
Platelet activating factors (AGEPC) from epidermal secretions of the Arabian Gulf catfish, Arius...
8
Biochimica et Biophysica Acta, 1091 (1991) 329-336 © 1991 ElsevierScience Publishers B.V. 0167-4889/91/$03.50 ADONIS 016748899100100Z 329 BBAMCR 12868 Platelet activating factors (AGEPC) from epidermal secretions of the Arabian Gulf catfish, Arius bilineatus, which stimulate wound healing Brent Summers 2, Jassim M. A1-Hassan 1, Martha Thomson 1, Alan Chun 2 and R.S. Criddle 2 t Department of Biochemistry, Faculty of Science, Kuwait University, Safat (KuwaiO and "Dep,~rtment of Biochemistry and Biophysics, University of California, Davis, CA (U.S.A.) (Received 29 June 1990) Key words: Plateletactivatingfactor; Catfish secretion; Wound healing High levels of platelet activating factor (PAF) activity were demonstrated by platelet aggregation and serotonin release assays to be present in fright induced epidermal secretions of the Arabian Gulf catfish, A rius biTineatus (Valenciennes, 1840). The PAF activity was purified by thin-layer chromatography. Mass spectral analysis combined with chemical and enzymatic modification of the purified PAF and inhibitor studies indicated that PAF activity was due to the presence of l.O.alkyi.2.acetyi-sn-glycero-3-phosphorylcholine (AGEPC) molecules. The total AGEPC concentration in the epider- mal secretions based on PAF assays was 8.10 .8 M, well above the threshold level for platelet activation which is near 5-10 -n M. Thus, stimulated epidermal secretory cells of Arius bilineatus supply platelet activating molecules at physiologically high concentrations to sites of injury. Introduction Secretions, elaborated largely from unicellular glands termed 'club cells', of the Arabian Gulf catfish, Arius bilineatus (Valenciennes, 1840), contain a mixture of enzymatically and pharmacologically active proteins and lipids [1,2]. Preparations from these secretions stimulate wound healing rates in test animals [3]. At times of shock or injury to the catfish, the club cells are rup- tured, and their contents released to the skin surface. This secretion, together with some additional epidermal components, forms a viscous skin coating which has been the subject of our previous studies [4]. We have recently demonstrated that there are four Ariid catfish present in the northern Arabian Gulf [5]. The most abundant species is Arius bilineatus, whose secretions have been the subject of our previous studies. This species has been consistantly misidentified by area Abbreviations: PAF, platelet activating factor; AGEPC, 1-O-alkyl-2- acetyl-sn-glycero-3-phosphorylcholines; TLC, thin-layer chromatog- raphy; FAB,fast atom bombardment; PDGF, platelet derived growth factor. Correspondence: R.S. Criddle, Department of Biochemistryand Bio- physics, Universityof California, Davis, CA, 95616, U.S.A. fish taxonomists as Arius thalassinus [5]. Consequently, all of our previous studies have utilized their incorrect nomenclature designating A rius bilineatus as A rius thalassinus. A. thalassinus is actually quite rare in Kuwaiti waters. The viscous skin secretions from Arius bilineatus are shown here to contain lipid components that stimu- late platelet activation. This results from the presence of 1-O-alkyl-2-acetyl-sn-glycero-3-phosphorylcholine (AGEPC) in the secretion at levels near 8.10 -8 M. AGEPC in skin secretions of catfish comes into contact with wound areas following injury to the fish and can therefore play a role in wound responses. AGEPC stimulates a wide range of responses which play important roles in the physiological processes accompanying the healing of wounds [6]. For example, AGEPC is a mediator of inflammation, has anti-hyper- tensive activity, induces contraction of smooth muscle, and is a potent inducer of platelet shape change, aggre- gation and secretion [7-9]. it is present in various mammalian blood cells [9]. Evidence indicates that there are specific receptor sites on platelets for this lipid and that occupation of these sites by AGEPC results in the stimulation of many coupled responses by platelets [10- 12]. Though a complete understanding of the mechanisms of AGEPC action on platelets remains to be elucidated,
-
Upload
brent-summers -
Category
Documents
-
view
216 -
download
0
Transcript of Platelet activating factors (AGEPC) from epidermal secretions of the Arabian Gulf catfish, Arius...

Biochimica et Biophysica Acta, 1091 (1991) 329-336 © 1991 Elsevier Science Publishers B.V. 0167-4889/91/$03.50 ADONIS 016748899100100Z
329
BBAMCR 12868
Platelet activating factors (AGEPC) from epidermal secretions of the Arabian Gulf catfish, Arius bilineatus, which stimulate
wound healing
Brent Summers 2, Jassim M. A1-Hassan 1, Martha Thomson 1, Alan Chun 2 and R.S. Criddle 2
t Department of Biochemistry, Faculty of Science, Kuwait University, Safat (KuwaiO and "Dep,~rtment of Biochemistry and Biophysics, University of California, Davis, CA (U.S.A.)
