
Prezentacja programu PowerPoint - protein.pl · dla genetyka funkcje wyznacza fenotyp mutanta, dla...
57
Molekularne przełączniki - nie wszystkie procesy przebiegają w komórce równocześnie – część działa cyklicznie, kontrolując zachowanie komórki i jej reakcje w zależności od różnorodnych sygnałów - takie białka, szczególnie często spotykane w obrębie szlaków przekazywania sygnałów, nazywane są molekularnymi przełącznikami - główną klasą MP są GTPazy (białka G), kontrolujące włączanie/wyłączanie większości procesów komórkowych
-
Upload
phungkhanh -
Category
Documents
-
view
217 -
download
0
Transcript of Prezentacja programu PowerPoint - protein.pl · dla genetyka funkcje wyznacza fenotyp mutanta, dla...

Molekularne przełączniki
- nie wszystkie procesy przebiegają w komórce
równocześnie – część działa cyklicznie,
kontrolując zachowanie komórki i jej reakcje
w zależności od różnorodnych sygnałów
- takie białka, szczególnie często spotykane
w obrębie szlaków przekazywania sygnałów,
nazywane są molekularnymi przełącznikami
- główną klasą MP są GTPazy (białka G),
kontrolujące włączanie/wyłączanie większości
procesów komórkowych

GNBP - guanine nucleotide binding protein
GAP - GTPase activating protein
GEF - guanine nucleotide exchange factor
GDI - guanine nucleotide dissociation inhibitor
GPR - G protein regulatory motif
ATP - energia dla enzymatycznych
reakcji metabolicznych, fosforylacji
w regulacji wewnątrzkomórkowej
i ruchu motorów molekularnych
GTP - głównie używany w regulacji
procesów z udziałem GNBP (z wyjąt-
kiem dynaminy, septyny, tubuliny
i czynnika elongacyjnego G)
Białka G

– zbudowane z podjednostek , i
– outside-in signaling
Guanine nucleotide binding proteins
– małe białka G, GNBP
– przekazywanie sygnałów wewnątrz komórki
– przełączniki molekularne
Heterotrimeryczne białka G (100 kDa)
Monomeryczne białka G (20-29 kDa)

Nadrodzina Ras
Ras podziały i różnicowanie
komórek
Arf/Sar tworzenie pęcherzyków,
aktywacja PLD
Rab
transport pęcherzykowy
i sekrecja
Ran transport jądrowy,
kontrola cyklu komórkowego
Rho reorganizacja cytoszkieletu,
podziały komórek

Small GTP-binding proteins
of the Rho subfamily (Rho,
Rac, Cdc42) function to
organize the actin
cytoskeleton, including:
filopodium extension,
lamellipodium formation,
generation of actin stress
fibers, focal adhesions
GTPazy z rodziny Rho

Rho GTPases
Cdc42 cells send out
exploratory filopodia
RhoA formation of actin stress fibers
and focal adhesions,
microtubule stabilization
Rac1 extensions of broad
sheet-like lamellipodia,
destabilization of microtubules

membrane
GAP – GTPase
activating protein
GEF – guanine
nucleotide exchange
factor
GDI – guanine
nucleotide
dissociation inhibitor
G protein
cycle
biological
response
effector
proteins
GEF
GDP
GTP
signal
R
GAP
GDP
GTP GDP
inactive
GD
I Rho
Rho GTP
active

http://www.sciencemag.org
Rho GTPases
amplification
integration
time control
precision (No. of genes)
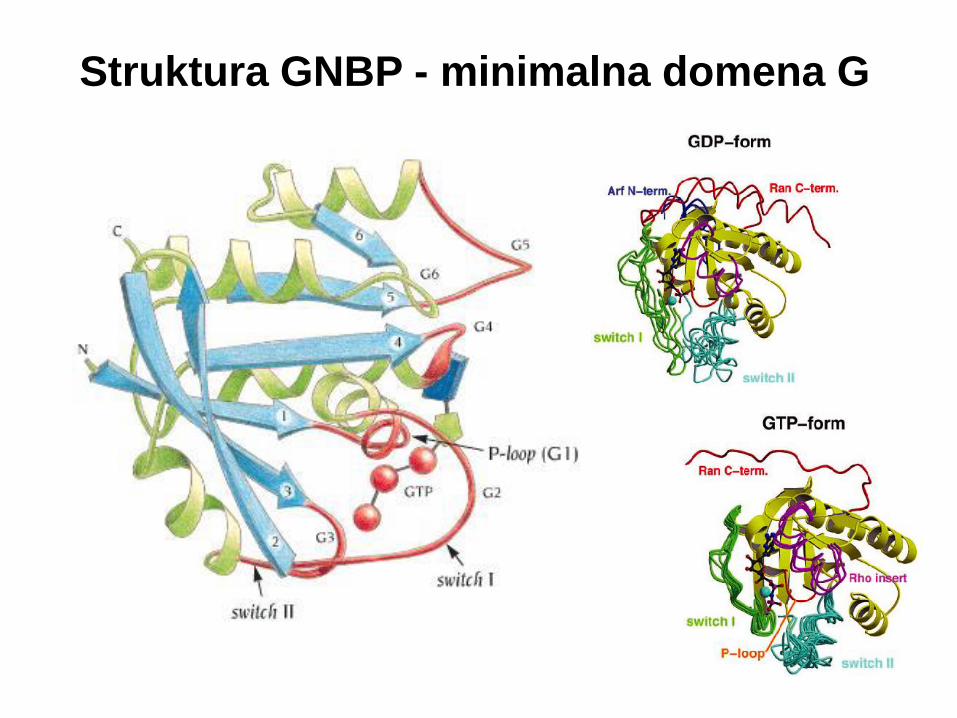
Struktura GNBP - minimalna domena G

