
Power point for BU on neuro transmmitters.
39
Power point for BU on neuro transmmitters. Note aspartate glutamate phenylalanine
Transcript of Power point for BU on neuro transmmitters.

Power point for BU on neuro transmmitters.
Note aspartate glutamate phenylalanine

• Excitatory amino acid transmitters• —are aliphatic amino acids that have two
acidic functions and one alpha-amino group.
• These are glutamate and aspartate

GLUTAMIC ACID (GLUTAMATE)

ASPARTIC ACID (ASPARTATE)

• 1. Glutamate (Figure 22-11)• —is a major excitatory neurotransmitter of the brain. It is
used by virtually all excitatory neurons in the CNS and is the neurotransmitter of 50% -60% of the neurons in the CNS.
• 50% -60 of brain synapses are glutamatergic. • Glutamate is thus the most common
neurotransmitter in the brain.• Glutamate or a structurally-similar chemical is an
excitatory neurotransmitter in many areas of the brain. • Stimulation of EAA (Excitatory amino acids) receptors
increases cation conductance, leading to depolarization, or stimulates phosphatidyl inositol turnover.
• Glutamate is always excitatory, usually due to simple receptors that increase the flow of positive ions by opening ion-channels

• Glutamate• —is the neurotransmitter of the cerebellar granule cell.• —is used by the corticobulbar and corticospinal tracts.• —is used by dorsal root ganglion cefls.• —is believed to be involved in long-term potentiation of
hippocampal neurons via N-methyl-D-aspartate (NMDA) receptors.
• —plays a role in kindling-induced seizures.• —plays a role in pain transmission (in A delta and C
fibers).• —Neocortical glutamatergic neurons project to the
striatum, the sub thalamic nucleus, and the thalamus. • The subthalamic nucleus projects glutamatergic fibers to
the globus pallidus. • Excitatory amino acids such as glutamate are thought to
be important in learning, memory and other brain functions.


• Glutamate is synthesized from the amino acid glutamine in the axon terminal and, after release, is rapidly taken up by transporters in presynaptic terminals and in astrocytes.
• Astrocytes then convert glutamate to glutamine; glutamine is released by the astrocytes, taken up, and reused by the presynaptic terminal.
• There are 3 classes of ionotropic glutamate receptors named for the compounds that activate them.
• a. 2-(Aminomethyl)phenylacetic acid (AMPA) and kainate receptors consist of 6 different proteins that form the 5 subunits around an ion channel.
• b. Activation of these receptors results in a rapid increase in the conductance of Na and K ions.
• c. The NMDA receptor which is possibly the most complicated of all neurotransmitter receptors consists of 5 subunits.
• d. Activation of the NMDA receptor results in an increase in the conductance of Na ions and Ca ions and the production of EPSPs.

• N-Methyl-D-Aspartate is a synthetic chemical not naturally found in biological systems, but it binds specifically to the NMDA glutamate receptor (receptors are frequently named for artificial substances that bind to the receptor with higher specificity than their natural neurotransmitter ligands).
• The NMDA receptor is the only known receptor which is regulated both by a ligand (glutamate) and by voltage.
• There are at least 5 binding sites which regulate NMDA receptor activity, ie, sites for (1) glutamate (2) glycine (3) magnesium (4) zinc and (5) a site that binds the hallucinogenic substance phencyclidine (PCP, "angel dust").
• Phencyclidine can induce psychosis -- an NMDA effect that is difficult to explain.
• NMDA receptors have a capacity for an activity-dependent increase in synaptic efficiency known as LTP (Long-Term Potentiation), which may be crucial to some forms of learning and memory.
• Inhibition of NMDA activity (and LTP) is believed to be an important part of the way ethanol affects brain functions.

