
Protein Folding Energetics, Kinetics and Models Oznur Tastan [email protected] Graduate Student...
50
Protein Folding Energetics, Kinetics and Models Oznur Tastan [email protected] Graduate Student Carnegie Mellon University
-
Upload
aubrie-merritt -
Category
Documents
-
view
232 -
download
0
Transcript of Protein Folding Energetics, Kinetics and Models Oznur Tastan [email protected] Graduate Student...

Protein FoldingEnergetics, Kinetics and Models
Oznur Tastan [email protected]
Graduate Student
Carnegie Mellon University

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 2
Lecture Outline
• Introduction: What is protein folding and why it is a problem?
• Globular Protein Folding Models
• Detection and characterization of denatured states, intermediates, such as the molten globule and comparison to folded states
• Kinetics and pathways
• Membrane Protein Folding Models
• 2-stage and 3-stage hypothesis
• New long range interaction hypothesis
• Summary

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 3
Lecture Outline
• Introduction: What is protein folding and why it is a problem?
• Globular Protein Folding Models
• Detection and characterization of denatured states, intermediates, such as the molten globule and comparison to folded states
• Kinetics and pathways
• Membrane Protein Folding Models
• 2-stage and 3-stage hypothesis
• New long range interaction hypothesis
• Summary

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 4
Our focus
in this lecture
http://www-nmr.cabm.rutgers.edu/academics/biochem694/2006BioChem412/Biochem.412_2-24-2006lecture.pdf

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 5
Folding is encoded in the amino acid sequence. Native state is the minimum energy state.
Addition of mercaptoethanol and urea
Removal of mercaptoethanol and urea
Native, catalytically active
state.
Native,catalytically active
state. Refolded correctly!
Unfolded; catalytically inactive.
Reduced disulfide bonds.
1/105 random chance
Anfinsen, 1973.
Anfinsen’s Experiment

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 6
The Protein Folding Problem:Writing the book of Protein Origami
http://www.idi.ntnu.no/grupper/KS-grp/microarray/slides/drablos/Fold_recognition/sld006.htm
Now collapse down hydrophobic core, and fold over Helix A to the
dotted line. Bring charged residues of ‘A’ into close proximity
of ‘B’ ..?
+membrane

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 7
How does a protein fold?Levinthal’s Paradox
Entropy
En
erg
y
Protein folding cannot be random-walk.
Simplest case: random-walk• Assume a chain of 100 amino acids.• Allow only 3 conformations.
- Possible conformations = 3100 ~ 1048
• Assume bond rotation rate 1014 sec.
- Reaching the native state would take: 1026 years ! Longer than the age of the universe!
Dill & Chan, 1997Levinthal, 1968

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 8
Why is protein folding problem difficult?
• Folding can be very fast, millisecond to second (slow folding is easier)
• Small energy changes between the denatured state to the native state ( 1-15 kcal/mol) - equivalent to the strength of a few hydrogen bonds
• The states populated along pathway are ensembles of structures
Comparison from multiple complementary techniques are required.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 9
The Three Protein Folding Models
Framework model
Nucleation condensation
model
Hydrophobic collapsemodel
http://www.makro.ch.tum.de/users/BFHZ/Scheibel/Scheibel%202003%20Bordeaux-1.pdf

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 10
Lecture Outline
• Introduction: What is protein folding and why it is a problem?
• Globular Protein Folding Models
• Detection and characterization of denatured states, intermediates, such as the molten globule and comparison to folded states
• Kinetics and pathways
• Membrane Protein Folding Models
• 2-stage and 3-stage hypothesis
• New long range interaction hypothesis
• Summary

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 11
The Native State
• A complex balance between: 1) Short-range local interactions
-intrinsic conformational preferences of the amino acids
2) Medium-range interactions
-stabilizing regions of secondary structure 3) Long-range interactions
- tertiary interactions determining the global fold
• Generally single conformation (with small fluctuations around the mean torsion angles).

