
POPULATION DYNAMICS OF RHIZOBIUM JAPONICUM AND RHIZOBIUM … II.pdf · POPULATION DYNAMICS OF...
118
POPULATION DYNAMICS OF RHIZOBIUM JAPONICUM AND RHIZOBIUM LEGUMINOSARUM IN HOST AND NON-HOST RHIZOSPHERES A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN AGRONOMY AND SOIL SCIENCE DECEMBER 1982 By Robert Baines Woolfenden II Thesis Committee: B. Ben Bohlool, Chairman Mitiku Habte Jake Halliday
Transcript of POPULATION DYNAMICS OF RHIZOBIUM JAPONICUM AND RHIZOBIUM … II.pdf · POPULATION DYNAMICS OF...

POPULATION DYNAMICS OF RHIZOBIUM JAPONICUM
AND RHIZOBIUM LEGUMINOSARUM IN HOST
AND NON-HOST RHIZOSPHERES
A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN AGRONOMY AND SOIL SCIENCE
DECEMBER 1982
By
Robert Baines Woolfenden II
Thesis Committee:
B. Ben Bohlool, Chairman Mitiku Habte Jake Halliday

We certify that we have read this thesis and that in our opinion
it is satisfactory in scope and quality as a thesis for the degree of
Master of Science in Agronomy and Soil Science.
THESIS COMMITTEE
.. ^ ^ Chairman

ACKNOWLEDGEMENTS
I am very grateful to Dr. B. Ben Bohlool for his guidance,
constructive criticisms, enthusiasm, and patience throughout all
phases of this research.
In addition, I am grateful to fellow graduate students,
Mark Kingsley, Renee Kosslak, Michael Sadowsky, and Paul Singleton,
for helpful suggestions and encouragement as the experiments pro
gressed.
This work was supported in part by Grant AID/ta-C-1207 (NifTAL
Project) from the United States Agency for International Development.
The conclusions reached within this study do not necessarily represent
the views of the granting agency.

TABLE OF CONTENTS
Page
ACKNOWLEDGEMENTS......................................... iii
LIST OF TABLES ......................................... v
LIST OF F I G U R E S ......................................... vi
LIST OF DIAGRAMS......................................... viii
CHAPTER 1. INTRODUCTION AND LITERATURE SURVEY ............ 1
CHAPTER 2. POPULATIONS OF RHIZOBIUM IN THE RHIZOSPHERESOF HOST AND NON-HOST PLANTS AT HARVEST......... 17
CHAPTER 3. POPULATIONS OF RHIZOBIUM IN THE RHIZOSPHERES OF HOST AND NON-HOST PLANTS DURING A 35-DAY GROWTH CYCLE....... ........................... 47
CHAPTER 4. RHIZOBIUM POPULATION DYNAMICS IN HOST AND NON-HOST RHIZOSPHERES DURING THE FIRSTNINE DAYS OF RHIZOSPHERE DEVELOPMENT........... 85
iv
CHAPTER 5. GENERAL DISCUSSION 107

V
Page
TABLE
2-1 Comparison of the recovery of added Rhizobiumfrom Kula loam soil by two methods.................. 30
4-1 Percentages of Rhizobium inoculant strains(combined) in the total rhizosphere populationof bacteria at 3, 5, 7, and 9 days.................. 100
4-2 Percentages of Rhizobium inoculant strains(combined) in the total rhizosphere populationof bacteria at planting, 24, and 48 hours........... 101
LIST OF TABLES

vi
££
31
32
33
34
35
36
38
39
39
59
60
61
62
63
64
65
66
LIST OF FIGURES
Counts of 4 Rhizobium strains at harvest 1
Counts of 4 Rhizobium strains at harvest 2
Counts of 4 Rhizobium strains at harvest 3
Counts of 4 Rhizobium strains at harvest 4
Acridine orange total counts of bacteria at harvests 2, 3, and 4..................
Soybean nodule occupancy by Rhizobium japonicum strains........................
Pea nodule occupancy by Rhizobium leguminosarum strains....................
R. japonicum nodule bacteria, stained with homologous fluorescent antibody . . .
R. 1eguminosarum nodule bacteria, stained with homologous fluorescent antibody . . .
Counts of 4 Rhizobium strains per gram of soil at 1 week ..........................
Counts of 4 Rhizobium strains per gram of soil at 2 weeks.........................
Counts of 4 Rhizobium strains per gram of soil at 3 weeks.........................
Counts of 4 Rhizobium strains per gram of soil at 4 weeks..........................
Counts of 4 Rhizobium strains per gram of soil at 5 weeks.........................
Counts of 4 Rhizobium strains per square centimeter of root area at 1 week........
Counts of 4 Rhizobium strains per square centimeter of root area at 2 weeks . . . .
Counts of 4 Rhizobium strains per square centimeter of root area at 3 weeks . . .

Page
FIGURE
3-9 Counts of 4 Rhizobium strains per squarecentimeter of root area at 4 weeks................ 67
3-10 Counts of 4 Rhizobium strains per squarecentimeter of root area at 5 weeks................ 68
3-11 Acridine orange total counts of bacteria pergram of soil over 5 weeks........................ 69
3-12 Acridine orange total counts of bacteria persquare centimeter of root over 5 weeks. ........ 70
3-13 Soybean nodule occupancy by R. japonicumstrains......................................... 72
3-14 Large pea nodule occupancy by R. leguminosarumstrains......................................... 73
3-15 Small pea nodule occupancy by R. leguminosarumstrains....................... 74
3-16 View of the experiment at 1 week................ . 75
3-17 View of the experiment at 3 weeks................ 77
3-18 View of the experiment at 5 weeks................ 77
4-1 Counts of 4 Rhizobium strains at 3 days........... 92
4-2 Counts of 4 Rhizobium strains at 5 days........... 93
4-3 Counts of 4 Rhizobium strains at 7 days........... 94
4-4 Counts of 4 Rhizobium strains at 9 days ........... 95
4-5 Acridine orange total counts of bacteriaat 3, 5, 7, and 9 days............................ 96
4-6 Counts of 4 Rhizobium strains in the pearhizosphere at planting, 24, and 48 hours......... 97
4-7 Counts of 4 Rhizobium strains in the soybeanrhizosphere at planting, 24, and 48 hours......... 98
vii
LIST OF FIGURES (Continued)

viii
FIGURE
4-8
LIST OF FIGURES (Continued)
Page
Acridine orange total counts of bacteria in the pea and soybean rhizospheres at planting,24, and 48 hours................................... 99

ix
Page
DIAGRAM
3-1 Positioning of watering device, and setup ofexperimental pot for watering from below.......... 54
3-2 Arrangement of cores and order of harvest forthe 35-day time course study...................... 55
LIST OF DIAGRAMS

CHAPTER 1
INTRODUCTION AND LITERATURE SURVEY

INTRODUCTION AND LITERATURE SURVEY
The increasing disparity between population growth and food
production has placed a burden on agriculturalists to devise more
efficient methods of food production. Therefore, the Rhizobium-
legume symbiosis has received renewed attention lately, as it becomes
more and more apparent that we cannot count on cheap fossil fuels to
supply our needs for fixed nitrogen. There has been a renascent
interest in biological nitrogen fixation in general, and in the
Rhizobium-legume symbiosis in particular, as a means to help provide
protein to the world's rapidly-expanding population.
The symbiosis is a partnership between soil bacteria of the
genus Rhizobium and plants of the family Leguminosae. The visual
manifestations of the symbiosis are legume root nodules which house
the rhizobia. Inside these nodules atmospheric dinitrogen gas is
enzymatically reduced initially to ammonia, thereby entering the
assimilation pathways of plants.
The term "rhizosphere" was coined by Hiltner in 1904 as "the
region of contact between soil and root, where the soil is affected
by the root".
Before any nodules are formed, however, a series of intricate
events occurs in the rhizosphere. Rhizobia multiply in the host
rhizosphere, somehow recognize the plants' roots as those of a host
legume, infect those roots, and eventually initiate nodule formation.
Specificity is the hallmark of the Rhizobium-legume symbiosis; only
certain species of rhizobia are able to infect the roots of partic
ular legume species. Although the relatioship between the bacteria

and the host legume has been studied in great detail, as yet the
basis for this specificity is not clear.
A number of authors (Bohlool and Schmidt, 1974; Dazzo and
Hubbel, 1975) have implicated lectins (sugar-specific proteins or
glycoproteins on the legume root surface) as a possible basis for
specificity. However, due to the lack of standardized techniques
for examining lectin binding, the notion of lectin-mediated speci
ficity is still controversial (Broughton, 1978, Schmidt, 1979).
Another possible contributor to specificity is the stimulation
of the growth of rhizobia in the rhizosphere of their homologous
host plants, over and above that of the normal rhizosphere flora,
and over other non-homologous rhizobia (Nutman, 1963, 1965). This
specific stimulation in the rhizosphere has been postulated to be
mediated by root exudates. Selective stimulation of rhizobia by the
roots of their legume hosts would be of great selective advantage to
both, and presumably could be operating in concert with lectins to
confer specificity to the symbiosis.
The objective of this chapter is to give a concise review of
the pertinent literature on the effects of root exudates on rhizobia,
and to examine specific stimulation by those exudates as a possible
contributor to specificity in the symbiosis.
Plant roots in general stimulate gram negative rods more than
other soil bacteria, and these constitute greater proportions of the
rhizosphere than the soil populations in general (Rovira and
McDougall, 1967). Krasil'nikov (1958) demonstrated that bacteria
which colonize the rhizosphere of one plant species do not necessarily

colonize that of other plant species. He also showed that not all
strains of Pseudomonas fluorescens have equal rhizosphere colonizing
abilities. One of the most consistent differences between bacteria
isolated from the rhizosphere and those isolated from fallow soil is
the requirement of the former for amino acids (Lochhead and Rouatt,
1955).
Starkey (1929) suggested the following factors to be involved in
the stimulation of microorganisms by plant roots; sloughed-off root
cells, moribund root hair and cortical cells, and soluble organic
nutrients released or actively exuded by intact roots. Much evidence
has been presented demonstrating the exudation of many organic materials
from healthy, intact plant roots (Scroth and Hildebrand, 1964; Rovira,
1962; 1965). These studies support the notion that sufficient
nutrients are exuded to support large rhizosphere populations of
microorganisms.
The legume rhizosphere has shown to be a particularly intense
zone of microbial activity, due to the nature and quantity of products
exuded by legume roots (Rovira, 1956a; 1962). According to the theory
of specific stimulation, legume exudates should stimulate rhizobia
able to infect them more than other rhizobia. This theory was detailed
by Nutman (1965).
Most of the early studies were conducted in aseptic solution,
agar, or sand culture, using one or a few strains of Rhizobium. The
following is a chronological review of some of the early studies.
Nutman (1953) grew clover, alfalfa, and vetch, singly and in
pairs consisting of either one or two plant species. Plants were
4

cultured aseptically in tubes on agar "slopes". In these experiments
the presence of an alfalfa plant in the same tube with a clover plant
and R. trifolii, plants were nodulated sooner than in tubes in which
there were two clover plants. Apparently, exudates from the alfalfa
roots were able to stimulate the nodulation of clover. Exudates
from inoculated clover plants uniformly inhibited the nodulation of
another clover, lucerne, or vetch plant in the same tube. This
inhibition was least in clover and greatest in vetch. Inhibition of
nodule formation on clover was obtained in tubes which had been
preplanted with clover and the earlier plantings removed. Nutman
interprets his results in terms of the secretion of some nodule-
inhibiting substance from the clover roots.
Several of the early examinations of plant root exudates in the
rhizosphere were carried out by Rovira (1956a, 1956b, 1961; Rovira
and Harris, 1961). In many of his studies, plants were cultured in
sterile sand, -instead of the sterile water or agar culture used by
several workers previously. Following growth, plants were carefully
removed, and exudates leached out of the sand. Rovira (1956a)
demonstrated the exudation of a host of amino compounds and sugars
from aseptically-grown oat and pea plants. Exudates from pea roots
were larger in quantity, and compounds were more numerous and varied
than those from the roots of oats. Early studies had been complicated
by the fact that only a small amount of rhizosphere soil was available
for study. To get around this problem, Rovira (1956b) established an
"artificial rhizosphere" in which non-sterile field soils were
saturated with root exudate solution collected from aseptically-grown
5

roots. Treatment was continued for 21 days, and an overall
stimulation of bacterial growth upon the addition of the exudates,
the bacteria consisting largely of gram negative rods, was taken as
evidence that his artificial rhizosphere had actually been successful.
The resulting populations of root-exudate-treated soils did not
increase the rate of organic matter decomposition in the soil.
However, the artificial rhizosphere population did promote the
decomposition of the more readily-available organic nutrients, such
as amino acids and glucose. Rovira (1961) compared the rhizospheres
of red clover and paspalum (a grass) with respect to numbers of R.
trifolii and total bacteria. In this study, non-sterile field soil
was used, and Rhizobium numbers were determined by the most probable
number (MPN) technique (Vincent and Waters, 1954). Bacterial numbers
were examined in response to the rhizospheres of the two plant
species and also to varying lime levels. Bacteria, including
rhizobia, were consistently present in larger numbers in the rhizo
sphere of clover than that of paspalum; ratios for Rhizobium were
about 5:1 (clover: paspalum) and for total bacteria, about 2:1.
Liming was beneficial to the rhizosphere populations of both plants.
The liming probably affected the growth of the microorganisms
directly and also indirectly, by first enhancing plant growth and
root exudation. In a further study, Rovira and Harris (1961)
examined the exudation of growth factors from peas, alfalfa, tomato,
and several clover species grown in sterile sand culture. Their aim
was to quantitatively assess the various B-group vitamins exuded by
these plant species. Biotin was found to be exuded in the largest
6

