
Proteomic Characterization of the Venom Of Five Bombus ... · for venom extraction. For venom...
17
FULL TITLE: PROTEOMIC CHARACTERIZATION OF THE VENOM OF FIVE BOMBUS (THORACOBOMBUS) SPECIES Nezahat Pınar Barkan a1 , Mustafa Bilal Bayazit b , Duygu Demiralp Özel c2 a Hacettepe University, Faculty of Science, Department of Biology, Ankara, Turkey b University of Lausanne, Department of Fundamental Neurosciences, Lausanne, Switzerland c Ankara University, Faculty of Engineering, Biomedical Engineering, Ankara, Turkey 1 Current Address: University of Gothenburg, Department of Oncology, Sahlgrenska Academy, Gothenburg, Sweden 2 Corresponding author: [email protected] not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was this version posted September 25, 2017. . https://doi.org/10.1101/193524 doi: bioRxiv preprint
Transcript of Proteomic Characterization of the Venom Of Five Bombus ... · for venom extraction. For venom...

FULL TITLE: PROTEOMIC CHARACTERIZATION OF THE VENOM OF FIVE BOMBUS
(THORACOBOMBUS) SPECIES
Nezahat Pınar Barkana1, Mustafa Bilal Bayazitb, Duygu Demiralp Özelc2
aHacettepe University, Faculty of Science, Department of Biology, Ankara, Turkey
bUniversity of Lausanne, Department of Fundamental Neurosciences, Lausanne,
Switzerland
cAnkara University, Faculty of Engineering, Biomedical Engineering, Ankara, Turkey
1Current Address: University of Gothenburg, Department of Oncology, Sahlgrenska
Academy, Gothenburg, Sweden
2Corresponding author: [email protected]
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Abstract
Venomous animals use venom; a complex biofluid composed of unique mixtures of proteins and
peptides, to act on vital systems of the prey or predator. In bees, venom is solely used for
defense against predators. However, the venom composition of bumble bees (Bombus sp.) is
largely unknown. Thoracobombus subgenus of Bombus sp. is a diverse subgenus represented
by 14 members across Turkey. In this study, we sought out to proteomically characterize the
venom of five Thoracobombus species by using bottom-up proteomic techniques. We have
obtained two-dimensional polyacrylamide gel (2D-PAGE) images of each venom sample. We
have subsequently identified the protein spots by using matrix assisted laser desorption
ionization / time of flight mass spectrometry (MALDI-TOF MS). We have identified 47 proteins for
Bombus humilis; 32 for B. pascuorum, 60 for B. ruderarius; 39 for B. sylvarum and 35 for B.
zonatus. Our analyses provide the primary proteomic characterization of five bumble bee
species’ venom composition.
Keywords: Bumble bees, MALDI-TOF MS, Proteomics, 2D-PAGE, Venom
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Introduction
In venomous animals, the use of venom or poison is one of the most essential mechanisms
used for either capturing prey or defending oneself against predators. In competing for
resources, venom is an adaptive trait and an example of convergent evolution. It is a complex
biofluid secreted by venom gland and is composed of unique mixture of proteins, enzymes,
peptides and biogenic amines that act on vital systems of the recipient (Calvete, 2009). In real
bees (Apoidea), largest family of bees with over 20.000 members (Fitzgerald, 2013), venom is
solely used for defense against predators (Arbuckle, 2017).
Among Apoidea, the majority of venom studies are focused on honey bee Apis mellifera (A.
mellifera). Bombus sp., which also belong in the same family as A. mellifera, are pollinators for
various native and cultivated plants (Free, 1993). The venom composition of Bombus sp. is
poorly understood.
Previous studies on Bombus sp. venom have mainly focused on characterization and isolation of
highly abundant single molecules using bio-assays and chemical sequencing via Edman
degradation (Choo, Lee, Yoon, Je, et al., 2010; Choo, Lee, Yoon, Kim, et al., 2010; Choo, Lee,
et al., 2012; Choo et al., 2011; Choo, Yoon, & Jin, 2012; Favreau et al., 2006; Kim et al., 2013;
Qiu et al., 2011; Qiu, Choo, Yoon, & Jin, 2012a, 2012b; Qiu et al., 2013; Wan et al., 2014).
Among the described peptides, bombolitins are unique to Bombus sp. and enhance
phospholipase A2 in liposome hydrolysis, in a similar fashion to melittin found in A. mellifera
venom and crabrolin and mastoparan found in vespid Vespa crabro venom (Argiolas & Pisano,
1985). Phospholipase A2 (Hoffman, El-Choufani, Smith, & De Groot, 2001; Hoffman & Jacobson,
1996; Van Vaerenbergh, Debyser, Smagghe, Devreese, & de Graaf, 2015), serine proteases
(Choo, Lee, Yoon, Kim, et al., 2010; Choo et al., 2011; Kim et al., 2013; Qiu et al., 2011; Qiu et
al., 2012b; Van Vaerenbergh et al., 2015) , serine protease inhibitors (Qiu et al., 2013; Wan et
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

al., 2014), acid phosphatases (Hoffman et al., 2001; Hoffman & Jacobson, 1996; Van
Vaerenbergh et al., 2015), arginine kinase (N. P. Barkan, Demiralp, & Aytekin, 2015),
hyaluronidase (Hoffman & Jacobson, 1996), putrescine (Tom Piek, 2013), citrate (Fenton et al.,
1995), defensin (Rees, Moniatte, & Bulet, 1997), and acethylcoline (T Piek, Veldsema-Currie,
Spanjer, & Mantel, 1983) have also been isolated from Bombus sp. venom.
