Prenatal Maternal Speech Influences Newborns’ Perception ...€¦ · Prenatal Maternal Speech...
18
INFANT BEHAVIOR AND DEVELOPMENT 9, 1X3-150 (1986) PrenatalMaternal Speech Influences Newborns’ Perception of Speech Sounds*. ANTHONY J. DECASPER AND MELANIE J. SPENCE University of North Carolina at Greensboro Pregnant women recited a particular speech passage aloud each day during their last 6 weeks of pregnancy. Their newborns were tested with an operant-choice procedure to determine whether the sounds of the recited passage were more reinforcing than the sounds of a novel passage. The previously recited passage was more reinforcing. The reinforcing value of the two passages did not differ for a matched group of control subjects. Thus, third-trimester fetuses experience their mothers’ speech sounds and that prenatal auditory experience can in- fluence postnatal auditory preferences. prenatal sensory experience auditory perception newborn perception fetal experience prenatal learning speech perception moternol voice Human newborns do not act like passive and neutral listeners. They prefer their own mothers’ voices to those of other females, female voices to male voices, and intrauterine heartbeat sounds to male voices, but they do not prefer their fathers’ voices to those of other males (Brazelton, 1978; DeCasper & Fifer, 1980; DeCasper & Prescott, 1984; Fifer, 1980; Panneton & DeCasper, 1984; Wolff, 1963). Why should newborns prefer some sounds over others? One hypothesis is that their auditory preferences are influenced by prenatal ex- perience with their mothers’ speech and heartbeats (DeCasper & Prescott, 1984). Several considerations suggest this hypothesis is plausible. Third-trimester fetuses hear, or are behaviorally responsive to, sound (e.g., Bernard & Sontag, 1947; Birnholz & Benacerraf, 1983; Grimwade, Walker, Bartlett, Gordon, & Wood, 1971; Johansson, Wedenberg, & Westin, 1964; Sontag & Wallace, 1935). Intrauterine recordings taken near term indi- cate that maternal speech and heartbeats are audible in utero (Querleu & Renard, 1981; Querleu, Renard, & Crepin, 1981; Walker, Grimwade, &Wood, 1971). Nonmaternal speech, for example male speech, is less audible because of attenuation by maternal tissue and/or masking by intrauterine sounds (Querleu & Renard, 1981; Querleu et al., 1981). * This research was supported by a Research Council Grant from the University of North Carolina at Greensboro and a generous equipment loan by Professor Michael D. Zeiler. We wish to thank the medical and administrative staff of Moses H. Cone Hospital, Greensboro, NC and, especially, the mothers and their infants for making this research possible. Thanks also to G. Gottlieb, R. Harter. R. Hunt, R. Panneton. K. Smith, and, especially, W. Salinger for their helpful comments on drafts of the manuscript. Portions of this paper were presented at the Third Biennial Interna- tional Conference on Infant Studies, March 1982. Austin, TX. Correspondence and requests for reprints should be addressed to Anthony J. DeCasper, Department of Psychology, University of North Carolina at Greensboro, Greensboro, NC 27412. 133
Transcript of Prenatal Maternal Speech Influences Newborns’ Perception ...€¦ · Prenatal Maternal Speech...

INFANT BEHAVIOR AND DEVELOPMENT 9, 1X3-150 (1986)
Prenatal Maternal Speech Influences Newborns’ Perception of Speech Sounds*.
ANTHONY J. DECASPER AND MELANIE J. SPENCE University of North Carolina at Greensboro
Pregnant women recited a particular speech passage aloud each day during their last 6 weeks of pregnancy. Their newborns were tested with an operant-choice procedure to determine whether the sounds of the recited passage were more reinforcing than the sounds of a novel passage. The previously recited passage was more reinforcing. The reinforcing value of the two passages did not differ for a matched group of control subjects. Thus, third-trimester fetuses experience their mothers’ speech sounds and that prenatal auditory experience can in- fluence postnatal auditory preferences.
prenatal sensory experience auditory perception newborn perception fetal experience prenatal learning speech perception moternol voice
Human newborns do not act like passive and neutral listeners. They prefer their own mothers’ voices to those of other females, female voices to male voices, and intrauterine heartbeat sounds to male voices, but they do not prefer their fathers’ voices to those of other males (Brazelton, 1978; DeCasper & Fifer, 1980; DeCasper & Prescott, 1984; Fifer, 1980; Panneton & DeCasper, 1984; Wolff, 1963). Why should newborns prefer some sounds over others? One hypothesis is that their auditory preferences are influenced by prenatal ex- perience with their mothers’ speech and heartbeats (DeCasper & Prescott, 1984). Several considerations suggest this hypothesis is plausible.
Third-trimester fetuses hear, or are behaviorally responsive to, sound (e.g., Bernard & Sontag, 1947; Birnholz & Benacerraf, 1983; Grimwade, Walker, Bartlett, Gordon, & Wood, 1971; Johansson, Wedenberg, & Westin, 1964; Sontag & Wallace, 1935). Intrauterine recordings taken near term indi- cate that maternal speech and heartbeats are audible in utero (Querleu & Renard, 1981; Querleu, Renard, & Crepin, 1981; Walker, Grimwade, &Wood, 1971). Nonmaternal speech, for example male speech, is less audible because of attenuation by maternal tissue and/or masking by intrauterine sounds (Querleu & Renard, 1981; Querleu et al., 1981).
* This research was supported by a Research Council Grant from the University of North Carolina at Greensboro and a generous equipment loan by Professor Michael D. Zeiler. We wish to thank the medical and administrative staff of Moses H. Cone Hospital, Greensboro, NC and, especially, the mothers and their infants for making this research possible. Thanks also to G. Gottlieb, R. Harter. R. Hunt, R. Panneton. K. Smith, and, especially, W. Salinger for their helpful comments on drafts of the manuscript. Portions of this paper were presented at the Third Biennial Interna- tional Conference on Infant Studies, March 1982. Austin, TX.
Correspondence and requests for reprints should be addressed to Anthony J. DeCasper, Department of Psychology, University of North Carolina at Greensboro, Greensboro, NC 27412.
133

134 DECASPER AND SPENCE
The newborns’ preference for their own mothers’ voices requires that they had some prior experience with her voice, but there is no evidence that the necessary experience occurred after birth. Fifer (1980) failed to find any relation between maternal-voice preference and postnatal age, whether the newborns roomed with their mother or in a nursery, or whether they were breast fed or bottle fed. Since the maternal voice is audible in utero, and since third-trimester fetuses can hear, perhaps the necessary experience occurred before birth. In contrast, newborns show no preference for their own fathers’ voices, even if they had explicit postnatal experience with his voice. Since male voices are not very audible in utero, perhaps the absence of a paternal-voice preference indi- cates the absence of prenatal experience with his voice (DeCasper & Prescott, 1984). The correlations between the presence or absence of specific-voice sounds before birth, and the presence or absence of specific-voice preferences after birth suggest that prenatal auditory experiences influence the earliest voice preferences.
