna~ajna karakteristika na IgG kajcrni linii) koi gi povrzuvaat te{kite verigi. Zna~ajna karakteristika na IgG kajlu|eto pretstavuvaat negovite 11 disulfidni vrski me|u verigite.
26

Imunoglobulin A (IgA)
Iako IgA so~inuva samo 10% do 15% od celiot imunoglobulin vo serumot, toj e dominantanata imunoglobulinska klasa vonadvore{nite sekreti (izla~oci) kako {to se dojkinoto mleko, plunkata, solzite i sluzta od bronhijalniot, genitourinarniot idegestivniot trakt. Vo serumot, IgA prvi~no postoi kako monomer, no polimernite formi (dimeri, trimeri i odredenitetrameri) ponekoga{ se gledaat, pri {to site sodr`at polipeptid so J veriga (vidi slika 4-17d). IgA od nadvore{nite la~ewa,nare~en sekretoren IgA, se sostoi od dimer ili tetramer, polipeptid so J veriga i polipeptidna veriga nare~ena sekretoren del(vidi slika 4-19a). Kako {to e objasneto podolu, sekretorniot del se dobiva od receptorot koj e odgovoren za prenesuvawetopolimeren IgA preku kleto~nite membrani. Polipeptidot so J veriga vo IgA e identi~en so onoj polipeptid koj se sretnuva kajpentamerniot IgM i ima sli~na funkcija pri olesnuvaweto polimerizacijata i na serumskiot IgA i na sekretorniot IgA.Sekretorniot del e polipeptid od 70000 Da koj e sozdaden od strana na epitelnite kletki vo mukoznata membrana. Toj se sostoi odpet imunoglobulinovidni domeni koj se vrzuva za domeni od Fk podra~jeto vo IgA dimerot. Ovoj soodnos se stabilizira sodisulfidna vrska pome|u pettiot domen od sekretorniot del i edna od verigite od dimerniot IgA.Dnevnoto proizvodstvo na sekretorni IgA e pogolemo otkolku proizvodstvoto na drugite klasi imunoglobilini. Plazma kletkitekoi la~at IgA se koncentrirani po povr{inata na mukoznata membrana. Dol` povr{inata na tenkoto crevo, na primer, ima pove}eod 2,5 h 1010 plazma kletki koi la~at IgA-broj koj iznenaduva~ki ja nadminuva celata populacija plazma kletki vo koseknata sr`,
f ! S j 5 15 I Alimfata i slezinata zaedno! Sekoj den, ~ovekot dostavuva od 5 g do 15 g sekretorenIgA vo mukoznite izla~oci.
(slika 4-19 Struktura i sozdavawe sekretoren IgA. (a) Sekretorniot IgA se sostoi od najmalku dve IgA molekuli, koi sekovalentno me|usebno povrzani preku J veriga i isto taka kovalentno se povrzani so sekretorniot del. Sekretorniot del sodr`ipet imunoglobulinovidni domeni koi se povrzani so dimeri~niot IgA so dislufidna vrska pome|u negoviot petti domen i eden odIgA te{kite verigi. (b) Sekretorniot IgA se formira za vreme na prenesuvaweto niz mukoznata membrana vo epitelnite kletki.Dimeri~niot IgA se vrzuva za poli-Ig receptorot vo bazolateralnata membrana od epitelnta kletka i e vnesen so receptorskiovozmo`ena endocitoza. Po prenesuvaweto na kompleksot receptor-IgA vrz povr{inata od lumenot, poli-Ig receptorot enzimskise rase~uva, osloboduvaj}i go sekretorniot del koj e vrzan za dimeri~niot IgA.