(Received 29 June 1990)
Key words: Platelet activating factor; Catfish secretion; Wound healing
High levels of platelet activating factor (PAF) activity were demonstrated by platelet aggregation and serotonin release assays to be present in fright induced epidermal secretions of the Arabian Gulf catfish, A rius biTineatus (Valenciennes, 1840). The PAF activity was purified by thin-layer chromatography. Mass spectral analysis combined with chemical and enzymatic modification of the purified PAF and inhibitor studies indicated that PAF activity was due to the presence of l.O.alkyi.2.acetyi-sn-glycero-3-phosphorylcholine (AGEPC) molecules. The total AGEPC concentration in the epider- mal secretions based on PAF assays was 8 . 1 0 .8 M, well above the threshold level for platelet activation which is near 5 -10 - n M. Thus, stimulated epidermal secretory cells of Arius bilineatus supply platelet activating molecules at physiologically high concentrations to sites of injury.
Introduction
Secretions, elaborated largely from unicellular glands termed 'club cells', of the Arabian Gulf catfish, Arius bilineatus (Valenciennes, 1840), contain a mixture of enzymatically and pharmacologically active proteins and lipids [1,2]. Preparations from these secretions stimulate wound healing rates in test animals [3]. At times of shock or injury to the catfish, the club cells are rup- tured, and their contents released to the skin surface. This secretion, together with some additional epidermal components, forms a viscous skin coating which has been the subject of our previous studies [4].
We have recently demonstrated that there are four Ari id catfish present in the northern Arabian Gulf [5]. The most abundant species is Arius bilineatus, whose secretions have been the subject of our previous studies. This species has been consistantly misidentified by area
Abbreviations: PAF, platelet activating factor; AGEPC, 1-O-alkyl-2- acetyl-sn-glycero-3-phosphorylcholines; TLC, thin-layer chromatog- raphy; FAB, fast atom bombardment; PDGF, platelet derived growth factor.
Correspondence: R.S. Criddle, Department of Biochemistry and Bio- physics, University of California, Davis, CA, 95616, U.S.A.
fish taxonomists as Arius thalassinus [5]. Consequently, all of our previous studies have utilized their incorrect nomenclature designating A rius bilineatus as A rius thalassinus. A. thalassinus is actually quite rare in Kuwaiti waters.
The viscous skin secretions from Arius bilineatus are shown here to contain lipid components that stimu- late platelet activation. This results from the presence of 1-O-a lky l -2 -ace ty l - sn -g lycero-3-phosphory lcho l ine (AGEPC) in the secretion at levels near 8 .10 -8 M. AGEPC in skin secretions of catfish comes into contact with wound areas following injury to the fish and can therefore play a role in wound responses.
AGEPC stimulates a wide range of responses which play important roles in the physiological processes accompanying the healing of wounds [6]. For example, AGEPC is a mediator of inflammation, has anti-hyper- tensive activity, induces contraction of smooth muscle, and is a potent inducer of platelet shape change, aggre- gation and secretion [7-9]. it is present in various mammalian blood cells [9]. Evidence indicates that there are specific receptor sites on platelets for this lipid and that occupation of these sites by AGEPC results in the stimulation of many coupled responses by platelets [10- 12].
Though a complete understanding of the mechanisms of AGEPC action on platelets remains to be elucidated,

330
studies increasingly indicate primary effects on metabo- lism of platelet phospholipids [13,14]. AGEPC induces rapid activation of phospholipase C, and subsequent degradation of inositol phospholipids, to form the metabolically active inositol phosphates and di- acylglycerols. Phospholipase A 2 is also activated to catalyze formation of lysophospho!ipids and fatty acids which may be used for the synthesis of prostaglandins, thromboxanes and leukotrienes [13-15].
AGEPC is commonly derived from basophils upon immunological challenge [16,17], and is also released from neutrophils [18], mast cells [19] and platelets [20] following responses to specific stimuli.
AGEPC-induced activation of platelets occurs at concentrations as low as 5 .10 -11 M. AGEPC may possibly be active in the stimulation of platelets at the level of one AGEPC molecule per platelet. Correspond- ingly, AGEPC is present in very low concentrations in human cells and is rapidly metabolized to inactive prod- ucts, making it difficult to directly quantify in vivo levels. The concentration of AGEPC in neutrophils has been estimated using a mass spectrum analytical method to measure the lysophospholipid breakdown products. Values reported were in the range of 20 to 40 mg AGEPC per 50.106 cells [21].
This paper details the characterization of a family of AGEPC molecules from a unique source which is of medical and pharmacological interest.
Materials and Methods
Sample collection. Catfish were caught on hook and line in Kuwaiti coastal waters. The epidermal secretions were collected by gentle scraping with a blade as de- scribed previously [1], were rapidly frozen on dry ice and stored at -80°C.
Fish collected for the initial studies of platelet activating activity in the epidermal preparations were obtained during August to October (summer samples), when temperatures of the coastal waters of Kuwait may reach 37°C. This shallow body of water undergoes large seasonal temperature variations, however, and other samples were collected during April to June when water temperatures range between 15-25°C (winter samples).