Consider that complex auditory stimuli can function as positive rein- forcers, neutral stimuli, or negative reinforcers of newborn behavior. Known reinforcers include vocal-group singing, solo female singing, prose spoken by a female, synthetic speech sounds, and intrauterine heartbeat sounds (Butter- field & Cairns, 1974; Butterfield & Siperstein, 1972; DeCasper, Butterfield, & Cairns, 1976; DeCasper & Carstens, 1981; DeCasper & Sigafoos, 1983). On the other hand, male speech and instrumental music lack reinforcing value, while white noise and faster-than-normal heartbeat sounds are aversive (Butter- field & Siperstein, 1972; DeCasper & Prescott, 1984; Salk, 1962). The differen- tial reinforcing effectiveness of these sounds seems to covary more with their similarity to sounds that were present in utero than with any general acoustic characteristic(s), which further suggests that prenatal auditory experience in- fluences postnatal auditory perception.
Finally, prenatal auditory experience has been shown to cause postnatal auditory preferences in a variety of infrahuman species (e.g., Gottlieb, 1981; Vince, 1979; Vince, Armitage, Walser, & Reader, 1982).
The hypothesis implies that prenatal experience with maternal speech sounds causes some property of the sounds to be differentially reinforcing after birth. Speech sounds enable at least two kinds of discriminations; some speech cues allow discrimination of language-relevant sounds, per se, or what is said, and some allow discrimination of the speaker or source of the speech sounds (Bricker & Pruzansky, 1976; Studdert-Kennedy, 1982). Thus, the pre- natal experience hypothesis implies that newborns prefer their own mothers’ voices, regardless of what she says, because of prenatal experience with her voice-specific cues. This implication, however, cannot be directly tested for obvious ethical and practical reasons. The hypothesis also implies that new- borns will prefer the acoustic properties of a particular speech passage if their mothers repeatedly recite that passage while they are pregnant.
We directly tested the latter implication in the following way. First, preg- nant women tape-recorded three separate prose passages. Then, they recited

PRENATAL AUDITORY LEARNING 135
one of the passages, their target passage, aloud each day during the last 6 weeks of pregnancy. After birth their infants were observed in an operant learning task where recordings of the target passage and a novel passage, one their mothers had recorded but had not recited, were both available as reinforcers. Then their relative reinforcing effectiveness was evaluated. If the prenatal ex- perience with the target passage increases its reinforcing value then: (a) the acoustic properties of the target passage will be more reinforcing than those of a novel passage; (b) the differential reinforcing value of the target passage should be carried by its language-relevant cues and, thus, should not require the presence of the infant’s own mother’s voice cues; and (c) the reinforcing values of the target and novel passages should not differ for control newborns who had never been exposed to either passage.
METHOD
Prenatal Phase
Pregnant Subjects. Thirty-three healthy women approximately 7 Yz months pregnant were recruited from childbirth preparation classes after being informed about the project. All were experiencing uncomplicated pregnancies.
Prenatal Procedures. After becoming familiar with three short children’s stories they tape-recorded all three. Recordings were made in a quiet room on an Akai 4000 stereophonic tape recorder. The tapes would be used as reinforcers in a postnatal learning task. Each woman was then assigned one of the stories as her target story. Assignment was made after all three had been recorded to prevent them from biasing the recording of their target, for example, by ex- aggerated intonation.
The women were instructed to read their target story aloud “two times through each day when you feel that your baby (fetus) is awake” and to “read the story in a quiet place so that your voice is the only sound that your baby can hear.” They maintained a log of their daily recitations and were occasion- ally checked by the researchers.
Story Materials. The stories were The King, the Mice, and the Cheese (Gurney & Gurney, 1965), the first 28 paragraphs of The Cat in the Hat (Seuss, 1957), and a story we called The Dog in the Fog, which was the last 28 para- graphs of The Cut in rhe Hut with salient nouns changed. The three stories were about equally long, they contained 579, 611, and 642 words, respectively. Each could be comfortably recited in about 3 min. Each was also composed from equal size vocabularies of 152, 142, and 154 words, respectively. Salient, high-frequency nouns common to at least two stories were changed. For exam- ple, cat and hat in The Cut became dog and fog in The Dog, and cat and dog from those stories became turtle and zebra in The King. The Cat contained 46 unique words (i.e., words that appeared only in The Cat), which accounted for 22% of the total word count; The Dog contained 57 unique words, which ac- counted for 22% of the total word count; and The King contained 85 unique

136 DKASPER AND SPENCE
words, which accounted for 44% of the total word count. All three stories con- tained common high-frequency words. For example, a, all, and, did, do, he, I, in, like, not, now, of, said, that, the, to, with, and you occurred at least three times in each. The common high-frequency words accounted for 43% of The Cat, 38% of the The Dog, and 36% of the The King. The remaining words oc- curred at least once in at least two of the stories. The stories also differed in prosodic qualities, such as patterns of syllabic beats. Thus, they differed in the acoustic properties of individual words as well as in prosody. The Cat and The Dog sounded more similar to each other than either did to The King, but we could readily identify the origin of short (several seconds) segments from all three.
Postnatal Phase
Experienced Newborns. Sixteen of the 33 fetal subjects completed test- ing as newborns. The 16 had been prenatally exposed to their target story an average of 67 times or for about 3.5 hours in all. They were tested at an average age of 55.8 hours (SD= 10). Each had to have had an uncomplicated full-term gestation and delivery, a birth weight between 3500-3900 grams, and APGAR scores of 8, 9, or 10 at 1 and 5 min after birth. If a subject was circumcised, he was not tested until at least 12 hours afterward. Parents gave informed consent for the testing and were invited to observe.
Seventeen infants were not tested or did not complete a test session: 5 because their mothers failed to return their logs, 4 because they encountered intrapartum or postpartum difficulties, 5 failed to meet state criteria at the time of testing or cried, and 3 subjects’ sessions were unavoidably interrupted.
Apparatus. Sessions occurred in a quiet, dimly lit room adjacent to the nursery. The infants lay supine in their bassinets and wore TDH-39 earphones, which were suspended from a flexible rod. They sucked on a regular feeding nipple with the hole enlarged to 1 mm. Rubber tubing connected the nipple to a Statham P23AA pressure transducer that was connected to a Grass poly- graph and solid state programming and recording components. Each infant heard a tape recording of his/her target story and a tape recording of a novel story, one of the others their mother had recorded but not recited. Both stories were recorded by the same woman, and each was played on separate channels of the stereo recorder. The tape ran continuously, and sound was electronically gated to the earphones by the automated programming equipment. Intensities averaged 70 dB SPL at the earphones.
Testing Procedures. Sessions began about 2.5 hours after a scheduled feeding in order to maximize the chance of obtaining an awake, alert, and cooperative infant (Cairns & Butterfield, 1974). Each infant was brought to a quiet-alert state before testing could begin (Wolff, 1966) and had to visually fixate and follow an experimenter’s face when he/she spoke to the infant. (If

PRENATAL AUDITORY LEARNING 137
the infant was not alert and did not fixate or follow, he/she was returned to the nursery, and another attempt was made after a later feeding.)
