
PLANT HEMAGGLUTININS WITH SPECIAL … HEMAGGLUTININS WITH SPECIAL REFERENCE TO A ... that of the...
18
PLANT HEMAGGLUTININS WITH SPECIAL REFERENCE TO A PREPARATION FROM THE-NAVY BEAN.* BY VERZ R. GODDARD AND LAFAYETTE B. MENDEL. (From the Laboratory of Physiological Chemistry, Yale University, New Haven.) (Received for publication, February 21, 1929.) Since the discovery of ricin by Kobert in 1887, considerable interest has been shown in the hemagglutinins obtainable from plants. The study of them has been somewhat overshadowed by the more recently discovered serum agglutinins, which they so remarkably resemble in action. They have, however, played an important part in immunological research throughout its history. Ricin, the extremely toxic hemagglutinin from the castor bean (Ricinus communis) was used by Ehrlich (1897) in his first in vitro studies of the reactions between antigen and antib0dy.l The living organism was no longer to be regarded as an indispens- able participant in the agglutinin-antiagglutinin reaction. Much remains to be done before the distribution of the hemag- glutinins in the plant world can be estimated. From the observa- tions made by Landsteiner and Raubitschek (1908), Mendel (1909), Eisler and Portheim (1909, 1926), Wienhaus (1909), and others, the substances appear to lack wide distribution.2 The early studies were confined to the toxic agglutinins: ricin, robin, cretin, and abrin. Later, non-toxic extracts were prepared in * The data reported in this paper are taken from the dissertation pre- sented by Verz R. Goddard in partial fulfilment of the requirements for the degree of Doctor of Philosophy, Yale University, 1927. r He showed that ricin neutralized by antiricin would not agglutinate rabbit corpuscles in a test-tube. From quantitative work, he concluded that this neutralization followed the law of multiple proportions. 2 A detailed summary of the principal plant sources which have been investigated with reference to the occurrence of hemagglutinins and of the animal species furnishing the blood corpuscles tested is included in the dissertation of Verz R. Goddard, Yale University, 1927. 417 by guest on May 25, 2018 http://www.jbc.org/ Downloaded from
Transcript of PLANT HEMAGGLUTININS WITH SPECIAL … HEMAGGLUTININS WITH SPECIAL REFERENCE TO A ... that of the...

PLANT HEMAGGLUTININS WITH SPECIAL REFERENCE TO A PREPARATION FROM THE-NAVY BEAN.*
BY VERZ R. GODDARD AND LAFAYETTE B. MENDEL.
(From the Laboratory of Physiological Chemistry, Yale University, New Haven.)
(Received for publication, February 21, 1929.)
Since the discovery of ricin by Kobert in 1887, considerable interest has been shown in the hemagglutinins obtainable from plants. The study of them has been somewhat overshadowed by the more recently discovered serum agglutinins, which they so remarkably resemble in action. They have, however, played an important part in immunological research throughout its history.
Ricin, the extremely toxic hemagglutinin from the castor bean (Ricinus communis) was used by Ehrlich (1897) in his first in vitro studies of the reactions between antigen and antib0dy.l The living organism was no longer to be regarded as an indispens- able participant in the agglutinin-antiagglutinin reaction.
Much remains to be done before the distribution of the hemag- glutinins in the plant world can be estimated. From the observa- tions made by Landsteiner and Raubitschek (1908), Mendel (1909), Eisler and Portheim (1909, 1926), Wienhaus (1909), and others, the substances appear to lack wide distribution.2 The early studies were confined to the toxic agglutinins: ricin, robin, cretin, and abrin. Later, non-toxic extracts were prepared in
* The data reported in this paper are taken from the dissertation pre- sented by Verz R. Goddard in partial fulfilment of the requirements for the degree of Doctor of Philosophy, Yale University, 1927.
r He showed that ricin neutralized by antiricin would not agglutinate rabbit corpuscles in a test-tube. From quantitative work, he concluded that this neutralization followed the law of multiple proportions.
2 A detailed summary of the principal plant sources which have been investigated with reference to the occurrence of hemagglutinins and of the animal species furnishing the blood corpuscles tested is included in the dissertation of Verz R. Goddard, Yale University, 1927.
417
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

448 Plant Hemagglutinins
various laboratories. The seeds of leguminous plants are es- pecially rich in potent products.
As blood cell clumping substances, phytohemagglutinins ap- parently have no relationship to the activity of the plant producing them. This is analogous to the well known fact that plants pro- duce enzymes having no obvious bearing upon the activities of the structures that yield them. The presence of pancreatic se- cretin in spinach and of urease in beans are obvious examples.
