
Physiological, morphological and growth responses to rhizosphere hypoxia by seedlings
14
Original article Physiological, morphological and growth responses to rhizosphere hypoxia by seedlings of North American bottomland oaks ES Gardiner JD Hodges Forest and Wildlife Research Center, Mississippi State University, Mississippi State, MS 36762, USA (Received 2 November 1994; accepted 29 June 1995) Summary — Bottomland oak species of the southern United States are distributed along topograph- ical gradients in floodplains. Because differences in soil aeration are associated with these gradients, we tested the hypothesis that oak species will exhibit diverging resistances to rhizosphere hypoxia. Four species which occupy different sites in floodplains, Quercus lyrata, Q laurifolia, Q phellos, and Q nigra, were used in two experiments designed to examine seedling responses during establishment and late in the first growing season. In both experiments, hypoxia tolerance was evaluated through mea- surements of gas exchange, biomass accumulation, and shoot and root growth. Evidence of oaks differing in resistance to rhizosphere hypoxia is presented, and results are discussed in relation to species distribution in floodplains. rhizosphere hypoxia / Quercus lyrata / Quercus laurifolia / Quercus phellos / Quercus nigra / eco- physiology Résumé — Réponses physiologiques et morphologiques, et de croissance à l’hypoxie rhizo- sphérique de chênes des plaines inondables d’Amérique du Nord. Certaines espèces de chênes du sud des États-Unis sont distribuées le long de gradients topographiques dans les plaines inon- dables. Du fait des différences dans l’aération des sols associées à ces gradients, nous avons testé l’hypothèse que ces espèces présentent des résistances différentes à l’hypoxie rhizosphérique. Quatre espèces occupant différents sites des plaines inondables (Quercus lyrata, Q laurifolia, Q phellos, et Q nigra) ont été soumises à deux expérimentations pour évaluer les réponses des plantes au début et à la fin de la première saison. Dans les deux études, la tolérance à l’hypoxie a été estimée par la mesure d’échanges gazeux, l’accumulation de biomasse, et la croissance de la pousse et de la racine. Des dif- férences de résistance à l’hypoxie de la rhizosphère ont été mises en évidence et les conséquences sur la distribution des espèces dans les plaines inondables ont été discutées. rhizosphère / hypoxie / Quercus lyrata / Quercus laurifolia / Quercus phellos / Quercus nigra / écophysiologie * Current address: Southern Hardwoods Laboratory, USDA, Forest Service, PO Box 227, Stoneville, MS 38776, USA
Transcript of Physiological, morphological and growth responses to rhizosphere hypoxia by seedlings

Original article
Physiological, morphological and growth responsesto rhizosphere hypoxia by seedlingsof North American bottomland oaks
ES Gardiner JD Hodges
Forest and Wildlife Research Center, Mississippi State University, Mississippi State, MS 36762, USA
(Received 2 November 1994; accepted 29 June 1995)
Summary — Bottomland oak species of the southern United States are distributed along topograph-ical gradients in floodplains. Because differences in soil aeration are associated with these gradients,we tested the hypothesis that oak species will exhibit diverging resistances to rhizosphere hypoxia. Fourspecies which occupy different sites in floodplains, Quercus lyrata, Q laurifolia, Q phellos, and Q nigra,were used in two experiments designed to examine seedling responses during establishment andlate in the first growing season. In both experiments, hypoxia tolerance was evaluated through mea-surements of gas exchange, biomass accumulation, and shoot and root growth. Evidence of oaksdiffering in resistance to rhizosphere hypoxia is presented, and results are discussed in relation tospecies distribution in floodplains.
rhizosphere hypoxia / Quercus lyrata / Quercus laurifolia / Quercus phellos / Quercus nigra / eco-physiology
Résumé — Réponses physiologiques et morphologiques, et de croissance à l’hypoxie rhizo-sphérique de chênes des plaines inondables d’Amérique du Nord. Certaines espèces de chênesdu sud des États-Unis sont distribuées le long de gradients topographiques dans les plaines inon-dables. Du fait des différences dans l’aération des sols associées à ces gradients, nous avons testél’hypothèse que ces espèces présentent des résistances différentes à l’hypoxie rhizosphérique. Quatreespèces occupant différents sites des plaines inondables (Quercus lyrata, Q laurifolia, Q phellos, et Qnigra) ont été soumises à deux expérimentations pour évaluer les réponses des plantes au début et àla fin de la première saison. Dans les deux études, la tolérance à l’hypoxie a été estimée par la mesured’échanges gazeux, l’accumulation de biomasse, et la croissance de la pousse et de la racine. Des dif-férences de résistance à l’hypoxie de la rhizosphère ont été mises en évidence et les conséquencessur la distribution des espèces dans les plaines inondables ont été discutées.
rhizosphère / hypoxie / Quercus lyrata / Quercus laurifolia / Quercus phellos / Quercus nigra /écophysiologie
* Current address: Southern Hardwoods Laboratory, USDA, Forest Service, PO Box 227, Stoneville,MS 38776, USA

INTRODUCTION
Microsite environments can profoundly influ-ence the composition and dispersion ofplants in communities. Soil surface hetero-geneity, proximity to mature plants, leaf lit-ter depth, and nutrient distributions are afew of the microsite attributes that can affectseed germination, establishment, andgrowth of herbaceous and woody plants(Harper et al, 1965; Beatty, 1984; Hartgerinkand Bazzaz, 1984; Fowler, 1986; Molofskyand Augspurger, 1992). Because alluvialsites are characteristically diverse in
edaphic, hydrologic, and biologic structure,several microsite types, which could poten-tially influence seedling establishment andgrowth, can be identified in forested wet-land communities (Marks and Harcombe,1981; Huenneke and Sharitz, 1986, 1990).