The infant was then placed supine in the bassinet and the earphones were locked in place. One researcher, who could not be seen by the infant and who was blind to the exact experimental condition in effect, held the nonnutritive nipple loosely in the infant’s mouth. Another monitored the equipment. The infant was then allowed 2 min to adjust to the situation and had to emit sucks having negative pressures of at least 20-mm Hg, a pressure normally exceeded by healthy infants. (If the infant failed to suck adequately, he/she was returned to the nursery, and another attempt was made after a later feeding.)
Testing began with 5 min of baseline sucking during which no voices were presented over the earphones. Unconstrained nonnutritive sucking occurs as groups or bursts of individual sucks separated by interburst intervals of several seconds. A sucking burst was defined as a series of individual sucks separated from one another by less than 2 s; when 2 s elapsed without a suck the equip- ment registered the end of the burst. Thus, interburst intervals (IBIS) began 2 s after the last suck of one burst and ended with the onset of the the first suck of the next burst. This criterion accurately captures the burst-pause pattern of newborns’ nonnutritive sucking (see Figure 1). IBIS tend to be unimodally dis- tributed for individual infants, and modal values vary between infants. The baseline was used to estimate the distribution and median value of each infant’s IBIS just before reinforcement began. Differential reinforcement of IBIS began after baseline had been established. (Hereafter, if the infant stopped sucking
Figure 1. Polygraph record of a newborn’s nonnutritive sucking. Wide horizontal marks indicate the onset and offset of o sucking burst. The time between the end of one burst and the beginning of the next denotes on interburst interval. Onset of the narrow event mork denotes that the time criterion, t seconds, has elapsed since the end of the last burst. Vertical lines indicate time in seconds.

138 DECASPER AND SPENCE
for two 1-min periods for any reason, he/she was returned to the nursery and not tested again.)
Reinforcement Contingencies. For eight randomly selected infants, suck- ing bursts that terminated IBIS equal to or greater than the infants’ baseline medians (t) produced the recording of a woman’s voice reciting the target story. Bursts terminating IBIS less than the baseline median were reinforced with the same woman’s recording of a novel story. Thus, only one of the two stories was presented binaurally with the first suck of a burst and remained on until the burst ended. Reinforcement contingencies were completely controlled by the solid-state equipment. Reinforcement contingencies were reversed for the other eight newborns, to control for the effects of any response bias that might arise from either of the contingencies or from changes in the behavioral dis- positions of the infants, for example, arousal or fatigue. Differential reinforce- ment lasted about 20 min.
The same differential reinforcement procedures were used in earlier voice- preference studies (DeCasper & Fifer, 1980; DeCasper & Prescott, 1984). The rationale is based on well established reinforcement procedures that differenti- ate the temporal properties of behavior: Differentially reinforcing a range of IBIS causes the shorter differentially reinforced IBIS to increase in frequency (see newborn studies by DeCasper & Fifer, 1980; DeCasper & Sigafoos, 1983; as well as animal studies by Anger, 1956; Catania, 1970; DeCasper & Zeiler, 1977; Malott & Cumming, 1964).
Subject Controls. Twelve control newborns matched to a prenatally ex- perienced counterpart on sex, race, and median interburst interval of baseline were also tested. They met the same selection criteria and were tested under ex- actly the same conditions as their counterparts, but their mothers had never recited any of the three stories.
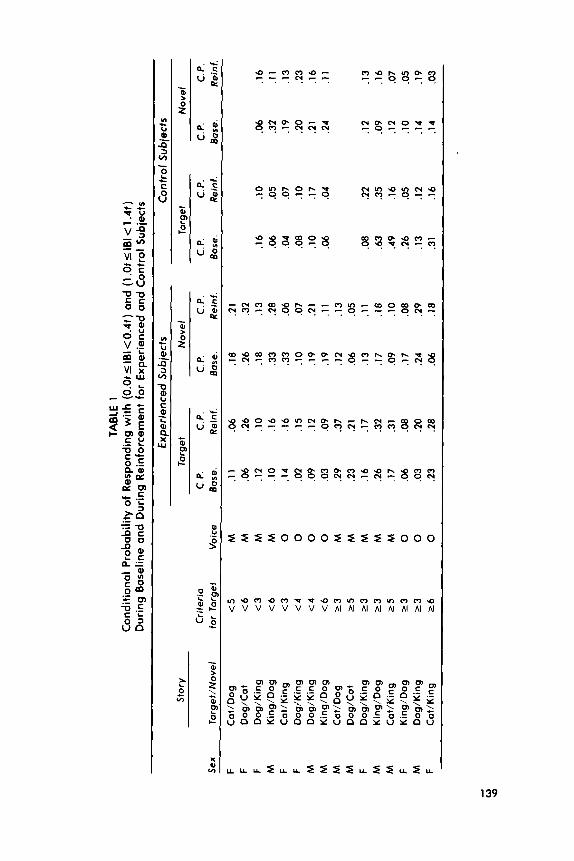
Other Experimental Controls. The influence of mother-specific voice cues on the reinforcing effects of the target stories was controlled by having nine newborns reinforced with recordings made by their own mother and seven with recordings made by some other infant’s mother. Both stories heard by an infant were recorded by the same woman to insure that their reinforcing value could not be unequally influenced by the speaker’s voice characteristics. The acoustic properties of any one story could not systematically influence the re- inforcing value of the target because each of the three stories had served as the target at least four times. No particular combination of target/novel pairings could systematically influence the reinforcing vaiue of the target because five of the six possible target/novel pairings occurred at least twice. Unpredictable subject loss prevented precise counterbalancing of voices and target/novel pairings (see Table 1).
TABL
E 1
Cond
itiona
l Pr
obab
ility
of Re
spon
ding
with
(O
.Ot
5 IB
I <0
.4t)
and
(1 .O
f 5
191
< 1.4
t) Du
ring
Base
line
and
Durin
g Re
infor
ceme
nt for
Ex
perie
nced
an
d Co
ntrol
Subje
cts
Expe
rienc
ed
Subje
cts
Contr
ol Su
bjec
ts
story
Targ
et
Nove
l Ta
rget
No
vel
Crite
ria
c. P.
c.
P.
c. P.
C.
P.
c. P.
Se
x c.
P.
c. P.
c.
P.
Torg
ef/N
ovel
fo
r To
rgef
vo
ice
Bose
. Re
inf.
Bose
. Re
inf.
Bose
. Re
lnf.
Base
. Re
inf.
