
Pflanzenphysiologisches Praktikum (nicht vertieft) Version ...
49
Pflanzenphysiologisches Praktikum (nicht vertieft) Version 2003/2004 Vorbemerkung: Es ist unbedingt erforderlich, sich auf den jeweiligen Kurstag vorzubereiten und die entsprechende Kursanleitung vor Versuchsbeginn sorgfältig zu lesen. Sehr zu empfehlen ist, ein Schema des Versuchsablaufs zu entwerfen. Wichtig: Bitte gehen Sie sorgsam mit den Glaswaren und Geräten um. Für Schäden an Sonderglaswaren bzw. grösseren Geräteschaden haftet jede/r Student/in selbst. Bitte beachten Sie zuverlässig die in den ausliegenden Gefahren- und Sicherheits- listen angegebenen Hinweise zur Entsorgung der Chemikalien. Die gekennzeichneten Abfallbehälter stehen unter den Abzügen. Über jede Aufgabe ist ein Protokoll zu schreiben. Dieses sollte folgende Abschnitte enthalten: 1. Eine kurze Einleitung in das Thema des Versuches 2. Versuchsdurchführung, soweit es nicht in der Anleitung steht, 3. Messdaten in übersichtlicher Form, 4. Darstellung der Ergebnisse in Graphiken bzw. Tabellen, kurze Darstellung der Ergebnisse in Worten 5. Interpretation des Versuchsergebnisses. Das Protokoll ist am nächsten Kurstag dem jeweiligen Betreuer(in) zum Testat vorzulegen. Sind Verbesserungen erforderlich, bitte immer alte Version wieder mitabgeben. An jedem Arbeitsplatz liegt eine Liste mit den vorhandenen Geräten, Glaswaren etc. aus. Bitte vor und nach dem Versuch prüfen, ob alles vorhanden ist. Am Ende des Versuchs bitte die gebrauchten Glaswaren ausleeren, abspülen und in die Abwaschwannen geben, sowie den Platz sauber und aufgeräumt verlassen.
Transcript of Pflanzenphysiologisches Praktikum (nicht vertieft) Version ...

Pflanzenphysiologisches Praktikum (nicht vertieft)
Version 2003/2004 Vorbemerkung: Es ist unbedingt erforderlich, sich auf den jeweiligen K urstag vorzubereiten und die entsprechende Kursanleitung vor Versuchsbeginn sorgfältig zu lesen. Sehr zu empfehlen ist, ein Schema des Versuchsablaufs zu ent werfen. Wichtig: Bitte gehen Sie sorgsam mit den Glaswaren und Geräten um. Für Schäden an Sonderglaswaren bzw. grösseren Geräteschaden haftet jede/ r Student/in selbst. Bitte beachten Sie zuverlässig die in den ausliegenden G efahren- und Sicherheits-listen angegebenen Hinweise zur Entsorgung der Chemikali en. Die gekennzeichneten Abfallbehälter stehen unter den Abzügen . Über jede Aufgabe ist ein Protokoll zu schreiben. Dieses so llte folgende Abschnitte enthalten: 1. Eine kurze Einleitung in das Thema des Versuches 2. Versuchsdurchführung, soweit es nicht in der Anleitung steht, 3. Messdaten in übersichtlicher Form, 4. Darstellung der Ergebnisse in Graphiken bzw. Tabellen , kurze Darstellung der
Ergebnisse in Worten 5. Interpretation des Versuchsergebnisses. Das Protokoll ist am nächsten Kurstag dem jeweiligen Betr euer(in) zum Testat vorzulegen. Sind Verbesserungen erforderlich, bitte imm er alte Version wieder mitabgeben. An jedem Arbeitsplatz liegt eine Liste mit den vorhand enen Geräten, Glaswaren etc. aus. Bitte vor und nach dem Versuch prüfen, ob alle s vorhanden ist. Am Ende des Versuchs bitte die gebrauchten Glaswaren ausleeren, ab spülen und in die Abwaschwannen geben, sowie den Platz sauber und aufgeräu mt verlassen.

2
Inhaltsverzeichnis
Seite
I. Der Wasserhaushalt der Pflanze.................. ........................................... 3
II. Der Mineralstoffhaushalt....................................................................... 11
III. Die Photosynthese und der Ferntransport von As similaten im Phloem 14
IV. Die Physiologie des Stoffwechsels - Enzyme als Biokatalysatoren 22 V. Die Steuerung der pflanzlichen Entwicklung durch Hormone und die Bewegungen der Pflanzen 30 VI. Die Dissimilation 42 VII. Anhang A. Übersicht über Kurstage und Versuche 47 B. Literatur 47 C. Pipettierübungen 48

3
I. Der Wasserhaushalt der Pflanze Drei Aspekte des pflanzlichen Wasserhaushalts homoiohydrer Pflanzen werden hier behandelt, nämlich der Wasserhaushalt der Zelle, d ie Transpiration und der Wurzeldruck. Das Wasserpotential Eine konzentrierte Zuckerlösung (Sirup) nimmt aus der Luf t Wasser auf und verdünnt sich dadurch. Die Lösung hat also ein Potential . Da es sich um Wasser handelt, nennt man es Wasserpotential. Umgekehrt hält eine Lösung das Lösemittel Wasser fest. Um ihr Wasser zu entreissen, muss man Energie aufwenden, d.h. man muss ihr Wasserpotential überwinden . Die Zuckerlösung versucht Wasser aufzunehmen weil sie einen „Mangel“ an Wa sser hat, deshalb gibt man in diesem Fall dem Wasserpotential ein negati ves Vorzeichen. Je numerisch negativer das Wasserpotential ist, umso größer i st der „Wassermangel“ und umso mehr kann der Umgebung Wasser entzogen werden. Reines Wasser hat definitionsgemäss kein Potential, sein Wasserpotential ist Null. Damit man die Wasserpotentiale verschiedener Systeme zuei nander in Beziehung setzen kann, gibt man sie in der Dimension Druck (bar oder MegaPascal) an. Warum dies notwendig und sinnvoll ist, wird bei der Un tersuchung des Wasserhaushaltes einer Zelle deutlich.
1. Der Wasserhaushalt der Zelle Lebende Zellen sind osmotische Systeme, da durch ihre sele ktiv permeable Cytoplasmamembran Wasser rasch diffundiert, während gelö ste Substanzen in der Regel nur sehr viel langsamer permeieren können (= Osmose). Bei Konzentrationsunterschieden zwischen den Lösungen auf beide n Seiten der Plasmamembran (z. B. Aussenmedium und Cytosol) strömt Wasse r vom Ort des niedrigeren osmotischen Potentials zum Ort des höheren. Ist das osmotische Potential des Zellsaftes grösser als das des umgebenden Mediums, so nimmt die Zelle solange Wasser auf, bis der G egendruck der sich dehnenden Zellwand so gross ist, dass eine weitere Volume nvergrösserung durch Wasseraufnahme nicht mehr möglich ist. Die Zelle ist dann maximal gespannt (turgeszent). In diesem Zustand ist der Druck der Zellwand auf den Protoplasten (Wanddruck, Druckpotential) identisch mit dem Druck des Prot oplasten auf die Zellwand (Turgordruck). Beide Drücke haben aber entgege ngesetzte Vorzeichen. Man kann also mit wenigen Parametern die osmotischen Eig enschaften der Zelle beschreiben. Es sind dies:
1. Die osmotische Potentialdifferenz ∆π = πz - πm, d. h. die Differenz zwischen
dem osmotischen Druck des Zellsafts (πz ) und dem osmotischen Druck des
Mediums (πm); ∆π nennt man auch den potentiellen osmotischen Druck einer Zelle.

4
2. Der Turgordruck P, das ist der Druck, den der Protopla st auf die Zellwand ausübt und sein Gegendruck, der Wanddruck. Dieser Druck wird im Gewebe durch die angrenzenden Zellen noch verstärkt (Gewebedruc k G).
3. Das Wasserpotential ΨΨΨΨ der Zelle (früher Saugkraft genannt), das gleich der Differenz zwischen der osmotischen Potentialdifferenz und d em Wanddruck (Druckpotential) ist.
Ist das Medium ausserhalb der Zelle reines Wasser (osmotische s Potential = 0),
so ist ∆π = πz , d. h. es zählt nur die Konzentration des Zellsaftes. Die Zusammenhänge zwischen den drei genannten Parameter n werden durch die Wasserpotentialgleichung beschrieben:
∆∆∆∆ΨΨΨΨ Zelle-Aussenmedium = - ∆∆∆∆ππππ + (P + G) d.h.die Wasserpotentialdifferenz (zwischen Zelle und Aussen medium) ist die Summe von osmotischer Druckdifferenz und Zellwand (+Geweb e-)druck. Dies kann durch das osmotische Zustandsdiagramm der Zelle veranschauli cht werden:
Die Zelle ist voll turgeszent, Die Zelle ist schlaff, maximales Zellvolumen, die Zellwand eingedellt, maximal gespannte Zellwand Plasmolyse Saugkraft = 0 maximale Saugkraft
Abb. I/1: Osmotisches Zustandsdiagramm einer pflanzlichen Zelle (gestrichelte Linie) (Quelle: Larcher, W.)
Begriffe: Ist im Vergleich zum Zellsaft das Aussenmedium
a) hypotonisch, so nimmt die Zelle solange Wasser auf, bi s ∆π = (P + G) ist;
Zellvolumen nimmt ab

