
Plasmodium falciparumookinetes require … falciparumookinetes require mosquito midgut chondroitin...
6
Plasmodium falciparum ookinetes require mosquito midgut chondroitin sulfate proteoglycans for cell invasion Rhoel R. Dinglasan* † , Aditi Alaganan*, Anil K. Ghosh*, Akio Saito ‡ , Toin H. van Kuppevelt § , and Marcelo Jacobs-Lorena* *Department of Molecular Microbiology and Immunology, Malaria Research Institute, Johns Hopkins Bloomberg School of Public Health, 615 North Wolfe Street, Baltimore, MD 21205; § Department of Biochemistry, Nijmegen Centre for Molecular Life Sciences, Radboud University Nijmegen Medical Centre, 6500 HB, Nijmegen, The Netherlands; and ‡ Department of Biochemistry, Kinki University School of Medicine, Osaka-Sayama, Osaka 589-8511, Japan Edited by Anthony A. James, University of California, Irvine, CA, and approved August 2, 2007 (received for review July 6, 2007) Malaria transmission entails development of the Plasmodium parasite in its insect vector, the Anopheles mosquito. Parasite invasion of the mosquito midgut is the critical first step and involves adhesion to host epithelial cell ligands. Partial evidence suggests that midgut oligosaccharides are important ligands for parasite adhesion; however, the identity of these glycans remains unknown. We have identified a population of chondroitin glycos- aminoglycans along the apical midgut microvilli of Anopheles gambiae and further demonstrated ookinete recognition of these glycans in vitro. By repressing the expression of the peptide-O- xylosyltransferase homolog of An. gambiae by means of RNA interference, we blocked glycosaminoglycan chain biosynthesis, diminished chondroitin sulfate levels in the adult midgut, and substantially inhibited parasite development. We provide evidence for the in vivo role of chondroitin sulfate proteoglycans in Plas- modium falciparum invasion of the midgut and insight into the molecular mechanisms mediating parasite–mosquito interactions. Anopheles glycosaminoglycans glycosytransferase malaria RNAi M alaria is caused by Plasmodium parasites, among which, Plasmodium falciparum inflicts the severest infection on human populations. The complex parasite life cycle includes developmental stages within both mammals and its obligatory insect vector, the Anopheles mosquito (1). Plasmodium gameto- cytes that are taken up in an infected blood meal transform into ookinetes in the mosquito midgut lumen. Ookinetes then mi- grate to the gut periphery where they are thought to recognize and adhere to midgut ligands. These steps directly precede cell invasion, traversal, and the differentiation of ookinetes into oocysts beneath the basal lamina. Each oocyst produces thou- sands of sporozoites that subsequently invade the mosquito salivary glands and are delivered to a vertebrate host during a succeeding blood meal. Clearly, ookinete invasion of midgut epithelia is the critical step for parasite establishment in the mosquito and, therefore, represents the best paradigm to de- velop novel transmission-blocking strategies (i.e., approaches that prevent parasite passage through the mosquito and there- fore impede the subsequent cascade of secondary infections in humans). Plasmodium ookinete molecules belonging to the Throm- bospondin-related adhesive protein (TRAP) family, which in- cludes the sporozoite surface protein, TRAP, and two ookinete proteins, the circumsporozoite and TRAP-related protein (CTRP) and the von Willebrand Factor A domain protein (WARP), have been demonstrated to bind to the glycosamino- glycan (GAG), heparin, in vitro (2). The major circumsporozoite protein (CSP) and TRAP both recognize heparan sulfate (HS) proteoglycans on the liver sinusoid, a critical step toward the establishment of parasite infection in humans (3, 4). Similarly, ookinete CTRP and WARP gene knockouts were shown to be incapable of invading the mosquito midgut, strongly implying their essential role in mosquito midgut cell invasion (5, 6). The common theme of GAG-ligand targeting by different Plasmo- dium stages suggest that there is conservation in the overall cell invasion strategy that occurs during parasite transit from the invertebrate to the vertebrate host and vice versa (7). However, there is no clear evidence for the existence of heparin or GAGs in the mosquito gut. Partial evidence suggests that negatively charged oligosaccha- rides are present along the apical microvilli of midgut epithelial cells and may act as adhesion ligands for ookinetes, however, the identity of these glycans remain unknown (8). Clearly, the identification and characterization of mosquito ligands is needed to further understand the mechanisms that mediate parasite– vector host interactions. We have now identified one set of mosquito midgut-specific oligosaccharide ligands to be sulfated chondroitin polysaccharides and provide evidence for their essential role in P. falciparum ookinete invasion in vivo. Results Chondroitin Sulfate (CS) Proteoglycans Localize to the Mosquito Midgut Microvilli. To determine whether ookinetes use GAGs as ligands for cell invasion, we first sought to resolve the presence of midgut microvillar GAGs by histochemical analysis. The cationic dye, Alcian blue stained sugar-fed mosquito midguts, thereby confirming the presence of anionic polysaccharides on microvilli apices (Fig. 1 a and b). At pH 1.0, alcian blue does not react with protonated carboxyl groups that may be present on microvilli glycoconjugates but reacts readily with ionized sulfate groups (9). MAb CS-56 (Fig. 1 c and d) and single-chain variable fragment (scFv) antibody IO3H10 staining (Fig. 1 e and f ) of midgut microvilli from blood-fed mosquitoes suggests that these anionic polysaccharides are analogous to CS. Midgut sections from blood-fed mosquitoes stained with scFv RB4EA12 (10) anti-HS antibodies showed basal lamina localization of HS proteoglycans (Fig. 1 g and h) (11). CS- and HS-modified proteoglycans are differentially distributed in polarized cells in mammals, wherein CS proteoglycans predominate in the apical surface and HS proteoglycans along the basal surface (12). CSGAGs are linear heteropolysaccharides of repeating gluco- ronic acid (GlcA) and N-acetylgalactosamine (GalNAc) dissa- Author contributions: R.R.D. designed research; R.R.D., A.A., and A.K.G. performed re- search; R.R.D., A.S., and T.H.v.K. contributed new reagents/analytic tools; R.R.D., A.S., T.H.v.K., and M.J.-L. analyzed data; and R.R.D. and M.J.-L. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Abbreviations: CS, chondroitin sulfate; DS, dermatan sulfate; GAG, glycosaminoglycan; HA, hyaluronic acid; HS, heparan sulfate; IRBC, infected RBC. † To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/cgi/content/full/ 0706340104/DC1. © 2007 by The National Academy of Sciences of the USA 15882–15887 PNAS October 2, 2007 vol. 104 no. 40 www.pnas.orgcgidoi10.1073pnas.0706340104
Transcript of Plasmodium falciparumookinetes require … falciparumookinetes require mosquito midgut chondroitin...