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 12
Random Coil and Denatured State
“Φ,Ψ angles of each residue is sterically independent”There should not exist any
non-local interactions.
Flory’s isolated pair hypothesis
Rg= RgNv
N = Length (Residues) v = Solvent viscosity
parameter
Rg values of 28 denatured proteins
obeys the Flory’s power law.Flory, 1969.
Rg values determined by SAXS
Sosnick, T.R., et al. 2004
03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 13
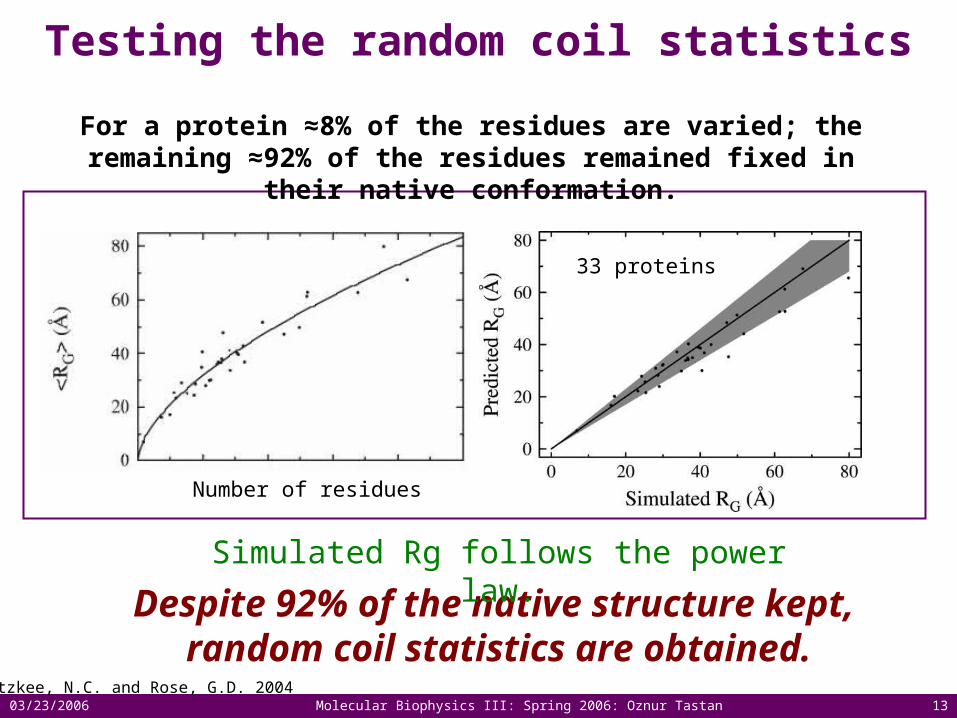
Testing the random coil statistics
Fitzkee, N.C. and Rose, G.D. 2004
Despite 92% of the native structure kept, random coil statistics are obtained.
Simulated Rg follows the power law.
33 proteins
Number of residues
For a protein ≈8% of the residues are varied; the remaining ≈92% of the residues remained fixed in their native conformation.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 14
The Denatured StateDoes Flory’s hypothesis hold?
Conformations of polyalanine chains are enumerated to test the hypothesis.
Pappu et.al 2003.
+ ={A,G,M,R,L,F,E,K,Q}
* = {J,P,O,I,o}
Flory’s hypothesis is not valid for polypeptide chains. Backbone conformations are limited by
additional steric clashes.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 15
Can we get a structure of the “denatured state”?
When the folded state breaks down:
1. The dispersion of all resonances decreases: - Extensive overlap of peaks.
2. Greater dynamic motions between residues: - Weak or eliminated NOEs between protons.
3. Ensemble of conformations: - Each NMR parameter reflects an average over a dynamic
ensemble of conformations.
Attainment of a high-resolution structure is not possible in the non-native state.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 16
NMR as a tool to study denatured states
67891011121H Chemical Shift (ppm )
105
110
115
120
125
130
15 N
Ch
emic
al S
hif
t (p
pm
)
7.08.09.010.0
112
114
116
118
120
122
124
126
128
1H Chemical Shift (ppm )
15N
Ch
emic
al S
hif
t (p
pm
)
Folded lysozyme: Unfolded lysozyme:
For small proteins, all backbone resonances can be resolved even in the denatured state.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 17
1. Measurement of NMR parameters in 15N-labeled unfolded protein
2. Comparison of NMR parameters
- unfolded with random coil parameters (sources:
- statistical analysis from unfolded peptides - random coil models (e.g. polymer model, Model-free analysis, Flory etc.)
- unfolded with folded state parameters- different degrees of unfolded states
•Chemical shifts
•Relaxation rates
•Heteronuclear NOE
•Dipolar couplings
•Scalar couplings
Which and how do we use NMR parameters?