Floodplains have little relief, but evenslight elevational changes can affect vege-tational cover (Tanner, 1986). Overcup oak(Quercus lyrata Walt), swamp laurel oak(Q laurifolia Michx), willow oak (Q phellosL), and water oak (Q nigra L) have beenobserved to occupy different topographicalmicrosites within floodplains of the south-ern United States. Q lyrata, a member ofthe subgenus Leucobalanus, grows on tem-poral sloughs or low flats in the floodplain(Hodges and Switzer, 1979; Tanner, 1986).These sites are frequently flooded, canremain saturated into the growing season,and exhibit poor internal soil aeration (Put-nam et al, 1960). Q lyrata has been classi-fied as the most flood-tolerant of the south-ern United States oaks (McKnight et al,1981). Q laurifolia is found on low flats nearthe periphery of temporal sloughs, and isamong the most flood-tolerant of oaks inthe subgenus Erythrobalanus (Putnam etal, 1960; Hodges and Switzer, 1979; McK-night et al, 1981). Q phellos, subgenus Ery-throbalanus, is typically found on low flats,but at a slightly higher elevation than Q lau-rifolia. It is classified as more flood-tolerant
than Q nigra (Putnam et al, 1960; Tanner,1986). Q nigra, the least flood-tolerant ofthese Erythrobalanus oaks, grows primarilyon high flats or low ridges which are theleast frequently flooded positions in thefloodplain (Hodges and Switzer, 1979; Tan-ner 1986). Soils on these sites are satu-rated for short periods and have the bestinternal aeration (Putnam et al, 1960).
Woody plants established on a floodplainmust survive periodic root inundation byfloodwater. Since oxygen is limiting in sat-urated soil (Ponnamperuma, 1984), terres-trial plants of wetlands must possess theability to cope or adjust to an anaerobic rhi-zosphere. The observed stratification ofoaks in bottomlands, and the apparentassociation of flood frequency, duration, andsoil aeration along the topographical gradi-ent has prompted this investigation of theecophysiological mechanisms influencingresistance to root hypoxia by floodplain treeregeneration in the genus Quercus. Thispaper reports on two experiments conductedto test the hypothesis that bottomland oakspecies exhibit different resistances to roothypoxia. The thought is that the specieswhich occupy the lowest sites, those siteswhich have aerobic soil for the shortest dura-tion in the growing season, will demonstratethe greatest resistance to root hypoxia. Thehypothesis was tested on seedlings duringthe establishment phase in Experiment 1,and it was tested on seedlings late in theirfirst growing season in Experiment 2.
MATERIALS AND METHODS
Experiment 1
Stratified Q lyrata, Q laurifolia, Q phellos, andQ nigra acorns were sown 2.5 cm deep in 164cm3 tubes containing sand. The tubes wereplaced in a germinator programmed for 8 h oflight at 30 °C and 16 h of darkness at 20 °C (Bon-ner and Vozzo, 1987). Sand was kept moist to

facilitate acorn germination. At epicotyl emer-gence, seedlings were transferred to a hydro-ponic network in a greenhouse where they wereestablished in a 0.1 strength Hoagland #2 nutrientsolution (pH = 5.5) (Jones, 1983).
The hydroponic network, built after a systemdescribed by Topa and McLeod (1986), consistedof two 208 L nutrient solution reservoirs, thirty18.9 L plastic pots, and appropriate plumbing.Each pot received non-circulating nutrient solutionso that 3.8 L of solution was replaced every 24 h.Nutrient solution in each pot was bubbled witheither O2 or N2 depending on treatment assign-ment (Topa and McLeod, 1986). N2 was used to
maintain dissolved oxygen (DO) concentrationat < 0.2 mg L-1 for pots designated to receivethe hypoxic rhizosphere treatment. O2 was usedto maintain solution DO concentration at
> 15 mg L-1 for pots receiving the normoxic rhi-zosphere treatment. O2 was used rather than airto ensure a homogeneously aerobic rhizosphere.
Four seedlings, one of each species, in thestage of epicotyl emergence were randomlyassigned to each of 30 pots. Plants were main-tained in nutrient solution for 54 days with half ofthe pots receiving hypoxic nutrient solution, andthe other half receiving normoxic nutrient solu-tion. At d6 of this experiment it was determinedthat seedlings receiving the hypoxic nutrient solu-tion would all soon die. Shoots were witheringand roots had not grown, so these seedlings weregiven a 24 day recovery period of normoxic nutri-ent solution. After leaves developed during therecovery period, seedlings once again receivedhypoxic nutrient solution for the remainder of theexperiment.