Plasmodium falciparum ookinetes require mosquitomidgut chondroitin sulfate proteoglycans forcell invasionRhoel R. Dinglasan*†, Aditi Alaganan*, Anil K. Ghosh*, Akio Saito‡, Toin H. van Kuppevelt§,and Marcelo Jacobs-Lorena*
*Department of Molecular Microbiology and Immunology, Malaria Research Institute, Johns Hopkins Bloomberg School of Public Health, 615 North WolfeStreet, Baltimore, MD 21205; §Department of Biochemistry, Nijmegen Centre for Molecular Life Sciences, Radboud University Nijmegen Medical Centre,6500 HB, Nijmegen, The Netherlands; and ‡Department of Biochemistry, Kinki University School of Medicine, Osaka-Sayama, Osaka 589-8511, Japan
Edited by Anthony A. James, University of California, Irvine, CA, and approved August 2, 2007 (received for review July 6, 2007)
Malaria transmission entails development of the Plasmodiumparasite in its insect vector, the Anopheles mosquito. Parasiteinvasion of the mosquito midgut is the critical first step andinvolves adhesion to host epithelial cell ligands. Partial evidencesuggests that midgut oligosaccharides are important ligands forparasite adhesion; however, the identity of these glycans remainsunknown. We have identified a population of chondroitin glycos-aminoglycans along the apical midgut microvilli of Anophelesgambiae and further demonstrated ookinete recognition of theseglycans in vitro. By repressing the expression of the peptide-O-xylosyltransferase homolog of An. gambiae by means of RNAinterference, we blocked glycosaminoglycan chain biosynthesis,diminished chondroitin sulfate levels in the adult midgut, andsubstantially inhibited parasite development. We provide evidencefor the in vivo role of chondroitin sulfate proteoglycans in Plas-modium falciparum invasion of the midgut and insight into themolecular mechanisms mediating parasite–mosquito interactions.
Anopheles � glycosaminoglycans � glycosytransferase � malaria � RNAi
Malaria is caused by Plasmodium parasites, among which,Plasmodium falciparum inflicts the severest infection on
human populations. The complex parasite life cycle includesdevelopmental stages within both mammals and its obligatoryinsect vector, the Anopheles mosquito (1). Plasmodium gameto-cytes that are taken up in an infected blood meal transform intoookinetes in the mosquito midgut lumen. Ookinetes then mi-grate to the gut periphery where they are thought to recognizeand adhere to midgut ligands. These steps directly precede cellinvasion, traversal, and the differentiation of ookinetes intooocysts beneath the basal lamina. Each oocyst produces thou-sands of sporozoites that subsequently invade the mosquitosalivary glands and are delivered to a vertebrate host during asucceeding blood meal. Clearly, ookinete invasion of midgutepithelia is the critical step for parasite establishment in themosquito and, therefore, represents the best paradigm to de-velop novel transmission-blocking strategies (i.e., approachesthat prevent parasite passage through the mosquito and there-fore impede the subsequent cascade of secondary infections inhumans).
Plasmodium ookinete molecules belonging to the Throm-bospondin-related adhesive protein (TRAP) family, which in-cludes the sporozoite surface protein, TRAP, and two ookineteproteins, the circumsporozoite and TRAP-related protein(CTRP) and the von Willebrand Factor A domain protein(WARP), have been demonstrated to bind to the glycosamino-glycan (GAG), heparin, in vitro (2). The major circumsporozoiteprotein (CSP) and TRAP both recognize heparan sulfate (HS)proteoglycans on the liver sinusoid, a critical step toward theestablishment of parasite infection in humans (3, 4). Similarly,ookinete CTRP and WARP gene knockouts were shown to beincapable of invading the mosquito midgut, strongly implying
their essential role in mosquito midgut cell invasion (5, 6). Thecommon theme of GAG-ligand targeting by different Plasmo-dium stages suggest that there is conservation in the overall cellinvasion strategy that occurs during parasite transit from theinvertebrate to the vertebrate host and vice versa (7). However,there is no clear evidence for the existence of heparin or GAGsin the mosquito gut.
Partial evidence suggests that negatively charged oligosaccha-rides are present along the apical microvilli of midgut epithelialcells and may act as adhesion ligands for ookinetes, however, theidentity of these glycans remain unknown (8). Clearly, theidentification and characterization of mosquito ligands is neededto further understand the mechanisms that mediate parasite–vector host interactions. We have now identified one set ofmosquito midgut-specific oligosaccharide ligands to be sulfatedchondroitin polysaccharides and provide evidence for theiressential role in P. falciparum ookinete invasion in vivo.
ResultsChondroitin Sulfate (CS) Proteoglycans Localize to the MosquitoMidgut Microvilli. To determine whether ookinetes use GAGs asligands for cell invasion, we first sought to resolve the presenceof midgut microvillar GAGs by histochemical analysis. Thecationic dye, Alcian blue stained sugar-fed mosquito midguts,thereby confirming the presence of anionic polysaccharides onmicrovilli apices (Fig. 1 a and b). At pH 1.0, alcian blue does notreact with protonated carboxyl groups that may be present onmicrovilli glycoconjugates but reacts readily with ionized sulfategroups (9). MAb CS-56 (Fig. 1 c and d) and single-chain variablefragment (scFv) antibody IO3H10 staining (Fig. 1 e and f ) ofmidgut microvilli from blood-fed mosquitoes suggests that theseanionic polysaccharides are analogous to CS. Midgut sectionsfrom blood-fed mosquitoes stained with scFv RB4EA12 (10)anti-HS antibodies showed basal lamina localization of HSproteoglycans (Fig. 1 g and h) (11). CS- and HS-modifiedproteoglycans are differentially distributed in polarized cells inmammals, wherein CS proteoglycans predominate in the apicalsurface and HS proteoglycans along the basal surface (12).CSGAGs are linear heteropolysaccharides of repeating gluco-ronic acid (GlcA) and N-acetylgalactosamine (GalNAc) dissa-
Author contributions: R.R.D. designed research; R.R.D., A.A., and A.K.G. performed re-search; R.R.D., A.S., and T.H.v.K. contributed new reagents/analytic tools; R.R.D., A.S.,T.H.v.K., and M.J.-L. analyzed data; and R.R.D. and M.J.-L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Abbreviations: CS, chondroitin sulfate; DS, dermatan sulfate; GAG, glycosaminoglycan; HA,hyaluronic acid; HS, heparan sulfate; IRBC, infected RBC.
