Photobleaching fluorescent actin in a fibroblast
17
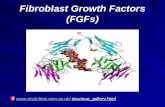
Alberts Fig. 16.56 Photobleaching fluorescent actin in a fibroblast • The fluorescent mark moves backward with respect to the front cell edge (and with respect to the substratum). Slow moving cell Y-L Wang et al., 1985. JCB 101:597-602 Experiment #1 Conclusion: 1. Actin meshwork is flowing backward 2 New actin is polymerized at the edge
-
Upload
conan-dickerson -
Category
Documents
-
view
38 -
download
0
description
Photobleaching fluorescent actin in a fibroblast. Experiment #1. The fluorescent mark moves backward with respect to the front cell edge ( and with respect to the substratum). Alberts Fig. 16.56. Slow moving cell. Conclusion: 1. Actin meshwork is flowing backward - PowerPoint PPT Presentation
Transcript of Photobleaching fluorescent actin in a fibroblast
Alberts Fig. 16.56
Photobleaching fluorescent actin in a fibroblast
• The fluorescent mark moves backward with respect to the front cell edge (and with respect to the substratum).
Slow moving cell
Y-L Wang et al., 1985. JCB 101:597-602
Experiment #1
Conclusion:1. Actin meshwork is flowing backward2 New actin is polymerized at the edge
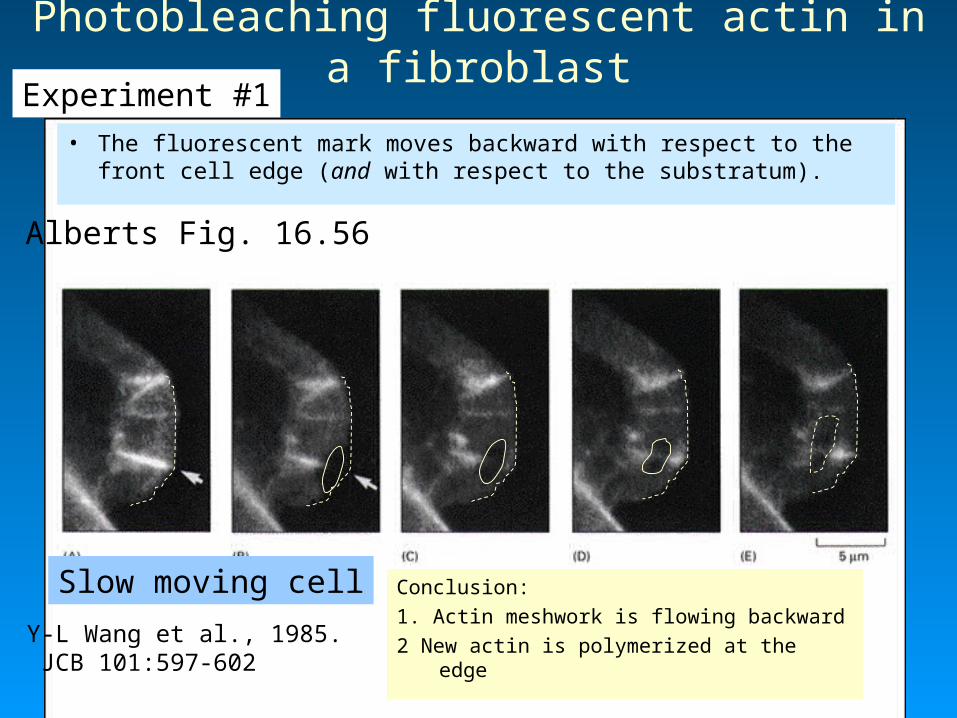
Alberts Fig. 16.58
Models of actin filament growth during protrusion: Treadmilling versus nucleation-release
Vic Small
Tim Mitchison
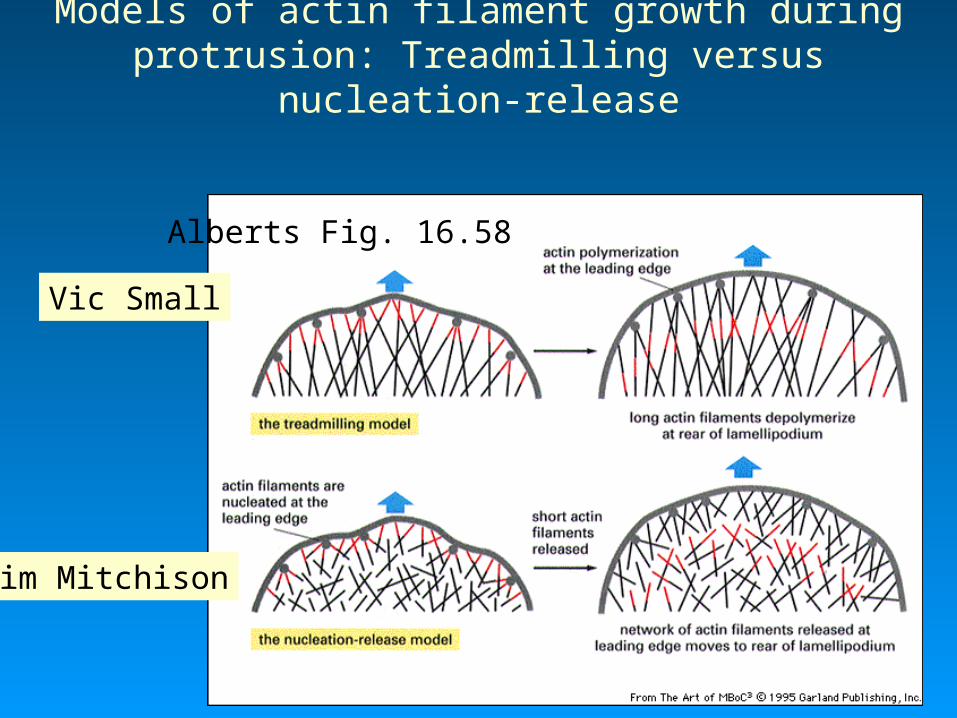
Photoactivation of fluorescence in a moving keratocyte