quantity, and was found in the rhizosphere of pea at 10 to 100 times
the concentration found in clover or tomato rhizospheres. Other
growth factors, notably pantothenate and niacin, were present, but in
amounts considered by the authors as unlikely to influence the growth
of microorganisms. In non-sterile sand culture, biotin and panto
thenate were seen to disappear rapidly, emphasizing the need for
strict asepsis in any studies of root exudates.
Studies by Rovira and coworkers emphasized the large variety and
quantity of substrates exuded by plant roots, particularly those of
legumes. However, most of these studies focused on the exudates
themselves rather than the particular bacteria stimulated by those
exudates. Because of this fact, most studies were carried out under
aseptic conditions in sand culture.
Elkan (1961) examined a non-nodulating, near isogenic soybean
variety to determine the reason for non-nodulation. In greenhouse
solution culture, he demonstrated that the root excretions from the
mutant non-nodulating line resulted in highly significant (p=.01)
decreases in nodulation of normal, nodulating plants. In addition,
the excretion resulted in decreased total nodule weight, total dry
weight, and total nitrogen per nodulating plant. Curiously, the
excretion did not inhibit growth of R. japonicum directly, nor did
it inhibit nodulation of other plant species by other rhizobia.
Tuzimura and Watanabe (1962a) examined the populations of
bacteria, fungi, actinomycetes, and specifically Agrobacterium
radiobacter and a Rhizobium spp. from Astragalus sinicus in the
rhizosphere of this plant. A non-sterile volcanic ash soil was used,
7

and rhizobia were enumerated by an MPN method. The astragalus plants
were able to support a reasonable population of astragalus rhizobia,
about 8.1 x 10** at flowering, and 10® per gram dry root at fruiting,
regardless of starting Rhizobium population. In a further study,
Tuzimura and Watanabe (1962b) followed populations of R. trifolii in
the rhizospheres of several leguminous plants, as well as the rhizo
spheres of non-legumes. Ladino clover, alfalfa, soybean, and peanut
were the legumes examined. The non-legumes were sudan grass and
upland rice. Rhizobium trifolii were consistently seen in larger
numbers in the rhizosphere of alfalfa, followed by peanut, followed
by soybeans, and, followed by clover. No statistical treatment of
the data was presented, and the authors state that "these findings
might not be necessarily valid, because the growth of the plants and
contents of soils which adhered to the root varied in each case."
Rhizosphere numbers of R. trifolii were, without exception, at least
two logs greater in the rhizosphere of legumes than in non-legume
rhizospheres.
Nutman (1963, 1965) suggested that "A given legume tends to
promote the multiplication of bacteria able to infect it more than
others", and, "Individual strains of nodule bacteria are more
strongly stimulated by those hosts they are able to infect than by
other legumes", citing only a reference by Wilson (1930) in support
of these statements. Dart and Mercer (1964) present the opposite
viewpoint. They state that there is no evidence that legume roots
selectively stimulate the growth of Rhizobium rather than other
organisms. Further, they state that the Rhizobium strains which
8

nodulate a particular legume are not preferentially stimulated in that
hosts rhizosphere over other Rhizobium strains, citing Krasil'nikov
(1958) and Purchase (private communication).
Rovira (1965) concluded that sufficient chromatographic
analyses had been performed on root exudates to indicate the wide
spectrum of compounds contained therein.
Peters and Alexander (1966) examined four different rhizobia in
the rhizosphere of alfalfa and Lotus corniculatus in aseptic solution
culture. Alfalfa was inoculated with R.. meliloti, R. trifolii, and
R. leguminosarum to see if alfalfa stimulated only its homologous
Rhizobium, R. meliloti. Despite differences in initial inoculum
size and in spite of the fact that R. meliloti alone induced
nodulation, populations of all rhizobia reached roughly 106 to
107 cells/ml of rooting medium after 1 week, and following that cell
numbers did not fall rapidly. These results, albeit in aseptic
solution culture, were not suggestive of specific stimulation. Also,
when alfalfa and Lotus corniculatus were inoculated with a mixed
culture of their respective microsymbionts, no selective interaction
between host and homologous organisms was seen. Peters and Alexander
(1965) suggested that the selectivity between microbe and host is
probably exerted first at individual receptor sites on host root
surfaces.
Van Egeraat (1975a) examined the growth of R. leguminosarum on
the rhizoplane (root surface) and in the rhizosphere of aseptically
growth pea seedlings, using sterile agar culture in petri plates.
He observed no bacterial growth on the main taproot, however, after
9

the emergence of secondary roots bacterial numbers increased where
the secondary roots emerged from the main taproot. Lateral roots of
the pea seedlings growing in sterile agar along the bottom of the
petri dish had a zone of bacterial growth some distance from the
roots. Van Egeraat (1975a) attributed this to a zone of growth
inhibition surrounding roots more closely. Of particular interest
in this study was the stimulation of rhizobia around the sites of
emergence ("wounds") of the lateral roots. Van Egeraat concludes
that young pea plants exude both growth stimulating and growth-
inhibiting compounds. Further, the growth-inhibiting compounds can
temporarily prevent the growth of R. leguminosarum in the immediate
vicinity of the pea roots. In a follow-up study, Van Egeraat (1975b)
demonstrated that R. 1eguminosarum grew equally well with homoserine
and glutamic acid as the nitrogen source, or as the sole source of
carbon, nitrogen, and energy. Strains of R. trifolii, R. phaseoli,
and R. meliloti behaved entirely differently. These three strains
could grow with glutamate as the only C and N source. With homo
serine, growth was extremely slow or absent, or in the case of R.
meliloti, considerably reduced. Van Egeraat suggested that homo
serine (which was found to comprise about 70% of the amino compounds
exuded by pea roots) might selectively stimulate the growth of R.
leguminosarum in the pea rhizosphere when a mixture of Rhizobium of
many species is present. This represents perhaps the most compelling
study in favor of specific stimulation. However, it was carried out
under totally aseptic conditions, and on the basis of these experi
ments it would be impossible to predict the fate of homoserine in the
10

rhizosphere of pea plants grown in the field. Homoserine is the first
compound to be suggested as a specific stimulant in the scientific
literature. Rovira (1965) stated that it would appear unlikely that
the the ubiquitous sugars and amino acids would provide the speci
ficity observed in the symbiosis, but rather the balance of these
compounds of the presence of exotic compounds peculiar to a particular
plant species. Van Egeraat (1975b) states that in the pea system,
just such a compound, homoserine, is present.
The studies on root exudates and the rhizosphere effect with
respect to rhizobia have either addressed the interaction in aseptic
systems, from which organisms can be conveniently plated and counted,
or have enumerated rhizobia indirectly by means of MPN counts (in the
case of non-sterile systems). However, either method has its share
of disadvantages, and thus early studies of individual species or
strains of Rhizobium in the rhizosphere of their homologous host
plants are not without criticism. What was needed was an adequate
methodology for strain-specific enumeration of rhizobia in natural
non-sterile rhizospheres, in which the full range of microbe-to-microbe
and plant-to-microbe interactions are taking place.
An autoecological approach, in which the numbers of several
different Rhizobium strains in the rhizospheres of several host
plants could be studied, is necessary to adequately address the
problem. The only method adequate for the study of a specific micro
organism directly in a natural soil environment is immunofluorescence
(Schmidt, 1979; Bohlool and Schmidt, 1980).
11

Reyes and Schmidt (1979) used membrane filter immunofluorescence
(Schmidt, 1974) to enumerate R. japonicum strain 123 in soil and in
rhizospheres of Minnesota field-grown soybeans. Rhizosphere effects
were modest. About 101* to 105 cells/gram of soil were observed in
soil adhering to plant roots. A comparably slight rhizosphere effect
was seen for corn. According to their data, strain 123 did not
follow what they termed the "scenario" of specific stimulation.
Further, these authors state that experimental support for the
specific stimulation hypothesis is meager, especially in terms of
rhizobial response to plant rhizospheres under natural soil conditions.
In a further study, Reyes and Schmidt (1981) used membrane filter
immunofluorescence for enumeration of, and immunofluorescence
examination of strain USDA 123 on root surfaces of field-grown
soybeans. Strain 123 was a consistent rhizosphere colonizer, but
failed to multiply as rapidly as the root system developed under
field conditions. Root surface populations declined from about
7 to 8 x 102 per square cm of root surface on day 9 to about 60 per
square cm on day 26. Average calculated cell density per square cm
was only a few hundred. They also examined the behavior of strain 123
in the rhizospheres of pot-grown soybeans. In this trial, cell
numbers and root surface populations were monitored every 4 days,
until nodule initiation. This greenhouse experiment confirmed the
findings of the field experiment. Strain 123 was seen in numbers of
a few hundred per square cm of root surface during early growth, and
declined to less than 100 per square cm with more extensive root
development. Also addition of a competing strain (USDA 138) had
12

little effect on the development of strain 123 in the soybean
rhizosphere. Even when 10 times as many 138 cells than 123 cells
were added, cell densities of both stabilized at a few hundred per
square cm of root surface, and each appeared to establish indepen
dently of the other. Immunofluorescence examination of unwashed
roots revealed that both strains were sparsely distributed on root
surfaces. Microscopic examination of the roots also revealed that
rhizobia were seldom seen in microcolonies, but rather as single,
double, or triple cells. In this study, no proliferation of
Rhizobium was observed, even at the junction of lateral and tap roots
where release of organic compounds is likely to occur due to the
wounds caused by the emergence of secondary roots (Van Egeraat,
1975a). No evidence was obtained in support of specific stimulation
as a prelude to nodulation. Strain 123 established in numbers
roughly equal to those of 138, even though 123 forms the vast
majority of the nodules. Performing rhizosphere counts on the basis
of root surface area makes the lack of specific stimulation in the
rhizosphere even more clear cut. Competitive advantages of
strain 123 over strain 138 were not obvious, however. These studies
by Reyes and Schmidt represent an important first step in examining
rhizobia in natural non-sterile field soil rhizospheres.
Exudates are important in establishing and maintaining the
rhizosphere population of microorganisms; bacteria in particular
seem to respond to the soil conditions around plant roots. In
aseptic systems, the interactions between exudates and bacteria
(particularly Rhizobium) have been studied by many investigators.
13

Results in sterile systems have been somewhat ambiguous. In
autoecological studies done under natural field soil conditions, no
evidence has to date been found in support of specific stimulation.
However, no study has examined several different species of rhizobia
in the rhizospheres of both homologous and non-homologous legumes.
An appropriately-designed autoecological study such as this could
adequately address the question of specific stimulation in the legume
rhizosphere.
The basis for specificity in the Rhizobium-legume symbiosis is
still a matter of debate. A better knowledge of this basis might
ultimately help to extend nodulation and nitrogen fixation capability
beyond the legumes to other grain crops, grasses, and cereals. Self-
sufficiency for nitrogen would be a highly desirable trait for any
crop plant.
14

BIBLIOGRAPHY
1. Bohlool, B. B. and E. L. Schmidt. 1974. Lectins: a possiblebasis for specificity in the Rhizobium-legume symbiosis.Science 185:269-271.
2. Bohlool, B. B. and E. L. Schmidt. 1980. The immunofluorescenceapproach in microbial ecology, pp. 203-241. In Advances in microbial ecology, Vol. 4. Plenum Publishing Corp., New York.
3. Broughton, W. J. 1978. Control of specificity in legume- Rhizobium interactions. J. Appl. Bact. 45:165-194.
4. Dart, P. J. and F. V. Mercer. 1964. The legume rhizosphere.Archiv fur Mikrobiologie 47:344-378.
5. Dazzo, F. B. and D. H. Hubbel. 1975. Cross-reactive antigens and lectins as determinants of symbiotic specificity in the Rhizobium-clover association. Appl. Microbiol. 30:1017-1033.
6. Van Egeraat, A. W. S. M. 1975a. The growth of Rhizobium leguminosarum on the root surface and in the rhizosphere of pea seedlings in relation to root exudates. Plant and Soil 42:367-379.
7. Van Egeraat, A. W. S. M. 1975b. The possible role of homoserinein the development of Rhizobium 1eguminosarum in the rhizosphere of pea seedlings. Plant and Soil 42:381-386.
8. Elkan, G. 1961. A nodulation-inhibiting root excretion from a non-nodulating soybean strain. Can. J. Microbiol. 7:851-856.
9. Krasil'nikov, N. A. 1958. Soil microorganisms and higher plants. Moscow Academy of Sciences, USSR. English edition. National Science Foundation.
10. Law, I. J. and B. W. Strijdom. 1977. Some observations on plant lectins and Rhizobium specificity. Soil Biol. Biochem. 9:79-84.
11. Lochhead, A. G. and J. W. Rouatt. 1955. The "rhizosphere effect" on the nutritional groups of soil bacteria. Soil Sci.Soc. Am. Proc. 19:48-49.
12. Nutman, P. S. 1953. Studies on the physiology of nodule formation. IV. The mutual inhibitory effects on nodule production of plants grown in association. Ann. Botany (London) 17:95-126.
13. Nutman, P. S. 1963. Factors influencing the balance of mutual advantage in legume symbiosis, pp. 51-71. In P. S. Nutman and B. Mosse (Eds.) Symbiotic Associations. Cambridge Univ. Press, Cambridge, England.

