
Presentación de PowerPointiesicaria.xtec.cat/sbg/Enzimas.pdfdel enzima. Modelo Ajuste inducido...
46
Javier Cámara
Transcript of Presentación de PowerPointiesicaria.xtec.cat/sbg/Enzimas.pdfdel enzima. Modelo Ajuste inducido...

Javier Cámara

¿Qué son los enzimas?-asa
Los enzimas son biocatalizadores, es decir, sustancias que en una reacción química aumentan notablemente su velocidad.
Su función no es hacer reacciones imposibles, sino únicamente acelerar las reacciones que espontáneamente podrían producirse.
Las reacciones catalizadas por enzimas llegan a ser desde un millón hasta 100 millones de veces más rápidas que la misma reacción no catalizada.

Rendimiento de los enzimas
7.7 X 106
1.9 X 106
1.0 X 109
1.9X 1011
6.0 X 1012
5.6 X 1014
1.0 X 106
5043005786095
1.3 X 10 –12.6 X 10 –54.3 X 10 –63.0 X 10 –91.0 X 10 –11
1.7 X 10 -13
Anhidrasa carbónicaCorismato mutasaTriosafosfato isomerasaCarboxipeptidasa AAMP nucleosidasaNucleasa estafilococal
RendimientoVelocidad de
reacción catalizada
Velocidad en ausencia de
enzimaEnzima

Características de los enzimas
- Casi todos son proteínas. Los ribozimas, son los únicos enzimas no proteicos.
- Tienen elevado peso molecular.
- No se consumen durante la reacción.
- Muy activos. Típicamente, cada molécula de enzima es capaz de transformar cada segundo de 100 a 1000 moléculas de sustrato en producto. El número de estas moléculas transformadas a producto por molécula de enzima en cada segundo, se conoce como el número de recambio.
- Actúan a temperatura ambiente.
- Son muy específicos.

Acción enzimáticaLa sustancia sobre la que actúa el enzima se llamasustrato.
El sustrato se une al enzima, formando el complejo sustrato-enzima.
S-ES + E
Una vez formados los productos el enzima puede comenzar un nuevo ciclo de reacción.
P + E
Ciclo de un enzima: sacarasa

Formación del complejo sustrato-enzima
enzima
La unión se realiza en una región concreta del enzima, llamada centro activo. El centro activo comprende (1) un sitio de unión formado por los aminoácidos que están en contacto directo con el sustrato y (2) un sitio catalítico, formado por los aminoácidos directamente implicados en el mecanismo de la reacción.
complejo sustrato-enzima enzima
productos
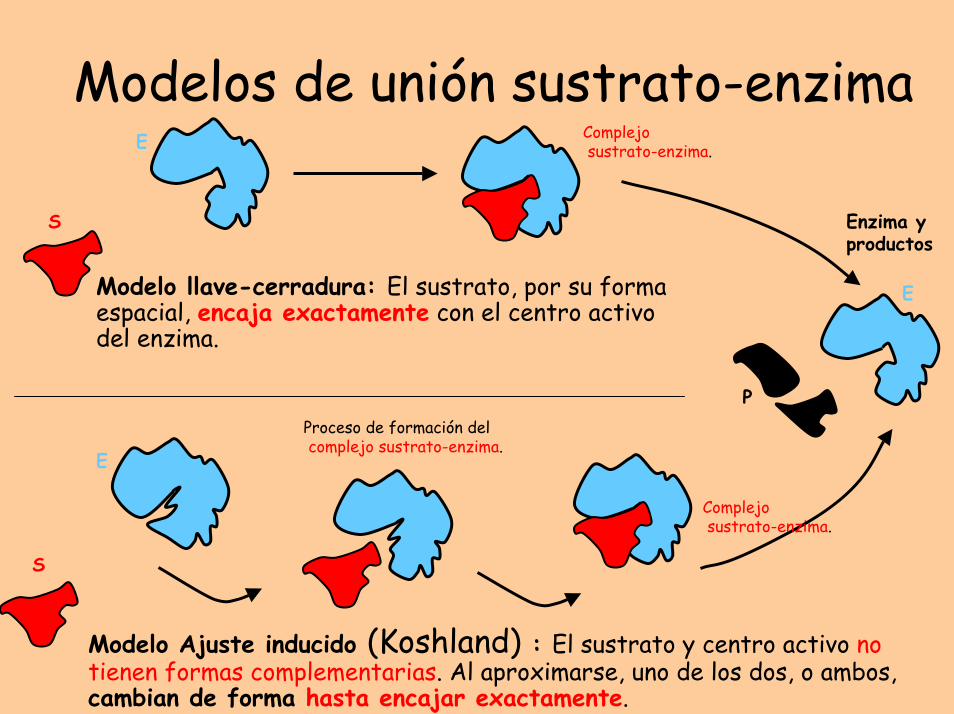
Modelos de unión sustrato-enzima
Modelo llave-cerradura: El sustrato, por su forma espacial, encaja exactamente con el centro activo del enzima.
Modelo Ajuste inducido (Koshland) : El sustrato y centro activo no tienen formas complementarias. Al aproximarse, uno de los dos, o ambos, cambian de forma hasta encajar exactamente.
Complejosustrato-enzima.
Proceso de formación delcomplejo sustrato-enzima.
Complejosustrato-enzima.
S
E
E
S Enzima y productos
E
P
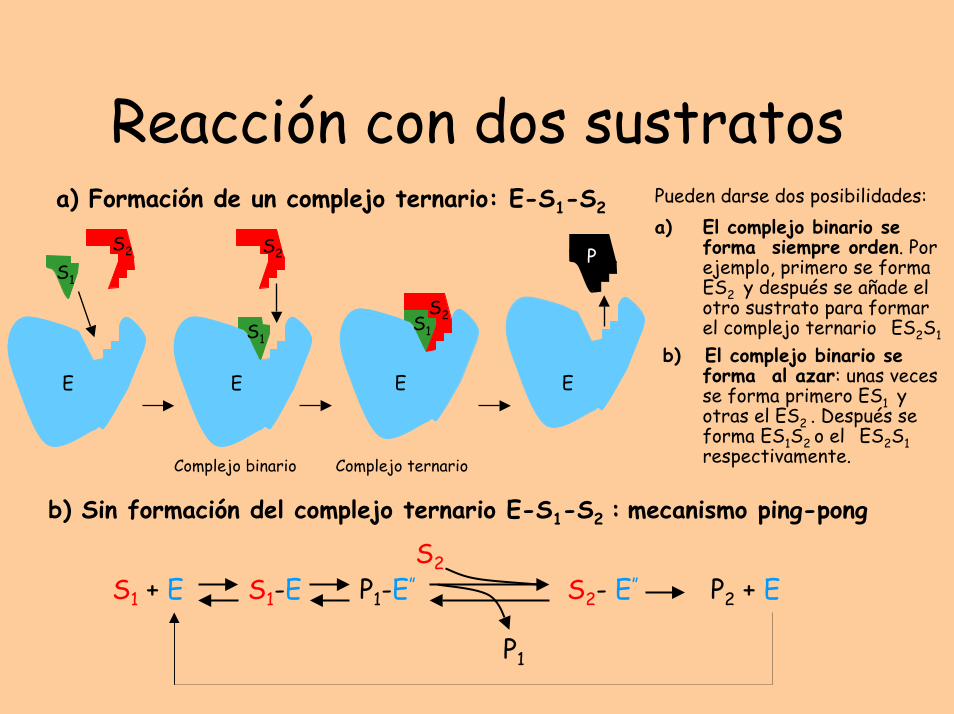
Reacción con dos sustratosa) Formación de un complejo ternario: E-S1-S2
Pueden darse dos posibilidades:a) El complejo binario se
forma siempre orden. Por ejemplo, primero se forma ES2 y después se añade el otro sustrato para formar el complejo ternario ES2S1
S1
S2
E
P
E
S1
S2
E
Complejo binario
S1
S2
E
Complejo ternario
b) El complejo binario se forma al azar: unas veces se forma primero ES1 y otras el ES2 . Después se forma ES1S2 o el ES2S1 respectivamente.
b) Sin formación del complejo ternario E-S1-S2 : mecanismo ping-pong
P1
S2S1 + E S1-E P1-E” P2 + ES2- E”

Nomenclatura1.- A los primeros enzimas que se descubrieron se les dio un nombre propio. Este nombre, los relacionaba, o no, con la procedencia anatómica donde fueron descubiertos: ptialina de la saliva, tripsina pancreática, rubisco...
2.- El primer intento de sistematizar la nomenclatura, fue utilizar un nombre con dos partes. La primera parte es el nombre del sustrato específico o general sobre el que actúan y la segunda parte es la terminación asa: ureasa, lipasa, proteasa...
3.- Al descubrirse que un mismo enzima puede realizar catalizar distintas reacciones, se amplió el nombre, añadiendo la función que realiza antes de la terminación asa, por ejemplo Isocítrico descarboxilasa. Al enzima rubisco, se le denomina ribulosa difosfato carboxilasa o bien ribulosa difosfato oxidasa, según la reacción que catalice.
4.- Un nuevo avance el nomenclatura se logra dando información complementaria, al indicar el coenzima que utiliza: malato NAD+ oxidoreductasa.
5.- La sistematización completa del nombre, se hace, nombrando a los enzimas con un numero clave (E.C.) que caracteriza al tipo de reacción según la clase (primer digito), subclase (segundo digito) y subsubclase (tercer digito). El cuarto digito es para lel enzima especifico. Asi, en la E.C. 2.7.1.1: el 2 indica que se trata de una transferasa, el 7 indica que transfiere fosfato, el primer 1que el alcohol es el aceptor del fosfato y el último 1 indica que es una hexoquinasa o ATPasa. También se le denomina D-hexosa-6-fosforotransferasa, enzima que cataliza la transferencia de fosfato desde el ATP al grupo hidroxilo de carbono 6 de la glucosa.