Uniwersalny przełącznik
wiązanie γ-fosforanu
przez grupy NH
łańcucha głównego
zachowywanych
ewolucyjnie reszt Thr35
i Gly60
uwolnienie grupy
fosforanowej powoduje
lokalną relaksację,
przekładającą się na
zmianę konformacyjną
pętli switch I i II
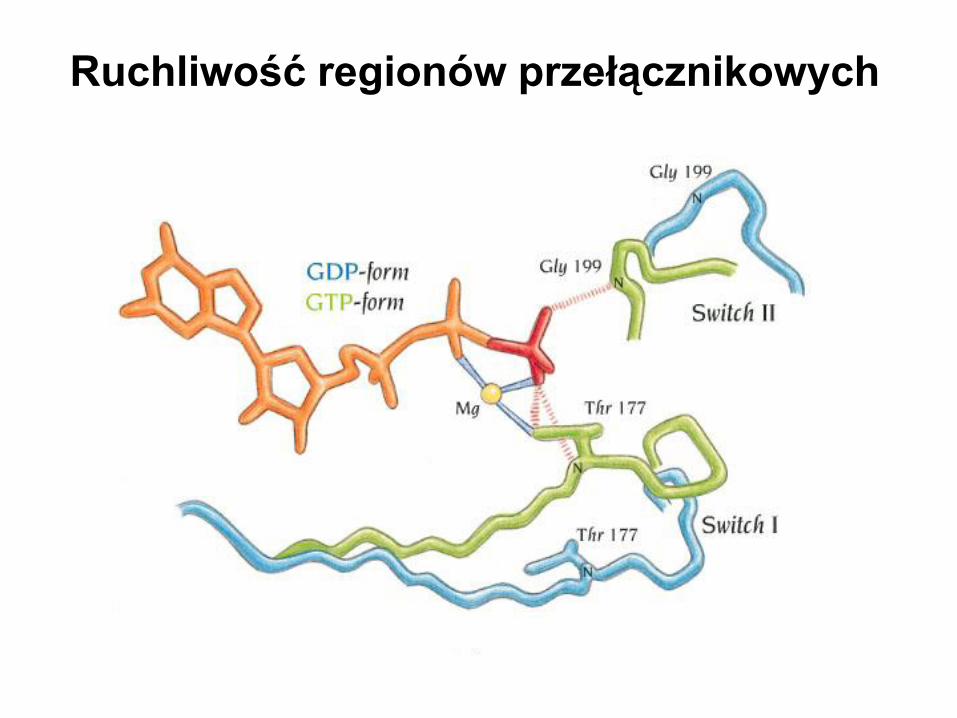
Ruchliwość regionów przełącznikowych

Ruchliwość regionów przełącznikowych

Vetter & Wittinghofer, 2000
G domain
Grupa gamma fosforanowa określa strukturę regionów przełącznikowych

Zmiany konformacyjne w białku Ran
w czasie hydrolizy GTP do GDP
Domena G

Dodatkowe domeny w GNBP i ich lokalizacja
względem domeny G
Rho - dodatkowy
13 aa, -helikalny
insert
Gi - niezależnie
zwijająca się
domena -helikalna
hGBP1 (human guanylate
binding protein) - wiele
dodatkowych elemen-
tów II rzęd. i C-ter-
minalna domena typu
coiled-coil
Czynnik elongacyjny Tu (EF-Tu), czynnik inicjacyjny (IF2/eIF5B),
czynnik elongacyjny G (EF-G) - mają odpowiednio dwie, trzy lub cztery
dodatkowe domeny

Mnogość oddziaływań domeny G z efektorami
Regiony przełącznikowe ‘switch’ są zawsze przynajmniej częściowo
zaangażowane w wiązanie efektora
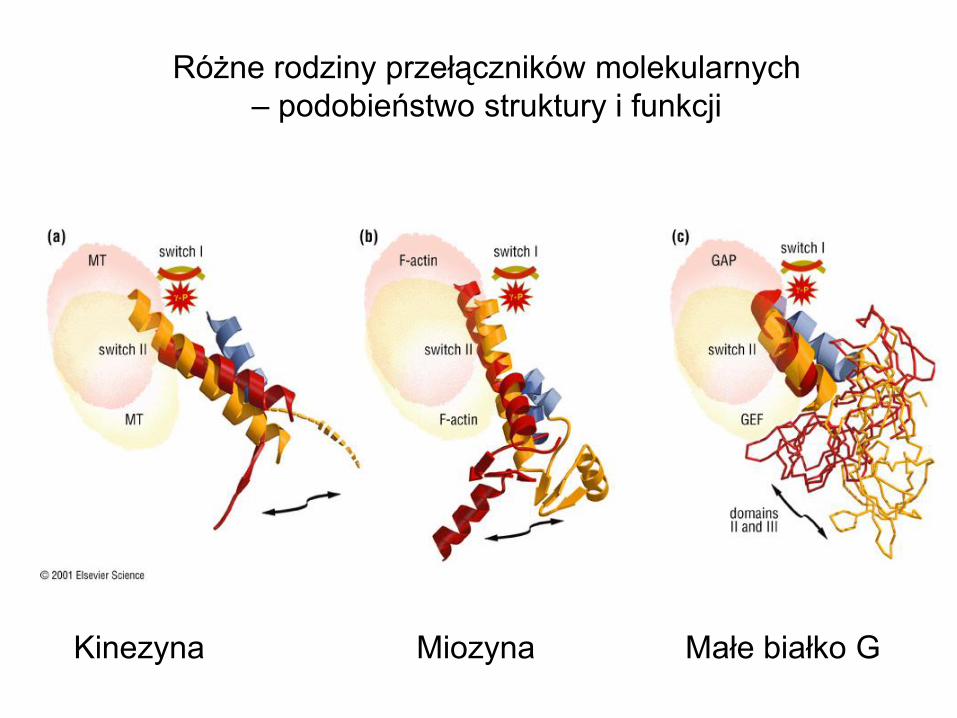
Różne rodziny przełączników molekularnych
– podobieństwo struktury i funkcji
Kinezyna Miozyna Małe białko G

Model for the motor
actions of muscle myosin

Models for the motor
actions of muscle kinesin

membrane
GAP – GTPase
activating protein
GEF – guanine
nucleotide exchange
factor
GDI – guanine
nucleotide
dissociation inhibitor
G protein
cycle
biological
response
effector
proteins
GEF
GDP
GTP
signal
R
GAP
GDP
GTP GDP
inactive
GD
I Rho
Rho GTP
active