F Co
t/Dog
<5
M
.ll
.0
6 .I8
.2
1 F
Dog/
Cot
<6
M
.06
.26
.26
.32
F Do
g/Ki
ng
<3
M
.12
.lO
.lB
.13
.16
.lO
36
M
.16
King
/Dog
<6
M
.lO
.1
6 .3
3 .2
8 .0
6 .0
5 .3
2 F
.I1
Cat/K
ing
<3
0 .I4
.1
6 .3
3 .0
6 .0
4 .0
7 .1
9 F
.13
Dog/
King
<4
0
.02
.15
.lO
.07
.00
.lO
.20
.23
M
Dog/
King
<4
0
.09
.12
.19
.21
.lO
.17
.21
.16
M
King
/Dog
<6
0
.03
.09
.19
.11
.06
.04
.24
.11
M
Cot/D
og
23
M
.29
.37
.12
.I3
M
Dog/
Cot
r5
M
.23
.21
.06
.05
F Do
g/Ki
ng
r3
M
.16
.17
.13
.ll
.00
.22
.12
.I3
M
King
/Dog
23
M
.2
6 .3
2 .I7
.I8
.6
3 .3
5 .0
9 M
.1
6 Ca
t/Kin
g 25
M
.I7
.3
1 .0
9 .lO
F
.49
.16
.12
.07
King
/Dog
23
0
.06
.00
.17
.08
.26
.05
.lO
.05
M
Dog/
King
r3
0
.03
.20
.24
.29
.13
.I2
.14
F .I9
Ca
t/Kin
g r6
0
.23
.28
.06
.18
.31
.16
.14
.03

140 DECASPER AND SPENCE
Data Analysis. Interburst intervals were read off the polygraph records. Times between the event marks signalling the end of one burst and the begin- ning of the next burst were measured and rounded down to the nearest whole second (see Figure 1). Thus, the scorers (AJD and MJS), who were highly prac- ticed, did not have to make detailed judgments about IBI values that might bias the data. Interscorer reliability approached 100%.
Next, each subject’s IBIS from the baseline and reinforcement phases were converted to a proportion of their time criterion (t). For example, if t =4 s then all 2-s IBIS had the value O.Sf, and if t = 6 s then 2-s IBIS had the value of 0.331. Converted IBIS were grouped into bins that were 0.21 s wide; Bin 1 con- tained IBIS between O.Ot and 0.2t s, Bin 2 contained IBIS between 0.2t and 0.4f S . . , and Bin 10 contained IBIS between 1.81 and 2.0t s. Bin 11 contained all &Is greater than 2.0f s. IBIS were assumed to be equally distributed within a bin. The conversion equates the relative size of IBIS across subjects and allows averaging over subjects.
RESULTS
Experienced Newborns
The hypothesis asserts that in utero exposure to the acoustic properties of the target story will make it more reinforcing than the novel story. If so, the rela- tive frequency of short IBIS should increase over baseline when reinforced by the target stories in the IBI<t condition and the relative frequency of IBIS slightly longer than the baseline median should increase when reinforced by target stories in the IBI > t condition.
Baseline IBI distributions were examined first in order to determine whether they differed between reinforcement contingencies. They did not dif- fer: A mixed ANOVA of the relative frequencies of baseline IBIS, with Contin- gencies (c t vs. > t) and Bin (l-10) as factors, indicated a significant effect of Bin, F(9, 126) = 13.3, p< X101. The effect merely confirms that the IBIS were unimodally distributed. Most important, there was no Contingency effect, F(1,14) p c 1 .O, and no Contingency x Bin interaction, F(9, 126) = 1.67, p > .lO.
The predictions of the hypothesis were first assessed by examining the differences between the relative frequencies of IBIS that occurred during base- line and those that occurred during reinforcement. Difference scores were entered into a mixed ANOVA with Contingencies (< t vs. > t) and Bin (l-10) as factors. There was no effect of Contingency, F( 1,14) < 1 .O, and a signifi- cant effect of Bin, F(9,126) = 5.48, p-c .025. Most important, there was a sig- nificant Contingency x Bin interaction, F(1,126) = 2.07, pc .05. Planned tests of simple effects confirmed that the interaction occurred because with the IBI < t contingency the relative frequency of short IBIS increased over baseline levels, while those of all other IBIS either decreased or did not change. With the IBI>t contingency the relative frequency of IBIS slightly greater than f seconds increased, while those of the others decreased or did not change. Any IBI between 0 and t seconds would have produced the target story under the

PRENATAL AUDITORY LEARNING 141
IBI < t contingency, and any IBI L t seconds would have produced it under the IBI > t contingency. But only the relative frequencies of the shorter IBIS rein- forced by the targets sytematically increased.
The differential reinforcement effects are more clearly revealed in the analysis of IBIS between O.Ot and 0.4t (the shorter IBIS) and those between 1 .Ot and 1.4t (IBIS slightly longer than t seconds). Conditional probabilities of baseline and reinforced IBIS in these classes were obtained by dividing the rela- tive frequency of IBIS in each class by the relative frequency of that class and all longer IBIS (see Table 1). This is a sensitive measure of temporally differ- entiated responding because: (a) it adjusts the inherently unequal opportunity for infants to emit equal numbers of short and long IBIS in a limited period of time; (b) it measures the probability that an infant will emit a particular class of IBIS given the opportunity to do so (cf. Anger, 1956; DeCasper & Fifer, 1980); and (c) it renders the conditional probabilities of IBIS between O.Ot and 0.4[, and those between 1.01 and 1.41, arithmetically independent of one another. The dependent variables for the target story and for the novel story were their reinforcement ratios: (conditional probabilities of IBIS during rein- forcement) divided by (conditional probability of IBIS during reinforcement) plus (conditional probability of IBIS during baseline).
The average values of baseline conditional probabilities of target-story IBIS and novel-story IBIS did not differ, t(15) = 1.37, p> .lO. However, their reinforcement ratios differed as expected. A mixed ANOVA with Contingency (<t vs. > t) and Interval (O.Ot-0.4t vs. 1 .Ot-1.4t) as factors, revealed no effect of Contingency, F( 1,14) < 1 .O, and no effect of Interval, F( 1,14) = 1.59, p > .20, but a significant Contingency x Interval interaction, F(l, 14) = 6.65, p< .025 (Figure 2). Target-story reinforcement ratios were larger than novel-story rein- forcement ratios, independent of the contingency and of the interval. The fact that 13 of the 16 infants had larger target ratios than novel ratios (p = .Ol 1 by the binomial test) and 13 of the 16 had target-story ratios greater than .50 indi- cates this result was typical. The individual-subject consistency implies that maternal voice cues were not necessary for producing the differential rein- forcement effect. Neither the target-story reinforcement ratios nor the differ- ence between the target ratios and novel ratios differed between the 9 infants who heard their own mothers’ voices and the 7 who heard unfamiliar voices, p-values of both f-tests > .lO.