On each of 3 days near the end of the 54 dayexperiment, leaf gas exchange variables weremeasured on three seedlings of each speciesand treatment. On each seedling, one medianleaf in a lag-stage flush was selected for mea-surement (Hanson et al, 1986). A LCA-3 CO2analyzer (The Analytical Development Co Ltd,UK) was used to measure CO2 exchange ratesand determine intercellular CO2 on these leavesunder saturating light (≥ 800 μmol m-2 s-1 pho-tosynthetically active photon flux density (PPFD)(Gardiner and Hodges, unpublished). Supple-mental light was provided with a fan-cooled, highpressure sodium lamp when ambient light wasnot saturating. Transpiration and stomatal con-ductance for these same leaves were measuredwith a Li-1600 Steady State Porometer (Licor Inc,Lincoln, NE, USA). All gas exchange measure-
ments were taken randomly for species and treat-ment combinations between 1000 and 1200 hourson each sample day. At the end of the experi-ment, leaf samples for carbon isotope analysiswere secured from five randomly chosen pots ineach treatment. Dried leaf material was groundto pass #40 mesh, and analyzed for stable carbonisotope ratios at the Bioscience Laboratory, Uni-versity of Utah, USA. Leaf tissue was analyzedfor stable isotope ratios because we thought thisvariable would serve as an integrated index ofstomatal aperature during rhizosphere hypoxia.
After the 54 day experiment, all 120 seedlingswere harvested and dissected into leaves, stemsand roots. These dried biomass components wereused as indices of proportional biomass accu-mulation, leaf weight ratio (LWR) = leaf weight/total plant weight, stem weight ratio (SWR) =stem weight/total plant weight, root weight ratio(RWR) = root system weight/total plant weight,root/stem ratio (R/S ratio) = root systemweight/stem weight. Relative height and diametergrowth were calculated from initial and final stemheights and diameters.
Experiment 2
Stratified acorns from Q lyrata, Q laurifolia, Q phel-los and Q nigra were planted in 0.9 L containersof a 50% potting soil: 50% sand mixture (v:v).Containers were placed in a greenhouse whereseedlings were grown for about 4 months. Ran-domly selected lag-stage seedlings were removedfrom their original containers, soil was carefullywashed away from roots, and these seedlingswere immediately transferred to a hydroponic net-work. Four seedlings, one of each species, wererandomly assigned to one of 40 pots in the hydro-ponic network. The network and nutrient solutionwere the same as those described in Experiment1. After 2 weeks of seedling acclimation, 20 potswere randomly assigned a 35 day treatment ofhypoxic nutrient solution, and 20 pots were main-tained in normoxic nutrient solution as a control.
Stomatal conductance and transpiration weremeasured 3 days before the treatment and ondays 1-10, 15, 20, 25, 30, and 35 of the treat-ment. These variables were measured on one
fully expanded leaf from four seedlings in thesame morphological stage while a portable lampmaintained PPFD on the leaf between 400 and800 μmol m-2 s-1. Stomatal conductance and

transpiration were measured randomly for speciesand treatment combinations between 0900 and
1100 hours on each sample day. Diurnal stomatalconductance and transpiration measurementswere randomly taken for each treatment andspecies combination at 2 h intervals beginningat 0600 hours, and finishing at 2000 hours on 4days during the last week of the experiment. Oneach selected seedling, one fully expanded leafon a predetermined flush was measured underambient light.
Following the experiment, 15 seedlings in eachtreatment and species combination were har-vested and dissected into leaves, stems, androots. LWR, SWR, RWR, and R/S ratios werecalculated from the dried biomass components.Relative height and diameter growth were calcu-lated from stem heights and diameters measuredon days 1 and 35.
For both experiments, data were analyzedaccording to a split-plot design with StatisticalAnalysis System software (SAS Version 6.04,SAS Institute, Cary, NC, USA). DO level was thewhole plot and species were the split-plot. If thetreatment x species interaction term was signifi-cant at a = 0.05, treatment combination meanswere separated with a LSD computed for the dif-ference between two whole plot means at thesame or different levels of the split-plot mean(Petersen, 1985). If the treatment x species inter-action term was not significant, it was pooled intothe error term to test significance of the treatmenteffect.
RESULTS
Experiment 1
Net photosynthesis and transpiration werereduced by 78 and 86%, respectively, onplants established in hypoxic nutrient solu-tion (table I). Decline in net photosynthesisand transpiration can be attributed to par-tial stomatal closure, because stomatal con-ductance decreased 84% for plants estab-lished in hypoxic nutrient solution (table I). Inthis experiment, leaf gas exchange did notvary by species under rhizosphere hypoxia.The δ13C for plants established in hypoxic
nutrient solution averaged about 1‰ below
that of plants established in hypoxic nutri-ent solution (table I). Calculated ratios ofnet photosynthesis/intercellular CO2 were
greatest for all oak seedlings established innormoxic nutrient solution, and averagedabout 81% less for seedlings establishedin hypoxic nutrient solution (table I).
