
Physiological Stress Reactivity in Late Pregnancy613938/FULLTEXT01.pdf · Physiological Stress...
72
ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2013 Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 887 Physiological Stress Reactivity in Late Pregnancy CHARLOTTE HELLGREN ISSN 1651-6206 ISBN 978-91-554-8636-5 urn:nbn:se:uu:diva-197441
-
Upload
nguyenxuyen -
Category
Documents
-
view
214 -
download
0
Transcript of Physiological Stress Reactivity in Late Pregnancy613938/FULLTEXT01.pdf · Physiological Stress...

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2013
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 887
Physiological Stress Reactivity inLate Pregnancy
CHARLOTTE HELLGREN
ISSN 1651-6206ISBN 978-91-554-8636-5urn:nbn:se:uu:diva-197441

Dissertation presented at Uppsala University to be publicly examined in Sal X,Universitetshuset, Biskopsgatan 3, Uppsala, Friday, May 17, 2013 at 09:15 for the degree ofDoctor of Philosophy. The examination will be conducted in English.
AbstractHellgren, C. 2013. Physiological Stress Reactivity in Late Pregnancy. Acta UniversitatisUpsaliensis. Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty ofMedicine 887. 70 pp. Uppsala. ISBN 978-91-554-8636-5.
During pregnancy, the basal activity is increased in both of our major stress response systems:the sympathetic nervous system and the hypothalamic-pituitary-adrenal axis. At the sametime, the reactivity towards stressors is reduced. These alterations sustain maternal and fetalhomeostasis, and are involved in the regulation of gestational length. Although the feto-placentalhormone synthesis produces the main endocrinological changes, also the central nervous systemundergoes adaptation. Together, these profound adjustments have been suggested to makewomen’s mental health more vulnerable during pregnancy and postpartum period. The aim ofthis thesis was to examine factors connected to physiological stress responses during the latepregnancy in relation to pain, labour onset, emotional reactivity, and mental health.
The first study examined the pain and sympathetic response during cold stress, in relationto time to delivery. Women with fewer days to spontaneous delivery had lower sympatheticreactivity, while no pain measure was associated with time to delivery.
In the second study, acoustic startle response modulation was employed to study emotionalreactivity during late gestation, and at four to six weeks postpartum. The startle responsewas measured by eye-blink electromyography, while the participants watched pleasant andunpleasant pictures, and positive and negative anticipation stimuli. A significant reduction instartle modulation by anticipation was found during the postpartum assessment. However, nostartle modulation by pleasant, or unpleasant, pictures was detected at either time-point.
The serum level of allopregnanolone, a neurosteroid implied in pregnancy-inducedhyporeactivity, was analysed in relation to self-reported symptoms of anxiety and depression.Although the participants reported low levels of depression, the women with the highestdepression scores had significantly lower levels of serum allopregnanolone. There was nocorrelation between allopregnanolone and anxiety scores.
In the fourth study, the cortisol awakening response was compared between women withdepression during pregnancy, women with depression prior to pregnancy, and women who hadnever suffered from depression. No group differences in cortisol awakening response duringlate pregnancy were found.
The results are in line with the previously described pregnancy-induced hyporesponsiveness,and add to the knowledge on maternal stress hyporeactivity, gestational length, and maternalmental health.
Keywords: acoustic startle response, allopregnanolone, antenatal, anxiety, cold pressor test,cortisol, cortisol awakening response, depression, electrodermal, estradiol, hypothalamic-pituitary-adrenal axis, postpartum, pregnancy, progesterone, skin conductance, stress,sympathetic nervous system
Charlotte Hellgren, Uppsala University, Department of Women's and Children's Health,Obstetrics and Gynaecology, Akademiska sjukhuset, SE-751 85 Uppsala, Sweden.
© Charlotte Hellgren 2013
ISSN 1651-6206ISBN 978-91-554-8636-5urn:nbn:se:uu:diva-197441 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-197441)

“nature, who delights in muddle and mystery, so that even now (the first of November 1927) we know not why we go upstairs, or why we come down again”
Orlando, Virginia Woolf


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Hellgren C., Åkerud H., Jonsson M., Sundström-Poromaa I. (2011) Sympathetic reactivity in late pregnancy is related to la-bour onset in women. Stress 14, 627-633.
II Hellgren C., Bannbers E., Åkerud H., Risbrough V., Sund-ström-Poromaa I. (2012) Decreased startle modulation during anticipation in the postpartum period in comparison to late pregnancy. Archives of Women’s Mental Health 15, 87-94.
III Hellgren C., Åkerud H., Skalkidou A., Bäckström T., Sund-ström-Poromaa I. Low serum allopregnanolone is associated with symptoms of depression in late pregnancy. Submitted.
IV Hellgren C., Åkerud H., Skalkidou A., Sundström-Poromaa I. Cortisol awakening response in late pregnancy in women with previous or ongoing depression. Submitted.
Reprints were made with permission from the respective publishers.


Contents
Introduction ................................................................................................... 11 Important endocrinological alterations in normal pregnancy and puerperium ............................................................................................... 11
Physiological and neuroendocrinological changes in pregnancy ........ 12 Physiological and neuroendocrinological changes in the postpartum period ................................................................................................... 13
The stress response system and its regulation during pregnancy and puerperium ............................................................................................... 14
The maternal HPA axis ........................................................................ 15 The maternal autonomic nervous system............................................. 16
Spontaneous labour onset ......................................................................... 18 Pain sensitivity during pregnancy and in the postpartum period.............. 19 Mood, cognition and mental health during pregnancy ............................. 20
Antenatal and postpartum depression and anxiety .............................. 20 Stress response regulation in depression and anxiety............................... 21 The skin conductance response ................................................................ 22 The cold pressor test ................................................................................. 23 The acoustic startle response and its modulation ..................................... 23 Allopregnanolone – in stress, mood, and pregnancy ................................ 24 The cortisol awakening response ............................................................. 26
Aims .............................................................................................................. 27
Materials and Methods .................................................................................. 28 Paper I-III ................................................................................................. 28
Psychiatric evaluation by self-report questionnaires and structured interview .............................................................................................. 28
Paper I ...................................................................................................... 29 Paper II ..................................................................................................... 30 Paper III .................................................................................................... 31 Paper IV.................................................................................................... 31
Summary of results ....................................................................................... 33 Paper I ...................................................................................................... 33 Paper II ..................................................................................................... 35 Paper III .................................................................................................... 37 Paper IV.................................................................................................... 38

Discussion ..................................................................................................... 39 Methodological considerations................................................................. 39
Participant selection bias ..................................................................... 39 Cross-sectional design ......................................................................... 39 Subjective and objective assessments of stress, pain, and affect ......... 40
Sympathetic stress response and labour onset .......................................... 41 β-endorphin, pain, and sympathetic response .......................................... 42 Startle modulation across pregnancy and postpartum .............................. 42 Allopregnanolone and mood during late pregnancy ................................ 44 Cortisol awakening response in currently and previously depressed pregnant women ....................................................................................... 45 Clinical implications and future perspectives .......................................... 47
Maternal stress systems in pregnancy complications .......................... 47 Maternal stress systems and fetal development ................................... 47 Reflections on cross species translation .............................................. 48
Conclusions ................................................................................................... 49
Sammanfattning på svenska .......................................................................... 50 Delstudie I............................................................................................ 50 Delstudie II .......................................................................................... 51 Delstudie III ......................................................................................... 51 Delstudie IV ......................................................................................... 52
Acknowledgments......................................................................................... 53
References ..................................................................................................... 56

Abbreviations
ACTH Adrenocorticotropic hormone ANOVA Analysis of variance AUC Area under the curve AVP Vasopressin BDNF Brain-derived neurotrophic factor BMI Body mass index CAR Cortisol awakening response CRH Corticotropin releasing hormone DSM-IV Diagnostic and statistical manual of mental disorders, 4th
edition EMG Electromyography EPDS Edinburgh postnatal depression scale dB Decibel GABA γ-aminobutyric acid HPA axis Hypothalamic-pituitary-adrenal axis hPL Human placental lactogen IAPS International affective picture system MADRS-S Montgomery-Åsberg depression rating scale – self as-
sessment MHPG 3-methoxy-4-phenyl glycol MINI Mini international neuropsychiatric interview pCRH Placental corticotropin releasing hormone POMC Pro-opiomelanocortin PVN Paraventricular nucleus of the hypothalamus SD Standard deviation SEM Standard error of the mean SPSS Statistical package for the social sciences SSRI Selective serotonin reuptake inhibitor STAI-S, -T Spielberger state trait anxiety inventory (-state, -trait) VAS Visual analogue scale


11
Introduction
Important endocrinological alterations in normal pregnancy and puerperium During pregnancy, the placenta produces increasing amounts of steroid hor-mones, peptides, and other factors which reset the maternal homeostasis [1]. The endocrine adaptations for childbirth and nursing result in the suspension of ovulation and the development and growth of the uterus, the placenta, and the foetus. In order to achieve this, readjustments of maternal immunology, stress physiology, and metabolism are required. The changes involve hor-mones and neurotransmitters which are known to have roles in mental health and disease. Several of the hormonal alterations have been suggested to alter women’s mood, cognition, and prevalence of psychiatric disorders during pregnancy and the postpartum period [2-4]. However, it is important to bear in mind that the majority of childbearing women maintain a good mental health at this turbulent time in their lives; the complex pattern of neuroendo-crine change must consequently be rich in elements which protect against mood disturbances.
It is primarily the placenta and the developing fetal endocrine system which govern the endocrinology of pregnancy. The feto-placental unit re-leases increasing amounts of hormones into the maternal bloodstream as it grows - a supply that is physically removed at delivery. A number of the maternal endocrine changes therefore follow a common trail. Oestrogens, progesterone, human placental lactogen (hPL), testosterone, corticotrophin releasing hormone (CRH), and cortisol all rise continuously through the 40 weeks of pregnancy, followed by a drastic drop at parturition. [1, 5-9].
All maternal organ systems have to adapt; this leads to well known preg-nancy side-effects like fatigue and sensitivity to infections, and less known effects, like enlarged adrenal glands [10, 11]. Sometimes the adaptations cause unwanted complications, such as hypertension [12]. The hormonal changes of pregnancy and childbirth also have effects on our major stress systems; the sympathetic nervous system and the hypothalamic-pituitary-adrenal (HPA) axis.

12
Physiological and neuroendocrinological changes in pregnancy The feto-placental hormone production regulates several pregnancy specific changes. Human placental lactogen and cortisol act to increase glucose and fatty acids in the circulation to meet the fetal energy demand [1]. Placental production also increases circulating levels of leptin, an appetite and energy expenditure regulating hormone with possible involvement in mood disor-ders [13]. Another important metabolic alteration is slightly decreased levels of free thyroid hormones [14, 15]. Thyroid hormones enable tissue energy consumption and clinical hypothyroidism is well known to cause depressive symptoms [15]. The net effect of the metabolic changes is a gradual increase of around 30% of the pre-pregnancy maternal basal metabolic rate [14, 16].
The high oestrogen level stimulates the renin-angiotensin system to in-crease maternal blood volume by around 40% [17]. As sympathetic nervous system tone increases and parasympathetic tone decreases, maternal heart rate and blood pressure also change. Some women develop a pathologically high sympathetic tone and pregnancy-induced hypertension [12].
Regulation of the maternal immunology is another important function of the steroid hormone surges [17]. As the foetus is perceived as non-self, the maternal immune system must be tranquil enough to allow the foetus to thrive, but it is still crucial to maintain a good defence against pathogens. Cortisol is routinely called immunosuppressive, but cortisol, rather, changes the balance between the two main components of the immune system. Thus, in parallel with the hypercortisolism of pregnancy, there is an up-regulation of the generalised inflammatory response, (i.e. monocytes and granulocytes) while there is a down-regulation of the antigen-specific component (i.e. T-cells) [11]. The placental tissue produces a plethora of cytokines which have been shown to interact with placental hormone synthesis. Cytokine signaling has also been implied in uterine contractions and rupture of membranes [18]. Long-term stress is known to compromise immune function, and the immu-nological aspect of depression has gained increasing research interest [19]. Accordingly, the immune adaptations of pregnancy, and immune-HPA inter-action, have been suggested to be of neuropsychological significance [20].
There are few studies on neurotransmitter adaptations inside the human central nervous system during pregnancy. However, analyses of cerebrospi-nal fluid (CSF) from women undergoing elective caesarean section show significantly lower levels of γ-amino-butyric acid (GABA), as well as the norepinephrine metabolite 3-methoxy-4-phenyl glycol (MHPG), than CSF from non-pregnant women [21]. Indirect evidence of central nervous changes also include decreased maternal serum levels of brain-derived neural growth factor (BDNF), both before and after delivery, and low BDNF is correlated with decreased serotonin levels in serum at both time-points [22]. A reduced availability of serum tryptophan, a serotonin precursor, is also evident during pregnancy [23]. Low GABA, serotonin, and BDNF are all considered bio-

13
markers for depression [21, 22], while most studies report CSF MHPG con-centrations to be equal in control and depressed populations [24]. The indi-rect human evidence is supported by studies in rodents, where pregnancy has been found to decrease serotonin, GABA, and norepinephrine content in brain tissue [25-27].
Physiological and neuroendocrinological changes in the postpartum period The serum concentrations of feto-placental hormones decline fast after partu-rition, and instead of abnormally high steroid hormones, postpartal oestro-gen, progesterone, and cortisol levels are below the normal non-pregnant/non-postpartal range [6, 8, 9]. Low oestradiol and progesterone concentrations persist when breast-feeding is continuous and prolactin is high [1]. The low HPA reactivity of pregnancy is still reduced during the hypocortisolism of lactation [28]. Ovulatory cycles with regular oestrogen and progesterone levels, and normal HPA axis reactivity, resume within a few weeks to months of non-lactation [29] [28]. Serum prolactin returns to normal within a few months after breast-feeding cessation [28]. Other hor-mones, like leptin, return to normal within weeks after delivery and do not depend on lactation status [6, 30].
To prepare for lactation is an important goal for the endocrine changes during pregnancy. Prolactin starts increasing during early pregnancy [31], induced by oestrogens [1]. CSF and plasma levels of prolactin in pregnant women at term are around ten times higher than in non-pregnant women [21]. When progesterone is withdrawn at parturition, the build-up of prolac-tin can induce milk production [31]. Furthermore, plasma prolactin crosses the blood brain barrier [32] and is responsible for decreasing postpartum stress responses, not only in the HPA axis activity, but also in the oxytocin “anti-stress” system [33, 34]. Prolactin is also immunomodulatory [35]. The immunological state during the postpartum period is characterized by an initial inflammatory response to delivery, and an exaggerated response has been postulated to increase the risk for postpartum depression [36].
The posterior pituitary peptide oxytocin is responsible for the ejection of milk in response to suckling. Oxytocin levels are higher in plasma, but not in CSF in pregnant compared to non-pregnant women [21]. Average plasma oxytocin does not increase from pregnancy levels in the postpartum period [37], although it transiently rises at nursing [38]. Oxytocin signalling within the brain is important for well-being and social attachment and has therefore been researched in relation to puerperal mood [39]. Since oxytocin is a nega-tive regulator of the HPA axis and plasma levels rise with nursing or merely holding an infant, oxytocin is believed to support maternal calm during the postpartum and lactation period [39]. However, the associations between

14
oxytocin, nursing, bonding, and emotional calm are not obvious, as oxytocin release within the brain seems to be independent of suckling [40] and periph-eral oxytocin does not seem to readily cross the blood-brain barrier [21, 41].
Several studies have described the normal postpartum as a depression-like state with reference to neuroendocrinological parameters. Serum indicators of serotonergic activity in healthy women a few days after delivery are low [42, 43], although the serotonergic indices seem to be restored to late pregnancy values at six weeks postpartum [43]. A few brain imaging studies have also been made in different stages of the postpartum period. For in-stance, magnetic resonance spectroscopy examination has indicated de-creased cortical GABA concentrations in healthy women several months into the postpartum period [44], while a positron emission tomography study has revealed elevated monoamine oxidase A at two weeks postpartum, i.e. in-creased breakdown of serotonin, norepinephrine, and dopamine [45].
The stress response system and its regulation during pregnancy and puerperium Both the HPA axis and the sympathetic nervous system are organized via signals from the hypothalamus, particularly the paraventricular nucleus (PVN) [46]. The PVN receives input from several other parts of the brain and the coordinated sympathetic (and parasympathetic) output from the PVN travels as a nerve signal to the target organs, including the adrenal medulla [47]. The sympathetic nervous system thus responds within seconds when increased attention or action is required, and reverts rapidly as the stimulus disappears [47]. The response includes cardiovascular changes, as well as increased sweat gland activity [1]. The stimulation of the adrenal medulla releases epinephrine into the circulation. Epinephrine increases heart rate, relaxes smooth muscles in the respiratory tract, and improves energy avail-ability [1].
The HPA axis reacts relatively slowly to stressful input. An increase in CRH and vasopressin (AVP) from the paraventricular nucleus of the hypo-thalamus causes a rise in adrenocorticotropic hormone (ACTH) secretion from the anterior pituitary, which in turn activates the adrenal cortices to release cortisol [1]. The peak in plasma and free salivary cortisol can be ob-served about 25 minutes after a laboratory stressor and baseline cortisol is not reached until about an hour after the stressful stimulus is ended [48]. Cortisol directs the metabolism towards increased acute energy usage and is immunomodulatory [1].
ACTH is not secreted in its bioactive form, but is cleaved from a larger polypeptide called pro-opiomelanocortin (POMC) [1]. Another POMC

15
cleavage product is β-endorphin, an analgesic opioid hormone, i.e. a balanc-ing element integrated in the HPA axis response [1].
The maternal HPA axis Optimal fertility is dependent on a balanced hypothalamic-pituitary adrenal axis. Increased levels of glucocorticoids have a direct inhibitory effect on reproductive tissues and will, at the same time, inhibit hormone production at all levels of the hypothalamic-pituitary-gonadal axis [49]. This means that long-term stress, or highly stressful life events, can impair fertility through amenorrhea or reduced sperm count [50].
Feto-placental production causes the baseline levels of HPA axis hor-mones to become elevated during pregnancy [8, 12, 51] so that in the third trimester, cortisol levels are around three times non-pregnant levels [8]. In contrast, the reactivity to acute stressful stimuli is dampened in late preg-nancy [51-53].
The long-term continuous increase of steroid hormones requires an un-coupling of the regular negative feedback systems. This is achieved by the feto-placental unit, which produces large amounts of oestrogens, progester-one, and CRH - among numerous other substances - to ultimately cause its own expulsion [54]. Instead of the negative feedback cortisol has on hypo-thalamic CRH production, cortisol has a positive feedback on placental CRH production, which in turn increases pituitary ACTH release [10, 55] (Figure 1). The maternal adrenals are enlarged during pregnancy, reflecting the in-creased production of cortisol [10]. Importantly, a substantial amount of cor-tisol is also produced by the fetal adrenal during the very last weeks of gesta-tion, probably as a signal for the maturation of fetal organs, as well as for the timing of parturition [56]. The feto-placental unit also synthesises the other HPA axis hormones ACTH and β-endorphin [57].
The altered HPA feedback is further evidenced by the fact that pharma-cological cortisol suppression by dexamethasone is reduced during late pregnancy and during the first weeks postpartum [58, 59].
Exogenous oestrogen decreases the sympathetic response to mental stress in healthy men [60] and perimenopausal women, and also decreases the HPA axis responsiveness in perimenopausal women [61]. Thus, also the increase in oestrogen may play a role in pregnancy-induced hyporesponsiveness.
While the baseline cortisol levels return to normal within a couple of days after parturition [62], the HPA axis hyporesponsiveness lingers in breastfeed-ing women [63]. It has been suggested that the lowered stress reactivity post-partum is important for maternal wellbeing [64] but an increased risk for immediate postpartum mood symptoms has also been found in relation to a higher degree of HPA axis dampening [65]. However, as in non-pregnant populations, the associations between HPA-measures and psychological variables during pregnancy have been contradictory. For example, higher,