• The fluorescent mark moves backward with respect to the front cell edge (but not with respect to the substratum).
Fast moving cell Theriot and Mitchison, 1991 Nature 352: 352-131
Fig. 18-19b

Treadmilling vs. nucleation-release
This is figure 4, from Theriot and Mitchison, 1991. It is posted under “papers” on HuskyCTImage quality is lost when converting to PDF

Rate of actin turnover is consistent with nucleation-release model
• (Small) EM studies show long actin filaments in lamellae of slow moving fibroblasts
• (Others) EM studies in rapidly moving cells show a dense meshwork of actin filaments of different lengths
• Photobleaching experiments show rate of actin filament turnover to be greater than expected if treadmilling of long filaments occurs
• BUT Small criticized interpretation of photobleaching experiments– Depolymerization of actin
filaments makes estimate of filament turnover artificially high

Current view
• Dendritic nucleation model (Mullins, 1998)• Confirmed by the discovery of Arp2/3 at leading edge of motile
cells• Varible lengths of F-actin in lamellae of motile cells• Treadmilling occurs in individual filaments that are nucleated
and released from these sites (if uncapped at both ends).• Treadmilling occurs in the whole mass of actin filament
meshwork
Question: If the rearward movement of bleached marks is due to treadmilling, then why do we observe faster rearward movement (w.r.t. substratum) in cells with slow rates of actin turnover?