16
14. Nutman, P. S. 1965. The relation between nodule bacteria and thelegume host in the rhizosphere and in the process of infection,pp. 231-247. In K. F. Baker and W. C. Snyder (Eds.) Ecology of soil-borne plant pathogens - prelude to biological control.Univ. Calif. Press, Berkeley.
15. Peters, R. J. and M. Alexander. 1966. Effect of legume rootexudates on the root nodule bacteria. Soil Sci. 102:380-387.
16. Reyes, V. G. and E. L. Schmidt. 1979. Population densities ofRhizobium japonicum strain 123 estimated directly in soil and rhizospheres. App. Environ. Microbiol. 37:854-858.
17. Reyes, V. G. and E. L. Schmidt. 1981. Populations of Rhizobiumjaponicum associated with the surfaces of soil-grown roots. Plant and Soil 61:71-80.
18. Rovira, A. D. 1956a. Plant root excretions in relation to therhizosphere effect. I. The nature of root exudates from oats and peas. Plant and Soil 7:178-194.
19. Rovira, A. D. 1956b. Plant root excretions in relation to therhizosphere effect. III. The effect of root nodule exudate on numbers and activity of microorganisms in soil. Plant and Soil 7:209-217.
20. Rovira, A. D. 1961. Rhizobium numbers in the rhizospheres ofred clover and paspalum in relation to soil treatment and numbers of bacteria and fungi. Aust. J. Agric. Res. 12:77-83.
21. Rovira, A. D. and J. R. Harris. 1961. Plant root excretions in relation to the rhizosphere effect. V. The exudation of B-group vitamins. Plant and Soil 14:119-214.
22. Rovira, A. D. 1962. Plant root exudates in relation to therhizosphere microflora. Soils Fertilizers 25:167-172.
23. Rovira, A. D. 1965. Plant root exudates and their influenceupon soil microorganisms, pp. 170-186. In K. F. Baker and W. C. Snyder (Eds.) Ecology of soil-borne plant pathogens - prelude to biological control. Univ. Calif. Press, Berkeley.
24. Rovira, A. D. and B. M. McDougall. 1967. Microbiological and biochemical aspects of the rhizosphere. pp. 417-463. InA. D. McLaren and G. F. Peterson (Eds.) Soil biochemistry. Marcel Dekker, New York.
25. Schmidt, E. L. 1974. Quantitative autoecological study of microorganisms in soil by immunofluorescence. Soil Sci. 118: 141-149.

17
26. Schmidt, E. L. 1979. Initiation of plant root-microbeinteractions. Ann. Rev. Microbiol. 33:355-376.
27. Scroth, M. N. and D. C. Hildebrand. 1964. Influence of plant exudates on root-infecting fungi. Ann. Rev. Phytopathology 2: 101-132.
28. Starkey, R. L. 1958. Interrelations between microorganisms andplant roots in the rhizosphere. Bact. Rev. 22:154-168.
29. Tuzimura, K. and I. Watanabe. 1962a. The growth of Rhizobium in the rhizosphere of the host plant. Ecological studies of root nodule bacteria (part 2). Soil Sci. Plant Nutri. (Tokyo) 8:19-24.
30. Tuzimura, K. and I. Watanabe. 1962b. The effect of various plants on the growth of Rhizobium. Ecological studies of root nodule bacteria (part 3). Soil Sci. Plant Nutri. (Tokyo) 8:13-17.
31. Vincent, J. M. and L. M. Waters. 1954. The root nodule bacteriaas factors in clover establishment in the red basaltic soils of the Lismore district, New South Wales. II. Survival and success of inocula in laboratory trials. Aust. J. Agric. Res. 5:61-76.
32. Wilson, J. K. 1930. Seasonal variation in the numbers of two species of Rhizobium in soil. Soil Sci. 30:289-296.

POPULATIONS OF RHIZOBIUM IN THE RHIZOSPHERES
OF HOST AND NON-HOST PLANTS AT HARVEST
CHAPTER 2

ABSTRACT
The growth of two strains of R. japonicum (strains USDA 110 and
CB 1809), and two strains of R. leguminosarum (strains Hawaii 5-0 and
Nitragin 92A3) was followed in the rhizospheres of soybean, pea, and
corn growing in non-sterile soil. Rhizosphere soil was sampled at
35 days over four successive growth cycles. The numbers of each strain
were determined by membrane filter immunofluorescence, using strain-
specific fluorescent antibodies. Nodule occupancy of the strains on
their appropriate host was determined by immunofluorescence. No
specific stimulation of rhizobia in the rhizospheres of their homologous
host plants was observed. Counts of all strains, as well as total
bacteria were generally in the following order: soybean rhizosphere>
pea>corn>fallow soil. Strain CB 1809 occupied a slightly higher
percentage of soybean nodules than USDA 110, whereas Nitragin 92A3
dominated Hawaii 5-0 in pea nodules. Fifteen percent of soybean
nodules were doubly infected, as were 3.5% of pea nodules. The four
strains together comprised 1.5 to 2.6% of the total rhizosphere
bacteria of the legumes and 2 to 9% of the total bacteria in the corn
rhizosphere. Rhizobia comprised 3 to 5% of the total bacteria in
fallow soil. These data are not suggestive of an overwhelming increase
of homologous rhizobia in the rhizospheres of their respective host
legumes. Specific stimulation of growth of Rhizobium in the legume
rhizosphere does not appear to be a contributor to specificity in the
Rhizobium-legume symbiosis under the conditions of this study.

INTRODUCTION
Soil bacteria of the genus Rhizobium are recognized by their
ability to form nitrogen-fixing nodules on the roots of many leguminous
plants. Rhizobia are specific with respect to the hosts they nodulate.
Only certain species of rhizobia are able to infect and nodulate the
roots of particular legumes. Although the relationship between rhizobia
and their host legumes has been studied in great detail, the basis for
this specificity is not completely clear (5,20).
Several possible mechanisms have been advanced to account for
specificity, involving both plant and bacterial components (5).
Bacterial chemotaxis might affect rhizosphere populations, as might
rhizosphere competence once rhizobia have arrived in the root zone
(21). Cellular recognition between host and bacterium is considered
by some as a mechanism for specificity. Plant proteins called lectins
are thought to interact specifically with rhizobia at the root surface
(3,9,22). Another proposed mechanism for specificity is the prefer
ential growth of the appropriate rhizobia in the rhizospheres of their
homologous host plants over and above that of other rhizobia (7,14,25).
Most studies of stimulation in the legume rhizosphere have been
carried out in aseptic systems, from which rhizobia can be conveniently
plated and counted (8,13,15). However extrapolation from aseptic
conditions to non-sterile field soil, in which the full range of
microbial interactions would be occurring, is unrealistic, and often
erroneous.
An indirect method of enumerating rhizobia in non-sterile soil is
the plant dilution assay (24). In this method, the plant itself is

used as a selective agent, and rhizobia enumerated by dilution-
extinction. This method is cumbersome, however, and does not lend
itself to ecological studies.
Direct study of specific bacterial strains in soil and rhizo
spheres has been prevented by the lack of an adequate methodology for
enumeration of bacteria in the particulate soil environment (4).
Bohlool and Schmidt (2) and Schmidt (19) solved many of the method
ological problems with the introduction of a quantitative membrane
filter immunofluorescence (MFIF) technique for enumeration of specific
microorganisms directly in soil. This work was extended by Kingsley
and Bohlool (11) who adapted the MFIF technique for use in tropical
soils. In the present report, MFIF is used to examine the population
dynamics of homologous and non-homologous rhizobia in the rhizospheres
of host and non-host plants.
21

MATERIALS AND METHODS
Preliminary Soil Examination
Kula loam soil, an Inceptisol (Typic Eutrandept, pH 6.5), was
chosen as the experimental soil. The soil was tested for the presence
of cross-reactive bacterial cells, fungal spores, and mycelia by the
membrane filter immunofluorescence (MFIE) method of Kingsley and
Bohlool (11). Kula soil was also tested for the presence of
indigenous R. japonicum and R. leguminosarum by inoculating soybean
and pea seedlings grown in flasks of sterile vermiculite. Seeds were
surface sterilized by shaking in a 4% solution of sodium hypochlorite
for 20 minutes followed by at least five rinses in sterile water, and
were germinated on .9% water agar. One gram of Kula soil was added
directly to the radicles of aseptically grown 2- to 3-day-old soybean
or pea seedlings. Plants were grown in a growth chamber for 28 days,
and roots checked for nodulation.
Rhizobium Strains
R. japonicum strains CB 1809 and USDA 110 and R. leguminosarum
strains Hawaii 5-0 and Nitragin 92A3 were obtained from the collection
of B. B. Bohlool of the University of Hawaii. Strains were maintained
on yeast-extract mannitol agar slants (24), with 1.0 g of yeast extract
(Difco Laboratories, Detroit, Michigan) substituted for yeast-water.
Experiments to Assess Recovery
Kula soil was inoculated with known numbers of rhizobia, and their
recovery assessed by the method of Kingsley and Bohlool (11), using
appropriate FAs.

Inoculum Preparation
Yeast-extract mannitol broth (24) cultures of each of the four
strains were grown to early stationary phase. Kula loam soil (250 g)
was amended to 1% with mannitol, adjusted to about -1/3 bar moisture
tension with distilled water, and autoclaved (121 C, 15 psi) for
45 minutes on two successive days. Ten ml aliquots of the above broth
cultures were then added aseptically, and these soil cultures incubated
at room temperature for 3 days (R. leguminosarum) or 8 days (R.
japonicum). MFIF counts were performed on these soil cultures and
aliquots of each added to 1 kg of non-sterile Kula soil to a level of
about 5 x 105 cells of each strain, per gram of moist soil. This
mixture was homogenized by manual shaking in a large plastic bag for
10 minutes. MFIF counts were performed on this intermediate dilution
of inoculant. The intermediate dilution was mixed with an additional
4 kg of non-sterile soil to give about 1 x 10s cells per gram (moist
soil) of each strain. Final mixing was done in a twin-shell dry
blender (The Patterson-Kelley Co., Inc., East Stroudsburg, Pa.) for
10 minutes.
Pots and Planting
Pots used were 1400 ml polypropylene enema containers (Resiflex,
hospital surplus) painted with a heavy coat of flat white paint. Each
pot contained 750 grams of non-sterile inoculated Kula soil (moist
weight). Soils were brought to about -1/3 bars of tension with
distilled water. Three-day-old seedlings of peas (wilt-resistant
Wisconsin Perfection), soybeans (Davis), and corn (Hawaiian Supersweet
23

#9) were planted. Seeds were pregerminated aseptically on .9% water
agar for 3 days. Three corn plants, four soybean plants, and five pea
plants were sown into their respective pots. Pots were planted in
triplicate. Three pots were set up as fallow controls (non-rhizosphere
soil). Following planting, the soil surface was covered with rinsed
white aquarium gravel (California Wonder Rock, Kordon Co., Hayward,
Ca.) to prevent undue soil heating and to inhibit the growth of algae
on the soil surface. The experiment was set up as a randomized
complete block.
Plant Growth Conditions
Harvest cycles were of approximately 35 days. The experiment
was not run longer because the release of bacteroids at nodule
senescence would certainly give the appearance of rhizosphere stimulation
of specific rhizobia (23) . Soil moisture tension was maintained at
about -1/3 bars by daily watering to a constant weight with distilled
water. Hoagland's N-free medium (10) was substituted for water once a
week. For the first harvest, plants were grown in a climate controlled
plexiglas house, onto which was attached a large air conditioner to
maintain temperatures of 27°C during the day and 21 C at night.
Subsequent cropping cycles were carried out in a greenhouse, where
temperatures were slightly higher. Four cropping cycles were performed.
Following a harvest, soil from the preceding harvest was placed back
into the same pots without further amendment, and the pots replanted
with the same crop.
24

Harvest and Sample Preparation
At harvest, the entire contents of each pot was carefully removed
and the loosely adhering soil gently shaken free of the root system.
Any large adhering soil clumps were also removed. Root systems with
adhering soil were then placed in 18-ounce Whirlpak bags for transport
to the laboratory. The root systems with rhizosphere soil (including
that which rubbed off the roots into the bags) were transferred to
square, wide mouth screwcap bottles (45x45x120 mm, approximate
capacity 230 ml). To these root and rhizosphere soil samples was
added 100 ml of .1% partially hydrolyzed gelatin (diluted in .1 M
dibasic ammonium phosphate, 11). Four drops of tween 80 (Sigma) were
added, and the bottles tightly capped and shaken on a Burrel Wrist-
Action shaker for 30 minutes at full power. Immediately after shaking,
25 ml of suspension was removed from each bottle and transferred to a
50 ml polycarbonate centrifuge tube. Samples were centrifuged gently
(700 x g, 5 min.) in a Sorvall centrifuge, to pellet soil particles.
Following centrifugation, supernatants were decanted into clean
screwcap tubes. The soil pellet, as well as the remaining soil and
soil solution was then rinsed into a pre-weighed aluminum tart pan for
soil dry weight determination. Pans were held at 105°C for 48 hours,
and weighed immediately after having cooled to room temperature. The
weight of the dried gelatin-ammonium phosphate preparation was deter
mined experimentally, and this value subtracted from soil weights.
Non-rhizosphere soil was examined using a modification of the
Kingsley/Bohlool MFIF procedure (11). Ten grams of non-rhizosphere
soil was placed in a 250 ml screwcap Erlenmeyer flask, and 1/3 of a
25

standard scintillation vial full of 3 mm diameter glass beads added.
To this was added 10 ml of 10% hydrogen peroxide (E. L. Schmidt,
personal communication). This mixture was shaken on a Burrel Wrist-
Action shaker for 10 minutes at full power. Flasks were capped some
what loosely, to prevent explosion. Following this step, 75 ml of
gelatin-ammonium phosphate (11) was added, and the mixture shaken for
an additional 20 minutes. Twenty-five ml of soil suspension was
removed from the flasks, transferred to 50 ml polycarbonate centrifuge
tubes, and centrifuged as above. Rhizosphere and non-rhizosphere
samples were diluted 1:10 and 1:2, respectively, before filtration for
FA counting.
Membrane Filter Immunofluorescence
Membrane filter counts were performed as in Kingsley and
Bohlool (11). Following filtration of the sample, filters were placed
on microscope slides, and the effective filtering area covered with
six drops of partially hydrolyzed gelatin-rhodamine isothiocyanate
conjugate (1). This served to reduce non-specific adsorption of
fluorescent antibody to soil colloids, and control background
fluorescence. RhITC-gelatin treated filters were then dried at 50°C
and held at that temperature until staining with the fluorescent
antibody. Filters were stained for 1 hour. Following staining,
filters were placed back on filter holders and immediately rinsed
with at least 150 ml of prefiltered .85% saline, then returned to
microscope slides, mounted with a coverslip in buffered glycerol
(pH 9), and observed. Microscopy was performed with a Zeiss standard
microscope 14 equipped with incident light illumination from an HBO 50
26