16
lower, as well as equal serum CRH in depressed, as compared to non-depressed pregnant women have been reported [66-69].
Rats also display an attenuated HPA axis response to stress during preg-nancy and lactation, including decreased negative feedback by glucocorti-coids [70]. The progesterone metabolite allopregnanolone has been suggested to contribute to the decreased HPA axis reactivity through increased opioid inhibition in the brainstem, resulting in reduced excitatory noradrenergic signals to the PVN [71]. Via GABAergic inhibition, allopregnanolone may also suppress PVN CRH neurons [71]. After parturition, the pregnancy-induced opioid inhibition ends, but HPA responsiveness is maintained dur-ing lactation, perhaps due to central prolactin and oxytocin responses to suckling [72].
The maternal autonomic nervous system In parallel with HPA axis changes, the hormonal changes of pregnancy and childbirth also affect our other major stress system: the sympathetic nervous system (SNS). In the third trimester, the basal muscle sympathetic nerve activity is almost doubled [12], while the skin conductance reactivity to a mental stressor, as well as to relaxation, is dampened [51, 73]. Diastolic blood pressure responses to physiological and psychological stressors are also reduced in the second trimester compared to before pregnancy [53]. In contrast, heart rate reactivity to a psychosocial stressor seems to be compa-rable in third trimester pregnant and non-pregnant women [74]. Salivary alpha-amylase, an indicator of autonomic nervous system activation, is an-other example of a stress response that is attenuated during pregnancy; the salivary alpha-amylase response to psychosocial stress is lower in second trimester pregnant women than in non-pregnant women, and even lower in the third trimester [75]. Correspondingly, the parasympathetic activation is decreased during rest [51]. A study describing cardiovascular and epineph-rine and norepinephrine responses to a cold pressor test in late pregnancy did not find a correlation between the physiological responses and the subjective pain measurements [76, 77]. However, whether the altered pain experience in late pregnancy is influenced by the dampened sympathetic reactivity is not clear.
Basal plasma epinephrine and norepinephrine levels do not appear to be generally altered during normal pregnancy, but there are several reports of reduced reactivity to physiological stressors [78]. In the postpartum period, lactation seems to reduce basal plasma levels of norepinephrine, but not im-pede sympathomedullary reactivity, as breastfeeding women have the same norepinephrine response to exercise as mothers of bottle-fed infants [28].
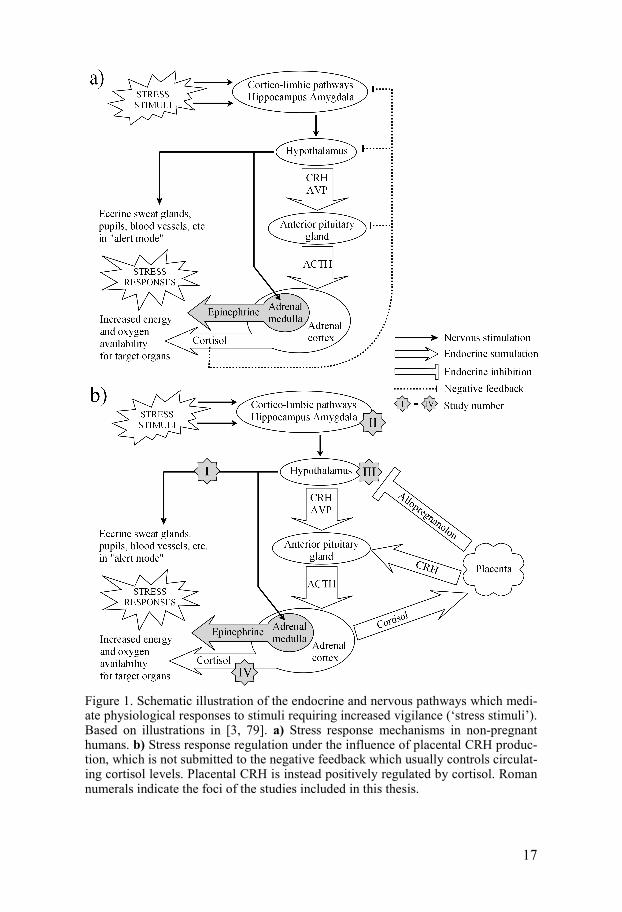
17
Figure 1. Schematic illustration of the endocrine and nervous pathways which medi-ate physiological responses to stimuli requiring increased vigilance (‘stress stimuli’). Based on illustrations in [3, 79]. a) Stress response mechanisms in non-pregnant humans. b) Stress response regulation under the influence of placental CRH produc-tion, which is not submitted to the negative feedback which usually controls circulat-ing cortisol levels. Placental CRH is instead positively regulated by cortisol. Roman numerals indicate the foci of the studies included in this thesis.

18
Spontaneous labour onset Although a human pregnancy is ideally 40 ± 2 weeks long, counting from the first day of the last menstrual period, there is a large variability [80]. Five to ten percent of all singleton births are pre-term, i.e. before 37 weeks of gestation, but only around half of the pre-term births are preceded by spon-taneous labour [81, 82]. Post-term pregnancy, when spontaneous labour has failed to initiate two weeks after the estimated date of delivery, precedes around 9% of all births in Sweden [83].
Several factors which contribute to labour onset have been identified. Uterine stretch, local prostaglandin release, fetal HPA axis activation, and oxytocin signalling are known to be involved, but the key which ultimately initiates labour at term is still unknown [57]. One theory is the “placental clock” hypothesis [84], which states that a sudden increase in bio-available placental corticotropic hormone (pCRH) in late pregnancy determines gesta-tional length by augmenting the prostaglandin and oxytocin signals which mediate uterine contractions. Maternal levels of pCRH have some predictive value regarding the risk of prematurity but when it comes to the more exact timing in term pregnancy pCRH is not of any practical use [85]. In non-primate mammals, a sudden drop in progesterone precedes parturition, while in humans and other primates the high progesterone levels are produced by the placenta and remain until after the delivery. The initiation of human la-bour has therefore been hypothesized to depend on functional progesterone withdrawal, through altered binding or expression of progesterone receptors [86]. The actions of progesterone may also be antagonized by the late preg-nancy surge in fetal cortisol production [87]. Oestrogens also seem to be important; lower oestradiol and, especially, higher oestriol/oestradiol ratio predicts successful response to post-term labour induction [88].
Different types of prostaglandins are involved both in inhibiting and me-diating uterine contractions and are also needed for cervical ripening [57]. Prostaglandin synthesis is in turn induced by oxytocin production within the uterus, which increases before parturition [57]. The contraction-induced stretch on the lower uterine segment during labour stimulates oxytocin re-lease, which enhances the contractions and additional oxytocin release in a positive feedback loop.
Involvement of the sympathetic nervous system in the regulation of la-bour is supported by the fact that lumbar sympathetic blockade reduces la-bour duration [89]. There is also a marked reduction in adrenergic innerva-tion of the uterus during pregnancy, a process which may have a role in the regulation of uterine contractions [90]. Whether the attenuation of sympa-thetic reactivity during pregnancy is involved in regulation of labour onset at term is not known.
Given the critical evolutionary significance, it is likely that labour initia-tion does not depend on one single factor, but that redundancy has evolved.

19
This is supported by the finding that oxytocin signalling is not requisite for successful parturition in mice [91, 92].
The reason behind spontaneous pre-term labour onset is often unknown. Especially in gestational week 33 and earlier, spontaneous labour is often associated with intrauterine infection [93]. The presence of bacteria induces maternal cytokines and fetal cortisol production and inhibits prostaglandin breakdown so that a premature prostaglandin peak occurs [93]. Other aeti-ologies include placental abrubtion, and abnormal uterine tension from poly-hydroamnion (excess amniotic fluid), or multiple foetuses [93].
Primiparity and genetic predisposition increases the risk for post-term pregnancy, but the aetiology is often unknown [88]. The most common rea-son for post-term pregnancy has been suggested to be faulty pregnancy dat-ing, and a high maternal body mass index (BMI) has also been identified as a risk factor [88, 94]. However, a high BMI is also a risk factor for incorrect pregnancy dating. In conclusion, much is known about the molecular mechanisms of labour regulation, but the levels of understanding are insuffi-cient for predicting labour onset.
Pain sensitivity during pregnancy and in the postpartum period Pain tolerance and/or pain thresholds are augmented in women during late pregnancy [76, 95, 96]. Experimental studies on human pregnancy-induced analgesia are few and differ largely in methodology; they also differ in which pain modality is studied (pressure, thermal etc.) and whether it is the pain tolerance, endurance or perceived pain intensity that is of interest. In general, there seems to be a lack of longitudinal studies and studies with large sample sizes. The regulation of opioid signalling in the spinal cord during human pregnancy is naturally difficult to assess. The established the-ory of pregnancy-induced analgesia is consequently based on rodent and pig models [97, 98]. Sows display pregnancy-induced analgesia a couple of days before parturition. The effect persists immediately postpartum and can then be blocked by an opioid antagonist [98]. The increase in pain tolerance in rats is also considered to depend mainly on alterations in opioid signalling; it has been shown that the dynorphin content increases selectively in the lum-bar spinal cord in rats in late pregnancy, and that methionine-enkephalin levels rise in the entire spinal cord [99]. Interestingly, some aspects of lower epidural and spinal analgesia requirements seem to be established in preg-nant women, although this phenomenon has been explained by mechanical reduction in the spinal and epidural spaces [100]. The late pregnancy rise in plasma β-endorphin has been suggested to contribute to relieve the stress and pain associated with the process of labour [101, 102]. The human placenta

20
produces small amounts of the opioid receptor agonists β-endorphin and dynorphin [57], but the increased systemic levels are due to HPA axis activ-ity. Baseline pregnancy levels of β-endorphin have previously been reported not to influence pain tolerance or pain intensity in response to a cold pressor test [77].
Mood, cognition and mental health during pregnancy The finding that physiological stress reactivity is progressively blunted dur-ing pregnancy is robust across experimental designs. On the other hand, the psychological reactivity to stress during the course of pregnancy remains elusive. The relatively few longitudinal subjects on the matter do not reveal any clinically significant changes in positive or negative affect [103], depres-sion [51], or anxiety [104] in healthy pregnant women. However, it has been argued that the psychological reactivity to highly stressful events, such as an earthquake, is lower during late pregnancy [105]. Less psychological distress in response to an experimental stressor in the third, than in the second tri-mester, has also been reported [106].
The popular concept of ‘pregnancy brain’ or ‘placenta brain’, i.e. that pregnant women become increasingly forgetful, unfocused, and emotionally labile, has little support in scientific literature [107]. Several smaller studies have reported deterioration in some aspects of memory function during pregnancy [4]. One of the rare longitudinal studies on cognitive function indicates only a statistically non-significant trend towards deterioration in one out of four tests of memory and cognition during pregnancy [107]. How-ever, there are some reports that pregnant women perceive their memory or other type of mental performance to be deteriorated during pregnancy, while they perform as well as non-pregnant controls [51, 108, 109]. An increase in subjective feelings of confusion and fatigue during pregnancy has been re-ported [51], but longitudinal studies about emotional processing are lacking. Since subjective ratings of emotionality during pregnancy and postpartum may be biased by cultural beliefs about the effects of pregnancy, it is valu-able to also use objective measures.
Rat mothers display increased aggression and a decrease in anxiety-like behaviours during both pregnancy and lactation [110]. It is also suggested that learning and memory are improved by pregnancy, lactation, and mere pup care [111]. Enhanced spatial memory function even appears to be main-tained a year after lactation [112].
Antenatal and postpartum depression and anxiety Although there are studies pointing to a high risk for onset or exacerbation of mood disorders in the puerperal period [113], the prevalence of depression

21
and anxiety may not be drastically higher at this time than during pregnancy or in the general female population [114-117]. Around 6-12% of all women are estimated to suffer from depression during pregnancy [113, 118] and the same figures are often used when estimating the frequency of postpartum depression [116, 119, 120]. However, by definition a postpartum depression starts after the delivery and few studies have pre-pregnancy or antenatal longitudinal data. One study suggests that 45% of women who are screen-positive for depression at six weeks after delivery, were also screen-positive prior to their pregnancy [121]. According to the strict definition, postpartum depression has its onset within four weeks after delivery [122], but the term is often used for depressive episodes which are ongoing at some point during the year after delivery. It is also debated whether antenatal depression and postpartum depression differ in symptomatology from major depressive dis-order in any other aspect than timing [123].
Perinatal anxiety disorders are less studied than perinatal depression, de-spite similar prevalence and high comorbidity. A meta-analysis indicated possible increases in new onset of panic disorder and generalised anxiety disorder during the postpartum period, compared to during pregnancy or in the general population, but largely stable courses of pre-existing anxiety disorders [124].
Regardless of the absolute prevalence and symptoms of depression and anxiety disorders during pregnancy and postpartum period, they present unique implications for pharmacological treatment and bonding with the infant, thus affecting the patient differently than depressive episodes during other times in life.
Previous depression is a major risk factor for postpartum depression [116], and perhaps an even more important risk factor for antenatal depression [123]. Socioeconomic factors such as lack of partner support, financial diffi-culties, or other life events are also very strong predictors for perinatal de-pression and anxiety [116, 125]. Still, during late pregnancy and the postpar-tum period, the biological systems known to be part of the depressive symp-tomatology are under a great strain [42-44, 65]; and a vulnerability to depres-sion could theoretically be more evident during this period.
Stress response regulation in depression and anxiety The heterogeneous aetiology, symptomatology, and treatment response pat-terns in patients with depression or anxiety are likely to prevent neuroscience from finding a single mechanism, or a single valid biomarker, for either de-pression or anxiety. However, alteration in HPA axis function is a frequent finding in major depression [126-128]. The general understanding is that ma-jor depression patients have a hyperactive HPA axis, and consequently a

22
higher cortisol production. A 2012 meta-analysis estimates the clinical sig-nificance to be modest, based on low overall effect sizes [129].
In generalised anxiety disorder and panic disorder, the cortisol output is thought to be exhausted by the chronic stress, and consequently be main-tained at a low level [130], but possibly with higher response to awakening [131].
Like the other HPA axis products, β-endorphin has also been investigated in relation to mood, although the results are inconclusive. In 2010, Yim and colleagues described a higher β-endorphin increase profile in women who were healthy during pregnancy, but experienced depression in the postpar-tum period [132].
The autonomic nervous system is also affected by mental illness. The sympathetic nerve activity is higher in major depression [133], and heart rate variability is lower [134]. There are several conflicting reports of autonomic function alterations in anxiety disorder patients, including increased and decreased basal levels and reactivity, and decreased habituation [135].
It is widely recognised that vulnerability to depression is partly heritable, and genetic variations in monoamine, brain derived neurotrophic factor, and HPA related genes, which increase susceptibility, have been identified [136-139]. There is also evidence that some biological markers of depression, including neurosteroid profile and cortisol reactivity, seem to linger in remit-ted patients [128, 140, 141]. Altered HPA axis reactivity may also precede the onset of a first depressive episode or a relapse [127]. These observations underline the importance of investigating the potential consequences of pre-vious depression on endocrinological function during pregnancy.
The skin conductance response In study I, skin conductance response was used as a measure of sympathetic reactivity, during a cold pressor test (see below). The first studies coupling changes in skin conduction to experimental stimuli were performed in 1888 by the French neurologist Charles Féré [142]. Since then, the technique has been used in a wide range of psychological and physiological experiments as well as in “lie detector” tests [51, 143, 144]. The skin conductance level de-pends on the activity of the eccrine sweat glands, which reflects the output of the sympathetic nervous system. The skin conductance response is not influ-enced by surrounding normal indoor temperature range or by local blood flow [142]. Eccrine sweat glands are distributed over most parts of the hu-man skin but are most abundant on palmar and plantar skin, where skin con-ductance is predominantly measured. Studies on, for example, attention, emotion, and pain have therefore employed the skin conductance response as a simple and non-invasive measure of sympathetic activity [142, 145].

23
During rest, skin conductance as a function of time is typically a negative slope with one to three non-specific peaks per minute. Within a few seconds of the perception of a sensory stimulus that calls for attention, a signal from the prefrontal cortex, via sympathetic innervation, causes the sweat glands to fill up. This will transiently render the skin more conductive, which is re-ferred to as the skin conductance response. A more intense or more interest-ing stimulus will be followed by a larger response – larger peaks, more fre-quent peaks and slower recovery [142]. The classical use of skin conductance in psychological experiments is to look at the latency, amplitude, and rise-time of individual peaks following transient exposure to discrete stimuli, such as pictures or words. Alternatively, skin conductance level or peak fre-quency can be used during continuous stimuli exposure, such as a stressful task [142]. Peak frequency has been used as primary outcome in studies of clinical pain and is related to subjective pain ratings [145-147]. Other ways to quantify skin conductance activity are skin conductance area under the curve, or rate of change. The rate of skin conductance change denotes the slope of a given time portion of the skin conductance curve, while the area under the curve is a function of the peak frequency and amplitude over time [148].
The cold pressor test The cold pressor test is frequently used as an experimental stressor and/or as a pain threshold assessment [76, 149-151]. The test typically implies that the subject is asked to submerge a hand or foot into cold water or crushed ice until the cold-induced pain becomes intolerable, while the physiological response of interest is concurrently measured. Alternatively, ice can be ap-plied to the subjects’ forehead [152]. Several researchers have applied the cold pressor test in pregnant and postpartum women. The findings indicate that pregnancy increases the pain tolerance [76] and attenuates the cortisol response to a cold pressor test [151].
The acoustic startle response and its modulation In study II, we attempted to assess emotional regulation through affective modulation of the acoustic startle response. The acoustic startle response is a defensive reflex response to a sudden aversive stimulus and is widely used in psychopharmacological animal research [153]. The human startle response can be experimentally assessed by measuring the contraction of musculus orbicularis oculi, the eye blink, in response to auditory or tactile stimuli. In rodents, the startle reflex is measured as whole body twitch. The startle re-flex is a simple brainstem reflex but it is modulated by impulses from several

24
other parts of the brain, such as the amygdalae. The amygdalae are nuclei within the limbic system which are known to be activated by emotional stimuli, especially fear and emotional faces [153, 154]. The baseline startle response is significantly increased in post-traumatic stress disorder [155, 156], generalised anxiety disorder [157], specific phobias [158, 159], and ob-sessive compulsive disorder [160, 161].
Anxiety, fear, and negative affect (threat of shock, aversive images) po-tentiate the startle magnitude, through activation of the amygdalae. Con-versely, startle can be attenuated by positive affect (reward, pleasant images) [157, 162]. In addition, the startle reflex is potentiated during the anticipation of highly emotional images compared to neutral images [163, 164]. Several anxiety disorders are associated with an altered affect-modulated startle re-sponse [155, 157, 165]. Depressed patients respond with lower modulation by affective images, as well as by anticipation stimuli [166, 167]. Interestingly, short term SSRI treatment also results in reduced affective startle modulation in healthy subjects [168]. Furthermore, tryptophan depletion has been re-ported to decrease the startle magnitude [169]. There is also a bi-phasic effect of cortisol on baseline startle in humans, but cortisol does not appear to alter the emotional modulation [170].
Some of the neurotransmitter systems which are believed to affect the startle response, such as GABAergic, opioid, and noradrenergic signalling [171], may be altered during pregnancy and the postpartum [71]. However, baseline startle response has previously been shown to be comparable in women in the third trimester of pregnancy and postpartum women [172]; but no study examining affective modulation of the startle response in pregnant or postpartum women has been published before.
Allopregnanolone – in stress, mood, and pregnancy Study III focuses on correlates of pregnancy levels of allopregnanolone, (3α-hydroxy-5α-pregnan-20-one). Allopregnanolone is a neuroactive steroid, i.e. a steroid with central nervous system activity which is also synthesised within the brain [173]. Allopregnanolone exerts its function via allosteric binding to GABAA-receptors which enhances the GABAergic signalling [174]. As GABA is the major inhibitory transmitter in the central nervous system, allopregnanolone has sedative, anxiolytic, and anti-convulsant prop-erties [173]. A functionally relevant amount of allopregnanolone is synthesised in the brain [175], but the main source of brain and serum allopregnanolone in non-pregnant women is the corpus luteum and adrenal cortex [176, 177]. The allopregnanolone synthesis in the adrenal cortex is stimulated in parallel with cortisol secretion, and thereby provides a negative regulation, or bal-ance, to the HPA axis stress response.