Platelet activating factor assays. PAF activities were measured both by aggregation, using the method of Born and Cross [22] with measurement of light-scatter- ing at 600 nm in a stirred cuvette of a Farrand Model MK1 fluorometer, and by the serotonin release assay described by Pinckard et al. [16]. Both assays were calibrated relative to the activity of authentic AGEPC purchased from Calbiochem.
Purification of PAF. PAF was isolated by thin-layer chromatography (TLC) using the methods of Pinckard et al. [16] and Satouchi et al. [23], with the use of one additional solvent system.
In a typical PAF preparation, A. bilineatus epider- mal secretions from about nine fish (21 ml containing about 12 mg total lipid) were extracted by homogeniza- tion of the thawed secretions with methanol (399 ml) at 0°C, in a Sorvall omni-mixer at full speed for 2 min. The homogenate was centrifuged to remove the solids (10000 ×g , 15 rnin). Chloroform (399 ml) and water (336 ml) were added to promote phase separation, and the chloroform-rich phase was dried under nitrogen. The residue was redissolved in chloroform/methanol (2:1) and purified by four successive TLC steps using 1000 micron prep-scored Analtech silica gel G plates. Following each TLC step, the margins of each plate were broken off and stained with iodine to visualize the lipids and standard AGEPC (Calbiochem). Fractions corresponding to each major lipid component, and to spaces between components, were scraped from the plates and the silica gel of each was twice eluted with chloroform/methanol/water (10 : 20 : 8) (total of 6 ml per plate) to recover samples for assay. Phase separation was promoted by the addition of chloroform (3 ml per plate), and water (3 ml per plate). The lower phase was retained and washed twice with methanol/water (10: 9) (6 ml per plate). The lower phase was dried under nitrogen and resuspended in chloroform/methanol (2 : 1) for the next step of purification or analysis. This first TLC step using a chloroform/methanol/water (65 : 35 : 6) solvent system (23 plates) separated the bulk of the lipids from PAF activity [16].
Active PAF fractions were chromatographed using methanol/dimethylsulfoxide (82:18) (ten plates) as a solvent system. This step separated most of the remain- ing contaminating lipids from PAF active fractions. TLC of active fractions was once again run (five plates) using chloroform/methanol/water (65 : 35:6). The low total lipid loading levels at this step allowed resolution of the components not initially possible with this solvent system during removal of the bulk lipids. The fourth chromatography step employed active fractions from step three on the low capacity, but high AGEPC resolv- ing methanol/water (2: 1) system of Satouchi [23] (two plates).
Recoveries of PAF at each step of purification were determined by addition of defined quantities of AGEPC to aliquots of each sample preparation. The recoveries of activity from the AGEPC augmented aliquots at each separation and recovery step were then used to quanti- rate PAF recovery for each sample.
Mass spectroscopy. Fast atom bombardment (FAB) mass spectrometry studies were performed using a ZAB-HS-2F mass spectrometer (VG Analytical). Ioni- zation of the sample was achieved by a 8 keV xenon atom bombardment using a thioglycerol matrix to pro- duce positive ions. Tandem mass spectrometry (MS-MS) on catfish epidermal PAF preparations have been pub- lished elsewhere [24].
Chemical deacylation. Studies of PAF activity and structure were carried out following deacylation of sam- ples using the methods of Hanahan et al. [17]. Reacyla- tion was carded out using acetic anhydride and the methods of Demopolous et al. [7]. Authentic PAF pre- parations were carried through the deacylation, reacyla- tion and purification steps to estimate recovery of sero- tonin release activity after each procedure.
Enzymic deacylation. Phospholipase A 2 (Sigma) was employed in the 'ether-buffer' system of Okuyama and Nojima [25] to catalyze hydrolysis of acyl groups in the 2-position of molecules with PAF activity. Rhizopus arrhizus lipase (Sigma) was employed as described by Mencia-Huerta et al. [19], as a test for acyl moieties at the 1-position.
Inhibition by antagonists. The specific competitive inhibitors of PAF action, CV3899 (1-O-hexadecyl- 2(lyso)-sn-glycerol-3-phosphocholine) 26] from Takeda Chemical, Japan and L652,731 (trans-2,5-bis(3,4,5-tri- methoxyphenyl) tetrahydrofuran) [27] from Merck, Sharp and Dohme, U.S.A., were suspended in ethanol and added to platelet suspensions (final concentration of ethanol was 0.570) 1 min prior to PAF addition in serotonin release assays, to determine their effects on catfish PAF activity. Inhibitory responses were com- pared with responses upon treatment of authentic AGEPC. Control reactions without inhibitors were car- ried out with 0.570 ethanol present.