†To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0706340104/DC1.
© 2007 by The National Academy of Sciences of the USA
15882–15887 � PNAS � October 2, 2007 � vol. 104 � no. 40 www.pnas.org�cgi�doi�10.1073�pnas.0706340104

charide units (GlcA�1–3GalNAc) (13). MAb CS-56 recognizesthe monosulfated GlcA-GalNAc (4S) disaccharide ‘‘A-unit’’ andGlcA-GalNAc (6S) ‘‘C-unit,’’ the disulfated GlcA (2S)-GalNAc(6S) disaccharide ‘‘D-unit’’, and octasaccharides thatcontained an internal trisulfated tetrasaccharide ‘‘A-D coreunit’’ composed of GlcA-GalNAc(4S)-GlcA(2S)-GalNAc(6S) inCS chains (Fig. 1i) (13). ScFv IO3H10 specifically recognizes the6-O-sulfated groups on the CS ‘‘C unit’’ as well as those on theCSD and CSE units (14).
Ookinetes Exhibit Binding Affinity to CSGAGs. To determinewhether P. falciparum ookinetes can bind to CSGAGs, weprobed an overnight ookinete culture with CSA, CSC, CSD, andCSE as well as dermatan sulfate (DS) and hyaluronic acid (HA)as controls (Fig. 2). Although the overnight cultures yielded fewmature form ookinetes (�12–16% of the total number ofparasites counted were mature ookinetes, �60–65% were late-retort ookinetes, and the remainder were a mixture of earlyretorts (Stage II) and zygotes), the cultures permitted theanalysis of CSGAG staining of the mature form ookinetes (themotile parasite stage in the mosquito midgut). We observed thatmature ookinetes exhibit binding affinity to CSA and CSE (Fig.2 a–f ) and that recognition of the CS ligands appears to bestructural-specific because ookinete binding to CSA was morediffused than CSE (Fig. 2 a and d), and affinity for CSC, CSD,or DS was not observed (data not shown). The CSD disaccharidediffers from CSA, CSC, and CSE motifs by the presence of asulfate group on C-2 of GlcA (Fig. 1i). Ookinete recognition forDS was analogous to what was observed for HA, which is
nonsulfated (Fig. 2 g–i). These data suggest that the presence ofGlcA and not IdoA is important for ookinete recognitionbecause GlcA replacement by IdoA in DS disaccharides leads to
Fig. 1. Histological analysis revealed polarized localization of CS and HS proteoglycans in An. gambiae mosquito midgut epithelial cells. (a and b) Hematoxylinand eosin-stained mosquito (sugar fed) midgut cross-sections were analyzed for the presence of acid mucopolysaccharides. Midgut sections were incubated inbuffer (pH 1.0) in the absence (a) or presence (b) of the polyvalent cationic dye, Alcian blue. Red arrow indicates apical microvilli staining with the dye. (c–h)Immunofluorescence analyses of midguts from blood-fed mosquitoes (24-h PBF) indicating polarized distribution of GAG species. (c) Bright-field image of theMAb CS-56 anti-CS-stained midgut section (d) showing midgut luminal staining. (e) Bright-field image of the anti-CS scFv antibody IO3H10-stained midgut section( f). (g) Bright-field image of the anti-HS scFv antibody RB4EA12-stained midgut section (h). Note that MAb CS-56 and scFv IO3H10 both corroborate the polarizedexpression of CSGAG to the apical microvilli. Arrows indicate preferential staining of the luminal interface between midgut epithelial cells and the blood mealor the basal lamina of the mosquito midgut. (i) CS disaccharide unit specificity of antibodies used in the analyses. MAb CS-56 has recognition specificity for CSA, C, and D units, and trisulfated (A-D units) on octasaccharide chains, whereas scFv IO3H10 has recognition specificity for CS C, D, and E units. MV, microvilli;LUMEN, midgut interior; BM, blood meal; E, midgut epithelial cell. DAPI, which was used to stain nuclei (blue), provides apical and basal orientation for thefluorescence images. Texas red-labeled secondary antibodies were used for detection and appear red. (Scale bars, �100 �m.)
Fig. 2. P. falciparum ookinetes bind to specific CS glycosaminoglycans(CSGAG). CSA (a–c) and CSE (d–f ) bind to mature ookinetes, whereas HA (g–i)do not. CSC, CSD, and DS produced similar results to HA. All GAGs werebiotinylated and detected with Alexa 488 conjugated to streptavidin (green).Note that CSE exhibited less diffused ookinete staining intensity than CSA. Thestructure of CSA, CSC, CSD, and CSE are shown in Fig. 1i, and DS and HA aredescribed in Results. Parasite morphology, along with an antibody to theookinete surface protein, Pfs25, was used to identify mature form ookinetes(red). DAPI was used to stain parasite nuclei (blue). (Scale bars, �10 �m.)
Dinglasan et al. PNAS � October 2, 2007 � vol. 104 � no. 40 � 15883
MIC
ROBI
OLO
GY

a loss of affinity and that GlcA needs to be in tandem withGalNAc, because HA (which is composed of GlcA-GlcNAcdisaccharides) also fails to bind to ookinetes. Similar results werealso observed for P. falciparum ookinetes that developed in vivo[supporting information (SI) Fig. 6].