25
Patients with major depression have been found to have low levels of al-lopregnanolone [178-180]. On the contrary, currently remitted women with major depression or bipolar disorder have higher serum allopregnanolone concentrations, regardless of treatment and co-morbidity [181]. Another study showed no difference in allopregnanolone between post-menopausal women with or without menopause-associated mood alterations [182]
The functional role of allopregnanolone in mood disorder pathophysiol-ogy is corroborated by the finding that successful treatment with selective serotonin reuptake inhibitors (SSRI) increases plasma and cerebrospinal fluid allopregnanolone [179, 180], although there are conflicting results [178].
There are also reports of low serum allopregnanolone in mild mood dis-turbance, such as the postpartum blues, which is a transient episode of de-pressed mood during the first week after childbirth [183]. In postmenopausal women, allopregnanolone has been shown to be related to negative mood in an inverted u-shaped curve, where levels of 1.5-2 nmol/l are associated with the highest magnitude of negative mood symptoms while concentrations below and above are associated with lower levels of negative mood [184]. During pregnancy, maternal serum concentrations of allopregnanolone rise to more than ten times the maximum menstrual cycle levels [185, 186], due to feto-placental synthesis. The fatigue experienced by many women during the first trimester has been proposed to be an effect of allopregnanolone [187]. An allopregnanolone serum concentration equivalent to that in the third tri-mester is sedative if intravenously administered to non-pregnant women [188], suggesting that tolerance develops during pregnancy [187].
A major issue when studying neuroactive substances in humans is access-ing central nervous system levels of the hormone of interest. Allopreg-nanolone crosses the blood-brain barrier, and although cerebrospinal fluid concentrations are around ten times lower, they are in strong correlation with serum levels of the steroid [189]. It can therefore be assumed that the varia-tions in serum allopregnanolone concentration during pregnancy are re-flected in the brain. More importantly, allopregnanolone serum levels were correlated with brain tissue levels in a human post mortem study [190].
In rats, exogenous allopregnanolone suppresses stress responses through up-regulation of opioid signalling in the brainstem, mimicking the effects of pregnancy [71]. Rodent studies further suggest that chronic stress, and the consequent HPA axis overload, can cause a depletion of serum and brain levels of allopregnanolone. Without the balancing effect of an adequate allo-pregnanolone response, the HPA axis function is thought to deteriorate fur-ther. In a series of experiments, Evans and colleagues have confirmed that exogenous allopregnanolone can protect rats against development of the behavioural deficits induced by the social isolation model of depression. Moreover, the continuous allopregnanolone supplementation also prevented the decline in hippocampal cell proliferation seen after social isolation [191].

26
Only one small study has investigated pregnancy allopregnanolone levels in clinically depressed pregnant women, and found no difference compared to controls [192]. However, the relationship between serum allopregnanolone and depressive symptoms outside the clinical range has not been studied.
The cortisol awakening response In study IV, the cortisol awakening response was used as a test of stress re-activity. The physiological secretion of cortisol is relatively high during morning hours, decreases during the day, and begins to slowly rise again a couple of hours after midnight [193]. Although ACTH and cortisol start to rise in the hours before waking, a marked increase in free cortisol (50 – 160%) is seen between the time of awakening and 30 – 45 minutes thereafter [194]. This increase, the cortisol awakening response was first described in 1997 [195]. Since the ACTH response to awakening is not as rapid, the corti-sol response is thought to depend on a sympathetically mediated increase in the adrenal splanchnic cortex sensitivity to ACTH [196]. The function of the awakening response is not established, although it has been suggested that it has a role in the transition from sleep inertia to alertness [196]. Despite the uncertain mechanism behind the cortisol awakening response, it has spurred a lot of research interest due to reports of its associations with mental health, as well as cardiovascular health, bone density, and perceived stress [194]. Interestingly, the cortisol awakening response has been reported to be in-creased not only in currently depressed, but also in formerly depressed indi-viduals, rendering it a potential vulnerability marker for, or scar after, de-pression [128]. The cortisol awakening response is also slightly higher in women than in men [197].
Although the absolute saliva cortisol concentrations are increased in late pregnancy, a distinct cortisol awakening response is still present in gesta-tional weeks 32-36 [198]. However, in analogy with the reduced stress reac-tivity to other stimuli [51, 151], Buss and colleagues have found a decrease in cortisol awakening response between the 17th and 31st week of pregnancy and also found an association between this dampening and the timing of delivery [199]. The cortisol awakening response has been reported to not be affected by antenatal symptoms of depression, but these studies did not take previous depressive episodes into account [200, 201].

27
Aims
I: to assess pain threshold and sympathetic nervous system response to cold pain in relation to onset of labour in healthy pregnant women II: to compare startle response modulation, during anticipation and during viewing of affective images, across late pregnancy and puerperium of healthy women III: to determine if the serum level of the neurosteroid allopregnanolone is correlated to self-reported symptoms of depression and anxiety in late preg-nancy IV: to compare the cortisol awakening response in late pregnancy in women with ongoing depression with currently healthy women, with and without a history of depression

28
Materials and Methods
Paper I-III Ninety-nine pregnant women were recruited via advertisement and primary care midwives, and visited the laboratory at the Department of Women’s and Children's Health in gestational week 36-40. The inclusion criteria were age >18 years, and an uncomplicated singleton pregnancy. The sequence of pro-cedures was: informed consent, venous blood sampling, interview about obstetric history and medication, psychiatric interview, self-report question-naires, startle modulation test, and cold pressor test. For study II, 31 women returned four to six weeks after delivery for a second visit, which included blood sampling, questionnaires, and a second startle modulation assessment.
Psychiatric evaluation by self-report questionnaires and structured interview There are several tools available to screen for psychiatric symptoms or ob-tain psychiatric diagnoses for research purposes. Two screening instruments to assess depressive symptoms which are commonly used in clinical practice in Sweden are the Montgomery-Åsberg Depression Rating Scale – Self-rated version (MADRS-S) and the Edinburgh Postnatal Depression Scale (EPDS). MADRS-S concerns the past three days and consists of nine items (e.g. ap-petite, pessimism, zest for life) rated from 0 to 6 [202]. MADRS-S is primar-ily designed to detect change and is therefore often used in clinical trials of antidepressants [203, 204]. No validated MADRS-S cut-off score is published for the detection of minor or major depression, but a score of 13 (90th per-centile) or more was considered as elevated depressive symptoms in study III.
The EPDS was developed for use in the postpartum period, and therefore avoids questions on somatic symptoms which can be confounded with nor-mal postpartum symptoms [205]. The EPDS contains ten items which can be rated on a scale from 0 to 3, based on the past seven days. The Swedish ver-sion of the EPDS has been validated during pregnancy [206] and during two to three months after delivery [207] with the suggested cut-off scores of ≥13 and ≥12 respectively to detect mild depression with positive predictive val-ues of 54% and 59%.

29
A questionnaire commonly used to evaluate anxiety is the Spielberger State-Trait Anxiety Inventory (STAI) [208]. The state and trait versions, STAI-S and STAI-T, assess current anxiety level and anxiety prone person-ality respectively. For both the STAI-S and -T, a cut-off score of >40 gives around 80% sensitivity and specificity for detecting anxiety disorders during pregnancy [114].
Most self-report questionnaires do not cover all criteria required for a psychiatric diagnosis, for example differential diagnoses and duration of symptoms. Therefore, a structured interview has higher specificity for de-tecting depression or anxiety disorders. The Mini International Neuropsy-chiatric Interview (MINI) is a structured interview based on DSM-IV criteria [209].
Paper I Ninety-three women were included in the analysis of skin conductance activ-ity during a cold pressor test in gestational week 36-40. The skin conduc-tance measurement from one woman was lost due to a technical error. Fur-ther, exclusion from analyses was warranted by current treatment with SSRI (n=2), or ongoing major depressive disorder or primary anxiety disorders detected by MINI (n=3). Furthermore, no included women reported use of analgesics within 24 hours of the test session. The cold pressor test consisted of one minute of holding the dominant hand in 35 ± 1 ºC water (baseline), followed by maximum one minute in 0 ± 1 ºC water (cold pressor).
Skin conductance was assessed via electrodes on the non-dominant hand with a Med-Storm skin conductance algesimeter (Med-Storm Innovation, Oslo, Norway). The skin conductance measurements were: area under the curve (AUC), peaks per second, and rate of change. The measurements were extracted for the first 30 seconds of baseline and cold pressor respectively (Figure 2). Pain threshold, cold endurance, and pain ratings were also meas-ured. Information on the number of days to delivery, and whether the onset was spontaneous, was obtained from medical records.
In addition to the data published in paper I, plasma levels of β-endorphin were also analysed. This was done using a radioimmunoassay using the EU-RIA-beta-endorphin kit (Euro-Diagnostica, Sweden) as described in [101].
Cold pressor test outcomes, including skin conductance measurements, were compared between women with less than two weeks and women with 14 days or more left to spontaneous delivery using Mann-Whitney U-tests. Linear relationships between days left to delivery and the skin conductance reactivity measures were also explored using linear regressions. SPSS Statis-tics for Windows, version 17 (SPSS Inc, Chicago), was used for statistical analyses in paper I and II.
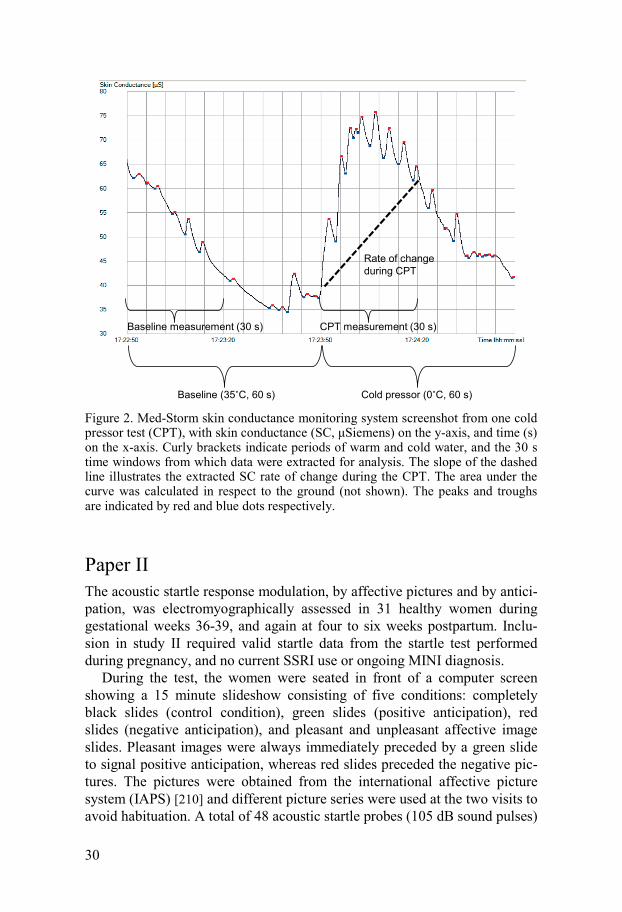
30
Figure 2. Med-Storm skin conductance monitoring system screenshot from one cold pressor test (CPT), with skin conductance (SC, μSiemens) on the y-axis, and time (s) on the x-axis. Curly brackets indicate periods of warm and cold water, and the 30 s time windows from which data were extracted for analysis. The slope of the dashed line illustrates the extracted SC rate of change during the CPT. The area under the curve was calculated in respect to the ground (not shown). The peaks and troughs are indicated by red and blue dots respectively.
Paper II The acoustic startle response modulation, by affective pictures and by antici-pation, was electromyographically assessed in 31 healthy women during gestational weeks 36-39, and again at four to six weeks postpartum. Inclu-sion in study II required valid startle data from the startle test performed during pregnancy, and no current SSRI use or ongoing MINI diagnosis.
During the test, the women were seated in front of a computer screen showing a 15 minute slideshow consisting of five conditions: completely black slides (control condition), green slides (positive anticipation), red slides (negative anticipation), and pleasant and unpleasant affective image slides. Pleasant images were always immediately preceded by a green slide to signal positive anticipation, whereas red slides preceded the negative pic-tures. The pictures were obtained from the international affective picture system (IAPS) [210] and different picture series were used at the two visits to avoid habituation. A total of 48 acoustic startle probes (105 dB sound pulses)
Baseline (35˚C, 60 s) Cold pressor (0˚C, 60 s)
Baseline measurement (30 s) CPT measurement (30 s)
Rate of changeduring CPT

31
were delivered binaurally by headphones. Twenty probes were programmed to occur during the control condition, and seven probes during positive an-ticipation stimuli, negative anticipation stimuli, pleasant image stimuli, and unpleasant image stimuli respectively. The acoustic probes and the electro-myographic recording of the eye blink startle responses were controlled by a commercial startle system (SR-HLAB, San Diego Instruments, San Diego, CA, USA). All women provided affective ratings of the pictures. Valence was rated on 10-cm visual analogue scales ranging from positive (0.0 cm) to negative (10.0 cm). Arousal ratings ranged between calm (0.0 cm) and aroused (10.0 cm). Serum levels of oestradiol and progesterone were ob-tained at both sessions.
Z-scores of the respective startle magnitudes were compared between sessions, within subjects in a repeated measures ANOVA. Differences in picture ratings, depression scores, and anxiety scores between pregnancy and postpartum states were compared with Mann–Whitney U-tests.
Paper III
Ninety-six women were included in Study III, two women were excluded due to current SSRI medication, and one did not provide a blood sample. Serum allopregnanolone was purified with celite chromatography and ana-lysed by radioimmunoassay at the Umeå Neurosteroid Research Center. Depression scores and anxiety levels were self-reported with the MADRS-S, and the STAI-S and STAI-T respectively. The cut-off for presence of de-pressive symptoms was set at a MADRS-S score of 13 or more, while the anxiety cut-off was 40 points for STAI-S and STAI-T.
Mann-Whitney U-tests were used for comparisons between groups, and linear regression was applied to enable control for potential confounders of the association between allopregnanolone self-rated depression scores. SPSS Statistics for Windows, version 20 (IBM Corp. Armonk, NY), was used for statistical analyses in papers III and IV.
Paper IV Women were recruited via e-mail, and participated in a set of neuropsy-chological tests at the Department of Women’s and Children's Health in gestational week 36-40, as a sub-study to the pregnancy cohort ‘Biology, Affect, Stress, Imaging, and Cognition in pregnancy and the puerperium’ (BASIC). All pregnant women in Uppsala County were invited to participate in BASIC at the time of their routine ultrasound screening in gestational week 16-18. Following informed consent, women were followed by web-based questionnaires at gestational week 17, gestational week 32, six weeks

32
postpartum, and six months postpartum. For the current study, 216 BASIC participants were recruited and attended a test session.
At the visit, the women received a cortisol awakening response sampling kit with instructions and a report sheet. Twin pregnancy, use of systemic steroid medication, and reported delay between awakening and the first sam-ple over 5 minutes warranted exclusion from the analysis. In all, the final analysis included salivary cortisol from 134 women, from time of morning awakening and 15, 30, and 45 minutes post-awakening. Ongoing depressive episodes were screened with the MINI-interview [209]. Previous depressive episodes were assessed by the question “Have you ever experienced a period of two weeks or more when you have felt depressed or uninterested in most things?” (analogous to MINI question A4a). If the woman answered ‘yes’, the interviewer asked her to consider the time-period when she felt depressed or uninterested (or ‘the worst one’ if she had experienced several such time-periods) and asked the questions in the depression module of the MINI in-terview. EPDS scores were obtained at the visit and from the web question-naires in gestational weeks 17 and 32.
The cortisol awakening response was compared between three groups: women who experienced depression during the current pregnancy, currently depression-free women with a previous experience of major or minor de-pression prior to the current pregnancy, and women who had never suffered from a depressive episode (controls). In the depressed group, we included women with a current depressive disorder (major or minor depression or dysthymia), or a history of major or minor depressive episode, where EPDS scores ≥ 13 at any time-point during pregnancy indicated that the prior (or last) episode had been ongoing during the present pregnancy. Women with a prior depressive episode had a history of at least one major or minor depres-sive episode according to the MINI interview, but not during the present pregnancy, as indicated by EPDS scores < 13 at all time-points (gestational week 17, 32 and at the visit). Control subjects were defined as having no current or prior depressive disorder, no current anxiety disorder (panic disor-der, generalised anxiety disorder, or social phobia) as assessed by MINI.
Salivary cortisol concentrations were analysed by electrochemilumines-cence immunoassay at the Department of Clinical Chemistry at Uppsala University Hospital. A linear mixed model with time-point as repeated measure, and group and time-point as fixed factors was applied to assess group differences in magnitude and shape of the cortisol curves.