Tipos de enzimasSegún su composición se pueden clasificar en tres grupos:
1.- Ribozimas: compuestos por pequeñas moléculas de ARN, que tienen actividad catalítica.
2.- Enzimas simples o estrictamente proteicos: son aquellos que están compuestos exclusivamente por aminoácidos.
3.- Enzimas complejos u holoenzimas: son los compuestos por una parte proteica (Apoenzima) y un grupo prostético, parte no formada por aminoácidos, (Cofactor).
HOLOENZIMA = APOENZIMA + COFACTOR Si es inorgánica se le denomina activador inorgánico. Ej. Mg2+ es necesario para las quinasas. El Zn2+ para las carboxipeptidasas...Si es una molécula orgánica se les denomina COENZIMA
La biotina o vit b8 es coenzima de los enzimas que transfieren CO2
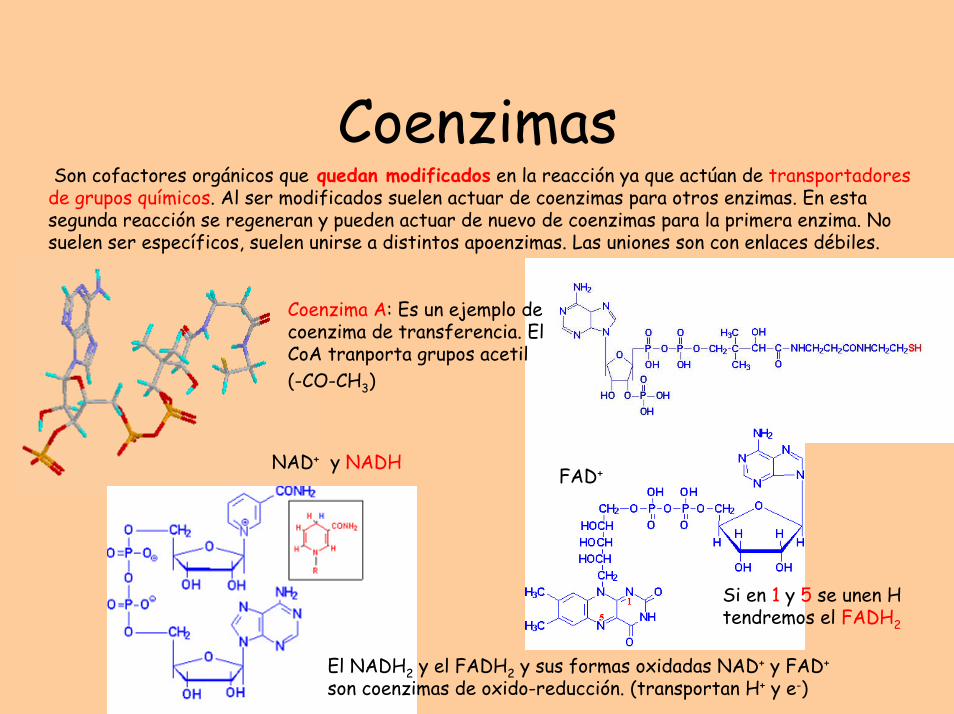
CoenzimasSon cofactores orgánicos que quedan modificados en la reacción ya que actúan de transportadores
de grupos químicos. Al ser modificados suelen actuar de coenzimas para otros enzimas. En esta segunda reacción se regeneran y pueden actuar de nuevo de coenzimas para la primera enzima. No suelen ser específicos, suelen unirse a distintos apoenzimas. Las uniones son con enlaces débiles.
Coenzima A: Es un ejemplo de coenzima de transferencia. El CoA tranporta grupos acetil (-CO-CH3)
NAD+ y NADH
El NADH2 y el FADH2 y sus formas oxidadas NAD+ y FAD+
son coenzimas de oxido-reducción. (transportan H+ y e-)
Si en 1 y 5 se unen H tendremos el FADH2
FAD+

Clasificación de los enzimas• Clase 1: Oxidorreductasas
• Clase 2: Transferasas
• Clase 3: Hidrolasas
• Clase 4: Liasas
• Clase 5: Isomerasas• Clase 6: Sintetasas

Clase 1: OxidorreductasasCatalizan reacciones de oxidorreducción, es decir, transferencia de hidrógeno (H) o electrones (e-) de un sustrato a otro. Según el sustrato sobre el que actúan o los coenzimas utilizados se diferencian 97 subclases diferentes. De ellas las dos más importantes son:
Oxidasas: transfieren sólo electrones
Deshidrogenasas: transfieren H utilizando coenzimas.
CH3 COOHCH
OH Ác. láctico
CH3 COOHC
O Ác. pirúvico
NAD+
NADH+H+oxidado
oxidado
reducidoe-e-
reducido
e- e-