5
b) isotonisch, es erfolgt keinerlei Netto-Wasserfluss c) hypertonisch, so gibt die Zelle solange Wasser aus dem Z ellsaft ans Medium
ab, bis das osmotische Potential des Protoplasten gleich de m des Mediums ist:
∆π = (P + G). Schrumpft dabei der Protoplast so sehr, dass er sich von der Zellwand ablöst, spricht man von Plasmolyse: Die Zelle w ird schlaff (P + G = 0), das Gewebe welkt. (siehe Abb.I/2: B-E). Bringt man die Zelle in diesem Zustand in ein hypotonisches Medium, so nimmt sie wieder Wasser au f und die Plasmolyse geht zurück (Deplasmolyse).
Abb.I/2: Der Verlauf der Plasmolyse einer Pflanzenzel le (h= HECHTsche Fäden), (Quelle :A. Wild)
Aufgabe: Bestimmung des osmotischen Potentials ππππ einer Zelle Der osmotische Wert des Zellsafts kann prinzipiell über zwei verschiedene Methoden gemessen werden: Zum einen kann man einen Pre sssaft aus dem Gewebe gewinnen und dessen osmotisches Potential durch Gefrierpunktserniedrigung bestimmen. Dabei muss beachtet werden, dass während der Presssaftgewinnung weder Wasser verdunstet (d . h. die Gewebe antrocknen), noch der Presssaft verdünnt wird, z. B. durch dem Gewebe anhaftendes Wasser. Zu diesem zählt auch Wasser in den Leit bahnen (Blattadern). Die andere Methode, die auch im Praktikum durchgeführt wird, misst das osmotische Potential der Zelle über die Grenzplasmolyse: Bei der Grenzplasmolyse ist das osmotische Potential des Med iums geringfügig negativer als das der Zelle. Um letzteres zu erhalten, b estimmt man das osmotische Potential desjenigen Mediums, bei dem 50 % de r Zellen Grenzplasmolyse zeigen. Zu bedenken ist dabei, dass a) dadurch die Verschiedenarti gkeit der Zellen selbst in einem homogen erscheinenden Gewebe vernachlässigt wir d, und dass b) man sicher sein muss, dass während der Beobachtungszeit die als Osm otikum verwendete Substanz nicht in die Zellen gelangt. Eine weitere Schwierigkeit ist, dass lichtmikroskopisch die Ab hebung des Protoplasten von der Zellwand schwer zu erkennen ist, wei l der Protoplast farblos ist. Deshalb bevorzugt man Zellen, die einen gefärbten Protoplasten besitzen, z. B. aus der unteren Epidermis von Rhoeo discolor. Die Farbe sitzt zwar ausschliesslich in der Vakuole, aber das Cytoplasma, das als sch maler Saum die Vakuole umschliesst, steht völlig im osmotischen Gleichgewicht mit der Vakuole, so dass die Plasmolyse der Zelle als Schrumpfen der Vakuole sichtbar ist.
Grenzplasmolyse

6
Durchführung des Versuchs: Als Osmotikum des Mediums werden Saccharoselösungen folgende r Konzentrationen verwendet: 0; 0,1; 0.2; 0,25; 0,30; 0,35; 0,40; 0,45; 0,50; 0,60; 0,70 M. Jeweils 2 Flächenschnittpräparate der unteren Bl attepidermis von Rhoeo discolor werden in ca. 5 ml dieser einzelnen Lösungen (in 10 ml Bechergläsern) eingelegt. Nach ca. 45 min, wenn sich das osmotische System stabilisiert hat, werden die Präparate in einem Tropfen der jeweilige n Lösung mit dem Mikroskop betrachtet. Bei jeder Konzentration wird der prozentua le Anteil der plasmolysierten Zellen ermittelt. Bitte beachten Sie, dass der Flächenschnitt einerseits nicht zu dick ist, denn man soll ja bequem die plasmolysierten Epidermiszellen auszähl en können, andererseits darf er aber auch nicht zu dünn sein, denn so nst werden die Epidermiszellen angeschnitten und laufen aus.
Auswertung: Die Ergebnisse werden in ein Diagramm (Abszisse: Konzentra tion; Ordinate: % plasmolysierte Zellen) eingetragen, und diejenige Kon zentration, bei der zu 50 % Grenzplasmolyse eintritt, wird aus dem Diagramm abgelese n.
Fragen und Überlegungen: a) Wie gross ist das osmotische Potential der Epidermiszelle n und wie groß ist
das Wasserpotential zum einen in Wasser, zum anderen in Sa ccharoselösung derjenigen Konzentration, bei der Grenzplasmolyse eintr itt?
c) Ist das osmotische Potential einer Zelle im Stadium de r Grenzplasmolyse identisch mit dem einer turgeszenten Zelle?
2. Der Wasserstrom durch die Pflanze Die Pflanze ist über ihre Organe Wurzel, Spross und Blät ter in den Feuchtigkeitsgradienten (Wasserpotentialgradienten) zwischen dem (relativ) feuchten Boden und der (relativ) trockenen Luft eingesp annt. Die Abbildung I/3 zeigt, dass zwischen den einzelnen Bereichen des Boden-Wurzel -Spross-Blatt-Luft - Weges ein kontinuierlich negativeres Wasserpotential vo rliegt. Von den Zellen der Blätter bis in die Wurzeln zieht sich ein zusammenhängender Wasserfaden. Er wird durch die Kohäsionskräfte der Wasserdi polmoleküle zusammengehalten, die bis zu 3 MPa betragen können. Da W asserdipolmoleküle andererseits durch die Adhäsionskräfte auch fest an die hyd rophilen Tracheen- und Tracheidenzellwände gebunden sind, gleitet der Wasse rfaden gleichsam in einer Röhre aus Wassermolekülen nach oben. Der Motor für das Hochsteigen des Wassers ist die Wasserabgabe an der Blattoberfläche durch T ranspiration, die infolge der Kohäsionskraft ein Nachströmen des Wassers bewir kt. Die Zugkraft für den Wasserfluss ist also letztlich das Wasser-Sättigungsdefizit der Luft. Sie muss jedoch nicht nur den hydrostatischen Druck der Wassersäule üb erwinden (bei einem 100 m hohen Baum ca. 0,9 MPa), sondern auch den Reibungswiderstand des Wasserstroms in den Gefässkapillaren. Man schätzt die insge samt

7
notwendigen Druckgradienten auf 0,03 MPa . m-1 . Voraussetzung für die
Aufrechterhaltung des Wasserflusses ist ein intakter Wasserfad en. Das Eindringen von Luft (Embolie oder Cavitation), entweder durch m echanische Verletzung oder durch Ausfrieren von gelöster Luft im Winter, muss entwe der verhindert oder (vielleicht durch Wurzeldruck) wieder repariert werden.
Abb. I/3: Das kontinuierlich negativer werdende Wassert potential (Quelle: Munk, K.)
Die Geschwindigkeit des Wassertransports im Xylem kann durch einen einfachen Versuch veranschaulicht werden, indem man einen Pflanzenst engel kurz in eine Farbstofflösung stellt und die während der Versuchszeit zur ückgelegte Wegstrecke des Farbstoffs ausmisst.
Aufgabe: Feststellung der Geschwindigkeit des Wasse rstroms im Xylem Der Stengel einer Sonnenblume wird mit einer neuen Rasierklinge glatt ca. 1 cm über dem Erdniveau abgeschnitten und mit dem Schnitten de in ein Reagenzglas mit 1 %iger Na-Fluorescein-Lösung gestellt. Nach 30 Sekun den werden die Blätter abgeschnitten (möglichst schnell um die Transpiration zu sto ppen) und das oberflächlich anhaftende Fluorescein von der Eintauchstel le des Stengels abgespült. Der Stengel wird dann in 1 cm Abschnitte zerl egt. Die Abschnitte werden mit dem apikalen Ende nach oben auf einer Glasp latte aufgereiht und unter einer UV-Lampe betrachtet. Na-Fluorescein leuchte t in UV-Licht hell auf. Es wird festgestellt, wie weit die Fluorescein-Lösung inner halb der Zeit, bis die Blätter entfernt worden waren, transportiert wurde. Unter de m Mikroskop soll ausserdem festgestellt werden, welche Zellarten hauptsächlich am Tr ansport beteiligt sind. Berechnen Sie die Flussgeschwindigkeit im Xylem.