(Osram) mercury vapor light source and a Zeiss fluorescein
isothiocyanate (FITC) filter pack. A Zeiss 63X Planapo oil immersion
objective was used. Duplicate filters were counted for each strain
from each pot, for a total of 96 filters per harvest. No correction
factor was used to account for the fact that recoveries are generally
about 70 to 95%.
Acridine Orange Total Counts
Estimations of the total rhizosphere population of bacteria were
made at each harvest using acridine orange. Dilutions of centrifuge
supernatants were made in water which was collected directly from a
glass still into an acid-washed flask. Samples were concentrated onto
membrane filters, and filters transferred to microscope slides.
Filters were stained directly on the slides with one drop of acridine
orange solution (Sigma, 1:30 000 in .1 M phosphate buffered saline
pH 7.2) previously filtered through a .2 micron membrane filter
(Gelman Acrodisc, Gelman, Ann Arbor, Mich.). Three filters were
counted per pot, for a total of 36 per harvest.
Nodule Typing
Following the extraction of the rhizosphere soil from roots, all
nodules were removed, squashed with forceps, and smeared on slides
for serological typing with fluorescent antibodies (18). Microscopy
was performed with incident light fluorescence illumination in
conjunction with transmitted-light phase contrast lighting.
27

Immunofluorescent Examination of Root Surfaces
Root surfaces were examined by immunofluorescence (6) to insure
that rhizobia bound to root surfaces were being released into suspension
for counting.
Photographs (Figs. 7 and 8) were made with Ektachrome 200 slide
film, and prints made from these slides.
28

RESULTS
The Kula soil was found to be a good experimental soil for the
application of immunofluorescence to the study of ecology of Rhizobium.
Recoveries of added inoculant rhizobia were consistently in the range
of 70 to 95% (Table 1). The hydrogen peroxide pre-treatment did not
change recovery of rhizobia significantly (Table 1) but reduced back
ground fluoresence, making rhizobia easier to count from non-rhizo
sphere soil. Cross-reactive bacteria were not a problem, and
cross-reactive fungal spores and mycelia were readily distinguishable
by their morphology. The soil contained no indigenous R. japonicum
capable of nodulating Davis soybeans. Nodules were observed on peas
tested with Kula soil, although very infrequently. These nodules were
small and white throughout, and bacteroids contained therein did not
react with FAs prepared for the strains used in this study.
Immunofluorescent examination of root surfaces indicated that the
majority of Rhizobium cells bound to roots were released into suspen
sion for counting.
Rhizosphere numbers of each of the four Rhizobium strains are
shown within each harvest across the three rhizospheres and fallow
soil (Figs. 1 to 4). Each point represents the mean of six enumerations
(duplicate filters from each of three replicate pots). Acridine orange
total counts for harvests 2 to 4 are presented in Figure 5. Each point
represents the mean of nine enumerations (triplicate filters from each
of three replicate pots).
Soybean nodules contained either of the two R. j aponicum strains
or both as illustrated in Figure 6. Pea nodules contained either of

30
Table 1. Recovery of added Rhizobium cells by the Kingsley and Bohlool method (11) and by a modification thereof*
Recovery method and strain
Cells recovered per gram of soil
Percentrecovery
Kingsley and Bohlool with flocculation of soil particles
USDA 110 4.5 X 1 0 6 70USDA 110 4.7 X 1 0 6 73USDA 110 4.7 X 1 0 6 73
Hawaii 5-0 4.2 X 106 77Hawaii 5-0 5.2 X 1 0 6 95Hawaii 5-0 5.0 X 1 0 6 92
Modified Kingsley and Bohlool with H2O2 pretreatment and centrifugation of soil particles
USDA 110 5.2 X 106 81USDA 110 5.1 X 106 80USDA 110 4.5 X 106 70
Hawaii 5-0 4.1 X 106 74Hawaii 5-0 4.4 X 106 81Hawaii 5-0 4.6 X 106 84
. *Average starting numbers of cells were USDA 110 6.4 x 106 and Hawaii 5-0 5.5 x 106. Recoveries were not significantly different (p=.05) by a Wilcoxon's signed rank test.

31
L.S.D, .05
FIG. 1 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 1

32
L.S.D. .05
STRAIN
USDA 110
R. japonicum
CB 1809
N1TAAB1N 92A3
R. leguminosarum
HAHA1I 5-0
5.2THTBmr TEH CHHR
CHOPFHEcmr
FIG. 2 COUNTS OF 4 RHIZOBIUM STRAINS (X 10*) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 2

33
L • S . D . .05
HITRACIN 9203
R. leguminosarum
HflHflll 5-0
.05
18.0SOYBEAN
CROP
FIG. 3 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 3

34
L • S - D • 05
NITRA61N 32R3
R. leguminosarum
HflHfllI 5-0
10.6
.05
FIG. 4 COUNTS OF 4 RHIZOBIUM STRAINS (X ltf) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 4

35
L .S .D. 05
HARVEST
FIG. 5 ACRIDINE ORANGE COUNTS OF BACTERIA IN 3 RHIZOPSHERES AND IN FALLOW SOIL (X 108)
PER GRAM OF OVEN DRY SOIL

36
HARVEST
10 20 30 40 50 80 70 80 90 tOOPERCENT
LEGENDi STRAINS SSR3S3 CB 1809 KSSSa MIXED V/////A USDA 110
FIG. 6 SOYBEAN NODULE OCCUPANCY BY RHIZOBIUM JAPONICUM STRAINS

the two inoculum R. leguminosarum strains or both as given in Figure 7.
Nodules were distributed throughout the root systems and were not
concentrated at the root crown. Mixed infections in nodules occurred
at an average of 15% for soybeans and 3.5% for peas. Of interest is
the occurrence of nodules on peas containing bacteroids unreactive
with either fluorescent antibody (Fig. 7). R. j aponicum bacteroids
were morphologically similar to broth cultured cells, whereas R.
leguminosarum bacteroids were enlarged relative to broth cultured
cells, and often branched (Figs. 8 and 9).
37

38
HARVEST
...I..""I.... I... "I.... I... ».|M ■■■ I ...p10 20 30 40 50 SO 70 80 90 100
PERCENTLEGEND i STRRINS m a m a HANA 11 5-0 1 U U J NIXED
B2ZZ3 NITRAGIN 92A3 ' i UNIDENTIFIED
FIG. 7 PEA NODULE OCCUPANCY BY RHIZOBIUM LEGUMINOSARUM STRAINS

39
FIG. 8 R. JAPONICUM NODULE BACTERIA, STAINED WITH HOMOLOGOUS FLUORESCENT ANTIBODY. CELLS ARE MORPHOLOGICALLY
SIMILAR TO BROTH CULTURED CELLS.(Scale = 10 ym)
FIG. 9 R. LEGUMINOSARUM NODULE BACTERIA, STAINED WITH HOMOLOGOUS FLUORESCENT ANTIBODY. CELLS ARE ENLARGED RELATIVE TO
BROTH CULTURED CELLS, AND OFTEN BRANCHED.(Scale = 10 ym)

* *

DISCUSSION
One of the factors suggested as an important contributor to
specificity in the Rhizobium-legume association is the selective
stimulation of the homologous Rhizobium species in the rhizosphere of
its homologous host (7,14,25).
However, in this study I found that the growth of R. japonicum
was not selectively stimulated in the soybean rhizosphere, and that
the growth of R. leguminosarum was not selectively stimulated in the
pea rhizosphere. With the exception of harvest 2, rhizosphere numbers
of all strains of rhizobia were in the following order: soybeans>
peas>corn>non-rhizosphere soil, although differences in rhizosphere
populations of the crops are not always significant (Figs. 1 to 4).
During growth cycle 2 the plants were subjected to a considerable
short-term heat stress, due to the unfortunate failure of the air
conditioning unit on the plexiglas house. At harvest 2, rhizosphere
populations of the four Rhizobium strains were different than at
harvests 1, 3, and 4 (Figs. 1 to 4). Exudation from pea roots might
have been enhanced due to the heat stress. Soybean rhizosphere
populations of Rhizobium were reduced at harvest 2 relative to the
other harvests (Figs. 1 to 4). Apparently exudation from soybean
roots was reduced by the heat stress, or an inhibitory substance was
exuded. Hawaii 5-0 and USDA 110 were stimulated significantly more
in the rhizosphere of peas than in other rhizospheres at harvest 2
(Fig. 2). CB 1809 and Nitragin 92A3 were stimulated to the same
extent in the rhizospheres of peas and soybeans at harvest 2 (Fig. 2).

R. japonicum strains CB 1809 and USDA 110 were present in similar
numbers in any particular rhizosphere at any harvest (Figs. 1 to 4).
These strains apparently had similar rhizosphere colonizing abilities.
Hawaii 5-0 was generally seen in significantly lower numbers than the
other three strains in all rhizospheres at each of the four harvests
(Figs. 1 to 4). Hawaii 5-0 was apparently not as capable of rhizo
sphere colonization as the other strains in the rhizospheres tested.
In non-rhizosphere soil, CB 1809 was consistently seen in higher
numbers than the other strains (Figs. 1 to 4). Because this phenomenon
persisted through four harvests, it appeared that CB 1809 was better
adapted than the other strains to root-free soil.
Populations of individual Rhizobium strains never rose above
about 2 - 3 x 107 per gram of rhizosphere soil, even when soil was
continuously cropped with the same plant species. Thus, it seems as
though the rhizosphere might have a limited carrying capacity for
Rhizobium.
Bacteria enumerated with acridine organge were significantly more
prevalent in the soybean rhizosphere than the pea rhizosphere, than
the corn rhizosphere at harvests 3 and 4 (Fig. 5). At harvest 2,
total bacterial numbers were higher in the pea rhizosphere than the
soybean rhizosphere, probably due to the heat stress mentioned
earlier (Fig. 5). This pattern of rhizosphere colonization reflects
that seen with rhizobia, with bacteria more prevalent in the soybean
rhizosphere, than the pea rhizosphere, than the corn rhizosphere, with
the exception of harvest 2 (Figs. 2 to 4). Thus, the stimulation of
Rhizobium reflects the stimulation of rhizosphere bacteria in general.
42

The soybean nodule occupancy data were consistent with soybean
rhizosphere counts of the two R.. japonicum strains, as neither strain
predominated (Figs. 1 to 4 and 6). In the pea rhizosphere, Nitragin 92A3
was seen in numbers averaging twice those of Hawaii 5-0 (Figs. 1 to 4).
However, 92A3 was found in nine times as many nodules as Hawaii 5-0
(Fig. 7). Nitragin 92A3 appeared to be a more competent rhizosphere
colonizer than Hawaii 5-0, but increased rhizosphere populations of
Nitragin 92A3 do not fully explain the competitive advantage of this
strain in nodulating peas.
Hawaii 5-0 has been shown to be a very effective competitor in
nodulating lentils grown in several tropical soils, including an
Inceptisol (S. N. May, M.S. Thesis, 1979, University of Hawaii).
Perhaps the competitive ability of strains varies with respect to
different host species or cultivars, or with different soil types.
From these data, it appeared as though specific stimulation in
the rhizosphere was not a contributor to specificity in the R.
japonicum/soybean and R. leguminosarum/pea associations under the
conditions of this study. Differences in the growth of rhizobia in
host and non-host rhizospheres were manifested at the strain level
rather than at the species level. Thus some strains might simply be
better rhizosphere colonizers than others, without regard for
bacterial or plant species.
43

LITERATURE CITED
1. Bohlool, B. B. and E. L. Schmidt. 1968. Nonspecific staining:its control in immunofluorescence examination of soil. Science 162:1012-1014.
2. Bohlool, B. B. and E. L. Schmidt. 1973. A fluorescent antibodytechnique for determination of growth rates of bacteria in soil. Bull. Ecol. Res. Comm. (Stockholm) 17:229-236.
3. Bohlool, B. B. and E. L. Schmidt. 1974. Lectins: a possiblebasis for specificity in the Rhizobium-legume symbiosis. Science 185:269-271.
4. Bohlool, B. B. and E. L. Schmidt. 1980. The immunofluorescenceapproach in microbial ecology, pp. 203-241. In M. Alexander (Ed.) Advances in microbial ecology. Vol. 4. Plenum Publishing Corp., New York.
5. Broughton, W. J. 1978. Control of specificity in legume- Rhizobium associations. J. Appl. Bacteriol. 45:165-194.
6. Broughton, W. J., Ursula Samrey, and B. Ben Bohlool. 1982. Competition for nodulation of Pisum sativum cv. Afghanistan requires live rhizobia and a plant component. Can. J. Micro. 28: 162-168.
7. Brown, Margaret E., R. M. Jackson, and S. K. Burlingham. 1968. Growth and effects of bacteria introduced into soil. InT. R. G. Gray and D. Parkinson (Eds.) The ecology of soil bacteria. Liverpool Univ. Press, Liverpool, pp. 531-551.
8. Van Egeraat, A. W. S. M. 1975. The growth of Rhizobium leguminosarum on the root surface and in the rhizosphere in relation to root exudates. Plant and Soil 42:367-379.
9. Graham, Terrence L. 1981. Recognition in Rhizobium-legume symbioses. In Kenneth L. Giles and Alan G. Atherly (Eds.)Biology of the rhizobiaceae. Academic Press, Inc., New York.
10. Hoagland, D. R. and D. I. Arnon. 1938. The water culture method for growing plants without soil. Calif. A9, Exp. Sta. Circular 347.
11. Kingsley, Mark T. and B. Ben Bohlool. 1981. Release of Rhizobium spp. from tropical soils and recovery for immunofluorescence enumeration. Appl. Environ. Micro. 42:241-248.