A possible physiological relationship between the agglutinin and the plant producing it has been indicated by Eisler and Portheim (1911, 1912, 1926) and by Schneider (1912). The hemagglutinin appears to be associated with the storage material of plants. Upon germination the seed loses its agglutinin which may pass over to the embryo (Russ and Oesterlin, 1921). It is interesting to note that certain plants which have reserve material in their stems, possess an agglutinating milk-sap. This is true of the Euphorbia studied by Eisler and Portheim. More recently, Mar- cusson-Begun (1926) has reported an agglutinin in the potato and has suggested the possibility of the occurrence of agglutinating substances in other root nodules.
The classic agglutinin-containing seed, that of the castor bean (Ricinus communis) was studied chemically by Osborne, Mendel, and Harris (1905), who made a separation of its proteins. Both the toxic and the hemagglutinative qualities were found to be as- sociated with the albumin, ricin, a name earlier indicated by Kobert for the toxic principle. Their method for the purification of ricin may be used for its preparation on a large scale (Karrer, Smirnoff, Ehrensperger, van Slooten, and Keller, 1924).
For many purposes a non-toxic agglutinin is of value. Such a substance might well prove of considerable practical importance as an aid in the preparation of antisera. In fact, a crude extract of the pea-bean (Phaseolus communis) is being used in the prepa- ration of certain of the anti hog cholera sera on a commercial scale (Dorset and Henley, 1916). It has been shown that where a clear, corpuscle-free serum is desired, the yield obtainable from a given volume of blood can be notably increased by the use of this agglutinin which causes the corpuscles to pack and makes possible a nearly perfect separation of cells and serum.
The present study involves the preparation of a non-toxic
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 449
hemagglutinin in a highly purified form and an investigation of the mechanism of its action in the hope of making the material more useful in research as well as for practical, serological work. The applicability of such a substance is, of course, not limited to immunological research. It can serve in various capacities in the study of the physical characteristics of erythrocytes and possibly of tissue cells in suspension. Wienhaus (1909) has shown that a bean extract would cause agglutination of pus, liver, and kid- ney cells.
Preparation of Hemagglutinin and Other Experimental Procedures.
For the preparation of the water-soluble protein fraction of the bean (Phaseolus communis) the method employed by Osborne, Mendel, and Harris in the preparation of ricin was closely fol- lowed. The influence of alterations in the integrity of the protein upon the hemagglutinating properties was also investigated.
After extraction of the defatted bean meal with sodium chloride solution (usually 3 per cent), removal of the undesired globulins by dialysis of the extract, repeated salting out of the desired pro- tein with ammonium sulfate, and dialysis of the precipitate to remove inorganic contaminants, a clear, straw-colored filtrate was obtained which, was evaporated in a current of air to dryness at 3540” on glass plates. The protein was then scraped from the plates, further desiccated over sulfuric acid, and stored in the form of a nearly white powder in glass bottles. Upon analysis the samples, with corrections for moisture and ash, varied in nitrogen content from 14.5 to 15.5 per cent. The material re- sponded positively to all the common protein color and precipita- tion tests. This substance had a high hemagglutinating potency. A sample which has been stored for over 2 years still shows (August, 1928) no appreciable loss in activity. Injections of as much as 8 mg. per kilo into rabbits and 600 per kilo into mice in- dicated that the material is non-toxic, in contrast to the com- parable ricin preparation from the castor bean for which a lethal dose as low as 0.002 mg. has been observed.
Macroscopic Agglutinatipn Method.-Many-macroscopic methods for detecting agglutination of bacteria have been published and a plea has been made for standardization of reports of agglutina- tion tests (Hadley, 1916). Most of these procedures are based
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

450 Plant Hemagglutinins
upon the Kolmer technique (Kolmer, 1923). Gardner’s gelatin standard method (Gardner, 1919) and Gates’ capillary tube method (Gates, 1921) represent special modifications. The diffi- culty in detecting agglutination appears to arise from the fact that it is a two phase reaction, the binding of the agglutinating agent occurring previous to flocculation. Gates (1922) has sug- gested a method for eliminating this time element in the agglutina- tion of bacteria. He hastened the process by centrifuging the organisms in contact with the agglutinin. A similar technique was recommended by Mudd (1927). The centrifuge method was found, in the experience of the writers, to give reasonably good results with erythrocytes also. After centrifugation agglutinated cells would remain in the bottom of an upturned tube or if ag- glutination were slight, would stream very slowly in comparison to the controls.