• Long-term potentiation (LTP) occurs at NMDA glutamate receptors (best studied in the hippocampus) and may be a mechanism used by CNS neurons in learning and memory.
• a. When the postsynaptic cell membrane is at its resting potential, glutamate does not bind with NMDA receptors and Ca ions are prevented from entering the cell because Mg ions block the channel.
• b. LTP occurs when 2 simultaneous presynaptic excitatory inputs depolarize the postsynaptic membrane and remove the Mg block of the NMDA receptor.
• c. An influx of Ca occurs, which results in increased protein synthesis, an increase in the strength of the NMDA receptor, and production of more NMDA receptors.
• d. The increased postsynaptic response may also result in the release of nitric oxide from the postsynaptic terminal; nitric oxide acts as a retrograde signal that directs the presynaptic terminal to release more glutamate in response to additional action potentials.
• Inhibition of NMDA activity (and LTP) is believed to be an important part of the way ethanol affects brain functions

• Long-term depression (LTD) (best studied in the cerebellum) results from synaptic transmission between excitatory parallel fibers (axons of granule cells) and Purkinje cells.
• a. Activated parallel fibers release glutamate, which binds with AMPA receptors and causes a brief depolarization of Purkinje cells.
• b. AMPA receptor binding generates second messengers that cause Ca to be released from the endoplasmic reticulum of the Purkinje cell and pumped out of the cell.
• c. The Ca release causes LTD by making the parallel fiber synapses less effective.

• Glutamate stimulation is terminated by a (chloride-independent) membrane transport system that is only used for re-absorbing glutamate & aspartate across the pre-synaptic membrane.
• Glutamate (like aspartate) re-enters the cell by a transporter driven by the high extracellular concentrations of Na+ and the high intracellular concentrations of K+.
• Sodium enters the cell along with the amino acids and potassium leaves the cell -- much the way a pulley couples the lifting of a light weight with the fall of a heavier weight.
• Thus, glutamate/asparate entry is indirectly powered by the ATP-driven Na+-K+-ase (sodium pump) which creates the high ion concentration gradients.

• DISEASE CORRELATIONS• Glutamate-induced excitotoxicity is implicated in the pathogenesis of
Alzheimer Disease, Huntington’s Disease, stroke, epilepsy and amyotrophic lateral sclerosis (ALS).
• Riluzole (Rilutek®), protects neurons from glutamate toxicity in animals and minimally slows progression of ALS.
• lschemic neurons in stroke patients may have gIutamate-jnduced excitotoxicjty, in which glutamate accumulates in the synaptic cleft, because the uptake process is energy dependent and is slowed by oxygen deprivation.
• Epilepsy,which is characterized by multiple seizures that arise from excessive neuronal activity in the cerebral cortex, may also cause glutamate-induced excitotoxicity of gamma-aminobutyric acid (GABA) neurons, increasing the likelihood of additional seizures.
• Excessive levels of glutamate result from an increase in the influx of Ca into the postsynaptic cell at N-methyl (NMDA) receptors. The in flux of Ca may result in the production of free radicals in the postsynaptic cell, which damage the membranes of its organelles and the plasma membrane. Disruption of the plasma membrane results in an influx of water, which leads to swelling and eventual lysis of the cell.
• Blockage of the NMDA Ca receptors that respond to glutamate may be effective in reducing neuronal loss in stroke patients.

• 2.Aspartate (see Figure 22-11)• —is a major excitatory transmitter of the
brain.• —is the transmitter of the climbing fibers of
the cerebellum.


• Like glycine, apartate is primarily localized to the ventral spinal cord.
• Like glycine, aspartate opens an ion-channel and is inactivated by reabsorption into the pre-synaptic membrane.
• Unlike glycine, however, apartate is an excitatory neurotransmitter, which increases the likelihood of depolarization in the postsynaptic membrane.
• Aspartate & glycine form an excitatory/inhibitory pair in the ventral spinal cord comparable to the excitatory/inhibitory pair formed by glutamate & GABA in the brain.
• Interestingly, the two excitatory amino acids -- glutamic acid & aspartic acid -- are the two acidic amino acids found in proteins, that have two carboxyl groups rather than one.