Schemat działania białka GEF
Seria szybkich i odwracalnych reakcji:
- binarny kompleks GNBP-nukleotyd
- trimeryczny kompleks GNBP-nukleotyd-GEF
- binarny kompleks GNBP-GEF
Równowaga jest przesuwana na korzyść
związanego GDP czy GTP przez
odpowiednie powinowactwo GDP i GTP
do GNBP; stężenie nukleotydów; powino-
wactwo i stężenie dodatkowych białek (jak
efektory które przesuwają równowagę
w stronę formy związanej z GTP
- strukturalnie białka GEF nie są spokrewnione
i szczegóły „zwalniania” GDP są różne. Jest
kilka mechanistycznych podobieństw:
oddziaływanie ze switch I i II oraz dostarczenie
reszt w obrębie pętli P.
- nie jest jasne, jaka jest kolejność wydarzeń
prowadzących do wymiany GDP/GTP, ani
która cześć nukleotydu jest zwalniana
pierwsza – zasada czy fosforan
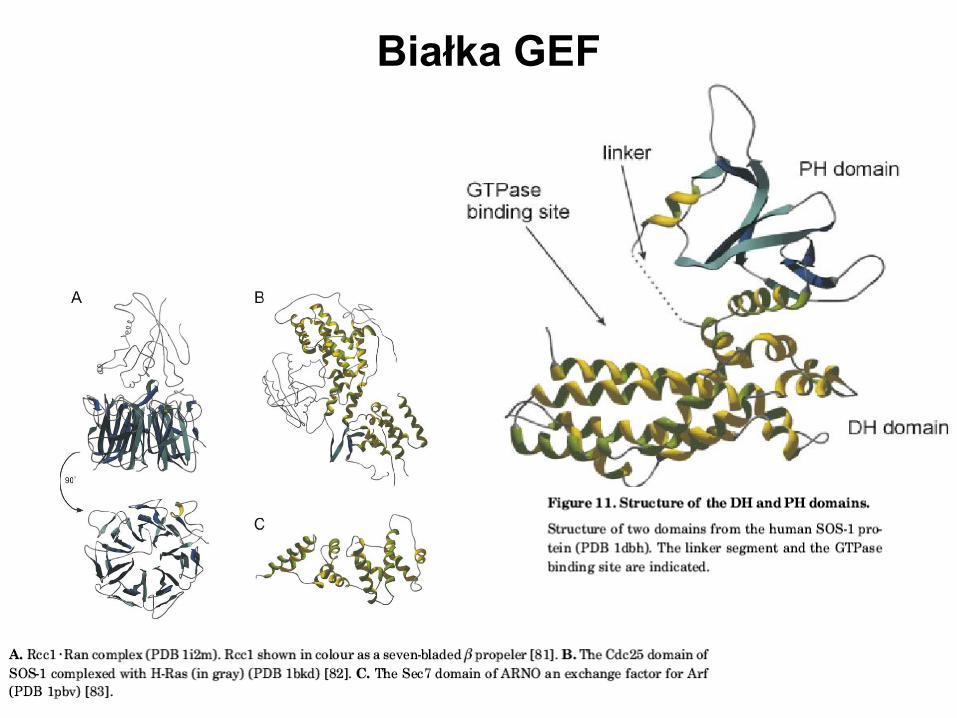
Białka GEF

GDI – inhibitor dysocjacji nukleotydu
- zupełnie różne sposoby
zwinięcia poznanych GDI
- wymagają do wiązania
prenylowanego końca GNBP
- kompleks GNBP-GDI tworzy
rezerwuar komórkowy GNBP
i pozwala na jego transport
do różnych błon w obrębie
komórki
- stąd rola GDI jako inhibitora
zdaje się być jedynie
przypadkową konsekwencją
Figure 7. Guanine nucleotide dissociation inhibitors. A. Rac2·RhoGDI (LyGDI) complex (PDB 1ds6). C-
terminal immunoglobulin-like domain is designated LyC, N-terminal - LyN, linker recognized by ICE
protease – ICE. B. Surface and electrostatic potential of LyGDI. Positively charged (basic) regions in blue
and negatively charged (acidic) ones in red. The hydrophobic cavity responsible for binding of the C-
terminally attached isoprenyl moiety is also shown. The switch I (SwI) and switch II (SwII) regions are
buried in the interaction site. C. RabGDI (PDB 1gnd). The GCD domain responsible for Rab binding is
shown.

membrane
GAP – GTPase
activating protein
GEF – guanine
nucleotide exchange
factor
GDI – guanine
nucleotide
dissociation inhibitor
G protein
cycle
biological
response
effector
proteins
GEF
GDP
GTP
signal
R
GAP
GDP
GTP GDP
inactive
GD
I Rho
Rho GTP
active

• GAP dostarcza reszt stabilizujących rejony przełącznikowe I i II, a głównie resztę Arg
stabilizującą Gln-61 (Gln-63 w RhoA), powodując tym samym odpowiednią orientację
cząsteczki wody do hydrolizy -fosforanu. W większości przypadków palec argininowy
(arginine-finger) jest niezbędny, ale są również inne kluczowo ważne dla aktywności
typu GAP reszty. Dla przykładu domena BH p85PI3-kinazy (BHPI3-K) zawiera
zachowywaną Arg, wiąże się do Rho, ale nie posiada aktywności typu GAP.
Wyjątek stanowią: Rap,
PSF, hGBP-1septyna,
dynamina gdzie zamiast
Gln występuje Thr, His,
reszta hydrofobowa
Domena GAP

Mechanizm działania GAP
GAP
GTPaza
GTP
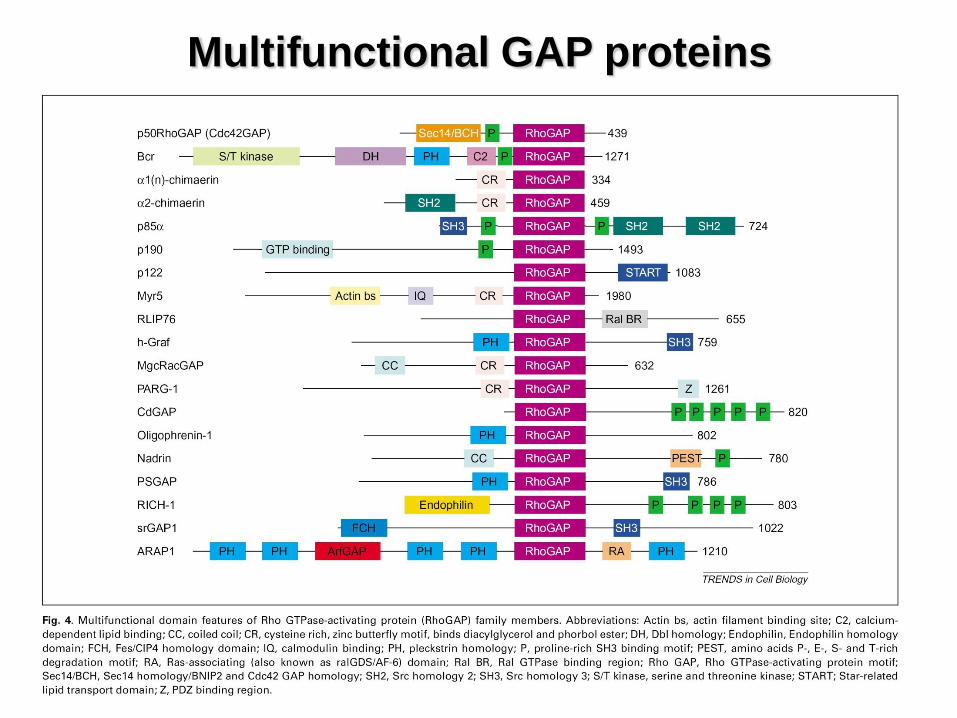
Multifunctional GAP proteins
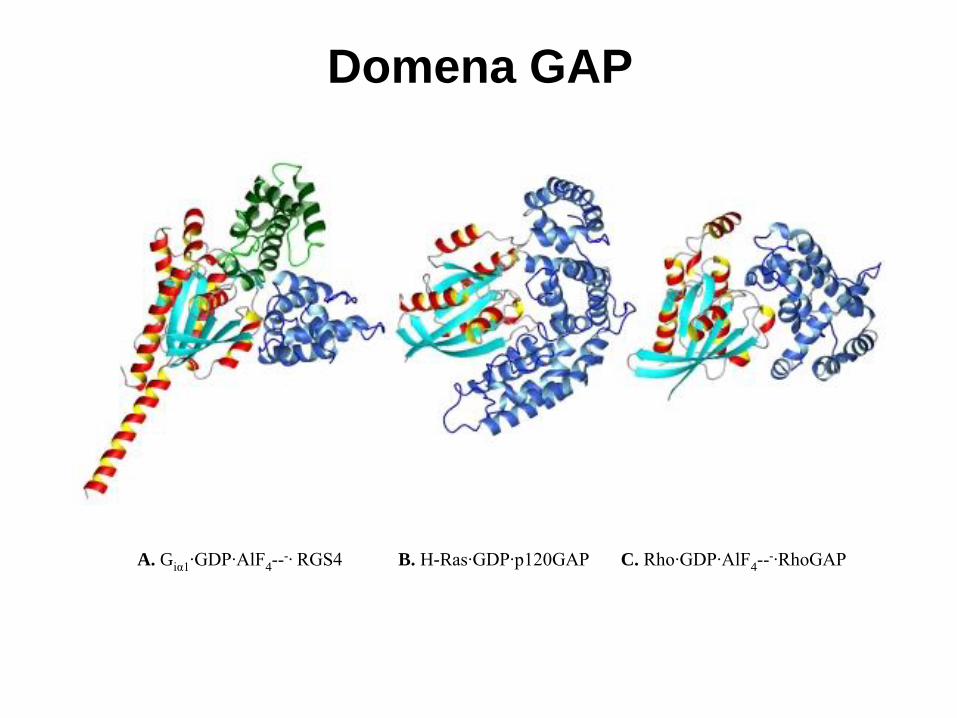
A. Giα1·GDP·AlF4-· RGS4 B. H-Ras·GDP·p120GAP C. Rho·GDP·AlF4
-·RhoGAP
Domena GAP