Clase 2: TransferasasCatalizan la transferencia de un grupo químico, distinto del hidrógeno, de un sustrato a otro. Hay 9 subclases.
Pi PiPi
ATP ADP
GlucosaGlucosa 6 fosfatoGlucoquinasa
Pi Pi
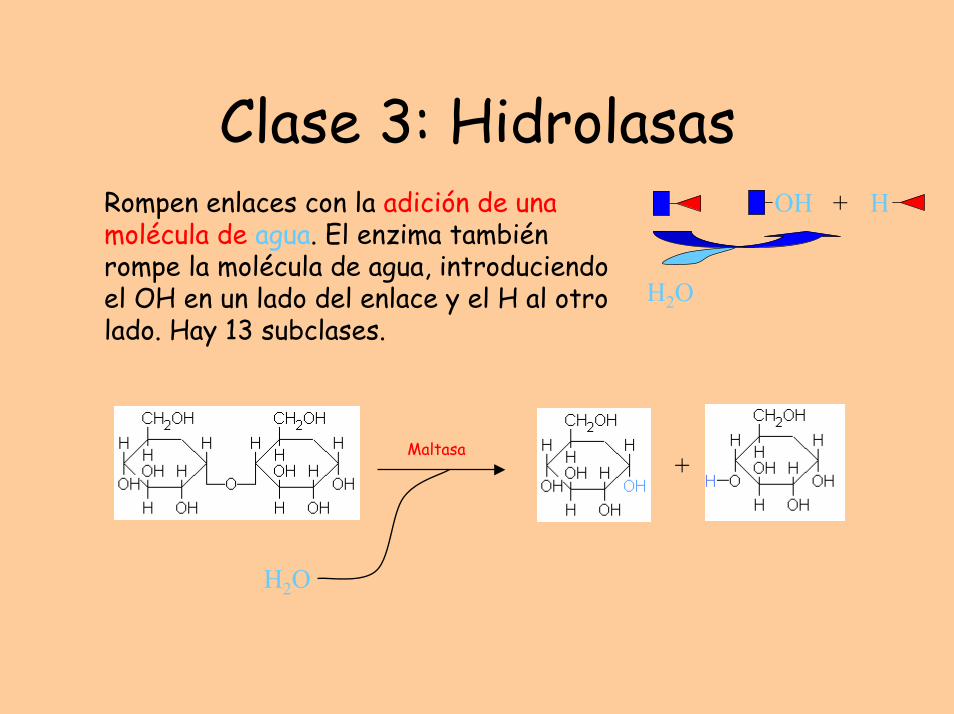
Clase 3: HidrolasasRompen enlaces con la adición de una molécula de agua. El enzima también rompe la molécula de agua, introduciendo el OH en un lado del enlace y el H al otro lado. Hay 13 subclases.
H2O
OH + H
+
H2O
Maltasa
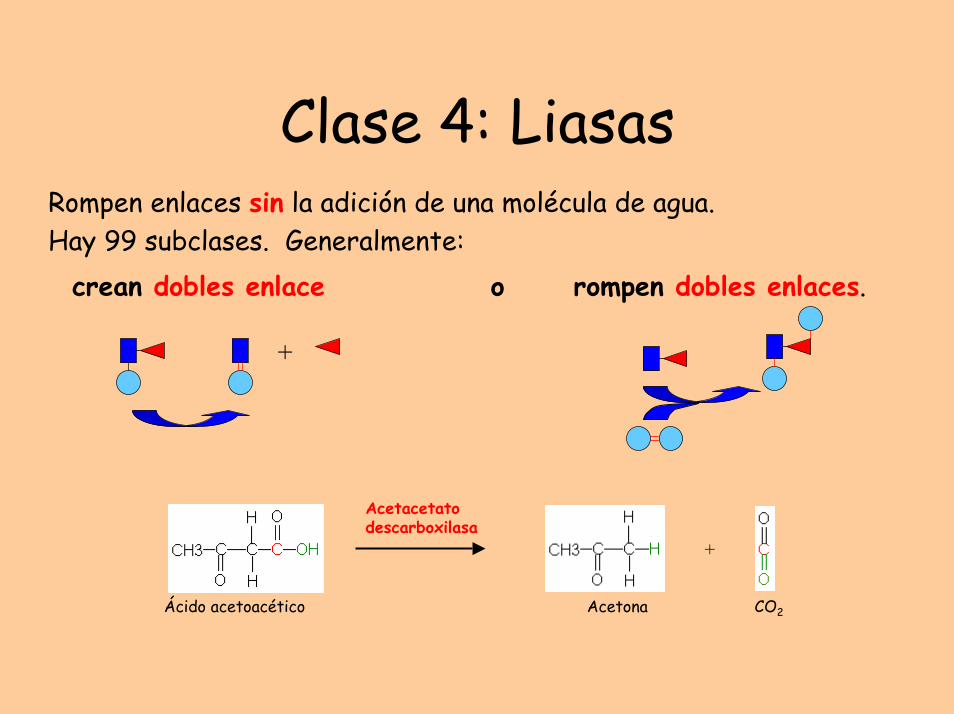
Clase 4: LiasasRompen enlaces sin la adición de una molécula de agua. Hay 99 subclases. Generalmente:
crean dobles enlace o rompen dobles enlaces.
+
Acetacetato descarboxilasa
+
Ácido acetoacético Acetona CO2
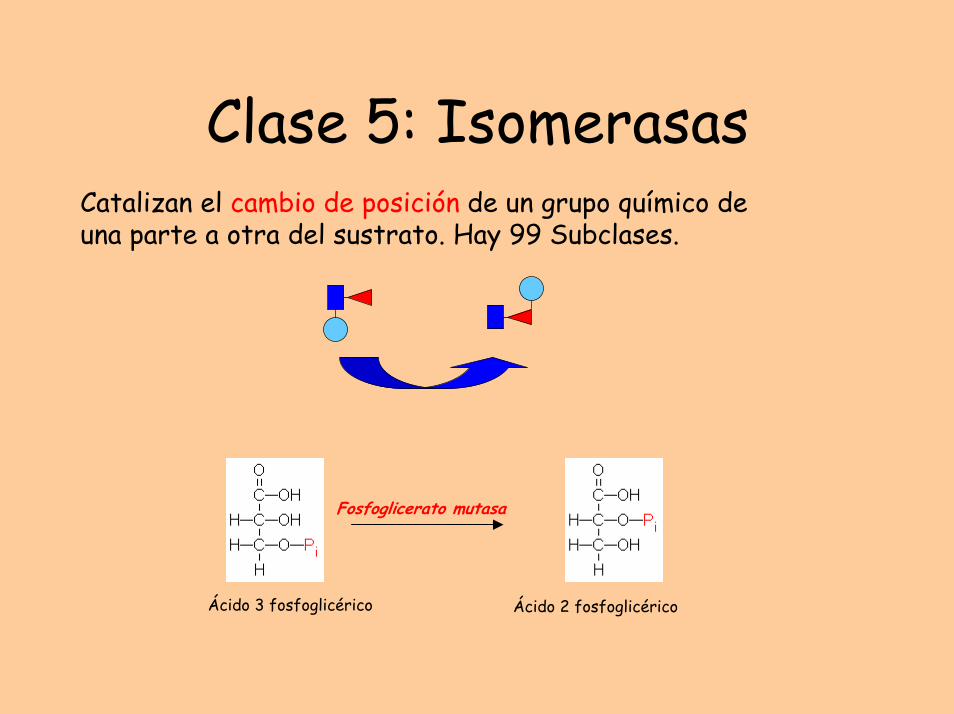
Clase 5: IsomerasasCatalizan el cambio de posición de un grupo químico de una parte a otra del sustrato. Hay 99 Subclases.
Ácido 3 fosfoglicérico Ácido 2 fosfoglicérico
Fosfoglicerato mutasa
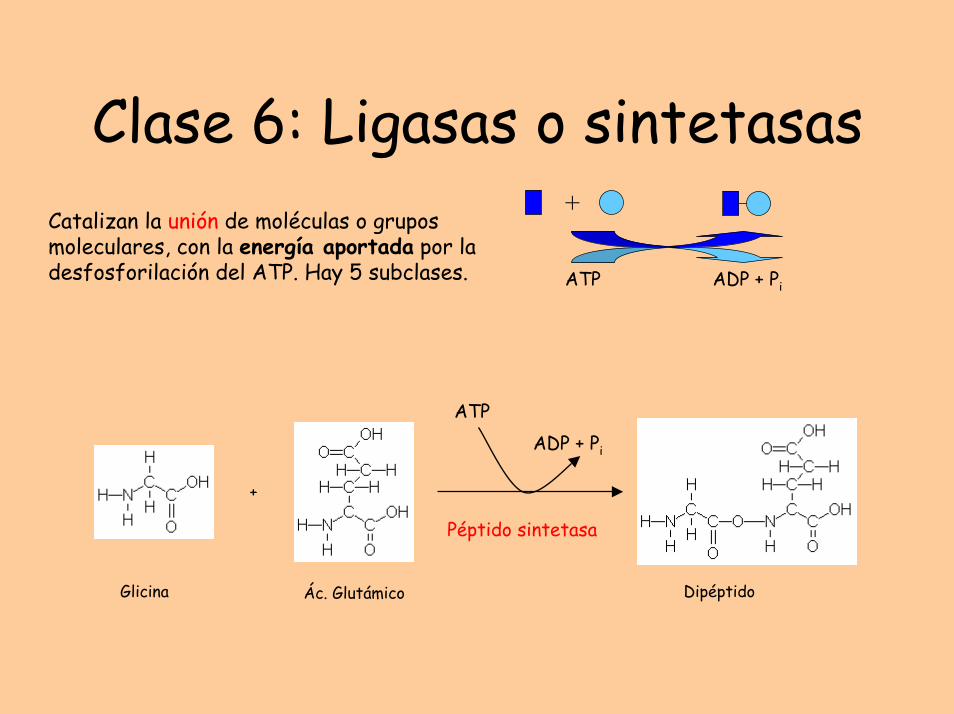
Clase 6: Ligasas o sintetasas+
ATP ADP + Pi
Catalizan la unión de moléculas o grupos moleculares, con la energía aportada por la desfosforilación del ATP. Hay 5 subclases.
Glicina Ác. Glutámico
+
Dipéptido
ATPADP + Pi
Péptido sintetasa
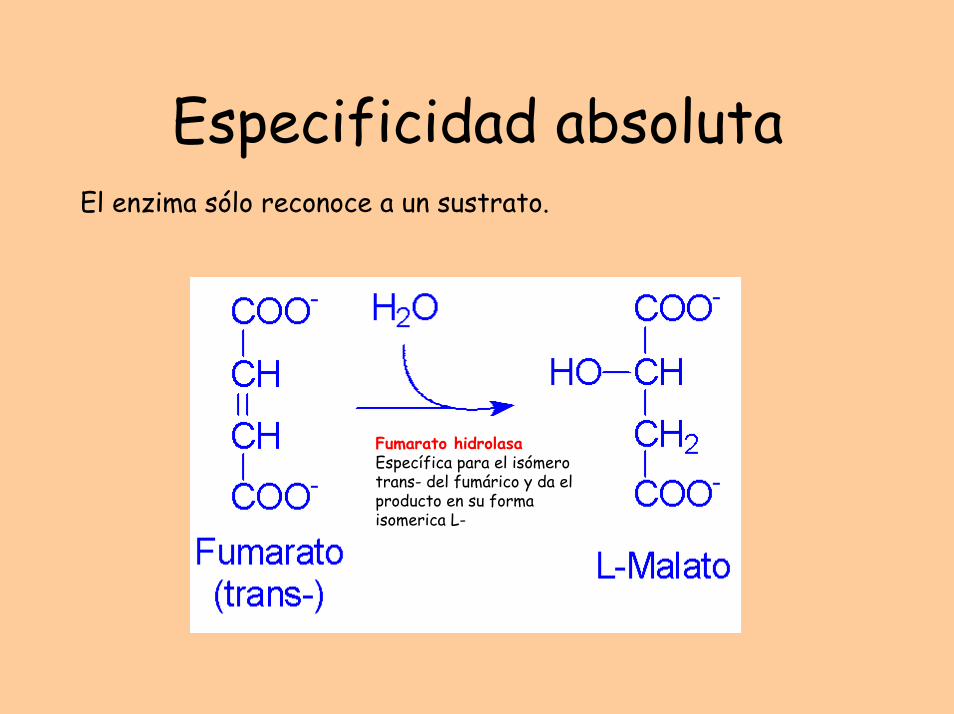
Especificidad absolutaEl enzima sólo reconoce a un sustrato.
Fumarato hidrolasaEspecífica para el isómero trans- del fumárico y da el producto en su forma isomerica L-