B kletkite koi sozdavaat IgA povlasteno se preseluvaat (domuvaat) vo subepitelnoto tkivo, kade se delat na plazma kletki koila~at IgA, koj cvrsto se vrzuva za receptorot na polimernite imunoglobulinski molekuli (slika 4-19b). Ovoj poli-Ig receptor seprika`uva vrz bazolateralnata povr{ina od pogolemiot del mukozen epitel (t.e., oblo`uvawe na digestivniot, respiratorniot igenitalniot trakt) i vrz `lezdeniot epitel od mle~nite, plunkovnite i solznite `lezdi. Po vruvaweto na polimerniot IgA zagenitalniot trakt) i vrz `lezdeniot epitel od mle~nite, plunkovnite i solznite `lezdi. Po vruvaweto na polimerniot IgA zapoli-Ig receptor, kompleksot receptor-IgA se prenesuva preku epitelnata bariera do lumenot. Prenesuvaweto na kompleksotreceptor-IgA vklu~uva receptorski ostvarena endocitoza na obblo`eni vdlabnatini i naso~eno prenesuvawe na vezikulite nizepitelnata kletka do luminalnata membrana, kade vezikulata se fuzira so plazminata membrana. Potoa, poli-Ig receptorotenzimski se rase~uva od membranata i stanuva sekretorendel, koj se vrzuva i se osloboduva zaedno so polimerniot IgA vomukoznite izla~oci. Sekretorniot del gi maskira oblastite koi se osetlivi na proteazno se~ewe vo zglobnoto podra~je odsekretorniot IgA, ovozmo`uvaj}i i na polimernata molekula da opstoi podolgo vo mukoznata sredina bogata so proteazi {to nebi bilo vozmo`no bez nego. Isto taka preku ovoj mehanizam pentamerniot IgM se trasportira vo mukoznite izla~oci, iakoso~inuva dosta pomal procent protivtelo vo mukoznite izla~oci otkolku IgA. Poli-Ig receptorot stapuva vo me|udejstvo so Jverigata i od polimerniot IgA i od IgM protivtelata.Sekretorniot IgA izvr{uva zna~ajna efektorna funkcija vrz povr{inata od mukoznite membrani, {to pretstavuvaat glavnivlezni mesta za pove}eto patogeni organizmi. Bidej}i e polimeren, sekretorniot IgA mo`e vkrsteno da povrze golemi protivgeniso pove}e epitopi. Vrzuvawe sekretorniot IgA za bakteriskite i virusnite povr{ini na protivgenite spre~uva patogenite da seprika~at za mukoznite kletki, spre~uvaj}i virusna infekcija i bakteriska kolonizacija. Kompleksite od sekretorniot IgA iprotivgenot lesno se zarobuvaat vo mukosot a potoa se otstranat od strana na cilijarnite epitelni kletki vo respiratorniottrakt ili so peristaltika vo crevata Se poka`a deka sekretorniot IgA ovozmo`uva zna~ajna linija na odbrana protiv bakteriitetrakt ili so peristaltika vo crevata. Se poka`a deka sekretorniot IgA ovozmo`uva zna~ajna linija na odbrana protiv bakteriitekako {to se Salmonella, Vibrio cholerae iNeisseria gonorrhoeae i virusi kako {to se polio, infunca i reovirusot.Mlekoto od gradite sodr`i sekretoren IgA i mnogu drugi molekuli koi pomagaat da se za{titi novoroden~eto protiv infekcijavo tekot na prvite meseci od `ivotot. Bidej}i imuniot sistem na novoroden~iwata ne e potpolno funkcionalen, doeweto igrazna~ajna uloga vo odr`uvawe zdravjeto kaj novoroden~iwata.
27

Imunoglobulin E (IgE)
Mo}nata biolo{ka aktivnost na IgE mu ozmozmo`uva da bide identifikuvan vo serumot i pokraj negovataisklu~itelno niska prose~na serumska koncentracija (0,3 mkg/ml). IgE protivtelata posreduvaat pri brzitehipersenzitivni reakcii koi se odgovorni za simptomite za alergija na polenov prav, astma, sipanici ianafilakti~en {ok. Prisustvoto na serumskiot del odgovoren za alergi~niot odgovor prvo be{e prika`ano vo1921 godina od strana na K. Prausnic i H. Kastner, koi vnatredermalno vnesle serum od alergi~no vo nealergi~nolice. Koga soodvetniot protivgen podocna bil vnesen na istoto mesto, se pojavil odgovor so otok i crvenilo.Ovaa odgovor. nare~ena P-K odgovor (nare~ena spored nejzinite osnova~i, Prausnic i Kastner), ja pretstavuva{eosnovata za prvata biolo{ka analiza za aktivnosta na IgE.Vistinskata identifikacija na IgE napravija K. i T. I{izaka vo 1966 godina. Tie zele serum od alergi~no lice iso nego imunizirale zajaci za da go podgotvat protividiotipski protivserumot. Na zaja~kiot protivserum mubilo ovozmo`eno da reagira so sekoja klasa od ~ove~kite protivtela koi bile poznati do toga{ (t.e., IgG, IgA,IgM i IgD). Na toj na~in, sekoe od poznatite protivizotipni protivtela bilo talo`eno i otstraneto od
j O b fprotivserumot na zajacite. Ona {to ostanalo bilo protiv izotipsko protivtelo specifi~no zaneidentifikuvana klasa protivtela. Se poka`a deka ova protivtelo celosno go blokira P-K odgovorot. Novotoprotivtelo bilo nare~eno IgE (poradi E protivgenot od polenot od ambrozija, {to pretstavuva mo}enpottiknuva~ na ovaa klasa protivtela).IgE se vrzuva za Fk receptorite na memebranite od krvnite bazofili i tkivni mastociti. Premostuvaweto nareceptorskite IgE molekuli so protivgen (alergen) predizvikuva bazofilite i mastocitite da gi preselatnivnite zrnca vo kon plazminata membrana i da ja oslobodat nivnata sodr`ina vo vonkleto~nata sredina, procespoznat kako obezzrnuvawe (degranulacija). Kako rezultat na toa, se osloboduvaat golem broj farmakolo{kiaktivni posrednici i predizvikuvaat alergiski manifestacii (slika 4-20). Obezzrnuvaweto na lokalnitemastociti pottiknata od IgE isto taka mo`e da oslobodi posrednici koi olesnuvaat sozdavawe razli~ni kletkikoi se potrebni za PZKC protivparazitskata odbrana (vidi poglavje 15).