In an attempt to devise a procedure, however, which would give completely consistent results in the quantitative study of hemagglutination, factors known to be significant in clumping and sedimentation were taken into consideration. Temperature, diameter of tube and length of liquid column, angle or slope of tube were regulated to give the best results obtainable. A chance observation led to the development of a method which proved invaluable in determination of agglutination titer.
It was found in agreement with Berczeller and Wastl (1923) that slanting the tube increased the rate of sedimentation. But it was noted, also, that after erythrocytes had been left in contact with agglutinin in chemically clean glass tubes, sloped at an angle of 40” and at a temperature of 40-44”, for 12 hours, the cells became firmly attached to the glass so that upon returning the tube to the vertical position they remained unmoved. This pe- culiar adherence of cells never occurred in the absence of hemag- glutinin nor when the latter was too dilute to bring about clump- ing detectable upon microscopic examination.
The ease by which hemagglutination is detected by this method is indicated in Fig. 1. Note the appearance of tubes in which the same volumes of rabbit erythrocytes, washed and suspended in physiological salt solution, were treated with increasing quan- tities of hemagglutinin. In the tube furthest left and where no agglutination occurred the corpuscles settled; the other tubes show
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 451
clearly the adherence of the red cells characteristic of agglutina- tion upon the glass walls.
After various modifications had been investigated, the follow- ing procedure was finally selected as most effective. Rabbit blood freshly removed from the ear vein and defibrinated was centrifuged until the serum could be removed. The corpuscles washed twice with 0.89 per cent sodium chloride solution were suspended therein. 2.5 cc. of packed corpuscles were suspended in a total volume of 100 cc. of the isotonic sodium chloride solu-
FIG. 1. Hemagglutination test; one negative (left), four positive agglu- tinations (right). Tubes contained (left to right): 0.0005, 0.0006, 0.0007, 0.0008, and 0.0009 mg. of hemagglutinin with 0.5 cc. of 2.5 per cent suspen- sion of rabbit erythrocytes in sodium chloride solution. (Each tube was made up to a total volume of 1 cc.)
tion. Aliquots of this suspension were used for the individual tests. As a rule 0.5 cc. of the corpuscle suspension was added to 0.5 cc. of 0.89 per cent sodium chloride solution containing varying amounts of the agglutinin preparation to be tested. The reaction was carried out in chemically clean glass tubes, 75 X 8 mm., placed in a rack which would keep them at an angle of 40” and incubated at 4044”, for 12 hours, Table I indicates the uniformity of results secured by this method of “agglutination titer” when tests were repeated on different days, with fresh suspensions of eryth-
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
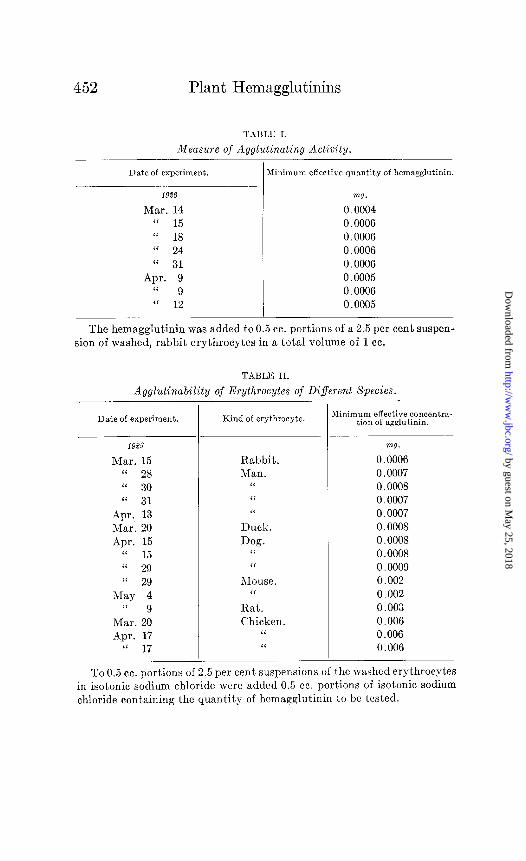
452 Plant Hemagglutinins
TAI3LlG I.
Measure of Agglutinating Activity.
Date of experiment. Minimum effective quantity of hemagglutinin.