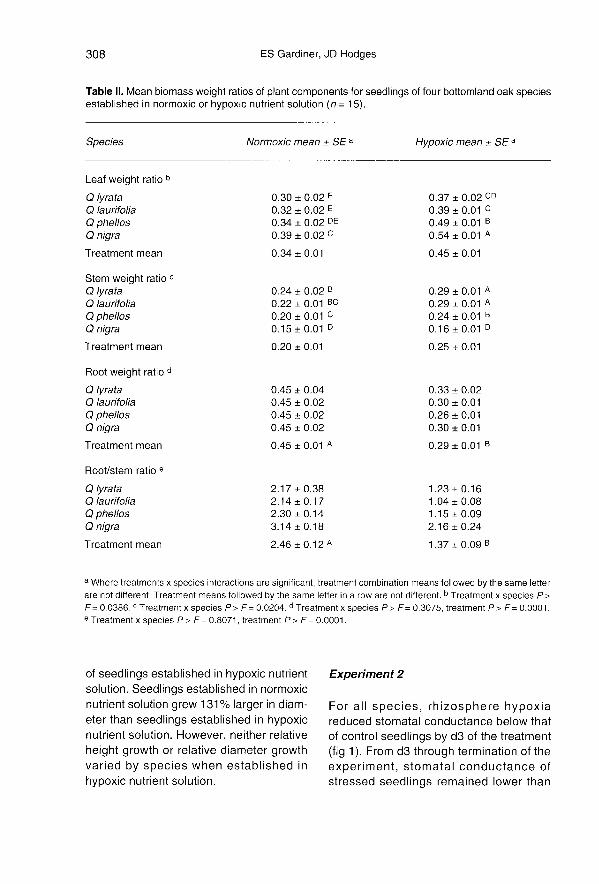
Total plant biomass of seedlings estab-lished in hypoxic nutrient solution was 40%less than that of seedlings established innormoxic nutrient solution. Biomass accu-mulation decreased 22% in leaves, 21 % instems, and 61 % in roots for all oak species.In addition to the depressed biomass accu-mulation, proportional biomass accumula-tion in plant components shifted for
seedlings established in hypoxic nutrientsolution (table II).
For all oaks, roots comprised about 38%of total plant biomass, but establishment inhypoxic nutrient solution reduced RWRs by36% (table II). Therefore, seedling estab-lishment in hypoxic nutrient solutionincreased proportional biomass accumula-tion in shoots, but relative change in SWRand LWR differed by species (table II).Q nigra showed an increase in LWR whenestablished in hypoxic nutrient solution, butthe other species showed an increase inLWR and SWR. When established in
hypoxic nutrient solution, Q nigra andQ phellos maintained proportionately moreleaf biomass than Q laurifolia and Q lyrata(table II). But Q lyrata and Q laurifolia main-tained proportionately more stem biomassthan Q phellos and Q nigra seedlings underrhizosphere hypoxia (table II). R/S ratios ofall species decreased about 44% whenestablished in hypoxic nutrient solution (tableII). Though total plant weight differedbetween rhizosphere treatments, R/S ratiowas weakly correlated with total plantbiomass (correlation coefficient = 0.24).
Relative height growth of seedlingsestablished in normoxic nutrient solutionwas 72% greater than relative height growth

of seedlings established in hypoxic nutrientsolution. Seedlings established in normoxicnutrient solution grew 131 % larger in diam-eter than seedlings established in hypoxicnutrient solution. However, neither relative
height growth or relative diameter growthvaried by species when established inhypoxic nutrient solution.
Experiment 2
For all species, rhizosphere hypoxiareduced stomatal conductance below that
of control seedlings by d3 of the treatment(fig 1). From d3 through termination of theexperiment, stomatal conductance ofstressed seedlings remained lower than
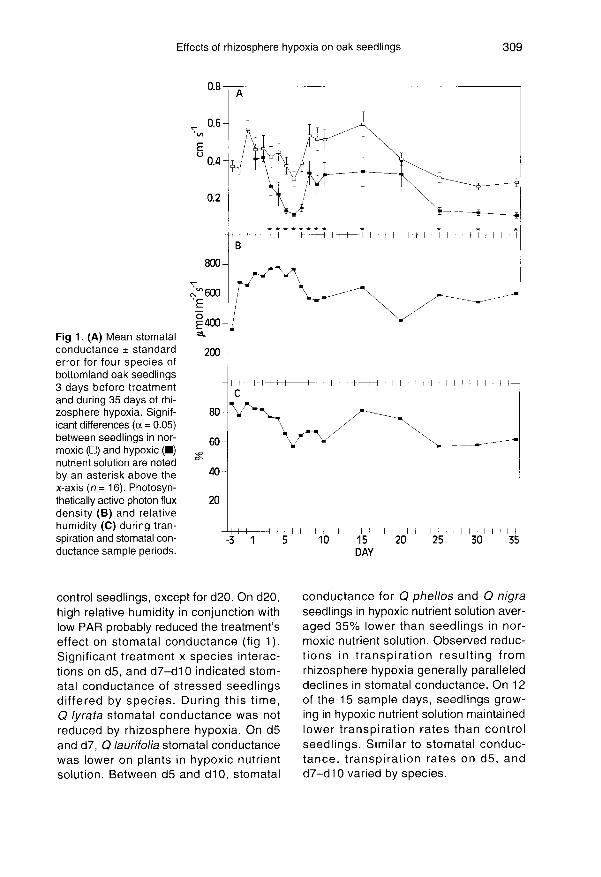
control seedlings, except for d20. On d20,high relative humidity in conjunction withlow PAR probably reduced the treatment’seffect on stomatal conductance (fig 1).Significant treatment x species interac-tions on d5, and d7-d10 indicated stom-atal conductance of stressed seedlingsdiffered by species. During this time,Q lyrata stomatal conductance was notreduced by rhizosphere hypoxia. On d5and d7, Q laurifolia stomatal conductancewas lower on plants in hypoxic nutrientsolution. Between d5 and d10, stomatal
conductance for Q phellos and Q nigraseedlings in hypoxic nutrient solution aver-aged 35% lower than seedlings in nor-moxic nutrient solution. Observed reduc-
tions in transpiration resulting fromrhizosphere hypoxia generally paralleleddeclines in stomatal conductance. On 12
of the 15 sample days, seedlings grow-ing in hypoxic nutrient solution maintainedlower transpiration rates than controlseedlings. Similar to stomatal conduc-tance, transpiration rates on d5, andd7-d10 varied by species.