The Anopheles gambiae GAG Biosynthetic Pathway and Cloning of theMosquito Peptide-O-Xylosyltransferase. CSGAG dissacharide unitsare attached to a tetrasaccharide core composed of a GlcA, twogalactose (Gal) units and a xylose (Xyl) residue (GlcA�1–3Gal�1–3Gal�1–4Xyl�1-Ser) that is glycosidically linked to thehydroxyl group of serines on proteoglycan polypeptides (15).CSGAGs vary in the degree and pattern of sulfation through thedifferential action of carbon 2-O, 4-O, and 6-O sulfotransferasesacting on GalNAc and the combination of A-E units (16). Inmammals, the addition of Xyl by a peptide-O-xylosyltransferase(OXT) to the Ser residue on a proteoglycan core initiates thesequential formation of the tetrasaccharide core (Fig. 3a, inbold). Unlike in mammals, for Drosophila (and putatively forother dipterans) the activity of a CS GalNAc-transferase I(GalNAcT-I) as opposed to an N-acetylglucosaminyl transferaseI (GlcNAcT-I) determine the initiation and progression of thechain toward either a CSGAG or HSGAG, respectively (16–18)(Fig. 3a). Anopheles homologs of Drosophila melanogaster GAGbiosynthetic enzymes, including OXT, as well as the genesencoding several proteoglycans have not been characterized (16,19). An An. gambiae genome database search using D. melano-gaster sequences suggested that two predicted OXT genes (EN-
SANGG00000025678; ENSANGG00000003504) exist in themosquito. However, similar to Drosophila (16), of the twohomologs identified, only the transcript for the latter gene(ENSANGG00000003504) could be amplified by PCR from anAn. gambiae midgut cDNA library (Fig. 3b). The complete An.gambiae O-xylosyltransferase (AgOXT1) sequence revealed atranscript size of 2.8 kb and a predicted 876-aa polypeptide (Fig.1 b and c). As with human and Drosophila OXTs, AgOXT1 is atype II transmembrane protein, with a WSC motif glycan-binding domain (GBD) and glycosyltransferase family 14 do-mains (Fig. 3 b and c), but lacks the characteristic Asp-Xaa-Asp(DXD) motif present in other family members and is involved incoordination of glycan substrates (16). The functional relevanceof the lack of a DXD motif in AgOXT1 is not readily apparent,although it has been shown that nematode OXT, which also lacksthis motif, is enzymatically active (15).
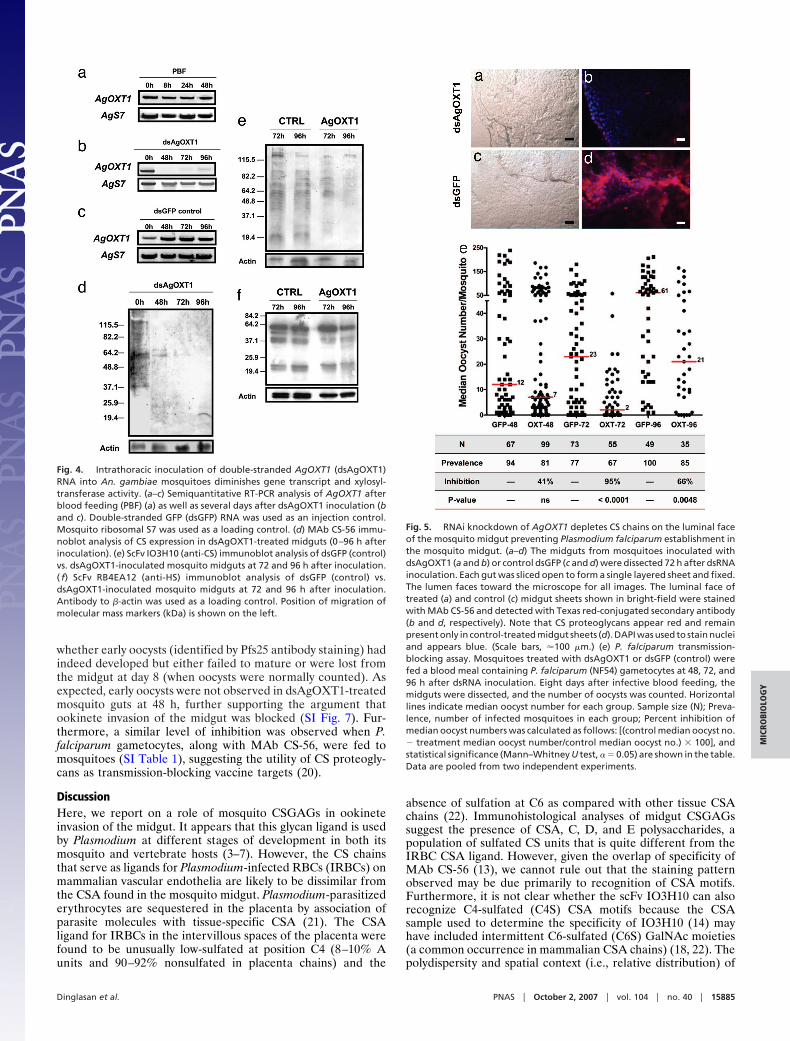
RNAi-Mediated Depletion of Midgut CS Blocks Parasite Invasion.Having confirmed the presence of sulfated CSGAGs on midgutmicrovilli and the ability of ookinetes to bind to this class ofpolysaccharides, we sought to determine the role of these ligandsduring midgut invasion. To do so, we produced double-strandedRNA corresponding to a 720-bp coding region of AgOXT1 (Fig.3b, gray arrows) and injected intrathoracically this construct intoadult female An. gambiae mosquitoes. Total RNA was collectedfrom blood-fed and sugar-fed adult mosquitoes at 48, 72, and96 h after inoculation with dsRNA. AgOXT1 mRNA is consti-tutively transcribed during normal blood feeding (Fig. 4a).However, we found that mRNA levels were reduced significantlybetween 48 and 72 h after inoculation by double-strandedAgOXT1 (dsAgOXT1), followed by modest recovery at 96 h (Fig.4b) as compared with dsRNA control-inoculated mosquitoes(Fig. 4c). The innate turnover of large proteoglycans is generallyrapid. Therefore, given these results, we expected that thereduction in AgOXT1 message would lead to a loss of transferaseactivity and, consequently, the depletion of CSGAGs becausethe cycling polypeptide can no longer be modified. CS-56 andscFv IO3H10 immunoblot analyses of guts from the correspond-ing RT-PCR time-points (Fig. 4 d and e) supported our hypoth-esis, wherein we observed that CSGAGs were significantlyreduced in the midgut. The multiple banding pattern observedin midguts from untreated and dsRNA control-injected mosqui-toes (Fig. 4 d and e), implies the occurrence of several chon-droitin proteoglycan species. However, at present, the identitiesof these midgut proteoglycan backbones remain unknown. In-terestingly, despite AgOXT1 knockdown in the midgut, HSbiosynthesis remained unaffected at both 72 and 96 h afterinoculation (Fig. 4f ), suggesting that, in comparison with apicalCS proteoglycans, basal lamina/extracellular matrix HS proteo-glycans may not turn over during the period before (0–3 days) orduring experimental infection (24 h after blood feeding) with P.falciparum ookinetes. Immunofluorescence staining of the lu-minal face of the mosquito midgut sheets at 72 h after dsRNAinjection (Fig. 5 a–d) confirmed the previous immunoblot data,wherein a marked decrease in signal was observed indsAgOXT1-injected mosquitoes.