Control Subjects
The following analysis of control-subject performances parallels that of the experienced subjects. The relative frequency distributions of baseline IBIS did not differ between reinforcement contingency conditions. A mixed ANOVA with Contingency (c t vs. > t) and Bin (l-10) as factors revealed a marginal effect of Bin, F(9,90) = 1.90, .lO<p> .05, but no effect of Contingency, F( 1,lO) < 1 .O, or of the Contingency x Bin interaction, F(9,90) = 1.38, p> .lO. The subsequent mixed ANOVA on the difference scores of IBIS that occurred
142 DKASPER AND SPENCE
.60
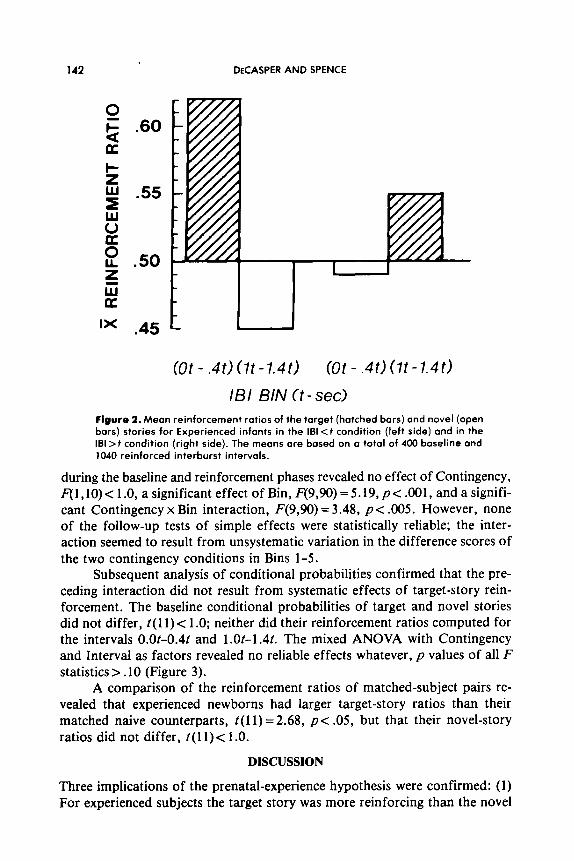
IBI BIN Ct - sec1 Figure 2. Mean reinforcement ratios of the target (hatched bars) and novel (open bars) stories for Experienced infants in the 161 <f condition (left side) and in the 161 >t condition (right side). The means are based on a total of 400 baseline and 1040 reinforced interburst intervals.
during the baseline and reinforcement phases revealed no effect of Contingency, F(1) 10) < 1 .O, a significant effect of Bin, F(9,90) = 5.19, p< .OOl, and a signifi- cant Contingency x Bin interaction, F(9,90) = 3.48, p c .005. However, none of the follow-up tests of simple effects were statistically reliable; the inter- action seemed to result from unsystematic variation in the difference scores of the two contingency conditions in Bins 1-5.
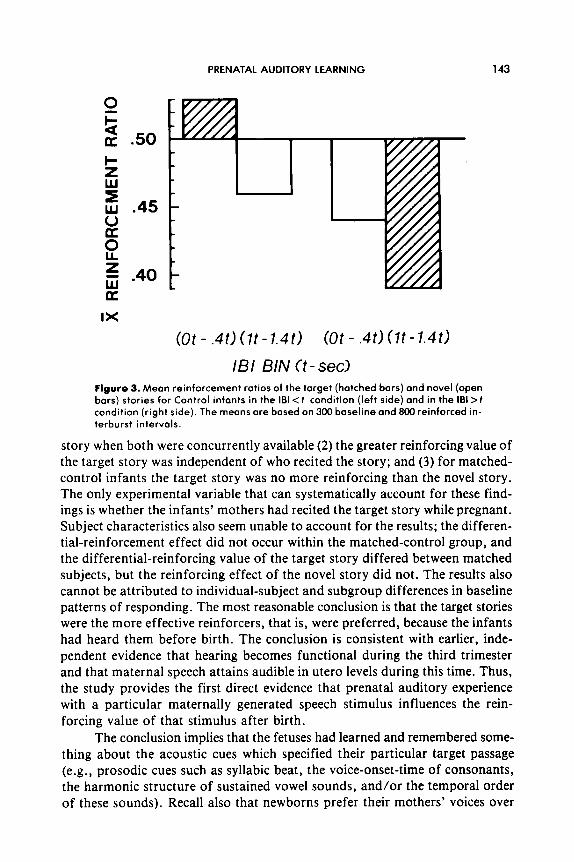
Subsequent analysis of conditional probabilities confirmed that the pre- ceding interaction did not result from systematic effects of target-story rein- forcement. The baseline conditional probabilities of target and novel stories did not differ, t( 11) < 1 .O; neither did their reinforcement ratios computed for the intervals O.Ot-0.4t and 1 .Ot-1.41. The mixed ANOVA with Contingency and Interval as factors revealed no reliable effects whatever, p values of all F statistics > .10 (Figure 3).
A comparison of the reinforcement ratios of matched-subject pairs re- vealed that experienced newborns had larger target-story ratios than their matched naive counterparts, t(l1) =2.68, p< .05, but that their novel-story ratios did not differ, t(1 1)~ 1 .O.
DISCUSSION
Three implications of the prenatal-experience hypothesis were confirmed: (1) For experienced subjects the target story was more reinforcing than the novel
PRENATAL AUDITORY LEARNING 143
IX
(0t - .4tmt-1.4t) (0t - .4tM-7.4t)
IBI BIN Ct-set) Figure 3. Mean reinforcement rotios of the target (hatched bars) and novel (open bars) stories for Control infants in the I81 <t condition (left side) and in the 181 >t condition (right side). The means are based on 300 baseline and 800 reinforced in- terburst intervals.
story when both were concurrently available (2) the greater reinforcing value of the target story was independent of who recited the story; and (3) for matched- control infants the target story was no more reinforcing than the novel story. The only experimental variable that can systematically account for these find- ings is whether the infants’ mothers had recited the target story while pregnant. Subject characteristics also seem unable to account for the results; the differen- tial-reinforcement effect did not occur within the matched-control group, and the differential-reinforcing value of the target story differed between matched subjects, but the reinforcing effect of the novel story did not. The results also cannot be attributed to individual-subject and subgroup differences in baseline patterns of responding. The most reasonable conclusion is that the target stories were the more effective reinforcers, that is, were preferred, because the infants had heard them before birth. The conclusion is consistent with earlier, inde- pendent evidence that hearing becomes functional during the third trimester and that maternal speech attains audible in utero levels during this time. Thus, the study provides the first direct evidence that prenatal auditory experience with a particular maternally generated speech stimulus influences the rein- forcing value of that stimulus after birth.