Diurnal patterns of stomatal conductancewere similar for all species, but magnitudediffered by DO treatment (fig 2). Stomatalconductance of control seedlings increasedto 0.26 ± 0.02 cm s-1 at 1000 hours,decreased around midday, and slightlyincreased at 1600 hours before falling atnight (fig 2). Stomatal conductance wasgreatest on stressed seedlings at 1000hours when it averaged 0.13 ± 0.02 cm s-1.At the 0800 hours sample period, stomatalconductance of Q laurifolia, Q phellos, andQ nigra seedlings growing in hypoxic nutri-
ent solution showed lower rates than control
seedlings. However, stomatal conductanceof Q lyrata was not influenced by the treat-ment at this sample time. For all species,stomatal conductance during the ’middayslump’ was 85% of the maximum conduc-tance rate in control seedlings, but it was
77% of the maximum conductance rate instressed plants. Diurnal patterns of transpi-ration paralleled those of stomatal conduc-tance.
For these older seedlings, roots were theonly plant component in which biomass

accumulation was significantly reduced by35 days of rhizosphere hypoxia. Root sys-tems grown in normoxic nutrient solution
averaged 47% more biomass than thosegrown in hypoxic nutrient solution (data notshown). In spite of the decrease in rootbiomass accumulation, total plant biomassaccumulation was not changed by 35 days
of rhizosphere hypoxia. However, shifts inproportional biomass accumulation werefound after 35 days of rhizosphere hypoxia.
LWRs varied by species within nutrientsolution type (table III). Q lyrata and Q nigraseedlings grown in hypoxic nutrient solu-tion had proportionally more leaf biomassthan plants grown in normoxic nutrient solu-

tion (table III). LWR for Q laurifolia was notchanged by rhizosphere hypoxia, but theLWR of Q phellos declined 19% in hypoxicnutrient solution. Q nigra showed the great-est LWR in both nutrient solutions (table III).For all oaks, SWR increased 44% under 35days of rhizosphere hypoxia (table III). Forthree of four oak species, RWR decreasedduring 35 days of rhizosphere hypoxia (tableIII). This shift placed Q nigra below the otherthree species in terms of the proportion ofroot biomass maintained under rhizospherehypoxia. For all species, R/S ratios were40% lower when seedlings were raised inhypoxic nutrient solution (table III). In this
experiment, two lines of evidence indicatedifferences in R/S ratios were allocation
responses rather than a function of total
plant biomass. First, total plant biomass didnot differ between plants grown in hypoxicand normoxic nutrient solution. And, the cor-relation between R/S ratio and total plantbiomass was very low (correlation coeffi-cient = -0.30).
Relative height and diameter growth forseedlings grown in hypoxic nutrient solu-
tion differed by species (table IV). Q lyratarelative height growth was small and did notdiffer in response to rhizosphere hypoxia.The lack of growth in this species may relateto its late-season phenology. Among theErythrobalanus species, Q laurifolia grewequally well in either solution. However,Q phellos and Q nigra seedlings in hypoxicsolution attained only 30% of the relativeheight growth of seedlings in normoxic solu-tion (table IV). Under rhizosphere hypoxia,average Q lyrata diameter growth was over3.5 times larger than for seedlings grown innormoxic solutions (table IV). Q laurifoliaseedlings in hypoxic solutions grew over1.8 times larger in diameter than seedlingsin normoxic solutions (table IV). Thirty-fivedays of rhizosphere hypoxia did not changerelative diameter growth for Q phellos andQ nigra seedlings (table IV).
DISCUSSION AND CONCLUSIONS
Short-term photosynthetic decline for plantsgrowing in saturated soil is often ascribed to

partial closure of stomates (Peterson andBazzaz, 1984; Pezeshki and Chambers,1985a, b; Tsukahara and Kozlowski, 1986).Stomatal closure can reduce net photo-synthesis by limiting the intercellular CO2concentration available for assimilation
(Kozlowski and Pallardy, 1984). In Experi-ment 1, significant decline in net photo-synthesis was observed concomitant toreduced stomatal aperture. However, non-stomatal limitations to photosynthesis mayalso be observed after extended periods ofroot hypoxia (McLeod et al, 1986; Dreyeret al, 1991; Beckman et al, 1992; Vu and
Yelenosky, 1992). Kozlowski and Pallardy(1984) suggested that nonstomatal limita-tion to photosynthesis during soil satura-tion may include decline in chlorophyll con-centration and changes in carboxylationenzymes. In Experiment 1, leaves of plantsestablished under rhizosphere hypoxiaappeared more chlorotic than the leaveson plants established in normoxic nutrientsolution. Two stronger lines of evidenceindicate that nonstomatal mechanisms con-
tributed to limiting net photosynthesis ofoaks established in hypoxic nutrient solu-tion. First, ratios of net photosynthesis/inter-cellular CO2 decreased under rhizospherehypoxia indicating a decline in assimilatedCO2 per unit of available CO2. And, adecrease in δ13C was observed on plantsestablished under rhizosphere hypoxia indi-cating greater enzymatic discriminationagainst 13C. Greatest discrimination wouldbe expected in leaves having the largestCi/Ca ratios (Farquhar et al, 1982). Forexample, if CO2 assimilation was reduced
relatively more than stomatal conductance(Guy and Wample, 1984). In this study,stomatal and possible nonstomatal mech-anisms appeared to limit net photosynthe-sis of oaks established in hypoxic nutrientsolution. However, all species showed asimilar decrease in net photosynthesiswhen established in hypoxic nutrient solu-tion.