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 18
1. Measurement of NMR parameters in 15N-labeled unfolded protein
2. Comparison of NMR parameters
- unfolded with random coil parameters (sources:
- statistical analysis from unfolded peptides - random coil models (e.g. polymer model, Model-free analysis, Flory etc.)
- unfolded with folded state parameters- different degrees of unfolded states
•Chemical shifts
•Relaxation rates
•Heteronuclear NOE
•Dipolar couplings
•Scalar couplings
Which and how do we use NMR parameters?

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 19
Persistence of native-like topology in the denatured states
Denatured proteins can preserve long range ordering, in conflict with the random-coil models.
Unf
olde
d
Folded
Shortie. et. al. 2001
SNase N-H Dipolar couplings

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 20
1. Measurement of NMR parameters in 15N-labeled unfolded protein
2. Comparison of NMR parameters
- unfolded with random coil parameters (sources:
- statistical analysis from unfolded peptides - random coil models (e.g. polymer model, Model-free analysis, Flory etc.)
- unfolded with folded state parameters- different degrees of unfolded states
•Chemical shifts
•Relaxation rates
•Heteronuclear NOE
•Dipolar couplings
•Scalar couplings
Which and how do we use NMR parameters?

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 21
Residue
0 25 50 75 100 125
R2
[s-1
]
0
2
4
6
8
10
12
Residue
0 25 50 75 100 125
Urea Water
Residue
0 25 50 75 100 125
R2
[s-1
]
0
2
4
6
8
10
12
Residue
0 25 50 75 100 125
Urea Water
1
||
int)(N
j
ji
rinsic eRiR
Random Coil Model of Segmental Motion
Residue
0 25 50 75 100 125
R2
[s-1
]
0
2
4
6
8
10
12
Residue
0 25 50 75 100 125
Urea Water
0
20 ||
x
b
xi
Ae
+ Gaussian Distributions of Deviations
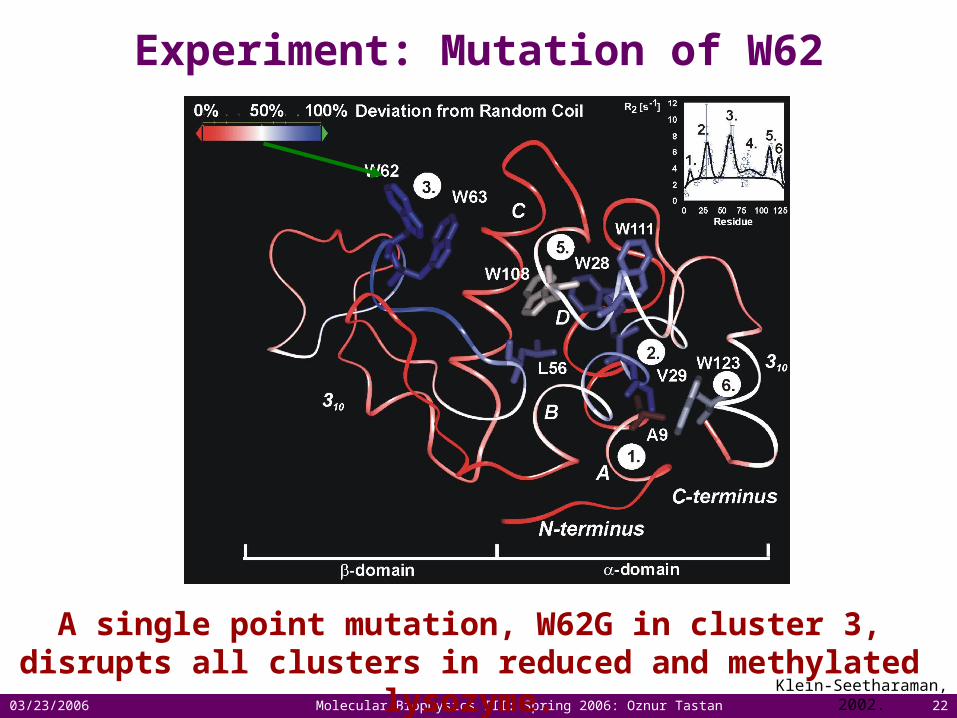
There are six clusters of residual structure in WL-SME.
1.
2.
3.
4.5.
6.
Residual structure in lysozyme
WL-SME in urea WL-SME in water
Klein-Seetharaman, 2002.
03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 22
A single point mutation, W62G in cluster 3, disrupts all clusters in reduced and methylated lysozyme.
Experiment: Mutation of W62
Klein-Seetharaman, 2002.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 23
The Molten Globule(MG) State
Molten globule is characterized by
1. Absence of specific tertiary contacts
2. presence of some secondary structure
3. Native-like compactness
4. Presence of hydrophobic core
Example: -lactalbumin
Molten globule observed in low pH