8
Fragen und Überlegungen: a) Wie gross ist die Strömungsgeschwindigkeit im Xylem?
b) Ist die im Versuch gemessene Geschwindigkeit bei gleichen Transpirations- bedingungen identisch mit der in einer nicht abgeschnit tenen Pflanze?
c) Wie wäre die Geschwindigkeit, wenn die Sonnenblume d oppelt so viele Xylem- gefässe hätte?
d) Was bedeutet "Geschwindigkeit", was bedeutet "Fluss"?
Das Potometer: Volumetrische Messung des Transpirat ionsstromes Der Transpirationsstrom kann auch volumetrisch mit einem P otometer ( oder Potetometer, s. Abb. I/4) gemessen werden. Mit dieser V orrichtung wird die im Zusammenhang mit der Transpiration einer Pflanze verbr auchte Wassermenge direkt ermittelt. Die Geschwindigkeit der Verringerung der Wassersäule im Messrohr (M) bei geschlossenem Hahn (H) ist ein direktes Mass für die transpirierte Wassermenge.
Abb. I/4: Das Po(te)tometer (Quelle: Schopfer, D.) 1. Bedingung Normale Raumtemperatur und normale Luftfeuchte
2. Bedingung: Normale Raumtemperatur und erhöhte Luftfeuchte (die Pflanze wird 5 min vor Messbeginn in nasses Filterpapier eingehüllt; dann Wasser vorrat aufgefüllt).
3. Bedingung: Normale Raumtemperatur und Wind (kalter Fön, 5 min vor Messbeginn fönen, Wasservorrat auffüllen, weiterfönen und messen) .
Ausführung : Das Potetometer wird über den Trichter bis zum Überlaufen mit Wasser gefüllt. Eine frisch abge-schnittene Sonnenblume wird, ohne den Stengel zu quetschen, in den rechten Schenkel eingeschraubt, wobei keine Luftblasen eingeschlossen werden dürfen. Dieser Bereich wird eng mit Parafilm umwickelt. Dann wird der Hahn (H) verschlossen, 5 Minuten abgewartet und dann die Geschwindigkeit der Wasserabnahme im Meßrohr bestimmt. Dabei wird die Pflanze nacheinander folgenden Bedingungen unterworfen:

9
Zu den Zeitpunkten 5, 10 und 15 min wird der Wasserver brauch abgelesen.
Auswertung: - Aus den Werten der Tabelle ist ein Diagramm "Wasserve rbrauch (in ml) gegen die Zeit" anzufertigen. - Interpretieren Sie das Diagramm
3. Der Wurzeldruck Auch wenn normalerweise der Wasserstrom in der Pflanze du rch die Transpiration getrieben wird, so muss es doch noch einen w eiteren Mechanismus des Wassertransports geben. Dies wird deutlich unter Bedin gungen, in denen keine Transpiration stattfinden kann, z. B. beim Frühjah rsaustrieb von blattabwerfenden Bäumen, oder beim "Bluten" eines St engelstumpfes. Ferner gibt es Pflanzen, die bei grosser Luftfeuchte Wasser in Tropfen form über die Blätter abgeben (Guttation). Verantwortlich für diese Phänome ne ist der Wurzeldruck, welcher eine osmotische Konsequenz der aktiven Aufnahmepro zesse in die Wurzel ist. Im Zentralzylinder wird durch aktive Transportprozesse (sel ektiv und energie-abhängig) eine erhöhte Teilchenkonzentration geschaffen, verglichen mit dem Aussenmedium der Wurzel. Hydrophobe Einlagerungen in d ie radialen Zellwände der Endodermis (Lignin) verhindern ein Rückströmen der konzentrierten Lösung aus dem Zentralzylinder. Wasser folgt dem Konzentrationsg radienten passiv (osmotisch) über die Zellen der Endodermis nach. Dies hat zur Folge, dass das Wasser im Zentralzylinder unter einem positiven Druck, de m Wurzeldruck, steht. Normalerweise tritt dieser positive Druck nicht in Erschein ung, da der Wassertransport durch Transpiration wesentlich rascher und v iel grösser ist. Nur in Fällen, in denen die Transpiration nicht abläuft, wir d er offensichtlich.
Aufgabe: Messung der Exsudationsrate bei Wurzeldruck Einer Boehmeria-Pflanze wird mit einer neuen Rasierklinge der Spross gl att unterhalb der Blätter abgeschnitten und an seiner Stel le eine 10 ml-Messpipette mit Hilfe einer kurzen, aber festen Schlauchverbindung a ufgesetzt. (Zuerst die Schlauchverbindung über den Stumpf ziehen, mit Wasser bi s oben hin auffüllen, und dann die Messpipette sorgfältig in die Schlauchverbi ndung einführen, bis die Verbindung dicht ist und Wasser an einer Markierung der Pipette steht.) In Abständen von jeweils 30 Minuten wird 2 Stunden lang die sezernierte Wassermenge in der Pipette gemessen. Die Wasserabgabe (in ml) wird in einem Diagramm gegen die Zeit aufgetragen.

10
Fragen und Überlegungen: a) Wie hoch ist die Pumpleistung aufgrund des Wurzeldrucks in einer
transpirierenden Pflanze, verglichen mit einer nicht tr anspirierenden?
b) Hängt der Wurzeldruck vom Wassergehalt des Bodens ab?
c) Spielt die Nährsalzkonzentration im Boden eine Rolle für den Wurzeldruck?
d) Kann Wurzeldruck durch Transport organischer Substanzen e ntstehen? Und ist das realistisch?
e) Kann Wurzeldruck entstehen in Situationen, in denen die Wurzel keinerlei gelöste Stoffe aus dem Boden aufnimmt?

11
II. Der Mineralstoffhaushalt Jedes Lebewesen benötigt bestimmte Mineralstoffe (Makro- und Spurenelemente), um leben zu können. In der Regel entziehen die Pflanzen die Nährstoffe dem Boden mit Hilfe des Wurzelsystems; sie können aber z. B. au ch über die Blätter Nährsalze aufnehmen (Blattdüngung). Die biologische Funktion von anorganischen Ionen ist viel fältig. Einige Beispiele seien kurz aufgeführt: 1. Als Baustoffe (Kiesel- und Kalkskelette vieler Einzelle r) oder Bestandteile von
Gerüstsubstanzen, wie Ca2+ und Mg2+ als Zellwandkomponenten. 2. Als Bestandteile von Wirkstoffen (Phosphat im ATP, Ei sen im Haem;
Magnesium im Chlorophyll; Kupfer in der Cytochromoxid ase). 3. Als Cofaktoren oder Effektoren bei enzymatischen Reakti onen (z. B. Mg-ATP-
Komplex bei ATP-abhängigen Reaktionen). 4. Zur Aufrechterhaltung von Ladungszuständen (elektrische n Feldern) an
Biomembranen (Reizleitung, Energiekonservierung, Transportvorgänge). 5. Zur Kontrolle des Quellungszustandes von Makromolekülen (Proteine und
Kohlenhydrate).
A) Selektive Ionenaufnahme Zellen können sowohl Ionen wie auch ungeladene Moleküle selektiv aus dem umgebenden Medium aufnehmen. Dabei stellt die Plasmam embran (das Plasmalemma) die Diffusionsbarriere dar, in die verschie dene Transportproteine (Carriermoleküle) eingebaut sind. Zwei verschiedene Mechanismen für die selektive Stoffaufn ahme sind bekannt: 1. Die Stoffaufnahme (Stoffabgabe) durch erleichterte Diffusion Treibende Kraft ist ein Konzentrationsgradient über e iner Membran. Der
Vorgang (Nettoflux) ist zu Ende, sobald auf beiden Sei ten der Membran die gleiche Konzentration erreicht ist.
2. Die aktive Stoffaufnahme (Stoffabgabe) Im Unterschied zu 1. kann bei der aktiven Stoffaufnahme - unter Energie-
verbrauch - ein Konzentrationsgradient über einer Memb ran aufgebaut werden, z. B. durch Anreicherung eines Stoffes im Innern der Zelle. Wird das energielieferende System der Zelle gehemmt (z. B. dur ch Entkopplung), so findet keine aktive Stoffaufnahme mehr statt.
Zu unterscheiden ist die selektive Stoffaufnahme vom sele ktiven Verbrauch eines Stoffes. So kann durch dauernden Verbrauch eines Stoffes (z. B. eines bestimmten Zuckers aus einem Zuckergemisch) selektiv ein Netto fluss dieses Stoffes in die Zelle erfolgen. Im Praktikum soll die Selektivität der Ionenaufnahme mi t einfachen Mitteln demonstriert werden. Hierfür verwendet man Salze, von denen bevorzugt das Kation oder das Anion aufgenommen wird. Bei der Kati onenaufnahme werden zum