Results
Results of assays for PAF activity in the methanolic extracts of Arius bilineatus epidermal secretions, and in the major subfractions produced during each step of the preparative procedures are presented in Table I. At each step of the purification, PAF activity was well resolved from the bulk of the lipid contaminants, allowing the majority of PAF activity to be retained. Catfish PAF activity comigrated with standard AGEPC in each of the three TLC chromatography systems. Based on the level of purified PAF require~i to stimulate 5070 sero-
TABLE !
Purification of PAF by TLC
Step Total Yield from Removed PAF by previous step for assay a analysis (nmol) (nmol)
First TLC 1.11 assumed to be 100~ 0.13
Second TLC 1.04 106~ 0.12 Third TLC 0.71 77~ 0.08 Fourth TLC 0.22 35~ -
a From 21 ml of epidermal preparation which contains about 2/3 epidermal secretions mixed with 1/3 sea water.
331
. m
I m . p ,
Time (minutesl
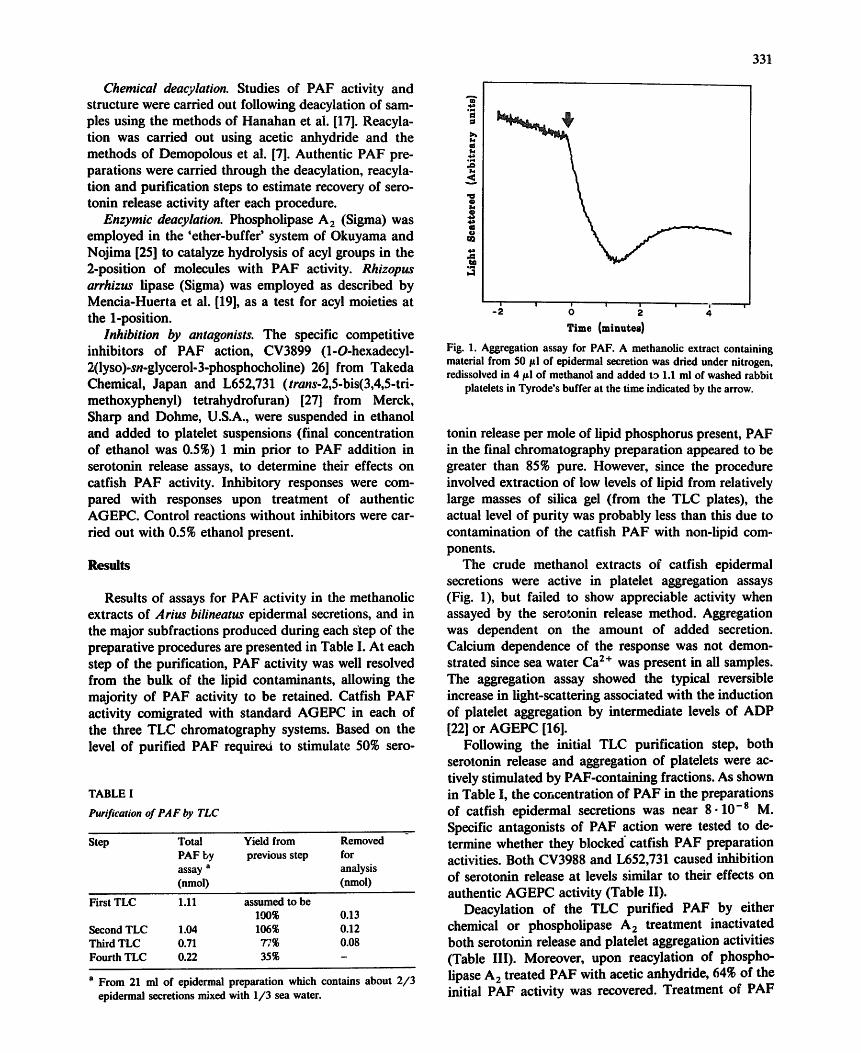
Fig. 1. Aggregation assay for PAF. A methanolic extract containing material from 50/tl of epidermal secretion was dried under nitrogen, redissolved in 4/tl of methanol and added ta 1.1 ml of washed rabbit
platelets in Tyrode's buffer at the time indicated by the arrow.
tonin release per mole of lipid phosphorus present, PAF in the final chromatography preparation appeared to be greater than 8570 pure. However, since the procedure involved extraction of low levels of lipid from relatively large masses of silica gel (from the TLC plates), the actual level of purity was probably less than this due to contamination of the catfish PAF with non-lipid com- ponents.
The crude methanol extracts of catfish epidermal secretions were active in platelet aggregation assays (Fig. 1), but failed to show appreciable activity when assayed by the serotonin release method. Aggregation was dependent on the amount of added secretion. Calcium dependence of the response was not demon- strated since sea water Ca z+ was present in all samples. The aggregation assay showed the typical reversible increase in light-scattering associated with the induction of platelet aggregation by intermediate levels of ADP [22] or AGEPC [16].
Following the initial TLC purification step, both serotonin release and aggregation of platelets were ac- tively stimulated by PAF-containing fractions. As shown in Table I, the concentration of PAF in the preparations of catfish epidermal secretions was near 8.10 -8 M. Specific antagonists of PAF action were tested to de- termine whether they blocked catfish PAF preparation activities. Both CV3988 and L652,731 caused inhibition of serotonin release at levels similar to their effects on authentic AGEPC activity (Table If).