(slika 4-20 Alergensko premostuvawe receptorski vrzan IgE vo mastocitite pottiknuva obezzrnuvawe,predizvikuvaj}i osloboduvawe supstancii (sini to~ki) koi posreduvaat alergiski projavi (manifestacii).
Imunoglobulin D (IgD)
IgD prvo be{e otkrien koga pacient dobi mnogukraten mielom ~ij mieloi~en protein ne mo`e{e da reagira soprotiv izotipen protivserum protiv izotipovite koi toga{ bea poznati: IgA, IgM i IgG. Koga zajacite beaimunizirani so ovoj mielomski protein, dobieniot protivserum se koriste{e za utvrduvawe istata klasaprotivtela pri niski nivoa vo normalen serum kaj lu|eto. Novata klasa, nare~ena IgD, ima serumskar r r ru j u r D rukoncentracija od 30 mkg/ml i so~inuva okolu 0,2% od celiot imunoglobulin vo serumot. IgD, zaedno so IgM, eglavniot membranski vrzan imunoglobulin prika`an vrz zrelite B keltki, a negovata uloga vo fiziologijata naB kletkite se istra`uva. Ne e utvrdena efektorna funkcija za IgD.
28

Protivgeni odrednici na imunoglobulinite
b } b f }Imunoglobulinskata molekuli, koga }e se vnesat vo drugi vidovi `ivotni, mo`at sami po sebe da funkcioniraat kako mo}ni imunogeni za pottiknuvaweprotivtelen odgovor. Takvi protiv-Ig protivtela pretstavuvaat mo}ni alatki za prou~uvawe razvojot na B kletkite i humoralnite imuni reakcii.Protivgenite odrednici, ili epitopi, na imunoglobulinskite molekuli se delat vo tri glavni kategorii: izotipni, alotipni i idiotipni odrednici,koi se locirani vo karakteristi~ni delovi od molekulata (slika 4-21).
(slika 4-21 Protivgeni odrednici za imunoglobulinite. Za sekoj tip na odrednica, prika`anani se op{tata lokacija na odrednicite vo protivtelnatamolekulata (levo) i dva primeri (sredina i desno). (a) izotipnite odrednici se odrednici vo konstantnite podra~ja koi pravat razlika pome|u sekojaIg klasa i potklasa vo vidovite. (b) Alotipnite odrednici se amino kiselinski mali razliki koi se kodirani od razli~ni aleli vo izotipnite geni.Alotipnite razliki mo`e da se otkrijat so sporeduvawe istata protivtelna klasa me|u razli~nite ~isti soevi. (v) Idiotipnite odrednici se sozdavaatso konformacija na amino kiselinskite sekvenci vo varijabilnite podra~ja od te{kite i od lesnite verigi koi se specifi~ni za sekoj protivgen. Sekojposeben odrednica e nare~en idiotop, a zbirot na posebnite idiotopi e idiotip.
Izotip
Izotipnite odrednici se odrednici vo konstantnoto podra~je koi zaedno ja odreduvaat sekoja klasa i potklasa te{ki verigi i sekoj vid i podvid odlesnite verigi kaj vidovite (vidi slika 421a). Sekoj izotip se kodira so poseben gen od konstantnoto podra~je, a site ~lenovi na vidot ja nosat istatagrupa geni vo konstantnoto podra~je (koi mo`at da vklu~uvaat pove}ekratni aleli). Kaj vidovite, sekoj obi~en poedinec }e gi prika`uva siteizotipovi vo serumot. Razli~ni vidovi nasleduvaat razli~ni geni vo konstantnite podra~ja i zatoa prika`uvaat razli~ni izotipovi. Spored toa, kogaprotivteloto od eden vid }e se vnese vo drug vid, izotipnite odrednici }e bidat prepoznaeni kako tu|i, pottiknuvaj}i protivtelen odgovor konizotipnite odrednici od tu|oto protivtelo. Protiv iziotipnoto protivtelo se koristi rutinski i za istra`uva~ki celi za odreduvawe klasata ilipotklasata na serumskot protivtelo koe se sozdava pri imunot odgovor ili za karakterizirawe na klasata membranski vrzani protivtela prisutni vrzB kletkite.