Bardzo podobna architektura regionów ‘switch’ I i II, podobna lokalizacja reszty
glutaminy i nałożenie cis-Arg z G , trans-Arg z Ras i RhoGAP, oraz reszty
tyrozyny z Ran.
Domena GAP
Nałożenie kompleksów białkowych:
Ran–RanBP1–RanGAP (czerwony)
Rho–RhoGAP (pomarańczowy)
Ras–RasGAP (zielony)
G –RGS (niebieski)

RasGAP – zółty, RhoGAP – czerwony,
RacExoS – niebieski, G RGS
Domena GAP

Toksyna ExoS z Pseudomonas aeruginosa
inaktywuje Rho „udając” GAP
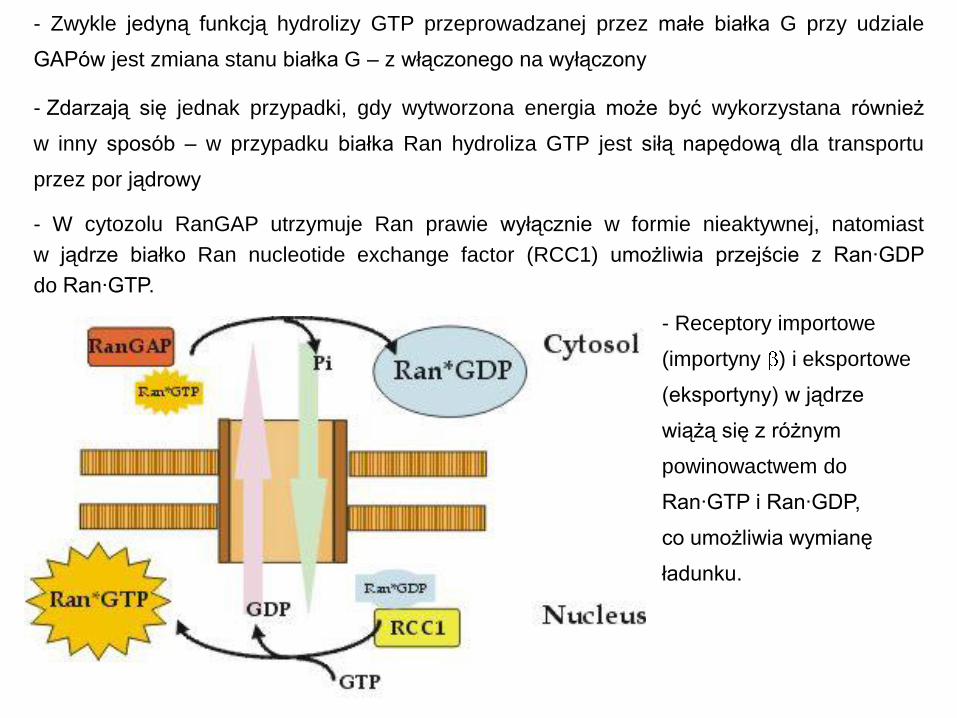
- Receptory importowe
(importyny ) i eksportowe
(eksportyny) w jądrze
wiążą się z różnym
powinowactwem do
Ran·GTP i Ran·GDP,
co umożliwia wymianę
ładunku.
- Zwykle jedyną funkcją hydrolizy GTP przeprowadzanej przez małe białka G przy udziale
GAPów jest zmiana stanu białka G – z włączonego na wyłączony
- Zdarzają się jednak przypadki, gdy wytworzona energia może być wykorzystana również
w inny sposób – w przypadku białka Ran hydroliza GTP jest siłą napędową dla transportu
przez por jądrowy
- W cytozolu RanGAP utrzymuje Ran prawie wyłącznie w formie nieaktywnej, natomiast
w jądrze białko Ran nucleotide exchange factor (RCC1) umożliwia przejście z Ran·GDP
do Ran·GTP.
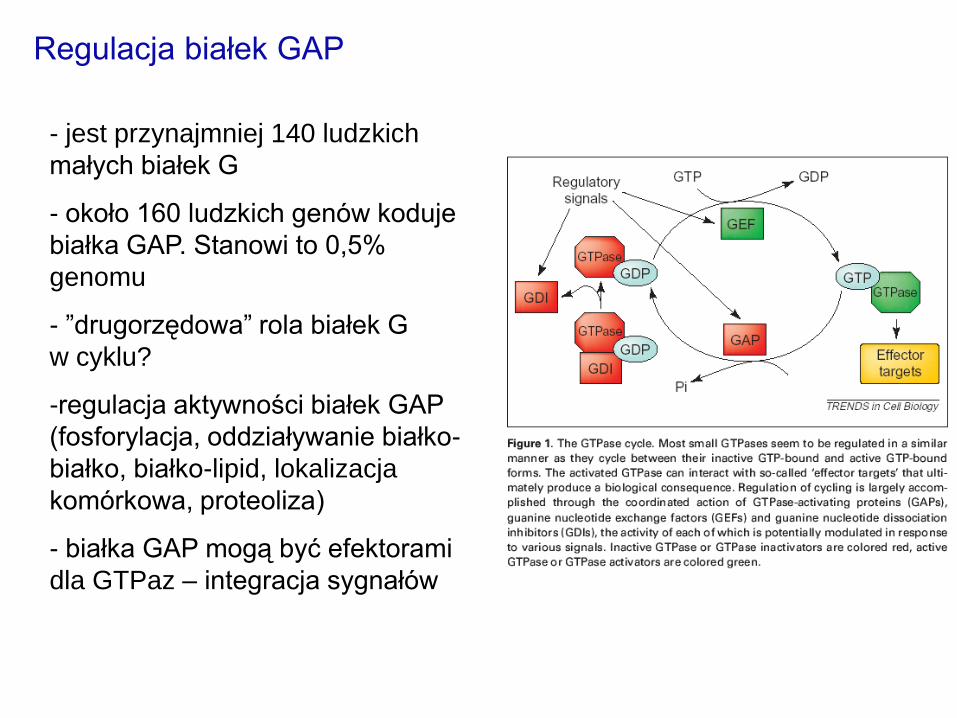
- jest przynajmniej 140 ludzkich
małych białek G
- około 160 ludzkich genów koduje
białka GAP. Stanowi to 0,5%
genomu
- ”drugorzędowa” rola białek G
w cyklu?
-regulacja aktywności białek GAP
(fosforylacja, oddziaływanie białko-
białko, białko-lipid, lokalizacja
komórkowa, proteoliza)
- białka GAP mogą być efektorami
dla GTPaz – integracja sygnałów
Regulacja białek GAP