Deacylation of the TLC purified PAF by either chemical or phospholipase A2 treatment inactivated both serotonin release and platelet aggregation activities (Table III). Moreover, upon reacylation of phospho- iipase A 2 treated PAF with acetic anhydride, 6470 of the initial PAF activity was recovered. Treatment of PAF

332
TABLE II
Effects of PAF Antagonists on activity of catfish PAF preparations
Test material Activity Antagonist (M) Activity a + antagonist
AGEPC, 5-10-1o M 100% b
Catfish PAF preparation c 84% d
CV3988 (5-10-'P) 4% 1,652,731 (10 -6 ) 11%
CV3988 (1-10 -~) 37% CV3988 (5-10-~) 7% 1,652,731 (10 -6 ) 9%
' Average of two determinations. b By definition. Assays by the method of Pinckard et al, [16], c Prepared through the third TLC step (Table 1). d Relative to corresponding value for 5.10-10 M AGEPC.
preparations with Type XI Rhizopus arrhizus lipase had no appreciable effect on activity.
While partially purified fractions showed serotonin release activity, the apparent absence of PAF-serotonin release activity in the initial methanolic extracts of catfish epidermat secretions suggested the presence of an inhibitor capable of blocking PAF-induced secretion of serotonin, but not platelet aggregation. Table IV summarizes evidence for the presence of inhibitors.
TABLE 111
Effects of chemical reactions on serotonin release catalyzed by catfish epidermal PAF preparations
Treatment of purified ~ Decrease in serotonin catfish PAF release activity a
None 0
PLA 2 treatment b 84 Chemical deacylation 100 PLA 2 treatment and reacylation 20 R, arrhizm treatment 7
Serotonin release assays were performed as described by Pinckard et at, ll$l,
b P L A 2 - Phospholipase A2.
Lipids extracted from the total catfish epidermal secre- tions had no serotonin release activity. When these lipids were fractionated by successive thin-layer chro- matography steps and recoveries of PAF-serotonin re- lease activity were quantitated, activity increased with each purification step. Following correction for absolute losses inherent in the purification procedures with the aid of internal standards, an increase of 240~ in sero- tonin release activity was observed from the first to the third chromatography steps.
Serotonin release stimulated by authentic PAF (3- 10-10 M) was also inhibited by components in the total catfish lipid extract. Inhibition was assayed by addition of potential inhibitory fractions from TLC separations to test solutions of authentic PAF followed by measure- ment of residual PAF activities. Fractions with R F values from 0.45 to 0.54 and from 0.92 to 1.0 in the first solvent system were strongly inhibitory. Thus, it ap- pears that at least one relatively non-polar inhibitor and one migrating with an R F near that of phosphatidyl choline are present in the crude lipid extract from catfish epidermal secretions.
In contrast to studies with lipid preparations from the total secretions, lipid extracts from a soluble protein fraction prepared from catfish epidermal secretions which were homogenized and centrifuged prior to freez- ing, were fully active in both platelet aggregation and serotonin release without further purification. Thus, the inhibitory compounds appear either to be bound to the insoluble proteins of the secretions or to be produced during the freezing and storage process.
Purified fractions from A. bilineatus epidermal secre- tions with PAF activity were examined by mass spec- trometry to determine whether activity was due to AGEPC. Unambiguous evidence has been obtained by tandem mass spectrometry [24] for the presence of AGEPC in the partially purified PAF preparations. Additionally, FAB mass spectroscopy provided infor- mation on ~ family of molecules in the preparations with m/z characteristics of AGEPC as shown in Fig. 2.
TABLE IV
Inhibition of PAF activity by inhibitors in epidermal preparation
Fraction Serotonin release activity (%)
Total lipid extract of epidermal secretion Recovery from 1st TLC step Remvery from 2rid TLC step Rc~ove.ry from 3rd TLC step 3 ,10- to M PAF 3-10 -1° M PAF+ total lipid extract from 200/zl epidermal secretion 3,10 - l ° M PAF+ fractions from TLC system No. 1 with R F -- 0.45 to 0.54 3-I0 - t ° M PAF+ fractions from TLC system No. 1 with R F -- 0.92 to 1.0
0 100% a 144% b
240% b 100% a
20% c 12~ 12~
" By definition. b Relative to activity recovered from the first TLC step. © Relative to the activity of 3-10 - t ° M pure PAF.

333
,OOll ] ,oo. .~.80 . . . 8O-
® eo- as
4o
< J , I ,J g < 20 ~ 20
0 0 ~ i
I 0 0
o K )
u ' ) i f )
3(~0 ' 5()0 ' 700 400 500 m / s m/z
100"
8O
• 60 ~J
i:I
q:l ¢I 4O =i ,.Q
.<
2O
B ~
i i i i
50O l l | i i
6O0
100"
80-
v
• 60 ii1
C q" m w
al "~ 40 I:1 :1 ,Q <
2O
o L, . . . . . I I I
I00 3C9 500 1 '00 m / ~ .