Especificidad de grupo
Actúan sobre el mismo grupo químico de moléculas.
Por ejemplo, la β-glucosidasa reconoce cualquier glúcido β perono a los α.
α fructosa β fructosa α glucosa β glucosa
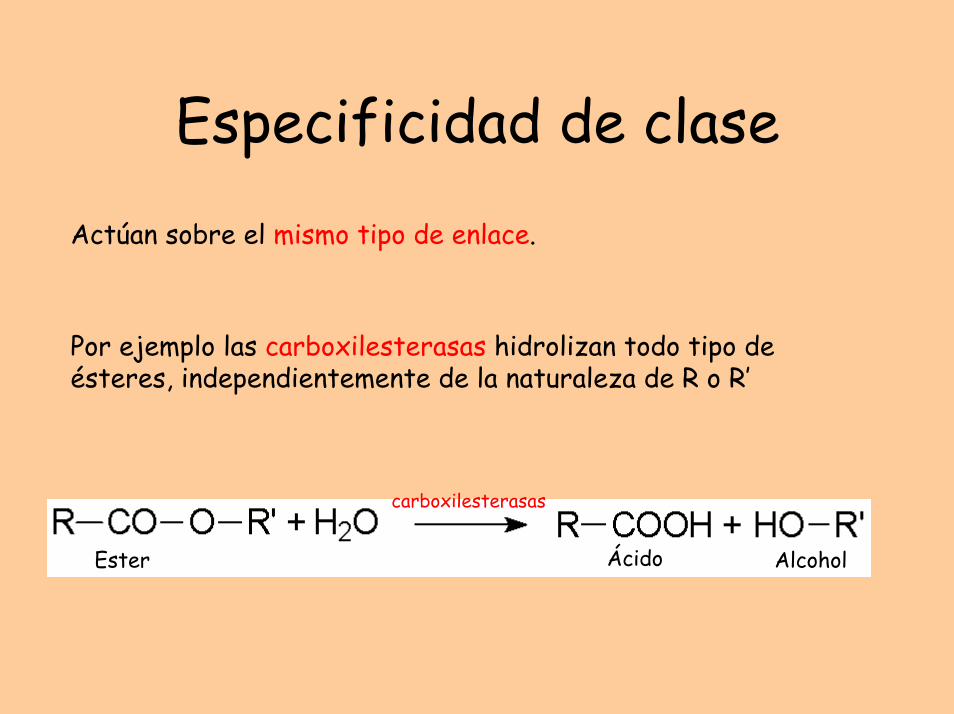
Especificidad de claseActúan sobre el mismo tipo de enlace.
Por ejemplo las carboxilesterasas hidrolizan todo tipo de ésteres, independientemente de la naturaleza de R o R’
carboxilesterasas
Ester Ácido Alcohol
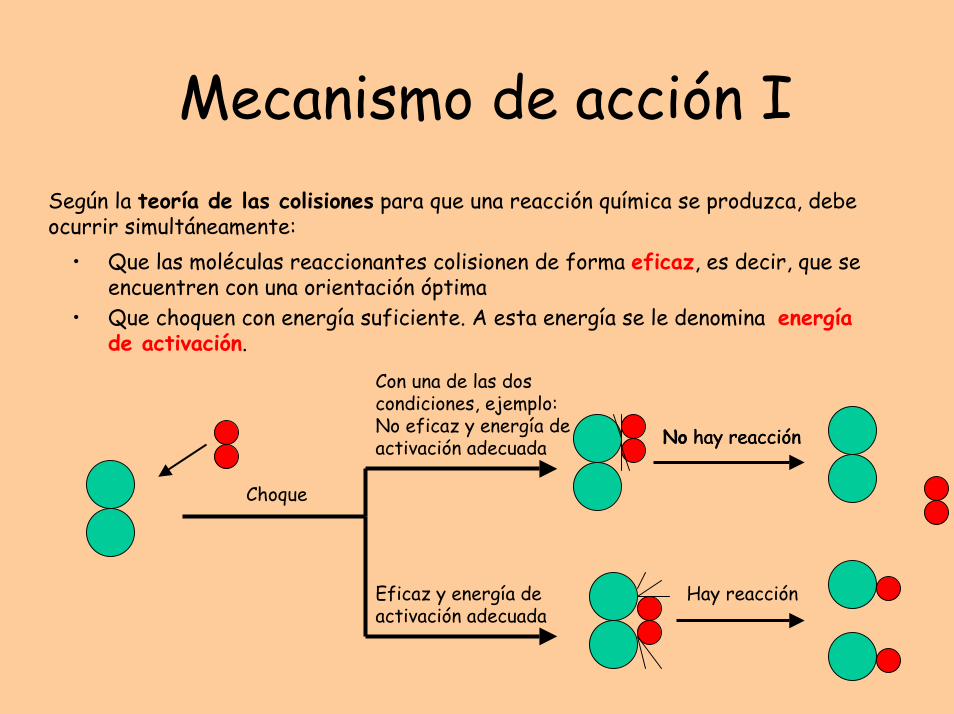
Mecanismo de acción ISegún la teoría de las colisiones para que una reacción química se produzca, debe ocurrir simultáneamente:
• Que las moléculas reaccionantes colisionen de forma eficaz, es decir, que se encuentren con una orientación óptima
• Que choquen con energía suficiente. A esta energía se le denomina energía de activación.
Choque
Eficaz y energía de activación adecuada
Con una de las dos condiciones, ejemplo: No eficaz y energía de activación adecuada No hay reacción
Hay reacción
No hay reacción
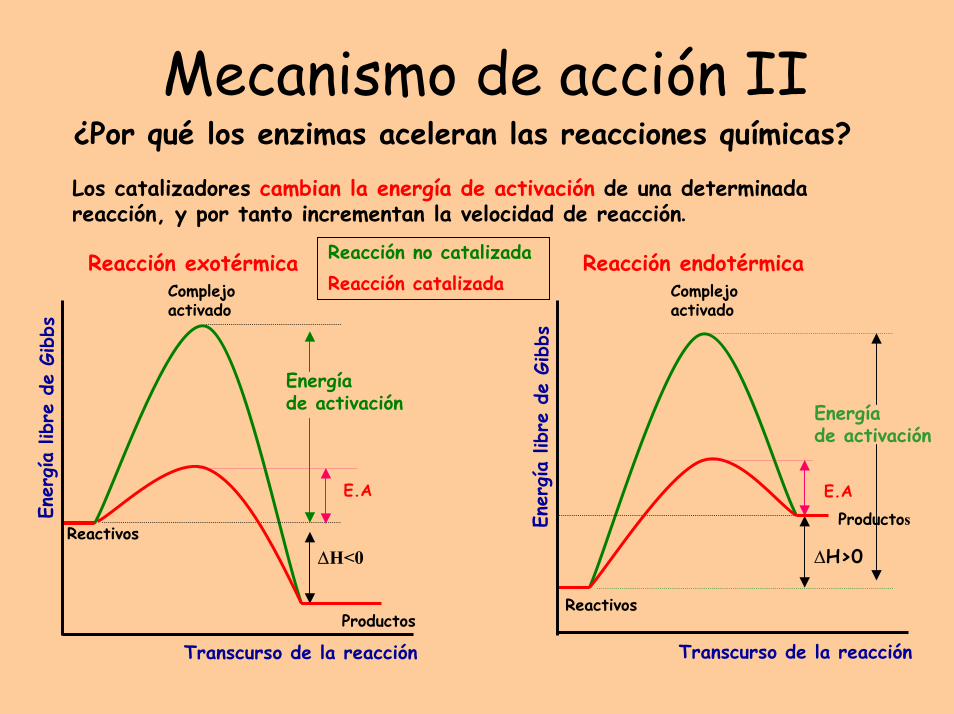
Mecanismo de acción II¿Por qué los enzimas aceleran las reacciones químicas?
Los catalizadores cambian la energía de activación de una determinada reacción, y por tanto incrementan la velocidad de reacción.
Reacción no catalizadaReacción catalizada
Transcurso de la reacción
Reacción exotérmica
Energía de activación
Reactivos∆H<0
Productos
E.A
Ener
gía
libre
de
Gibb
s
Complejoactivado
Reacción endotérmica
Transcurso de la reacción
Energía de activación
Complejoactivado
Reactivos
∆H>0
Productos
E.A
Ener
gía
libre
de
Gibb
s
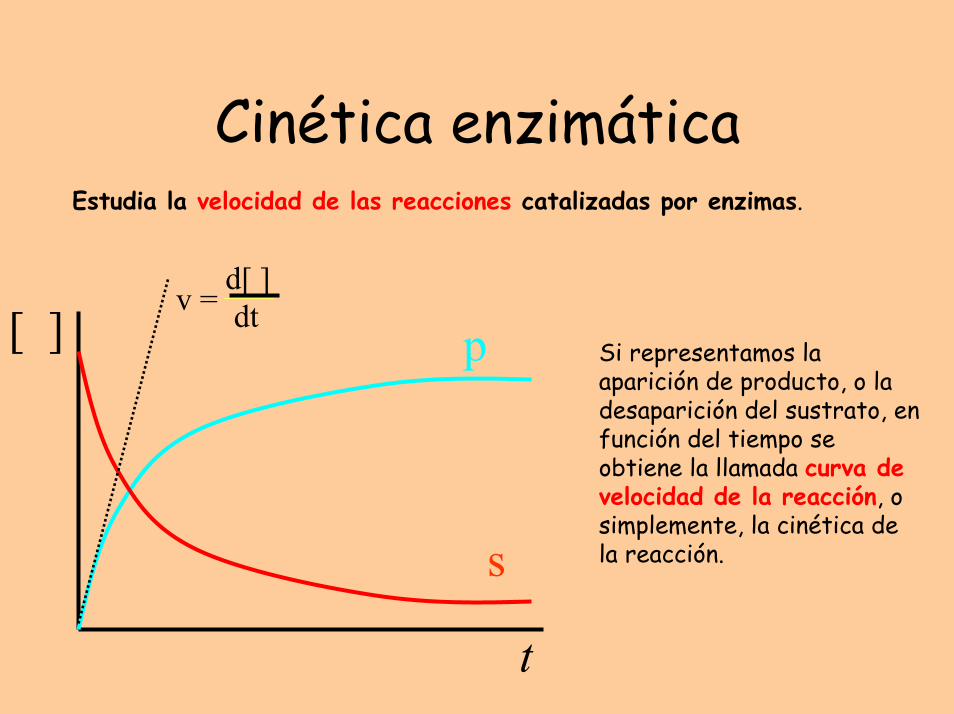
Cinética enzimáticaEstudia la velocidad de las reacciones catalizadas por enzimas.
t
s
p[ ]d[ ]dtv =
Si representamos la aparición de producto, o la desaparición del sustrato, en función del tiempo se obtiene la llamada curva de velocidad de la reacción, o simplemente, la cinética de la reacción.