45
12. Krasil'nikov, N. A. 1958. Soil microorganisms and higher plants. Acad. Sci. USSR Moscow. English ed. National Science Foundation.
13. MacGregor, A. N. and M. Alexander. 1972. Comparison of nodulating and non-nodulating strains of Rhizobium trifolii. Plant and Soil 36:129-139.
14. Nutman, P. S. 1965. The relation between nodule bacteria and the legume host in the rhizosphere and in the process of infection. In K. F. Baker and W. C. Snyder (Eds.) The ecology of soil-borne plant pathogens, pp. 231-247. Univ. Calif. Press, Berkeley.
15. Peters, R. J. and M. Alexander. 1966. Effect of legume exudateson the root nodule bacteria. Soil Sci. 102:380-387.
16. Purchase, H. F. and P. S. Nutman. 1957. Studies on the physiology of nodule formation. VI. The influence of bacterial numbers in the rhizosphere on nodule formation. Ann. Bot. Cond., N. S. 21:439.
17. Rovira, A. D. and J. R. Harris. 1961. Plant root excretions inrelation to the rhizosphere effect. V. The excretion of B-group vitamins. Plant and Soil 14:199.
18. Schmidt, E. L., R. 0. Bankole, and B. B. Bohlool. 1968.Fluorescent antibody approach to study of rhizobia in soil. J.Bact. 95:1987-1992.
19. Schmidt, E. L. 1974. Quantitative autoecological study of microorganisms in soil by immunofluorescence. Soil Science 118:141-149.
20. Schmidt, E. L. 1978. Ecology of the legume root nodule bacteria,pp. 269-303. In Y. R. Dommergues and S. V. Krupa (Eds.) Interactions between non-pathogenic soil microorganisms and plants. Elsevier Scientific Publishing Co., Amsterdam.
21. Schmidt, E. L. 1979. Initiation of plant "root-microbe interactions. Ann. Rev. Microbiol. 33:355-376.
22. Schmidt, E. L. and B. B. Bohlool. 1981. The role of lectins insymbiotic plant-microbe interactions. In W. Tanner andF. A. Loewus (Eds.) Encyclopedia of plant physiology, New Series Vol. 13 B. Plant Carbohydrates II. Springer-Verlag, Berlin, Heidelberg.
23. Tsien, H. C., P. S. Cain, and E. L. Schmidt. 1977.' Viability ofRhizobium bacteroids. Appl. Environ. Micro. 34:854-856.
24. Vincent, J. M. 1970. A manual for the practical study of rootnodule bacteria. International Biological Programme Handbook no. 15. 'Blackwell Scientific Publications, Oxford, England.

46
25. Vincent, J. M. 1974. Root nodule symbiosis with Rhizobium.pp. 265-341. In A. Quispel (Ed.) The biology of nitrogen fixation. North Holland Publishing Co., Amsterdam.

POPULATIONS OF RHIZOBIUM IN THE RHIZOSPHERES OF HOST
AND NON-HOST PLANTS DURING A 35-DAY GROWTH CYCLE
CHAPTER 3

ABSTRACT
The growth of two R. j aponicum strains (USDA 110 and CB 1809) and
two R. leguminosarum strains (Hawaii 5-0 and Nitragin 92A3) was fol
lowed in the rhizospheres of soybean, pea, and corn plants growing in
non-sterile soil. Rhizosphere soil was sampled at weekly intervals
over a 35-day growth period. Rhizosphere numbers of each strain were
determined by membrane filter immunofluorescence, using strain-
specific fluorescent antibodies. Numbers of the four strains were
highest in the soybean rhizosphere at 1 week, averaging 2.0 x 107 per
gram of soil. Numbers of the four strains peaked in the pea rhizo-
sphere at 4 weeks, averaging 1.5 x 10 per gram of soil. At 1, 2, and
3 weeks, numbers per gram of soil of all strains were significantly
higher in the soybean rhizosphere than in the pea rhizosphere, than in
the corn rhizosphere, than in fallow soil. At week 4, pea rhizosphere
numbers per gram of soil of all strains significantly surpassed soybeans
rhizosphere numbers (1.5 x 107 and 1.2 x 107, respectively). At 5
weeks, rhizosphere numbers of all strains were in the following order:
pea>corn>soybean>fallow soil. Differences between Rhizobium numbers
in the rhizospheres of the three crops were more difficult to detect
on a root surface area basis. Numbers of all strains declined at 5
weeks per gram of soil and per square cm of root. No specific
stimulation of rhizobia by their legume hosts was observed. Strains
Nitragin 92A3, CB 1809, and USDA 110 were more successful rhizosphere
colonizers than Hawaii 5-0. Acridine orange total counts of bacteria
were highest on the basis of soil weight in the rhizospheres of the
three crops at 1 week, and following a decline at 2 weeks, remained

relatively constant for the remainder of the experiment. Acridine
orange total counts on the basis of root surface area were variable,
and patterns were not evident. Percentages of soybean nodules
occupied by the two R. j aponicum strains reflected the rhizosphere
populations of the strains, as neither predominated. Mixed
infections were observed at an average of 7%. The majority of large
nodules on peas contained Nitragin 92A3, and a lesser number,
Hawaii 5-0. Sixteen percent of large pea nodules contained R.
leguminosarum bacteroids unreactive with fluorescent antibodies
prepared against the two inoculant strains. In small pea nodules,
either one or both of the two inoculant strains were present in most
nodules at harvests 3 and 4. An unidentified strain or strains
occupied 74% of the small pea nodules at harvest 5. Mixed infections
in pea nodules varied from 4 to 10% in small and large nodules,
respectively. I found no evidence to support the theory of specific
stimulation of rhizobia by their homologous legume hosts in the R.
j aponicum/soybean and R. leguminosarum/pea associations.
49

INTRODUCTION
Rhizobia are distinguished from other soil bacteria by their
unique ability to form nitrogen-fixing nodules on the roots of many
leguminous plants. Rhizobia are specific with respect to the hosts
they nodulate. Only certain species of Rhizobium are able to infect
and nodulate the roots of particular legumes. Although the relation
ship between rhizobia and their host legumes has been studied in
great detail, the basis for this specificity is not completely clear
(3,16).
Several possible mechanisms have been proposed to account for
specificity, involving both plant and bacterial components (3).
Cellular recognition between host and bacterium is considered by
some as a mechanism for specificity. Plant proteins called lectins
are thought to interact specifically with rhizobia at the root
surface (6,17).
Another mechanism proposed as a possible contributor to
specificity in the Rhizobium-legume symbiosis is the preferential
growth of the appropriate Rhizobium in the rhizosphere of its
homologous host plant over and above that of other non-homologous
rhizobia, and over other soil microorganisms (3,10,19).
Most studies of stimulation in the legume rhizosphere have been
carried out in aseptic systems, from which rhizobia can be conve
niently plated and counted (5,9,11). Extrapolation from aseptic
conditions to non-sterile field soil, in which the full range of
microbial interactions would be occurring, is unrealistic, and
probably erroneous.

In a study described earlier (Chapter 2) rhizosphere numbers of
four strains of Rhizobium were determined in homologous and non-
homologous legume rhizospheres over four successive 35-day growth
cycles. Rhizobium populations were assessed on the basis of soil
weight by membrane filter immunofluorescence with strain-specific
fluorescent antibodies (1,8,15). Specific stimulation of homologous
rhizobia by their legume hosts was not observed, and rhizosphere
colonization by rhizobia reflected more strain differences than
species differences. It is possible however, that specific stimulation
could have occurred earlier in the growth cycle, possibly just prior
to the onset of nodulation. If this was the case, an experiment in
which rhizosphere sampling and enumeration was done at 35 days would
probably overlook specific stimulation.
Rovira (14) suggested that microbial densities in the rhizo
sphere on a unit soil weight basis be used with caution, since
rhizosphere soil weights are affected by plant species, soil moisture,
soil structure, and handling of the root during recovery of rhizo
sphere soil. He suggested that rhizosphere numbers be expressed on
the basis of root weight. Reyes and Schmidt (13) suggested that an
even more conservative method would be to express microbial densities
on the basis of root surface area, as roots of various plant species
vary in size, type and growth characteristics. They conducted a
series of experiments (12,13) to determine if stimulation of R.
japonicum strain 123 in the soybean rhizosphere is a possible
contributor to this strain’s extraordinary success in nodulating
soybeans in soils of the Midwest (7). They were unable to detect
any specific stimulation of strain 123 in the soybean rhizosphere.
51

What follows is the description of an experiment in which
membrane filter immunofluorescence was used to assess the population
dynamics of homologous and non-homologous rhizobia in the rhizospheres
of host and non-host plants over one 35-day growth cycle, with har
vests at weekly intervals. Immunofluorescence counts of rhizobia and
acridine orange counts of total bacteria are presented on the basis
of root surface area as well as rhizosphere soil weight.
52

MATERIALS AND METHODS
Rhizobium Strains and Inoculum Preparation
Kula loam soil, an Inceptisol, was prepared as described
previously (Chapter 2). R. j aponicum strains CB 1809 and USDA 110,
and R. leguminosarum strains Hawaii 5-0 and Nitragin 92A3 were
obtained from the collection of B. B. Bohlool of the University of
Hawaii. Strains were maintained on yeast-extract mannitol agar
slants (18) with 1.0 g of yeast extract (Difco Laboratories, Detroit,
Michigan) substituted for yeast-water. Inoculum was prepared as
previously described (Chapter 2). Initial levels of the four
Rhizobium strains per gram of moist soil were as follows as determined
by membrane filter immunofluorescence: R. japonicum CB 1809,
4.7 x 105, and USDA 110 4.6 x 105, R. leguminosarum Hawaii 5-0,
3.8 x 105 and Nitragin 92A3 4.3 x 105.
Pots and Planting
Pots were 25 cm in diameter and 20 cm high and were painted with
a heavy coat of flat white paint. A watering device was placed into
each pot to provide water to the plants in the cores as evenly as
possible (Diagram 1). The pots were than each filled with 5.5 kg of
moist Kula soil. Cores of p.v.c. pipe of various sizes were then
driven into the soil in the pattern outlined in Diagram 2. The soil
inside of the cores was loosened slightly with a stout wire. The
soil was adjusted to about -1/3 bars of moisture tension with
deionized water. Three-day-old soybean, pea, and corn seeds (pre
germinated as described before) were planted, one per core. Following
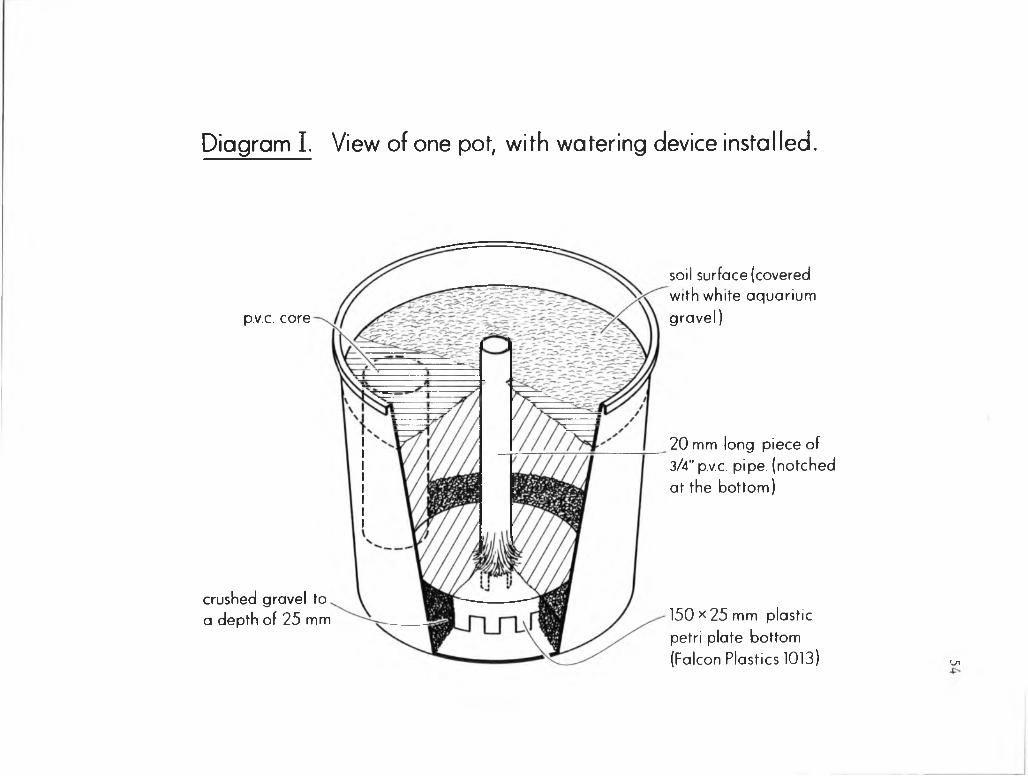
Diagram I. View of one pot, with watering device installed.
20 m m long piece of 3/4" p.v.c. pipe, (notched at the bottom)
p.v.c. core
soil surface (covered with white aquarium gravel)
crushed gravel to a depth of 25 m m 150 x 25 m m plastic
petri plate bottom (Falcon Plastics 1013) Ui

55
Diagram 2. Arrangement of cores and order of harvest for the 35 - day time course study.
Perimeter of the watering device
Legend1. V'dia x 4" long p.v.c. pipe, 4. IV2" dia x 5" long p.v.c. pipe,harvested at 1 week harvested at 4" weeks.
2. 1" dia x 5"long p.v.c. pipe, 5. 2" dia x 5" long p.v.c. pipe,harvested at 2 weeks. harvested at 2 weeks.
3. 1" dia x 5" long p.v.c. pipe, harvested at 3 weeks.