12
Ladungsausgleich Protonen abgegeben, bei der Anionenau fnahme werden zusammen mit den Anionen Protonen aufgenommen. Im erst en Fall wird das Medium also angesäuert, im zweiten Fall alkalisiert. Sal ze, die durch selektive Ionenaufnahme zu einer Ansäuerung des Mediums führen, si nd physiologisch saure Salze (z. B. (NH 4)2SO4, nur NH4
+ wird aufgenommen), solche die eine Alkalisierung des Milieus zur Folge haben, sind physiologi sch basische Salze (z. B. KNO3, nur NO3
- wird aufgenommen). Die Selektivität der Ionenaufnahm e lässt sich in einem solchen Fall sehr einfach durch die pH-Veränder ung im Medium nachweisen.
Aufgabe: Messung der Ionenaufnahme (Stickstoffaufna hme) aus (NH 4)2SO4 und KNO3 durch Wurzeln von Weizenkeimlingen. An Wurzeln von Weizenkeimlingen wird (qualitativ) die selektive Ionenaufnahme von Ammonium bzw. Nitrat mit einem farbigen pH-Indika tor veranschaulicht. Als Indikator wird Bromkresolpurpur (giftig) verwendet, d as sich oberhalb von pH 6,8 purpur und unterhalb von pH 5,2 gelb färbt.
Durchführung: Es werden 3 Agarplatten hergestellt: 1. Agar in Aqua dest. gelöst (Kontrolle) 2. Agar in 5 mM (NH 4)2SO4 - Lösung 3. Agar in 10 mM KNO 3 – Lösung 50 ml 1,0 % Agar (0,5 g Agar / 50 ml entweder Aqua dest., (NH4)2SO4 oder KNO3) werden in einem 250 ml Erlenmeyerkolben auf einem he izbaren Rührer unter Rühren auf ca. 50°C erwärmt. Dann wird der pH-Wert m it 0,02 N HCl auf pH 5,5 eingestellt. Bitte beachten Sie dabei, dass der pH-Wert auch temperaturabhängig ist und daher vielleicht eine Temperaturkorrektur am pH -Meter notwendig sein kann. Dann wird der Agar kurz zum Kochen gebracht, in Pla stikschalen gefüllt und auf Raumtemperatur abgekühlt. Ca. 7 Tage alte Weizenpflanzen werden auf die Agarpla tten gelegt (Wurzeln nicht verletzen) und im Brutschrank bei 30°C und hoher Luftfe uchte (wassergetränktes Filterpapier hineinlegen) über Nacht stehengelassen. Am nächsten Morgen werden die Agarplatten mit 0,04 % Bromkresolpurpur-Lö sung pH 5,5 angefärbt. Beschreiben und diskutieren Sie das Ergebnis.
Aufgabe: Messung der Ionenaufnahme (Stickstoffaufna hme) aus (NH 4)2SO4 und KNO3 durch einzellige Algen (Chlorella). Mit einer Suspension von Chlorella wird die Änderung des pH-Werts im Medium auf Grund der Ionenaufnahme in Zeitabständen gemessen.

13
Die Ernte der Algen wird vom Assistenten durchgeführt: (Die Algensuspension wird in grossen Stahlbechern 5 min bei 3.000 U/min (ca. 1000 x g ) abzentrifugiert. Der Überstand wird abgegossen, die Algen in dest. Wasser aufgenommen, durch vorsichtiges Schwenken von der Wand gelöst und ein weiteres Mal abzentrifugiert. Nach Suspension in ca. 40 ml 5 mM KCl und 10 mM Glukose und 3 h Schütteln können sie weiterverarbei tet werden.)
Durchführung des Versuchs: Drei 100 ml Erlenmeyerkolben werden nach unterem Schem a mit der Algensuspension und Wasser beschickt, dann wird die Suspension mit 0.02 N HCl bzw. NaOH in allen Gefässen auf pH 5 eingestellt. Die p H-Bestimmung erfolgt stets in einem 25 ml Becherglas. Dann werden die Salzlösu ngen zugegeben, nochmals der pH gemessen, und notiert, und im Erlenmeye rkolben auf der Schüttelmaschine geschüttelt. Nach jeweils 15 min., insgesam t 5 mal, wird eine pH-Bestimmung durchgeführt. Dabei wird wiederum der g esamte Inhalt des Erlenmeyerkolbens in ein 25 ml Becherglas gegossen und un ter Schwenken nach ca. 1 min. der pH-Wert abgelesen. Achten Sie darauf, d ass die gesamte pH-Elektrode eintaucht. Bitte beachten: Lösungen müssen vor der Zugabe von (NH4)2SO4 - bzw KNO 3 - Lsg. auf pH 5 eingestellt werden !
Kontrolle
+ NH4 -Aufn.
- NO3-Aufn.
Algensuspension (ml) 8 8 8 H2O (ml) 8 6 6 Mit 0.02 N HCl bzw. NaOH auf pH 5 einstellen
pH 5 pH 5 pH 5
40 mM (NH4)2SO4 -Lsg. (ml) - 2 - 80 mM KNO3 -Lsg. (ml) - - 2
Auswertung: Die gemessenen pH-Werte werden in einem Diagramm gege n die Zeit aufgetragen. Das Ergebnis wird diskutiert.
Fragen und Überlegungen: a) Welche prinzipiellen Möglichkeiten sehen Sie, um die pH-Wert Veränderungen zu erklären? Wie könnte man experimentell diese Möglichkeiten unter scheiden? b) Hat die Ladung des Ions Folgen für seinen Transport? Begründen Sie die Antwort !

14
III. Die Photosynthese und der Ferntransport von As similaten im Phloem
Die Photosynthese
Der Prozess der Photosynthese lässt sich vereinfacht in Lichtr eaktionen (Sauerstoffentwicklung, NADP-Reduktion, ATP-Bildung) und Dunkelreaktionen (Kohlendioxid-Fixierung unter Verbrauch des im Licht g ebildeten NADPH+H+ und ATP) zerlegen. Die Lichtreaktionen finden an der Thyla koidmembran der Chloroplasten statt, die Dunkelreaktionen (Calvin-Zyklus) im Stroma. Die Endprodukte der Photosynthese sind Stärke und Saccharose. S tärke wird im Chloroplasten gebildet, Saccharose im Cytoplasma. Saccharo se ist die Transportform der Photosyntheseprodukte, sie wird im Phl oem zur Versorgung der nicht photosynthetisch aktiven Gewebe weitergeleitet. Als Summenformel lässt sich die Photosynthese so wiedergebe n:
6 CO2 + 12 H2O C 6H12O6 + 6 O2 + 6 H2O
A. Nachweis von Saccharose und Stärke als Photosynt heseprodukte
1. Saccharose Von einer belichteten und einer 24 Stunden lang verd unkelten Ricinuspflanze (ca. 4 Wochen alt) werden Blattscheiben von ausgewachsenen Bl ättern (ca. 500 mg, genaues Gewicht notieren) in kleine Stücke geschnitten und in insgesamt 5 ml 80 % Ethanol gemörsert (mit 2 ml anfangen, die restlichen ml zum Nachspülen von Mörser und Pistill verwenden). Der Extrakt wird in Zen trifugenbecher überführt und 5 min. in der Tischzentrifuge bei 4000 Upm zentrifugier t. Der Überstand, der u.a. auch die Saccharose enthält, wird mit einer Pasteurpipet te abgenommen und für die Saccharosebestimmung verwendet. Die Saccharosebestimmung wird mit dem Anthrontest durchge führt. Mit ihm können reduzierende Zucker nachgewiesen werden (wie zum Be ispiel Glukose und Fruktose, nicht jedoch Saccharose). Prinzip: Zuerst werden die im Extrakt vorhandenen redu zierenden Zucker durch alkalische Behandlung zerstört, danach wird die Saccharose d urch 80%ige Schwefelsäure in Glukose und Fruktose gespalten, die dann mit Anthron nachgewiesen werden können. Eichkurve: 50 µl Saccharoselösung (Ausgangskonzentration 2 mM) verschied ener Konzentration (siehe Tabelle) werden mit 50 µl 30 %ig er KOH 10 min lang gekocht (Vorsicht ! Löcher in den Deckel stechen). Nach dem Abkühlen werden 1,5 ml
Licht