-aktywność p190-B
RhoGAP jest stymulowana
oddziaływaniem z Rnd3 –
kaskadowe oddziaływanie
GTPaz mediowane przez
białka GAP, podobnie jak
w przypadkach opisanych
dla białek GEF
- wewnątrzcząsteczkowo:
poprzez domenę P (w p120
RasGAP)
Regulacja poprzez oddziaływanie z białkami
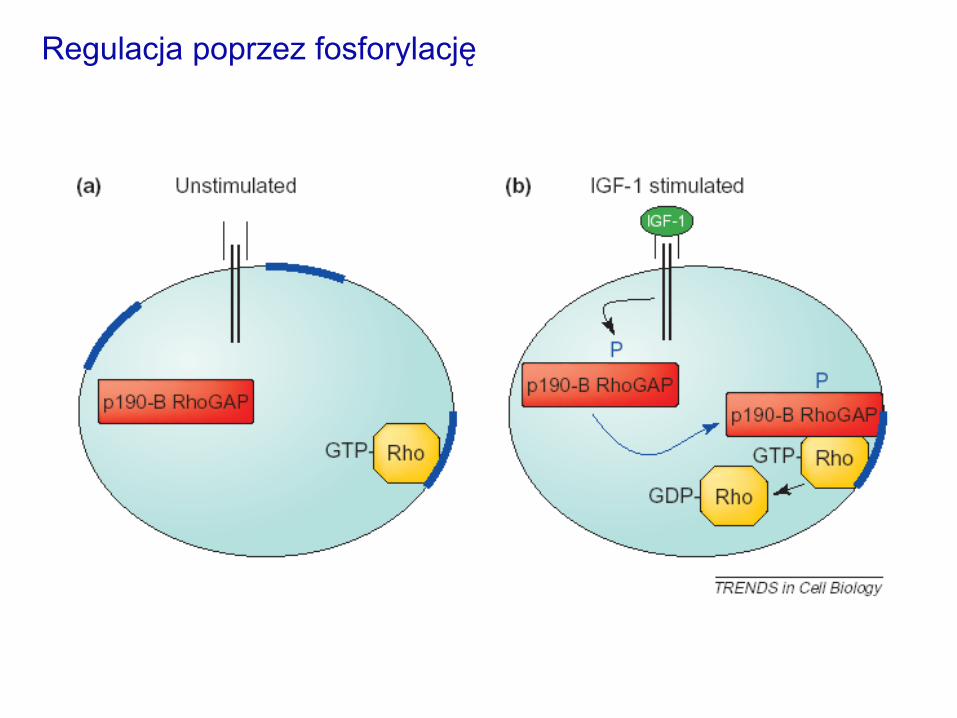
Regulacja poprzez fosforylację

http://www.sciencemag.org
Regulacja białek GAP
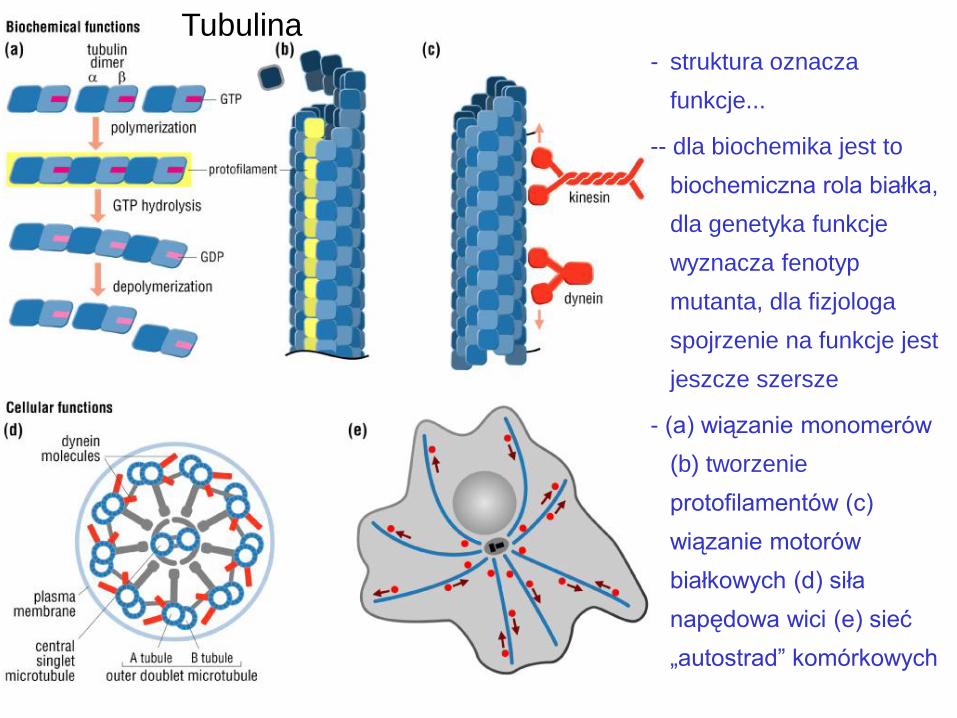
Tubulina - struktura oznacza
funkcje...
-- dla biochemika jest to
biochemiczna rola białka,
dla genetyka funkcje
wyznacza fenotyp
mutanta, dla fizjologa
spojrzenie na funkcje jest
jeszcze szersze
- (a) wiązanie monomerów
(b) tworzenie
protofilamentów (c)
wiązanie motorów
białkowych (d) siła
napędowa wici (e) sieć
„autostrad” komórkowych