planting, the soil surface was covered with rinsed white aquarium
gravel, to prevent undue soil heating and to inhibit the growth of
algae.
Plant Growth Conditions
The soil moisture tension was maintained at about -1/3 bars
throughout the experiment by watering to a constant weight with
deionized water. Water was poured into the central upright tube of
the watering device. In this manner, water was supplied to the roots
in the cores as evenly as possible from the perimeter of the base of
the device, rather than from a single central location. The experi
ment was set up as a randomized complete block with three replications
and carried out under natural lighting in a greenhouse.
Harvest and Sample Preparation
At harvests 1, 2, and 3, four cores were removed from each pot
as outlined in Diagram 2. At harvests 4 and 5, two cores were
removed from each pot (Diagram 2). Root systems were separated from
the cores by gently tapping the cores on the inside of a metal pan.
Soil that adhered to the root system after gentle shaking was
considered to be rhizosphere soil. All other soil was returned to
the cores in the pots and gently compacted. Non-rhizosphere soil was
removed from the cores as above, pooled, mixed, and a 10-gram sample
taken. The remainder was placed back into the cores as described
above.
Bacteria were released from root surfaces and rhizosphere soil
particles as previously described (Chapter 2). At harvest 1, 50 ml
56

of the gelatin-ammonium phosphate mixture (8) was used for extractions
rather than 100 ml, because of the small amount of rhizosphere soil
present.
Root Surface Area Estimation
Root surface area was estimated by the method of Carlson (4), as
modified by Reyes and Schmidt (13) for greenhouse-grown soybeans.
Enumeration of Bacteria
Membrane filter counts were performed as described by Kingsley
and Bohlool (8). Duplicate filters were counted for each strain from
each pot, for a total of 96 filters per harvest.
Acridine orange total counts were made at each harvest as
previously described. Three filters were counted per pot, for a
total of 36 per harvest.
Nodule Typing
All nodules were removed, squashed, smeared on microscope
slides, and serologically typed with fluorescent antibodies as
previously described (Chapter 2).
57

RESULTS
Rhizosphere numbers of each of the four inoculant strains are
presented at weekly intervals on the basis of soil weight in Figures 1
to 5. Numbers of all strains per gram of rhizosphere soil are in the
following order at 1, 2, and 3 weeks: soybean>pea>corn>non-rhizosphere
soil (Figs. 1 to 3). At 4 weeks rhizobia per gram of soil had declined
in the soybean rhizosphere, but increased in that of peas, signifi
cantly surpassing soybean rhizosphere numbers (Fig. 4). Rhizobium
numbers per gram of soil of all strains were higher in the soybean
rhizosphere than that of corn at 4 weeks, with the exception of
Hawaii 5-0 (Fig. 4). At 5 weeks, numbers of rhizobia per gram of soil
were in the following order: pea>corn>soybean>non-rhizosphere soil.
Rhizosphere numbers of each of the four strains are presented at
weekly intervals on the basis of root surface area in Figures 6 to 10.
Differences in rhizosphere populations of Rhizobium between the crops
were more difficult to detect on the basis of root surface area,
owing to the variability in the estimations of root surface area
(Figs. 6 to 10).
Rhizosphere bacteria enumerated using acridine orange fluores
cence microscopy were in the following order: soybean>pea>corn>non-
rhizosphere soil. However, the variability of the acridine orange
total counts was high, and differences between crops and non-rhizo
sphere soil are often not significant (Fig. 11). Differences between
rhizosphere populations of bacteria per square cm of root between
crops were even more difficult to detect, with the only significant
differences at 5 weeks (Fig. 12).

59
FIG. 1 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 1
* denotes L.S.D. of less than 0.05

60
L.S.D. .05
0 .0* 0 .0*
STRAIN
R. japonicum
NITRAGIN
R. leguminosarum
HANAII 5-0
FIG. 2 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 2
.2
' .05
* denotes L.S.D. of less than 0.05

61
L.S.D. .05
n . s. 0.3 0.1 0 .0*
CROP
USDR 110
R. japonicum
CB 1808
HflHflll 5-0
N1TRRGIN 92R3
R. leguminosarum
FIG. 3 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 3
* denotes L.S.D. of less than 0.05

62
L.S.D. .05
N1TRA81N 9203
R. leguminosarum
HflHflll 5-0
STRAIN
USDA 110
R. iaponicum
CB 1808
L.S.D
FALLONCROP
FIG. 4 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 4
* denotes L.S.D. of less than 0.05

63
L.S.D. .05
n.s. 0.1 0.1
L.S.D
STRAIN
USDA 110
R. japonicum
CB 1809
NITnASIN 92B3
R. leguminosarumHAMAI1 5-0
FIG. 5 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT HARVEST 5

64
L.S.D..05
1.4 0.3
/I / n.s.
n. s.
L.S.D. .05
n.s.
FIG. 6 COUNTS OF 4 RHIZOBIUM STRAINS (X 10*) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES AT HARVEST 1

65
L.S.D. 05
CROP
L.S.D.
U3DR 110
R. japonicum
CB 1B09
NITRAGIN 92A3
R. leguminosarum
HAWAII 5-0
U. 7
FIG. 7 COUNTS OF 4 RHIZOBIUM STRAINS (X 104) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES AT HARVEST 2

66
L.S.D. .05
n.s.
US0A 110
R. japonicum
CB 1809
NITRAGIN 92R3
CROP
n.s. L.S.D.
n.s.R. leguminosarum
HflNRlI 5-0
FIG. 8 COUNTS OF 4 RHIZOBIUM STRAINS (X 104) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES AT HARVEST 3
S .
.05

67
FIG. 9 COUNTS OF 4 RHIZOBIUM STRAINS (X 10A) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES AT HARVEST 4

STRAIN
n.s. 0.3
1.3L.S.D.
NITRAGIN 82A3
R. leguminosarum
CROP
USOA 110
R. japonicum
CB 1808
12.4
12.6
FIG. 10 COUNTS OF 4 RHIZOBIUM STRAINS (X 10A) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES AT HARVEST 5

69
HAHVE3T
FIG. 11 ACRIDINE ORANGE TOTAL COUNTS OF BACTERIA (X 108) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL

HARVEST
FIG. 12 ACRIDINE ORANGE TOTAL COUNTS OF BACTERIA (X 107) PER SQUARE CENTIMETER OF ROOT AREA
IN 3 RHIZOSPHERES

Soybean nodules contained the two inoculant R. japonicum strains
as outlined in Figure 13. Mixed infections were observed at an
average of 7%. Large and small nodules formed on pea plants. Large
nodules were elongated and red inside. Small nodules were nearly
spherical, and white throughout. Large and small nodules contained
the two inoculant R. 1eguminosarum strains as well as an unidentified
strain or strains as outlined in Figures 14 and 15. Mixed infections
occurred in large and small pea nodules at an average of 10 and 5%,
respectively.
Photographs of the experiment are presented at 1, 3, and 5 weeks
in Figures 16, 17, and 18.
71

72
HARVEST
1 2
3
4
510 20 30 40 50 60 70 80 90 100
PERCENTLEGENO> STRAINS ESSES CB 1809 B C T MIXED USDA 110
FIG. 13 SOYBEAN NODULE OCCUPANCY BY RHIZOBIUM JAPONICUM STRAINS

73
HARVEST
2
3
4
5
- - - - - -
8 §10 20 30 40 50 60 70 80 90 100
PERCENTLEGENDi STRAINS l w w a i HAWAII 5-0
NITRAGIN 92A3LAAAAJ m i x e d i 1 UNIDENTIFIED
FIG. 14 LARGE PEA NODULE OCCUPANCY BY RHIZOBIUM LEGUMINOSARUM STRAINS

74
HARVEST
3
4
5
tO 20 30 40 50 80 70 80 90 tOOPERCENT
LEGENDt STRAINS HAWAII 5-0 lxaaaj MIXEDZZZZZZZ NITRAGIN 92A3 i 1 UNIDENTIFIED
FIG. 15 SMALL PEA NODULE OCCUPANCY BY RHIZOBIUM LEGUMINOSARUM STRAINS

. 16 VIEW OF THE EXPERIMENT AT 1 WEEK. GROWTH OF THE EXPERIMENTAL PLANTS WAS UNIFORM WITHIN EACH SPECIES,
AND WITHIN AND BETWEEN REPLICATE POTS.

76

77
FIG. 17 VIEW OF THE EXPERIMENT AT 3 WEEKS. PLANTS GREW WELL DESPITE THE CONFINEMENT OF THEIR ROOT SYSTEMS BY CORES.CORN PLANTS ARE STARTING TO SHOW NITROGEN DEFIEIENCY
SYMPTOMS (CHLOROSIS OF OLDER GROWTH), WHEREAS THE LEGUMES ARE GREEN AND HEALTHY.
FIG. 18 VIEW OF THE EXPERIMENT AT 5 WEEKS. AT 5 WEEKS, PEAS WERE FILLING PODS AND SOYBEANS HAD FLOWERED. THE NITROGEN
DEFICIENCY OF CORN WAS MORE PRONOUNCED, AS OLDER LEAVES WERE SEVERELY CHLOROTIC OR NECROTIC.

78

DISCUSSION
One of the factors proposed as a possible contributor to
specificity in the Rhizobium-legume symbiosis is the selective
stimulation of the homologous Rhizobium species in the rhizosphere of
its homologous host (3,10). In the previous study (Chapter 2) I
found that species of rhizobia were not specifically stimulated in
the rhizospheres of their host at 35 days. I designed this study to
examine the population dynamics of rhizobia in host and non-host
rhizospheres at weekly intervals during the growth of the plants.
Reyes and Schmidt (13) suggested that root surface area might be a
better basis for expressing rhizosphere populations of rhizobia, so
root area was estimated, and data presented on the basis of root area
as well as soil weight.
As was seen previously (Chapter 2), rhizosphere stimulation of
host and non-host rhizospheres was dependent more on the Rhizobium
strains than on the particular species of Rhizobium. At 1, 2, and
3 weeks, numbers per gram of soil of the strains were in the follow
ing order: soybean rhizosphere>pea>corn>non-rhizosphere soil
(Figs. 1 to 3). Differences between crops at these harvests were
all significant at the .05 level. At harvests 4 and 5 soybean
rhizosphere* numbers decreased (Figs. 4 and 5). At harvest 5 both pea
and corn rhizospheres harbored significantly more rhizobia of all
strains than the soybean rhizosphere (Fig. 5). This sharp decline
of all strains in the soybean rhizosphere might be an artifact of
having grown the plants in cores. At 4 and 5 weeks much of the
soybean root system had emerged from the bottom of the cores, and

roots proliferated rapidly in the bottom of the pots. The lowered
numbers in the rhizospheres of peas and soybeans at 5 weeks probably
reflect the fact that roots were growing rapidly, and enmeshing
rhizosphere soil faster than the growth of the inoculant strains
could keep up.
Differences in rhizosphere populations of rhizobia in the rhizo
spheres of the three crops were harder to detect on the basis of root
surface area. Patterns of rhizosphere colonization were similar to
those observed when rhizosphere populations were examined on a soil
weight basis (Figs. 6 to 10). Significant differences between crops
were only observed at 2 and 5 weeks (Figs. 7 and 10). Variability
between root surface area estimations was high. This variability
precluded the detection of differences between Rhizobium numbers in
the rhizospheres of the three crops for harvests 1, 3, and 4 (Figs. 6,
8, and 9). Significant differences were observed between strains on
the basis of root surface area across the five harvests, however
differences were not indicative of specific stimulation (Figs. 6 to
10). As was seen previously (Chapter 2), numbers of the two R.
japonicum strains were generally similar, and similar to numbers of
R. leguminosarum strain Nitragin 92A3. R. leguminosarum strain
Hawaii 5-0, however, is seen in significantly lower numbers than the
other three strains in all rhizospheres and across all five harvests
(Figs. 1 to 10). This would indicate rhizosphere colonization more
on the basis of differences between strains, rather than species
differences, with Hawaii 5-0 being less adapted to the conditions in
the rhizospheres of all plants.
80

As was seen previously (Chapter 2), soybean nodule occupancy data
approximated the rhizosphere counts of USDA 110 and CB 1809, as
neither dominated (Fig. 13). In large pea nodules, Nitragin 92A3
dominated Hawaii 5-0 with the exception of harvest 2, when percentages
of the two strains were nearly equal (Fig. 14). Nodule occupancy of
small pea nodules was different. At harvest 3, both inoculant strains
occupied nodules in approximately equal percentages, and unidentified
bacteroids occupied a nearly equal percentage (Fig. 15). However,
Hawaii 5-0 occupied more small nodules than Nitragin 92A3 at harvests
4 and 5, and at harvest 5 unidentified bacteroids predominated
(Fig. 15).
Hawaii 5-0 has been shown to be very competitive in nodulating
lentils in many tropical soils, including an Inceptisol (S. N. May,
M.S. Thesis, 1979, University of Hawaii). However, Hawaii 5-0 was
not as competitive as Nitragin 92A3 in nodulating peas. Perhaps the
soil or host plant influence the competition between strains for
nodulation of legumes.
Plants grew well in the Kula soil, despite having their root
systems confined to cores. Lack of uniformity within or between
replicates was not a problem (Figs, 16, 17, and 18). At 5 weeks, the
corn was slightly chlorotic relative to the legumes, probably due to
a nitrogen deficiency, as no amendments (other than rhizobia) were
made to the soil (Fig. 18).
In this study, no evidence was found in support of specific
stimulation of R. j aponicum in the rhizosphere of soybeans or R.
1eguminosarurn in the rhizosphere of peas. Rhizosphere colonization
81

by rhizobia reflected more strain differences than species
differences.
Rhizosphere numbers of rhizobia were quite high in the soybean
and pea rhizospheres at 1 week, increasing from about 4 - 5 x 105 per
gram of moist soil at zero time to 1 - 2 x 107 per gram of oven dry
soil at 1 week. Although no specific responses were observed on the
basis of rhizosphere growth during this study, it is possible that
specific interactions occurred before the first sampling time, at
1 week. This possibility will be investigated in a later experiment.
82