33
Summary of results
Paper I The mean number of skin conductance fluctuations (peaks) per second, the rate of change, and the area under the curve all increased significantly during the cold pressor test compared to the baseline measurement (all p-values < 0.001, Wilcoxon signed-rank tests), indicating that the cold pressor activated the sympathetic nervous system. Furthermore, the level of sympathetic acti-vation depended on perceived pain; positive correlations were found be-tween self-reported pain intensity (VAS-score) and skin conductance area under curve (r = 0.241, p = 0.026), and with rate of skin conductance change (r = 0.354, p = 0.001). VAS-score was also weakly correlated to the number of skin conductance fluctuations per second (r= 0.209, p= 0.055). In addi-tion, the skin conductance rate of change was significantly higher in women with cold endurance under 60 seconds.
Skin conductance activity during the cold pressor test decreased signifi-cantly with fewer days left to spontaneous parturition in simple linear regres-sions (area under the curve: r = -0.313, p = 0.007 and rate of change: r = -0.319, p = 0.006). The number of days left to delivery was normally distrib-uted, but since no skin conductance variables followed normal distribution the regressions’ residual distributions were checked for normality, and found to meet this criterion (Shapiro-Wilk W > 0.95).
In line with the negative correlation, women with fewer than 14 days left to parturition (median) had significantly lower skin conductance response than women with two to four weeks left. The difference in skin conductance area under curve (AUC) approached significance (Table 1).
34
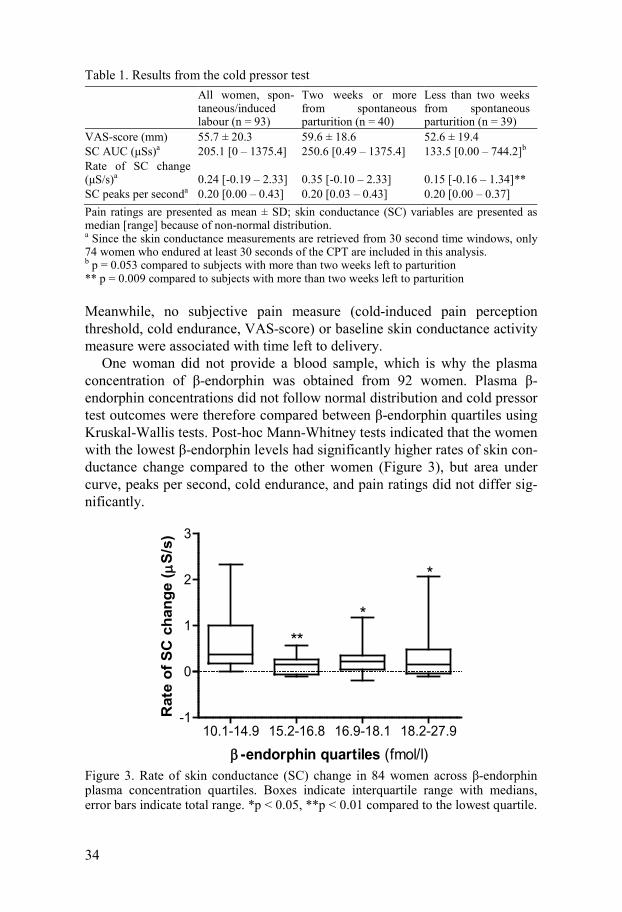
Table 1. Results from the cold pressor test
All women, spon-taneous/induced labour (n = 93)
Two weeks or more from spontaneous parturition (n = 40)
Less than two weeks from spontaneous parturition (n = 39)
VAS-score (mm) 55.7 ± 20.3 59.6 ± 18.6 52.6 ± 19.4 SC AUC (μSs)a 205.1 [0 – 1375.4] 250.6 [0.49 – 1375.4] 133.5 [0.00 – 744.2]b Rate of SC change (μS/s)a 0.24 [-0.19 – 2.33] 0.35 [-0.10 – 2.33] 0.15 [-0.16 – 1.34]** SC peaks per seconda 0.20 [0.00 – 0.43] 0.20 [0.03 – 0.43] 0.20 [0.00 – 0.37] Pain ratings are presented as mean ± SD; skin conductance (SC) variables are presented as median [range] because of non-normal distribution. a Since the skin conductance measurements are retrieved from 30 second time windows, only 74 women who endured at least 30 seconds of the CPT are included in this analysis. b p = 0.053 compared to subjects with more than two weeks left to parturition ** p = 0.009 compared to subjects with more than two weeks left to parturition
Meanwhile, no subjective pain measure (cold-induced pain perception threshold, cold endurance, VAS-score) or baseline skin conductance activity measure were associated with time left to delivery.
One woman did not provide a blood sample, which is why the plasma concentration of β-endorphin was obtained from 92 women. Plasma β-endorphin concentrations did not follow normal distribution and cold pressor test outcomes were therefore compared between β-endorphin quartiles using Kruskal-Wallis tests. Post-hoc Mann-Whitney tests indicated that the women with the lowest β-endorphin levels had significantly higher rates of skin con-ductance change compared to the other women (Figure 3), but area under curve, peaks per second, cold endurance, and pain ratings did not differ sig-nificantly.
Figure 3. Rate of skin conductance (SC) change in 84 women across β-endorphin plasma concentration quartiles. Boxes indicate interquartile range with medians, error bars indicate total range. *p < 0.05, **p < 0.01 compared to the lowest quartile.
10.1-14.9 15.2-16.8 16.9-18.1 18.2-27.9-1
0
1
2
3
**
*
*
β -endorphin quartiles (fmol/l)
Rat
e of
SC
cha
nge
( µS/
s)
35
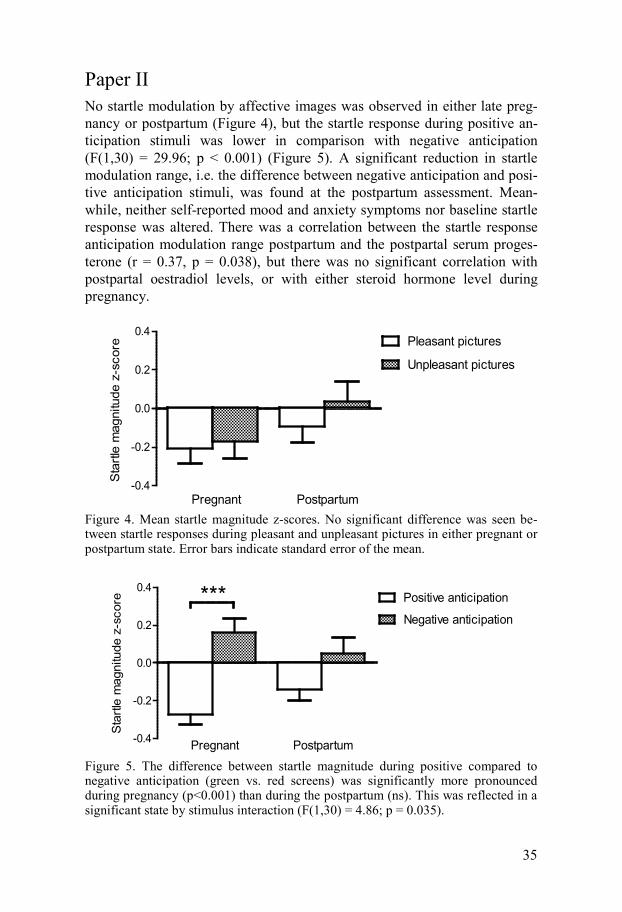
Paper II No startle modulation by affective images was observed in either late preg-nancy or postpartum (Figure 4), but the startle response during positive an-ticipation stimuli was lower in comparison with negative anticipation (F(1,30) = 29.96; p < 0.001) (Figure 5). A significant reduction in startle modulation range, i.e. the difference between negative anticipation and posi-tive anticipation stimuli, was found at the postpartum assessment. Mean-while, neither self-reported mood and anxiety symptoms nor baseline startle response was altered. There was a correlation between the startle response anticipation modulation range postpartum and the postpartal serum proges-terone (r = 0.37, p = 0.038), but there was no significant correlation with postpartal oestradiol levels, or with either steroid hormone level during pregnancy.
Figure 4. Mean startle magnitude z-scores. No significant difference was seen be-tween startle responses during pleasant and unpleasant pictures in either pregnant or postpartum state. Error bars indicate standard error of the mean.
Figure 5. The difference between startle magnitude during positive compared to negative anticipation (green vs. red screens) was significantly more pronounced during pregnancy (p<0.001) than during the postpartum (ns). This was reflected in a significant state by stimulus interaction (F(1,30) = 4.86; p = 0.035).
-0.4
-0.2
0.0
0.2
0.4
Sta
rtle
mag
nitu
de z
-sco
re
Pregnant Postpartum
Pleasant pictures
Unpleasant pictures
-0.4
-0.2
0.0
0.2
0.4
Pregnant Postpartum
Positive anticipation
Negative anticipation***
Sta
rtle
mag
nitu
de z
-sco
re

36
The women rated the unpleasant images slightly more negative and the pleasant images slightly more positive at the postpartum visit. They also indicated increased arousal for unpleasant, but not pleasant, images at their postpartum visit (Table 2).
Table 2. VAS-ratings of valence and arousal of the affective pictures (mean ± SD)
Late pregnancy Postpartum
Valence, pleasant images 1.9 ± 0.9 1.6 ± 0.7 * Valence, unpleasant images 7.8 ± 1.1 8.1 ± 1.0 * Arousal, pleasant images 2.9 ± 1.8 2.6 ± 1.8 Arousal, unpleasant images 5.7 ± 1.9 6.4 ± 1.6 ** * p < 0.05; ** p < 0.01, compared with late pregnancy
37
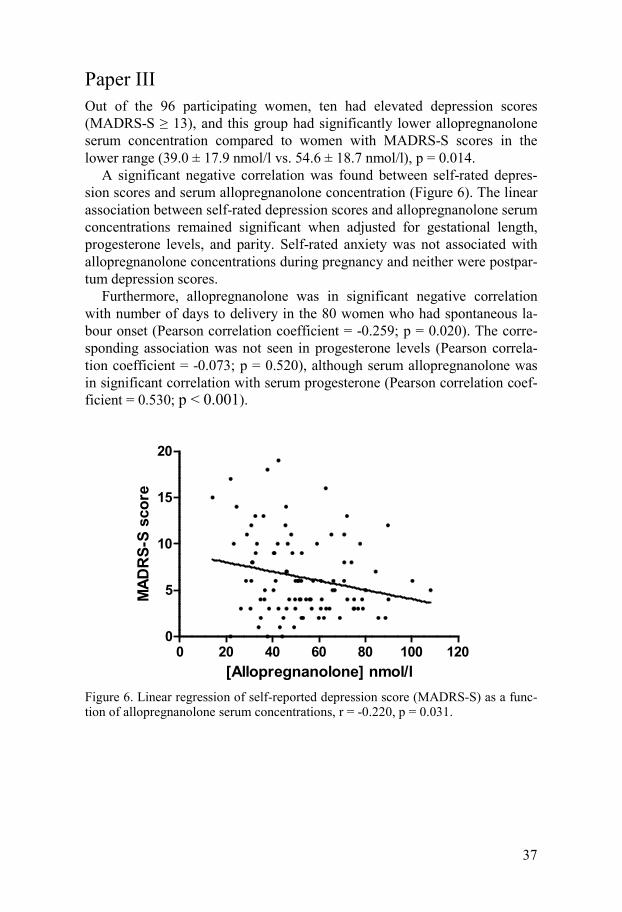
Paper III Out of the 96 participating women, ten had elevated depression scores (MADRS-S ≥ 13), and this group had significantly lower allopregnanolone serum concentration compared to women with MADRS-S scores in the lower range (39.0 ± 17.9 nmol/l vs. 54.6 ± 18.7 nmol/l), p = 0.014.
A significant negative correlation was found between self-rated depres-sion scores and serum allopregnanolone concentration (Figure 6). The linear association between self-rated depression scores and allopregnanolone serum concentrations remained significant when adjusted for gestational length, progesterone levels, and parity. Self-rated anxiety was not associated with allopregnanolone concentrations during pregnancy and neither were postpar-tum depression scores.
Furthermore, allopregnanolone was in significant negative correlation with number of days to delivery in the 80 women who had spontaneous la-bour onset (Pearson correlation coefficient = -0.259; p = 0.020). The corre-sponding association was not seen in progesterone levels (Pearson correla-tion coefficient = -0.073; p = 0.520), although serum allopregnanolone was in significant correlation with serum progesterone (Pearson correlation coef-ficient = 0.530; p < 0.001).
Figure 6. Linear regression of self-reported depression score (MADRS-S) as a func-tion of allopregnanolone serum concentrations, r = -0.220, p = 0.031.
0 20 40 60 80 100 1200
5
10
15
20
[Allopregnanolone] nmol/l
MAD
RS-
S sc
ore
38
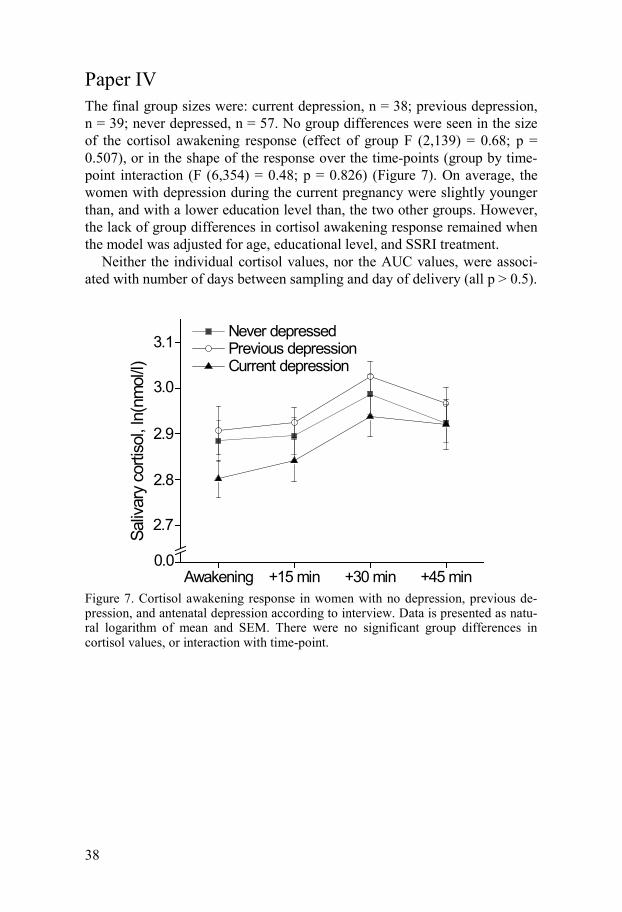
Paper IV The final group sizes were: current depression, n = 38; previous depression, n = 39; never depressed, n = 57. No group differences were seen in the size of the cortisol awakening response (effect of group F (2,139) = 0.68; p = 0.507), or in the shape of the response over the time-points (group by time-point interaction (F (6,354) = 0.48; p = 0.826) (Figure 7). On average, the women with depression during the current pregnancy were slightly younger than, and with a lower education level than, the two other groups. However, the lack of group differences in cortisol awakening response remained when the model was adjusted for age, educational level, and SSRI treatment.
Neither the individual cortisol values, nor the AUC values, were associ-ated with number of days between sampling and day of delivery (all p > 0.5).
Figure 7. Cortisol awakening response in women with no depression, previous de-pression, and antenatal depression according to interview. Data is presented as natu-ral logarithm of mean and SEM. There were no significant group differences in cortisol values, or interaction with time-point.
Awakening +15 min +30 min +45 min0.0
2.7
2.8
2.9
3.0
Saliv
ary
corti
sol,
ln(n
mol
/l)
Never depressed Previous depression Current depression
3.1

39
Discussion
Methodological considerations Participant selection bias Participants in study I-III were recruited via advertisement and thus actively volunteered to participate. This imposes a self-selection bias which resulted in a sample with lower parity and higher education in comparison with the general population of pregnant women. Although the BASIC study, which was the recruitment basis for study IV, has a population-based recruitment strategy, the participants in BASIC are also more likely to be primiparous, and to have a university education (Sundström-Poromaa, unpublished obser-vations). Moreover, few of the participating women are born outside of Sweden. Although this has implications for the generalisability of the results, a high level of homogeneity is also a prerequisite for explorative studies with relatively small sample sizes. Both labour onset and stress system regulation during pregnancy are known to be affected by socioeconomic status, migra-tion, and minority status [93, 211]. To account for these factors, substantially larger study populations are required. However, strategies to recruit study populations which truly represent the target population are essential for ex-panding the knowledge on stress system regulation during pregnancy.
Cross-sectional design In studies I, III, and IV, we used cross-sectional designs, which only allow for comparisons between individuals. Since we have studied factors which change rapidly during pregnancy and puerperium, longitudinal designs would have increased the possibility to draw firm conclusions from the re-sults. However, also in study II, where we had longitudinal data, the inter-pretation would have been facilitated if we had also performed tests pre-pregnancy and/or after the return of regular ovulation. This would require a lot of time and large starting sample sizes, as attrition due to pregnancy complications and participant drop-out could be substantial. Repeated meas-urements also introduce habituation.

40
Subjective and objective assessments of stress, pain, and affect A recent review found a general low correspondence between measures of psychological experience and objective measurements of stress (cortisol and cardiovascular reactivity) [212]. Regarding cortisol, one reason for lack of correlation may be that the baseline ultradian pulsatility of cortisol secretion renders single cortisol analyses rather uninformative [213].
Compared to the determination of steroid hormones in plasma and saliva, the autonomous nervous system parameters, such as skin conductance, have the advantages of immediacy and high temporal resolution. The psycho-physiologic relevance of the skin conductance response is further indicated by the observation that both higher VAS-scores and lower cold endurance were associated with increased skin conductance reactivity in study I. Skin conductance activity have previously been shown to be associated with rat-ings of postoperative pain [145], and can thus be thought of as an objective measure of pain. However, any salient stimuli or emotion, positive or nega-tive, will induce sympathetic activation. Thus, the method is better suited for the study of experimental pain.
As pain stimulus, we employed the cold pressor test. It is a relatively mild laboratory stressor which has been proven to be well tolerated by pregnant women [76]. However, we do not know how the reactivity to cold stress re-lates to the reactivity towards other types of stressors, such as mental arith-metic, public speech, or physical exercise. It is possible that the responses to different types of tests have different relevance for maternal well-being. However, also study designs involving repeated testing with a variety of stressors during pregnancy could be considered unethical.
Among psychophysiological reactivity measures, the startle response is unique in the sense that it can be reduced by appetitive stimuli, and increased by fear and anxiety. It has therefore been seen as an objective measure of anxiety, fear, and/or amygdala activation [157]. Fear- and anxiety-potentiation of startle is used in pharmacological animal models of anxiety, but the translation to human state and trait anxiety has been questioned by prominent startle researchers [214]. However, the lack of significant startle modulation by pleasant and unpleasant images during pregnancy and the postpartum period is surprising in relation to existing literature [162] and our previous experience. By use of the same experimental setup, we have previ-ously observed a significant startle modulation also during pleasant com-pared to unpleasant images in healthy women during the follicular and luteal phase of the menstrual cycle [215]. However, also in that study, the effect of the anticipation stimuli was stronger than the effect of the affective pictures. A contributing factor could be that the images were shown for a shorter time than the anticipation stimuli, as the paradigm used in the study is primarily designed to assess anticipation. Another important issue is the quality of the affective pictures. Although they are validated to induce emotion and

41
arousal, this validation is made in a population of American college students, and for the most part many years ago [210]. Several demographic and con-textual variables therefore differ from our study participants. The low arousal, compared the normative scores, indicated in the ratings from our participants is likely to have contributed to the weak effect of picture va-lence.
The picture ratings were the only subjective ratings of emotional reactiv-ity that we obtained from the women. The mean valence ratings only changed marginally, but the slightly increased mean arousal towards un-pleasant pictures during the postpartum visit are in line with a previous ob-servation of lower affective response during late pregnancy [105].
Sympathetic stress response and labour onset We found an association between sympathetic stress response and days to spontaneous delivery, within the last month of pregnancy in a relatively large sample of 74 individuals. Several earlier studies have shown reduced stress responses in women in late pregnancy, and in the postpartum period, compared to early pregnancy or non-pregnant controls. The studies vary considerably in study design, some are longitudinal during pregnancy [51, 106] or across pregnancy and puerperium [151, 216] and some compare sepa-rate groups of women in different trimesters and/or puerperium with non-pregnant controls [28, 75]. No previous study has, to our knowledge, shown a decrease in stress reactivity during human pregnancy within such a narrow time span. However, when it comes to pain, one study by Cogan and Spin-nato [96] did repeated testing at the end of pregnancy and found a rise in pressure pain thresholds in six women during the last 16 days before parturi-tion. Unlike Cogan and Spinnato [96], we did not find an effect of time to delivery in the subjective pain measures (pain threshold, cold endurance, and VAS-score). This discrepancy is most likely due to the cross-sectional de-sign of our study, where the inter-individual variability in pain experience is not accounted for. It is also possible that sensation of cold pain is differently regulated in pregnancy than pressure pain. Since we had a ceiling effect, in that the majority of women endured the maximum 60 seconds of the cold pressor test, one possibility is also that an effect on cold endurance would be visible if the upper time limit was extended.
We were unable to detect any relation between baseline skin conductance measures and time to parturition. Hence, it is unlikely that our findings are secondary to an increasing sympathetic tone. While our cross-sectional study presents inter-individual differences in sympathetic nervous system reactiv-ity, a longitudinal study design could render more insight into the patterns of pregnancy-induced stress adaptation over the course of pregnancy and puer-perium. Our interpretation of the results is that the sympathetic stress re-

42
sponse is continuously decreasing during the last weeks before giving birth, while the baseline sympathetic level remains stable. Since no similar asso-ciation was seen in absolute cortisol, or awakening response, in relation to time to delivery (study IV), skin conductance or sympathetic response may offer a more sensitive parameter for the investigation of stress response regu-lation before labour. If the relationship is present also in post-term pregnan-cies, skin conductance assessment would have a potential to predict the woman’s chances for having a spontaneous onset of labour, or successful response to labour induction.
β-endorphin, pain, and sympathetic response High β-endorphin levels have been suggested to increase pain tolerance dur-ing pregnancy [102]. A study in our lab has also indicated an increase in an-algesic requirement during labour in women with low circulating levels of β-endorphin in gestational week 37 [101]. Nevertheless, pregnancy-induced analgesia is thought to depend on signalling within the central nervous sys-tem, and plasma levels of β-endorphin cannot be seen as indicative of central opioid analgesic activity [217]. The higher sympathetic activation in women with relatively low β-endorphin that we observed during the cold pressor test could theoretically be a sign of peripheral opioid analgesia [218]. However, our results are very preliminary and need to be replicated. Only one out of three skin conductance variables was significantly elevated in the women with the lowest β-endorphin, and, in line with Saisto et al. [77], the endor-phin level was not correlated with any subjective measure of pain.
Startle modulation across pregnancy and postpartum We found a reduced startle modulation by image anticipation at four to six weeks postpartum, compared to during late pregnancy, in a within-subjects design. The lack of a habituation effect on modulation in the same paradigm in healthy non-pregnant women contradicts the possibility that repeated test-ing accounted for the attenuation during anticipation [215]. Also, the pres-ence of an effect of emotional images in non-pregnant women [215] indicates that the emotional startle modulation in the women in the present study was reduced, in late pregnancy as well as during the postpartum period. Reduced startle modulation during anticipation, as well as by affective images, has been observed in depressed subjects [164, 167], and may therefore be viewed as indicative of emotional dysfunction or a pro-depressive state, in line with other “depression-like” alterations in CNS signalling indices found in preg-nancy [21-23] and the postpartum period [42-44]. However, the subjects in this study did not suffer from depression, and neither do the majority of

43
postpartum women. It is therefore more likely that reduced startle modula-tion in the peripartum represents an adaptive change in emotional reactivity at a time of rapid neuroendocrine changes.
As mentioned in the introduction, the qualitative and quantitative differ-ences in any type of psychophysiological reactivity between late pregnancy - with its high baseline activity, and the postpartum period - with low baseline activity, are sparsely studied, although both states are thought to be charac-terised by hyporeactivity. The lower extent of startle modulation by negative anticipation which was detected postpartum may thus be attributed to the factors suggested to cause postpartum hyporeactivity, i.e. central nervous system oxytocin and prolactin [33, 34]. Since we did not obtain any direct indices of oxytocin, prolactin, or HPA signalling, we can only speculate on this matter. As mentioned in the introduction, the amygdalae are proposed to be the key structure for affective modulation of startle. Breastfeeding women have reduced right amygdala activation in response to arousing pictures at three to six months postpartum, compared to nulliparous controls. However, the amygdala activation is not affected by nasal oxytocin administration in either group, suggesting that acute oxytocin concentration is not mediating this difference [219]. Instead, the relative progesterone deficiency of the postpartum may play a part, as progesterone administration has been re-ported to increase amygdala activation to emotional stimuli in cycling women [220]. This could be argued to be supported by the fact that we saw an association between postpartal serum progesterone and the decreased startle modulation to image anticipation in the postpartum period. However, since ovulation is suppressed during early postpartum and lactation, the variation in serum progesterone (and its neuroactive metabolites) is depend-ent on HPA axis activity [221]. It is therefore possible that the reduced modulation is secondary to the decreased HPA activity.
Studies comparing baseline startle in postpartal rats with diestrous rats have yielded contradictory results. One study argues lactation-induced startle attenuation [222], whereas another found an increase in startle [223]. Rats subjected to rapid progesterone withdrawal display an enhanced baseline startle response [224]. In contrast, no difference in baseline startle response was detected between pregnant and postpartum states in this study or be-tween pregnant women and women examined on postpartal days one to seven in a previous study from our lab [172]. This highlights the importance of caution when translating animal data to human conditions, especially in the field of reproduction.