We then determined how the loss of luminal CSGAGs wouldaffect parasite development. Control and treated dsRNA-inoculated mosquitoes were provided a P. falciparum-infectedblood meal at 48, 72, and 96 h after inoculation (PI). The medianPlasmodium oocyst numbers were reduced by 95% (P � 0.0001)in the dsAgOXT1 group in comparison with dsGFP controls forthe 72 h PI cohorts (Fig. 5e). For the 96 h PI cohort, inhibitionwas reduced modestly to 66% (P � 0.0048), which may be a resultof transcript/enzyme recovery as was observed for this time pointby RT-PCR and immunoblot (Fig. 4 b, d, and e). We alsoexamined mosquito midguts from dsAgOXT1-treated mosqui-toes (72 h time point) at 48 h after blood feeding to determine
Fig. 3. Anopheles gambiae peptide O-xylosyltransferase (AgOXT1) mRNAtranscript, predicted protein structures, and its role in GAG biosynthesis. (a)RNA-meditated interference strategy to inhibit GAG biosynthesis. We hypoth-esized that RNAi knockdown of the AgOXT1will result in the depletion ofGAGs because of the loss of the tetrasaccharide core common to both heparanand CS chains. (b) The single predicted transcript for AgOXT1. Gray arrowsindicate primer sites used in the amplification of the double-stranded Ag-OXT1. (c) A comparison of the predicted protein structures of the enzyme fromAn. gambiae (Ag) and Drosophila melanogaster (Dm). White rectangle, trans-membrane domain; cross-hatched rectangle, a DXD motif. GBD, glycan-binding domain; GlycoT14, glycosyltransferase family 14 domain; Gal, galac-tose; GlcA, glucoronic acid; Xyl, xylose; GalT-I, galactosyltransferase-I; GalT-II,galactosyltransferase-II; GalNAcT-I, N-acetylgalactosaminyltransferase-I; Glc-NAcT-I, N-acetylglucosaminyltransferase-I.
15884 � www.pnas.org�cgi�doi�10.1073�pnas.0706340104 Dinglasan et al.
whether early oocysts (identified by Pfs25 antibody staining) hadindeed developed but either failed to mature or were lost fromthe midgut at day 8 (when oocysts were normally counted). Asexpected, early oocysts were not observed in dsAgOXT1-treatedmosquito guts at 48 h, further supporting the argument thatookinete invasion of the midgut was blocked (SI Fig. 7). Fur-thermore, a similar level of inhibition was observed when P.falciparum gametocytes, along with MAb CS-56, were fed tomosquitoes (SI Table 1), suggesting the utility of CS proteogly-cans as transmission-blocking vaccine targets (20).
DiscussionHere, we report on a role of mosquito CSGAGs in ookineteinvasion of the midgut. It appears that this glycan ligand is usedby Plasmodium at different stages of development in both itsmosquito and vertebrate hosts (3–7). However, the CS chainsthat serve as ligands for Plasmodium-infected RBCs (IRBCs) onmammalian vascular endothelia are likely to be dissimilar fromthe CSA found in the mosquito midgut. Plasmodium-parasitizederythrocytes are sequestered in the placenta by association ofparasite molecules with tissue-specific CSA (21). The CSAligand for IRBCs in the intervillous spaces of the placenta werefound to be unusually low-sulfated at position C4 (8–10% Aunits and 90–92% nonsulfated in placenta chains) and the
absence of sulfation at C6 as compared with other tissue CSAchains (22). Immunohistological analyses of midgut CSGAGssuggest the presence of CSA, C, D, and E polysaccharides, apopulation of sulfated CS units that is quite different from theIRBC CSA ligand. However, given the overlap of specificity ofMAb CS-56 (13), we cannot rule out that the staining patternobserved may be due primarily to recognition of CSA motifs.Furthermore, it is not clear whether the scFv IO3H10 can alsorecognize C4-sulfated (C4S) CSA motifs because the CSAsample used to determine the specificity of IO3H10 (14) mayhave included intermittent C6-sulfated (C6S) GalNAc moieties(a common occurrence in mammalian CSA chains) (18, 22). Thepolydispersity and spatial context (i.e., relative distribution) of
Fig. 4. Intrathoracic inoculation of double-stranded AgOXT1 (dsAgOXT1)RNA into An. gambiae mosquitoes diminishes gene transcript and xylosyl-transferase activity. (a–c) Semiquantitative RT-PCR analysis of AgOXT1 afterblood feeding (PBF) (a) as well as several days after dsAgOXT1 inoculation (band c). Double-stranded GFP (dsGFP) RNA was used as an injection control.Mosquito ribosomal S7 was used as a loading control. (d) MAb CS-56 immu-noblot analysis of CS expression in dsAgOXT1-treated midguts (0–96 h afterinoculation). (e) ScFv IO3H10 (anti-CS) immunoblot analysis of dsGFP (control)vs. dsAgOXT1-inoculated mosquito midguts at 72 and 96 h after inoculation.( f) ScFv RB4EA12 (anti-HS) immunoblot analysis of dsGFP (control) vs.dsAgOXT1-inoculated mosquito midguts at 72 and 96 h after inoculation.Antibody to �-actin was used as a loading control. Position of migration ofmolecular mass markers (kDa) is shown on the left.