LITERATURE CITED
Bohlool, B. B. and E. L. Schmidt. 1973. A fluorescent antibodytechnique for determination of growth rates of bacteria in soil.Bull. Ecol. Res. Comm. (Stockholm) 17:229-236.
Broughton, W. J. 1978. Control of specificity in legume-Rhizobium associations. J. Appl. Bacteriol. 45:165-194.
Brown, Margaret E., R. M. Jackson, and S. K. Burlingham. 1968. Growth and effects of bacteria introduced into soil. In T. R. G. Gray and D. Parkinson (Eds.) The ecology of soil bacteria. Liverpool Univ. Press, Liverpool, pp. 531-551.
Carlson, J. B. 1969. Estimating surface area of soybean root systems. J. Minn. Acad. Sci. 36:16-19.
Van Egeraat, A. W. S. M. 1975. The growth of Rhizobiumleguminosarum on the root surface and in the rhizosphere in relation to root exudates. Plant and Soil 42:367-379.
Graham, Terrence L. 1981. Recognition in Rhizobium symbioses.In Kenneth L. Giles and Alan G. Atherly (Eds.) Biology of the Rhizobiaceae. Academic Press, Inc., New York.
Ham, G. E., L. R. Frederick, and L. C. Anderson. 1971. Serogroups of Rhizobium j aponicum in soybean nodules sampled in Iowa. Agron.J. 63:69-72.
Kingsley, Mark T. and B. B. Bohlool. 1981. Release of Rhizobium spp. from tropical soils and recovery for immunofluorescence enumeration. Appl. Environ. Micro. 42:241-248.
MacGregor, A. N. and M. Alexander. 1972. Comparison of nodulating and non-nodulating strains of Rhizobium trifolii. Plant and Soil 36:129-139.
Nutman, P. S. 1965. The relation between nodule bacteria in the rhizosphere and in the process of infection. In K. F. Baker and W. C. Snyder (Eds.) The ecology of soil-borne plant pathogens, pp. 231-247. Univ. Calif. Press, Berkeley.
Peters, R. J. and M. Alexander. 1966. Effect of legume exudates on the root nodule bacteria. Soil Sci. 102:380-387.
Reyes, V. G. and E. L. Schmidt. 1979. Population densities of Rhizobium japonicum strain 123 estimated directly in soil and rhizospheres. Appl. Environ. Micro. 37:854-858.

84
13. Reyes, V. G. and E. L. Schmidt. 1981. Populations of Rhizobiumj aponicum associated with the surfaces of soil-grown roots. Plant and Soil 61:71-80.
14. Rovira, A. D. 1961. Rhizobium numbers in the rhizosphere of redclover and paspalum in relation to soil treatment and numbers ofbacteria and fungi. Aust. J. Agric. Res. 12:77-83.
15. Schmidt, E. L. 1974. Quantitative autoecological study of microorganisms in soil by immunofluorescence. Soil Sci. 118:141-149.
16. Schmidt, E. L. 1978. Ecology of the legume root nodule bacteria,pp. 269-303. In Y. R. Dommergues and S. V. Krupa (Eds.) Interactions between non-pathogenic soil microorganisms and plants. Elsevier Scientific Publishing Co., Amsterdam.
17. Schmidt, E. L. and B. B. Bohlool. 1981. The role of lectins inplant-microbe interactions, pp. 658-677. In W. Tanner andF. A. Loewus (Eds.) Encyclopedia of plant physiology, New Series, Vol. 4. Plant carbohydrates II. Springer-Verlag, Berlin, Heidelberg.
18. Vincent, J. M. 1970. A manual for the practical study of rootnodule bacteria. International Biological Programme Handbook no. 15. Blackwell Scientific Publications, Oxford, England.
19. Vincent, J. M. 1974. Root nodule symbiosis with Rhizobium.pp. 265-341. In Quispel (Ed.) The biology of nitrogen fixation. North Holland Publishing Co., Amsterdam.

RHIZOBIUM POPULATION DYNAMICS IN HOST AND NON-HOST RHIZOSPHERES
DURING THE FIRST NINE DAYS OF RHIZOSPHERE DEVELOPMENT
CHAPTER 4

ABSTRACT
The growth of two R. japonicum strains (USDA 110 and CB 1809) and
two R. leguminosarum strains (Hawaii 5-0 and Nitragin 92A3) was fol
lowed in non-sterile rhizospheres. In one study, growth of these
strains was followed in the rhizospheres of soybeans and peas at
planting, 24, and 48 hours, and rhizosphere growth rates estimated.
Mean doubling times for the strains were 12 to 13 hours in the soybean
rhizosphere and 9 to 11 hours in the pea rhizosphere. Total counts of
bacteria measured by acridine orange staining indicated that other
bacteria were not as strongly stimulated by the rhizosphere as
rhizobia in the early stages of plant growth. In another study,
rhizosphere populations of the four strains were followed in the
rhizospheres of soybeans, peas, and corn, as well as in non-rhizo
sphere soil at 3, 5, 7, and 9 days. Rhizosphere numbers of Rhizobium
increased from their starting levels of 4 - 9 x 10s per gram of moist
soil to 1 - 2 x 107 per gram of moist soil at 3 days, and declined
thereafter. Populations of bacteria enumerated with acridine orange
remained relatively constant. This early burst of the growth of
rhizobia might indicate that they responded faster to the conditions
of the rhizosphere than the bacterial population at large, but were
diluted out as the root systems expanded rapidly. No specific stimu
lation of the growth of rhizobia was detected in the rhizospheres of
their host plants. Rhizosphere populations reflected strain differ
ences more than species differences, in all crops and at all harvests.

INTRODUCTION
Soil bacteria of the genus Rhizobium are distinguished from other
genera by their ability to form nitrogen-fixing nodules on the roots
of leguminous plants. Rhizobia are specific with respect to the hosts
they nodulate. Only certain species of Rhizobium are able to infect
and nodulate the roots of particular legumes. Although the relation
ship between host and bacterium has been studied in great detail, the
basis for this specificity is not completely clear (1,7).
One proposed mechanism as a possible contributor to specificity
is the preferential growth of the appropriate Rhizobium in the
rhizosphere of its homologous host plant over other soil bacteria and
rhizobia (2,4,9).
In studies described earlier (Chapters 2 and 3), rhizosphere
numbers of four Rhizobium strains were determined at harvest across
four 35-day harvests, as well as at weekly intervals during one 35-day
growth cycle. I observed no specific stimulation of growth of
Rhizobium species in the rhizospheres of their homologous host plants
in these studies. When rhizosphere populations were sampled once a
week, numbers of all strains increased from their starting levels of
about 4 x 105 per gram of soil to a level of about 1 x 107 per gram of
soil at 1 week, and generally declined thereafter.
Following is the description of two experiments in which population
dynamics of homologous and non-homologous rhizobia were followed in the
rhizospheres of host and non-host plants. Rhizosphere populations were
sampled in one study at planting, 24, and 48 hours, and in another
study at 3, 5, 7, and 9 days.

MATERIALS AND METHODS
Inoculum Preparation and Rhizobium Strains
Fresh Kula loam soil, prepared as described previously, was used
for the 9-day time course study. Initial levels of each of the strains
per gram of moist soil were as follows, as determined by immuno
fluorescence: R. j aponicum USDA 110, 8.3 x 105 and CB 1809, 9.2 x 105,
_R. leguminosarum Nitragin 92A3, 4.2 and 105 and Hawaii 5-0, 3.9 x 105.
For the early rhizosphere growth rate study, about 5 x 109 cells of
each strain (as estimated by optical density) were transferred from
yeast-extract mannitol broth (8) culture and blended with 2000 grams
of fresh Kula soil, to give about 2.5 x 106 cells per gram of moist
soil. This intermediate dilution was blended with more moist Kula soil
275 g of intermediate dilution to 5225 g of bulk soil. This 1:20
dilution was expected to give about 1.25 x 105 cells per gram of moist
soil. Actual starting numbers at planting were slightly higher than
this estimate, as follows: USDA 110, 4.3 x 10s, CB 1809, 5.1 x 105,
Nitragin 92A3, 4.3 x 105 and Hawaii 5-0, 4.7 x 105. Strains were
obtained from the collection of B. B. Bohlool of the University of
Hawaii, and were grown and maintained as described previously.
Pots and Planting
Pots were 25 cm in diameter and 20 cm high, and were painted with
a heavy coat of flat white paint. Each was filled with 5.5 kg of
moist Kula soil. Into each pot was planted 80 seeds. Soybeans (Davis)
peas (Wilt-resistant Wisconsin Perfection), and sweetcorn (Hawaiian
Supersweet #9) were the crops. In the growth rate study, only peas and
soybeans were planted. Seeds were not pregerminated. Following

planting, the soil surface was covered with rinsed white aquarium
gravel to prevent undue soil heating and the growth of algae.
Deionized water was added to bring the soil to a moisture tension of
about -1/3 bars.
Soil moisture tension was maintained at about -1/3 bars by
watering to constant weight with deionized water. Each experiment was
set up as a randomized complete block in three replications and carried
out in a greenhouse under natural lighting conditions. Soil temper
ature during the afternoon was about 30°C.
Harvest and Sample Preparation
For the 9-day time course, I harvested 15 to 20 plants from each
pot at 3, 5, 7, and 9 days after the planting of dry seeds. Forty
plants were dug out (by hand) from each pot at 24 and 48 hours in the
growth rate study. Rhizosphere soil was considered to be that which
adhered to seeds and root system after gentle shaking. Plants and
root systems were placed into 18-ounce Whirlpak bags for transport to
the laboratory. A sample of approximately 10 grams was removed from
the non-rhizosphere soil pots at each time point during the 9-day time
course. Ten-gram samples were removed from pea and soybean pots
immediately before the planting of the growth rate study.
Bacteria were released from root surfaces and rhizosphere soil
particles for enumeration as previously described (Chapter 2). At
harvest 1 of the 9-day time course, 50 ml of the extractant mixture
(3) was used rather than 100 ml, because of the small amount of rhizo
sphere soil present. Twenty and 50 ml of extractant were used for the
24-and 48-hour enumerations, respectively, of the growth rate study.
89

Membrane Filter Immunofluorescence and Acridine Orange Total Counts
90
Membrane filter counts were performed as described by Kingsley
and Bohlool (3). Duplicate filters were counted for each strain from
each pot, for a total of 96 filters per harvest in the 9-day time
course and 48 per harvest of the growth rate study.
I made acridine orange counts at each harvest as previously
described (Chapter 2). Three filters were counted per pot, for a
total of 36 per harvest in the time course and 18 per harvest in the
growth rate study.

RESULTS
Rhizosphere numbers of each of the four inoculant strains are
presented at 3, 5, 7, and 9 days in Figures 1 to 4. Acridine orange
total counts at the above time points are presented in Figure 5.
Populations of each of the four inoculant strains are presented
at planting, 24, and 48 hours in the pea and soybean rhizospheres in
Figures 6 and 7. Estimates of mean doubling times of inoculant strains
are also presented in Figures 6 and 7. At 24 hours, soybeans had
radicles about 1 cm long, and peas were swollen, but radicles had not
yet extended. At 48 hours, soybeans and peas had radicles of about
6 cm and 1 cm, respectively. Rhizosphere populations of bacteria
estimated by acridine orange total counts are presented at planting,
24, and 48 hours in Figure 8.
Percentages of Rhizobium (the four inoculant strains combined) in
the rhizosphere population of bacteria are given for each study in
Tables 1 and 2, respectively.
Nodules were not observed on the roots of the legumes, due to the
short duration of the experiment.
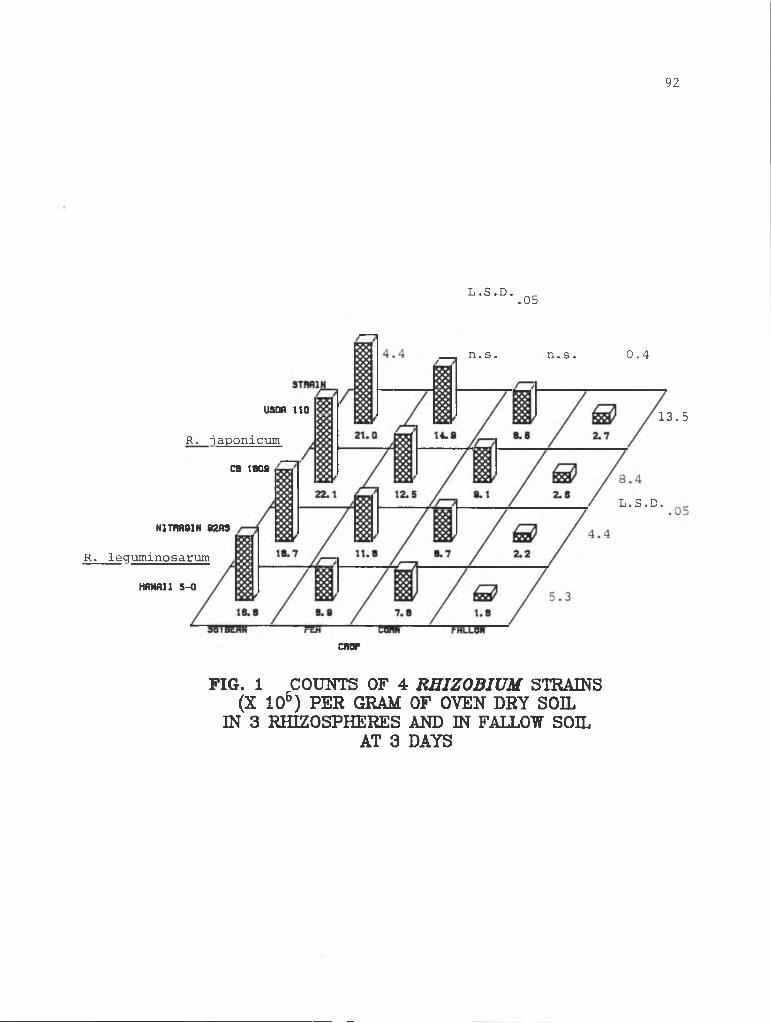
92
L.S.D. .05
n.s. n.s. 0.4
L.S.D
13.5
chop
USDA ITO
R. japonicumCB 1809
N1TIW61N 92H3
R. leguminosarumHAHA11 5-0
FIG. 1 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT 3 DAYS