Partial stomatal closure can also account
for decreased transpiration under anaerobicsoil conditions, because transpiration isoften observed to decline without a con-
current change in leaf water potential(Pezeshki and Chambers, 1986; Osonubiand Osundina, 1987). Quick stomatalresponse to soil flooding decreased tran-spiration within 2 days of soil flooding forQ pagoda (Raf), Q michauxii (Nutt), and Qmacrocarpa (Michx) (Tang and Kozlowski,1982b; Pezeshki and Chambers, 1985a,1986). In Experiment 1, transpiration ofseedlings established in hypoxic nutrientsolution appeared to be limited primarily bystomatal closure, and it decreased similarlyfor all species. For the older seedlings,greatest stomatal disfunction was recordedbetween d5 and d10 of the stress. Duringthis period, Q lyrata seedlings were effectedthe least. Q phellos and Q nigra showedthe greatest decline in stomatal function
during this period.Diurnal patterns of stomatal conduc-
tance and transpiration observed in thesecond experiment were in general agree-ment with published results on otherspecies grown in flooded soil. Stomatalconductance is often reduced during mid-day, but the degree of reduction is typicallygreatest in stressed plants (Tang andKozlowski, 1982a,b). Relative to controlplants, seedlings grown in hypoxic nutri-ent solution exhibited lower stomatal con-
ductance and transpiration rates, and pro-portionally greater midday fluctuations ofthese parameters. Treatment effects onQ lyrata stomatal function were realizedlater in the day than on the other threespecies.
Rhizosphere hypoxia induced morpho-logical changes in leaves, stems, and rootsfor seedlings of both age classes. Roothypoxia can stimulate changes in leaf mor-phology by promoting abscission, reducingexpansion rate, or preventing leaf forma-tion (Sena Gomes and Kozlowski, 1980b;

Smit et al, 1990; Terazawa and Kikuzawa,1994). Leaf abscission was not observedin either experiment, but LWRs increasedfor all plants receiving rhizosphere hypoxia.In both experiments, Q lyrata and Q laurifoliamaintained the smallest proportion of leafbiomass under rhizosphere hypoxia, whileQ nigra maintained the highest LWR underrhizosphere hypoxia.
Stems of seedlings established in hypoxicnutrient solution were 20% smaller in mass
than those of seedlings established in nor-moxic nutrient solution. Q lyrata and Q lau-rifolia maintained a larger proportion of stembiomass than Q phellos and Q nigra, whenestablished under rhizosphere hypoxia.SWRs of the older seedlings were similarfor all species under rhizosphere hypoxia.Other hardwood mesophytes usually showa reduction in stem dry weight resulting fromroot hypoxia (Hosner and Boyce, 1962;Dickson et al, 1965; Tang and Kozlowski,1982b), but 4 months of soil flooding did notchange RWR of Q pagoda seedlings (Joneset al, 1989).
Root biomass is often reduced duringshort-term soil flooding, even on hydricspecies such as Taxodium distichum L(McLeod et al, 1986). Roots may showreduced biomass in anaerobic soil because
of reduced growth or decomposition (Hookand Brown, 1973; Angeles et al, 1986). Inthese experiments, evidence of root systemdecomposition was not visible. So, differ-ences in root biomass accumulation areattributed to root growth differences betweentreatments. During seedling establishment,RWR was decreased similarly for all
species. But in hypoxic nutrient solution,Q nigra maintained the proportionatelysmallest root system among the olderseedlings. Several authors have argued thatreduced root growth on seedlings in floodedsoil will decrease post-flood drought toler-ance of these seedlings (Tang andKozlowski, 1982a,b; Sena Gomes andKozlowski 1980b). In this study, decreased
root mass and relatively high proportions ofleaf mass may predispose Q phellos andQ nigra to slow post-flood recovery.
In both experiments, proportionalbiomass accumulation was shifted towardsthe shoot. R/S ratio of plants receivinghypoxic nutrient solution were lower thanthose of plants receiving normoxic nutrientsolution. This occurred apart fromdecreases in total plant biomass on thestressed seedings. Topa and McLeod(1986) found a similar root-shoot responsefor conifer seedlings which were grown inanaerobic nutrient solution. In contrast, two
Eucalyptus species showed proportionatelymore root growth than shoot growth, thiswas primarily a result of adventitious rootgeneration and leaf abscission under con-ditions of anaerobic soil (Sena Gomes andKozlowski 1980a).