Recently, the whole genome sequencing of European large earth bumblebee B. (Bombus)
terrestris has become publically accessible. The accessibility to the genome sequence,
combined with the use of mass spectrometry, paved the way for deeply analyzing the venom
proteome composition of this species. This study has identified 55 new molecules in the B.
terrestris venom. Moreover, 72% of identified molecules had homologs in A. mellifera venom
indicating that while there are some species-specific differences, these two bee species share
major defense mechanisms. B. terrestris is the only member of the Bombus sp. whose venom
proteome composition has been characterized.
In bumble bees, color patterns show similarity among species or are highly variable within
species. Therefore, classification based on coat color can be unreliable (P. Williams, 2007). This
is especially true for the Thoracobombus subgenus which consists of 14 species in Turkey that
are morphologically difficult to separate. To overcome this, we have previously used landmark
based geometric morphometrics on their wing structure (N. P. A. Barkan, A. M., 2013). Other
approaches, use of pheromone analysis (Hovorka, Valterova, Rasmont, & Terzo, 2006; Terzo,
Valterová, & Rasmont, 2007; Terzo, Valterova, Urbanova, & Rasmont, 2003; Urbanová,
Valterová, Rasmont, & Terzo, 2002) and DNA based molecular techniques (S. Cameron, Hines,
& Williams, 2007; S. A. Cameron, Hines, & Williams, 2006; S. A. Cameron & Williams, 2003; Du
et al., 2015; Kawakita et al., 2003; Kawakita et al., 2004; P. H. Williams et al., 2012) have also
been used in differentiating bumble bee species.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Turkey is regarded as one of the countries with the highest amount of species richness both in
number and diversity in the West-Palaearctic region. The main aim of this study is to determine
and compare the venom protein profiles of different Thoracobombus species of Turkey using
bottom-up proteomic strategies. With the adoption of these approaches, not only will we
characterize the venom proteome of Thoracobombus species for the first time, but we will also
highlight the proteomic differences within the subgenus that may help in differentiating between
the species.
2. Methods
2.1. Venom Extraction
B. humilis (Illiger), B. pascuorum (Scopoli), B. ruderarius (Müller), B. sylvarum (Linnaeus), and
B. zonatus (Smith) were collected from Ankara, Bolu, Çankırı and Kayseri provinces from Middle
Anatolian Region during June, July, August and September in 2013 and 2014 (Table S1), with
the permission from the Republic of Turkey, Ministry of Forestry and Water Affairs Directorate of
Nature Conservation & National Parks, and the Republic of Turkey, Ministry of Food, Agriculture
& Livestock. Alive specimens were transported to Hacettepe University Department of Biology
for venom extraction. For venom extraction, the venom reservoir is removed from the stinger in a
sterile environment with forceps and the membrane is disrupted afterwards. From each
individual, this method yielded 2-5 µl of venom which were collected in a cryotube and
transported in liquid nitrogen. For proteomic analyses venom samples were pooled, while for
spectroscopic analyses, populations were pooled separately. Collected samples were stored at -
80°C until use.
2.2. Proteomic Analysis
2.2.1. Protein Quantification and Two-Dimensional Gel Electrophoresis
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Venom of aforementioned bumble bee species was collected from 20 individuals per species.
Protein concentrations of the venom samples were obtained by using the Bradford assay
(Bradford, 1976). Venom was dissolved in 20 µl rehydration buffer [7M urea, 2 M thiourea, 4%
CHAPS (w/v), 1% ampholytes (pH 3-10), 10 mM DTT and trace amount of bromophenol blue].
2D-PAGE was performed as previously described (Igci & Demiralp, 2012) in three technical
replicates and a total of 50 µg of protein was loaded per gel.
2.2.2. 2D-PAGE Analysis
2D gel electrophoresis (2DE) images from venom samples of 5 different Thoracobombus
species were analyzed using PDQuest Software Advanced 8.0.1 (Bio-Rad, CA, USA). PDQuest
software examines each spot on every gel image and it picks a master gel containing most
spots. All spots in each gel image is then matched onto the master gel. Each post is annotated
with an identification number (SSP). For quantifying spots, four different normalization options
are performed including total density in gel images, local regression, total quantity in valid spot
and mean of log ratios. In the present study, 2DE image analysis is performed for both detecting
common and/ or unique spots between samples and for quantifying spot intensity which
accounts for the amount of protein in that spot. All spots were matched manually and ‘total
density in gel images’ normalization was performed. Two-way ANOVA was performed to detect
significancies in spot intensities (IBM Corp. Released 2010. IBM SPSS Statistics for Windows,
Version 19.0. Armonk, NY: IBM Corp.).