15
Anthron-Reagens dazugegeben (gut mischen) und 10 min la ng bei 40°C im Wasserbad inkubiert. Durch Reduktion wird das gelbe Anthr on grünblau. Die Extinktion der Proben wird in Halbmikroliter-Küvetten bei 623 nm gegen H 2O gemessen. Tragen Sie die Konzentration gegen die Extinktion bei 623 nm auf. Pipettierschema der Eichkurve für Saccharose:
Konzentration [mM] 0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
Saccharose [µl] 0 5 10 15 20 25 30 35 40 45 50
Wasser [µl] 50 45 40 35 30 25 20 15 10 5 0
Von den Proben der Blattextrakte werden je 2 x 50 µl (Doppelprobe), wie bei der Eichkurve beschrieben, behandelt. Die Probe des belichtet en Blattes muss vorher 1:3 mit H2O verdünnt werden (dh. 1 Teil Probe (50 µl ), 2 Teile H2O (100 µl)). Berechnen Sie, wieviel µmol Saccharose/g Frischgewicht in den belichteten bzw. verdunkelten Blättern vorhanden waren.
2. Stärke: Von der belichteten und der verdunkelten Ricinuspflanze wird je ein Blattstück auf den Stärkegehalt überprüft. Stärke wird mit Jodkaliumjo did nachgewiesen; dabei färbt sich Stärke dunkelblau. Um die Farbreaktion besser se hen zu können, muss erst das Chlorophyll aus den Blättern entfernt werden. Dazu werden die Blattstücke in kochenden 80%igen Ethanol gelegt, mit Alu folie abgedeckt und gekocht, bis sie weiss sind. Sie werden kurz mit H 2O abgespült und dann in Jodkaliumjodid gelegt.
Beschreiben und diskutieren Sie Ihre Beobachtungen.
B) Chromatographische Trennung der Chloroplastenfar bstoffe
Allgemeines: Als Chromatographie bezeichnet man ein Verfahren, bei dem zur Trennung eines Stoffgemisches die Unterschiede in der Wanderungsgeschwindi gkeit der einzelnen Komponenten in einem System aus stationärer und bewegl icher Phase benutzt werden. Aufgrund der verschiedenen Mechanismen, die die Trennung hervorrufen, unterscheidet man u. a. folgende Arten: a) Adsorptionschromatographie:
Verschieden starke (reversible) Bindung der Komponenten an die Oberfläche eines Adsorbens. Als Adsorbens werden anorganische Oxide, H ydroxide und Carbonate (z. B. Al2O3, MgO, Ca(OH)2, CaCO3) oder gepulverte organische

16
Stoffe (z. B. Stärke, Polyamid) verwendet, als mobile Phase organische Lösungsmittel (z. B. Benzin, Benzol, Aceton, Alkohole oder Gemische davon).
b) Verteilungschromatographie: Verteilung der zu trennenden Substanzen zwischen 2 flüssige n Phasen, von denen eine (meist Wasser) durch Bildung von Hydrathülle n an einem Träger (z. B. Zellulose im Papier) stationär ist.
c) Ionenaustauschchromatographie: Reversible Bindung ionisierter Verbindungen an gegensi nnig geladene Gruppen eines Austauschers.
In der Praxis liegt oft eine Kombination dieser Mögli chkeiten vor, wobei eine von ihnen dominiert. Welche Art der Chromatographie man im Einzelfall verwendet und welche Systeme von Träger und Lösungsmittel man wählt, richtet sich nach der Natur der zu trennenden Stoffe. Für die Trennung lipophiler Stoffe, wie z. B. der Pl astidenpigmente, eignet sich am besten die Adsorptionschromatographie. Man kann dabei da s Adsorbens entweder als Säule (Säulenchrom.) oder als dünne Schicht auf eine r Trägerfolie oder Glasplatte (Dünnschichtchrom.) verwenden. In diesem Prakti kum wird das zweite Verfahren gewählt, das sich besser zur Untersuchung kleinster Stoffmengen eignet.
Ausführung:
1. Vorbereitung der Chromatographiekammer: Das Laufmittel besteht aus 200 ml Petrolether (Siedebe reich 40 - 60°C), 24 ml Isopropanol und 0,5 ml Aqua dest. (es wird gestellt). C a. 120 ml des Gemisches wird in die Kammer gegossen, deren Kammerwände vorher mit Filterpapier ausgekleidet werden. Die Kammer wird mit einer Glaspla tte geschlossen, wobei zur Abdichtung Exsikkatorfett auf den Kammerrand gestriche n wird. Diese Vorbereitung sorgt dafür, dass die Kammer vor Verwendu ng mit Laufmitteldämpfen gesättigt ist. Achtung: Das Entwicklungsgemisch ist explosiv! Kein Feuer!!! Unter dem Abzug arbeiten !!
2. Extraktion der Pigmente: 3 g Blätter von der Sprossspitze einer Sonnenblumen-Pfl anze werden kleingeschnitten und unter Zugabe einer Spatelspitze CaC O3 (zur Neutralisierung des sauren Zellsaftes) und etwas Seesand mit 3 ml kaltem A ceton in einer vorgekühlten (Kühlschrank!) Reibschale zerrieben. Nach der ersten, groben Zerstörung der Blattstruktur wird das Mörsern mit weiter en 3 ml Aceton fortgesetzt. Der Brei wird 5 min lang in der Tischzentrifuge bei vo ller Geschwindigkeit zentrifugiert. Der klare, tiefgrüne Überstand, der all e Pigmente enthält, wird in ein Reagenzglas gefüllt und in Eis gestellt.

17
3. Dünnschichtchromatographie: Eine Dünnschichtfertigplatte (Trennschicht Kieselgel) wird für die chromatographische Trennung verwendet. Eine Startlinie wird mit einem sehr weichen Bleistift 2 cm über dem unteren Rand markiert ( Trägerschicht dabei nicht verletzen!). Entlang dieser Linie werden 2 Auftragszone n von jeweils 4 cm Länge für die Probelösung ebenfalls mit dem Bleistift markier t, wobei zwischen jeder Auftragszone wie auch von den linken und rechten Platten kanten ein Abstand von mindestens 2 cm einzuhalten ist. Die Pigmentlösung wird a ls möglichst schmales Startband auf jede der Auftragszonen aufgetragen, wob ei einmal 100 µl und einmal 200 µl appliziert werden. Dies erfolgt durch vo rsichtiges Auftragen (tröpfchenweise) des Extraktes mit einer verstellbaren Pi pette und gleichzeitigem vorsichtigem Fönen. Die Trägerschicht soll beim Auftragen so wenig wie möglich verletzt werden. Die trockene Dünnschichtplatte wird in die Kammer gestell t (Auftragsende unten; Trägerschicht nach vorne gerichtet !) und die Kammer ver schlossen (auf Dichtigkeit achten !) und verdunkelt. Im Zuge des Höherst eigens des Laufmittels auf der Trägerschicht kann die Trennung der Farbstoffe v erfolgt werden. Wenn die Laufmittelfront ca. 2 cm vom oberen Rand der Platte a ngelangt ist (nach ca. 45 Minuten), wird die Platte aus der Kammer genommen. D ie Positionen der Laufmittelfront und der Pigmentbanden werden sofort (!) mit einem Bleistift markiert. Die Farben der Pigmentbanden werden notier t und der Abstand der Laufmittelfront und der einzelnen Pigmentzonen von de r Startlinie aus gemessen. Die Pigmente werden anhand der beigefügten Skizze iden tifiziert. -- -- -- -- -- -- -- -- --- -- -- -
- Front
___________________ β-Carotin ..…..……......….…............
. Oxidationsprodukt des Chlorophylls
____________________ Chlorophyll a ____________________ Chlorophyll b ____________________ Lutein ......................................
......................................
......................................
) ) Xanthophylle )
____________________ Start
Beispiel einer Auftrennung der Chloroplastenfarbstoffe durch Dünnschicht-chromatographie an Kieselgel mit dem beschriebenen Entw icklungsgemisch.
Auswertung: Die Ergebnisse der dünnschichtchromatographischen Trennung werden mass-stabsgerecht dargestellt. Dabei werden die Rf-Werte (re lative Wanderungsstrecken) der beobachteten Pigmentbanden angegeben.
frontLaufmittel
Pigments
der
des
Strecke
Strecke
gtezurückgele
gtezurückgeleRf
=

18
Deuten irgendwelche Banden auf Zerstörungsprodukte von Pigmenten hin? Wenn ja, welche? Warum soll die Trennung im Dunkeln stattfinden?
Der Ferntransport von Assimilaten im Phloem Der Langstreckentransport der Assimilate (vor allem Sacchar ose) vom Ort ihrer Synthese (Mesophyllzellen des grünen Blattes) zum Ort ihr es Verbrauchs (wachsendes Gewebe, Wurzel, Spross, Früchte etc.) überbrückt in relativ kurzer Zeit weite Strecken. Dies ist nur möglich, weil dafür ei gene Leitbahnen (Siebröhren-Geleitzellenkomplex) entwickelt worden sind. Betrachtet man diesen Ferntransport näher, lässt er sich in drei Schritte unter gliedern: -Phloembeladung: aus den Mesophyllzellen (source) in den Siebröhren-Geleitzellen-Komplex, -Ferntransport: in den Siebröhren, und -Phloementladung: aus den Siebröhren in die verbrauch enden (parenchymatischen) Zellen (sink).
Phloembeladung Da die Saccharosekonzentration in den Mesophyllzellen (1 - 50 mM) geringer ist als im Siebröhren-Geleitzellen-Komplex (200 - 400 mM) , muss man einen aktiven Aufnahmeprozess in die Siebröhren annehmen. Es ist noch u nklar, ob die Saccharose symplastisch, d. h. von Zelle zu Zelle durch cytop lasmatische Verbindungen in der Zellwand (Plasmodesmen), in die S iebröhren gelangt oder ob sie über die Plasmamembran der Mesophyllzelle in den Ze llwandbereich gelangt, dort bis zum Siebröhren-Geleitzellen-Komplex diffundiert und an der Plasmamembran des Komplexes von einem Transportprotein aufgenommen wird (apoplastischer Weg). Wahrscheinlich ist, dass der erste Teil des Weges symplastisch erfolgt, bis zum Phloemparenchym, die Saccharose dort in den Apoplasten gelangt und von einem Saccharosecarrier aktiv in die Siebröhre transportiert wird. Die aktive Saccharoseaufnahme ist an die Aufnahme von Protonen gekoppelt (H+-Saccharose-Symport).
Ferntransport 1930 wurde von E. MÜNCH die Druckstromhypothese entwickelt, die bis heute als Erklärung für den Mechanismus des Ferntransports in den Si ebröhren gilt . Die aktive Aufnahme von Saccharose in die Siebröhren ist mit dem passiven Nachströmen von Wasser aus dem benachbarten Xylem verbund en; dadurch vergrössert sich das Flüssigkeitsvolumen an der Phloembelade stelle und es entsteht ein Druck. Da die aktive Abgabe von Saccharose au s den Siebröhren ebenfalls mit einer passiven Abgabe von Wasser verbunden ist, entsteht so in der Siebröhre ein Druckgradient und ein Massenfluss von in Wa sser gelöster Saccharose (und allen anderen in der Siebröhre vorhand enen Substanzen). Dieser Vorgang erklärt auch, warum der Transport im Phloem an lebende Zellen gebunden ist, denn nur sie besitzen selektivpermeable Pla smalemma-Membranen, welches die Voraussetzung für ein osmotisches System ist.