Fig. 2. FAB mass spectra of (A) purified catfish PAF, (B) the expanded region of purified catfish PAF containing the PAF molecu- lar ions and (C) standard PAF from Calbiochem. The base ion peak at m/z 184 is characteristic of the phosphorylcholine moiety. Signals at m/z 552 and 524 in the standard correspond to C18 and C16
derivatives of PAF.
The spectrum in Fig. 2A shows that the PAF prepara- tion from catfish epidermal secretions is by no means pure, but that a base peak occurs at m/z 184 and a family of homologous peaks differing in 14 mass units
m
i t ) to
i
600
Fig. 3. FAB mass spectt~ m of purified PAF from fish caught in early spring (cold water adapte~ The high m/z region of the spectrum is presented, showing a family of molecular ions with m/z 2 mass units less than corresponding peaks in warm water fish (compare Fig. 2B).
is present in the range 510 to 636. Fig. 2B presents the region from m/z 460 to 640 in an expanded form. The FAB-MS spectrum of standard AGEPC (Calbiochem) is shown in Fig. 2C. The standard preparation con- tained both octadecyl (m/z 552) and hexadecyl (m/z 524) alkyl chains. The base peak at m/z 184 in both the sample and standard spectra was a phosphorylcholine moiety.
The results in Fig. 2B are consistent with a family of AGEPC molecules in the catfish epidermal secretions. A form with a C18 alkyl chain (m/z 552) appears most abundant, while C15, C16, C17, C19 and C20 homologs were all present in significant quantities.
Samples of epidermal secretions collected from catfish adapted to cold winter waters exhibited total levels of PAF activity in serotonin release assays com- parable to those described above. However, PAF activ-
40
Winter
Summer 3O
~: 2o q:l
, Q <
10
i
16 17 18 19 20 Alkyl Chain Length
Fig. 4. The distribution of alkyl chain lengths from summer PAF and winter PAF.

334 100
80,
I ) g 60,
40,
20,
o . L . , . . Jllhl-Jl -.Ju. 4OO
,i[ • 50O
l.,l.,..,h., .h 600
m/z Fig. 5, FAB mass spectrum of a chemically deacylated catfish PAF preparation. A family of molecular ions with masses 42 units less than
the corresponding molecular ions of untreated PAF is shown.
ity migrated with slightly higher mobilities on TLC than the previously studied summer samples and standard AGEPC. Examination of PAF structures in this pre- paration by FAB mass spectrometry again showed con- siderable impurities, but revealed a homologous family of molecules in the high mass region of the spectrum. m / z values in this case were two mass units lower than the values observed for the summer preparation (Fig. 3). These molecules were presumed to correspond to the mono-unsaturated forms of the summer PAF molecules. This interpretation would be consistent with the in- creased mobility of PAF activity on TLC. The distribu- tions of alkyl carbon chain lengths in the preparations of PAF from summer and from winter samples were compared based on relative signal heights on FAB spectra (Fig. 4). The overall distribution of alkyl chain lengths among summer (saturated)and winter (un- saturated) chains was not greatly different.
Base-hydmlyzed PAF preparations were examined by FAB mass spectrometry. A family of molecules was found in the high mass region of the spectrum which differed in mass from the molecules in the starting material by 42 mass units (Fig. 5).
Discussion
The existence of platelet activation activity in Arius bilineatus epidermal secretions has been established. PAF activity levels measured in the secretion were more than 1000-times in excess of threshold levels for stimu- lation of the activation of platelets. This PAF activity must be either constantly present at high levels in the secretory epidermal club cells, or be rapidly synthesized following a shock induced stimulus. In either case, these molecules must encounter rather stable conditions within the secretions to remain unmetabolized for at least the 10 to 15 min required to harvest the fish, collect the secretions, then freeze and eventually thaw
the sample. It appears that molecules causing PAF activity in the A. bilineatus secretion are not in contact with appreciable levels of enzymes which could cause degradation.
The initial evidence that the A. bilineatus secretions contained factors which could activate platelets was stimulation of platelet aggregation (Fig. 1). Since the crude catfish secretion was inactive in stimulation of serotonin release, it appeared that either the catfish PAF was different from AGEPC, or that endogenous molecules which interfered with certain AGEPC-in- duced activities were present. A test for endogenous 'inhibitors' of AGEPC action revealed the presence of factors which inhibited serotonin release. These factors which blocked serotonin release, but not platelet aggre- gation, do not correspond to the acid labile, proteina- ceous AGEPC-degrading inhibitors previously de- scribed in rabbit plasma [16]. Rather, they are methanol extractable compounds which modify the responsive- ness of platelets to both catfish epidermal PAF prepara- tions and authentic AGEPC. These compounds have not been studied further.