Dopamine

• DOPAMINE . • Characteristics- dopamine• —is a catecholamine.• —can be identified by the marker tyrosine hydroxylase. • —is a neurotransmitter that inhibits the transmission of nerve
impulses--in the substantia nigra, basal ganglia, and corpus striatum of the brain
• —plays a role in cognitive motor, and neuroendocrine functions.• —is depleted in Parkinson disease; so low levels of dopamine
are associated with Parkinson's disease.• —has increased production in schizophrenics (dopamine
hypothesis of schizophrenia)• — is controlled by the enzyme Monamine Oxidase (MAO-B)
which inhibits its production• — since nicotine inhibits MAO it therefore raises level of
dopamine with euphoric results.• like serotonin, dopamine apparently is a neurotransmitters only
within the CNS.
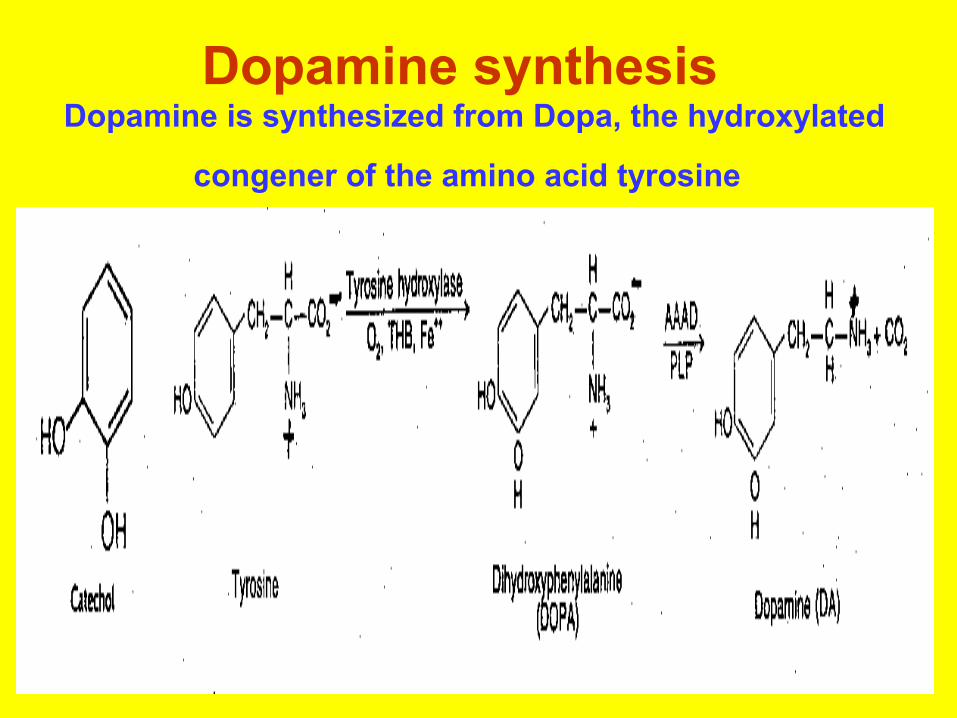
Dopamine synthesis Dopamine is synthesized from Dopa, the hydroxylated
congener of the amino acid tyrosine

• The amino acid tyrosine is not an essential amino acid because it can be synthesized in the liver from phenylalanine by the enzyme phenylalanine hydroxylase.
• But it cannot be synthesized in the brain, and therefore must enter the brain by the large neutral amino acid transporter, which also transports phenylalanine, tryptophan, methionine and the branch-chained amino acids.
• These amino acids all compete for the transporter, so a large quantity of one of the other amino acids in the blood stream could greatly limit the amount of tyrosine entering the brain.
• One case in which this occurs is when there is a liver deficiency of phenylalanine hydroxylase. In that case, phenylalanine reaches high concentrations in the blood and monopolizes the large neutral amino acid transporter, producing the mental retardation of phenylketonuria.

• Once in the brain, tyrosine can be converted to DihydrOxyPhenylAlanine (DOPA) by the tyrosine hydroxylase enzyme using oxygen, iron and TetraHydroBiopterin (THB) as co-factors.
• High concentrations of dopamine inhibit tyrosine hydroxylase activity through an influence on the THB co-factor.
• DOPA is converted to dopamine by Aromatic Amino Acid Decarboxylase (which is fairly nonspecific insofar as it will decarboxylate any aromatic amino acid) using PyridoxaL Phosphate (PLP) as a co-factor.
• This reaction is virtually instantaneous unless there is a Vitamin B6 deficiency.
• Dopamine & epinephrine are primarily inhibitory neurotransmitters that produce arousal. This may sound paradoxical, but the most likely explanation for this effect is that the postsynaptic cells for catecholamines themselves are inhibitory.
• There are 3-4 times more dopaminergic cells in the CNS than adrenergic cells.
• Dopamine in the caudate nucleus facilitates posture, whereas dopamine in the nucleus accumbens is associated with an animal's speed (and pleasure).