2.2.3. In-gel enzymatic digestion, Matrix-Assisted laser desorption ionization/ time
of flight (MALDI-TOF) mass spectrometry and peptide mass fingerprinting (PMF)
analysis
In gel trypsinization of protein spots was performed as previously described (Igci & Demiralp,
2012). Obtained peptides were loaded on MALDI-TOF plates using ZipTip® Pipette Tips
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

(Millipore). Tryptic peptides were dissolved in sample solvent containing 0.1% and 50%
acetonitrile and mixed with an equal volume of matrix solution containing alpha-cyano-4-
hydroxycinnamic acid, 0.1% TFA and 75% acetonitrile. 1.5 µl of these mixtures were spotted
onto a target plate and analyzed with MALDI-TOF mass spectrometer (Water, UK) operated in
positive-ion reflectron mode. Obtained spectra were then examined in MASCOT search engine
(http://www.matrixscience.com/cgi/search_form.pl?FORMVER=2&SEARCH=PMF). Organisms
taxonomically related to bumble bees were selected as bumble bees are not present in the
database. “Other Metazoa” was selected as the taxonomical group as it is a diverse group
including all venomous animals and Drosophila sp., the taxonomically closest species to bumble
bees. Both Swissprot and NCBInr databases were used to maximize the number of hits.
Identified proteins were categorized into protein families using Swissprot database.
3. Results
3.1. 2D-PAGE Analysis of Thoracobombus Venom
2DE gel images were obtained from each venom sample (Fig 1).
In Thoracobombus matchset 87 protein spots were manually matched in 2DE images of B.
humilis, B. pascuorum, B. sylvarum, B. ruderarius and B. zonatus venom. Spot quantities were
calculated and significant spots were detected statistically. Among these protein spots, 21 spots
were present in each species. Common and differentially expressed protein spots are
summarized (Table S2).
3.2. Mass-spectrometry of Thoracobombus Venom
From the 2DE gel images, we have spectrometrically characterized 47 proteins for B. humilis; 32
for B. pascuorum; 60 for B. ruderarius; 39 for B. sylvarum and 35 for B. zonatus (Fig. S1 and
Table S3-7). Putative toxins are categorized according to their function and subcellular location
(Van Vaerenbergh et al., 2015) (Figure 1). Toxins previously identified in B. terrestris and A.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

mellifera venoms are used as references (Table 2) (Van Vaerenbergh, Debyser, Devreese, & de
Graaf, 2014; Van Vaerenbergh et al., 2015). Major putative toxins; phospholipase A2, venom
protease, venom acid phosphatase and hyaluronidase were detected in venom of all five
analyzed Thoracobombus species. Arginine kinase, a known component of spider and parasitoid
wasp venom, was also identified in all studied species. Detection of serine protease snake (B.
ruderarius and B. sylvarum), peptidases (B. humilis and B. sylvarum), and kunitz-type serine
protease inhibitor (B. humilis and B. sylvarum), were limited to certain species.
3.3. Species Specific Expression of Putative Venom Toxins
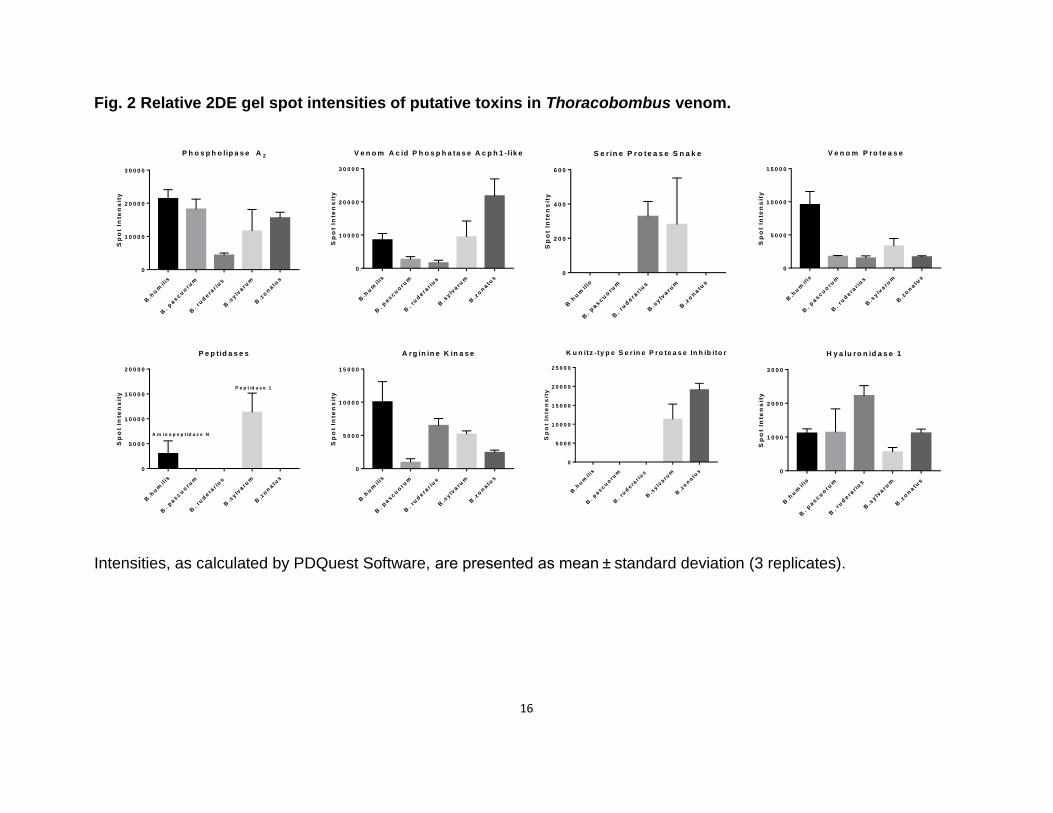
Next, we compared the intensities of 2DE gel spots corresponding to putative toxins in each
sample (Fig. 2). According to these spot intensities, several toxins display species specific
expression patterns (Fig. 3). The expression of phospholipase A2, is the lowest in B. ruderarius
venom among studied species. B. zonatus venom displays the highest venom acid phosphatase
expression. B. pascuorum and B. ruderarius are characterized by a relatively low venom acid
phosphatase expression. B. humilis and B. pascuorum display the most and the least abundant
arginine kinase profile, respectively. B. humilis displays the highest venom protease expression.