Modelo de Michaelis-MentenEn 1913 Leonor Michaelis y Maude Menten postulatron la teoríageneral de la acción enzimática, basada en la formación delcomplejo SE.

1.-Deducciones de M-M Michaelis y Menten propusieron que las reacciones catalizadas enzimáticamente ocurren en dos etapas: En la primera etapa se forma el complejo enzima-sustrato y en la segunda, el complejo enzima-sustrato da lugar a la formación del producto, liberando el enzima libre
S + E SE E + PK1
K2
K3
Dado que concentración total de enzima, [ET] es constante a lo largo de la reacción y que presenta dos formas: el enzima libre (E) y el enzima unido al sustrato (ES), se puede decir que:
[ET] = [E] + [SE]
v1 = k1[S] [ET] - k1 [S] [SE]Sustituyendo en la primera ecuación,
O lo que es lo mismo [E] = [ET] - [SE]
En este esquema, k1, k2 y k3 son las constantes cinéticas individuales de cada proceso y también reciben el nombre de constantes microscópicas de velocidad. Según esto, podemos afirmar que:
v1 = k1 [S]·[E] v2 = k2 [SE] v3 = k3 [SE]
la transformamos en la siguiente:

2.-Deducciones de M-M S + E
K1
K2
K3SE E + PDe la reacción:
deducimos que la velocidad de formación del complejo enzima-sustrato (v1) es igual a la de su disociación (v2+ v3), por tanto: v1 = v2 + v3
k1[S] [ET] - k1 [S] [SE]= k2 [SE] + k3 [SE] O lo que lo mismo:
Despejando [SE], queda que:[Et]·[S]
(k2+k3)/k1 + [S][SE] =
Si todas las constantes
las agrupamos en una: Km = (k2 +k3) / k1 queda:
[Et]·[S]
Km + [S][SE] =
A la constante KM se le denomina constante de Michaelis-Menten.
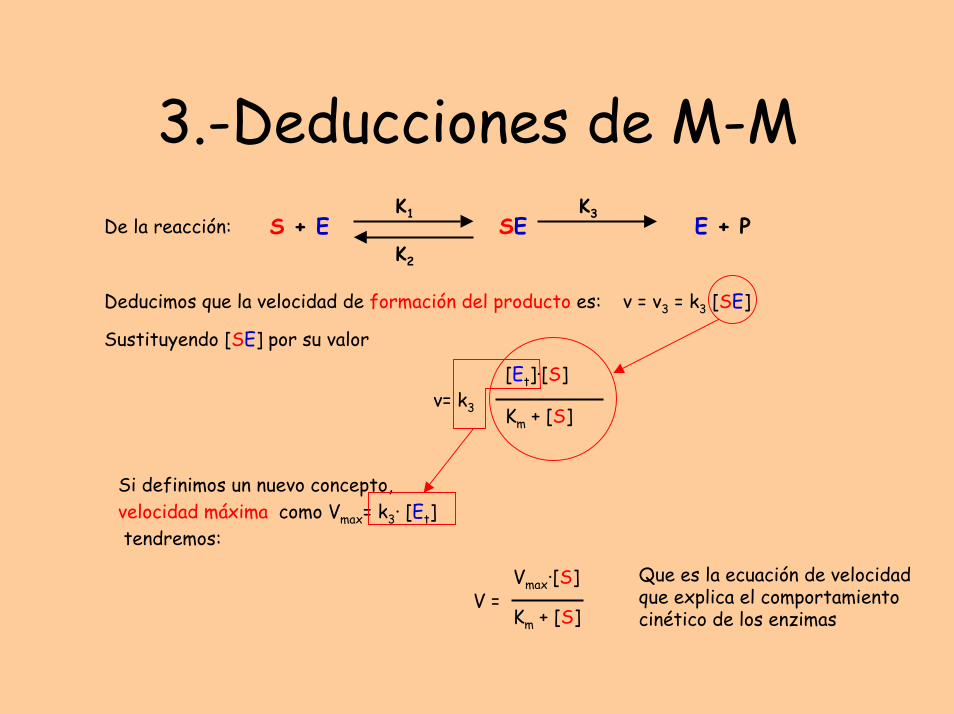
3.-Deducciones de M-MS + E
K1
K2
K3SE E + PDe la reacción:
Deducimos que la velocidad de formación del producto es:
Si definimos un nuevo concepto, velocidad máxima como Vmax= k3· [Et]tendremos:
v = v3 = k3 [SE]
[Et]·[S]
Km + [S]v= k3
Sustituyendo [SE] por su valor
Que es la ecuación de velocidad que explica el comportamiento cinético de los enzimas
V =Vmax·[S]
Km + [S]
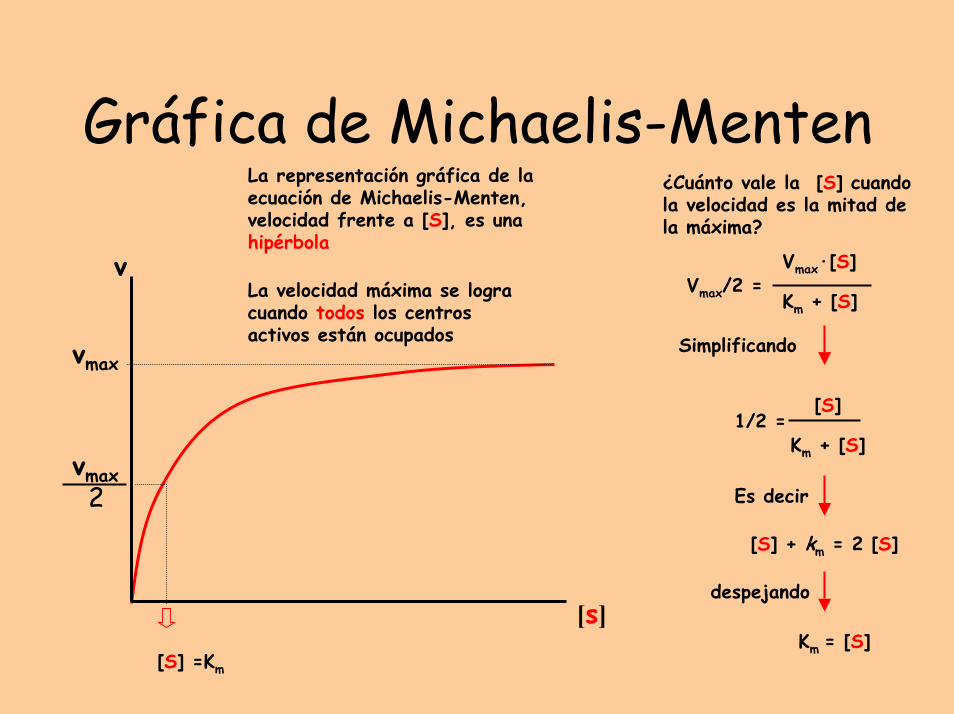
Gráfica de Michaelis-Menten
[s]
v
La representación gráfica de la ecuación de Michaelis-Menten, velocidad frente a [S], es una hipérbola
vmax2
?[S] =Km
vmax
La velocidad máxima se logra cuando todos los centros activos están ocupados
¿Cuánto vale la [S] cuando la velocidad es la mitad de la máxima?
Vmax/2 =Vmax·[S]
Km + [S]
Simplificando
[S]1/2 =
Km + [S]
Es decir
[S] + km = 2 [S]
despejando
Km = [S]

Significado de KmKm es la concentración de sustrato para la cual la velocidad de reacción es la mitad de la velocidad máxima.
El valor de Km da idea de la afinidad del enzima por el sustrato.A menor Km, mayor afinidad del enzima por el sustrato. Este hecho tiene fácil explicación si tenemos en cuenta que Km se define como (k2+k3/k1), donde las reacciones 2 y 3 destruyen el complejo ES, mientras que la reacción 1 lo forma. Así, si Km es grande, el complejo ES es inestable pues predomina la tendencia a destruirlo (poca afinidad hacia el sustrato), y si Km es pequeña, el complejo ES es estable, ya que predomina la tendencia a formarlo (gran afinidad hacia el sustrato).
La Km del sustrato natural es menor que la de los sustratos análogos. Si dos sustratos del mismo enzima tienen distinta Km, el que presente mayor Kmtiene menor afinidad por el enzima, y la reacción transcurre siempre a menor velocidad que con el sustrato de menor Km, salvo a concentraciones saturantes de sustrato, donde la V = Vmax.
Los valores de Km de muchos enzimas son próximos a los de la concentración fisiológica de sus sustratos, de forma que pequeñas variaciones en la [S] pueden suponer grandes cambios en la velocidad de toda una ruta metabólica.