Soil saturation during the growing sea-son typically reduces shoot growth of older,mesophytic seedlings (Dickson et al, 1965;Gill, 1970; Topa and McLeod, 1986).Though most of the energy and carbonrequired for initial shoot growth is providedby the acorn (Crow, 1988), rhizospherehypoxia limited shoot growth of all oaks dur-ing seedling establishment (Experiment 1).Thus, a functional root system is requiredto realize potential shoot growth of seedlingsduring establishment. For the older
seedlings, relative growth responses to rhi-zosphere hypoxia varied by species. Rela-tive height growth of Q phellos and Q nigrawere reduced 69% in hypoxic nutrient solu-tion. However, Q lyrata and Q laurifolia rel-ative diameter growth increased under rhi-zosphere hypoxia. Increased diametergrowth has been observed on other speciesgrown in flooded soil, and attributed to stemhypertrophy (Yamamoto et al, 1987). In thisstudy, rhizosphere hypoxia may haveinduced enlargement of the basal portionof stems, but there was not excessive stem
hypertrophy as reported for other speciessubjected to soil flooding (Gardiner, 1994).

Micromorphological examinations havefailed to reveal aerenchyma formation bythese and other bottomland oaks of the
southern United States (Pezeshki, 1991;Gardiner, 1994).
Results from the first experiment indi-cated early exposure to hypoxic nutrientsolution was equally lethal to all establish-ing species. However, seedlings allowedto recover demonstrated that shifts in plantmorphology may be important for resis-tance to root hypoxia during seedling estab-lishment. High LWRs along with significantreductions in root mass place Q phellosand Q nigra as the least resistant to roothypoxia during seedling establishment.Species resistance to rhizosphere hypoxiaduring seedling establishment could not beseparated through physiological or growthresponses. For the older seedlings, Q lyrataand Q laurifolia showed early physiologi-cal resistance, a morphology most con-ducive to survival under rhizospherehypoxia, and better shoot growth thanQ phellos and Q nigra. These findings donot completely explain bottomland oak dis-tribution in floodplains, and the results mayhave been different if seedlings were raisedin soil rather than nutrient solution. How-
ever, these findings indicate that low oxygenpartial pressures around roots can influ-ence bottomland oak physiology, morphol-ogy and growth. Thus, it potentially playsa strong role in determining establishment,survival and growth of bottomland oakregeneration.
ACKNOWLEDGMENTS
We thank the many people who assisted withdesign and implementation of this research. Twoanonymous reviewers are thanked for their con-structive comments on an earlier draft of this
manuscript. This article has been approved forpublication as Journal Article No FA-035-0695 ofthe Forest and Wildlife Research Center, Mis-sissippi State University.
REFERENCES
Angeles G, Evert RF, Kozlowski TT (1986) Develop-ment of lenticels and adventitious roots in flooded
Ulmus americana seedlings. Can J For Res 16, 585-590
Beatty SW (1984) Influence of microtopography andcanopy species on spatial patterns of forest under-story plants. Ecology 65, 1406-1419
Beckman TG, Perry RL, Flore JA (1992) Short-termflooding affects gas exchange characteristics ofcontainerized sour cherry trees. Hort Sci 27, 1297-1301
Bonner FT, Vozzo JA (1987) Seed biology and tech-nology of Quercus. USDA, Forest Service, Gen TechRep SO-66
Crow TR (1988) Reproductive mode and mechanisms forself-replacement of northern red oak (Quercusrubra) - a review. For Sci 34, 19-40
Dickson RE, Hosner JF, Hosley NW (1965) The effectsof four water regimes upon the growth of four bot-tomland tree species. For Sci 11, 299-305
Dreyer E, Colin-Belgrand M, Biron P (1991) Photosyn-thesis and shoot water status of seedlings from dif-ferent oak species submitted to waterlogging. Ann SciFor 48, 205-214
Farquhar GD, O’Leary MH, Berry JA (1982) On the rela-tionship between carbon isotope discrimination andthe intercellular carbon dioxide concentration in
leaves. Aust J Plant Physiol 9, 121-137
Fowler NL (1986) Microsite requirements for germinationand establishment of three grass species. Am MidlNat 115, 131-145
Gardiner ES (1994) Physiological responses of four bot-tomland oak species to root hypoxia. PhD Disser-tation, Mississippi State University
Gill CJ (1970) The flooding tolerance of woody species- a review. For Abst 31, 671-688
Guy RD, Wample RL (1984) Stable carbon isotope ratiosof flooded and nonflooded sunflowers (Helianthusannuus). Can J Bot 62, 1770-1774
Hanson PJ, Dickson RE, Isebrands JG, Crow TR, DixonRK (1986) A morphological index of Quercusseedling ontogeny for use in studies of physiologyand growth. Tree Physiol 2, 273-281
Harper JL, Williams JT, Sagar GR (1965) The behaviourof seeds in soil. I. The heterogeneity of soil surfacesand its role in determining the establishment of plantsfrom seed. J Ecol 53, 273-286
Hartgerink AP, Bazzaz FA (1984) Seedling-scale envi-ronmental heterogeneity influences individual fit-ness and population structure. Ecology 65, 198-206