1926 VW.
Mar. 14 0.0004 “ 15 0.0006 (‘ 18 0.0006 “ 24 0.0006
“ 31 0.0006 Apr. 9 0.0005
“ 9 0.0006 “ 12 0.0005
The hemagglutinin was added to 0.5 cc. portions of a 2.5 per cent suspen- sion of washed, rabbit erythrocytes in a total volume of 1 cc.
TABLE II.
Agglutinability of Erythrocytes of Different Species.
Date of experiment.
1923
Mar. 15 “ 28 “ 30 “ 31
Apr. 13 Mar. 20
Apr. 15 8‘ 15 “ 29
“ 29 May 4
(‘ 9 Mar. 20
Apr. 17 “ 17
Kind of erythrocyte.
Rabbit. Man.
“
I‘ ‘I
Duck. Dog.
“
“
Mouse. “
Rat.
Chicken. “
‘<
Minimum effective concentra- tion of agglutinin.
mg.
0.0006 0.0007 0.0008
0.0007 0.0007 0.0008
0.0008 0.0008 0.0009
0.002 0.002
0.003 0.006
0.006 0.006
To 0.5 cc. portions of 2.5 per cent suspensions of the washed erythrocytes in isotonic sodium chloride were added 0.5 cc. portions of isotonic sodium
chloride containing the quantity of hcmagglutinin to be tested.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 453
rocytes. In Table II the results of comparing the agglutination titers of different species of animals are shown.
From Tables I and II it can be seen that the dilution at which the agglutinin is still active is, for rabbit erythrocytes, 1 to 6,000,OOO or by analogy with ordinary immunological terminology this figure could be called its titer, since clumping is effected in a total volume of 1 cc. of solution containing only 0.0006 mg. of agglutinin by dry weight. It is active in lower concentration than the purified bean agglutinin “phasin” as reported by Wien- haus in 1909. His product was, however, only partially soluble, having suffered considerable denaturation during the process of preparation. It seems probable that the soluble fraction of his material was highly active. In contrast to Wienhaus’ phasin, the bean agglutinin reported here gave remarkably similar results with the washed corpuscles of the various species tested-es- pecially with those of man, rabbit, dog, and duck. The eryth- rocytes of the hen, more difficult to clump than the other species tried, required only 10 times as much agglutinin as was necessary for rabbit.
In&hence of Medium upon Hemagglutination.
In&Siting Factors.-Not only artificial media, but the natural medium of the erythrocytes, i.e. blood serum, shows a strong in- fluence upon agglutination of the corpuscles. This was noted by Landsteiner and Raubitschek in their early work. Raubitschek (Kraus and Levaditi, 1911) was of the opinion that the inhibiting influence of serum is due to the presence of its proteins. He noted the selective affinity of erythrocytes for certain substances, such as peptones and albumoses. Agglutinated cells shaken and incubated with peptone solution became unagglutinated. Our own observation supports Raubitschek’s conclusion regarding the retarding influence of protein material upon the agglutination re- action. Table III indicates the effect of egg white upon the action of the agglutinin preparation. In these experiments the standard method already described was not employed. Whole defibrinated blood in 0.1 cc. portions was treated with solutions of bean hemag- glutinin dissolved in 0.89 per cent sodium chloride in a total volume of 1 cc. Column 1 shows the effect of the hemagglutinin alone; the results in Column 2 were secured when egg white was
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

454 Plant Hemagglutinins
introduced just after the addition of the hemagglutinin, whereas in experiments in Column 3 the egg white was added just prior to the addition of hemagglutinin. When the corpuscles were first treated with egg white they failed to agglutinate in the presence
TABLE III.
InJEuence of Egg White upon Agglutination of Whole, DeJibrinated Blood.
mg.
0.05 0.06 0.07 0.08 0.09
Effect upon hemagglutination.
Blood +&;magglu- Blood + hemagglu- Blood + 1 cc. egg white tinin + 1 cc. egg white. + hemagglutinin.
(1) (2) (3)
- - -
+ - -
+ + -
+ + -
+ + -
TABLE IV.
Inhibitory Influence of Serum upon Hemagglutination.
Suspension.
vol. per cent
2.5 2.5 2.5 5.0 5.0 5.0 5.0
-
-
Kind of erythrocytes.
Rabbit. “ “ “
Duck. “ “
7-
None.
Rabbit. Turtle. Rabbit. None. Duck. Hen.
cc.
0.012 0.012 0.025
0.025 0.025
I 1
_-
-
Winimum effec- ,ive quantity of
agglutinin.
w.