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 24
The Molten Globule(MG) State
1. Absence of specific tertiary contacts
2. Presence of some secondary structure
3. Native-like compactness
4. Presence of hydrophobic core Kuwajima, K. 1989.
native pH5.4
MG-state pH2
unfolded state (in 9M urea,pH2)

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 25
The Molten Globule(MG) State
1. Absence of specific tertiary contacts
2. Presence of some secondary structure
3. Native-like compactness
4. Presence of loosely packed hydrophobic core.
Rg
Native 15.7+0.2
MG 17.2+0.3
Unfolded 30.0+0.7
Katoka, 1997.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 26
The Molten GlobuleSignificance for Protein Folding Mechanism
Disordered polypeptide collapse into the molten globule. According to one view,
http://www.bmb.psu.edu/courses/bmb401H/Chapter7and8.pdf

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 27
How do small single-domain proteins fold?
• 20 small proteins(< 100 aa) are showed to fold :
– simple two-state folding kinetics
– show variation in their folding rates(microseconds to seconds)
– all structures has to pass the transition state in order to reach the native state
Dobson, 2003

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 28
Kinetics of Two-State Folding
ΔGŦ-D = GŦ - G°D = - RTlnkfold ΔGN-D = G°N – GŦ = - RTlnkunfold
λ = kfold + kunfold
[GdnHCl]
Chevron Plot of CI2
ln λ
kfold kunfold
An indicator of 2-state kinetics.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 29
Ф-value AnalysisCharacterization of the Transition State(TS)
Ф=1: site of mutation is native-like in TS.
Ф=0: site of mutation is unfolded in TS.
Fractional Ф value: partial structure in TS.
TS cannot be isolated or studied directly.
Reproduced form
Systematically introduce mutations in the native protein.
Infer structure of TS from the energetics of the folded state (mutant versus wild-type).
Ф = ΔΔGŦ-D / ΔΔGN-D

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 30
Transition State Analysis:Case Study I: Chymotrypsin inhibitor(CI2)
In the transition state of CI2 three residues with Ф-values >0.5 come
together :A16,L49,I57.
A hydrophobic core supporting the nucleation-condensation
mechanism

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 31
Complex pathwaysCase study II: hen lysozyme
Thus cannot be approximated with simple 2-state kinetics.
Most proteins (>100 aa) fold with observable intermediates.
lysozyme
Dill & Chan et al.1997
03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 32
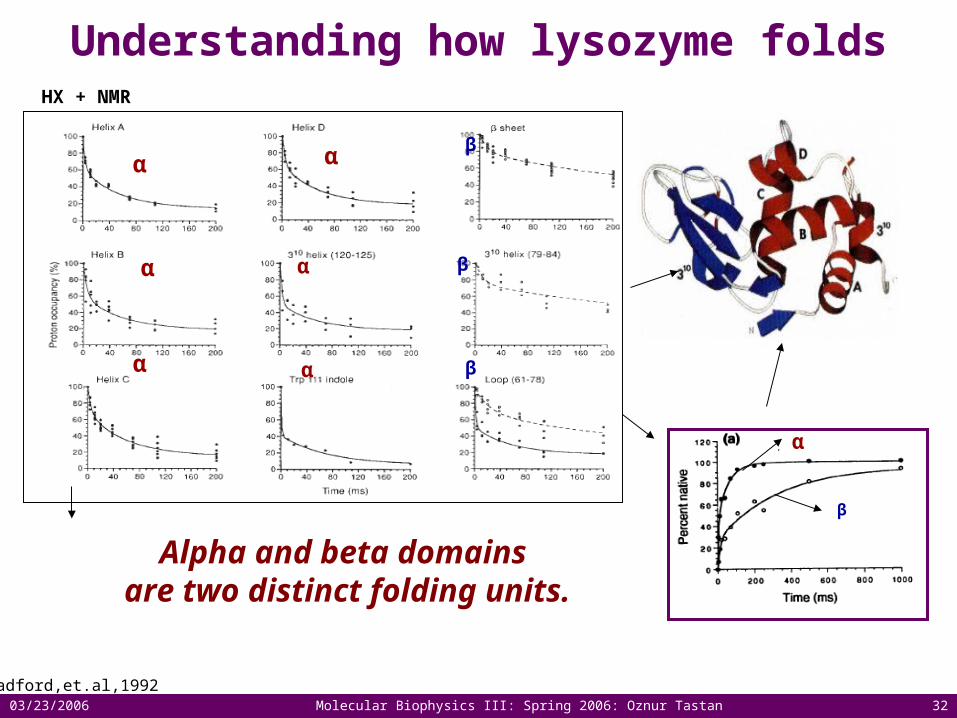
Understanding how lysozyme folds
Radford,et.al,1992
α
α
β
β
β
α
α
α
Alpha and beta domains are two distinct folding units.
α
HX + NMR
α
β