19
Phloementladung Der Mechanismus der Phloementladung scheint stark gewebespe zifisch zu sein. Da in den Siebröhren eine höhere Saccharosekonzentration vorliegt als im Sink-Gewebe, wäre ein aktiver Abgabeprozess nicht unbedingt e rforderlich, die Saccharose könnte auch entsprechend ihrem Konzentrationsgrad ienten (passiv) in die verbrauchenden Zellen gelangen. Der Weg kann wied er, wie bei der Beladung, symplastisch oder apoplastisch sein, ist aber offensichtlich ge webespezifisch. In wachsendem Gewebe wird Saccharose wahrscheinlich symplastisch e ntladen; bei der Entladung in Speichergewebe (wie Samen, Speicherp arenchymen von Zuckerrübe und Kartoffel etc.) ist ein apoplastischer Schri tt zwischengeschaltet.
Phloemsaftzusammensetzung Der Hauptbestandteil des Siebröhrensaftes ist Saccharose, d och werden auch Aminosäuren und andere Stoffe im Phloem transportiert ; als Gegenion für Säuren dient vor allem Kalium. Ausser den genannten Verbindu ngen wurden im Phloemsaft ATP, Nucleinsäuren,Hormone, Vitamine und Pr oteine gefunden.
Abb. III/1: Münch’sche Druckstromtheorie: Modellvorstellung und physiologische Relevanz
(Quelle: Taiz & Zeiger)
Aufgabe a): Modellversuch zur Demonstration der Mass enströmung beim Phloemtransport
Ausführung: Der Versuch wird gemäss Abb. III/2 aufgebaut. In die li nke mit Wasser gefüllte Saugflasche wird eine konzentrierte, mit Neutralrot gef ärbte Saccharoselösung in

20
einen Dialyseschlauch eingebracht, der dem Phloem im Bel adungsbereich (z. B. im ausgewachsenen Blatt) entspricht. In der rechten Flasche befindet sich dagegen ein nur mit Wasser gefüllter Dialyseschlauch, der dem Phloem im Entladungsbereich (z. B. in der Wurzel) entspricht. Aufgr und des hohen osmotischen Potentials der Saccharoselösung nimmt diese aus i hrer unmittelbaren Umgebung durch die semipermeable Membran des Dialyseschla uches (durchlässig für Wasser, aber nur schwer für die gelösten Stoffe) Wasser auf. Beobachten Sie, was passiert, und beschreiben Sie es.
Abb. III/2: Versuchsanordnung zur Demonstration der Masse nströmung im Phloem nach Münch Siehe auch Abb. III/1
Diskussion:
1. Wie lange hält der Transport von “Phloem”-Substanze n in diesem Modellversuch an und warum kommt es zum Stillstand?
2. Welcher Unterschied besteht zur intakten Pflanze bezügli ch der Gleichgewichts- einstellung?
3. Erklären Sie aufgrund der Gesetzmässigkeiten, die diese m Versuch zugrunde

21
liegen, wann ein Phloemtransport in der Aufwärts richtung stattfinden kann.
Aufgabe b): Phloemsaftgewinnung und -analyse Ricinus gehört zu den wenigen Pflanzen, von denen man Phloem saft durch Anschneiden der Siebröhren gewinnen kann. Die meisten P flanzen besitzen einen schnell wirkenden Mechanismus, um verletzte Siebröhren zu v erschliessen. Bei Ricinus scheint dieser Mechanismus nur langsam wirksam zu werde n, so dass man einige Zeit Phloemsaft aus angeschnittenen Siebröhr en abnehmen kann.
Ausführung Mit einer scharfen Rasierklinge wird leicht in die Rinde des Ricinusstammes geritzt (vom Assistenten zeigen lassen). Der heraustretende Phloem saft wird in einer geeichten 5µl Kapillare bis zur Marke aufgefangen (bei einer gut exudierenden Pflanze kann eine zweite Kapillare gefüllt werden). De r Phloemsaft wird mit einem auf die Kapillare aufgesetzten Schlauch in ein Fiolax-R öhrchen mit 495 µl Wasser ausgedrückt. Durch Wiederansaugen der Flüssigkeit und Ausdrü cken des Schlauches wird die Kapillare zweimal gespült. Von dieser Probe wird eine Saccharosebestimmung, wie beim Versuch Photosynthese beschrie ben, durchgeführt.
Auswertung Berechnen Sie die Konzentration (mM) an Saccharose im P hloemsaft und vergleichen Sie sie mit der Konzentration im Blatt (µm ol/g Frischgewicht ≅ mM, da sich das Frischgewicht aus ca. 10 % Trockenmasse und 90 % Wasser zusammensetzt).