Chemical and physical evidence that the A. bilineatus platelet activating factor activity may be due to the presence of AGEPC is summarized: (i) the active material co-migrated with authentic AGEPC in three separate high-resolution TLC solvent systems optimized to separate AGEPC from other lipids; (ii) isolated PAF preparations, though not completely pure, gave FAB-MS spectra having molecular ions and major fragments which had masses corresponding to those of authentic AGEPC. The molecular ions from ,n /z 510 to 636 appeared to correspond to a family of AGEPC homo- logs having alkyl chain lengths from 15 to 24 carbons. The base peak was invariably at m / z 184, indicating the presence of the phosphorylcholine moiety; (iii) base or p h o s p h o l i p a s e A 2 hydrolysis of the purified A. bilin- eatus PAF caused complete inactivation. This is pre- sumed to occur by removal of an essential group esteri- fled at position 2 of glycerol; (iv) following chemical deacylation, the mass of the putative AGEPC molecular ions were reduced by 42 Da, consistent with the loss of a 2-acetyl group; (v) reacylation with acetic acid anhydride restored activity, confirming the required role of an ester moiety for activity; (vi) treatment of the PAF preparation with R. arrhizus lipase had no effect on activity, suggesting the presence of an ether rather than an ester linkage at the 1-position of glycerol; and (vii) specific antagonists of PAF action effectively in- hibited stimulation of serotonin release by catfish PAF.
"thus, the PAF activity purified from A. bilineatus epidermal preparations consisted of a family of glycerol based lipid molecules with R1 alkyl chains of varying lengths, phosphorylcholine moieties, and acetyl ester groups at position 2. These observations and the known highly specific structural requirements for PAF stimu-

lated platelet activation, served to establish that the molecules isolated from the epidermal secretions were 1-O-alkyl-2-acetylglyceroether-3-phosphorylcholine. Moreover, tandem MS studies by Plattner et al. [24] of partially purified epidermal PAF preparations unam- biguously showed the presence of AGEPC.
The apparent change in the degree of alkyl chain saturation between summer and winter samples prob- ably reflects a seasonal difference in the degree of unsaturation of the lipids of the microorganisms which are at the base of the catfish's food chain. Major changes in the lipid compositions of catfish have been demon- strated to occur with changes in diet [28-30]. The presence of PAF preparations with relatively large quantities of alkyl chains with odd numbers of carbon atoms is not unprecedented [31].
The occurrence of AGEPC in catfish epidermal secretions poses the question of what biological func- tion(s) utilize high levels of AGEPC in a fright induced secretion which coats the exterior of the fish. The best current hypothesis for a biological role for AGEPC comes from consideration of other activities known to be present in the secretions. The secretions contain a mixture of components with activities which may in- fluence the rates of wound healing responses [4]. Per- haps the high concentrations of AGEPC which are present could aid in this process. A role for AGEPC in wound healing has been previously suggested by S'eppa et al. [6].
The precise roles which AGEPC could play in wound healing remain uncertain. However, secretion of high levels of AGEPC from A. bilineatu~" epidermal cells as a fright induced response seems more than coincidental. It is well known that cells migrate into a wound area in defined sequences [32], and that their migration may be influenced by AGEPC. It is generally thought that the phagocytic cells are attracted to materials present in the wound [33-35]. Fibroblasts, which arrive somewhat later, exhibit a specific chemotactic response to platelet derived growth factor (PDGF) [6]. Moreover, PDGF act,~ as a potent mitogen in stimulating fibroblast pro- liferation [36]. Therefore, it is reasonable that the high levels of AGEPC present at wound sites in A. bilineatus epidermal tissues could stimulate activation of platelets and elicit secretion of PDGF. The released PDGF in turn may accelerate the rate of wound healing by both chemotactic and mitogenic activities.
Acknowledgements
The authors acknowledge with gratitude the support rendered for our project by His Highness Sheikh Jaber AI Ahmad AI-Sabah 'The Amir of Kuwait'. We also thank Sheikh Nasser Mohammad AI Abroad AI-Sabah 'Minister of Social Affairs and Labour' for his help and support. We wish to express our gratitude to A. Daniel
335
Jones and Kei Miyano at the U.C. Davis Facility for Advanced Instrumentation for their technical assis- tance. The ZAB-HS-2F mass spectrometer (VG Analyti- cal) was purchased in part with NIH funds (Grant No RR01460-01). We also thank Robert Stack and Ron Plattner of the U.S.D.A. Agricultural Research Service for collaborative efforts on tandem-MS spectrometry. We thank Professor S.J. Wakil for his constant help, advice, scientific discussions and purchase of reagents, and Mr. Mohammad AI-Qattan for his help with sam- ple collection. Special thanks are due to Kuwait Foun- dation for the Advancement of Science (Grant No. 86-04-01) and Kuwait University (Grant No. SBO 17) for partial support of this project.
References
! AI-Hassan, J.M., Thomson, M. and Criddle, R.S. (1982) Marine Biology 70, 626-629.