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 33
The details?
α domain is structured independently of the β
domain in the early stages of folding.
HX labeling &
EMS
Far-UV CD
Large secondary structure is formed
within the milliseconds of folding.
unprotected
β
α
Dobson,et.al. 1994
Single exponential, the α domain forms before the near UV develops.
The tertiary contacts are not fixed yet.
Near-UV CD

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 34
Intrinsic Trp flouresence
A change in the some or all of the Trp occurs in early collapse in later
intermediates and on formation of the native structure.
Quenching of flouresence by iodine
Exclusion of water in the early stages of folding
Binding of ANS
Maximal emission in the early stages.
A relatively loosely packed, condensed state exists in the
early stages of reaction.Dobson,et.al. 1994

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 35
Folding pathway of lysozyme
Very rapidly alpha domain forms.
Hydrophobic interior develops. Few tertiary interactions.
Protective structure evolves. Dynamic and
fluctuating beta domain.
Alpha and beta domains are stabilized
Major >30%Minor ~10%
Dobson,et.al. 1994
03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 36
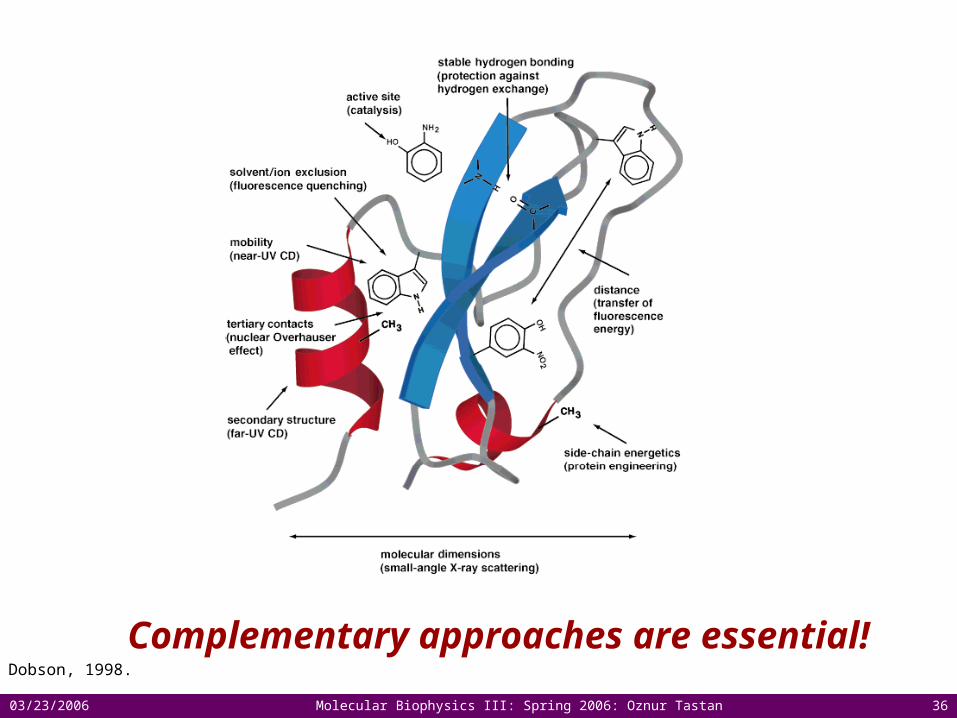
Complementary approaches are essential! Dobson, 1998.
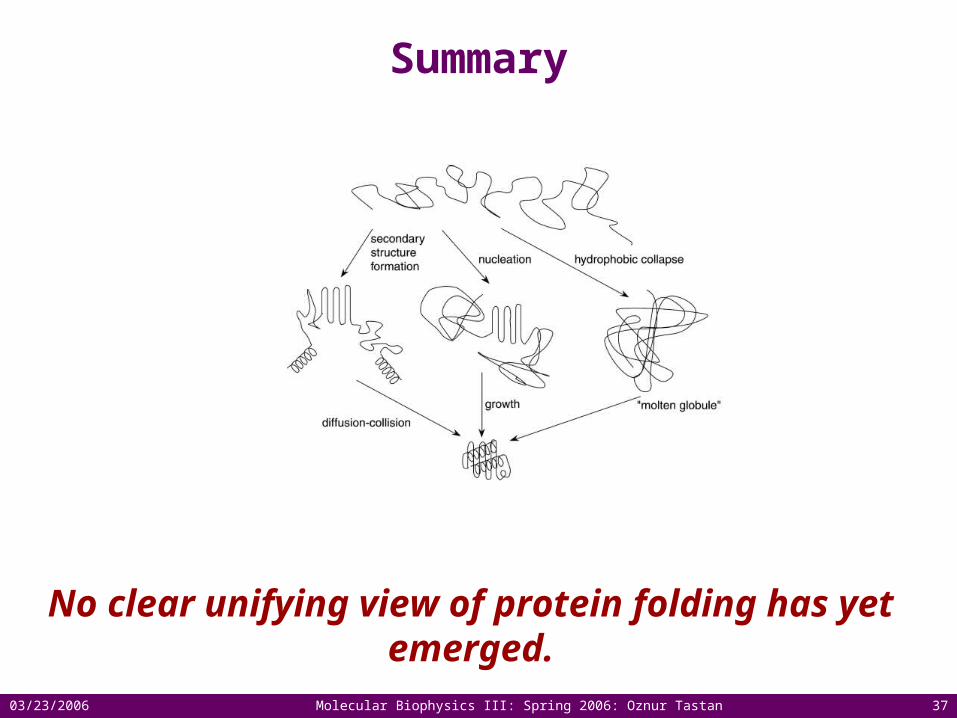
03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 37
Summary
No clear unifying view of protein folding has yet emerged.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 38
Membrane Protein FoldingModel systems
α-helical bundles β-barrels
The lipid environment
OmpABacteriorhodopsinMammalian Rhodopsin