93
L.S.D. .05
CROP
L.S.D.
usoa 110
R. japonicumCB 1809
N1TIW91N 8203
R. leguminosarum
HflHflll 5-0
FIG. 2 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT 5 DAYS

94
FIG. 3 COUNTS OF 4 REIZOBIUM STRAINS (X 105) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT 7 DAYS

95
L.S.D. .05
CROP
STRAIN
USDA 110
R. japonicumCB 1809
HAWAII 5-0
N1TRA61N 92A3
R. leguminosarum
L.S
FIG. 4 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL AT 9 DAYS

96
HARVEST
FIG. 5 ACRIDINE ORANGE TOTAL COUNTS OF BACTERIA (X 108) PER GRAM OF OVEN DRY SOIL
IN 3 RHIZOSPHERES AND IN FALLOW SOIL

97
FIG. 6 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN THE PEA RHIZOSPHERE AT PLANTING, 24, AND 48 HOURS
(10.0)
(8.9)
7)
(hours)
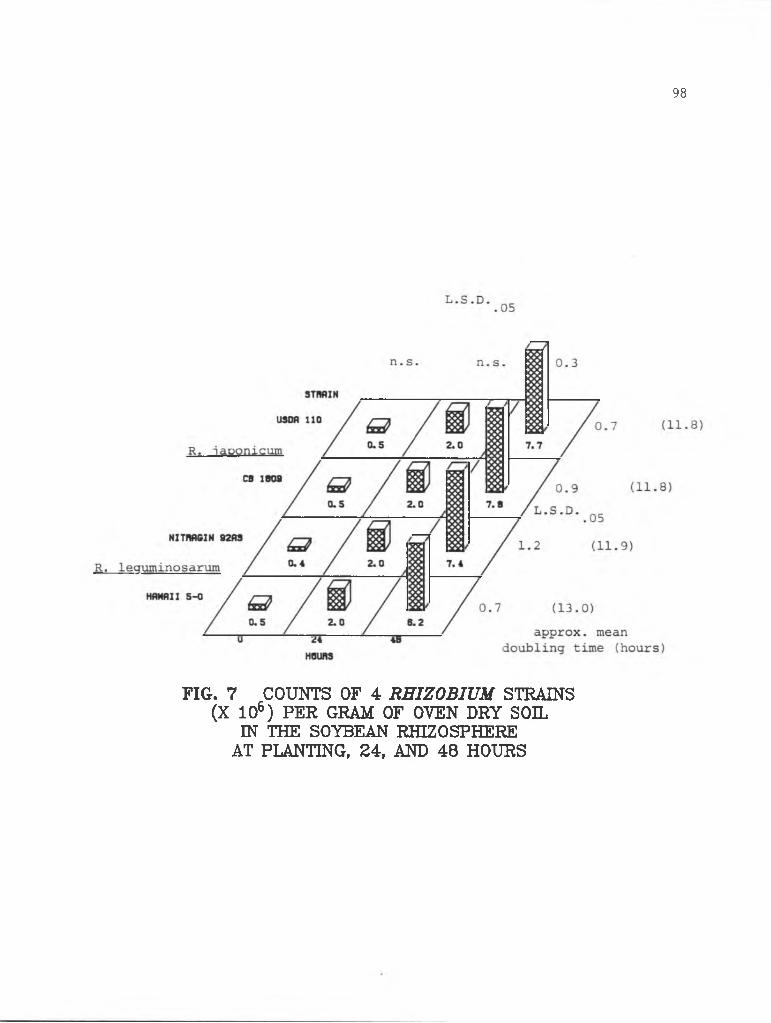
98
FIG. 7 COUNTS OF 4 RHIZOBIUM STRAINS (X 106) PER GRAM OF OVEN DRY SOIL
IN THE SOYBEAN RHIZOSPHERE AT PLANTING, 24, AND 48 HOURS

99
L.S.D. .05
FIG. 8 ACRIDINE ORANGE TOTAL COUNTS OF BACTERIA (X 108) PER GRAM OF OVEN DRY SOIL IN PEA AND SOYBEAN RHIZOSPHERES

Table 1. Percentages of identified Rhizobium in the rhizosphere population of bacteria at 3, 5,
7, and 9 days
Time from planting (days) 3 5 7 9
Rhizosphere
soybean 12 6 5 5
pea 12 10 5 7
corn 9 3 3 4
non-rhizosphere 5 4 4 4soil

101
Table 2. Percentages of identified Rhizobium in the rhizosphere population of bacteria at
planting, 24, and 48 hours
Time from planting (hours) 0 24 48
Rhizosphere
soybean 1.0 1.3 5.6
pea 0.8 2.0 8.0

DISCUSSION
Specific stimulation in the rhizosphere is proposed as a possible
contributor to specificity in the Rhizobium-legume symbiosis (2,4,9).
In two earlier studies (Chapters 2 and 3), population dynamics of two
R. japonicum and two R. leguminosarum strains was followed in host and
non-host rhizospheres. I sampled rhizosphere populations at 35 days
in one experiment and at weekly intervals during one 35-day growth
cycle in another. In these studies, as well as those of Reyes and
Schmidt (5,6) on the soybean R. japonicum strain 123 association, no
specific stimulation was detected in the rhizosphere.
Rhizosphere numbers of rhizobia were in the following order:
soybean>pea>corn at 3, 5, and 7 days, although differences between
rhizosphere populations between crops were not always significant
(Figs. 1 to 3). At 9 days, pea rhizosphere numbers of all strains
significantly surpassed those for soybean, with the exception of
strain Nitragin 92A3 (Fig. 4). Rhizobia per gram of soybean rhizo
sphere soil declined over the four harvests (Figs. 1 to 4). In the
pea rhizosphere rhizobia declined for the first week (Figs. 1 to 3),
then increased slightly at 9 days (Fig. 4). Numbers of Rhizobium per
gram of corn rhizosphere soil were rarely significantly different from
numbers in non-rhizosphere soil at 3, 5, 7, and 9 days. As was seen
in Chapters 2 and 3, numbers of the two R. japonicum strains were
nearly equal in any particular rhizosphere at any given harvest.
Rhizosphere numbers of Hawaii 5-0 were sometimes significantly lower
than numbers of the other three inoculant strains, but differences
were usually slight, less than the two-fold differences seen in

Chapter 2 (Figs. 1 to 4). Apparently the competitive advantage of the
other strains over Hawaii 5-0 was not as great during early development
of the rhizosphere.
In the growth rate study, rhizobia responded rapidly to the condi
tions of the rhizosphere, and numbers of each strain increased about
1 1/2 logs in the first 48 hours (Figs. 6 and 7). Growth rates in the
early rhizosphere were roughly equal for both the fast-growing (R.
leguminosarum) and the slow-growing (R. japonicum) strains. This
suggests that there might be limiting nutrients or growth factors
which restrict the growth of rhizobia, regardless of their growth rate
in broth culture.
The soybean rhizosphere did not specifically stimulate the growth
of R. j aponicum, nor did the pea rhizosphere specifically stimulate R.
leguminosarum (Figs. 1 to 4, 6 and 7). In the 9-day time course, as
was seen previously (Chapter 3) numbers of all inoculant strains of
rhizobia in all rhizospheres were highest at the first time point (3
and 7 days for the two studies, respectively). At 3 days rhizobia
comprised their highest percentages of the rhizosphere population of
bacteria, relative to acridine orange total counts (Tables 1 and 2).
It appears that rhizobia responded to the conditions of the early
rhizosphere with a burst of rapid growth. Rhizobia multiplied rapidly
in the developing seedling rhizosphere, and reached peak populations,
on the basis of soil weight, even before the emergence of shoots from
the soil. Thus, the early events in the rhizosphere may play an
important role in the nodulation of legumes, as proposed by Kosslak,
Dowdle, Sadowsky, and Bohlool (8th North American Rhizobium Conference,
103

1981. Abstracts). However, patterns of growth do not reflect
selective stimulation of rhizobia by their homologous hosts (Figs. 1 to
4, 6 and 7). Soybean and pea rhizospheres stimulated the growth of
rhizobia more strongly than the corn rhizosphere during the first nine
days of growth (Fig. 1). This was also the case for bacteria in
general (Fig. 5).
Rhizobial numbers declined after this initial burst of growth,
probably because the plant root systems expanded and enmeshed soil
faster than rhizobia could colonize it.
These data support the work of Reyes and Schmidt (5,6), who
examined the growth of R. japonicum strain 123, a highly competitive
soybean strain, both on the basis of rhizosphere soil weight (5) and on
the basis of root surface area (6). They found no evidence to support
the concept of specific stimulation as a possible contributor to
specificity. In the first study (5), rhizosphere numbers of strain 123
per gram of soil peaked at 21 days, and declined at 28 days. In the
second experiment (6), they observed a decline in numbers of strain 123
in the soybean rhizosphere between days 16 and 30, on the basis of root
area. Reyes and Schmidt (6) postulated that the rhizosphere population
of strain 123 is unable to keep up with the rapidly-expanding soybean
root system.
It is possible that specific stimulation might have occured at
microsites along the plant root system. If this were the case, the
rhizosphere sampling and enumeration methodology such as the one used
for these studies might be inadequate to detect the stimulation. This
highly, localized stimulation seems unlikely for the case of the soybean
rhizosphere, however. Reyes and Schmidt (6) examined soil adhering
104

to unwashed soybean roots and found populations of two strains of R.
j aponicum to be uniform and low (6).
Acridine orange total counts were relatively constant throughout
the 9-day time course (Fig. 5). An early burst of growth of bacteria
was observed in the first 48 hours. Rhizobia grew more rapidly than
the other soil bacteria, but because of the high initial numbers of
other bacteria, they always comprised the vast majority of the rhizo
sphere population (Tables 1 and 2). This early burst of growth was
not specific with respect to legume host or Rhizobium species.
However, this early burst of growth in the rhizosphere might be a
factor in ensuring the successful establishment of the Rhizobium-
legume symbiosis.
105

LITERATURE CITED
Broughton, W. J. 1978. Control of specificity in legume- Rhizobium associations. J. Appl. Bacteriol. 45:165-194.
Brown, Margaret E., R. M. Jackson, and S. K. Burlingham. 1968. Growth effects of bacteria introduced into soil. In T. R. G. Gray and D. Parkinson (Eds.) The ecology of soil bacteria. Liverpool Univ. Press, Liverpool, pp. 531-551.
Kingsley, Mark T. and B. B. Bohlool. 1981. Release of Rhizobium spp. from tropical soils and recovery for immunofluorescence enumeration. Appl. Environ. Micro 42:241-248.
Nutman, P. S. 1965. The relation between nodule bacteria in the rhizosphere and in the process of infection. In K F. Baker and W. C. Snyder (Eds.) The ecology of soil-borne plant pathogens, pp. 231-247. Univ. Calif. Press, Berkeley.
Reyes, V. G. and E. L. Schmidt. 1979. Population densities of Rhizobium japonicum strain 123 estimated directly in soil and rhizospheres. Appl. Environ. Micro. 37:854-858.
Reyes, V. G. and E. L. Schmidt. 1981. Populations Rhizobium japonicum associated with the surfaces of soil-grown roots. Plant and Soil 61:71-80.
Schmidt, E. L. 1978. Ecology of the legume root nodule bacteria,pp. 269-303. In Y. R. Dommergues and S. V. Krupa (Eds.) Interactions between non-pathogenic soil microorganisms and plants. Elsevier Scientific Publishing Co., Amsterdam.
Vincent, J. M. 1970. A manual for the practical study of theroot nodule bacteria. International Biological Programme Handbookno. 15. Blackwell Scientific Publications, Oxford, England.
Vincent, J. M. 1974. Root nodule symbiosis with Rhizobium. pp. 265-431. In Quispel (Ed.) The biology of nitrogen fixation. North Holland Publishing Co., Amsterdam.

CHAPTER 5
GENERAL DISCUSSION

GENERAL DISCUSSION
From these data it would appear that specific stimulation in the
rhizospheres of peas and soybeans of their homologous Rhizobium species
was not seen under the conditions of these studies. Bacteria, including
rhizobia, were generally more strongly stimulated by the soybean
rhizosphere than the pea rhizosphere. The rhizosphere of sweetcorn
harbored more bacteria than fallow soil, but less bacteria than the
legume rhizospheres.
All strains grew rapidly in the early periods of rhizosphere
development. Numbers peaked at about 3 days in the pea and soybean
rhizospheres, and declined thereafter as the plant root systems expanded
rapidly. Nodulation patterns reflected rhizosphere populations of the
Rhizobium strains, but rhizosphere differences alone could not account
for the differences in nodule occupancy.
Rhizosphere populations of Rhizobium reflected more strain differ
ences than species differences. Perhaps rhizobia colonize rhizospheres
without regard to plant species, and some strains are better rhizo
sphere colonizers than others.
Curiously, rhizosphere populations of each strain never got above
about 2 - 3 x 107 cells per gram of soil even when soil was continuously
cropped with one plant species. Thus, it seems as though the rhizosphere
might have a limited carrying capacity for Rhizobium.
The rapid early growth of Rhizobium in the rhizosphere was
surprising. Although the growth of all inoculant strains was similar in
the early rhizosphere in these experiments, this might not be the case
for all strains in all rhizospheres.

Further work should concentrate on the documentation of the rapid
growth of rhizobia in the early rhizosphere. It seems unlikely that
this early growth would comprise a mechanism for host-bacterium
specificity, but perhaps the competitive advantage of some strains over
others is a result of rapid early growth in the rhizosphere.
The basis for specificity in the Rhizobium-legume association
remains uncertain. In these studies, specific stimulation of Rhizobium
species in the rhizospheres of their homologous host plants was not
observed.
109


