Hyaluronidase 1 expression is more than two folds higher in B. ruderarius venom compared to
the other species.
4. Discussion
Identification of Thoracobombus subgenus represents a challenge as its members are
morphologically similar. In our previous study, we had applied landmark based geometric
morphometrics to distinguish its members from each other more accurately. In this current study,
we provide the primary proteomic characterization of the venom profile of five bumble bee
species belonging to this subgenus. Several putative venom toxins are detected in
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Thoracobombus which were in accordance with B. terrestris and A. mellifera venom.
Furthermore, we outline the species specific expression patterns of putative toxins.
In Hymenoptera, the defensive behavior of social groups is similar. Venom is used for protecting
the colonies rather than for capturing prey. Therefore, the venom composition of these groups
are expected to have common overall pattern. It is recently illustrated that the venom profile of B.
terrestris and A. mellifera largely overlap, while also retaining certain species specific
differences.
Accordingly, in Thoracobombus venom we have identified putative venom toxins collectively
used by several Hymenoptera species to debilitate predators such as; phospholipase A2,
arginine kinase, acid phosphatase (venom acid phosphatase Acph-1), serine protease (serine
protease snake), peptidases (peptidase 1 and aminopeptidase N), hyaluronidase (hyaluronidase
1) and serine protease inhibitors (kunitz-type serine protease inhibitor).
While the Thoracobombus venom composition may be similar to other Hymenoptera species in
terms of major toxins identified, it is important to note that the venom may be tailored for the
individual species’ needs with regards to abundancies of protein families present. Accordingly, a
change in the protein family profile of snake venom has been described upon nutritional shift.
Here, we have compared the 2DE gel images obtained from different Thoracobombus species
and highlighted the species specific expression differences of venom proteins. In particular, our
results indicate that expression levels of putative toxins may be used in differentiating between
species. Within the Thoracobombus subgenus, B. humilis is morphologically very similar to B.
pascuorum; while B. ruderarius and B. sylvarum resemble each other in terms of morphology
and coat color. On the other hand, B. zonatus with its characteristic yellow coat color is easily
distinguished from others. We have collected B. ruderarius and B. sylvarum at the highest
altitudes (>2000m) and B. zonatus at the lowest (~ 1000m). Interestingly, we have found that
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

morphologically similar species differentiate in their expression of putative venom toxins, even in
a similar habitat.
In future studies, different populations of same species can be used to assess the direct effect of
altitude, vegetation, and prey population on the venom composition of bees. In this study we
utilized 2DE gel images and MALDI-TOF mass spectrometry for assessing the expression levels
of putative toxins, which may have its disadvantages. For example, we could not detect
bombolitin, unique bumble bee peptides, in our samples. As bombolitins are very small peptides
(10 kilo Daltons), we may have failed to detect these peptides with our equipment. More
sensitive quantitative mass-spectrometry approaches may be necessary to confirm the
correlation between microenvironment and the venom profile of bumble bees.
Profiling bumble bee venom can have pharmaceutical benefits. Venom peptides can be orally
active and transported through the blood-brain barrier and through mammalian cell membranes.
Commercial venom-derived drugs are available for the treatment of major diseases such as
diabetes, stroke, and hypertension (King, 2011). Bumble bee venom may have the potential to
serve as an alternative source of these drugs which are mostly snake venom-derived.
Understanding the species specific expression patterns of toxins can also benefit the specific-
compound oriented pharmacological analyses.
In conclusion, we provide the primary proteomic characterization of a diverse bumble bee
subgenus, and illustrate species specific differences in toxin expression.
Acknowledgments
This study was part of the PhD thesis of Nezahat Pinar Barkan. The experiments of this study
were performed completely in Ankara University Biotechnology Institute. We are grateful to
Selen Peker, Naşit İğci, Beycan Ayhan and Hatice Yıldızhan for their great help during
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

laboratory work. We would especially like to thank Seçil Karahisar Turan for her great support
and guidance throughout the study.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

References
Arbuckle, K. (2017). Evolutionary Context of Venom in Animals. In A. Malhotra (Ed.), Evolution of Venomous Animals and Their Toxins (pp. 3-31). Dordrecht: Springer Netherlands.