Wiązanie substratu (MAPKK-2)
do czynnika L toksyny wąglika
(anthrax toxin lethal factor)
Rozpoznanie, komplementarność i centra aktywne
- miejsca wiążące ligandy i centra
aktywne są tworzone w trakcie
zwijania białka poprzez nieidealne
pakowanie się łańcuchów bocznych
- regiony te tworzą mikrośrodowiska,
które znacznie różnią się
od otaczającego rozpuszczalnika (jeśli
reszty budujące kieszeń są hydrofobowe,
jej wnętrze przypomina rozpuszczalnik
organiczny i może ona wiązać np. lipidy)
- kompromis energetyczny kumulacji
reszt o tych samych własnościach
(np. przy tworzeniu naładowanych łatek)
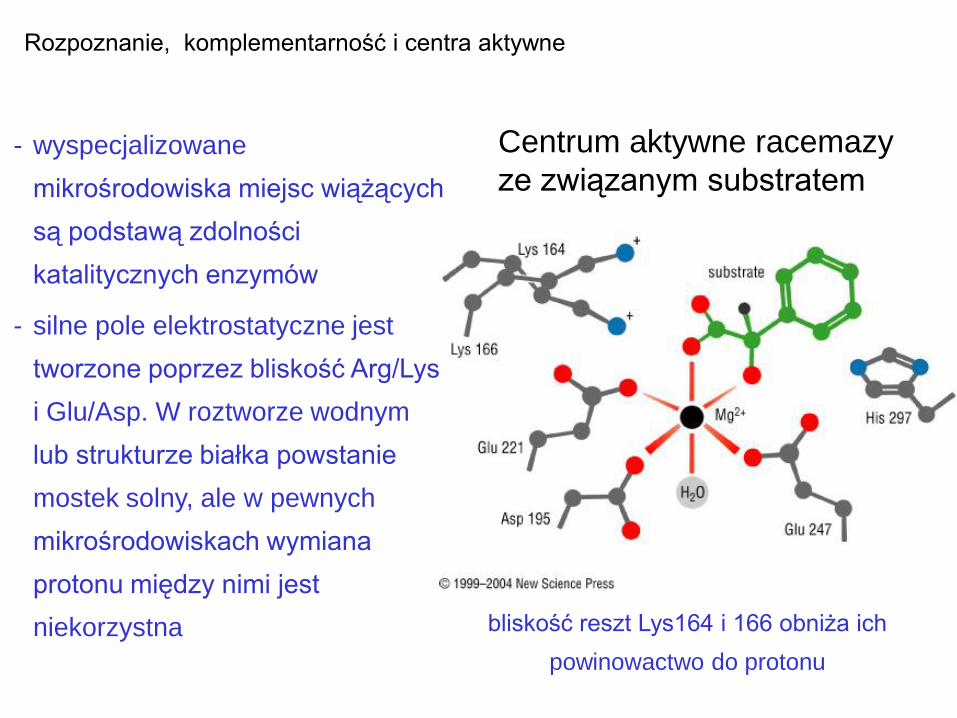
Centrum aktywne racemazy
ze związanym substratem
Rozpoznanie, komplementarność i centra aktywne
- wyspecjalizowane
mikrośrodowiska miejsc wiążących
są podstawą zdolności
katalitycznych enzymów
- silne pole elektrostatyczne jest
tworzone poprzez bliskość Arg/Lys
i Glu/Asp. W roztworze wodnym
lub strukturze białka powstanie
mostek solny, ale w pewnych
mikrośrodowiskach wymiana
protonu między nimi jest
niekorzystna bliskość reszt Lys164 i 166 obniża ich
powinowactwo do protonu

Białka są cząstkami elastycznymi
- najszybsze ruchy
to wibracje atomów
i rotacja grup metylowych
- tranzycje z jednego
rodzaju zwinięcia
do drugiego zachodzą
niezwykle rzadko
- fluktuacje konformacyjne
w strukturze domen
zachodzą jedynie lokalnie
(ligandy mogą powodować
lokalne uporządkowanie,
zwinięcie się fragmentu białka)

Izomeraza triozofosforanowa
- 8 reszt 10Å
- po związaniu liganda, pętla
„na dwóch zawiasach” zamyka
miejsce wiązania i wchodzi z nim
w interakcje, jednocześnie
osłaniając ligand
od rozpuszczalnika
- ruchy pozostałej części białka są
dużo mniejsze
(„rigid-body movement”)

Mioglobina kaszalota
zacieniowana zgodnie
ze stopniem elastyczności
-białka nazwano cząsteczkami
„półciekłymi” – ruch atomów
większy niż w ciałach stałych
(np. NaCl), ale mniejszy niż
w wodzie
-im ciemniejsze zacieniowanie,
tym bardziej sztywny atom
-powierzchnia nie jest jednakowo
dynamiczna
-grupy metylowe i reszty
aromatyczne wykazują ruchy
kolektywne

Aminotransferaza Asp
forma zamknięta i otwarta
-ruchy indukowane wiązaniem liganda
– o najwyższej wadze funkcjonalnej
-rearanżacja pojedynczej reszty vs.
ruch całej domeny
-indukowane dopasowanie –
związanie substratu powoduje
przejście z formy nieaktywnej
w aktywną (w tym przypadku z 10o
przesunięciem mniejszej domeny
w kierunku głównej części cząsteczki)
siłą sprawczą dla induced fit
jest mostek solny między
Arg domeny mobilnej
a grupą karboksylową
liganda (Asp)