44
Allopregnanolone and mood during late pregnancy We observed that women who had higher depression scores displayed sig-nificantly lower allopregnanolone serum concentrations. This was also mir-rored in a negative association between late pregnancy serum allopreg-nanolone and self-rated depression scores.
In disagreement with our results, one previous study examining clinically depressed and euthymic pregnant women found no difference in allopreg-nanolone levels [192]. One explanation for the discrepant findings could be that the group sizes of Pearson-Murphy and colleagues were too small to detect the relatively modest effect of allopregnanolone suggested by our results. In addition, Pearson-Murphy and colleagues studied a population of women with first onset of clinical depression during pregnancy; our subjects represent a relatively healthy pregnant population with varying degrees of depressed mood, in most cases outside the range of clinical disease. How-ever, the allopregnanolone precursor 5α-dihydroprogesterone (5α-DHP) was elevated in the depressed women in the Pearson-Murphy study, indicating a related neurosteroid imbalance. While the conflicting results underline the lack of direct clinical value of allopregnanolone during pregnancy, they also provide evidence for the importance of a general neurosteroid balance. Preg-nancy and postpartum period entail large neuroendocrine changes, and often increased stress. It is possible that the elevation of neuroactive progesterone metabolites, with their inhibitory potential, is one of the factors which ensure that most women maintain good mental health throughout these challenges [183, 192], but is of less relevance for the development of clinical depression during pregnancy [192].
Serum concentration is not an ideal measure when studying the effect of allopregnanolone on mood, which is most likely determined by local changes in neurosteroid synthesis and GABAA receptor subunit composition within the brain [225]. However, the association we observed suggests that the massive increase in peripheral feto-placental allopregnanolone synthesis has the potential to affect the central nervous system, and to some extent protect women from negative mood symptoms. Allopregnanolone readily crosses the blood-brain barrier, therefore the serum levels of the steroid are correlated with brain tissue levels [190]. In addition, allopregnanolone is accumulated in certain brain areas in rats [226].
In non-pregnant women, allopregnanolone release is coupled to HPA-axis activity. Allopregnanolone decreases the HPA-response in pregnant rodents [71] and in addition to the direct GABA-enhancing effect, allopregnanolone administration affects the transcription of genes involved in the HPA-axis regulation in rats [227]. Thus, there is a possibility that the association seen in our study is mediated by HPA-axis regulation effects on mood [66].
Based on rodent studies [228, 229], we had expected that high serum allo-pregnanolone would also be a protective factor against symptoms of anxiety.

45
We found no such effect, perhaps because of our relatively non-anxious study population. It must also be mentioned that we asked about state anxi-ety (referring to the momentary anxiety at the visit), and trait anxiety (refer-ring to anxiety prone personality). We did not obtain self-rated symptoms of anxiety in the last days, which would be more analogous to the MADRS-S depression ratings.
We found a significant association between higher serum allopreg-nanolone concentration and fewer days left to spontaneous delivery. Since allopregnanolone is a metabolite of progesterone, this correlation could be assumed to be secondary to the continuous placental progesterone produc-tion. However, we did not observe a correlation between progesterone levels and days to parturition. Allopregnanolone and closely related compounds have previously been implied to be involved in the timing of delivery, but the mechanism likely involves the relative proportions of several steroids [186].
One limitation to our study is the rather healthy study population; a MADRS-S score of 13 or more does not indicate a pathological level of de-pressed mood. Thus, future studies with larger numbers of women suffering from depressed mood are needed to prove the validity of the association with serum allopregnanolone.
Cortisol awakening response in currently and previously depressed pregnant women In line with Shea et al. (2007), we found no difference between the controls and the women with depression during pregnancy. Furthermore, our results gave no support for the hypothesis that biological vulnerability to, or scar after, a depressive episode is visible in the cortisol awakening response of pregnant women with a history of depression, as has been shown in non-pregnant individuals [128].
A meta-analysis of differences in salivary cortisol between depressed and healthy study participants suggests that patients with depression have slightly higher absolute morning cortisol values than healthy controls, al-though this difference does not seem to be clinically significant [129]. The largest study on cortisol awakening response in non-pregnant individuals [128] found around 2.5 nmol/l higher absolute cortisol peak levels in subjects with previous, or ongoing, depression. Since the current study was smaller, we may not have had power to detect a difference in that size range. Thus, we cannot rule out that a small difference in cortisol awakening response may exist between women with ongoing depression and controls. Given the very high inter-individual variation, even in healthy subjects, it is unlikely that a group difference of 2-3 nmol/l is clinically relevant.

46
Extensive assessments of HPA axis function, such as dexamethasone suppression or ACTH-provocation are not feasible during pregnancy due to probable effects on the foetus [230]. The cortisol awakening response offers an opportunity to study cortisol reactivity without subjecting the participants to experimental stressors. However, to disentangle the effects of depression and pregnancy on individual patterns of HPA axis function, it may be neces-sary to consider genetic variations in HPA axis related genes [231], as well as subtypes of depression [3, 232]. It is also possible that an HPA axis vulner-ability is significant and visible in the cortisol awakening response only in patients with severe depression during pregnancy. Still, one study has found significantly higher evening (12 h after awakening) cortisol at the end of the second trimester in 13 women with major depression, compared to 33 non-depressed controls [67]; this observation underlines the link between HPA axis function and depression during pregnancy.
It has been suggested that morning cortisol values are less informative, as they represent the highest levels of the day, and that their inter-individual variation, especially during pregnancy, may be limited by a “ceiling effect” [233]. However, as exemplified in this study, the inter-individual variation in cortisol awakening response is relatively large.
The strengths of this study include a careful group designation based on a structured interview concerning current depressive symptoms, as well as previous episodes, and repeated self-report questionnaires during pregnancy. The participating women were also in a narrow gestational age range, during the last weeks of term pregnancies, which should have minimised the varia-tion caused by the increasing dampening of cortisol reactivity with gesta-tional age. We also excluded all samples where the wake-up sample was collected more than five minutes after the reported wake up-time. However, the lack of further control over participants’ coherence to the sampling pro-tocol must be considered a limitation.
We conclude that there seems to be no essential difference in cortisol awakening response in gestational weeks 36-40, between women with or without previous, or ongoing, depressive episodes.

47
Clinical implications and future perspectives The studies in this thesis have a predominantly basic research approach and primarily concern healthy, pregnant women. The changes in psychophysi-ological and endocrine reactivity during pregnancy are indeed inherently interesting as they represent unique examples of the plasticity of human ho-meostasis. Moreover, increased knowledge of normal variation is crucial to provide a background for deviations with connections to mental health and vulnerability, as touched upon in the included studies. An interesting point is that not only exaggerated reactivity, but also very insufficient reactivity, can be indicative of dysregulation, and thus both ends of the spectrum should be analysed in relation to the middle [234].
As repeatedly stated above, there is a lack of longitudinal studies. Obvi-ously, to have the best theoretical and statistical power for studying any changes during pregnancy and postpartum, it is necessary to obtain pre-pregnancy data, although this may be time-consuming. Also, additional stud-ies following the same individuals over several time-points during and after pregnancy, and after the return to cycling status, are needed.
Maternal stress systems in pregnancy complications Serious pregnancy complications with possible links to dysregulation of HPA and sympathetic activity, such as pregnancy-induced hypertension, gestational diabetes, and pre-term labour, affect many women [12, 85, 235]. Further study of reactivity measures in these patient groups may offer oppor-tunities for prediction of these complications, or relevant outcome measures for preventive interventions.
Maternal stress systems and fetal development The main research activity in the field of maternal stress systems is aimed at consequences for the intrauterine milieu. The blunting of stress responses during pregnancy is suggested to protect the foetus from excess cortisol and vascular constriction compromising uterine blood flow. The fetal production of cortisol is known to be essential for fetal maturation [17], but excess glu-cocorticoids can have adverse effects on the development of the fetal HPA-axis [236, 237], and consequently offspring health through adolescence and adulthood. The mediator of the relationship between antenatal depression, and anxiety, and later child development has therefore been suggested to be excess cortisol exposure. However, the association between maternal distress and maternal cortisol values during pregnancy is not evident; as exemplified in study IV, and by previous studies [238, 239]. In this area of study, too, intra-individual variation may give more insight [200]. Importantly, prenatal

48
anxiety has been linked to decreased placental capacity to protect the foetus from circulating glucocorticoids [240].
Reflections on cross species translation Although stress and pain regulation show common features in human and rodent pregnancy, rodents and humans differ greatly in, for example, gesta-tional length, number of offspring, placental hormone synthesis, and timing of progesterone withdrawal [57, 87], factors which potentially affect labour initiation. Rodent studies emphasise an increase in aggressive behaviour, and a decrease in anxious behaviour in both pregnant and lactating females [110], findings which are not clearly demonstrated in humans. There are also dif-ferences in the social organisation of caring for the offspring. Furthermore, corticosterone reactivity to experimental stressors in rats is in proportion with their total diurnal corticosterone variation, while human cortisol re-sponses to experimental stressors in humans are more subtle [213]. These factors deserve thought when comparing rodent data to endocrine and behav-ioural parameters in humans.

49
Conclusions
The sympathetic stress response is continuously decreasing, even in the last weeks before giving birth at term, while the baseline sympathetic level seems to be stable during the final month. Measurement of skin conductance is a sensitive and non-invasive means to assess sympathetic stress activity, and is an interesting tool for increasing the knowledge about sympathetic reactions during pregnancy. Given the complex interactions between stress reactivity and labour initiation, this subject deserves more research attention.
Healthy women with stable anxiety and depression scores have decreased startle modulation by anticipation in the puerperium in comparison with late pregnancy. Since altered startle response modulation has previously been associated with anxiety and depressive states, this result provides a back-ground for further studies examining emotional modulation in women who develop anxiety and/or depressive disorders during pregnancy and the post-partum period. Our finding of relatively low serum allopregnanolone in late pregnant women with moderate depressive symptoms, compared with women with little depressive symptoms, is in line with a growing body of evidence for the importance of neurosteroid balance in mood regulation. The fact that our study population was fairly healthy and had low levels of anxiety hinders generalisation to women with ongoing psychiatric disorders. A history of depression, or ongoing depression, is not an important determi-nant for cortisol awakening response during late pregnancy. Thus, pregnancy seems to smooth out, or reverse, the HPA alterations associated with depres-sion in non-pregnant individuals. Further studies examining cortisol reactiv-ity in relation to depression during pregnancy will be needed to consider additional determinants, like depression subtypes and HPA axis related gene polymorphisms.

50
Sammanfattning på svenska
Under en graviditet anpassas den maternella fysiologin på flera sätt för att kroppen ska kunna ta hänsyn till fostrets behov. Den fysiologiska anpass-ningen framkallas genom hormoner som produceras av moderkakan och av fostret. Parallellt höjs också aktiviteten i hypotalamus-hypofys-binjure- (HPA-) axeln, som styr produktionen av stresshormonet kortisol. Även den sympatiska delen av det autonoma nervsystemet ökar sin basaktivitet. Dock minskar reaktiviteten (svaret vid stress) både i det sympatiska nervsystemet och i HPA-axeln. Man har även kunnat påvisa förändringar i det centrala nervsystemet i sen graviditet och under postpartumperioden. Omställningar-na förändrar den gravida kvinnans metabolism och immunförsvar och har också visats vara viktiga för såväl reglering av förlossningsstarten som fost-rets utveckling. Eftersom stressfysiologiska avvikelser (som till exempel högt kortisol) har kopplats samman med psykisk ohälsa hos icke-gravida har det föreslagits att dessa förändringar utsätter gravida kvinnors psykiska hälsa för en påfrestning som kan orsaka depression och ångest. De flesta kvinnor mår dock bra under graviditet och efter förlossning, vilket tyder på att det bland alla förändringar som sker även ingår element som skyddar mot skad-liga effekter.
Målet med denna avhandling var att undersöka några aspekter av stressre-aktivitet under slutet av graviditeten i relation till förlossningsstart, emotio-nell reaktivitet och psykisk hälsa. I samtliga delstudier deltog kvinnor med högst fem veckor kvar till beräknat förlossningsdatum.
Delstudie I Förutom den sänkta stressreaktiviteten verkar också smärtkänsligheten minska under sen graviditet, både hos människor och hos andra däggdjur. Hur smärtkänslighet upplevs är dock högst subjektivt och kan vara svårt att mäta. Hudkonduktans, det vill säga aktiviteten i de eccrina svettkörtlarna i handflator och på fotsulor, styrs helt av sympatiska nervsystemet och har föreslagits vara ett objektivt mått på smärtintensitet som kan komplettera självrapportering. I delstudie ett undersöktes smärta och sympatiskt svar vid köldstress i relation till hur många dagar kvinnorna hade kvar till förloss-ning. Sympatisk aktivitet (hudkonduktans) mättes medan kvinnan höll en hand i iskallt vatten. Vi mätte också hur lång tid det tog innan kvinnan kände köldsmärta samt hur lång tid hon stod ut med att hålla handen i vattnet.

51
Kvinnan fick sedan skatta hur stark smärtan varit. Det visade sig att det sympatiska svaret var lägre hos de kvinnor som hade färre dagar kvar till en spontan förlossningsstart, medan smärtkänslighet inte visade något samband med tid till förlossning. Detta tyder på att den sympatiska reaktiviteten minskar under de sista veckorna före förlossningen, även om detta inte helt kan bevisas i en tvärsnittsstudie. Ytterligare studier på sympatisk reaktivitet i förhållande till förlossningsstart kan kanske ge bättre metoder för att förut-säga tidpunkten för spontan förlossning.
Delstudie II Trots att de förändringar som sker i det centrala nervsystemet under sen gra-viditet och under amningsperioden har liknats vid de avvikelser som påvisats hos deprimerade individer finns det inte så mycket kunskap om hur den emotionella regleringen påverkas. Ett sätt att studera emotionell reglering är att undersöka hur den blinkreflex som uppstår vid ett plötsligt ljud påverkas av positiva och negativa känslostimuli. Blinkreflexen är en hjärnstamsreflex, som automatiskt skyddar ögonen vid plötsliga sinnesintryck. Blinkreflexen är alltså inte viljestyrd, men blir kraftigare vid rädsla och negativa känslor. Detta eftersom den bland annat påverkas av aktiveringen av amygdala, det limbiska systemets rädslocenter. Unikt för blinkreflexen är att den också dämpas av starka positiva, eller appetitiva, känslotillstånd. I den andra del-studien mättes emotionell blinkreflex-modulering under slutet av gravidite-ten och några veckor efter förlossningen. Som känslostimuli visades behag-liga och obehagliga bilder på en datorskärm. Dessutom var skärmen grön före de behagliga bilderna och röd före de obehagliga bilderna för att stimu-lera positiv respektive negativ förväntan. Vi såg ingen blinkreflex-modulerande effekt av de behagliga och obehagliga bilderna under något av de två testtillfällena, vilket tyder på minskad modulering jämfört med icke-gravida kvinnor. Däremot var blinkreflexens magnitud signifikant högre vid negativ än vid positiv förväntan under graviditet men inte vid postpartumbe-söket. Detta kan tolkas som att den emotionella blinkreflex-moduleringen är dämpad både under graviditet och postpartumperiod och även ytterligare dämpad under postpartumperioden än under graviditeten. Dock såg vi inte att den dämpade emotionella moduleringen skulle hänga samman med hur kvinnorna mådde eller med hur de uppfattade bildernas känslomässiga inne-håll.
Delstudie III Allopregnanolon är en neuroaktiv progesteronmetabolit som har uppmätts i lägre nivåer i blod och cerebrospinalvätska hos deprimerade individer än hos icke-deprimerade. Under en graviditet mer än tiofaldigas serumnivån av allopregnanolon och i djurförsök har allopregnanolon visats mediera den

52
minskade HPA-axel-reaktiviteten under dräktighet och digivning. I den tred-je delstudien analyserades serumnivåer av allopregnanolon i relation till självskattade ångest- och depressionssymptom under sen graviditet. Studie-deltagarna rapporterade över lag låg förekomst av ångest- och depressions-symptom, men de kvinnor som hade högst depressionsskattningar visade sig ha signifikant lägre allopregnanolonnivåer än resten av gruppen. Ångest-skattningarna var däremot inte associerade med serumkoncentrationerna av allopregnanolon. Resultaten skulle kunna tyda på att den höga nivån av allo-pregnanolon under graviditeten till viss del skyddar mot de påfrestningar som de övriga endokrinologiska förändringarna kan utgöra för den maternel-la mentala hälsan.
Delstudie IV Stresshormonet kortisol stiger hastigt vid uppvaknande och är som högst cirka 30 minuter efteråt. Detta kortisolsvar har visat sig vara större hos de-primerade patienter, och är även högt hos friska individer som tidigare varit deprimerade. Kortisolsvaret vid uppvaknande försvagas till viss del under graviditeten, i takt med att basalnivån stiger, men är fortfarande påvisbart under sen graviditet. I delstudie fyra mättes kortisolsvaret vid uppvaknande hos tre grupper av kvinnor under sen graviditet: kvinnor som lidit av depres-sion under graviditeten, nu friska kvinnor som tidigare lidit av depression samt kvinnor som aldrig drabbats av depression. Det visade sig att det inte fanns någon statistisk signifikant skillnad i kortisolsvar, eller skillnad i abso-lut kortisolnivå, mellan grupperna. Detta tyder på att det ökade kortisolsvar som visats under och efter en depression hos icke-gravida inte följer med under graviditeten.

53
Acknowledgments
This work was carried out at the Department of Women’s and Children’s Health, Uppsala University. I would like to express my gratitude to the people who made it possible, and worthwhile. Thank you! All participating women who have given me time and blood; many of you even gave your babies to me for a while. Together you contributed with the most important part of the studies. Inger Sundström-Poromaa, Professor at the Department of Women’s and Children’s Health, and my main supervisor. Thank you for constantly inter-rupting whatever is on your hands to supervise me on more or less work related problems. Once in your office, it always turned out there never really was a problem! Helena Åkerud, Associate Professor at the Department of Women’s and Children’s Health, and my co-supervisor. You have truly co-supervised me with a rare efficiency and supported me from the first day. Also, thank you for always taking my problems seriously! Ove Axelsson and Jan Gustafsson, former and current head of the Depart-ment of Women’s and Children’s Health, for the possibility to perform my doctoral studies at the department. Victoria Risbrough, Assistant professor at the Department of Psychiatry, University of California, for providing the affective startle paradigm and startle knowledge. Alkistis Skalkidou, Associate Professor at the Department of Women’s and Children’s Health, co-author, and reseledare extraordinaire. Your generous sharing of knowledge and smiles is an inspiration. Dr. Maria Jonsson, co-author, for the possibility to participate in research in a clinical setting.