Cálculo de Km y de Vmax
• Para determinar gráficamente los valores de KM y Vmax es más sencillo utilizar la representación doble recíproca (1/v0 frente a 1/[S]0), ya que es una línea recta.
De esta forma, a partir de los datos experimentales se puede calcular gráficamente, los valores de KM y Vmax de un enzima para diversos sustratos.
Km=1/2Si -2=-1/Km
Si 3=1/vmax Vmax=1/3
Es una recta en la cual:•La pendiente es KM/Vmax
•La abscisa en el origen (1/v0 = 0) es -1/KM
•La ordenada en el origen (1/[S]0 = 0) es 1/Vmax
-1/Km
1/Vmax
1/V
1/[S]
Pendiente= KM / Vmax
Esta representación doble recíproca recibe el nombre de representación de Lineweaver-Burk.

Unidades de la velocidad de reacción
Se define la unidad de actividad enzimática (U) como la cantidad de enzima que cataliza la conversión de 1 µmol de sustrato en un minuto. (1 µmol/min).
El Sistema Internacional de unidades (SI) define la unidad de actividad enzimática como la cantidad de enzima que transforma 1 mol de sustrato por segundo. Esta unidad se llama katal (kat).
Como 1 mol son 106 µmoles y 1 minuto son 60 segundos, resulta que1 katal = 60 x 106 U.
Esta unidad es muy grande, de forma que se utilizan frecuentemente los submúltiplos como el microkatal (µkat = 10-6 kat) o el nanokatal (nkat = 10-9 kat).

Regulación de las reacciones enzimáticas
La actividad de los enzimas puede regularse por:
1.- Expresión génica.
2.- Efecto del pH.
3.- Efecto de la temperatura.
4.- Presencia de inhibidores.
5.- Modulación alostérica.
6.- Modulación por proteolisis.
7.- Modulación isoenzimas.
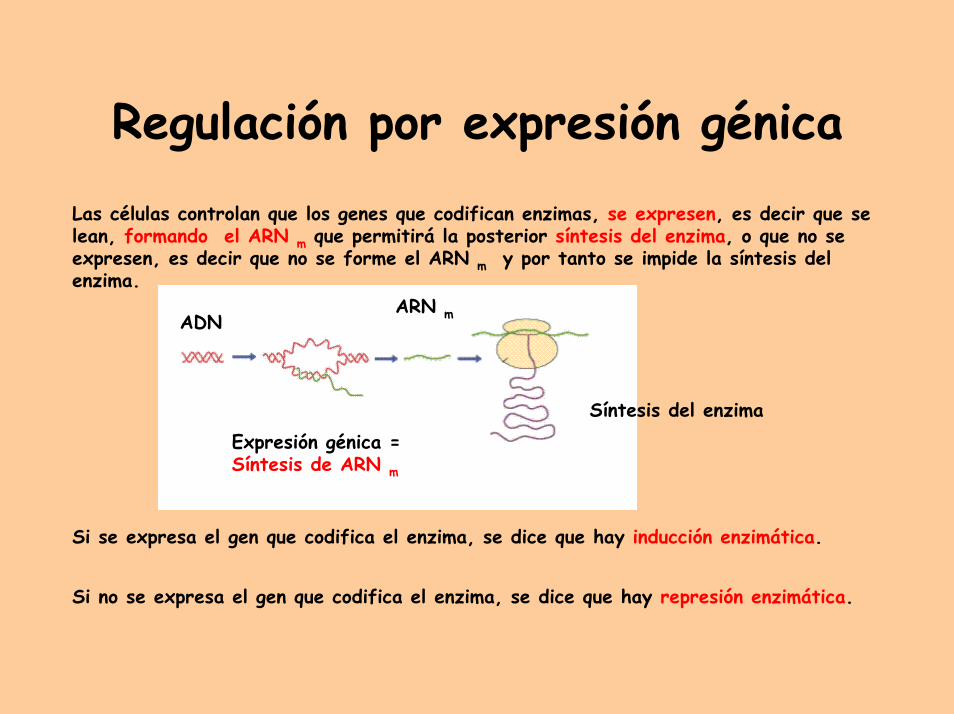
Regulación por expresión génica
ADN
Expresión génica =Síntesis de ARN m
ARN m
Síntesis del enzima
Las células controlan que los genes que codifican enzimas, se expresen, es decir que se lean, formando el ARN m que permitirá la posterior síntesis del enzima, o que no se expresen, es decir que no se forme el ARN m y por tanto se impide la síntesis del enzima.
Si se expresa el gen que codifica el enzima, se dice que hay inducción enzimática.
Si no se expresa el gen que codifica el enzima, se dice que hay represión enzimática.

Regulación por efecto del pHLos enzimas, en las cadenas laterales de sus aminoácidos, poseen grupos químicosionizables (carboxilos -COOH; amino -NH2; tiol -SH; imidazol, etc.). Según el pH del medio estos grupos se ionizan de forma distinta, pueden tener carga eléctrica positiva, negativa o neutra. Como la forma espacial de las proteínas depende, en parte, de las cargas eléctricas de sus restos, habrá un pH en el cual la conformación será la más adecuada para la actividad catalítica. Este es el llamado pH óptimo.
100%
pH1 2 3 4 5 6 7 8 9 10 11 12
V
50%
Si cambia el pH del medio, cambia la forma espacial del enzima y esto cambiala velocidad de la reacción.
9,7Arginasa
7,8Fumarasa
7,7Tripsina
7,6Catalasa
6,8Ureasa
1,5Pepsina
pHóptimoEnzima
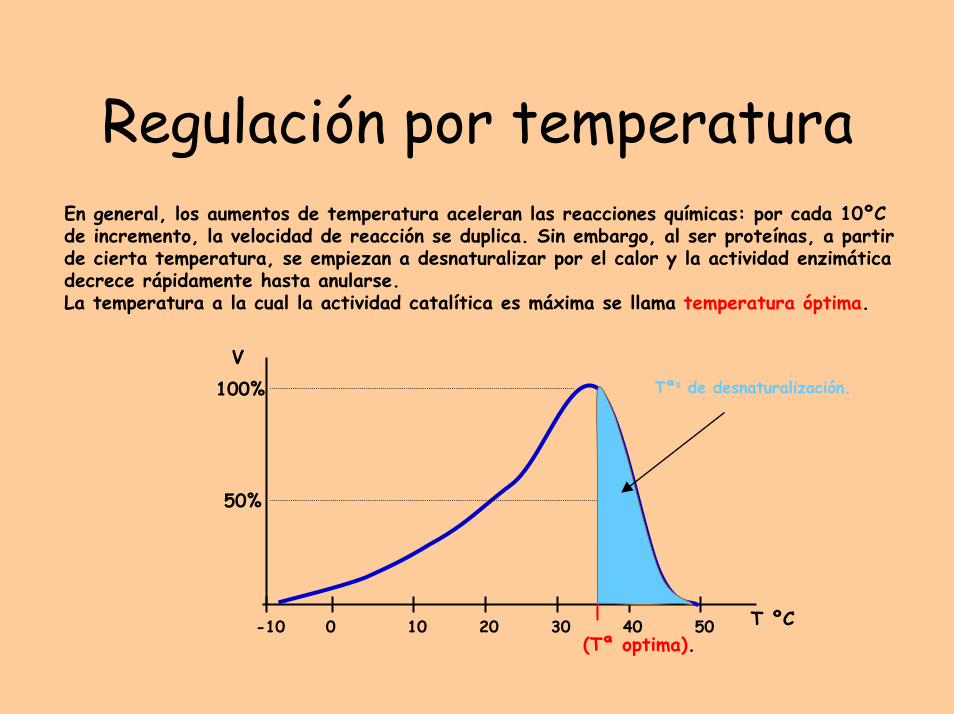
Regulación por temperaturaEn general, los aumentos de temperatura aceleran las reacciones químicas: por cada 10ºC de incremento, la velocidad de reacción se duplica. Sin embargo, al ser proteínas, a partir de cierta temperatura, se empiezan a desnaturalizar por el calor y la actividad enzimáticadecrece rápidamente hasta anularse.La temperatura a la cual la actividad catalítica es máxima se llama temperatura óptima.
100%
T ºC-10 0 10 20 30 40 50
V
50%
(Tª optima).
Tªs de desnaturalización.