Różnice w temperaturowej zależności aktywności dehydrogenazy 3-
fosforanu D-gliceraldehydu (GAPDH)
z dwóch organizmów są pochodną ich sztywności
Różna elastyczność/sztywność białek
w zależności od ich funkcji i pochodzenia (białka termofilne)
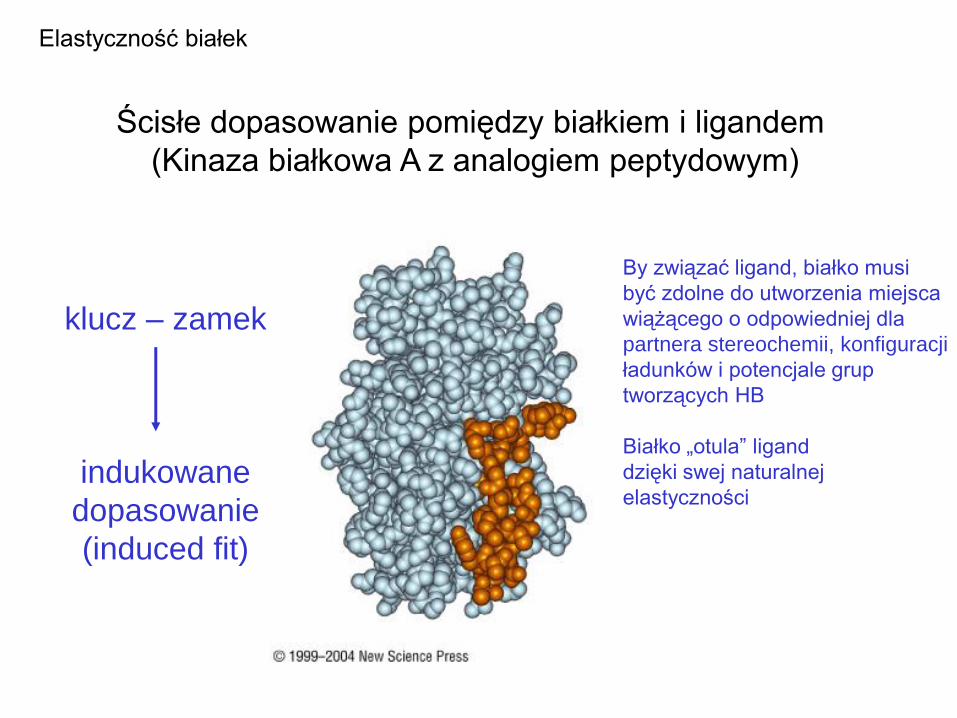
Ścisłe dopasowanie pomiędzy białkiem i ligandem
(Kinaza białkowa A z analogiem peptydowym)
Elastyczność białek
klucz – zamek
indukowane
dopasowanie
(induced fit)
By związać ligand, białko musi
być zdolne do utworzenia miejsca
wiążącego o odpowiedniej dla
partnera stereochemii, konfiguracji
ładunków i potencjale grup
tworzących HB
Białko „otula” ligand
dzięki swej naturalnej
elastyczności

proteaza HIV w kompleksie z trzema różnymi inhibitorami
powodującymi zamknięcie „klapy”
(a) haloperidol (b) crixivan (c) peptydowy analog substratu
-elastyczność białek zapewnia możliwość dopasowania
konformacyjnego do wiązanego liganda (pod warunkiem,
że dojdzie do utworzenia wystarczająco wielu korzystnych oddziaływań,
przewyższających energetyczny koszt dopasowania strukturalnego)
Dopasowanie białka do różnych ligandów
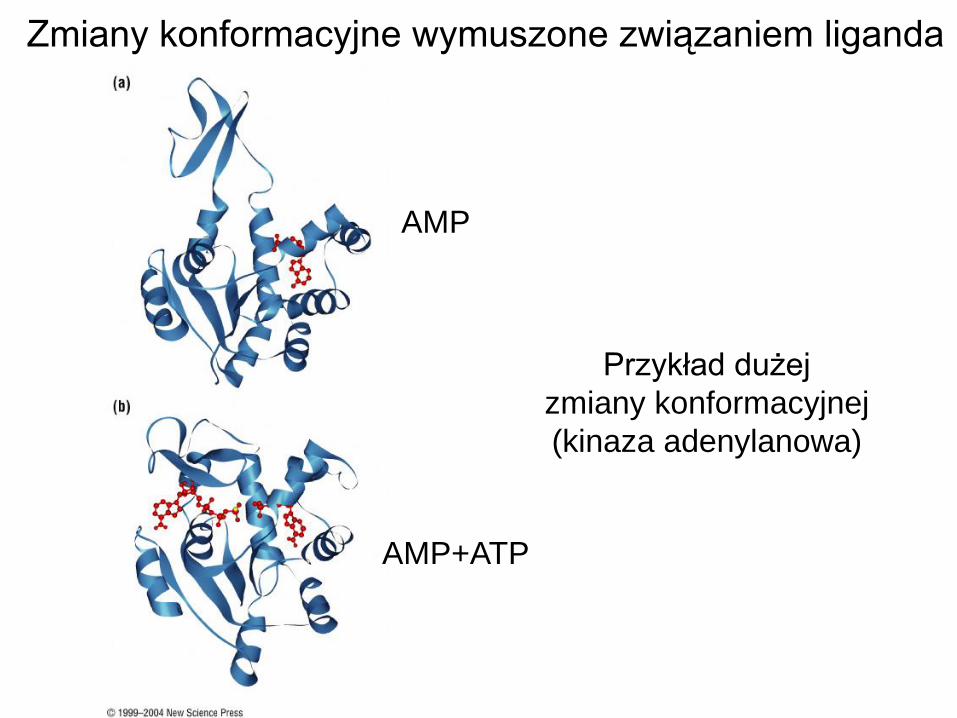
Przykład dużej
zmiany konformacyjnej
(kinaza adenylanowa)
AMP
AMP+ATP
Zmiany konformacyjne wymuszone związaniem liganda

Kompleks hGH z dwoma cząsteczkami receptora,
dwie niezależne powierzchnie oddziaływania hGH
z dwiema identycznymi cząsteczkami receptora
-oddziaływanie poprzez jedną dużą
powierzchnię zagrzebania (setki Å2)
lub kilka oddzielonych obszarów
-trudne do przewidzenia (setki kontaktów)
-ścisłe dopasowanie,
wiele punktów kontaktu
-najczęściej ligand wiązany jest przez
wystające pętle lub w obrębie dużego
zagłębienia (dodatkowe dopasowanie
kształtu), ale zdarzają się też całkiem
płaskie interfejsy oddziaływania
Miejsca wiązania makrocząsteczek – wklęsłe, wypukłe lub płaskie