0.0006 0.002 0.006 0.003 0.0008 0.002 0.006
0.5 cc. portions of a suspension of washed corpuscles were used in a total volume of 1 cc. with and without additions of equal volumes of serum, the suspension being diluted with isotonic saline solution to a total volume of 1 cc. Before dilution to final volume, serum was added in a proportion equivalent to the actual volume of the corpuscles.
of far greater quantities of hemagglutinin than were readily effec- tive without the egg white. When the egg white was added after the corpuscles and the agglutinin had had even a brief opportunity to act the influence was less marked.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 455
The effect of adding traces of ‘blood serum to washed corpuscles can be seen from Table IV, a summary of a series of experiments in which washed corpuscles suspended in saline were compared with those to which serum of the same or foreign species had been added. Serum of the turtle and the hen were particularly in- hibitory. For rabbit serum the ratio between the requisite
Injluence of Sodium Chloride upon Hemagglutination.
Date of experiment. Erythrocyte suspen- sion. Agglutinin added.
199s vol. per cent VW.
Apr. 10 5.0 0.002 “ 10 5.0 0.005 “ 10 10.0 0.005
May 15 2.5 0.02 “ 17 2.5 0.03 “ 17 2.5 0.04 “ 16 2.5 0.05
Apr. 9 2.5 0.05 May 17 2.5 0.08
“ 17 2.5 0.08 “ 16 2.5 0.1 “ 17 2.5 0.1 “ 16 2.5 0.15
- Minimum effective
weight of N&l.
ml.
>0.7 0.4 0.4 0.3 0.2 0.2 0.2 0.5? 0.2 0.2* 0.2 0.2 0.2
Summary of a series of experiments in each of which the amount of hemagglutinin was kept constant and sodium chloride was varied in differ- ent test samples until agglutination was obtained. Samples consisted of 0.5 cc. portions of a suspension of washed corpuscles in isotonic sucrose, diluted to a total volume of 1 cc. with an isotonic mixture containing known weight of hemagglutinin and the quantity of sodium chloride to be tested.
*After the corpuscles of this series had been allowed to react with agglu- tinin for 4 hour, they were washed with sucrose solution to remove excess agglutinin, resuspended in sucrose solution, and titrated, with the result indicated.
amounts of agglutinin for clumping equivalent volumes of eryth- rocytes in whole, diluted blood and in sodium chloride solution was approximately 5 : 3.
Favoring InJluence.-Electrolytes are of course factors in the medium essential for hemagglutination. Rona and Gyorgy (1920) concluded from their work on the influence of electrolytes upon
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

456 Plant Hemagglutinins
ricin agglutination that a definite cation series could be estab- lished by arranging the salts in the order of their values. With the hemagglutinin reported here, it was found that when the concentration of electrolyte was greatly lowered, excessively large amounts of agglutinin were required. Thus, corpuscles washed three times in 0.25 M sucrose solution required 10 times as much hemagglutinin as those washed and suspended in isotonic sodium chloride solution. After washing the cells twelve times in isotonic sucrose solution, it was no longer possible to cause agglutination with any concentration of protein used.
In order to study the relationship between concentration of hemagglutinin and quantity of electrolyte, a series of media was prepared containing mixtures of varying concentrations of sodium chloride and sucrose selected to keep them isotonic with rabbit erythrocytes. These were used in a series of “titration experi- ments” summarized in Table V. To 0.5 cc. portions of a 2.5 per cent suspension of rabbit erythrocytes in isotonic sucrose were added 0.5 cc. portions of sucrose solution containing the amount of agglutinin indicated with varying quantities of sodium chloride. Table V records the minimum weight of sodium chloride in the presence of which agglutination is brought about in each case. The “titration” was not carried far enough to determine the concentration of sodium chloride necessary to permit agglutina- tion of the corpuscles with 0.002 mg. of protein. It was greater than 0.72 mg. of sodium chloride. As the weight of hemag- glutinin increased, the requirement for sodium chloride decreased until the value of hemagglutinin reached a certain level, i.e. 0.02 mg. Beyond this value, the quantity of sodium chloride required was shown to be constant-O.2 mg. Isotonic salt solution would con- tain in an equal volume about 10 times this concentration of sodium chloride.
Chemical Alteration of the Protein.
The question as to whether the hemagglutinating property is dependent upon the integrity of the protein has been approached by a study of the effects of chemical changes upon the latter.