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 39
Denaturation of bacteriorhodopsin
Effects of Urea and Guanidinium Hydrochloride:
almost none, not even on tertiary structure.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 40
Denaturation of Bacteriorhodopsin
NativeSDS
Formic acid
Secondary structure remains even in SDS.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 41
Why is it so difficult to disrupt secondary structure in membrane
proteins?

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 42
Thermodynamic considerations
The engaging of polar backbone in H-bonds is favorable.
White & Wimley,1999.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 43
Refolding of Bacteriorhodopsin in the lipid bilayers
BR can assemble in to the native structure when helices are inserted into the membrane independently.
Retinal reconstituted:
%90 of the activity regenerated.
Refolded: fragments have near-identical
helix content.
1. C-1 and C-2 in SDS 2. C-1 and C-2 +lipid 3. Retinal omitted.
Popot, 1987.
C-1C-2

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 44
The two-stage hypothesis: Independent helix intermediate
1st Stage:
Helix folds Independently
2nd Stage:
Final packing and interactions between
the helices are formed.New 3rd Stage:
Biding of prosthetic groups, folding of loops,
oligomerization…
Two stage hypothesis may not hold in all cases.
Engelman &Popot, 1990.Engelman & Popot, 2003. Figures from Klein-Seetharaman,2005.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 45
HR unfolded with AFM
Cooperative unfolding barriers are observed, in conflict with the 2-stage hypothesis.
Helix G unfolded in two stepsHelixF
Helix E unfolds with helix D
Short cytoplasmic segment
Helix C
B-C loop
Helix B
Helix A unfolds in two steps
Part of
helix E
Cisneros, 2005.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 46
Folding Core Prediction of Rhodopsin
Agreement with the mutational data (>%90 ).
Evidence for the significance of long-range interactions in the folding of
rhodopsin.
Gaussian Network Model (GNM) FIRST
Folding core lies in the EC-TM domain interface.
Rader. et.al 2004