Hodges JD, Switzer GL (1979) Some aspects of theecology of southern bottomland hardwoods. In: North

America’s Forests: Gateway To Opportunity. Pro-ceedings of the 1978 Joint Convention of the Societyof American Foresters and the Canadian Instituteof Forestry, Society of American Foresters, Wash-ington, DC, 360-365
Hook DD, Brown CL (1973) Root adaptations and rela-tive flood tolerance of five hardwood species. ForSci 19, 225-229
Hosner JF, Boyce SG (1962) Tolerance to water satu-rated soil of various bottomland hardwoods. For Sci
8, 180-186
Huenneke LF, Sharitz RR (1986) Microsite abundanceand distribution of woody seedlings in a South Car-olina cypress-tupelo swamp. Am Midl Nat 115, 328-335
Huenneke LF, Sharitz RR (1990) Substrate heterogeneityand regeneration of a swamp tree, Nyssa aquatica.Am J Bot 77, 413-419
Jones JB Jr, (1983) A Guide for the Hydroponic and Soil-less Culture Grower. Timber Press, Portland, OR
Jones RH, Sharitz RR, McLeod KW (1989) Effects offlooding and root competition on growth of shadedbottomland hardwood seedlings. Am Midl Nat 121,165-175
Kozlowski TT, Pallardy SG (1984) Effects of flooding onwater, carbohydrate, and mineral relations. In: Flood-ing and Plant Growth (TT Kozlowski, ed), AcademicPress Inc, Orlando, FL, 165-193
Marks PL, Harcombe PA (1981) Forest vegetation ofthe Big Thicket, southeast Texas. Ecol Monog 51,287-305
McKnight JS, Hook DD, Langdon OG, Johnson RL(1981) Flood tolerance and related characteristicsof trees of the bottomland forests of the southernUnited States. In: Wetlands of Bottomland Hard-wood Forests (JR Clark, J Benforado, eds), Else-vier Scientific Pub Co, New York, 29-69
McLeod KW, Donovan LA, Stumpff NJ, Sherrod KC(1986) Biomass, photosynthesis and water use effi-ciency of woody swamp species subjected to flood-ing and elevated water temperature. Tree Physiol2, 341-346
Molofsky J, Augspurger CK (1992) The effect of leaf lit-ter on early seedling establishment in a tropical for-est. Ecology 73, 68-77
Osonubi O, Osundina MA (1987) Stomatal responsesof woody seedlings to flooding in relation to nutrientstatus in leaves. J Exp Bot 38, 1166-1173
Petersen RG (1985) Design and Analysis of Experi-ments. Marcell Dekker Inc, New York
Peterson DL, Bazzaz FA (1984) Photosynthetic andgrowth responses of silver maple (Acer saccharinumL) seedlings to flooding. Am Midl Nat 112, 261-272
Pezeshki SR (1991) Root responses of flood-tolerantand flood-sensitive tree species to soil redox condi-tions. Trees 5, 180-186
Pezeshki SR, Chambers JL (1985a) Responses of cher-rybark oak seedlings to short-term flooding. For Sci31, 760-771
Pezeshki SR, Chambers JL (1985b) Stomatal and pho-tosynthetic response of sweet gum (Liquidambarstyraciflua) to flooding. Can J For Res 15, 371-375
Pezeshki SR, Chambers JL (1986) Variation in flood-induced stomatal and photosynthetic responsesof three bottomland tree species. For Sci 32, 914-923
Ponnamperuma FN (1984) Effects of flooding on soils.In: Flooding and Plant Growth (TT Kozlowski, ed),Academic Press Inc, Orlando, FL, 9-45
Putnam JA, Furnival GM, McKnight JS (1960) Man-agement and Inventory of Southern Hardwoods.USDA Agricultural Handbook, 181
Sena Gomes AR, Kozlowski TT (1980a) Effects of flood-ing on Eucalyptus camaldulensis and Eucalyptusglobulus seedlings. Oecologia 46, 139-142
Sena Gomes AR, Kozlowski TT (1980b) Responses ofPinus halepensis seedlings to flooding. Can J ForRes 10, 308-311
Smit BA, Neuman DS, Stachowiak ML (1990) Roothypoxia reduces leaf growth. Plant Physiol 92, 1021-1028
Tang ZC, Kozlowski TT (1982a) Physiological, mor-phological, and growth responses of Platanus occi-dentalis seedlings to flooding. Plant Soil 66, 243-255
Tang ZC, Kozlowski TT (1982b) Some physiologicaland morphological responses of Quercus macro-carpa seedlings to flooding. Can J For Res 12, 196-202
Tanner JT (1986) Distribution of tree species in Louisianabottomland forests. Castanea 51, 168-174
Terazawa K, Kikuzawa K (1994) Effects of flooding onleaf dynamics and other seedling responses inflood-tolerant Alnus japonica and flood-intolerantBetula platyphylla var japonica. Tree Physiol 14,251-261
Topa MA, McLeod KW (1986) Responses of Pinusclausa, Pinus serotina and Pinus taeda seedlingsto anaerobic solution culture. I. Changes in growthand root morphology Physiol Plant 68, 523-531
Tsukahara H, Kozlowski TT (1986) Effect of floodingand temperature regime on growth and stomatalresistance of Betula platyphylla var japonicaseedlings. Plant Soil 92, 103-112
Vu JC, Yelenosky G (1992) Photosynthetic responses ofrough lemon and sour orange to soil flooding, chill-ing, and short-term temperature fluctuations duringgrowth. Environ Exp Bot 32, 471-477
Yamamoto F, Kozlowski TT, Wolter KE (1987) Effect offlooding on growth, stem anatomy, and ethylene pro-duction of Pinus halepensis seedlings. Can J ForRes 17, 69-79




![[] Microbial Ecology of the Rhizosphere(BookFi.org)](https://static.fdocuments.net/doc/165x107/55cf944c550346f57ba106f1/-microbial-ecology-of-the-rhizospherebookfiorg.jpg)