Regulación por inhibidoresLos inhibidores son substancias que se unen al enzima disminuyendo o anulando su actividad. Hay dos tipos:
1.- Inhibición irreversible: El inhibidor se fija permanentemente, por medio de enlaces covalentes, al lugar activo de la enzima, o lo desnaturaliza impidiendo su actividad. Por ejemplo, los llamadosgases nerviosos actúan sobre los enzimas que participan en la transmisión del impulso nervioso, de forma que este proceso no puede tener lugar, produciéndose parálisis o muerte.
2.- Inhibición reversible: El inhibidor se fija temporalmente, por medio de enlaces débiles, al enzima dificultando su acción. Pueden ser de tres tipos: Inhibidores competitivos, no competitivos y acompetitivos
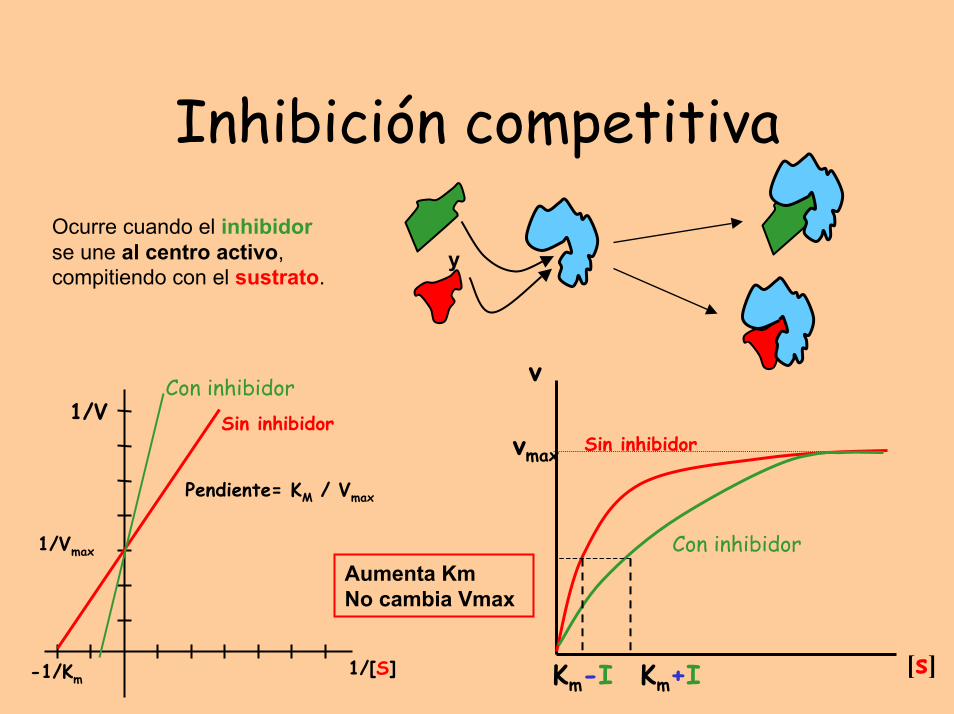
Inhibición competitivaOcurre cuando el inhibidorse une al centro activo, compitiendo con el sustrato.
-1/Km
1/Vmax
1/V
1/[S]
Pendiente= KM / Vmax
Con inhibidorSin inhibidor
[s]
v
vmax Sin inhibidor
Con inhibidor
Km+IKm-I
y
Aumenta KmNo cambia Vmax
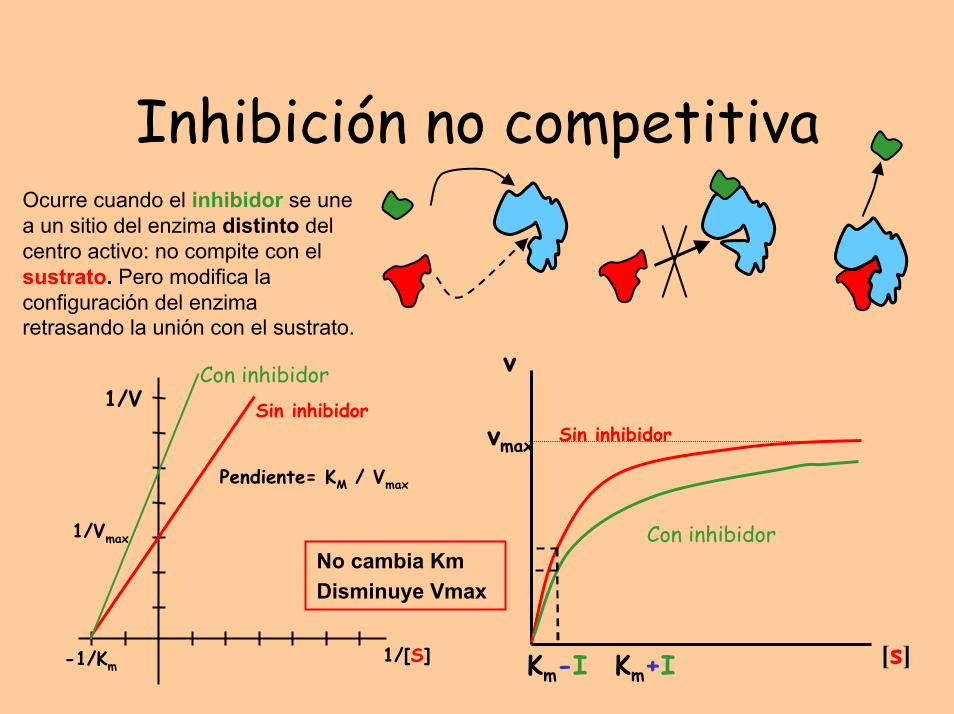
Inhibición no competitivaOcurre cuando el inhibidor se une a un sitio del enzima distinto del centro activo: no compite con el sustrato. Pero modifica la configuración del enzima retrasando la unión con el sustrato.
-1/Km
1/Vmax
1/V
1/[S]
Pendiente= KM / Vmax
Con inhibidorSin inhibidor
vmax
No cambia KmDisminuye Vmax
[s]
v
Sin inhibidor
Con inhibidor
Km+IKm-I

Inhibición acompetitivaOcurre cuando el inhibidor se une al complejo sustrato-enzima, retrasando la liberación del producto.
Complejo sustrato-enzima
-1/Km
1/Vmax
1/V
1/[S]
Pendiente= KM / Vmax
Con inhibidorSin inhibidor
Disminuye KmDisminuye Vmax
[s]
v
vmax Sin inhibidor
Con inhibidor
Km-IKm+I