Alotip
Iako site ~lenovi od vidovite ja nasleduvaat istata grupa izotpni geni, za del od genite postojat mnogu aleli (vidi slika 4-21b). Ovie aleli kodiraatamino kiselini so mali razliki, nare~eni alotipni odrednici, koi se javuvaat kaj del, no ne kaj site, ~lenovi od vidovite. Zbirot od poedine~nitealotipni odrednici prika`an od protivtelnata odrednica e alotip. Kaj lu|eto, alotipovite se karakteristi~ni za site ~etiri IgG potklasi, za ednapotklasa od IgA i za κapa lesnata veriga. Alotipovite vo gama verigata se nare~eni Gm obele`uva~i. Utvrdeni se najmalku 25 razli~ni Gm alotipovi.Tie se imenuvani po klasata i potklasata, po {to sledi brojot na alelite, G1m(1), G2m(23), G3m(11), G4m(4a). Od dvete potklasi na IgA, samo potklasatau , d r j , ( ), ( ), ( ), ( ) d d ,IgA2 ima alotipovi, A2m(1) i A2m(2). Kapa lesnata veriga ima tri alotipovi, nare~eni κapam(1), κapam(2) i κapam(3). Sekoja od ovie alotipniodrednici pretstavuva razlika vo edna do ~etirite amino kiselini koi se kodiraat so razli~ni aleleli od istiot gen.Protivteloto za alotpnite odrednici mo`e da se sozdade so vnesuvawe protivtelo od eden ~len na vidot vo drug ~len od istiot vid koj nosi razli~nialotipni odrednici. Protivteloto za alotipnite odrednici ponekoga{ se sozdava od strana na majkata pri bremenosta kako odgovor na alotpniteodrednici od embrionskite imunoglublini od tatkoto. Protivtelata za alotpnite odrednici mo`at isto taka da nastanat od transfuzija na krv.
Idiotip
Unikatnata amino kiselinska sekvenca kaj VT i VL domenite od dadeno protivtelo mo`e da funkcionira ne samo kako mesto za vrzuvawe protivgen tukuisto taka kako grupa protivgeni odrednici. Idiotipnite odrednici nastanuvaat od sekvencata na varijabilnite podra~ja vo te{kite i vo lesniteverigi. Sekoja oddelna protivgena odrednica na varijabilnoto podra~je e imenuvano kako idiotop (vidi slika 4-21v). Sekoe protivtelo }e prika`epove}e idiotopi, od koi del se vistinskata oblast za vrzuvawe protivgeni, a del gi so~inuvaat sekvencite vo varijabilnite podra~ja nadvor od oblastaza vrzuvawe. Zbirot na posebnite idiotopi e nare~en idiotip na protivteloto.Bidej}i protivtelata koi se sozdadeni od edine~na B kletka nastanuva od istiot klon koj ima identi~ni sekvenci vo varijabilnite podra~ja, site goimaat istiot idiotip. Protiv idiotipskoto protivtelo se sozdava so vnesuvawe protivtela koi imaat minimalni varijacii vo nivnite izotipovi i
^alotipovi, taka {to mo`e da se prepoznae razlikata vo idiotipovite. ^esto se koristi homogeno protivtelo kako {to e mielomskiot protein ilimonoklonsko protivtelo. Vnesuvaweto takvo protivtelo vo primatelot koj e genetski identi~en so daritelot }e dovede do sozdavawe protividiotipsko protivtelo za idiotpskite odrednici.
29

B-kleto~en receptor
Na imunolozite dolgo vreme ne im be{e jasno kako Ig koj se vrzuva za membranata(mIg) vo B kletkite prenesuva aktivira~ki signal po kontakt so protivgenot.Dilemata e vo toa {to site izotipovi na mIg imaat mnogu kratki citoplazminiopa{ki: citoplazminite opa{ki od mIgM i mIgD sodr`at samo tri aminokiselini, krajot na mIgA - 14 amino kiselini, a kraevite na mIgG i mIgE - 28amino kiselini. Vo sekoj slu~aj, citoplazminata opa{ka e premnogu kratka za damo`e da se povrze so vnatrekleto~nite signalni molekuli (na pr., tirozinska
G )kinaza i G proteini).