• Both dopamine & norepinephrine are catabolized (degraded) by a two-step process involving the enzymes MonoAmineOxidase (MAO) and Catechol-O-MethylTransferase (COMT).
• COMT is primarily active in the synapses, and uses & S-Adenosyl Methionine (SAM) as a methyl-group donor.
• MAO is primarily active in the pre-synaptic terminal against catecholamines that are not safely enclosed in storage vesicles.
• Normally, COMT only catabolizes about 10% of synaptic catecholamine, since catecholamine synaptic activity is primarily terminated by re-uptake into the pre-synaptic neuron terminal.
• MAO accounts for a much larger portion of catecholamine metabolism.
• The darkly pigmented neurons in the pars compacta of the substantia nigra accounts for 80% of the dopamine in the brain.
• The dark pigment neuromelanin is a dopamine polymer that makes the substantia nigra appear black.
• Motor control in the striatum (caudate nucleus and putamen) is thought to involve a balance between inhibitory dopaminergic (D2) and excitatory cholinergic neurons.
• NOTE: Dopamine is degraded by monoamine oxidase A in the brain and monoamine oxidase B outside the CNS and by catechol-O-methyl transferase (COMT).

DOPAMINE RECEPTORS• There are two primary dopamine receptor-types:
D1 (stimulatory) and D2 (inhibitory), both of which act through G-proteins.
• Both subtypes reside in numerous regions of the brain.
• Activation of either subtype inhibits the rate of neuronal firing.
• D2 receptors often occur on the dopaminergic neurons, partially for the purpose of providing negative feedback.
• These so-called autoreceptors can inhibit both dopamine synthesis and release.

• The binding of dopamine to D1-receptors stimulates the activity of Adenylyl Cyclase (AC), which converts ATP to cyclic AMP (cAMP), a second messenger which binds to Protein Kinase A (PKA).
• PKA then modulates the activity of various proteins by the addition of phosphate.

• No specific D1 agonists have been identified.
• We have no drugs to specifically or selectively block the activity of dopamine at D1 receptors.
• Apomorphine is a D2 agonist.

DOPAMINE RECEPTORS

DOPAMINERGIC TRACTS• There are 4 main dopaminergic tracts or pathways in the brain: • (1) the nigrostriatial tract from the substantia nigra to the corpus striatum
accounts for most of the brain's dopamine ( loss or destruction of these dopaminergic (i.e., dopamine-releasing) neurons gives rise to the clinical manifestations of Parkinson's Disease )
• (2) the tuberohypophyseal (or tuberoinfundibular) tract from the arcuate nucleus of the hypothalamus to the portal vessels of the infundibulum near the pituitary stalk, which has a controlling effect on the release of the hormones prolactin through tonic inhibition via D2 receptors; —Released dopamine inhibits the release of prolactin from the adeno hypophysis
• (3) the mesolimbic tract from the ventral tegmental area projects to all cortical and subcortical structures of the limbic system;
• —is linked to behavior and schizophrenia.• . (4) the mesocortical tract from the ventral tegmental area to the
neocortex, particularly the prefrontal area. Dopamine cells project topographically to the areas they innervate.
• NOTE that Neurons of the chemoreceptor trigger zone of the medulla, which controls vomiting are also dopaminergic.

• The mesolimbic & mesocortical dopaminergic systems are thought to play an important role in motivation, by attaching cognition of incentive significance to stimuli.
• In experiments on animals that are motivated to electrically self-stimulate themselves with electrodes implanted in their brains, dopamine is the mediating neurotransmitter for the locus ceruleus, lateral hypothalamus, ventral tegmental area and sulcal prefrontal cortex (but not the nucleus accumbens or substantia nigra).

• Cocaine particularly increases dopaminergic activity in the mesolimbic areas of the brain by inhibiting dopamine re-uptake in the ventral tegmental area and the nucleus accumbens.
• Amphetamine not only inhibits re-uptake, but also releases dopamine from most brain regions.
• Both cocaine & amphetamine produce feelings of psychological energy & arousal, associated with diminished appetite & need for sleep. Both cocaine & amphetamine can lead to visual & tactile hallucinations as well as paranoid thinking, although the psychotic effects of amphetamine may also be mediated by increased serotonin release.
• Chronic amphetamine users seem to lose a capacity for normal pleasure -- which has been correlated with neuron degeneration in the mesolimbic area.