Heat Coagulation.-The preparation studied could be denatured by heat when in solution. Below 68” the protein did not separate from aqueous solution. With slow heating, coagulation would
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 457
begin at 68”, but the greater part of the coagulable protein sepa- rated out between 75” and 81”. The filtrate from the coagulum at the higher temperature, showed a greatly lowered potency to agglutinate rabbit erythrocytes. At 100” coagulation became so complete that no agglutinating power remained. Heating the dry preparation to the same temperature was without influence upon solubility or potency of the material unless the heating
TABLE VI.
InfEuence of Peptic Digestion upon Activity of Hemagglutinating Protein.
Minimum effective quantityt;;iemagglu-
mg.
Untreated protein..................................... 0.0008
Dissolved in pepsin-HCl and neutralized at once.. . . i
0.0008
0.0007 After 1 hr. digestion.. . . . . . . . . . . . . . . . . . . . . . 0.002
‘L 14 hrs. “ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0.004
“ 3 days “ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0.007 “ 6 “ “ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Activity lost.
To 0.1 gm. portions of the protein weighed out exactly and placed in
test-tubes were added 2 cc. of 0.1 N HCI solution and 2 cc. of 1 per cent pepsin solution with 2 cc. of water. The tubes were stoppered and incu- bated in a constant temperature water bath at 37.5” for the periods of time
indicated in the summary. At the end of the incubation period, the solu-
tion was exactly neutralized by adding the calculated quantity of sodium hydroxide. The sodium chloride content was increased to make the solu-
tion exactly isotonic to rabbit erythrocytes after dilution to 20 cc. with water. Further dilutions were made with isotonic sodium chloride and the agglutinating potency of the preparations with rabbit erythrocytes were
determined by the method already outlined. The minimum effective weight of hemagglutinin was expressed in terms of dry weight of the protein before digestion.
process was continued for a number of hours. Then the change was only slight. After 4 days of heating at 100” the agglutinating potency was lowered to one-thirtieth that of the original protein. Wienhaus has reported a similar finding with his phasin; heating the dry preparation for 2$ hours at 130” caused only a partial destruction of potency.
Acid Hydrolysis.-Short exposures to weak acid produced no
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

458 Plant Hemagglutinins
apparent effect upon the protein. When 0.1 gm. portions were treated with 1 cc. of 0.1 N hydrochloric acid and neutralized within 5 minutes, solubility and agglutinating potency were unaffected. Upon longer exposures, the protein gradually separated out in an insoluble form with an accompanying loss in agglutinating value. Thus after 3 days exposure to 0.1 N hydrochloric acid, the requi- site weight of protein for clumping a given volume of erythrocyte suspension was increased to 5 times the amount required before the exposure to acid.
Peptic Digestion.-This also destroys the agglutinating po- tency. Hemagglutinating protein was dissolved in pepsin-HCI. When the solution was immediately neutralized no change in the minimum effective hemagglutinating dose of the protein could be detected. With the progress of digestion, however, the agglutinat- ing activit,y of the solution was lowered; and at the end of a considerable period of incubation, it seemed to be entirely lost. The preceding summary of protocols (Table VI) illustrates these observations.
Mode of Action of Hemagglutinin.
There are indications that the hemagglutinins of plant origin resemble those of the serum. Comparable reactions appear to affect both groups of substances similarly. Both serum and plant hemagglutinins require electrolyte for their action; in both cases, a “fixation” reaction occurs between the agglutinin and the oor- puscle prior to the flocculation which is dependent upon some physicochemical action of salts. By referring to Table V, the reader will note that rabbit erythrocytes may be washed with sucrose solution after exposure to hemagglutinin and still retain enough of the “fixed” material to clump upon addition of sodium chloride. The electrolyte is needed to complete the reaction. Wells (1925) has spoken of agglutination as a process of “salting out” in which electrolytes are of great importance. He points out further that one might think of the antigen-antibody (or agglutinin-corpuscle) reaction as resulting in the formation of an electrically amphoteric, colloidal suspension. The ions of elec- trolyte, then, cause precipitation by discharging the particles un- equally. Evidence bearing upon this theory has been given by the work of Coulter (1920) who determined the isoelectric point
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel
TABLE VII.
459
Variations in Amount of Hemag MI; utinin Fixed by Corpuscles.
Date of Kind of experiment. erytbrooyte. For 3
successive portions.
Mar. 15 “ 18 “ 24
Apr. 9 “ 13
Mar. 12 rlpr. 3
“ 12 Mar. 25
“ 18
Rabbit.
sol. per cent mg. mg. m7.