Fig. 5. RNAi knockdown of AgOXT1 depletes CS chains on the luminal faceof the mosquito midgut preventing Plasmodium falciparum establishment inthe mosquito midgut. (a–d) The midguts from mosquitoes inoculated withdsAgOXT1 (a and b) or control dsGFP (c and d) were dissected 72 h after dsRNAinoculation. Each gut was sliced open to form a single layered sheet and fixed.The lumen faces toward the microscope for all images. The luminal face oftreated (a) and control (c) midgut sheets shown in bright-field were stainedwith MAb CS-56 and detected with Texas red-conjugated secondary antibody(b and d, respectively). Note that CS proteoglycans appear red and remainpresent only in control-treated midgut sheets (d). DAPI was used to stain nucleiand appears blue. (Scale bars, �100 �m.) (e) P. falciparum transmission-blocking assay. Mosquitoes treated with dsAgOXT1 or dsGFP (control) werefed a blood meal containing P. falciparum (NF54) gametocytes at 48, 72, and96 h after dsRNA inoculation. Eight days after infective blood feeding, themidguts were dissected, and the number of oocysts was counted. Horizontallines indicate median oocyst number for each group. Sample size (N); Preva-lence, number of infected mosquitoes in each group; Percent inhibition ofmedian oocyst numbers was calculated as follows: [(control median oocyst no.� treatment median oocyst number/control median oocyst no.) � 100], andstatistical significance (Mann–Whitney U test, � � 0.05) are shown in the table.Data are pooled from two independent experiments.
Dinglasan et al. PNAS � October 2, 2007 � vol. 104 � no. 40 � 15885
MIC
ROBI
OLO
GY

CSA and CSC, CSD, or CSE on chondroitin chains vary greatlyand have been found to be tissue- or species-specific (13, 14, 18,and 22). Therefore, although it remains possible that CSA maybe the major sulfated chondroitin unit in the mosquito midgut,analysis of the relative position of CSA units along the chain (i.e.,disaccharide repeat sequence) is essential to get a completeunderstanding of CSGAG structure.
Evidence for the putative importance of spatial context forCSA is highlighted by the observation that the CSA motif forIRBCs sequestering in the placenta is a dodecasaccharide do-main consisting of two to four A-units interspersed with non-sulfated or C-units. These data suggest that IRBC-bindingaffinity for CSGAGs is highly structural-specific. We observedthat mature ookinetes also bind to CSGAGs in a structural-specific manner. However, it appears that GalNAc (C4S) or(C6S), although necessary to confer binding affinity, may not bethe only binding determinants on CSGAGs. It appears thatsulfate and carboxylate position on GalNAc and GlcA, respec-tively, and not necessarily the presence of charge alone isimportant for binding to ookinetes. For example, epimerizationof C-5 in GlcA produces the L-iduronate (IdoA) moiety presentin DS (23). Although GalNAc of DS maintains the 4S modifi-cation, IdoA abrogates ookinete recognition (presumablythrough its axial carboxylate group). Replacing GalNAc withGlcNAc produces HA, which appears to be poorly recognized byookinetes. Because of the inherent difficulty of producing manymature-stage ookinetes in vitro (24), it is difficult to assessCSGAG affinity by alternative methods (e.g., ELISA). However,because mature ookinetes are the final, motile stage, andmosquito CSGAGs are not present in the blood-meal bolus butare localized to the midgut apical microvilli, then we can inferfrom the data that mature ookinetes, which are involved inmidgut adhesion, use midgut CSGAGs in the invasion process.Note that CS chains exist as repeating disaccharide polymers andthat the use of biotinylated disaccharides in our binding assaysmay have underestimated the affinity and complexity of CSGAGrecognition by ookinetes. Taken together, the results suggestthat CS disaccharide sequence and relative sulfate position arecritical structural determinants for ookinete recognition.
Plasmodium sporozoites have been shown to recognize liverHS proteoglycans through the major surface protein CSP andTRAP (3, 4). The related ookinete protein, CTRP can bind toheparin in vitro. However, heparin and HSs are quite dissimilarand both differ greatly from CSGAGs. Therefore, it remains tobe seen whether CTRP is the cognate receptor on ookinetes formidgut CSGAGs. It should be noted that recombinant CTRPbinding may not directly reflect what occurs in vivo. Alterna-tively, other ookinete surface lectin-like molecules may also beinvolved in CSGAG recognition.
Because peptide-O-xylosyltransferase is involved in the firststep of GAG biosynthesis, depletion of AgOXT1 resulted in adramatic decrease in GAG expression and marked inhibition ofparasite development in the mosquito. We also targeted theGalNAcT-1 by RNAi and observed a reduction in transcriptabundance without the consequent reduction in CSGAG-modified molecules in the midgut (data not shown). As expected,parasite development was also unaffected. We predict thattargeting other enzymes in the biosynthetic pathway will producesimilar results, presumably because of functional redundancy ofrelated glycosyltransferases (25) in the gut.
Analysis of the role of glycans in mediating Plasmodiummerozoite invasion of erythrocytes revealed the existence of asialic acid-independent adhesion pathway (26). Preliminary ev-idence suggests that Plasmodium ookinetes also exhibit alterna-tive midgut invasion mechanisms (ref. 27 and R.R.D., unpub-lished results). Therefore, RNAi leakiness notwithstanding, if aGAG-independent midgut invasion pathway exists, then it ispossible that, for at least one parasite subpopulation, other
ookinete lectin-like molecules (28) work in concert with theircognate midgut surface proteins to compensate for the loss ofGAG ligands. In the context of our experiments, other midgutmembrane glycan ligands that are left unaffected by RNAiknockdown include neutral and sulfated glycans and proteindomains on glycolipids and glycoproteins, respectively. It is alsopossible that in the absence of critical sulfated glycans, protein–protein interactions may be sufficient for facilitating parasiterecognition, adhesion, and invasion of midgut epithelial cells.Given the possible diversity of midgut invasion mechanisms,future work should examine different parasite–mosquito speciescombinations to determine the role, if any, that CSGAGs play inthese interactions.