22
IV. Die Physiologie des Stoffwechsels - Enzyme als Biokatalysatoren
1. Allgemeines: Die Zelle benötigt Katalysatoren, um ihre metastabile n Verbindungen zur Reaktion zu bringen. Sie produziert hierfür eigene Biokatalysato ren (Enzyme), die sich durch eine hohe Spezifität und Effektivität auszeichnen. Als Ka talysatoren werden sie während der Reaktion nicht verändert, sie unterliegen jedoch - wie alle Zellbausteine - einem schnelleren oder langsameren Umsat z (“turnover”). Ihre Katalysatorfunktion erfüllen die Biokatalysatoren durch Bildung eines Enzym-Substratkomplexes (ES), der entweder zu Enzym plus Produkt reagiert oder einfach wieder zu den Ausgangsverbindungen dissoziiert:
Im Enzym-Substratkomplex werden die Reaktanten durch Bin dung an das Enzym (Ionen- oder Wasserstoffbrücken) in Reaktionsbereitschaft ge bracht und dadurch die Aktivierungsenergie erniedrigt.
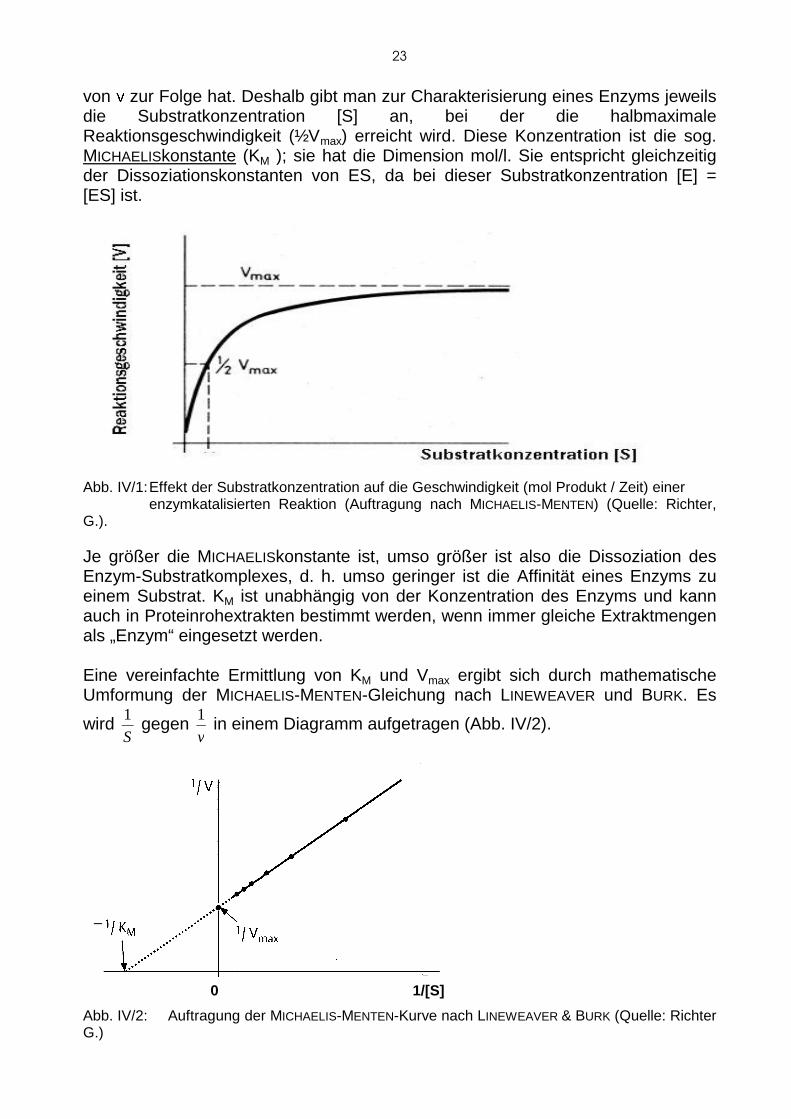
a. Charakterisierung der katalytischen Funktion Ein Katalysator verschiebt nicht das chemische Gleichgewicht, er beschleunigt lediglich dessen Einstellung. Deshalb besteht ein wichtige s Merkmal einer katalysierten Reaktion darin, dass die Reaktionsgeschwindigke it (bei nicht limitierendem Substrat) der Katalysatormenge proportional ist. Nach der Theorie von M ICHAELIS und MENTEN ist die Geschwindigkeit dieser Komplexbildung wesentlich grösser als die des Zerfalls von ES in E + P (k 1 >> k3 ). Vernachlässigt man k4 (welches meist sehr klein ist), so bestimmt also k 3 die gesamte Reaktionsgeschwindigkeit. Unter dieser oft zutreffe nden Voraussetzung ist der Umsatz von S nach P der Konzentration des Enzymsubst ratkomplexes [ES] direkt proportional und die Reaktionsgeschwindigkeit somit ein Maß für [ES]. Erhöht man bei konstanter Enzymmenge die Substratkonzentration [S], so ist auch eine Erhöhung der ES-Konzentration und folglich e ine Vergrösserung der Reaktionsgeschwindigkeit (v) zu erwarten. Dies lässt sich solan ge fortsetzen, bis alle Enzymmoleküle als Enzym-Substratkomplex vorliegen. D ann kann man durch Erhöhung der Substratkonzentration keine Steigerung der Reaktionsgeschwindigkeit mehr erzielen. Anders ausgedrückt he isst das: Eine enzymatisch katalysierte Reaktion zeigt (bei konstanter Enzym menge) das charakteristische Merkmal der Substratsättigung: (Abb. IV/1). Die Sättigungskonzentration, bei der die maximale Reakt ionsgeschwindigkeit (Vmax) erreicht ist, ist von Enzym zu Enzym und bei einer Reakt ion mit mehreren, verschiedenen Substraten für jedes Substrat verschieden. A llerdings lässt sich die sättigende Konzentration aus dem abgebildeten Diagramm (Auftragung nach MICHAELIS und MENTEN, Abb. IV/1) nicht genau ablesen, weil im sättigungsnah en Bereich eine grosse Änderung der Substratkonzentration nu r eine kleine Änderung
23
von ν zur Folge hat. Deshalb gibt man zur Charakterisierung e ines Enzyms jeweils die Substratkonzentration [S] an, bei der die halbmaxi male Reaktionsgeschwindigkeit (½Vmax) erreicht wird. Diese Konzentration ist die sog. MICHAELISkonstante (KM ); sie hat die Dimension mol/l. Sie entspricht gleichzei tig der Dissoziationskonstanten von ES, da bei dieser Substratko nzentration [E] = [ES] ist.
Abb. IV/1: Effekt der Substratkonzentration auf die Geschwindigkeit (mol Produkt / Zeit) einer enzymkatalisierten Reaktion (Auftragung nach M ICHAELIS-MENTEN) (Quelle: Richter, G.). Je größer die M ICHAELISkonstante ist, umso größer ist also die Dissoziation des Enzym-Substratkomplexes, d. h. umso geringer ist die Affi nität eines Enzyms zu einem Substrat. KM ist unabhängig von der Konzentration des Enzyms und kann auch in Proteinrohextrakten bestimmt werden, wenn imme r gleiche Extraktmengen als „Enzym“ eingesetzt werden. Eine vereinfachte Ermittlung von K M und Vmax ergibt sich durch mathematische Umformung der MICHAELIS-MENTEN-Gleichung nach LINEWEAVER und BURK. Es
wird S
1 gegen
v
1 in einem Diagramm aufgetragen (Abb. IV/2).
0 1/[S]
Abb. IV/2: Auftragung der M ICHAELIS-MENTEN-Kurve nach LINEWEAVER & BURK (Quelle: Richter G.)

24
Von der MICHAELIS-MENTEN Gleichung
SKSv
vM
+
⋅= max wird der Kehrwert gebildet
SvSK
vM
⋅
+=
max
1 und zerlegt zu
vSv
Kv
M
maxmax
111 +⋅
⋅=
Es wird S
1 gegen
v
1 in einem Diagramm aufgetragen (Abb. IV/2). Die Ger ade
schneidet die Ordinate im Abschnitt vmax
1 und die Abszisse im Abschnitt K M
1− .
b. pH-Optimum Die Aktivität der meisten Enzyme ist vom pH-Wert des Rea ktionsgemisches abhängig, sie weisen ein sog. pH-Optimum auf. Vielfach sind diese Optimumskurven Glockenkurven. Die pH-Empfindlichkeit der Enzyme wird durch folgende A spekte verständlich: 1. Bindung von Protonen an ionisierbare Gruppen der Substratbindestelle des aktiven Zentrums oder an andere Stelle des Proteins, da durch wird dann seine Molekülstruktur und Wirksamkeit verändert 2. Bindung oder Dissoziation von Protonen an das Substra t. Das pH-Optimum eines Enzyms ist nicht notwendigerweise mit dem pH-Wert des Zellmilieus identisch. Enge pH-Optima mit steilen Flanke n können ein Mittel zur Regulation der Enzymaktivität durch kleine pH-Verschiebun gen in der Zelle darstellen. Ein bekanntes Beispiel ist die lichtabhängige pH-Verschiebung im Chloroplastenstroma von pH 7 (im Dunkeln) nach pH 8 (im Licht). Zur Bestimmung des pH-Optimums eines Enzyms ist es nötig, dass immer Substra tsättigung vorliegt, da sich in vielen Fällen der K M-Wert mit dem pH verschiebt.
c. Cofaktoren: Coenzyme und prosthetische Gruppen (“prosthetisch” = angefügt) Bei vielen Enzymen hängt die Funktion nur vom Enzympro tein ab, viele benötigen aber zur spezifischen Katalysatorwirkung einen zusätzlichen Cofaktor. Sind die Cofaktoren fest an ein Enzymmolekül gebunden, so spricht man von ihnen als prosthetische Gruppe, wie z. B. Metallionen oder das Flavin der Succinat-Dehydrogenase. Sind sie dissoziabel und somit zwische n mehreren Enzymen austauschbar, so definiert man sie als Coenzyme (z. B. Coenzym A bei der Pyruvatdecarboxylierung und Citratsynthese). Das Co faktoren benötigende Enzymsystem (Holoenzym) besteht also entweder aus:

25
Prosthetischer Gruppe plus Apoenzym (= 1 Molekül) oder au s Coenzym und Apoenzym (= 2 Moleküle). Cofaktoren sind keine Katalysatoren sondern Co-Substrate, denn sie werden bei der Reaktion verändert (z. B. NAD + → NADH + H+).
2. Untersuchung von Enzymen
a. Extraktion des Enzyms Um ein Enzym zu untersuchen, muss dieses aus den Zellen (Alg en, Hefe) oder Geweben extrahiert werden. Bei wasserlöslichen Enzymen g eschieht dies durch Aufschluss des biologischen Materials in Wasser oder geeignet en Pufferlösungen, wobei zuerst die strukturelle Intaktheit der Zelle aufge löst werden muss (z. B. mit Mörser, Ultra-Turrax, French Press oder Ultraschall). Das entstehende sog. Homogenat wird dann abzentrifugier t um unlösliche Zellbestandteile vom löslichen Extrakt (= Rohextrakt) zu trennen. Viele membran-gebundene, d. h. nicht lösliche Enzyme können jedoch nur unter Verwendung von Detergentien (z. B. Triton der Na-Dodecylsulfat: SDS) solubilisiert werden.
b. Enzym-Test Für den Nachweis der Enzymeigenschaft eines Proteins stellt man sich ein geeignetes in-vitro-System zusammen, welches das Substrat ( die Reaktanten) der Reaktion und, falls nötig, Cofaktoren bei günstigen pH- und Temperaturbedingungen enthält. Die Zugabe von Enzym oder eines das Enzym enthaltenden Extraktes katalysiert dann die Reaktion, wo bei das gebildete Produkt als Beweis für die abgelaufene Reaktion gilt. Um sicher zu sein, dass die Reaktion tatsächlich durch das Enzym katalysiert wurde, muss eine Kont rollprobe mit denaturiertem (auf 100 °C erhitztem) Protein angesetzt werden. Es darf dann, zumindest bei reinen Enzympräparaten, keine Reaktion stat tfinden. Da Enzyme in der Regel sehr spezifisch sind, ist es zunächst gleichgültig, o b der verwendete Proteinextrakt noch andere Enzymaktivitäten enthält.
c. Amylase Zur Untersuchung eines Enzyms wird im Praktikum die Amylase verwendet. Amylasen (Amylum = Stärke) sind hydrolytische Enzyme, die glucosidische α1-4-Bindungen in der Stärke spalten. Im Gegensatz zu Glucosida sen setzen Amylasen als kleinstes Endprodukt Maltose frei. Man unterscheidet 2 Typen von Amylasen:
1. αααα-Amylasen, die das Polyglucan an verschiedenen Stellen, d.h. mehr oder weniger zufällig attackieren. Da sie die Stärke von inne n heraus spalten, nennt man sie Endoamylasen. Endprodukt des Abbaus von α-Amylase ist αααα-Maltose (und etwas Glucose), beim Abbau von Amylopectin wird ne ben der α-Maltose auch Isomaltose (Glucose α1-6-Glucosid) frei.
2. ββββ-Amylasen, die das Polyglucan vom reduzierenden Ende her abbauen und dabei ββββ-Maltose freisetzen. Beim Abbau von Amylopektin werden alle Ke tten