Argiolas, A., & Pisano, J. J. (1985). Bombolitins, a new class of mast cell degranulating peptides from the venom of the bumblebee Megabombus pennsylvanicus. Journal of Biological Chemistry, 260(3), 1437-1444.
Barkan, N. P., Demiralp, D. O., & Aytekin, A. M. (2015). Comparison of Venom Extraction Methods of Bombus (Bombus) terrestris (Hymenoptera: Apidae: Bombus) by Using Bottom-up Proteomic Strategies. Current Proteomics, 12(2), 80-86.
Barkan, N. P. A., A. M. (2013). Systematical studies on the species of the subgenus Bombus (Thoracobombus) (Hymenoptera: Apidae: Bombus Latreille) in Turkey. Zootaxa, 3737(2), 167-183.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, 72, 248-254.
Calvete, J. J. (2009). Venomics: digging into the evolution of venomous systems and learning to twist nature to fight pathology. J Proteomics, 72(2), 121-126. doi:10.1016/j.jprot.2009.01.018
Cameron, S., Hines, H., & Williams, P. (2007). A comprehensive phylogeny of the bumble bees (Bombus). Biological Journal of the Linnean Society, 91(1), 161-188.
Cameron, S. A., Hines, H. M., & Williams, P. H. (2006). Molecular phylogeny of the bumble bee subgenus Pyrobombus (Hymenoptera: Apidae: Bombus) with insights into gene utility for lower-level analysis. Invertebrate systematics, 20(3), 289-303.
Cameron, S. A., & Williams, P. H. (2003). Phylogeny of bumble bees in the New World subgenus Fervidobombus (Hymenoptera: Apidae): congruence of molecular and morphological data. Molecular phylogenetics and evolution, 28(3), 552-563.
Choo, Y. M., Lee, K. S., Yoon, H. J., Je, Y. H., Lee, S. W., Sohn, H. D., & Jin, B. R. (2010). Molecular cloning and antimicrobial activity of bombolitin, a component of bumblebee Bombus ignitus venom. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 156(3), 168-173.
Choo, Y. M., Lee, K. S., Yoon, H. J., Kim, B. Y., Sohn, M. R., Roh, J. Y., . . . Woo, S. D. (2010). Dual function of a bee venom serine protease: prophenoloxidase-activating factor in arthropods and fibrin (ogen) olytic enzyme in mammals. PLoS One, 5(5), e10393.
Choo, Y. M., Lee, K. S., Yoon, H. J., Qiu, Y., Wan, H., Sohn, M. R., . . . Jin, B. R. (2012). Antifibrinolytic role of a bee venom serine protease inhibitor that acts as a plasmin inhibitor. PloS one, 7(2), e32269.
Choo, Y. M., Qiu, Y., Yoon, H. J., Lee, K. Y., Sohn, H. D., & Jin, B. R. (2011). Enzymatic properties of a bee venom serine protease from the bumblebee Bombus ignitus. Journal of Asia-Pacific Entomology, 14(3), 249-251.
Choo, Y. M., Yoon, H. J., & Jin, B. R. (2012). Effects of the bumblebee (Bombus ignitus) venom serine protease inhibitor on serine protease and phospholipase A 2 of B. ignitus venom. Journal of Asia-Pacific Entomology, 15(4), 543-545.
Du, Q., Bi, G., Zhao, E., Yang, J., Zhang, Z., & Liu, G. (2015). Complete mitochondrial genome of Bombus terrestris (Hymenoptera: Apidae). Mitochondrial DNA, 1-2. doi:10.3109/19401736.2015.1089568
Favreau, P., Menin, L., Michalet, S., Perret, F., Cheneval, O., Stöcklin, M., . . . Stöcklin, R. (2006). Mass spectrometry strategies for venom mapping and peptide sequencing from crude venoms: case applications with single arthropod specimen. Toxicon, 47(6), 676-687.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Fenton, A. W., West, P. R., Odell, G. V., Hudiburg, S. M., Ownby, C. L., Mills, J. N., . . . Shannon, S. B. (1995). Arthropod venom citrate inhibits phospholipase A 2. Toxicon, 33(6), 763-770.
Fitzgerald, K. T. (2013). Chapter 49 - Insects—Hymenoptera A2 - Peterson, Michael E. In P. A. Talcott (Ed.), Small Animal Toxicology (Third Edition) (pp. 573-588). Saint Louis: W.B. Saunders.