The conclusion implies that the fetuses had learned and remembered some- thing about the acoustic cues which specified their particular target passage (e.g., prosodic cues such as syllabic beat, the voice-onset-time of consonants, the harmonic structure of sustained vowel sounds, and/or the temporal order of these sounds). Recall also that newborns prefer their mothers’ voices over

144 DECASPER AND SPENCE
that of another female, when both speak the same novel material (DeCasper & Fifer, 1980; Fifer, 1980). The present results add to the evidence indicating that the maternal-voice preference also originated in utero. If so, then fetuses also register some specific information about their mothers’ voices (e.g., spectra of nasals and vowels, glottal frequency and spectrum, and/or the temporal characteristics of pitch, intensity, and formants) (Bricker & Pruzansky, 1976). The specific acoustic cues that register in utero and which influence subsequent perception of speech and voice sounds are not known at present. However, whether language-relevant cues or voice-specific cues play an active role in newborns’ perception has now been shown to depend upon: (a) which class of cues are differentially available, (b) the infants’ prenatal experience with the cues, and (c) the circumstances attending postnatal perception (e.g., behavioral contingencies, infant state, or the presence or absence of other sounds).
The present study suggests noninvasive, ethically acceptable methods to further study the effects of prenatal auditory stimulation on postnatal auditory function and development, especially the development of speech perception. Such research might also benefit clinical treatment of the perinate, for exam- ple, by aiding in the diagnosis of fetal condition and by providing information for designing environments of preterm infants.
Some Post Hoc Considerations
Learning is generally and most satisfactorily inferred from a change in perfor- mance rather than from absolute measures of performance. However, change scores-the difference scores and reinforcement ratios used in this study-are almost always inversely related to prelearning performance, the baseline proba- bilities of responding (cf. Glass & Stanley, 1970, p. 182). The present discussion focuses on the extent to which the preceding inferences about differential- reinforcement effects were influenced by the relation between baseline levels of performance and the difference scores and reinforcement ratios. The issue is salient here because the hypothesis asserts that reinforcement would differen- tially affect specific IBIS whose baseline probabilities varied considerably.
The abscissa of Figure 4 shows the mean conditional probabilities of base- line IBIS for each of the eight subconditions represented in Table 1. The mean baseline conditional probabilities of IBIS between O.Ot and 0.4t (subconditions l-4 in Figure 4) are lower than the mean baseline conditional probabilities of IBIS between 1 .Ot and 1.4t (subconditions 5-8). They differ simply because baseline IBIS between O.Ot and 0.4t come from the left of a unimodal distribu- tion and IBIS between 1 .Ot and 1.4t come from near the median of the distribu- tion. The primary means of experimentally controlling for the influence of these baseline differences was to counterbalance the reinforcers associated with the IBI < t and IBI > I contingencies: As Figure 4 suggests, and as reported earlier, when the values of baseline probabilities are pooled over IBI < f and the IBI > I contingencies (1 with 6; 2 with 5; 3 with 8; 4 with 7) the average baseline probabilities do not differ.
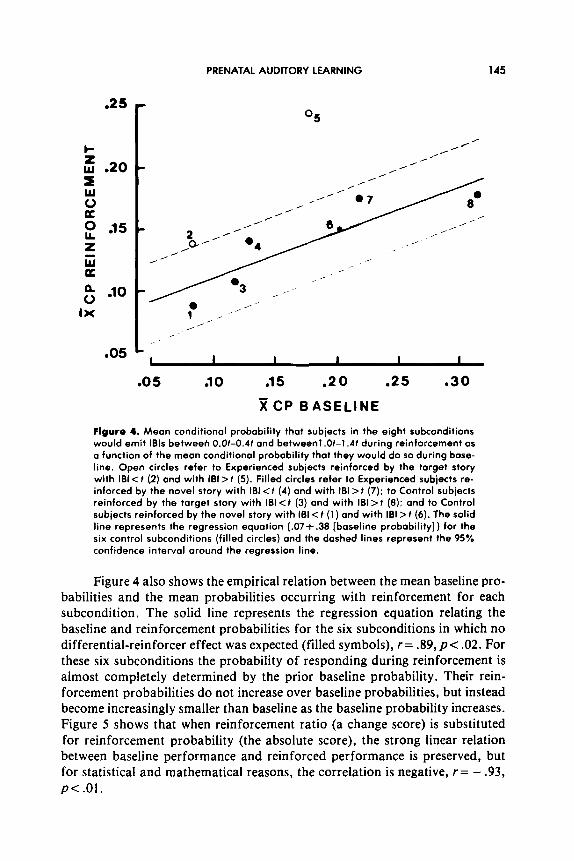
.25
I-
z .20 I
ti
5 .15 z” iii a
$ .lO
IX
PRENATAL AUDITORY LEARNING 145
.05 .l 0 .15 .20 .25 .30
Ti CP BASELINE
Flgure 4. Mean conditional probability that subjects in the eight subconditions would emit IBIS between O.Ot-0.41 and betweenl.Ot-1.41 during reinforcement OS a function of the mean conditional probability that they would do so during base- line. Open circles refer to Experienced subjects reinforced by the target story with IBI < t (2) and with lBl> t (5). Filled circles refer to Experienced subiects re- inforced by the novel story with IBI <t (4) and with IBI >t (7); to Control subjects reinforced by the target story with IBI < t (3) and with 181 > f (8): and to Control subjects reinforced by the novel story with IBI < t (1) and with 181 > t (6). The solid line represents the regression equation (.07+.3B [baseline probability]) for the six control subconditions (filled circles) and the dashed lines represent the 95% confidence interval around the regression line.
Figure 4 also shows the empirical relation between the mean baseline pro- babilities and the mean probabilities occurring with reinforcement for each subcondition. The solid line represents the regression equation relating the baseline and reinforcement probabilities for the six subconditions in which no differential-reinforcer effect was expected (filled symbols), r= .89, p< .02. For these six subconditions the probability of responding during reinforcement is almost completely determined by the prior baseline probability. Their rein- forcement probabilities do not increase over baseline probabilities, but instead become increasingly smaller than baseline as the baseline probability increases. Figure 5 shows that when reinforcement ratio (a change score) is substituted for reinforcement probability (the absolute score), the strong linear relation between baseline performance and reinforced performance is preserved, but for statistical and mathematical reasons, the correlation is negative, r= - .93, p< .Ol.