2 AI-Hassan, J.M., Thomson, M., Ali, M., Fayad, S., Elkhawad, A., Thulesius, O. and Criddle, R.S. (1986) Toxicon 24, 1009-1017.
3 AI-Hassan, J.M., Thomson, M. and Criddle, R.S. (1983) Lancet 1-83, 1044.
4 AI-Hassan, J.M., Thomson, M., Aft, M. and Criddle, R.S. (1987) J. Toxicol. Toxin Rev. 6, 1-43.
5 AI-Hassan, J.M., Clayton, D.A., Criddle, R.S. and Thomson, M. (1988) J. Nat. History (British Museum) 22, 473-487.
6 Seppa, H., Grotendorst, G., Seppa, S., Schiffmann, E. and Martin, G.R. (1982) J. Cell Biol. 92, 584-588.
7 Demopoulos, C.A., Pinckard, R.N. and Hanahan, D.J. (1979) J. Biol. Chem. 254, 9355-9358.
8 Benveniste, J., Henson, P.M. and Cochrane, C.G. (1912) J. Exp. Med. 136, 1356-1377.
9 Lee, T.C. and Snyder, F. (1985) in Phospholipids and cellular regulation (Kuo, J.F., ed.), CRC Press, Boca Raton.
10 Valone, F.H., Coles, E., Reinhold, V.R. and Goetzl, E.J. (1982) J. lmmunol. 129, 1637-1641.
11 Valone, F.H. (1984) Immunology 52, 169-174. 12 Nishihira, J., lshibashi, T., lmai, Y. and Muramatsu, T. (1985)
Tohoku J. Exp. Med. 147, 145-152. 13 ShuHa, S.D. and Hanahan, D.J. (1983) Arch. Biochem. Biophys.
227, 626-629. 14 Billah, M.M. and Lapetina, E.G. (1983) Proc. Natl. Aead. Sci.
USA 80, 965-968. 15 Granstrom, E. (1983) Adv. Pain Res. Ther. 5, 605-615. 16 Pinckard, R.N., Farr, R.S. and Hanahan, DJ. (1979) J. lmmunol.
123, 1847-1857. 17 Hanahan, D.J., Demopoulos, C.A., Liehr, J. and Pinckard, R.N.
(1980) J. Biol. Chem. 255, 5514-5516. 18 Clark, P.O., Hanahan, D.J. and Pinckard, R.N. (1980) Biochim.
Biophys. Acta 628, 69-75. 19 Mencia-Huerta, J.M., Lewis, R.A., Razin, E. and Austin, K.F.
(1983) J. Immunol. 131, 2958-2964. 20 Chignard, M., Le Cordie, J.P., Tance, M., Vargaftig, B.B. and
Benveniste, J. (1979) Nature 279, 799-800. 21 Haroldson, P.E., Clay, K.L. at~d Murphy, R.C. (1985) Annual
conference on mass spectroscopy and allied topics 33, 11. 22 Born, G.V.R. and Cross, M.J. (1963) J. Physiol. 168, 178-183. 23 Satouchi, K., Pinckard, R.N. and Hanahan, D.J. (1981) Arch.
Biochem. Biophys. 211,683-688. 24 Piattner, R.D., ~tack, R.J., $umntet~,, B., AI.Hassan, J.M. and
Criddle, R.S. (1988) Org. Mass Spectrometry 23, 834-840. 25 Okuyama, H. and Nojima, S. (1965) J. Biochem. 57, 529-538.

336
26 Terashita~ Z., Tsushima, S., Yoshioka, Y., Nomura, H., Inada, Y. and Nishikawa, K. (1983) Life Sci. 32, 1975-1982.
27 Hwan 8, S.B., Biftu, T., Doebber, M.H., Lain, M.H.T., Wu, M.S. and Shen, T.Y. (1985) Prostaglandins 30, 689.
28 Mustafa, F.A. and Medeiros, D.M. (1986) J. Food Sci. 50, 585-588. 29 Chanmugan, P., Boudreau, M. and Hwang, D.H. (1986)J. Food
Sei. 51, 1556-1557. 30 AI-Hassan, J.M., Afzal, M., Ali, M., Thomson, M., Fatims, T.,
Fayad, S. and Criddle, R.S. (1986) Comp. Biochem. Physiol. 85B, 41--47.
31 Lenihan, D.J., Greenberg, N. and Lee, T.C. (1985) Comp. Bio- chem. Physiol. 81C, 81-86.
32 Ross, R. (1968) Biol. Rev. 43, 51-96. 33 Kay, A.B., Pepper, D.S. and McKenzie, R. (1974) Brit. J. Haema-
tol. 27, 669-677. 34 Kaplan, A.P., Goetzl, E.J. and Austin, K.F. (1973) J. Clin. Invest.
52, 2591-2595. 35 Gallin, J.l. and Kaplan, A.P. (1974) J. hmnunol. 113, 1928-1934. 36 Ross, R. and Vogel, H. (1978) Cell 14, 203-210.


