Free, J. (1993). Insect pollination of crops. In: Academic Press, London, 684pp. Hoffman, D. R., El-Choufani, S. E., Smith, M. M., & De Groot, H. (2001). Occupational allergy to
bumblebees: Allergens of Bombus terrestris. Journal of Allergy and Clinical Immunology, 108(5), 855-860. doi:10.1067/mai.2001.119029
Hoffman, D. R., & Jacobson, R. S. (1996). Allergens in hymenoptera venom XXVII: Bumblebee venom allergy and allergens. Journal of Allergy and Clinical Immunology, 97(3), 812-821. doi:10.1016/S0091-6749(96)80159-X
Hovorka, O., Valterova, I., Rasmont, P., & Terzo, M. (2006). Male cephalic labial gland secretions of two bumblebee species of the subgenus Cullumanobombus (Hymenoptera: Apidae: Bombus latreille) and their distribution in central Europe. Chem Biodivers, 3(9), 1015-1022. doi:10.1002/cbdv.200690099
Igci, N., & Demiralp, D. O. (2012). A preliminary investigation into the venom proteome of Macrovipera lebetina obtusa (Dwigubsky, 1832) from Southeastern Anatolia by MALDI-TOF mass spectrometry and comparison of venom protein profiles with Macrovipera lebetina lebetina (Linnaeus, 1758) from Cyprus by 2D-PAGE. Arch Toxicol, 86(3), 441-451. doi:10.1007/s00204-011-0763-5
Kawakita, A., Sota, T., Ascher, J. S., Ito, M., Tanaka, H., & Kato, M. (2003). Evolution and phylogenetic utility of alignment gaps within intron sequences of three nuclear genes in bumble bees (Bombus). Molecular Biology and Evolution, 20(1), 87-92.
Kawakita, A., Sota, T., Ito, M., Ascher, J. S., Tanaka, H., Kato, M., & Roubik, D. W. (2004). Phylogeny, historical biogeography, and character evolution in bumble bees (Bombus: Apidae) based on simultaneous analysis of three nuclear gene sequences. Molecular Phylogenetics and Evolution, 31(2), 799-804.
Kim, J. S., Choi, J. Y., Lee, J. H., Park, J. B., Fu, Z., Liu, Q., . . . Parker, B. L. (2013). Bumblebee venom serine protease increases fungal insecticidal virulence by inducing insect melanization. PloS one, 8(4), e62555.
King, G. F. (2011). Venoms as a platform for human drugs: translating toxins into therapeutics. Expert Opin Biol Ther, 11(11), 1469-1484. doi:10.1517/14712598.2011.621940
Piek, T. (2013). Venoms of the Hymenoptera: biochemical, pharmacological and behavioural aspects: Elsevier.
Piek, T., Veldsema-Currie, R., Spanjer, W., & Mantel, P. (1983). Acetylcholine and an unidentified, muscle-contracting factor in the venom of the bumblebee, Bombus terrestris L. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology, 75(2), 351-356.
Qiu, Y., Choo, Y. M., Yoon, H. J., Jia, J., Cui, Z., Wang, D., . . . Jin, B. R. (2011). Fibrin (ogen) olytic activity of bumblebee venom serine protease. Toxicology and applied pharmacology, 255(2), 207-213.
Qiu, Y., Choo, Y. M., Yoon, H. J., & Jin, B. R. (2012a). Molecular cloning and antibacterial activity of bombolitin isolated from the venom of a bumblebee, Bombus terrestris. Journal of Asia-Pacific Entomology, 15(1), 21-25.
Qiu, Y., Choo, Y. M., Yoon, H. J., & Jin, B. R. (2012b). Molecular cloning and fibrin (ogen) olytic activity of a bumblebee (Bombus hypocrita sapporoensis) venom serine protease. Journal of Asia-Pacific Entomology, 15(1), 79-82.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint

Qiu, Y., Lee, K. S., Choo, Y. M., Kong, D., Yoon, H. J., & Jin, B. R. (2013). Molecular cloning and antifibrinolytic activity of a serine protease inhibitor from bumblebee (Bombus terrestris) venom. Toxicon, 63, 1-6.
Rees, J. A., Moniatte, M., & Bulet, P. (1997). Novel antibacterial peptides isolated from a European bumblebee, Bombus pascuorum (Hymenoptera, Apoidea). Insect biochemistry and molecular biology, 27(5), 413-422.
Terzo, M., Valterová, I., & Rasmont, P. (2007). Atypical Secretions of the Male Cephalic Labial Glands in Bumblebees: The Case of Bombus (Rhodobombus) mesomelasGerstaecker (Hymenoptera, Apidae). Chemistry & biodiversity, 4(7), 1466-1471.
Terzo, M., Valterova, I., Urbanova, K., & Rasmont, P. (2003). De la nécessité de redécrire les phéromones sexuelles des mâles de bourdons [Hymenoptera: Apidae, Bombini] publiées avant 1996 pour leur utilisation en analyse phylogénétique. Phytoprotection, 84(2), 39-49.
Urbanová, K., Valterová, I., Rasmont, P., & Terzo, M. (2002). Znaèkovací feromony: slovení sekretu labiální vlázy samcù Bombus magnus.[Marking pheromones of bumblebees: composition of the labial gland secretion of males of Bombus magnus]. Chemické listy, 96, 918-919.