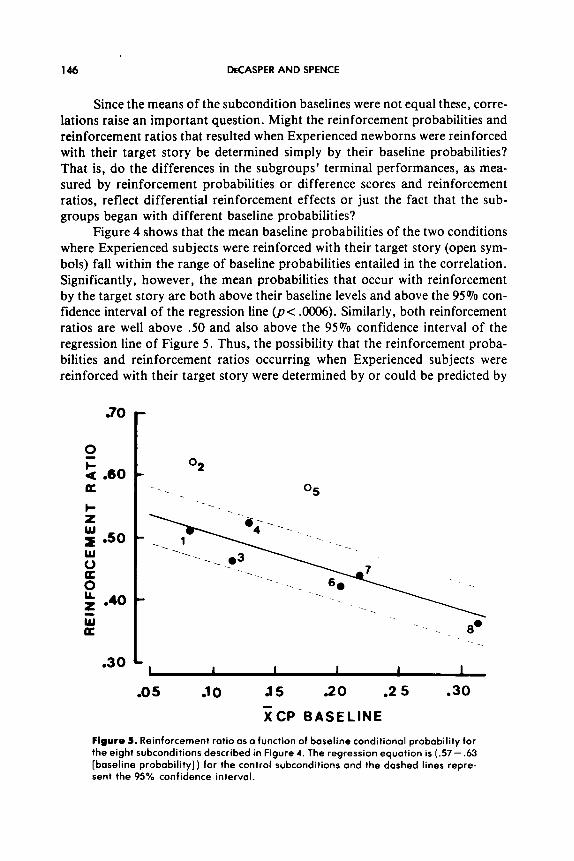
146 DECASPER AND SPENCE
Since the means of the subcondition baselines were not equal these, corre- lations raise an important question. Might the reinforcement probabilities and reinforcement ratios that resulted when Experienced newborns were reinforced with their target story be determined simply by their baseline probabilities? That is, do the differences in the subgroups’ terminal performances, as mea- sured by reinforcement probabilities or difference scores and reinforcement ratios, reflect differential reinforcement effects or just the fact that the sub- groups began with different baseline probabilities?
Figure 4 shows that the mean baseline probabilities of the two conditions where Experienced subjects were reinforced with their target story (open sym- bols) fall within the range of baseline probabilities entailed in the correlation. Significantly, however, the mean probabilities that occur with reinforcement by the target story are both above their baseline levels and above the 95% con- fidence interval of the regression line @C .0006). Similarly, both reinforcement ratios are well above SO and also above the 95% confidence interval of the regression line of Figure 5. Thus, the possibility that the reinforcement proba- bilities and reinforcement ratios occurring when Experienced subjects were reinforced with their target story were determined by or could be predicted by
.70
0
2 .60 a
L
r” .50
Y
i5 P .40 I
iii a
.05 30 15 30 .2 5 .30
iCP BASELINE
Figure 3. Reinforcement ratio as o function of baseline conditional probobility for the eight subconditions described in Figure 4. The regression equotion is (.57- .63 [baseline probobility]) for the control subconditions and the doshed lines repre- sent the 95% confidence interval.

PRENATAL AUDITORY LEARNING 147
their baseline probabilities can be rejected. The favored alternative hypothesis, of course, is that prenatal experience increased the reinforcing effectiveness of their target stories: The effect of prenatal experience with the reinforcer was to increase the conditional probability of reinforced responding by 40% over the level predicted by baseline in the IBI c f condition and by 76% in the IBI > t condition. Reinforcement ratios were increased by 20% and 26% over the levels predicted by baseline performance.
It may still be argued, however, that the preceding analysis was based on subgroup means and that the pattern of individual-subject baseline probabili- ties within the subgroups was biased toward producing difference scores and reinforcement ratios that supported the prenatal hypothesis. That is, if the baseline probability of each Experienced subject reinforced with the target story had been the same as the baseline of a control subject, then their rein- forcement probabilities and reinforcement ratios might not differ.
The following analyses addressed this possibility by comparing selected groups of subjects after matching individual infants on baseline probabilities. Subject matching was accomplished by applying the following three rules: (1) baseline probabilities had to be within f .02 of each other; (2) if possible, the subjects were to have the same reinforcement contingency; and (3) if more than one match was possible, pairs were matched so as to minimize the differ- ence between conditional probabilities that occurred with reinforcement. No other factors were considered.
In the first comparison, 10 of the 16 baseline probabilities produced by Experienced infants in the conditions where they were reinforced with the tar- get story were matched to the baseline probabilities of 10 of the 16 Experienced subjects in the conditions where they were reinforced with the novel story. The 10 baseline pairs were: (.06/.06), (.06/.06), (.09/.09), (.lO/.lO), (.12/.12), (.14/.13), (.16/.17), (.17/.17), (.23/.24), and (.26/.26). The mean baseline probability for each group was .14. The mean probability occurring with rein- forcement by the target story (.20) was greater than that occurring with rein- forcement by the novel story (.15), t((9)=2.61, pc.01 (l-tail I test) T=4.5, p < .025 (Wilcoxen test). the mean probability of responding with target-story reinforcement was greater than the baseline mean, f(9) =2.99, p< .005; T=3, p < .Ol, but the mean probability of responding with novel-story reinforcement did not differ from baseline, t(9) = .58; T= 22.5. In addition, the mean occur- ring with target-story reinforcement was well above the 95% confidence inter- val of Figure 4, but the mean occurring with novel-story reinforcement was well within the interval. The reinforcement ratio of the target story was well above the 95% confidence interval of Figure 5, but the reinforcement ratio for the ,novel-story was within the interval.
Six infants from each reinforcement condition could not be matched. The mean of the unmatched baselines for the target-story condition was .12; the mean occurring with reinforcement was .18. For the novel-story infants

148 * DECASPER AND SPENCE
these means were .22 and .17, respectively. The reinforcement mean and rein- forcement ratio occurring with target-story reinforcement were above the 95% confidence intervals of Figures 4 and 5, but the analogous measures resulting from reinforcement with the novel story were well within the confidence inter- vals.
Next, baselines of Experienced subjects who were reinforced with the tar- get story were matched to baselines of Control subjects reinforced with the target story. Nine pairs could be formed: (.03/.04), (.06/.06), (.06/.06), (.09/.08), (.lO/.lO), (.14/.13), (.16/.16), (.26/.26), and (.29/.31). The baseline mean of each group was .13. The mean reinforcement probability for the Experienced subjects (.19) was greater than that of Control infants (.lO), r(8) = 2.58,p< .Ol; T= 1, p < .005. The Experienced subjects’ reinforcement probabilities were larger than baseline probabilities, t (8) = 2.94, p< .005; T= 0, pc .005, but the Control subjects’ were not, f(8) = 1.19, p> .lO; T= 15. Here, too, the Experi- enced subject’s mean reinforcement probability and reinforcement ratio were both well above the 95% confidence intervals of Figures 4 and 5. The reinforce- ment probability of the Control group was within the interval of Figure 4. Their reinforcement ratio, however, fell below the 95% confidence interval of Figure 5, even though the members of the group had exactly the same baseline probabilities as their Experienced counterparts.
Seven Experienced subjects’ and three Control subjects’ baselines could not be matched. The baseline means of these subjects are .13 and .40, respec- tively. Their respective means occurring with reinforcement by the target story were .19 and .20. The reinforcement mean and reinforcement ratio of the Ex- perienced subjects both lay well above the 95 % confidence intervals of Figures 4 and 5. Analogous scores for Control subjects were within the intervals.
Finally, Experienced subjects reinforced with the novel story were matched to Control subjects who were also reinforced with the novel story. The nine pairs of baseline probabilities were: (.06/.06), (.09/.09), (.lO/.lO), (.12/.12), (.13/.13), (.19/.20), (.19/.19), (.24/.24), and (.33/.32). The mean for each group was .16. Neither the between-group difference nor the changes from baseline were statistically reliable (all t values < 1 .O; all T values > 16). All re- inforcement means and reinforcement ratios fell within the confidence intervals of Figures 4 and 5. Seven Experienced infants and three Control infants could not be matched. Their respective baseline means were .19 and .16, and their respective reinforcement means were .18 and .13. All reinforcement means and reinforcement ratios were within the confidence intervals of Figures 4 and 5.