2.5 0.0006 0.006 0.008
0.0006 0.006 0.006 0.0006 0.002* 0.0006 0.006 0.008 0.0005 0.006 0.01
5.0 0.0007 0.009 0.01 0.0005 0.003 0.003 0.0008 0.006 0.01
10.0 0.0009 0.003 20.0 0.005 0.008
“ 28 -4pr. 13
Mar. 22 Apr. 17
Man. 2.5 0.0007 0.007 5.0 0.002 0.02 0.05t
Chicken. 2.5 0.006 0.01 0.006 0.03 0.08 0.006 0.04
Mar. 20 Duck. 2.5 0.0006 0.006 0.006 20.0 0.005 0.03f
Apr. 15 Dog. 2.5 0.0008 0.006 0.009 0.0008 0.007
5.0 0.003 0.009 0.03t
To 0.5 cc. portions of erythrocytes of suspensions indicated in isotonic sodium chloride were added increasing quantities of hemagglutinin; and the volume was made up to 1 cc. with isotonic sodium chloride. After the corpuscles had clumped and sedimented, 0.5 cc. of the supernatant medium was removed and tested for agglutinating potency with a fresh suspension of corpuscles. Thus the amount of protein required in the initial dose for clumping two successive and three successive portions of corpuscles was determined.
dinimum effective quantity of agglutinin:
*This value was obtained by using a 1.25 per cent suspension of corpuscles.
t This value was obtained by using a 2.5 per cent suspension of corpuscles.
1 This value was obtained by using a 10.0 per cent suspension of corpuscles.
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

460 Plant Hemagglutinins
for normal sheep erythrocytes and showed that by the process of sensitization, the isoelectric point was moved in the direction of neutrality. Thus flocculation was made possible nearer neu- trality, since particles in suspension are most easily aggregated and sedimented at their isoelectric points.
The two types of agglutinin also have similar fixation processes. According to the well established rule of Eisenberg and Volk (foreseen by Bordet) “for the same mass of cells, the absolute quan- tity fixed is directly proportional to the concentration of antibody; the relative quant,ity inversely” (Wells, 1925). In this connection, we found it interesting to calculate from data collected the maxi- mum quantity of our agglutinin held by the cells of a given volume of suspension and to compare this with the minimum effective quantity of agglutinin for this same cell mass. With a uniform suspension of corpuscles in a series of tubes, the quantity of ag- glutinin was varied from the minimum requirement upward. By the standard procedure, not until 0.006 mg. or 10 times the mini- mum effective concentration had been used was there sufficient agglutinin free in the supernatant medium to cause clumping of a second unit of corpuscles (see Table VII). A liberal allowance for incomplete removal of unbound agglutinin being made, the bound fraction was at least 8 times the minimum required for clumping the mass of cells used. Corpuscles, then, do not combine with one and only one given quantity of a plant hemagglutinin. The amount “fixed” depends upon the quantity available in the medium.
Chemical Nature of Hemagglutinin.
The interpretation given by Eisler and Portheim (1926) to results which they obtained from a study of the hemagglutinin of Phaseolus multiJEorus was that the active substance was not itself protein but was adsorbed onto the albumin fraction of the bean. They compared the increase in agglutinating value of the maturing seed with the increase in quantity of the various protein fractions and found that the former greatly exceeded the increase in total protein or in the albumin fraction itself (which contained agglutinin in higher concentration than any of the other fractions). In contrast to this conclusion, all the evidence afforded by the work reported here points toward the protein nature of the
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel
hemagglutinin. We observed that the various phases of the denaturation process affecting the albumin preparation were ac- companied by diminution of the agglutinating potency. Thus, the slow, denaturing effect of acids would in time destroy the agglutinin as completely as would the rapid process of heat coagu- lation. Neither was it possible to digest the protein by enzyme action without a concomitant loss of agglutinating potency. One would not necessarily expect this to happen were the agglutinin merely adsorbed onto the protein since it might then be liberated and be made available through the process of digestion. Peptic digestion changed the effectiveness of the agglutinin along with the elastic influence upon the protein molecule. By a peptic hydrolysis of 6 days duration, the agglutinin was rendered inert. Since from proteins with an agglutinating quality an individual hemagglutinating fraction has never been separated, and since chemical changes in the protein itself are accompanied by changes inthe agglutinating potency, there is no reason for assuming two closely associated entities rather than one endowed with an unusual property. Moreover, the protein itself is reactive in such low concentration that if the agglutinin were a separate entity oc- curring as a contaminant, its effective concentration would be astonishingly minute.