Van Vaerenbergh, M., Debyser, G., Devreese, B., & de Graaf, D. C. (2014). Exploring the hidden honeybee (Apis mellifera) venom proteome by integrating a combinatorial peptide ligand library approach with FTMS. J Proteomics, 99, 169-178. doi:10.1016/j.jprot.2013.04.039
Van Vaerenbergh, M., Debyser, G., Smagghe, G., Devreese, B., & de Graaf, D. C. (2015). Unraveling the venom proteome of the bumblebee (Bombus terrestris) by integrating a combinatorial peptide ligand library approach with FT-ICR MS. Toxicon, 102, 81-88. doi:10.1016/j.toxicon.2013.10.002
Wan, H., Kim, B. Y., Lee, K. S., Yoon, H. J., Lee, K. Y., & Jin, B. R. (2014). A bumblebee (Bombus ignitus) venom serine protease inhibitor that acts as a microbial serine protease inhibitor. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 167, 59-64.
Williams, P. (2007). The distribution of bumblebee colour patterns worldwide: possible significance for thermoregulation, crypsis, and warning mimicry. Biological Journal of the Linnean Society, 92(1), 97-118.
Williams, P. H., An, J., Brown, M. J. F., Carolan, J. C., Goulson, D., Huang, J., & Ito, M. (2012). Cryptic Bumblebee Species: Consequences for Conservation and the Trade in Greenhouse Pollinators. PLoS ONE, 7(3), e32992. doi:10.1371/journal.pone.0032992
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint
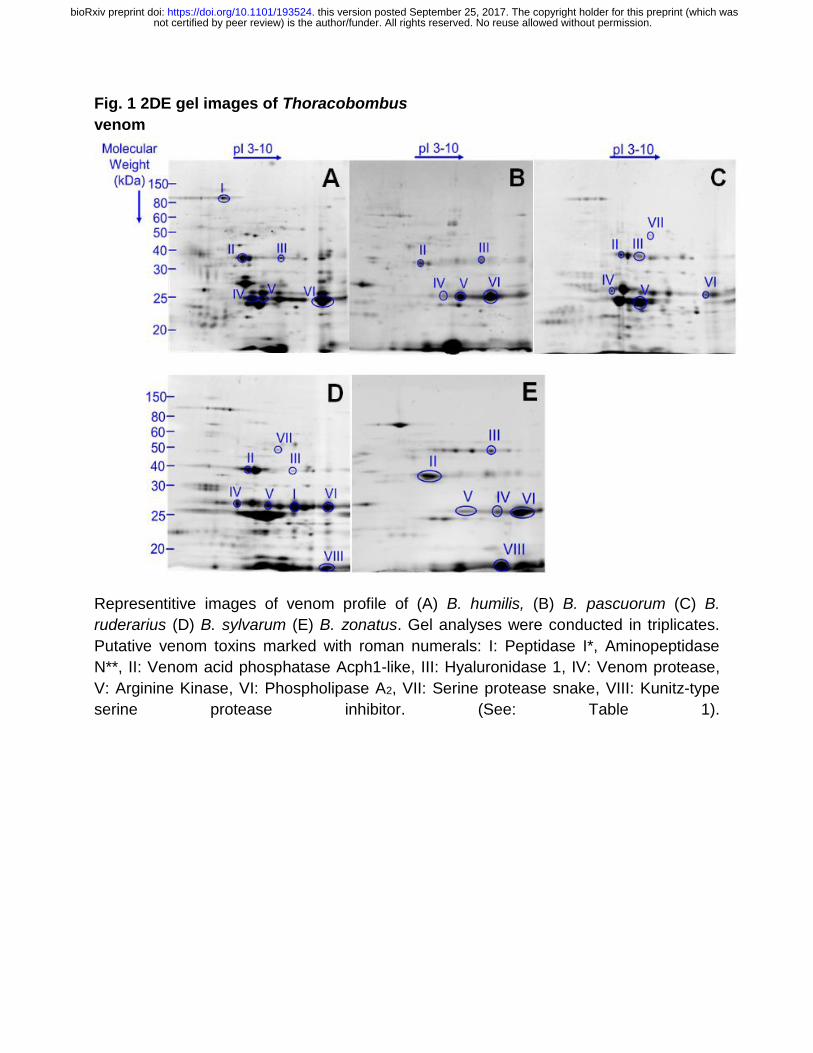
Fig. 1 2DE gel images of Thoracobombus
venom
Representitive images of venom profile of (A) B. humilis, (B) B. pascuorum (C) B.
ruderarius (D) B. sylvarum (E) B. zonatus. Gel analyses were conducted in triplicates.
Putative venom toxins marked with roman numerals: I: Peptidase I*, Aminopeptidase
N**, II: Venom acid phosphatase Acph1-like, III: Hyaluronidase 1, IV: Venom protease,
V: Arginine Kinase, VI: Phospholipase A2, VII: Serine protease snake, VIII: Kunitz-type
serine protease inhibitor. (See: Table 1).
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint
16
Fig. 2 Relative 2DE gel spot intensities of putative toxins in Thoracobombus venom.