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 47
New Model: Long Range Interaction intermediate
Klein-Seetharaman,2005.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 48
References• Anfinsen, C.B. (1973) "Principles that govern the folding of protein chains." Science 181 223-230.
• Cisneros, D.A., D. Oesterhelt, and D.J. Muller, Probing origins of molecular interactions stabilizing the membrane proteins halorhodopsin and bacteriorhodopsin. Structure, 2005. 13(2): p. 235-42.
• Dobson, C.M.Sali A., and Karplus, M., "Protein Folding: A Perspective from Theory and Experiment", Angew. Chem. Int. Ed. Eng. 37, 868-893 ( 1998).
• Dobson, C.M. "Protein Folding and Misfolding", Nature 426, 884-890 ( 2003).• Dobson, C.M., P.A. Evans, and S.E. Radford, Understanding how proteins fold: the lysozyme story so far. Trends Biochem Sci,
1994. 19(1): p. 31-7. • Engelman, D. M., Chen, Y., Chin, C. N., Curran, R., Dixon, A. M., Dupuy, A, Lee, A., Lehnert, U., Matthews, E., Reshetnyak, Y., Senes, A.,
Popot, J-L. “Membrane Protein Folding: Beyond the Two Stage Model” FEBS Lett. (2003) 555:122-5.
• Flory, P. J. (1969) Statistical Mechanics of Chain Molecules (Wiley, New York).• Fitzkee, N.C. and Rose, G.D. (2004). Reassessing random-coil statistics in unfolded proteins. Proc. Natl. Acad. Sci. 101: 12497–12502.• Klein-Seetharaman, J., Oikawa, M.,Wirmer, J., Duchardt, E., Ueda, T., Imoto, T., Smith, L.J., Dobson, C. and Schwalbe, H. (2002) Long-Range
Interactions within a Non-Native Protein. Science 295, 1719-1722. • Klein-Seetharaman, J. (2005) Dual role of interactions between membranous and soluble portions of helical membrane receptors for folding and
signaling. Trends in Pharmacological Science 26(4), 183-189 • Kuwajima, K. (1989). The molten globule state as a clue for understanding the folding and cooperativity of globular-protein structure. Proteins:
Struct. Funct. Genet. 6: 87–103.• Kataoka, M., Y. Hagihara, K. Mihara, and Y. Goto. (1993). Molten globule of cytochrome c studied by the small angle X-ray scattering. J. Mol.
Biol. 229:591-596.• Radford, S. E., Dobson, C. M. & Evans, P. A. The folding of hen lysozyme involves partially structured• intermediates and multiple pathways. Nature 358, 302-307 (1992).• Pappu, R. V. , Srinivasan, R. & Rose, G. D. (2000) Proc. Natl. Acad. Sci. USA 97, 12565-12570.• Popot, J-L and Engelman D.M."Membrane Protein Folding and Oligomerization: The Two-Stage Model“ Biochemistry (1990), 29 (17), 4031-7.• Popot J.L., Gerchman S.E., Engelman D.M. (1987) Refolding of bacteriorhodopsin in lipid bilayers. A thermodynamically
controlled two-stage process. J. Mol. Biol. 198:655-76• Shortle D, Ackerman MS. (2001) Persistence of native-like topology in a denatured protein in 8 M urea. Science. Jul 20;293(5529):487-9. • White S. H. and Wimley, W. C. (1999). Membrane protein folding and stability: Physical principles. Annu. Rev. Biophys. Biomol. Struct. 28:319-
365. • http://www.otago.ac.nz/humannutrition/dietetics/gfx/philosophy.jpg, March 22, 2006.
• http://www-nmr.cabm.rutgers.edu/academics/biochem694/2006BioChem412/Biochem.412_2-24-2006lecture.pdf, March 22, 2006.
• http://www.makro.ch.tum.de/users/BFHZ/Scheibel/Scheibel%202003%20Bordeaux-1.pdf, March 22, 2006.

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 49
Acknowledgements
Dr. Judith Klein-Seetharaman
Dr. Sanford Leuba
&
The class of MB3 (Spring 2006)

03/23/2006 Molecular Biophysics III: Spring 2006: Oznur Tastan 50
QUESTIONS?
http://www.otago.ac.nz/humannutrition/dietetics/gfx/philosophy.jpg


