54
Torbjörn Bäckström, Professor at the Department of Obstetrics and Gyne-cology at Umeå University and co-author. Thank you for contributing with experience and knowledge on neurosteroids. Matts Olovsson, Professor at the Department of Women’s and Children’s Health. Thank you for always checking if I am alright if you find me work-ing outside of office hours. Anneli Stavreus-Evers, Associate Professor at the Department of Women’s and Children’s Health. Your colourfulness and experience really add to our working environment. Elin Bannbers, fellow småflicka, för att du har trampat upp min väg, och för att du stått och väntat med jämnt humör och ytterkläderna på i samman-lagt 700 timmar under dessa år. Tack också för roliga minnen från resorna och för att du förmanar mig så att jag aldrig äter så gammal mat igen. Lena Moby, tack för att du höll reda på mig när jag var ny och förvirrad. Från dig kan man alltid få både känslomässigt stöd och ett ärligt svar! Ulla, Maggan och Vera, tack för att ni lämnade efter er en labbkultur med mycket fika och humor, som vi gör vårt bästa för att förvalta. Ingegerd Clason, tack för att du lyst upp många gråa dagar i centrifugrum-met med din humor! Alla andra! Malin Helmestam och Eva Davey, mina närmsta grannar på känslokontoret, jag är otroligt tacksam för att ni inte bara stått ut med mitt ständiga snack, utan också går in för att dela mina frustrationer och glädje-ämnen - flera gånger i timmen. Ett extra tack till Malin för hennes brinnande engagemang mot plast. Theodora Kunovac-Kallak, som alltid är hjälpsam och lojal, samt skrattar gott åt mina tillkortakommanden. Malin Gingnell, denna renässansmänniska, som står för både seriös rådgivning och kreativa upptåg. Karin Lindgren, vars skönsång och människokärlek tränger igenom betong. Katja Junus, ofta vitklädd, ofta svartklädd, alltid nyanserad. Fati-mah Dabo, tack för intressanta diskussioner under tiden du arbetade hårt inför din disputation. Sara Sylvén, tack för glada tillrop och möten kring BASIC. Erika Comasco, tack för spännande samarbete, jag hoppas att det blir fler. Helena Volgsten, Helena Kaihola, Anna-Karin Hannerfors, Stavros Iliadis och Jocelien Olivier, tack för alla vetenskapliga och mer personliga diskussioner. Christine, Malin O och Narges, tack för era alltför kortvariga insatser på labbet, och trevliga sykvällar.

55
Jane och Ann-Sofie, tack för att ni ryckt in och tagit blodprov och pratat om livet i centrifugrummet.
Alla nuvarande och tidigare kollegor i och kring gruppen och labbet, tack för seminarier, journal clubs och härligt resesällskap. Administratörerna, som fixar allt inom ekonomi, data och ”papper” på nolltid – och verkar ha oändligt tålamod. Alla studenter och praktikanter, som varit på labb och hjälpt till med mina projekt, eller frågat mig frågor som fått mig att inse att jag ändå tagit mig framåt i statistiken och forskningsmetodiken under årens lopp. Alla engagerade själar i ESMeralda och alla ungdomar som kommer till läxläsningstillfällena och ger mig perspektiv på att ha spenderat en dag på att skriva ut etiketter på fel sida. Mina ”civila” vänner, som gett mig styrka och glädje genom telefon på mina promenader till och från jobbet, med gratinerad hummer i Ulleråker, på Kringlan i Göteborg, med Idolkväll på Kantorn, i Toskåne, i joggingspåret i stadsskogen, i Flogstaringens koloniförening, genom att ta ord på allvar och se det komiska i allt som händer, eller med dumplings och vykort. Siv, Göran, Maria och Tove Bergström med familjer, tack för att ni för-gyller mitt liv med fikastunder, svamputflykter, kalas, reparationer, bilskjuts, telefonrådgivning, etcetera. Tack mina föräldrar, Eva Hellgren och Håkan Runeson, för allt - framför allt för mina syskon. Tack också till mina syskon, Ian, Eric och Sanna, som gör mig så stolt och glad. Tack också resten av familjen: Ana och Stig, most-rar, faster, onklar och kusiner för all uppmuntran. Tina, tack för kärlek och för att du hjälper mig med bildhantering, och för att vi hjälps åt med allt annat som är viktigt i livet.

56
References
1. Greenstein, B. and Wood, D.F., The endocrine system at a glance. 2nd ed. 2006, Malden, Mass., USA: Blackwell Pub. 120 p.
2. Bloch, M., Daly, R.C., and Rubinow, D.R. Endocrine factors in the etiology of postpartum depression. Compr Psychiatry., 2003. 44(3): 234-46.
3. Kammerer, M., Taylor, A., and Glover, V. The HPA axis and perinatal depression: a hypothesis. Arch Womens Ment Health., 2006. 9(4): 187-96.
4. Buckwalter, J.G., Buckwalter, D.K., Bluestein, B.W., and Stanczyk, F.Z. Pregnancy and post partum: changes in cognition and mood. Prog Brain Res, 2001. 133: 303-19.
5. Dorr, H.G., Heller, A., Versmold, H.T., Sippell, W.G., Herrmann, M., Bidlingmaier, F., and Knorr, D. Longitudinal study of progestins, mineralocorticoids, and glucocorticoids throughout human pregnancy. J Clin Endocrinol Metab, 1989. 68(5): 863-8.
6. Schubring, C., Englaro, P., Siebler, T., Blum, W.F., Demirakca, T., Kratzsch, J., and Kiess, W. Longitudinal analysis of maternal serum leptin levels during pregnancy, at birth and up to six weeks after birth: relation to body mass index, skinfolds, sex steroids and umbilical cord blood leptin levels. Horm Res, 1998. 50(5): 276-83.
7. Stalla, G.K., Bost, H., Stalla, J., Kaliebe, T., Dorr, H.G., Pfeiffer, D., von Werder, K., and Muller, O.A. Human corticotropin-releasing hormone during pregnancy. Gynecol Endocrinol, 1989. 3(1): 1-10.
8. Jung, C., Ho, J.T., Torpy, D.J., Rogers, A., Doogue, M., Lewis, J.G., Czajko, R.J., and Inder, W.J. A longitudinal study of plasma and urinary cortisol in pregnancy and postpartum. J Clin Endocrinol Metab, 2011. 96(5): 1533-40.
9. Harris, B., Lovett, L., Newcombe, R.G., Read, G.F., Walker, R., and Riad-Fahmy, D. Maternity blues and major endocrine changes: Cardiff puerperal mood and hormone study II. BMJ., 1994. 308(6934): 949-53.
10. Mastorakos, G. and Ilias, I. Maternal and fetal hypothalamic-pituitary-adrenal axes during pregnancy and postpartum. Ann N Y Acad Sci, 2003. 997: 136-49.
11. Luppi, P. How immune mechanisms are affected by pregnancy. Vaccine, 2003. 21(24): 3352-7.
12. Greenwood, J.P., Scott, E.M., Stoker, J.B., Walker, J.J., and Mary, D.A. Sympathetic neural mechanisms in normal and hypertensive pregnancy in humans. Circulation, 2001. 104(18): 2200-4.
13. Pasco, J.A., Jacka, F.N., Williams, L.J., Henry, M.J., Nicholson, G.C., Kotowicz, M.A., and Berk, M. Leptin in depressed women: cross-sectional and longitudinal data from an epidemiologic study. J Affect Disord, 2008. 107(1-3): 221-5.
14. Lof, M., Olausson, H., Bostrom, K., Janerot-Sjoberg, B., Sohlstrom, A., and Forsum, E. Changes in basal metabolic rate during pregnancy in relation to changes in body weight and composition, cardiac output, insulin-like growth

57
factor I, and thyroid hormones and in relation to fetal growth. Am J Clin Nutr, 2005. 81(3): 678-85.
15. Speroff, L. and Fritz, M.A., Reproduction and The Thyroid, in Clinical gynecologic endocrinology and infertility. 2005, Lippincott Williams & Wilkins: Philadelphia. p. x, 1334 p.
16. Damjanovic, S.S., Stojic, R.V., Lalic, N.M., Jotic, A.Z., Macut, D.P., Ognjanovic, S.I., Petakov, M.S., and Popovic, B.M. Relationship between basal metabolic rate and cortisol secretion throughout pregnancy. Endocrine, 2009. 35(2): 262-8.
17. Pepe, G.J. and Albrecht, E.D. Actions of placental and fetal adrenal steroid hormones in primate pregnancy. Endocr Rev, 1995. 16(5): 608-48.
18. Bowen, J.M., Chamley, L., Keelan, J.A., and Mitchell, M.D. Cytokines of the placenta and extra-placental membranes: roles and regulation during human pregnancy and parturition. Placenta, 2002. 23(4): 257-73.
19. Maes, M. Depression is an inflammatory disease, but cell-mediated immune activation is the key component of depression. Prog Neuropsychopharmacol Biol Psychiatry, 2011. 35(3): 664-75.
20. Blackmore, E.R., Moynihan, J.A., Rubinow, D.R., Pressman, E.K., Gilchrist, M., and O'Connor, T.G. Psychiatric symptoms and proinflammatory cytokines in pregnancy. Psychosom Med, 2011. 73(8): 656-63.
21. Altemus, M., Fong, J., Yang, R., Damast, S., Luine, V., and Ferguson, D. Changes in cerebrospinal fluid neurochemistry during pregnancy. Biol Psychiatry, 2004. 56(6): 386-92.
22. Lommatzsch, M., Hornych, K., Zingler, C., Schuff-Werner, P., Hoppner, J., and Virchow, J.C. Maternal serum concentrations of BDNF and depression in the perinatal period. Psychoneuroendocrinology, 2006. 31(3): 388-94.
23. Schrocksnadel, H., Baier-Bitterlich, G., Dapunt, O., Wachter, H., and Fuchs, D. Decreased plasma tryptophan in pregnancy. Obstet Gynecol, 1996. 88(1): 47-50.
24. Placidi, G.P., Oquendo, M.A., Malone, K.M., Huang, Y.Y., Ellis, S.P., and Mann, J.J. Aggressivity, suicide attempts, and depression: relationship to cerebrospinal fluid monoamine metabolite levels. Biol Psychiatry, 2001. 50(10): 783-91.
25. Smolen, A., Smolen, T.N., and van de Kamp, J.L. Alterations in brain catecholamines during pregnancy. Pharmacol Biochem Behav, 1987. 26(3): 613-8.
26. Smolen, A., Smolen, T.N., and Han, P.C. Alterations in regional brain GABA concentration and turnover during pregnancy. Pharmacol Biochem Behav, 1993. 44(1): 63-9.
27. Glaser, J., Russell, V.A., and Taljaard, J.J. Rat brain hypothalamic and hippocampal monoamine and hippocampal beta-adrenergic receptor changes during pregnancy. Brain Res, 1992. 577(2): 293-9.
28. Altemus, M., Deuster, P.A., Galliven, E., Carter, C.S., and Gold, P.W. Suppression of hypothalmic-pituitary-adrenal axis responses to stress in lactating women. J Clin Endocrinol Metab, 1995. 80(10): 2954-9.
29. Jackson, E. and Glasier, A. Return of ovulation and menses in postpartum nonlactating women: a systematic review. Obstet Gynecol, 2011. 117(3): 657-62.
30. Butte, N.F., Hopkinson, J.M., and Nicolson, M.A. Leptin in human reproduction: serum leptin levels in pregnant and lactating women. J Clin Endocrinol Metab, 1997. 82(2): 585-9.

58
31. Pang, W.W. and Hartmann, P.E. Initiation of human lactation: secretory differentiation and secretory activation. J Mammary Gland Biol Neoplasia, 2007. 12(4): 211-21.
32. Torner, L., Toschi, N., Pohlinger, A., Landgraf, R., and Neumann, I.D. Anxiolytic and anti-stress effects of brain prolactin: improved efficacy of antisense targeting of the prolactin receptor by molecular modeling. J Neurosci, 2001. 21(9): 3207-14.
33. Torner, L., Toschi, N., Nava, G., Clapp, C., and Neumann, I.D. Increased hypothalamic expression of prolactin in lactation: involvement in behavioural and neuroendocrine stress responses. Eur J Neurosci, 2002. 15(8): 1381-9.
34. Heinrichs, M., Meinlschmidt, G., Neumann, I., Wagner, S., Kirschbaum, C., Ehlert, U., and Hellhammer, D.H. Effects of suckling on hypothalamic-pituitary-adrenal axis responses to psychosocial stress in postpartum lactating women. J Clin Endocrinol Metab, 2001. 86(10): 4798-804.
35. Ben-Jonathan, N., Mershon, J.L., Allen, D.L., and Steinmetz, R.W. Extrapituitary prolactin: distribution, regulation, functions, and clinical aspects. Endocr Rev, 1996. 17(6): 639-69.
36. Corwin, E.J. and Pajer, K. The psychoneuroimmunology of postpartum depression. J Womens Health (Larchmt), 2008. 17(9): 1529-34.
37. Levine, A., Zagoory-Sharon, O., Feldman, R., and Weller, A. Oxytocin during pregnancy and early postpartum: individual patterns and maternal-fetal attachment. Peptides, 2007. 28(6): 1162-9.
38. Dawood, M.Y., Khan-Dawood, F.S., Wahi, R.S., and Fuchs, F. Oxytocin release and plasma anterior pituitary and gonadal hormones in women during lactation. J Clin Endocrinol Metab, 1981. 52(4): 678-83.
39. Slattery, D.A. and Neumann, I.D. No stress please! Mechanisms of stress hyporesponsiveness of the maternal brain. J Physiol, 2008. 586(2): 377-85.
40. Amico, J.A., Challinor, S.M., and Cameron, J.L. Pattern of oxytocin concentrations in the plasma and cerebrospinal fluid of lactating rhesus monkeys (Macaca mulatta): evidence for functionally independent oxytocinergic pathways in primates. J Clin Endocrinol Metab, 1990. 71(6): 1531-5.
41. Takagi, T., Tanizawa, O., Otsuki, Y., Sugita, N., Haruta, M., and Yamaji, K. Oxytocin in the cerebrospinal fluid and plasma of pregnant and nonpregnant subjects. Horm Metab Res, 1985. 17(6): 308-10.
42. Bailara, K.M., Henry, C., Lestage, J., et al. Decreased brain tryptophan availability as a partial determinant of post-partum blues. Psychoneuroendocrinology, 2006. 31(3): 407-13.
43. Doornbos, B., Fekkes, D., Tanke, M.A., de Jonge, P., and Korf, J. Sequential serotonin and noradrenalin associated processes involved in postpartum blues. Prog Neuropsychopharmacol Biol Psychiatry, 2008. 32(5): 1320-5.
44. Epperson, C.N., Gueorguieva, R., Czarkowski, K.A., Stiklus, S., Sellers, E., Krystal, J.H., Rothman, D.L., and Mason, G.F. Preliminary evidence of reduced occipital GABA concentrations in puerperal women: a 1H-MRS study. Psychopharmacology (Berl), 2006. 186(3): 425-33.
45. Sacher, J., Wilson, A.A., Houle, S., Rusjan, P., Hassan, S., Bloomfield, P.M., Stewart, D.E., and Meyer, J.H. Elevated brain monoamine oxidase A binding in the early postpartum period. Arch Gen Psychiatry, 2010. 67(5): 468-74.
46. Ulrich-Lai, Y.M. and Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat Rev Neurosci, 2009. 10(6): 397-409.

59
47. Kaltsas, G.A. and Chrousos, G.P., The Neuroendocrinology of Stress, in Handbook of Psychophysiology, Cacioppo JT, T.L., Berntson GG, Editor. 2007, Cambridge University Press. p. 303-318.
48. Kudielka, B.M., Schommer, N.C., Hellhammer, D.H., and Kirschbaum, C. Acute HPA axis responses, heart rate, and mood changes to psychosocial stress (TSST) in humans at different times of day. Psychoneuroendocrinology, 2004. 29(8): 983-92.
49. Chrousos, G.P., Torpy, D.J., and Gold, P.W. Interactions between the hypothalamic-pituitary-adrenal axis and the female reproductive system: clinical implications. Ann Intern Med, 1998. 129(3): 229-40.
50. Kalantaridou, S.N., Zoumakis, E., Makrigiannakis, A., Lavasidis, L.G., Vrekoussis, T., and Chrousos, G.P. Corticotropin-releasing hormone, stress and human reproduction: an update. J Reprod Immunol, 2010. 85(1): 33-9.
51. DiPietro, J.A., Costigan, K.A., and Gurewitsch, E.D. Maternal psychophysiological change during the second half of gestation. Biol Psychol, 2005. 69(1): 23-38.
52. Schulte, H.M., Weisner, D., and Allolio, B. The corticotrophin releasing hormone test in late pregnancy: lack of adrenocorticotrophin and cortisol response. Clin Endocrinol (Oxf), 1990. 33(1): 99-106.
53. Matthews, K.A. and Rodin, J. Pregnancy alters blood pressure responses to psychological and physical challenge. Psychophysiology, 1992. 29(2): 232-40.
54. Challis, J.R.G., Matthews, S.G., Gibb, W., and Lye, S.J. Endocrine and paracrine regulation of birth at term and preterm. Endocr Rev, 2000. 21(5): 514-50.
55. Robinson, B.G., Emanuel, R.L., Frim, D.M., and Majzoub, J.A. Glucocorticoid stimulates expression of corticotropin-releasing hormone gene in human placenta. Proc Natl Acad Sci U S A, 1988. 85(14): 5244-8.
56. Murphy, B.E. Human fetal serum cortisol levels related to gestational age: evidence of a midgestational fall and a steep late gestational rise, independent of sex or mode of delivery. Am J Obstet Gynecol, 1982. 144(3): 276-82.
57. Speroff, L. and Fritz, M.A., The Endocrinology of Pregnancy, in Clinical gynecologic endocrinology and infertility. 2005, Lippincott Williams & Wilkins: Philadelphia. p. x, 1334 p.
58. Nolten, W.E. and Rueckert, P.A. Elevated free cortisol index in pregnancy: possible regulatory mechanisms. Am J Obstet Gynecol, 1981. 139(4): 492-8.
59. Owens, P.C., Smith, R., Brinsmead, M.W., et al. Postnatal disappearance of the pregnancy-associated reduced sensitivity of plasma cortisol to feedback inhibition. Life Sci, 1987. 41(14): 1745-50.
60. Del Rio, G., Velardo, A., Zizzo, G., Avogaro, A., Cipolli, C., Della Casa, L., Marrama, P., and MacDonald, I.A. Effect of estradiol on the sympathoadrenal response to mental stress in normal men. J Clin Endocrinol Metab, 1994. 79(3): 836-40.
61. Komesaroff, P.A., Esler, M.D., and Sudhir, K. Estrogen supplementation attenuates glucocorticoid and catecholamine responses to mental stress in perimenopausal women. J Clin Endocrinol Metab, 1999. 84(2): 606-10.
62. Allolio, B., Hoffmann, J., Linton, E.A., Winkelmann, W., Kusche, M., and Schulte, H.M. Diurnal salivary cortisol patterns during pregnancy and after delivery: relationship to plasma corticotrophin-releasing-hormone. Clin Endocrinol (Oxf), 1990. 33(2): 279-89.
63. Altemus, M., Redwine, L.S., Leong, Y.M., Frye, C.A., Porges, S.W., and Carter, C.S. Responses to laboratory psychosocial stress in postpartum women. Psychosom Med, 2001. 63(5): 814-21.