(slika 4-22 Osnovna struktura na B - kleto~niot receptor (BKR). Ovoj protivgen-vrzuva~ki receptor e sostaven od membranski vrzani imunoglobulini (mIg) idisulfidno povrzani heterodimeri nare~eni Ig-a/Ig-b. Sekoj heterodimer sodr`iimunoglobulin zdoplena struktura i citoplazmini opa{ki mnogu podolgi od oniekaj mIg. Kako {to e prika`ano na slikata, molekulata na mIg e povrzana so edenheterodimer Ig-a/Ig-b. (referenca)r r (r f r )Odgovorot na ovaa zagatka e deka mIg ne go so~inuva celiot receptor za vrzuvaweprotivgeni vo B kletkite. Tuku, B-kleto~niot receptor (BKR) e transmembranskiproteinski kompleks koj e sostaven od mIg i disulfidno vrzani heterodimerinare~eni Ig-alfa/Ig-beta. Molekulite od ovoj heterodimer se povrzuvaat so mIgmolekula za da formiraat BKR (slika 4-22). Ig-alfa verigata ima dolgacitoplazmna opa{ka koja sodr`i 61 amino kiselina, opa{kata na Ig-beta sodr`i48 amino kiselini. Opa{kite na Ig-alfa i Ig-beta se dovolno dolgi za da stapatvo me|udejstvo so vnatrekleto~nite signalni molekuli. Otkrivaweto naheterodimerot Ig-alfa/Ig-beta od strana na Majkl Ret i negovite kolegi voranite 90-ti godini od dvaesettiot vek zna~itelno go zgolemi razbiraweto zaaktivirawe na B kletkite, {to e detalno objasneto vo poglavje 11.
30

Fk receptorite se vrzuvaat za Fk podra~jata od protivtelata
Iako biosintezata i povr{inskoto prika`uvawe imunoglobulini e unikatno za potekloto na B kletkite, mnogukletki imaat membranski glikoproteini nare~eni Fk receptori (FkR) koi imaat afinitet za Fk delovite odizla~enite protivtelni molekuli. Ovie receptori se neophodni za mnogu biolo{ki funkcii na protivtelata. Fkreceptorite se odgovorni za dvi`eweto na protivtelata niz membranite i prenesuvaweto IgG od majkata voembrionot preku placentata. Ovie receptori isto taka ovozmo`uvaat pasivno dobivawe protivtelo od mnoguvidovi na kletki, vklu~uvaj}i B i T limfociti, neutrofili, mastociti, eozinofili, makrofagi i kletkiprirodni ubijci. Spored toa, Fk receptorite ovozmo`uvaat sredstva so koi protivtelata - produktite odsteknatiot imun sistem - mo`at da anga`iraat takvi klu~ni kleto~ni elemnti od vrodeniot imunitet kako {to semakrofagite i kletkite prirodni ubijci. Anga`iraweto protivgeni vrzani za protivtelo od strana na Fkreceptorite na makrofagite i neutrofilite ovozmo`uva efektiven signal za efikasnata fagocitoza(opsonizacija) na protivgen protivtelnite kompleksi. Vo odnos na pottiknuvaweto takvi efektorni funkciikako {to e opsonizacijata ili PZKC, protivgen posreduvanoto premostuvawe na protivtelata vrzani za FkRmo`e da sozdade imunoregulatorni signali koi vlijaat vrz aktivacijata na kletkite pottiknuvaatmo`e da sozdade imunoregulatorni signali koi vlijaat vrz aktivacijata na kletkite, pottiknuvaatdiferencirawe i, vo odredeni slu~ai, nadolno gi reguliraat kleto~nite odgovori.
Postojat mnogu razli~ni Fk receptori (slika 4-23). Receptorot poli-Ig e neophoden za prenesuvawe polimerniteimunoglobulini (IgA i do odreden stepen, pentamerniot IgM) preku epitelnite povr{ini. Kaj lu|eto,neonatalniot Fk receptor (FkRN) prenesuva pove}e IgG od majkata do embrionot pri bremenosta i isto taka igrauloga pri reguliraweto na serumskite nivoa na IgG. Za mnogu klasi od IgG ima otkrieno Fk receptori. Taka, imaFkalfaR receptor koj se vrzuva za IgA, FkepsilonR receptor koi se vrzuva za IgE (vidi isto taka slika 4-20) inekolku razli~ni FkgamaR (R1, R2-A, R2-B1, R2-B2, R3) sposobni da vrzat IgG i negovite potklasi koi sesretnuvaat kaj lu|eto. Vo mnogu slu~ai, premostuvaweto na ovie receptori so vrzuvawe na protivgenprotivtelnite kompleksi, doveduva do pottiknuvawe kaskada za prenesuvawe signalot i rezultira so takvoodnesuvawe kako fagocitoza ili PZKC. Fk receptorot ~esto e del od signalno prenesuva~kiot kompleks koj{tovklu~uva u~estvo na drugi pomaga~ki polipeptidni verigi. Kako {to e prika`ano na slika 4-23, ova mo`e dau u u dru d r r , dvklu~uva par gama verigi ili, vo slu~ajot na IgE receptorot, poslo`en spoj na dve gama verigi i beta veriga.Spojot na vonkleto~niot receptor so vnatrekleto~nata signalno prenosna edinica se sre}ava kaj B-kleto~niotreceptor (slika 4-22) i pretstavuva centralna karakteristika na T-kleto~niot receptoren kompleks (vidipoglavje 9).