• Schizophrenia is thought to be due to an overstimulation of D2 receptors in the mesolimbic and mesocortical systems.
• Evidence for the "excess dopamine" theory of schizophrenia comes largely from the fact that D2 antagonist drugs alleviate the symptoms, whereas substances which increase D2 stimulation, such as amphetamines, can induce psychotic symptoms (which are reversible with D2 antagonists).

DOPAMINE PATHWAYS



• Other actions of dopamine include • the induction of vomiting by stimulation of
D2 cells in the chemoreceptor trigger zone,• stimulation of growth hormone release by
D2 receptors, and • increased exploration & locomotion
(thought to be connected to dopaminergic activity in the nucleus accumbens).
• Sexual behavior in the male is increased by dopamine agonists, whereas sexual behavior in the female is increased by dopamine antagonists.

• Perception of time-intervals is believed to be mediated by spiny neurons located in the striatum of the basal ganglia. Timing begins with a burst of dopamine and ends with a recognized signal.
• Marijuana slows subjective time by lowering dopamine available, whereas cocaine and methamphetamine accelerates the sense of time by increasing dopamine availability.
• (Adrenaline and stress hormones can also "make seconds feel like hours".)
• The natural brain amine phenylethylamine (PEA, found in chocolate) has been associated with the "love-excitement" of sexual attraction & emotional infatuation.
• PEA concentrations are normally highest in the nucleus accumbens (a "reward center")followed by the frontal & cingulate cortices.
• Levels spike during orgasm and ovulation. PEA is very similar to amphetamine in chemical structure and may likewise act by causing dopamine release, but endorphin release may be a significant effect.
• PEA is preferentially oxidized by MonoAmine Oxidase-B (MAO-B), which may account for the anti-depressant effects of selegiline.

Two diseases occuring in the striatum and substantia nigra.
• Parkinson disease• —results from degeneration of dopaminergic neurons
found in the pars compacta of the substantia nigra.• —results in a reduction of dopamine in the striatum
and in the sub stantia nigra.• —results in the formation of Lewy bodies,
intraneuronal inclusions in the substantia nigra.• Huntington disease (Huntington chorea)• —results from a loss of ACh- and GABA-containing
neurons in the striatum (caudatoputamen).• —results in a loss of GABA in the striatum and
substantia nigra.• PLEASE NOTE THE DIFFERENCES

• What is Parkinsons disease? • Parkinsons disease is a progressive debilitating,
degenerative neurological disorder which causes the premature loss of dopamine producing nerve cells in the brain
• The disease is caused by decreased dopamine neurotransmission in the nigrostriatal pathway secondary to degradation of dopaminergic neurons that project from the substantia nigra (black substance) to the striatum (caudate and putamen).
• When the degeneration of nerves called the substantia nigra (in the centre of the brain) is such that 80% of dopamine is lost, the symptoms of Parkinsons disease are produced.

This syndrome covers several diseases of different etiologies which affect primarily the pigmented neuronal groups including the substantia nigra, locus ceruleus, dorsal motor nucleus of cranial nerve X and the substantia innominata.
• Most cases are sporadic. It affects about one in 500 people; approximately one percent of people over 60 have Parkinsons. Parkinson’s Disease affects a half million Americans.

• The causes of Parkinson’s is not completely understood
• PARKINSONS disease may • occur naturally, or be • drug induced• Naturally Occuring Parkinsons• Pathophysiology : Parkinson’ disease is movement
disorder due to dysfunction in the basal ganglia and associated brain structures as a result of a decrease in dopamine levels.
• Affected CNS levels : striatal dopamine & degeneration of dopaminergic neurons nigrostriatal inhibit activity of striatal GABAergic neurons,
• Reduction of normal dopaminergic neurotransmission leads to excessive excitatory actions of cholinergic neurons ( striatal GABAergic neurons)
• As a result the dopamine & acetylcholine levels in the brain become out of balance.
![Neuro Assessment for Scalp the Non-Neuro Nurse … · Neuro Assessment for the Non-Neuro Nurse Terry M. Foster, RN, ... Microsoft PowerPoint - Neuro Grand Forks ND [Read-Only] Author:](https://static.fdocuments.net/doc/165x107/5b88746b7f8b9a301e8d8c76/neuro-assessment-for-scalp-the-non-neuro-nurse-neuro-assessment-for-the-non-neuro.jpg)

