Dwa kompleksy białko-DNA
(a) represor genu toksyny dyfterytu (rozpoznanie przez helisa-zwrot-helisa)
(b) czynnik transkrypcyjny Gal4 (rozpoznanie przez pętlę stabilizowaną przez Zn)
Miejsca wiązania – wystające helisy i pętle

cyt P450 ze związanym substratem
Wiązanie liganda we wnętrzu
„kieszeni” białka
-elastyczność białek
pozwala znaleźć się w ich
wnętrzu bardzo dużym ligandom
-miejsce wiązania liganda często
można zidentyfikować jako „jamkę”
w strukturze wolnego białka

Dimeryczna dehydrogenaza
bakteryjna 3-izopropylomaleinianu
Położenie miejsc wiążących
Miejsca katalityczne często
występują na styku domen
strukturalnych
i/lub podjednostek

Słabe oddziaływania prowadzą
do łatwej wymiany partnerów
w ścieżce sygnalizacyjnej
(partner swapping)
STAT – signal transducer and
activator of transcription

Wymiana domen w białku kapsydu wirusa papilloma
służy stabilizacji trimeru (domain swapping)
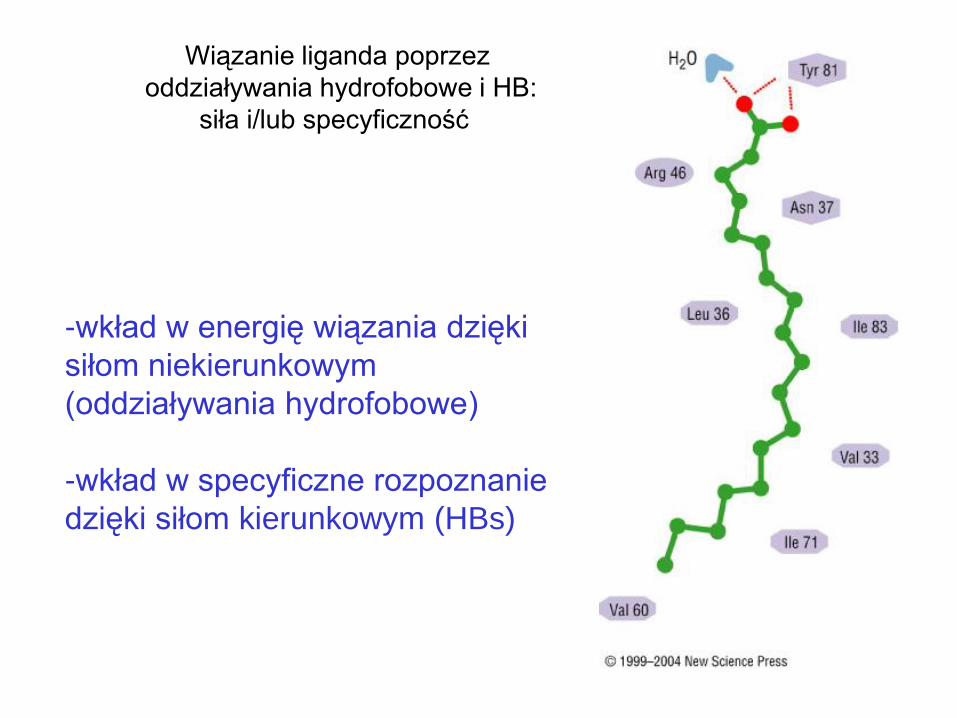
Wiązanie liganda poprzez
oddziaływania hydrofobowe i HB:
siła i/lub specyficzność
-wkład w energię wiązania dzięki
siłom niekierunkowym
(oddziaływania hydrofobowe)
-wkład w specyficzne rozpoznanie
dzięki siłom kierunkowym (HBs)

Białka strukturalne
- komórka jest tworem silnie ustrukturyzowanym a
zarazem dynamicznym
- nadanie i utrzymanie kształtu komórek i organelli,
a także jego zmiany w odpowiedzi na bodźce
zewnętrzne, zależą od białek strukturalnych
- kształt samego tylko rybosomu zależy od ponad
100 różnych białek
- wiele struktur tworzonych przez białka ma
charakter tymczasowy, np. skrzepy krwi, włókna
naprężeniowe

Oddziaływania w białkach
strukturalnych -rusztowania wyłącznie białkowe (α - kolagen,
β - jedwab, elastyna, keratyna, wirusy)
lub mieszane (chrząstka)
-często cross-linking łańcuchów białkowych
(kolagen, elastyna)
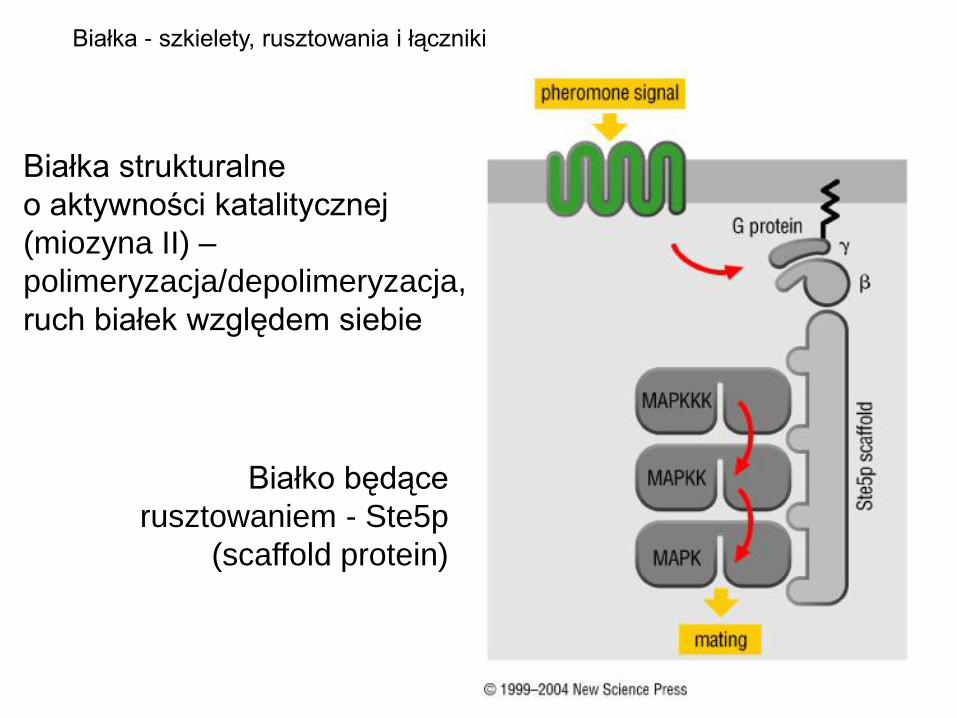
Białko będące
rusztowaniem - Ste5p
(scaffold protein)
Białka - szkielety, rusztowania i łączniki
Białka strukturalne
o aktywności katalitycznej
(miozyna II) –
polimeryzacja/depolimeryzacja,
ruch białek względem siebie


