(slika 4-23 Strukturata na brojni ~ove~ki Fk receptori. Polipeptidite koi se vrzuvaat za Fk receptorite seprika`ani vo sino, a, tamu kade {to se prisutni, pomaga~kite signalno prenosni polipeptidi se prika`ani vozeleno. Jamkite vo ovie strukturi pretstavuvaat delovi od molekulata so karakteristi~na struktura odimunoglobulinski dipli. Ovie molekuli se pojavuvaat na plazma membranata od razli~ni vidovi na kletki kako{to se kleto~nite povr{inski protivgeni i, kako {to e prika`ano na slikata, mnogu od niv se imenuvani so KDimiwa (od klasteri na diferencijacija, vidi dodatok 1). FkgamaR2 ima tri razli~ni vidovi, A, B1 i B2, koi serazlikuvaat po nivnite vnatrekleto~ni podra~ja.
31

Imunoglobulinska superfamilija
Strukturite na razli~ni imunoglobulinski te{ki i lesni verigi koi bea prethodno objasneti delat nekolkufunkcii, {to uka`uva na toa deka tie imaat zaedni~ki evoluciski predhodnik. Posebno, site klasi na te{ki ilesni verigi imaat strukturata na imunoglobulinski zdiplen domen (vidi slika 4-8). Prisustvoto na ovaakarakteristi~na struktura kaj site imunoglobulinski te{ki i lesni verigi uka`uva na toa deka genite koi gikodiraat niv poteknuvaat od zaedni~ko prvobitno gensko kodirawe polipeptid od okolu 110 amino kiselini.Genskoto duplirawe i podocne`noto odvojuvawe mo`elo da sozdade razli~ni geni vo te{kite i vo lesnite verigi.Se poka`a deka golem broj membranski proteini poseduvaat eden ili pove}e podra~ja koi se homogeni zaimunoglobulinskiot domen. Sekoj od ovie membranski proteini se klasificira kako ~len na imunoglobulinskasuperfamilija. Izrazot superfamilija se koristi za imenuvawe proteini ~ii soodvetni geni proizleguvaat odzaedni~kiot prvobiten gen koj ja kodira osnovanata struktura na domenot. Ovie geni se razvile nezavisno i nedelat genska povrzanost ili funkcija. Slednive proteini, vo odnos na samite imunoglobulini, se pretstavitelni~lenovi na imunoglobulinskata superfamilija (slika 4-24):
Heterodimer Ig-alfa/Ig-beta, del od B-kleto~niot receptorReceptor poli-Ig, koj pridonesuva za sekretorniot del vo IgA i IgMT-kleto~en receptorT-kleto~ni pomaga~ki proteini, vklu~uvaj}i gi KD2, KD4, KD8, KD28 i gama, delta i epsilon verigi od KD3Klasa 1 i klasa 2 GTK molekulibeta - mikroglobulin, nepromenliv protein povrzan so klasa 1 GTK molekuliteRazli~ni kleto~no adhezivni molekuli. vklu~uvaj}i VCAM-1, MKAM-1, MKAM-2 i LFA-3.Trombociten faktor za rast
Brojni drugi proteini, del od niv opi{ani vo drugi poglavja, isto taka pripa|aat na imunoglobulinskatasuperfamilija.S#u{te ne e izvr{ena rendgenska kristalografska analiza na site ~lenovi od imunoglobulinskatasuperfamilija. No i pokraj toa, prvi~nata sekvenca na amino kiselinite od ovie proteini uka`uva na toa dekasite tie go sodr`at obi~niot imunoglobulinski zdiplen domen. Konkretno, site ~lenovi od imunoglobulinskatasuperfamilija sodr`at najmalku edna ili pove}e verigi od okolu 110 amino kiselini, sposobni za podreduvawe voobitkanite lenti na protivparalenite beta plo~ki, obi~no so nepromenliva me|uveri`na disulfidna vrska kojazatvora jamka koja od 50 do 70 ostatoci.
(slika 4-24 Nekoi ~lenovi od imunoglobulinska superfamilija, grupa na strukturno povrzani, obi~nomembranski vrzani glikoproteini. Vo site prika`ani slu~ai osven za beta - mikroglobulinot,
b j bkarboksilnokrajniot del od molekulata e pricvrsten za membranata.