60
64. Carter, C.S., Altemus, M., and Chrousos, G.P. Neuroendocrine and emotional changes in the post-partum period. Prog Brain Res., 2001. 133: 241-9.
65. Magiakou, M.A., Mastorakos, G., Rabin, D., Dubbert, B., Gold, P.W., and Chrousos, G.P. Hypothalamic corticotropin-releasing hormone suppression during the postpartum period: implications for the increase in psychiatric manifestations at this time. J Clin Endocrinol Metab, 1996. 81(5): 1912-7.
66. Rich-Edwards, J.W., Mohllajee, A.P., Kleinman, K., Hacker, M.R., Majzoub, J., Wright, R.J., and Gillman, M.W. Elevated midpregnancy corticotropin-releasing hormone is associated with prenatal, but not postpartum, maternal depression. J Clin Endocrinol Metab, 2008. 93(5): 1946-51.
67. O'Keane, V., Lightman, S., Marsh, M., Pawlby, S., Papadopoulos, A.S., Taylor, A., Moore, R., and Patrick, K. Increased pituitary-adrenal activation and shortened gestation in a sample of depressed pregnant women: a pilot study. J Affect Disord, 2011. 130(1-2): 300-5.
68. Meltzer-Brody, S., Stuebe, A., Dole, N., Savitz, D., Rubinow, D., and Thorp, J. Elevated corticotropin releasing hormone (CRH) during pregnancy and risk of postpartum depression (PPD). J Clin Endocrinol Metab, 2011. 96(1): E40-7.
69. Yim, I.S., Glynn, L.M., Dunkel-Schetter, C., Hobel, C.J., Chicz-DeMet, A., and Sandman, C.A. Risk of postpartum depressive symptoms with elevated corticotropin-releasing hormone in human pregnancy. Arch Gen Psychiatry, 2009. 66(2): 162-9.
70. Windle, R.J., Wood, S.A., Kershaw, Y.M., Lightman, S.L., and Ingram, C.D. Adaptive changes in basal and stress-induced HPA activity in lactating and post-lactating female rats. Endocrinology, 2013. 154(2): 749-61.
71. Brunton, P.J. and Russell, J.A. Allopregnanolone and suppressed hypothalamo-pituitary-adrenal axis stress responses in late pregnancy in the rat. Stress, 2011. 14(1): 6-12.
72. Brunton, P.J., Russell, J.A., and Douglas, A.J. Adaptive responses of the maternal hypothalamic-pituitary-adrenal axis during pregnancy and lactation. J Neuroendocrinol, 2008. 20(6): 764-76.
73. DiPietro, J.A., Mendelson, T., Williams, E.L., and Costigan, K.A. Physiological blunting during pregnancy extends to induced relaxation. Biol Psychol, 2012. 89(1): 14-20.
74. De Weerth, C., Wied, G.D., Jansen, L.M., and Buitelaar, J.K. Cardiovascular and cortisol responses to a psychological stressor during pregnancy. Acta Obstet Gynecol Scand, 2007. 86(10): 1181-92.
75. Nierop, A., Bratsikas, A., Klinkenberg, A., Nater, U.M., Zimmermann, R., and Ehlert, U. Prolonged salivary cortisol recovery in second-trimester pregnant women and attenuated salivary alpha-amylase responses to psychosocial stress in human pregnancy. J Clin Endocrinol Metab, 2006. 91(4): 1329-35.
76. Saisto, T., Kaaja, R., Ylikorkala, O., and Halmesmaki, E. Reduced pain tolerance during and after pregnancy in women suffering from fear of labor. Pain, 2001. 93(2): 123-7.
77. Saisto, T., Kaaja, R., Helske, S., Ylikorkala, O., and Halmesmaki, E. Norepinephrine, adrenocorticotropin, cortisol and beta-endorphin in women suffering from fear of labor: responses to the cold pressor test during and after pregnancy. Acta Obstet Gynecol Scand, 2004. 83(1): 19-26.
78. de Weerth, C. and Buitelaar, J.K. Physiological stress reactivity in human pregnancy--a review. Neurosci Biobehav Rev, 2005. 29(2): 295-312.
79. Buckingham, J.C., Glucocorticoids, Effects of Stress on, in Encyclopedia of stress, Fink, G., Editor. 2000, Academic Press: San Diego.

61
80. Kieler, H., Axelsson, O., Nilsson, S., and Waldenstrom, U. The length of human pregnancy as calculated by ultrasonographic measurement of the fetal biparietal diameter. Ultrasound Obstet Gynecol, 1995. 6(5): 353-7.
81. Tucker, J. and McGuire, W. Epidemiology of preterm birth. BMJ, 2004. 329(7467): 675-8.
82. Martin, J.A., Hamilton, B.E., Ventura, S.J., Osterman, M.J., Kirmeyer, S., Mathews, T.J., and Wilson, E.C. Births: final data for 2009. Natl Vital Stat Rep, 2011. 60(1): 1-70.
83. Roos, N., Sahlin, L., Ekman-Ordeberg, G., Kieler, H., and Stephansson, O. Maternal risk factors for postterm pregnancy and cesarean delivery following labor induction. Acta Obstet Gynecol Scand, 2010. 89(8): 1003-10.
84. McLean, M., Bisits, A., Davies, J., Woods, R., Lowry, P., and Smith, R. A placental clock controlling the length of human pregnancy. Nat Med, 1995. 1(5): 460-3.
85. Wadhwa, P.D., Garite, T.J., Porto, M., Glynn, L., Chicz-DeMet, A., Dunkel-Schetter, C., and Sandman, C.A. Placental corticotropin-releasing hormone (CRH), spontaneous preterm birth, and fetal growth restriction: a prospective investigation. Am J Obstet Gynecol, 2004. 191(4): 1063-9.
86. Zakar, T. and Hertelendy, F. Progesterone withdrawal: key to parturition. Am J Obstet Gynecol, 2007. 196(4): 289-96.
87. Karalis, K., Goodwin, G., and Majzoub, J.A. Cortisol blockade of progesterone: a possible molecular mechanism involved in the initiation of human labor. Nat Med, 1996. 2(5): 556-60.
88. Torricelli, M., Novembri, R., Voltolini, C., et al. Biochemical and biophysical predictors of the response to the induction of labor in nulliparous postterm pregnancy. Am J Obstet Gynecol, 2011. 204(1): 39 e1-6.
89. Leighton, B.L., Halpern, S.H., and Wilson, D.B. Lumbar sympathetic blocks speed early and second stage induced labor in nulliparous women. Anesthesiology, 1999. 90(4): 1039-46.
90. Brauer, M.M. Cellular and molecular mechanisms underlying plasticity in uterine sympathetic nerves. Auton Neurosci, 2008. 140(1-2): 1-16.
91. Lee, H.J., Caldwell, H.K., Macbeth, A.H., Tolu, S.G., and Young, W.S., 3rd A conditional knockout mouse line of the oxytocin receptor. Endocrinology, 2008. 149(7): 3256-63.
92. Nishimori, K., Young, L.J., Guo, Q., Wang, Z., Insel, T.R., and Matzuk, M.M. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proc Natl Acad Sci U S A, 1996. 93(21): 11699-704.
93. Goldenberg, R.L., Culhane, J.F., Iams, J.D., and Romero, R. Epidemiology and causes of preterm birth. Lancet, 2008. 371(9606): 75-84.
94. Denison, F.C., Price, J., Graham, C., Wild, S., and Liston, W.A. Maternal obesity, length of gestation, risk of postdates pregnancy and spontaneous onset of labour at term. BJOG, 2008. 115(6): 720-5.
95. Carvalho, B., Angst, M.S., Fuller, A.J., Lin, E., Mathusamy, A.D., and Riley, E.T. Experimental heat pain for detecting pregnancy-induced analgesia in humans. Anesth Analg., 2006. 103(5): 1283-7.
96. Cogan, R. and Spinnato, J.A. Pain and discomfort thresholds in late pregnancy. Pain, 1986. 27(1): 63-8.
97. Gintzler, A.R. and Liu, N.J. The maternal spinal cord: biochemical and physiological correlates of steroid-activated antinociceptive processes. Prog Brain Res, 2001. 133: 83-97.

62
98. Jarvis, S., McLean, K.A., Chirnside, J., Deans, L.A., Calvert, S.K., Molony, V., and Lawrence, A.B. Opioid-mediated changes in nociceptive threshold during pregnancy and parturition in the sow. Pain, 1997. 72(1-2): 153-9.
99. Medina, V.M., Wang, L., and Gintzler, A.R. Spinal cord dynorphin: positive region-specific modulation during pregnancy and parturition. Brain Research, 1993. 623(1): 41-6.
100. Fassoulaki, A., Gatzou, V., Petropoulos, G., and Siafaka, I. Spread of subarachnoid block, intraoperative local anaesthetic requirements and postoperative analgesic requirements in Caesarean section and total abdominal hysterectomy. Br J Anaesth, 2004. 93(5): 678-82.
101. Dabo, F., Nyberg, F., Qin, Z., Sundstrom-Poromaa, I., and Akerud, H. Plasma levels of beta-endorphin during pregnancy and use of labor analgesia. Reprod Sci, 2010. 17(8): 742-7.
102. Varrassi, G., Bazzano, C., and Edwards, W.T. Effects of physical activity on maternal plasma beta-endorphin levels and perception of labor pain. Am J Obstet Gynecol, 1989. 160(3): 707-12.
103. Giesbrecht, G.F., Letourneau, N., Campbell, T., and Kaplan, B.J. Affective experience in ecologically relevant contexts is dynamic and not progressively attenuated during pregnancy. Arch Womens Ment Health, 2012. 15(6): 481-485.
104. Paoletti, A.M., Romagnino, S., Contu, R., et al. Observational study on the stability of the psychological status during normal pregnancy and increased blood levels of neuroactive steroids with GABA-A receptor agonist activity. Psychoneuroendocrinology, 2006. 31(4): 485-92.
105. Glynn, L.M., Wadhwa, P.D., Dunkel-Schetter, C., Chicz-Demet, A., and Sandman, C.A. When stress happens matters: effects of earthquake timing on stress responsivity in pregnancy. Am J Obstet Gynecol, 2001. 184(4): 637-42.
106. Entringer, S., Buss, C., Shirtcliff, E.A., Cammack, A.L., Yim, I.S., Chicz-DeMet, A., Sandman, C.A., and Wadhwa, P.D. Attenuation of maternal psychophysiological stress responses and the maternal cortisol awakening response over the course of human pregnancy. Stress, 2010. 13(3): 258-68.
107. Christensen, H., Leach, L.S., and Mackinnon, A. Cognition in pregnancy and motherhood: prospective cohort study. Br J Psychiatry, 2010. 196(2): 126-32.
108. Christensen, H., Poyser, C., Pollitt, P., and Cubis, J. Pregnancy may confer a selective cognitive advantage Journal of reproductive and infant psychology, 1999. 17: 7-25.
109. Janes, C., Casey, P., Huntsdale, C., and Angus, G. Memory in pregnancy. I: Subjective experiences and objective assessment of implicit, explicit and working memory in primigravid and primiparous women. J Psychosom Obstet Gynaecol, 1999. 20(2): 80-7.
110. Macbeth, A.H. and Luine, V.N. Changes in anxiety and cognition due to reproductive experience: a review of data from rodent and human mothers. Neurosci Biobehav Rev, 2010. 34(3): 452-67.
111. Kinsley, C.H., Madonia, L., Gifford, G.W., et al. Motherhood improves learning and memory. Nature, 1999. 402(6758): 137-8.
112. Lemaire, V., Billard, J.M., Dutar, P., George, O., Piazza, P.V., Epelbaum, J., Le Moal, M., and Mayo, W. Motherhood-induced memory improvement persists across lifespan in rats but is abolished by a gestational stress. Eur J Neurosci, 2006. 23(12): 3368-74.
113. Vesga-Lopez, O., Blanco, C., Keyes, K., Olfson, M., Grant, B.F., and Hasin, D.S. Psychiatric disorders in pregnant and postpartum women in the United States. Arch Gen Psychiatry, 2008. 65(7): 805-15.

63
114. Grant, K.A., McMahon, C., and Austin, M.P. Maternal anxiety during the transition to parenthood: a prospective study. J Affect Disord, 2008. 108(1-2): 101-11.
115. Evans, J., Heron, J., Francomb, H., Oke, S., and Golding, J. Cohort study of depressed mood during pregnancy and after childbirth. BMJ, 2001. 323(7307): 257-60.
116. O'Hara, M.W. and Swain, A.M. Rates and risk of postpartum depression - A meta-analysis. International Review of Psychiatry, 1996. 8(1): 37-54.
117. Andersson, L., Sundstrom-Poromaa, I., Bixo, M., Wulff, M., Bondestam, K., and aStrom, M. Point prevalence of psychiatric disorders during the second trimester of pregnancy: a population-based study. Am J Obstet Gynecol, 2003. 189(1): 148-54.
118. Andersson, L., Sundstrom-Poromaa, I., Wulff, M., Astrom, M., and Bixo, M. Depression and anxiety during pregnancy and six months postpartum: a follow-up study. Acta Obstet Gynecol Scand., 2006. 85(8): 937-44.
119. Milgrom, J., Gemmill, A.W., Bilszta, J.L., et al. Antenatal risk factors for postnatal depression: a large prospective study. J Affect Disord, 2008. 108(1-2): 147-57.
120. Heron, J., O'Connor, T.G., Evans, J., Golding, J., and Glover, V. The course of anxiety and depression through pregnancy and the postpartum in a community sample. J Affect Disord, 2004. 80(1): 65-73.
121. Josefsson, A., Berg, G., Nordin, C., and Sydsjo, G. Prevalence of depressive symptoms in late pregnancy and postpartum. Acta Obstet Gynecol Scand., 2001. 80(3): 251-5.
122. American Psychiatric Association. and American Psychiatric Association. Task Force on DSM-IV., Diagnostic and statistical manual of mental disorders : DSM-IV. 4th ed. 1994, Washington, DC: American Psychiatric Association. xxvii, 886 p.
123. Altemus, M., Neeb, C.C., Davis, A., Occhiogrosso, M., Nguyen, T., and Bleiberg, K.L. Phenotypic differences between pregnancy-onset and postpartum-onset major depressive disorder. J Clin Psychiatry, 2012. 73(12): e1485-91.
124. Ross, L.E. and McLean, L.M. Anxiety disorders during pregnancy and the postpartum period: A systematic review. J Clin Psychiatry., 2006. 67(8): 1285-98.
125. Andersson, L., Sundstrom-Poromaa, I., Wulff, M., Astrom, M., and Bixo, M. Implications of antenatal depression and anxiety for obstetric outcome. Obstet Gynecol, 2004. 104(3): 467-76.
126. Peeters, F., Nicholson, N.A., and Berkhof, J. Cortisol responses to daily events in major depressive disorder. Psychosom Med, 2003. 65(5): 836-41.
127. Vrshek-Schallhorn, S., Doane, L.D., Mineka, S., Zinbarg, R.E., Craske, M.G., and Adam, E.K. The cortisol awakening response predicts major depression: predictive stability over a 4-year follow-up and effect of depression history. Psychol Med, 2013. 43(3): 483-93.
128. Vreeburg, S.A., Hoogendijk, W.J., van Pelt, J., et al. Major depressive disorder and hypothalamic-pituitary-adrenal axis activity: results from a large cohort study. Arch Gen Psychiatry, 2009. 66(6): 617-26.
129. Knorr, U., Vinberg, M., Kessing, L.V., and Wetterslev, J. Salivary cortisol in depressed patients versus control persons: a systematic review and meta-analysis. Psychoneuroendocrinology, 2010. 35(9): 1275-86.

64
130. Steudte, S., Stalder, T., Dettenborn, L., Klumbies, E., Foley, P., Beesdo-Baum, K., and Kirschbaum, C. Decreased hair cortisol concentrations in generalised anxiety disorder. Psychiatry Res, 2011. 186(2-3): 310-4.
131. Vreeburg, S.A., Zitman, F.G., van Pelt, J., et al. Salivary cortisol levels in persons with and without different anxiety disorders. Psychosom Med, 2010. 72(4): 340-7.
132. Yim, I.S., Glynn, L.M., Schetter, C.D., Hobel, C.J., Chicz-Demet, A., and Sandman, C.A. Prenatal beta-endorphin as an early predictor of postpartum depressive symptoms in euthymic women. J Affect Disord, 2010. 125(1-3): 128-33.
133. Veith, R.C., Lewis, N., Linares, O.A., et al. Sympathetic nervous system activity in major depression. Basal and desipramine-induced alterations in plasma norepinephrine kinetics. Arch Gen Psychiatry, 1994. 51(5): 411-22.
134. Kemp, A.H., Quintana, D.S., Gray, M.A., Felmingham, K.L., Brown, K., and Gatt, J.M. Impact of depression and antidepressant treatment on heart rate variability: a review and meta-analysis. Biol Psychiatry, 2010. 67(11): 1067-74.
135. Berntson, G.G., Sarter, M., and Cacioppo, J.T. Anxiety and cardiovascular reactivity: the basal forebrain cholinergic link. Behav Brain Res, 1998. 94(2): 225-48.
136. Schulze, T.G., Muller, D.J., Krauss, H., et al. Association between a functional polymorphism in the monoamine oxidase A gene promoter and major depressive disorder. Am J Med Genet, 2000. 96(6): 801-3.
137. Kaufman, J., Yang, B.Z., Douglas-Palumberi, H., Grasso, D., Lipschitz, D., Houshyar, S., Krystal, J.H., and Gelernter, J. Brain-derived neurotrophic factor-5-HTTLPR gene interactions and environmental modifiers of depression in children. Biol Psychiatry, 2006. 59(8): 673-80.
138. Caspi, A., Sugden, K., Moffitt, T.E., et al. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science, 2003. 301(5631): 386-9.
139. Dekker, M.J., Tiemeier, H., Luijendijk, H.J., et al. The effect of common genetic variation in 11beta-hydroxysteroid dehydrogenase type 1 on hypothalamic-pituitary-adrenal axis activity and incident depression. J Clin Endocrinol Metab, 2011. 97(2): E233-7.
140. Ahrens, T., Deuschle, M., Krumm, B., van der Pompe, G., den Boer, J.A., and Lederbogen, F. Pituitary-adrenal and sympathetic nervous system responses to stress in women remitted from recurrent major depression. Psychosom Med, 2008. 70(4): 461-7.
141. Girdler, S.S., Lindgren, M., Porcu, P., Rubinow, D.R., Johnson, J.L., and Morrow, A.L. A history of depression in women is associated with an altered GABAergic neuroactive steroid profile. Psychoneuroendocrinology, 2012. 37(4): 543-53.
142. Dawson, M.E., Schell, A.M., and Filion, D.L., The electrodermal system, in Handbook of Psychophysiology, Cacioppo JT, T.L., Berntson GG, Editor. 2007, Cambridge University Press. p. 159-181.
143. Dube, A.A., Duquette, M., Roy, M., Lepore, F., Duncan, G., and Rainville, P. Brain activity associated with the electrodermal reactivity to acute heat pain. Neuroimage, 2009. 45(1): 169-80.
144. MacLaren, V.V. A quantitative review of the guilty knowledge test. J Appl Psychol, 2001. 86(4): 674-83.

65
145. Ledowski, T., Bromilow, J., Paech, M.J., Storm, H., Hacking, R., and Schug, S.A. Monitoring of skin conductance to assess postoperative pain intensity. Br J Anaesth, 2006. 97(6): 862-5.
146. Choo, E.K., Magruder, W., Montgomery, C.J., Lim, J., Brant, R., and Ansermino, J.M. Skin conductance fluctuations correlate poorly with postoperative self-report pain measures in school-aged children. Anesthesiology, 2010. 113(1): 175-82.
147. Hullett, B., Chambers, N., Preuss, J., Zamudio, I., Lange, J., Pascoe, E., and Ledowski, T. Monitoring electrical skin conductance: a tool for the assessment of postoperative pain in children? Anesthesiology, 2009. 111(3): 513-7.
148. Dawson ME, S., AM, Filion DL, The electrodermal system, in Handbook of Psychophysiology, Cacioppo JT, T.L., Berntson GG, Editor. 2000, Cambridge University Press. p. 200-221.
149. Wolf, S. and Hardy, J.D. Studies on Pain. Observations on Pain Due to Local Cooling and on Factors Involved in the "Cold Pressor" Effect. J Clin Invest, 1941. 20(5): 521-33.
150. Greenwood, J.P., Stoker, J.B., Walker, J.J., and Mary, D.A. Sympathetic nerve discharge in normal pregnancy and pregnancy-induced hypertension. J Hypertens, 1998. 16(5): 617-24.
151. Kammerer, M., Adams, D., Castelberg Bv, B., and Glover, V. Pregnant women become insensitive to cold stress. BMC Pregnancy Childbirth, 2002. 2(1): 8.
152. Grewen, K.M. and Light, K.C. Plasma oxytocin is related to lower cardiovascular and sympathetic reactivity to stress. Biol Psychol, 2011. 87(3): 340-9.
153. Davis, M., Falls, W.A., Campeau, S., and Kim, M. Fear-potentiated startle: a neural and pharmacological analysis. Behav Brain Res., 1993. 58(1-2): 175-98.
154. Davis, M. and Shi, C. The extended amygdala: are the central nucleus of the amygdala and the bed nucleus of the stria terminalis differentially involved in fear versus anxiety? Ann N Y Acad Sci., 1999. 877: 281-91.
155. Grillon, C., Ameli, R., Goddard, A., Woods, S.W., and Davis, M. Baseline and fear-potentiated startle in panic disorder patients. Biol Psychiatry., 1994. 35(7): 431-9.
156. Pissiota, A., Frans, O., Fredrikson, M., Langstrom, B., and Flaten, M.A. The human startle reflex and pons activation: a regional cerebral blood flow study. Eur J Neurosci., 2002. 15(2): 395-8.
157. Grillon, C. and Baas, J. A review of the modulation of the startle reflex by affective states and its application in psychiatry. Clin Neurophysiol., 2003. 114(9): 1557-79.
158. Hamm, A.O., Cuthbert, B.N., Globisch, J., and Vaitl, D. Fear and the startle reflex: blink modulation and autonomic response patterns in animal and mutilation fearful subjects. Psychophysiology., 1997. 34(1): 97-107.
159. Pissiota, A., Frans, O., Michelgard, A., Appel, L., Langstrom, B., Flaten, M.A., and Fredrikson, M. Amygdala and anterior cingulate cortex activation during affective startle modulation: a PET study of fear. Eur J Neurosci, 2003. 18(5): 1325-31.
160. Hoenig, K., Hochrein, A., Quednow, B.B., Maier, W., and Wagner, M. Impaired prepulse inhibition of acoustic startle in obsessive-compulsive disorder. Biol Psychiatry., 2005. 57(10): 1153-8.
161. Kumari, V., Kaviani, H., Raven, P.W., Gray, J.A., and Checkley, S.A. Enhanced startle reactions to acoustic stimuli in patients with obsessive-compulsive disorder. Am J Psychiatry., 2001. 158(1): 134-6.