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
P h o s p h o lip a s e A 2
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
V e n o m A c id P h o s p h a ta s e A c p h 1 - lik e
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
2 0 0
4 0 0
6 0 0
S e r in e P r o te a s e S n a k e
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
V e n o m P ro te a s e
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
P e p t id a s e s
Sp
ot
Inte
ns
ity
A m in o p e p t id a s e N
P e p t id a s e 1
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
A rg in in e K in a s e
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
5 0 0 0
1 0 0 0 0
1 5 0 0 0
2 0 0 0 0
2 5 0 0 0
K u n itz - ty p e S e r in e P ro te a s e In h ib ito r
Sp
ot
Inte
ns
ity
B.h
um
ilis
B. p
ascu
oru
m
B. ru
dera
r iu
s
B.s
ylv
aru
m
B.z
on
atu
s
0
1 0 0 0
2 0 0 0
3 0 0 0
H y a lu ro n id a s e 1
Sp
ot
Inte
ns
ity
Intensities, as calculated by PDQuest Software, are presented as mean ± standard deviation (3 replicates).
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint
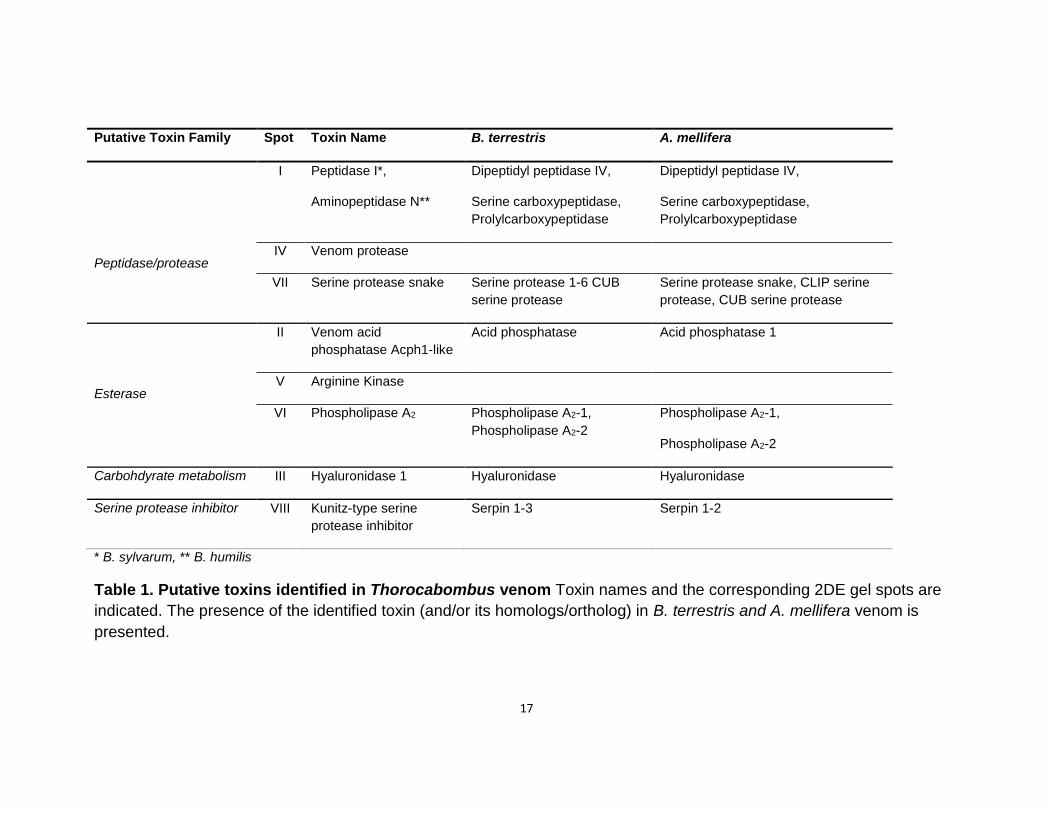
17
Putative Toxin Family Spot Toxin Name B. terrestris A. mellifera
Peptidase/protease
I Peptidase I*,
Aminopeptidase N**
Dipeptidyl peptidase IV,
Serine carboxypeptidase,
Prolylcarboxypeptidase
Dipeptidyl peptidase IV,
Serine carboxypeptidase,
Prolylcarboxypeptidase
IV Venom protease
VII Serine protease snake Serine protease 1-6 CUB
serine protease
Serine protease snake, CLIP serine
protease, CUB serine protease
Esterase
II Venom acid
phosphatase Acph1-like
Acid phosphatase Acid phosphatase 1
V Arginine Kinase
VI Phospholipase A2 Phospholipase A2-1,
Phospholipase A2-2
Phospholipase A2-1,
Phospholipase A2-2
Carbohdyrate metabolism III Hyaluronidase 1 Hyaluronidase Hyaluronidase
Serine protease inhibitor VIII Kunitz-type serine
protease inhibitor
Serpin 1-3 Serpin 1-2
* B. sylvarum, ** B. humilis
Table 1. Putative toxins identified in Thorocabombus venom Toxin names and the corresponding 2DE gel spots are
indicated. The presence of the identified toxin (and/or its homologs/ortholog) in B. terrestris and A. mellifera venom is
presented.
not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which wasthis version posted September 25, 2017. . https://doi.org/10.1101/193524doi: bioRxiv preprint


