32

Monoklonski protivtela
K } } f B jKako {to spomenavme prethodno, pove}eto protivgeni nudat pove}e epitopi i zatoa pottiknuvaat razmno`uvawe i diferencirawe na mno{tvo B-kleto~ni klonovi, pri {to sekoj eizveden od B kletkata koja prepoznava poseben epitop. Serumskite protivtelata koi se dobivaat od toa se heterogenic, so~inuvaj}i me{avina od protivtela, pri {to sekoe especifi~no za eden epitop (slika 4-25). Takviot odgovor na poliklonsko protivtelo ja olesnuva lokalizacijata, fagocitozata i komplementarno posredenoto ragraduvawe protivgen,na toj na~in ima jasni prednosti za organizamot in vivo. Za `al, heterogeni~nosta na protivteloto koe ja zgolemuva imunolo{kata za{tita in vivo ~esto ja namaluva delotvornosta naprotivserumot za razli~ni in vitro upotrebi. Za pove}eto istra`uva~ki, dijagnosti~ki i terapevtski celi, se preferiraat monoklonski protivtela, izvedeni od eden klon i koispored toa se specifi~ni za eden epitop.
(slika 4-25 Voobi~aeniot poliklonski protivserum sozdaden kako odgovor na kompleks protivgen sodr`i me{avina od monoklonski protivtela, pri {to sekoe e specifi~no za eden od~etirite epitopi prika`ani kaj protivgenot (vmetnato). Na sproti toa, monoklonskoto protivtelo koe e izvedeno od edna plazma kletka, e specifi~no za eden epitop vo kompleksniotprotivgen. Ovde se prika`ani konturite na osnoviot metod za dobivawe monoklonsko protivtelo.
Ne e izvodlivo direktno biohemisko pro~istuvawe monoklonskoto protivtelo od preparat na poliklonski protivtela. Vo 1975 god., Xorx Koler i Cezar Mil{tajn osmislija metod zapodgotvuvawe monoklonsko protivtelo, {to brzo stana edna od imunite klu~ni tehnologii. So spojuvawe (fuzija) normalno aktivirana, protivtelno sozdava~ka B kletka so mielomskakletka (kancerozna plazma kletka), tie mo`ea da sozdadat hibridna kletka, nare~ena hibridom, koja gi poseduva{e svojstva na bessmrten rast {to gi ima mielomskata kletka iizla~uva{e protivtelo koe be{e sozdadeno od B kletkite (vidi slika 4-25). Dobienite klonovite od hibridomnite kletki, koi la~at golemi koli~ini monoklonski protivtela, mo`atneograni~eno da se kultiviraat. Razvojot na tehnikite za sozdavawe monoklonski protivtela, detalite koi se opi{ani vo poglavje 22, na imunolozite im dade mo}na i raznovidnaistra`uva~ka alatka. Zna~eweto na rabotata na Koler i Mil{tajn be{e priznaeno koga i dvajcata bea nagradeni so Nobelova nagrada.
Monoklonskite protivtela imaat zna~ajni klini~ki upotrebi
Se poka`a deka monoklonskite protivtela se dosta korisni kako dijagnosti~ki, prika`uva~ki i terapevtski reagensi vo klini~kata medicina. Pome|u pove}eto dijagnosti~kireagensi so monoklonski protivtela koi se dostapni vo momentov se nao|aat proizvodite za otkrivawe bremenost, dijagnosticirawe brojni patogeni mikroorganizmi, merewetonivoata vo krvta na razli~ni lekovi, protivgeni za histokompitabilna sovpadlivost i otkrivawe protivgenite ra{ireni od nekoi tumori.Radioozna~enite monoklonski protivtela isto taka mo`e da se koristat in vivo za otkrivawe ili locirawe tumorski protivgeni, ovozmo`uvaj}i rana dijagnoza na odredeni prvi~niili metastatski tumori kaj pacienti. Na primer, monoklonskoto protivtelo za kletki od rak na dojkata se bele`i so jod-131 i se vnesuva vo krvta za otkrivawe ra{irenosta na tumorotvo okolnite limfni jazli. Ovaa tehnika za monoklonsko prika`uvawe mo`e da otkrie metastaza na rak na dojkata koj ne bi se otkril so drugi, pomalku ~uvstvitelni tehniki naskenirawe.Kako {to e objasneto vo klini~kiot fokus na strana 98, upotrebata na protivtelata kako terpavetski agensi ima dolga istorija. Od neodamna dostapnosta na monoklonskite protivtelai sposobnosta za nivno ühumaniziraweû so tehniki za genetsko modificirawe i dade nov pottik na ovaa oblast. Ranite obidi za upotreba na gluv~e{ki