66
162. Vrana, S.R., Spence, E.L., and Lang, P.J. The startle probe response: a new measure of emotion? J Abnorm Psychol, 1988. 97(4): 487-91.
163. Sabatinelli, D., Bradley, M.M., and Lang, P.J. Affective startle modulation in anticipation and perception. Psychophysiology, 2001. 38(4): 719-22.
164. Dichter, G.S., Tomarken, A.J., and Baucom, B.R. Startle modulation before, during and after exposure to emotional stimuli. Int J Psychophysiol, 2002. 43(2): 191-6.
165. Ray, W.J., Molnar, C., Aikins, D., Yamasaki, A., Newman, M.G., Castonguay, L., and Borkovec, T.D. Startle response in generalised anxiety disorder. Depress Anxiety, 2009. 26(2): 147-54.
166. Dichter, G.S. and Tomarken, A.J. The chronometry of affective startle modulation in unipolar depression. J Abnorm Psychol, 2008. 117(1): 1-15.
167. Mneimne, M., McDermut, W., and Powers, A.S. Affective ratings and startle modulation in people with nonclinical depression. Emotion, 2008. 8(4): 552-9.
168. Harmer, C.J., Shelley, N.C., Cowen, P.J., and Goodwin, G.M. Increased positive versus negative affective perception and memory in healthy volunteers following selective serotonin and norepinephrine reuptake inhibition. Am J Psychiatry, 2004. 161(7): 1256-63.
169. Norra, C., Becker, S., Herpertz, S.C., and Kunert, H.J. Effects of experimental acute tryptophan depletion on acoustic startle response in females. Eur Arch Psychiatry Clin Neurosci, 2008. 258(1): 1-9.
170. Buchanan, T.W., Brechtel, A., Sollers, J.J., and Lovallo, W.R. Exogenous cortisol exerts effects on the startle reflex independent of emotional modulation. Pharmacol Biochem Behav, 2001. 68(2): 203-10.
171. Koch, M. The neurobiology of startle. Prog Neurobiol., 1999. 59(2): 107-28. 172. Kask, K., Backstrom, T., Gulinello, M., and Sundstrom-Poromaa, I. Lower
levels of prepulse inhibition of startle response in pregnant women compared to postpartum women. Psychoneuroendocrinology., 2008. 33(1): 100-7.
173. Paul, S.M. and Purdy, R.H. Neuroactive steroids. Faseb J, 1992. 6(6): 2311-22. 174. Majewska, M.D., Harrison, N.L., Schwartz, R.D., Barker, J.L., and Paul, S.M.
Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science, 1986. 232(4753): 1004-7.
175. Puia, G., Mienville, J.M., Matsumoto, K., Takahata, H., Watanabe, H., Costa, E., and Guidotti, A. On the putative physiological role of allopregnanolone on GABA(A) receptor function. Neuropharmacology, 2003. 44(1): 49-55.
176. Genazzani, A.D., Luisi, M., Malavasi, B., et al. Pulsatile secretory characteristics of allopregnanolone, a neuroactive steroid, during the menstrual cycle and in amenorrheic subjects. Eur J Endocrinol, 2002. 146(3): 347-56.
177. Ottander, U., Poromaa, I.S., Bjurulf, E., Skytt, A., Backstrom, T., and Olofsson, J.I. Allopregnanolone and pregnanolone are produced by the human corpus luteum. Mol Cell Endocrinol, 2005. 239(1-2): 37-44.
178. Morgan, M.L., Rapkin, A.J., Biggio, G., Serra, M., Pisu, M.G., and Rasgon, N. Neuroactive steroids after estrogen exposure in depressed postmenopausal women treated with sertraline and asymptomatic postmenopausal women. Arch Womens Ment Health, 2010. 13(1): 91-8.
179. Romeo, E., Strohle, A., Spalletta, G., di Michele, F., Hermann, B., Holsboer, F., Pasini, A., and Rupprecht, R. Effects of antidepressant treatment on neuroactive steroids in major depression. Am J Psychiatry, 1998. 155(7): 910-3.
180. Uzunova, V., Sheline, Y., Davis, J.M., Rasmusson, A., Uzunov, D.P., Costa, E., and Guidotti, A. Increase in the cerebrospinal fluid content of neurosteroids

67
in patients with unipolar major depression who are receiving fluoxetine or fluvoxamine. Proc Natl Acad Sci U S A, 1998. 95(6): 3239-44.
181. Hardoy, M.C., Serra, M., Carta, M.G., Contu, P., Pisu, M.G., and Biggio, G. Increased neuroactive steroid concentrations in women with bipolar disorder or major depressive disorder. J Clin Psychopharmacol, 2006. 26(4): 379-84.
182. Barbaccia, M.L., Lello, S., Sidiropoulou, T., et al. Plasma 5alpha-androstane-3alpha,17betadiol, an endogenous steroid that positively modulates GABA(A) receptor function, and anxiety: a study in menopausal women. Psychoneuroendocrinology, 2000. 25(7): 659-75.
183. Nappi, R.E., Petraglia, F., Luisi, S., Polatti, F., Farina, C., and Genazzani, A.R. Serum allopregnanolone in women with postpartum "blues". Obstet Gynecol, 2001. 97(1): 77-80.
184. Andreen, L., Sundstrom-Poromaa, I., Bixo, M., Nyberg, S., and Backstrom, T. Allopregnanolone concentration and mood--a bimodal association in postmenopausal women treated with oral progesterone. Psychopharmacology (Berl). 2006. 187(2): 209-21.
185. Dombroski, R.A., Casey, M.L., and MacDonald, P.C. 5-Alpha-dihydroprogesterone formation in human placenta from 5alpha-pregnan-3beta/alpha-ol-20-ones and 5-pregnan-3beta-yl-20-one sulfate. J Steroid Biochem Mol Biol, 1997. 63(1-3): 155-63.
186. Parizek, A., Hill, M., Kancheva, R., et al. Neuroactive pregnanolone isomers during pregnancy. J Clin Endocrinol Metab, 2005. 90(1): 395-403.
187. Turkmen, S., Backstrom, T., Wahlstrom, G., Andreen, L., and Johansson, I.M. Tolerance to allopregnanolone with focus on the GABA-A receptor. Br J Pharmacol, 2011. 162(2): 311-27.
188. Timby, E., Balgard, M., Nyberg, S., et al. Pharmacokinetic and behavioral effects of allopregnanolone in healthy women. Psychopharmacology (Berl), 2006. 186(3): 414-24.
189. Kancheva, R., Hill, M., Novak, Z., Chrastina, J., Velikova, M., Kancheva, L., Riha, I., and Starka, L. Peripheral neuroactive steroids may be as good as the steroids in the cerebrospinal fluid for the diagnostics of CNS disturbances. J Steroid Biochem Mol Biol, 2010. 119(1-2): 35-44.
190. Bixo, M., Andersson, A., Winblad, B., Purdy, R.H., and Backstrom, T. Progesterone, 5alpha-pregnane-3,20-dione and 3alpha-hydroxy-5alpha-pregnane-20-one in specific regions of the human female brain in different endocrine states. Brain Res, 1997. 764(1-2): 173-8.
191. Evans, J., Sun, Y., McGregor, A., and Connor, B. Allopregnanolone regulates neurogenesis and depressive/anxiety-like behaviour in a social isolation rodent model of chronic stress. Neuropharmacology, 2012. 63(8): 1315-26.
192. Pearson Murphy, B.E., Steinberg, S.I., Hu, F.Y., and Allison, C.M. Neuroactive ring A-reduced metabolites of progesterone in human plasma during pregnancy: elevated levels of 5 alpha-dihydroprogesterone in depressed patients during the latter half of pregnancy. J Clin Endocrinol Metab, 2001. 86(12): 5981-7.
193. Wilhelm, I., Born, J., Kudielka, B.M., Schlotz, W., and Wust, S. Is the cortisol awakening rise a response to awakening? Psychoneuroendocrinology, 2007. 32(4): 358-66.
194. Clow, A., Thorn, L., Evans, P., and Hucklebridge, F. The awakening cortisol response: methodological issues and significance. Stress, 2004. 7(1): 29-37.
195. Pruessner, J.C., Wolf, O.T., Hellhammer, D.H., Buske-Kirschbaum, A., von Auer, K., Jobst, S., Kaspers, F., and Kirschbaum, C. Free cortisol levels after

68
awakening: a reliable biological marker for the assessment of adrenocortical activity. Life Sci, 1997. 61(26): 2539-49.
196. Clow, A., Hucklebridge, F., Stalder, T., Evans, P., and Thorn, L. The cortisol awakening response: more than a measure of HPA axis function. Neurosci Biobehav Rev, 2010. 35(1): 97-103.
197. Vreeburg, S.A., Kruijtzer, B.P., van Pelt, J., et al. Associations between sociodemographic, sampling and health factors and various salivary cortisol indicators in a large sample without psychopathology. Psychoneuroendocrinology, 2009. 34(8): 1109-20.
198. de Weerth, C. and Buitelaar, J.K. Cortisol awakening response in pregnant women. Psychoneuroendocrinology, 2005. 30(9): 902-7.
199. Buss, C., Entringer, S., Reyes, J.F., Chicz-DeMet, A., Sandman, C.A., Waffarn, F., and Wadhwa, P.D. The maternal cortisol awakening response in human pregnancy is associated with the length of gestation. Am J Obstet Gynecol, 2009. 201(4): 398 e1-8.
200. Giesbrecht, G.F., Campbell, T., Letourneau, N., Kooistra, L., and Kaplan, B. Psychological distress and salivary cortisol covary within persons during pregnancy. Psychoneuroendocrinology, 2012. 37(2): 270-9.
201. Shea, A.K., Streiner, D.L., Fleming, A., Kamath, M.V., Broad, K., and Steiner, M. The effect of depression, anxiety and early life trauma on the cortisol awakening response during pregnancy: preliminary results. Psychoneuroendocrinology, 2007. 32(8-10): 1013-20.
202. Svanborg, P. and Asberg, M. A new self-rating scale for depression and anxiety states based on the Comprehensive Psychopathological Rating Scale. Acta Psychiatr Scand, 1994. 89(1): 21-8.
203. Montgomery, S.A. and Asberg, M. A new depression scale designed to be sensitive to change. Br J Psychiatry., 1979. 134: 382-9.
204. Cunningham, J.L., Wernroth, L., von Knorring, L., Berglund, L., and Ekselius, L. Agreement between physicians' and patients' ratings on the Montgomery-Asberg Depression Rating Scale. J Affect Disord, 2011. 135(1-3): 148-53.
205. Cox, J.L., Holden, J.M., and Sagovsky, R. Detection of postnatal depression. Development of the 10-item Edinburgh Postnatal Depression Scale. Br J Psychiatry., 1987. 150: 782-6.
206. Rubertsson, C., Borjesson, K., Berglund, A., Josefsson, A., and Sydsjo, G. The Swedish validation of Edinburgh Postnatal Depression Scale (EPDS) during pregnancy. Nord J Psychiatry, 2011. 65(6): 414-8.
207. Wickberg, B. and Hwang, C.P. The Edinburgh Postnatal Depression Scale: validation on a Swedish community sample. Acta Psychiatr Scand, 1996. 94(3): 181-4.
208. Spielberger, C.D., State-trait Anxiety Inventory for Adults. Sampler Set, Manual Set, Scoring Key 1983, Palo Alto: Consulting Psychologists Press Inc. .
209. Sheehan, D.V., Lecrubier, Y., Sheehan, K.H., et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry, 1998. 59 Suppl 20: 22-33;quiz 34-57.
210. Lang, P.J., Bradley, M.M., and Cuthbert, B.N. International affective picture system (IAPS): Affective ratings of pictures and instruction manual. Technical Report A-6. University of Florida, Gainesville, FL., 2005.
211. Chen, Y., Holzman, C., Chung, H., Senagore, P., Talge, N.M., and Siler-Khodr, T. Levels of maternal serum corticotropin-releasing hormone (CRH) at midpregnancy in relation to maternal characteristics. Psychoneuroendocrinology, 2010. 35(6): 820-32.

69
212. Campbell, J. and Ehlert, U. Acute psychosocial stress: does the emotional stress response correspond with physiological responses? Psychoneuroendocrinology, 2012. 37(8): 1111-34.
213. Young, E.A., Abelson, J., and Lightman, S.L. Cortisol pulsatility and its role in stress regulation and health. Front Neuroendocrinol, 2004. 25(2): 69-76.
214. Baas, J.M., Mol, N., Kenemans, J.L., Prinssen, E.P., Niklson, I., Xia-Chen, C., Broeyer, F., and van Gerven, J. Validating a human model for anxiety using startle potentiated by cue and context: the effects of alprazolam, pregabalin, and diphenhydramine. Psychopharmacology (Berl), 2009. 205(1): 73-84.
215. Bannbers, E., Kask, K., Wikstrom, J., Risbrough, V., and Sundstrom Poromaa, I. Patients with premenstrual dysphoric disorder have increased startle modulation during anticipation in the late luteal phase period in comparison to control subjects. Psychoneuroendocrinology, 2011. 36(8): 1184-92.
216. Nisell, H., Hjemdahl, P., Linde, B., and Lunell, N.O. Sympatho-adrenal and cardiovascular reactivity in pregnancy-induced hypertension. I. Responses to isometric exercise and a cold pressor test. Br J Obstet Gynaecol, 1985. 92(7): 722-31.
217. Baker, D.G., West, S.A., Orth, D.N., et al. Cerebrospinal fluid and plasma beta-endorphin in combat veterans with post-traumatic stress disorder. Psychoneuroendocrinology, 1997. 22(7): 517-29.
218. Stein, C. and Lang, L.J. Peripheral mechanisms of opioid analgesia. Curr Opin Pharmacol, 2009. 9(1): 3-8.
219. Rupp, H.A., James, T.W., Ketterson, E.D., Sengelaub, D.R., Ditzen, B., and Heiman, J.R. Lower sexual interest in postpartum women: relationship to amygdala activation and intranasal oxytocin. Horm Behav, 2013. 63(1): 114-21.
220. van Wingen, G.A., van Broekhoven, F., Verkes, R.J., Petersson, K.M., Backstrom, T., Buitelaar, J.K., and Fernandez, G. Progesterone selectively increases amygdala reactivity in women. Mol Psychiatry., 2008. 13(3): 325-33.
221. Wirth, M.M., Meier, E.A., Fredrickson, B.L., and Schultheiss, O.C. Relationship between salivary cortisol and progesterone levels in humans. Biol Psychol, 2007. 74(1): 104-7.
222. Toufexis Lactation-induced reduction in rats' acoustic startle is associated with changes in noradrenergic neurotransmission. Behav Neurosci. 1999 Feb;113(1):176-84., 1999.
223. Byrnes, E.M., Bridges, R.S., Scanlan, V.F., Babb, J.A., and Byrnes, J.J. Sensorimotor gating and dopamine function in postpartum rats. Neuropsychopharmacology., 2007. 32(5): 1021-31.
224. Gulinello, M., Orman, R., and Smith, S.S. Sex differences in anxiety, sensorimotor gating and expression of the alpha4 subunit of the GABAA receptor in the amygdala after progesterone withdrawal. Eur J Neurosci, 2003. 17(3): 641-8.
225. Smith, S.S., Shen, H., Gong, Q.H., and Zhou, X. Neurosteroid regulation of GABA(A) receptors: Focus on the alpha4 and delta subunits. Pharmacol Ther, 2007. 116(1): 58-76.
226. Zhu, D., Birzniece, V., Backstrom, T., and Wahlstrom, G. Dynamic aspects of acute tolerance to allopregnanolone evaluated using anaesthesia threshold in male rats. Br J Anaesth, 2004. 93(4): 560-7.
227. Patchev, V.K., Hassan, A.H., Holsboer, D.F., and Almeida, O.F. The neurosteroid tetrahydroprogesterone attenuates the endocrine response to stress and exerts glucocorticoid-like effects on vasopressin gene transcription in the rat hypothalamus. Neuropsychopharmacology, 1996. 15(6): 533-40.

70
228. Bitran, D., Hilvers, R.J., and Kellogg, C.K. Anxiolytic effects of 3 alpha-hydroxy-5 alpha[beta]-pregnan-20-one: endogenous metabolites of progesterone that are active at the GABAA receptor. Brain Res, 1991. 561(1): 157-61.
229. de Brito Faturi, C., Teixeira-Silva, F., and Leite, J.R. The anxiolytic effect of pregnancy in rats is reversed by finasteride. Pharmacol Biochem Behav, 2006. 85(3): 569-74.
230. Waffarn, F. and Davis, E.P. Effects of antenatal corticosteroids on the hypothalamic-pituitary-adrenocortical axis of the fetus and newborn: experimental findings and clinical considerations. Am J Obstet Gynecol, 2012. 207(6): 446-54.
231. Mahon, P.B., Zandi, P.P., Potash, J.B., Nestadt, G., and Wand, G.S. Genetic association of FKBP5 and CRHR1 with cortisol response to acute psychosocial stress in healthy adults. Psychopharmacology (Berl), 2013. 2012 Dec 30. [Epub ahead of print].
232. Lamers, F., Vogelzangs, N., Merikangas, K.R., de Jonge, P., Beekman, A.T., and Penninx, B.W. Evidence for a differential role of HPA-axis function, inflammation and metabolic syndrome in melancholic versus atypical depression. Mol Psychiatry, 2012. Oct 23. doi: 10.1038/mp.2012.144. [Epub ahead of print].
233. Christian, L.M. Physiological reactivity to psychological stress in human pregnancy: current knowledge and future directions. Prog Neurobiol, 2012. 99(2): 106-16.
234. Lovallo, W.R. Do low levels of stress reactivity signal poor states of health? Biol Psychol, 2011. 86(2): 121-8.
235. Poyhonen-Alho, M., Viitasalo, M., Nicholls, M.G., Lindstrom, B.M., Vaananen, H., and Kaaja, R. Imbalance of the autonomic nervous system at night in women with gestational diabetes. Diabet Med, 2010. 27(9): 988-94.
236. Davis, E.P. and Sandman, C.A. Prenatal psychobiological predictors of anxiety risk in preadolescent children. Psychoneuroendocrinology, 2012. 37(8): 1224-33.
237. O'Connor, T.G., Ben-Shlomo, Y., Heron, J., Golding, J., Adams, D., and Glover, V. Prenatal anxiety predicts individual differences in cortisol in pre-adolescent children. Biol Psychiatry, 2005. 58(3): 211-7.
238. Voegtline, K.M., Costigan, K.A., Kivlighan, K.T., Laudenslager, M.L., Henderson, J.L., and Dipietro, J.A. Concurrent levels of maternal salivary cortisol are unrelated to self-reported psychological measures in low-risk pregnant women. Arch Womens Ment Health, 2013. 16(2): 101-8.
239. Salacz, P., Csukly, G., Haller, J., and Valent, S. Association between subjective feelings of distress, plasma cortisol, anxiety, and depression in pregnant women. Eur J Obstet Gynecol Reprod Biol, 2012.
240. O'Donnell, K.J., Bugge Jensen, A., Freeman, L., Khalife, N., O'Connor, T.G., and Glover, V. Maternal prenatal anxiety and downregulation of placental 11beta-HSD2. Psychoneuroendocrinology, 2012. 37(6): 818-26.


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 887
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine.
Distribution: publications.uu.seurn:nbn:se:uu:diva-197441
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2013


