Our data has established the role of glycans as ligands thatmediate parasite invasion of the mosquito midgut. However, themechanisms by which glycans are used for cell adhesion or cellinvasion are still poorly understood. The sequence and structureof midgut CS chains as well as identification of cognate lectin-like molecules on ookinetes remain to be analyzed.
After this manuscript was submitted for publication, a paperby Sinnis et al. [Sinnis P, Coppi A, Toida T, Toyoda H,Kinoshita-Toyoda A, Xie J, Kemp MM, Linhardt RJ (2007)J Biol Chem, in press] appeared that further supported ourfindings that chondroitin and HS polysaccharides are present inmosquito midguts.
Materials and MethodsImmunofluorescence Microscopy of Midgut Sections. Midguts ormidgut sheets from blood-fed or sugar-fed mosquitoes were fixedin 4% paraformaldehyde overnight at 4°C. Whole guts wereembedded in paraffin for sectioning, and midgut sheets wereused as whole mounts. Sections were blocked (PBS/3% BSA)overnight at 4°C. Each section was probed with either MAbCS-56 (1:50) (Sigma, St. Louis, MO) overnight at 4°C or theperiplasmic fraction of RB4EA12 (29) containing anti-HSsingle-chain variable fragment (scFv) antibody (1:100) for 2 h atroom temperature. MAb CS-56 was detected by Texas red-conjugated �-mouse IgM secondary antibody (Invitrogen, Carls-bad, CA). Anti-HS scFv was detected with a mouse anti-VSV tagMAb, followed by detection with Texas red-conjugated anti-mouse secondary antibody. The slides were mounted withSlowfade reagent containing DAPI (Molecular Probes/Invitrogen) and visualized by microscope.
CS–Ookinete-Binding Assays. An 18- to 19-d-old P. falciparumNF54 gametocyte culture was transferred to a flask and diluted1:5 in complete ookinete medium (24, 27) supplemented with15% heat-inactivated fetal calf serum and incubated at 19°C for24 h with rocking. Transformed ookinetes were purified bytreatment with Lympholyte (Cedarlane Laboratories, Ontario,Canada) and Percoll density gradient. Parasites were smearedonto glass slides, air-dried, and then fixed in 4% paraformalde-hyde for 1 h. Biotinylated CSA, CSC, CSD, CSE, DS, and HA(each at 10 �g/ml) were used to probe the slides [Saito et al. (30)and Saito et al. (31)]. CS probes were detected by Alexa-488conjugated Streptavidin (Molecular Probes/Invitrogen). Ooki-netes were identified by morphology and the use of anti-Pfs25monoclonal antibody followed by detection using Texas red-conjugated anti-mouse secondary antibody (Molecular Probes/Invitrogen). The slides were mounted with Slowfade reagentcontaining DAPI (Molecular Probes/Invitrogen) and imagescaptured as above.
Identification of Anopheles gambiae O-Xylosyltransferase Homologs.We used the D. melanogaster amino acid sequence for peptide-O-xylosyltransferase 1 DmPOXT1 (16) to BLAST the EnsemblAn. gambiae genome database v.37 (www.ensembl.org/Anopheles�gambiae/index.html). The complete sequence of all
15886 � www.pnas.org�cgi�doi�10.1073�pnas.0706340104 Dinglasan et al.

Anopheles OXT homologs was determined from ORF scanningof genomic data 5�-upstream of the predicted cDNAs found inthe database. Primers corresponding to AgOXT1 (1557F, 5�-CAGCACAAGAAGCTGTTCTTTGGC-3� and 2337R, 5�-AGCTTCCGTGACGAGAAATCGTGT-3�) were used toscreen an An. gambiae cDNA library.
SDS/PAGE and Western Blot Analysis. Mosquito midgut lysates (n �5–10 female midguts) were fractionated on a 10% Tris-Glycinegel and transferred to nitrocellulose membranes. Transblotswere probed with mouse MAb CS-56 �-CS (IgM) (Sigma)diluted 1:100 or anti-chondroitin O-6 sulfate antibodies (1:100).Primary antibodies were detected by use of alkaline phos-phatase-conjugated �-mouse IgM antibody (1:5,000) and chemi-luminescence by using CDP-Star substrate with Nitroblock II(PerkinElmer, Wellesley, MA).
Semiquantitative RT-PCR. The AgOXT1 primers, 1557F and2337R, were used to amplify a 780-bp product corresponding tothe C terminus of AgOXT1 from our An. gambiae midgut cDNAlibrary. The PCR product was cloned into the pCR2.1 TOPO(Invitrogen). For RT-PCR, 1 �g of total midgut RNA (n � 10females) collected from each time point after blood feeding wasreverse-transcribed following standard protocols. AgOXT1 tran-script abundance was determined by using the primers above.The amplified An. gambiae ribosomal protein gene (AgS7)product was used as a loading control (AgS7F, 5�-TGCTG-CAAACTTCGGCTAT-3�, and AgS7R, 5�-CGCTATGGTGT-TCGGTTCC-3�). Amplification using Herculase polymerase(Stratagene, La Jolla, CA) was performed as follows: 94°C for 2min, 94°C for 45 s, 55°C for 45 s, and 72°C for 45 s, with a finalextension at 72°C for 10 min (30 cycles). Cloned PCR productswere confirmed by DNA sequencing.
RNA-Mediated Interference (RNAi). AgOXT dsRNA was producedby using the primers above with the T7 polymerase promoter
sequence, 5�-TAATACGACTCACTATAGGG-3�, appended tothe 5� termini of each primer and transcribed in vitro by using theMegascript RNAi kit (Ambion, Austin, TX) as per the manu-facturer’s protocols. The dsRNA solution was diluted to 6 �g/�lin injection buffer (10 mM Tris). An. gambiae females (5–6 d old)were anesthetized and inoculated intrathoracically with 400 ng ofdsRNA or control GFP dsRNA. Mosquitoes were maintainedfor 2, 4, or 5 days before being fed a blood meal containing P.falciparum gametocytes (see below).