After equating the baseline probability of IBIS of individual infants in specific conditions, the only consistent finding was that the target story was the more effective reinforcer for Experienced infants. In sum, the results of this study cannot be accounted for by differences in the baseline values of subcon- ditions or individual subjects. The previous conclusion can be retained: The postnatal reinforcing value of a speech passage is increased by prenatal experi- ence with the passage.

PRENATAL AUDITORY LEARNING 149
REFERENCES
Anger, D. (1956). The dependence of interresponse times upon the relative reinforcement of dif- ferent interresponse times. Journal of Experimental Psychology, 52. 145-161.
Bernard, J., & Sontag, L.W. (1947). Fetal reactivity to tonal stimulation: A preliminary report. Journal of Generic Psychology, 70, 205-210.
Birnholz, J.C., & Benacerraf, B.R. (1983). The development of human fetal hearing. Science, 222, 517-519.
Brazelton, T. B. (1978). The remarkable talents of the newborn. Birth & Family Journal, 5,4-10. Bricker, P.D., & Pruzansky, S. (1976). Speaker recognition. In N.J. Lass (Ed.), Contemporary
issues in experimenral phonetics. New York: Academic. Butterfield, E.C., & Cairns, G.F. (1974). Whether infants perceive linguistically is uncertain, and
if they did its practical importance would be equivocal. In R.L. Scheifelbush & L.L. Lloyd (Eds.), Language perspectives: Acquisition, retardation, and intervention. Baltimore, MD: University Park Press.
Butterfield, E.C., & Siperstein, G.N. (1972). Influences of contingent auditory stimulation upon nonnutritional sucking. In J. Bosma (Ed.), Oral sensation and percepfion: The mouth of the infant. Springfield, IL: Charles C. Thomas.
Cairns, G., & Butterfield, E.C. (1974). Assessing infants auditory functioning. In B.Z. Fried- lander, G.M. Sterritt, & G.C. Kirk (Eds.), Exceptionalinfant: Vol. 3. New York: Brunner/ Mazel.
Catania, C.C. (1970). Reinforcement schedules and psychophysical judgments: A study of some temporal properties of behavior. In W.N. Schoenfeld (Ed.), The theory of reinforcement schedules. New York: Appleton-Century-Crofts.
DeCasper, A.J., Butterfield, E.C., & Cairns, G.F. (1976). The role of contingency relafions in
speech discrimination by newborns. Paper presented at the Fourth Biennial Conference on Human Development, Nashville, TN.
DeCasper, A.J., & Carstens, A.A. (1981). Contingencies of stimulation: Effects on learning and emotion in neonates. Infant Behavior and Development. 4, 19-35.
DeCasper, A.J., & Fifer, W.P. (1980). Of human bonding: Newborns prefer their mother’s voices. Science, 208, 1174-1176.
DeCasper, A.J., & Prescott, P.A. (1984). Human newborns’ perception of male voices: Prefer- ence, discrimination and reinforcing value. Developmental Psychobiology, 17, 481-491.
DeCasper, A.J., & Sigafoos, A.D. (1983). The intrauterine heartbeat: A potent reinforcer for newborns. Infant Behavior and Development, 6, 19-25.
DeCasper, A.J., & Zeiler, M.D. (1977). Time limits for completing fixed ratios: IV. Components of the ratio. Journal of the Experimental Analysis of Behavior, 27, 235-244.
Fifer, W.P. (1980). Early attachment: Maternal voice preferences in one- and three-day old in- fants. Unpublished doctoral dissertation, University of North Carolina at Greensboro.
Glass, G.V., & Stanley, J.C. (1970). Statistical methods in education andpsychology. Englewood Cliffs, NJ: Prentice-Hall.
Gottlieb, G.G. (1981). Roles of early experience in species-specific perceptual development. In R.N. Aslin, J.R. Alberts, & M.R. Petersen (Eds.), Development of perceplion. New York: Academic.
Grimwade, J.C., Walker. D.W., Bartlett, M., Gordon, S., & Wood, C. (1971). Human fetal heart rate change and movement in response to sound and vibration. American Journal of Ob- sretrics and Gynecology, 109, 86-90.
Gurney, N., & Gurney, E. (1965). The king, fhe mice, and fhe cheese. New York: Beginner Books/ Random House.
Johansson, B., Wedenberg, E., & Westin, B. (1964). Measurement of tone response by the human fetus: A preliminary report. Acta Ofo-laryngologica, 57, 188-192.
Malott, R.W., & Cumming, W.W. (1964). Schedules of interresponse time reinforcement. Psycho- logical Record, 14, 21 l-252.

150 . DECASPER AND SPENCE
Panneton, R.K.. & DeCasper, A.J. (1984). Newbornsprefer intrauterine heartbeat sounds to male voices. Paper presented at the International Conference on Infant Studies, New York.
Querleu, D., & Renard, K. (1981). Les perceptions auditives du foetus humain. Medicine & Hy- giene, 39, 2102-2110.
Querleu. D., Renard. K., & Crepin, G. (1981). Perception auditive et reactivite foetale aux stimu- lations sonores. Journal de Gynecologie Obstetrique et Biologic de la Reproduction, 10, 307-314.
Salk. L. (1962). Mothers’ heartbeat as an imprinting stimulus. Transactions of the New York Academy of Science, 24, 753-763.
Seuss, D. (1957). The cat in the hut. New York: Beginner Books/Random House. Sontag, L.W., &Wallace, R. (1935). The movement response of the human fetus to sound stimuli.
Child Development, 6, 253-258. Studdert-Kennedy, M. (1982). The beginnings of speech. In K. Immelmann, G.W. Barlow, L.
Petrinovich, & M. Main (Eds.), Behauiorul development. Cambridge: Cambridge Univer- sity Press.
Vince, M.A. (1979). Postnatal effects of prenatal sound stimulation in the guinea pig. Animal Behaviour, 27, 908-919.
Vince, M.A., Armitage, S.E., Waiser, E.S., & Reader, M. (1982). Postnatal consequence of pre- natal sound stimulation in the sheep. Behovior, 8I, 128-139.
Walker, D., Grimwade, J., & Wood, C. (1971). Intrauterine noise: A component of the fetal en- vironment. American Journal of Obstetrics and Gynecology. 109, 91-95.
Wolff, P.H. (1963). Observations on the early development of smiling. In B.M. Foss (Ed.), Deter- minants of infant behavior (Vol. 2). New York: Wiley.
Wolff, P.H. (1966). The causes, controls and organization of behavior in the neonate. Psycho- logical Issues, 5 (1, Monograph No. 1).
1 January 1985; Revised 20 December 1985 W