When actin polymerization is inhibited the actin meshwork continues to move rearward
Mechanical inhibition of retrograde flow
(see movie)

• In fast moving cells contractile forces at the front edge are low compared to the strength of adhesions
• No rearward actin flow – so that most newly polymerized actin contributes to protrusion – bleached marks are stationary w.r.t. substratum
• In slow moving cells contractile forces at the front are high (but not higher) compared to the strength of adhesions
• Actin flows rearward because most newly polymerized actin “feeds” the flow – protrusion is limited - bleached marks move back w.r.t. substratum
Adhesions allow mechanical coupling between a contractile cytoskeleton and
the substratum
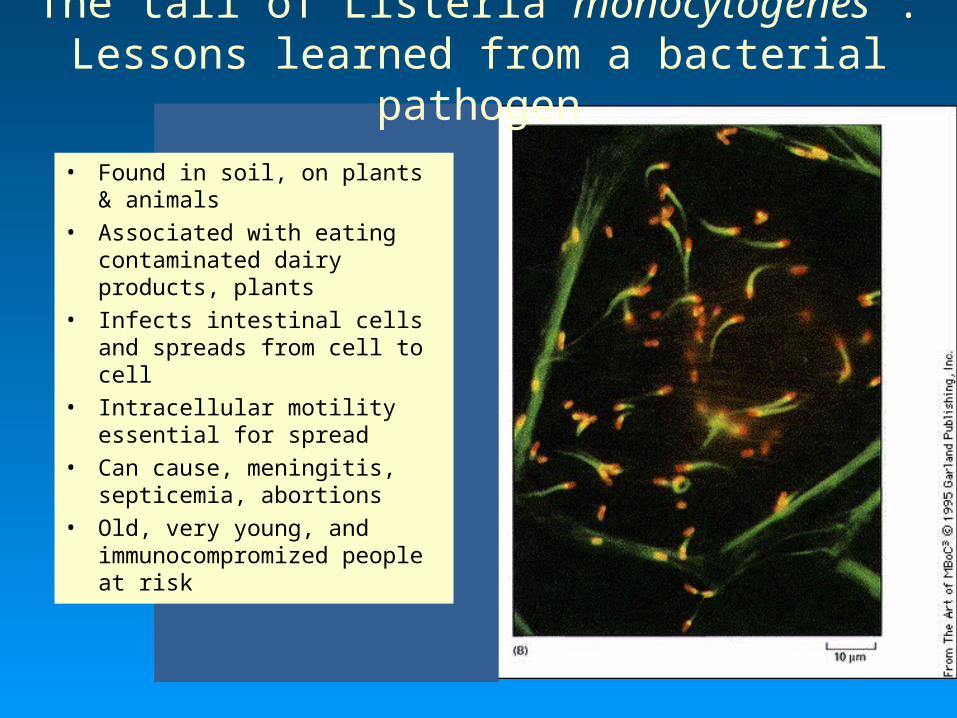
The tail of Listeria monocytogenes : Lessons learned from a bacterial pathogen
• Found in soil, on plants & animals
• Associated with eating contaminated dairy products, plants
• Infects intestinal cells and spreads from cell to cell
• Intracellular motility essential for spread
• Can cause, meningitis, septicemia, abortions
• Old, very young, and immunocompromized people at risk