Parasite Membrane Feeding Assay. P. falciparum gametocyte cul-tures from isolate NF54 were maintained as at the JohnsHopkins Malaria Research Institute Parasite Core facility. Cul-tures were harvested 15–17 d after initiation. The culture waswashed and brought up in normal human serum (InterstateBlood Bank, Memphis, TN) plus human RBCs at 0.3% game-tocytemia in a 45% hematocrit. Infective blood was delivereddirectly into glass, water-jacketed membrane feeders warmed to38°C. Mosquitoes were allowed to blood feed for 20 min. Themosquitoes were maintained for 8 d, and only midguts from fullygravid females from each cage were further analyzed because eggdevelopment serves as a proxy for blood feeding to repletion.The dissected midguts were stained with 0.1% mercurochrometo visualize oocyst development (the read-out for successfulparasite invasion). Three independent experiments were per-formed for each corresponding RNAi time point.
Statistical Analyses. Nonparametric statistical analysis was used toevaluate the difference in median oocyst intensity betweenexperimental and control groups (Mann–Whitney U test, one-tailed, � � 0.05) using the STATVIEW 5.0 software (SASInstitute, Cary, NC).
This work was supported by National Institute of Allergy and InfectiousDiseases, National Institutes of Health (NIH) Grants R01AI031478 andRR00052. R.R.D. is a NIH Ruth L. Kirschstein National ResearchService Award scholar (5F32AI068212–02).
1. Ghosh A, Edwards MJ, Jacobs-Lorena M (2000) Parasitol Today 16:196–201.2. Li F, Templeton TJ, Popov V, Comer JE, Tsuboi T, Torii M, Vinetz JM (2004)
J Biol Chem 279:26635–26644.3. Pinzon-Ortiz C, Friedman J, Esko J, Sinnis P (2001) J Biol Chem 276:26784–26791.4. Pradel G, Garapaty S, Frevert U (2002) Mol Microbiol 45:637–651.5. Templeton TJ, Kaslow DC, Fidock DA (2000) Mol Microbiol 36:1–9.6. Yuda M, Yano K, Tsuboi T, Torii M, Chinzei Y (2001) Mol Biochem Parasitol
116:65–72.7. Dinglasan RR, Jacobs-Lorena M (2005) Infect Immun 73:7797–7807.8. Dinglasan RR, Fields I, Shahabuddin M, Azad AF, Sacci JB, Jr (2003) Infect
Immun 71: 6995–7001.9. Lev R, Spicer SS (1964) J Histochem Cytochem 12:309.
10. Dennissen MA, Jenniskens GJ, Pieffers M, Versteeg EM, Petitou M,Veerkamp JH, van Kuppevelt TH (2002) J Biol Chem 277:10982–10986.
11. Avnur Z, Geiger B (1984) Cell 38:811–822.12. Tveit H, Dick G, Skibeli V, Prydz K (2005) J Biol Chem 280:29596–29603.13. Ito Y, Hikino M, Yajima Y, Mikami T, Sirko S, von Holst A, Faissner A, Fukui
S, Sugahara K (2005) Glycobiology 15:593–603.14. Smetsers TF, van de Westerlo EM, ten Dam GB, Overes IM, Schalkwijk J, van
Muijen GN, van Kuppevelt TH (2004) J Invest Dermatol 122:707–716.15. Mizuguchi S, Uyama T, Kitagawa H, Nomura KH, Dejima K, Gengyo-Ando
K, Mitani S, Sugahara K, Nomura K (2003) Nature 423:443–448.16. Wilson IB (2002) J Biol Chem 277:21207–21212.17. Costa-Filho A, Souza ML, Martins RC, dos Santos AV, Silva GV, Comaru MW,
Moreira MF, Atella GC, Allodi S, Nasciutti LE, et al. (2004) Insect Biochem MolBiol 34:251–260.
18. Medeiros GF, Mendes A, Castro RA, Bau EC, Nader HB, Dietrich CP (2000)Biochim Biophys Acta 1475:287–294.
19. Pinto DO, Ferreira PL, Andrade LR, Petrs-Silva H, Linden R, Abdelhay E,Araujo HM, Alonso CE, Pavao MS (2004) Glycobiology 14:529–536.
20. Dinglasan RR, Valenzuela JG, Azad AF (2005) Insect Biochem Mol Biol35:1–10.
21. Fried M, Domingo GJ, Gowda CD, Mutabingwa TK, Duffy PE (2006) ExpParasitol 113:36–42.
22. Achur RN, Valiyaveettil M, Gowda DC (2003) J Biol Chem 278:11705–11713.23. Trowbridge JM, Gallo RL (2002) Glycobiology 12:117R–25R.24. Carter EH, Suhrbier A, Beckers PJ, Sinden RE (1987) Parasitology 95 (Pt
1):25–30.25. Ten Hagen KG, Tran DT, Gerken TA, Stein DS, Zhang Z (2003) J Biol Chem
278:35039–35048.26. Duraisingh MT, Maier AG, Triglia T, Cowman AF (2003) Proc Natl Acad Sci
USA 100:4796–4801.27. Dinglasan RR, Kalume DE, Kanzok SM, Ghosh AK, Muratova O, Pandey A,
Jacobs-Lorena M (2007) Proc Natl Acad Sci USA, in press.28. Trueman HE, Raine JD, Florens L, Dessens JT, Mendoza J, Johnson J,
Waller CC, Delrieu I, Holders AA, Langhorne J, et al. (2004) J Parasitol90:1062–1071.
29. ten Dam GB, Hafmans T, Veerkamp JH, van Kuppevelt TH (2003) J HistochemCytochem 51:727–739.
30. Saito A, Munakata H (2004) Electrophoresis 25:2452–2460.31. Saito A, Munakata H, Satoh K (2002) Connect Tissue Res 43:1–7.
Dinglasan et al. PNAS � October 2, 2007 � vol. 104 � no. 40 � 15887
MIC
ROBI
OLO
GY


