monoklonski protivtela zalekuvawe lu|e se soo~ija so mo`nosta od silen odgovor protiv ovie tu|i proteini. Najnovite produkti se sozdavaat vo ~ove~ki sistemi ili se genetski modificirani (vidi poglavje 5)za vklu~uvawe V podra~jata ili KDR od protivtela koi ne se od lu|eto voK podra~jata i skeletot na ~ove~kite protivtela, namaluvaj}i ja mo`nosta za pokrenuvawe imun odgovorprotiv niv.Postoi raste~ki broj na priznaeni terapevtski protivtela i nad stotici novi protivtela vo patekite za ravoj na lekovite. Tie pridonesuvaat nekolku milijardi dolari od proda`ba.Glavnite podra~ja kade terapijata so protivtela se poka`a korisna se lekuvaweto na rak i lekuvawe artritski naru{uvawa. Proivodite Rituksan (generi~ko ime, rituksimab) zadr j d r j r r u r u r r ru u r d u ( r , r u )lekuvawe Nehoxkinov limfom i Remikejd (generi~ko ime, infliksimab) ili Humira (adalinumab) za revmatski artrit se pome|u najmnogu upotrebuvanite. Generi~kite imiwa nalekovite go otslikuvaat vidot na protivteloto. Na primer, sufiksot -umab ozna~uva ~ove~ko monoklonski protivtelo, dodeka -imab ozna~uva himeri~no protivtelo, koe sodr`isekvenci i od lu|eto i od drugite vidovi.
Abzimite se monoklonski protivtela koi kataliziraat reakcii
Vrzuvaweto protivteloto za negoviot protivgen na mnogu na~ini e sli~no na vrzuvaweto na enzimot za negoviot supstrat. I vo dvata slu~ai vrvuvaweto vklu~uva slaba, nekovalentnavrska i poseduva visoka specifi~nost i ~esto visok afinitet. Ona {to gi razlikuva me|udejstvoto na protivteloto so protivgenot od me|udejstvoto na enzimot so supstratot e toa {toprotivteloto ne gi menuva kovalentnite vrski na protivgenot, dodeka enzimot katalizira hemiska promena vo negoviot supstrat. Sepak, kako i enzimite, protivtelata so soodvetnaspecifi~nost mo`e da ja stabilizraat preodnata sostojba na vrzniot supstrat, na toj na~in namaluvaj}i ja aktivaciskata energija za hemisko modificirawe supstratot.Sli~nostite pome|u me|udejstvoto protivgen protivtelo i me|udejstvoto enzim-supstrat go postavija pra{aweto za toa dali odredeni protivtela mo`e da se odnesuvaat kako enzimi ida kataliziraat hemiski reakcii. Za ispituvawe ovaa mo`nost, se sintetizira{e kompleks hapten-nosa~ pri {to haptenot strukturno nalikuva{e na preodnata sostojba na eden esterkoj pretrpuva hidroliza. Kletkite od slezinata kaj glu{ecot koj e imuniziran so ovoj analog vo preodna sostojba bea izme{ani so mielomski kletkite za sozdavawe monoklonskiprotivhaptensko monoklonski protivtela. Koga na ovie monoklonski protivtela bea inkubirani so ester supstrat, del od niv ja zabrzaa hidrolizata za okolu 1000 pati, tie seodnesuvaa kako enzim koj obi~no ja katalizira hidrolizata na supstratot. Kataliti~nata aktivnost na ovie protivtela be{e visoko specifi~na: tie hidrolziraa samo esteri ~ijastruktura vo preodna sostojba dosta nalikuva{e na analogot vo preodna sostojba koj se koriste{e kako hapten pri imunizirawe so kowugatot. Ovie kataliti~ni protivtela beanare~eni abzimi spored nivnata dvojna uloga kako protivtela i enzimi.r r d d j u rGlavnata cel na istra`uvaweto so katalizirawe protivtela e sozdavwe baterija od abzimi koi gi delat peptidnite vrski na odredeni amino kiselinski ostatoci, kako {torestrikcionite enzimi ja se~at DNK na specifi~ni mesta. Takvite enzimi se od neprocenliva vrednost pri strukturnata i funkcionalnata analiza na proteinite. Dopolnitelno,mo`no e sozdavawe abzimi so sposobnost za razgraduvawe sosiroci vo krvta ili da rase~at virusni glikoproteini vo specifi~ni mesta, na toj na~in blokiraj}i ja virusnata infekcija.Za `al, kataliti~nite protivtela koi gi rase~uvaat peptidnite vrski na proteinite mnogu e te{ko da se dobijat. Golem del od istra`uvawata koi vo momentov se vr{at se posveteni naovoj zna~aen no te`ok problem.
33


