Modulación alostérica IEl término alostérico significa “otro sitio” y también “otra forma”. Los enzimas alostéricos son aquellos, que presentan dos sitios y dos formas.
Los sitios son lugares donde se pueden unir sustancias. Uno es el centro activo, al que puede unirse el sustrato y otra el centro regulador, al que puede unirse, de forma reversible, otra sustancia llamada ligando.
Las formas son dos conformaciones espaciales, que están en equilibrio químico. Una es la tensa, en la que sustrato no se puede unir, forma inactiva y la otra es la relajada, en la que el sustrato se puede unir.
Centro activo con afinidad al sustrato
Centro regulador
Forma relajadaForma tensa
Centro regulador
Centro activo sin afinidad al sustrato
Si el ligando y el sustrato son substancias distintas, al enzima alostérico se le llamaherotrópico, en caso contrario, es decir, si son la misma sustancia, se denominan homotrópico.
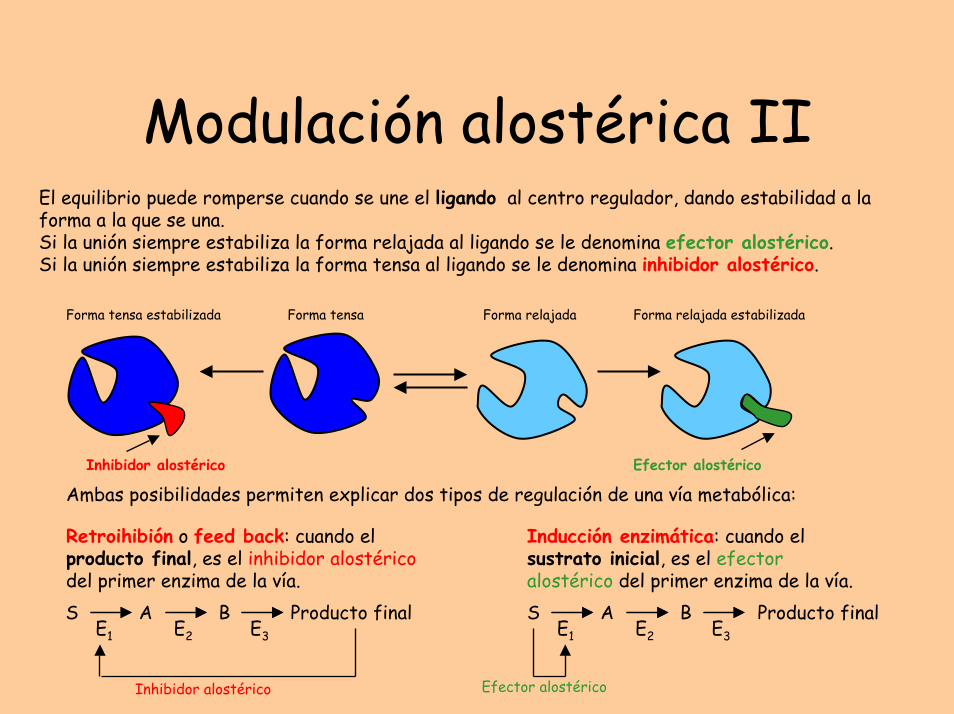
Modulación alostérica IIEl equilibrio puede romperse cuando se une el ligando al centro regulador, dando estabilidad a la forma a la que se una.Si la unión siempre estabiliza la forma relajada al ligando se le denomina efector alostérico. Si la unión siempre estabiliza la forma tensa al ligando se le denomina inhibidor alostérico.
Forma relajadaForma tensaForma tensa estabilizada Forma relajada estabilizada
Efector alostéricoInhibidor alostérico
Ambas posibilidades permiten explicar dos tipos de regulación de una vía metabólica:
S A B Producto final
Inhibidor alostérico
E1 E2 E3
Retroihibión o feed back: cuando el producto final, es el inhibidor alostéricodel primer enzima de la vía.
Efector alostérico
S A B Producto final E1 E2 E3
Inducción enzimática: cuando el sustrato inicial, es el efector alostérico del primer enzima de la vía.
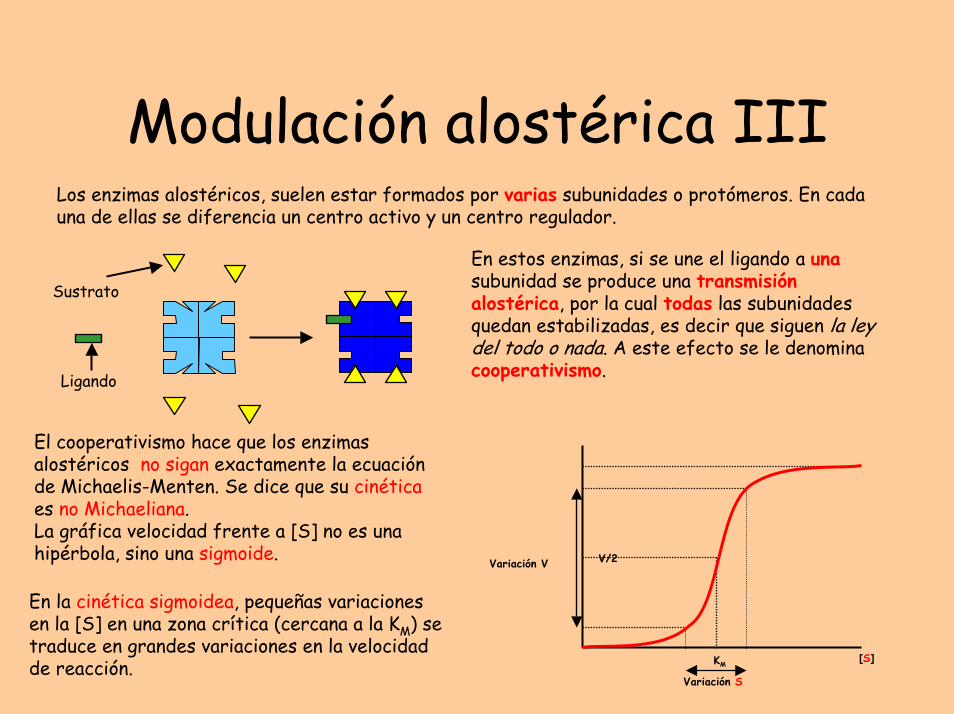
Modulación alostérica IIILos enzimas alostéricos, suelen estar formados por varias subunidades o protómeros. En cada una de ellas se diferencia un centro activo y un centro regulador.
En estos enzimas, si se une el ligando a unasubunidad se produce una transmisión alostérica, por la cual todas las subunidadesquedan estabilizadas, es decir que siguen la ley del todo o nada. A este efecto se le denomina cooperativismo.
Sustrato
Ligando
El cooperativismo hace que los enzimas alostéricos no sigan exactamente la ecuación de Michaelis-Menten. Se dice que su cinética es no Michaeliana.La gráfica velocidad frente a [S] no es una hipérbola, sino una sigmoide.
KM
V/2
Variación S
Variación V
[S]
En la cinética sigmoidea, pequeñas variaciones en la [S] en una zona crítica (cercana a la KM) se traduce en grandes variaciones en la velocidad de reacción.
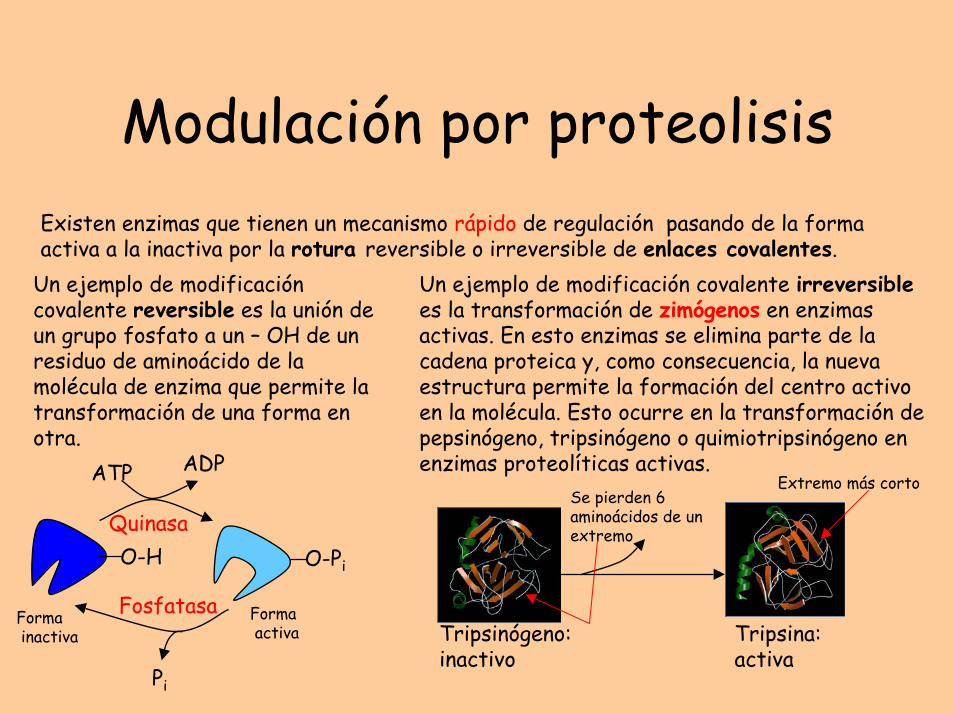
Modulación por proteolisisExisten enzimas que tienen un mecanismo rápido de regulación pasando de la forma activa a la inactiva por la rotura reversible o irreversible de enlaces covalentes.
Un ejemplo de modificación covalente irreversiblees la transformación de zimógenos en enzimas activas. En esto enzimas se elimina parte de la cadena proteica y, como consecuencia, la nueva estructura permite la formación del centro activo en la molécula. Esto ocurre en la transformación de pepsinógeno, tripsinógeno o quimiotripsinógeno en enzimas proteolíticas activas.
Tripsinógeno:inactivo
Se pierden 6 aminoácidos de un extremo
Tripsina:activa
Extremo más corto
Un ejemplo de modificación covalente reversible es la unión de un grupo fosfato a un – OH de un residuo de aminoácido de la molécula de enzima que permite la transformación de una forma en otra.
O-H
ATP ADP
Pi
O-Pi
Quinasa
Fosfatasa Formaactiva
Formainactiva

Modulación isoenzimasSe llaman isozimas o isoenzimas a los enzimas que catalizan la misma reacciónpero que pueden distinguirse por alguna característica física, estructural, inmunológica, etc. Su origen es genético.
Las diferencias que presentan, se traducen en ligeros cambios en sus propiedades, de forma que cada isozima se adapta perfectamente a la función que debe realizar.
Se diferencian isoenzimas en función de:
- el tipo de tejido: Por ejemplo, la Hexokinasa, presenta, al menos, cuatro formas distintas:
- Hepática- Cerebral- Muscular- Eritrocitaria
- el compartimento celular donde actúa: Por ejemplo, la malato deshidrogenasadel citoplasma es distinta de la de la mitocondria.
- el momento concreto del desarrollo del individuo: Por ejemplo, algunos enzimas de la glicolisis del feto son diferentes de los mismos enzimas en el adulto.



