Movie
• Listeria rocketing in infected cell
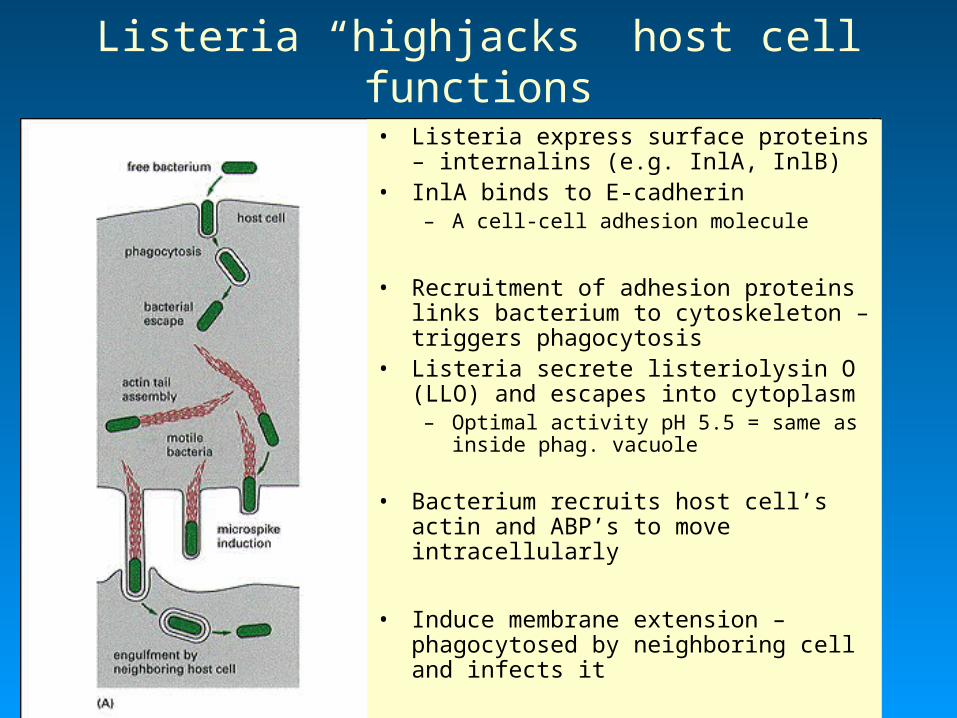
Listeria “highjacks” host cell functions• Listeria express surface proteins –
internalins (e.g. InlA, InlB)• InlA binds to E-cadherin
– A cell-cell adhesion molecule
• Recruitment of adhesion proteins links bacterium to cytoskeleton – triggers phagocytosis
• Listeria secrete listeriolysin O (LLO) and escapes into cytoplasm– Optimal activity pH 5.5 = same as
inside phag. vacuole
• Bacterium recruits host cell’s actin and ABP’s to move intracellularly
• Induce membrane extension – phagocytosed by neighboring cell and infects it

Advantages of studying Listeria
• 1. Doesn’t have the drawbacks of other whole cell systems– e.g. Some cytoskeletal mutations are lethal – Functions of many ABPs are redundant– Difficult to reconstitute cell motility – it requires a plasma
membrane
• 2. No plasma membrane• 3. Motility can be reconstituted in vitro• 4. The tail of Listeria is analogous to a lamellipodium of a
moving cell
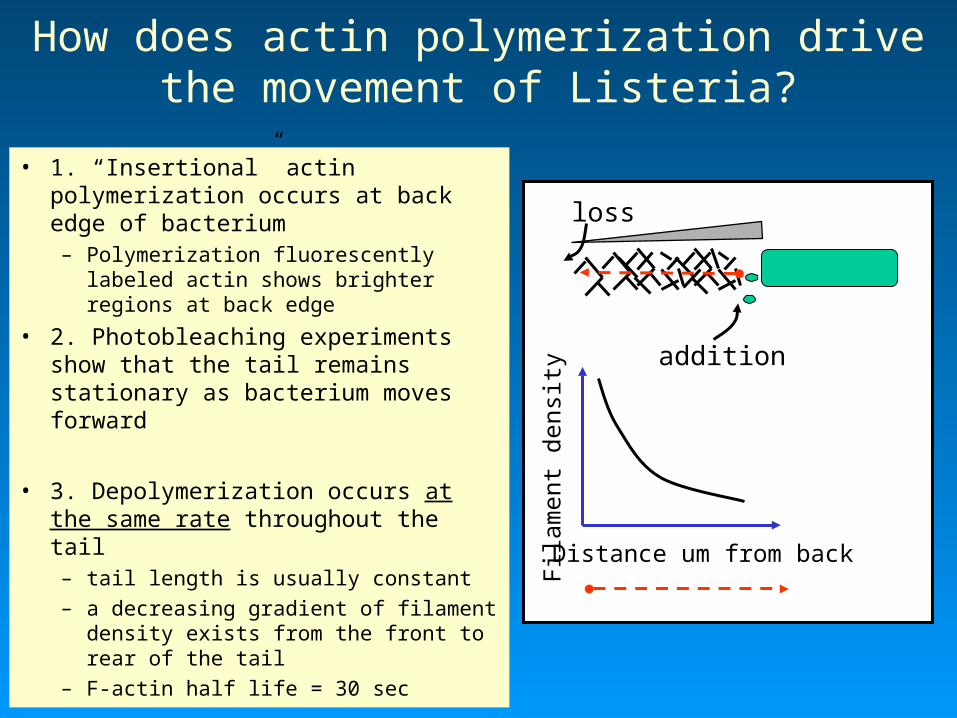
How does actin polymerization drive the movement of Listeria?