Practical Considerations.
The outcome of this study suggests that hemagglutinating preparations of the sort that we have investigated may have a larger field of usefulness in the preparation of therapeutic serums than has hitherto been accorded to them. It should prove com- paratively simple and inexpensive to prepare in adequate quanti- ties readily soluble specimens of the bean albumin that can be duly sterilized and preserved in dry form for long periods ready for instantaneous use. The stoichiometric relations studied indi- cate that the quantities required for corpuscular agglutination are comparatively small. Furthermore, through preliminary tests on the species of blood to be agglutinated, it should be possible to estimate for large lots the dosage so selected that an excess nf unfixed hemagglutinating protein will not remain in the serum. Danger of anaphylactic reactions through the retention of a
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Plant Hemagglutinins
foreign protein like the bean albumin in a therapeutic serum ought thus to be successfully avoided.
SUMMARY.
A non-toxic, highly potent, soluble, hemagglutinating protein having the characteristics of an albumin was prepared in dry form from navy beans (Phaseolus communis). A quantitative, macroscopic method for measuring hemagglutination was devised and used for studying the variables affecting the reaction. In- dispensability of electrolytes and the inhibiting influence of certain proteins, notably those of egg albumin and the serum proteins were demonstrated. Chemical changes in the protein which led to denaturation or hydrolytic cleavage were shown to be ac- companied by a lessened hemagglutinative potency. The mode of action, the chemical nature, and some practical aspects relating to the application of the procedures to the production of thera- peutic serums are discussed.
BIBLIOGRAPHY.
Berczeller, L., and Wastl, H., Biochem. Z., 142, 524 (1923). Coulter, C. B., J. Gen. Physiol., 3,309 (1920).
Dorset, M., andHenley, R. R., J. Agric. Research, 6, 333 (1916). Ehrlich, P., Fortschr. Med., 15,41 (1897).
Eisler, M., and Portheim, L., 2. Immunitiitsforsch., Orig., 1, 151 (1909). Eisler, M., and Portheim, L., Ber. deutsch. bot. Ges., 29,419 (1911). Eisler, M., and Portheim, L., Centr. Bakt., 1. AU., Orig., 66, 309 (1912).
Eisler, M., and Portheim, L., 2. Immunitiitsforsch., 47,59 (1926). Gardner, A. D., Lancet, 1,21 (1919). Gates, F. L., J. Am. Med. Assn., 77,2054 (1921).
Gates, F. L., J. Ezp. Med., 35, 63 (1922). Hadley, P., J. Immunol., 2, 463 (1916). Karrer, P., Smirnoff, A. P., Ehrensperger, H., van Slooten, J., and Keller,
M., 2. physiol. Chem., 135, 129 (1924).
Kobert, R., Lehrbuch der Intoxikationen, Stuttgart (1902). Kolmer, J. A., Infection, immunity and biological therapy, Philadelphia
and London (1923). Kraus, R., and Levaditi, C., Handbuch der Technik und Methodik der
Immunitatsforschung, Jena (1911). Landsteiner, K., and Raubitschek, H., Centr. Bakt., f. AU., Orig., 46,
660 (1908).
Marcusson-Begun, H., 2. Immunittitsforsch., 46,49 (1926). Mend& L. B., Arch. Jisiol., 7, 168 (1909). Mudd, S., J. Immunol., 13, 113 (1927).
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

V. R. Goddard and L. B. Mendel 463
Osborne, T. B., Mendel, L. B., and Harris, I. F., Am. J. Physiol., 14, 259 (1905).
Rona, P., and Gyiirgy, P., Biochem. Z., 106,120 (1920). Russ, V. K., and Oesterlin, E., Biochem. Z., 114,258 (1921). Schneider, E. C., J. Biol. Chem., 11, 47 (1912). Wells, H. G., The chemical aspects of immunity, New York (1925). Wienhaus, O., Biochem. Z., 18,228 (1909).
by guest on May 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Verz R. Goddard and Lafayette B. MendelBEAN
PREPARATION FROM THE NAVYSPECIAL REFERENCE TO A
PLANT HEMAGGLUTININS WITH
1929, 82:447-463.J. Biol. Chem.
http://www.jbc.org/content/82/2/447.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
ml#ref-list-1
http://www.jbc.org/content/82/2/447.citation.full.htaccessed free atThis article cites 0 references, 0 of which can be by guest on M
ay 25, 2018http://w
ww
.jbc.org/D
ownloaded from


