• 1. “Insertional” actin polymerization occurs at back edge of bacterium– Polymerization fluorescently
labeled actin shows brighter regions at back edge
• 2. Photobleaching experiments show that the tail remains stationary as bacterium moves forward
• 3. Depolymerization occurs at the same rate throughout the tail– tail length is usually constant– a decreasing gradient of filament
density exists from the front to rear of the tail
– F-actin half life = 30 sec
Distance um from backFi
lam
ent d
ensi
ty
addition
loss
How does actin polymerization become localized at one end of the
bacterium?• 1.Identification of nucleation factors• 2. Symmetry breaking (later)
• In the early 90’s used a genetic screen in mutant Listeria that could not form tails, and “normal” ones
• Found a single gene actA - encodes a bacterial surface protein ActA
• Can induce tail formation in:– Immotile Listeria, other bacteria, polystyrene beads

Act A is required for tail formation
• Does not bind directly to actin
• Looked for proteins that localized to the back edge of bacterium that are not seen in the tail.
• 1. Found VASP (vasodilator-stimulated phosphoprotein)– discovered by immunofluorescence studies – Binds to proline rich region of Act-A – Known to be associated with F-actin and focal adhesions in lamellipodia
• 2. Profilin – binds VASP• VASP and profilin accelerate filament elongation but are not nucleators
– Evidence: Actin clouds form in profilin depleted cytoplasmic extracts– VASP-actin complexes have no nucleating activity
• How is elongation accelerated?
• Poly proline regions bind multiple VASP molecules– Evidence: Bacterial speed is proportional to number of proline-rich repeats in
ActA• GFP-profilin concentration at back edge is proportional to speed
Bact. Memb. anchor sequence
N term C term
Signal peptide Proline-rich repeats
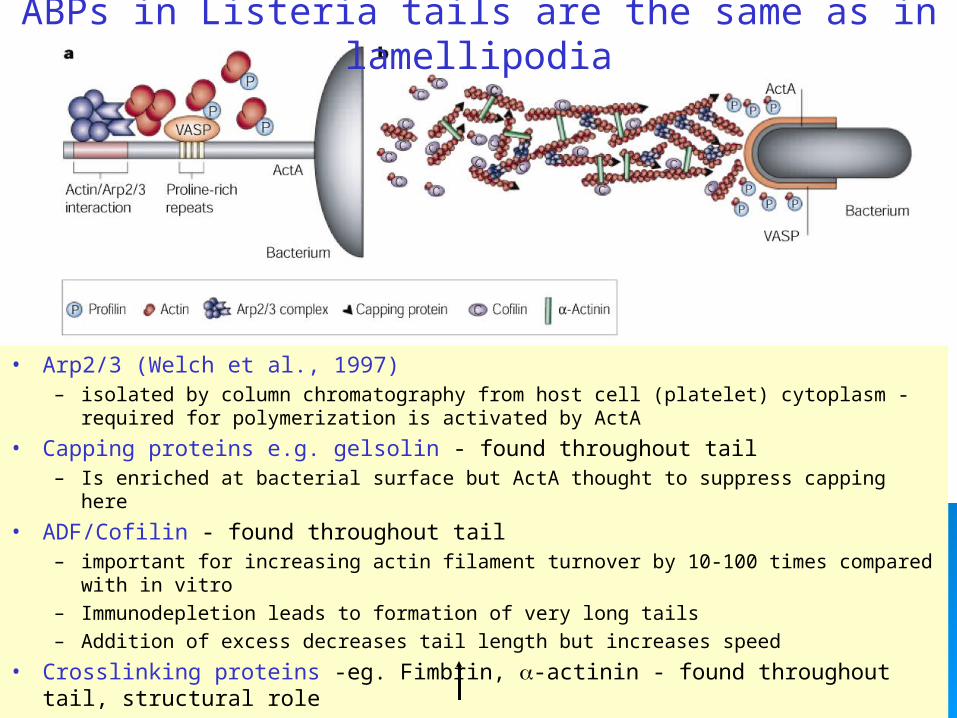
ABPs in Listeria tails are the same as in lamellipodia
• Arp2/3 (Welch et al., 1997)– isolated by column chromatography from host cell (platelet) cytoplasm -
required for polymerization is activated by ActA
• Capping proteins e.g. gelsolin - found throughout tail – Is enriched at bacterial surface but ActA thought to suppress capping here
• ADF/Cofilin - found throughout tail – important for increasing actin filament turnover by 10-100 times compared
with in vitro– Immunodepletion leads to formation of very long tails– Addition of excess decreases tail length but increases speed
• Crosslinking proteins -eg. Fimbrin, -actinin - found throughout tail, structural role– introduction of dom. negative fragment stops bacteria movement
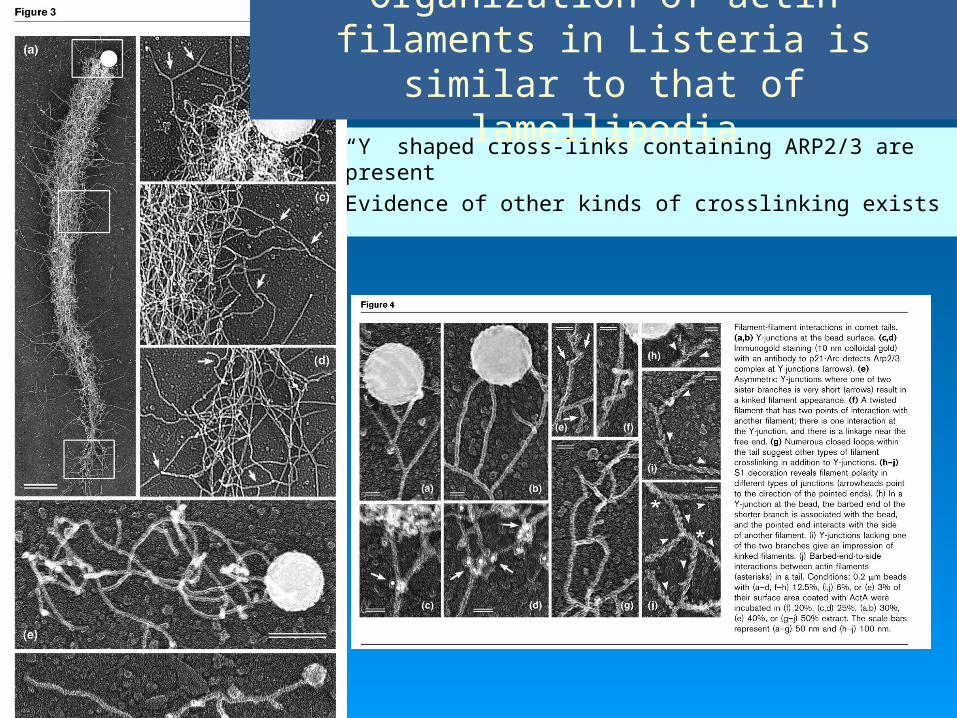
• “Y” shaped cross-links containing ARP2/3 are present
• Evidence of other kinds of crosslinking exists
Organization of actin filaments in Listeria is similar to that of
lamellipodia





![CYTOSKELETON NEWS - fnkprddata.blob.core.windows.net · Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)]](https://static.fdocuments.net/doc/165x107/609edd2b88630103265d18ee/cytoskeleton-news-dynamic-remodeling-of-the-actin-cytoskeleton-ie-rapid-cycling.jpg)