26
bis zur ersten α1-6-Bindung gespalten. Der nicht abbaubare, meist hochpolymere Rest bildet das sog. β-Grenzdextrin.
Neben den beiden Typen der Amylasen gibt es noch spezifi sche, die α1-6-Bindungen in verzweigten Homoglucanen spaltende Enzyme: sog. “Debranching” Enzyme, deren bekanntestes das R-Enzym ist. Sie produzieren aus de m verzweigten Homoglucan die unverzweigte Amylose oder hö here Oligodextrine. Der Nachweis der Amylase-Aktivität kann auf zweierlei Wei se geführt werden: 1. durch den Nachweis des Verschwindens der Stärke, geführt mit Hilfe der Jod-
Stärke-Reaktion, und 2. durch den Nachweis der bei der Hydrolyse entstehenden reduzierenden
Gruppen (bei vollständigem Abbau: Halbacetalgruppe de r Maltose). Von der Theorie der Enzymkatalyse her gesehen, ist der N achweis der reduzierenden Gruppen vorzuziehen, da er das Produkt der enzymatischen Reaktion erfasst.
Amylasen sind ubiquitär verbreitet: Bekannt sind die Sp eichel- und Bauchspeichelamylasen des Menschen, wichtig sind aber auch di e Amylasen in keimenden Samen (Getreide), in Früchten (Bananen) und in Blättern (nächtlicher Abbau der Assimilationsstärke). In der Regel kommen die A mylasen aber nicht einzeln vor, sondern immer als Gemisch der verschiedenen T ypen. Im Praktikum wird ein Proteinextrakt aus keimenden Weizensamen verwen det: In ihm finden sich α- und β-Amylase sowie das “Debranching” Enzym.
Aufgabe: Nachweis der Amylaseaktivität und Optimier ung des Testsystems sowie Bestimmung des pH-Optimums. Ausführung im Überblick a) Extraktion der Weizenkörner und Herstellung des Rohex trakts. b) Optimierung des Testsystems, d.h. Bestimmung der günsti gsten Enzymmenge
(hierzu ist eine Eichkurve für den Nachweis reduzierender Gruppen mit dem DNSS-Reagenz zu erstellen).
c) Bestimmung des pH-Optimums.
a) Extraktion der Amylase aus Weizenkörnern: Vorbemerkung: Amylase ist ein recht “robustes”, lösliches Enzym, das durch Aufschluss von Weizenkeimlingen in Wasser ohne Puffer- oder sonstige Schutzsubstanzen gewonnen werden kann. Durchführung: 2 Keimlinge (Weizen oder Gerste) werden von Wurzeln u nd Koleoptilen befreit und mit 10 ml dest. Wasser und ein er Spatelspitze Sand in einer Reibschale zerrieben. Zunächst soll das Mörsern mit ca. 3 ml Wasser begonnen werden und erst bei schon weitgehender Zerstörung der K örner die übrigen ca. 7 ml Wasser nach und nach hinzugefügt werden. Der Brei wir d durch 6-fach gelagerten Mull in ein kleines Becherglas abfiltriert u nd dann in ein Zentrifugenglas

27
überführt. Mit einem zweiten, wassergefüllten Zentrifu genglas wird das Homogenat auf der Waage austariert und in der Tischzentrifuge 10 min. lang bei höchster Geschwindigkeit abzentrifugiert. Der Überstand ist der Pr otein-Rohextrakt. Er wird für die weiteren Versuche verwendet und im Eisbad aufb ewahrt.
b) Optimierung des Testsystems Die Bestimmung der amylolytischen Aktivität geschieht in 2 Schritten: a) Spaltung von Amylopektin (Präparat: “Lösliche Stärke nach Zulkowski”) mit
einer definierten Menge Enzympräparat bei definierte m pH-Wert (Puffer) und definierter Temperatur (Wasserbad);
b) Abbruch der enzymatischen Reaktion durch Zusatz des DNSS- Reagenz’, anschliessende Reaktion der reduzierenden Gruppen mit DNS S (in der Hitze), sowie Quantifizierung (nach Abkühlen) mittels Spektralpho tometrie.
Vom Rohextrakt wird eine Verdünnungsreihe hergestellt (gut mischen): unverdünnt 1:10 1:20 1:30 1:40 1:50
Rohextrakt (ml)
dest. Wasser (ml)
x
0
1
9
0.5
9.5
0.3
9.7
0.25
9.75
0.20
9.80
Von der Verdünnung 1:10 werden 5 ml 5 min. lang im kochenden Wasserbad denaturiert; der Überstand dieser Lösung dient als Kontrolle. Zur Ermittlung der günstigsten Enzymkonzentration für di e Ermittlung des pH-Optimums wird der Test nach folgendem Schema in 10 ml R eagenzgläsern aufgebaut: Ansatz-Nr. 1 2 3 4 5 6 7
1% Stärkelösung, mit Phosphat-
puffer (0.2 M, pH 5,2) 1:2 verdünnt
1.5
ml
1.5
ml
1.5
ml
1.5
ml
1.5
ml
1.5
ml
1.5
ml
Temperaturangleich der Proben im 37 °C Wasserbad (ca. 3 -5 min., gelegentlich schütteln)
0.5 ml Rohextrakt
bzw. Verdünnung
bzw. gekochter Rohextrakt
+
1:10
1:20
1:30
1:40
1:50
+
10 min. Amylolyse bei 37°C
Stop mit 2 ml DNSS-Reagens, Reagenzgläser mit Alu-Fol ie verschliessen
5 min (genau) ins kochende Wasserbad
Abkühlen auf Zimmertemperatur
Extinktion bei 540 nm gegen Ansatz Nr. 7 messen

28
Gleichzeitig kann die Maltose-Eichkurve (siehe unten) erst ellt werden. Das Versuchsergebnis wird in einem Diagramm (µmol Maltose gegen Verdünnung) dargestellt.
Der DNSS-Test Bei der amylolytischen (= hydrolytischen) Spaltung der Stärke werden reduzierende Gruppen freigesetzt. Diese lassen sich mittels 3,5-Dinitrosalicylsäure quantifizieren: Das Halbacetal reduziert eine der Nitro gruppen zur Aminogruppe:
Die Nitroaminosalicylsäure besitzt ein von der Dinitrosal icylsäure verschiedenes Absorptionsmaximum bei 540 nm. Da das Endprodukt des amylolytischen Stärkeabbaus Maltose i st, wird die Eichkurve mit diesem Disaccharid erstellt.
Maltose-Verdünnungsreihe für die Eichkurve Es wird eine 3 mM Maltose-Lösung gestellt. Davon wird f olgende Verdünnungsreihe (je 10 ml) hergestellt ( nach dem Pipettieren gut mischen! ).
Maltose-Lösung (ml) 4 3 2 1,5 1 0,5 0
dest. Wasser (ml) 6 7 8 8,5 9 9,5 10
Konzentration (mM) 1,2 0,9 0
µmol in1 ml 1,2 0,9 0
c) pH-Optimum Für die Bestimmung des pH-Optimums der amylolytischen St ärkespaltung wird eine stättigende Substratkonzentration, eine optimale E nzymkonzentration und eine mittlere Temperatur (37°C) gewählt. Als optimal e Enzymkonzentration wird diejenige Konzentration ausgewählt (evtl. aus dem Diag ramm extrapoliert), die etwa ein
�E von 0,8 ergibt. Die Phosphatpuffer im Bereich zwischen pH 3,6 und
7,8 werden gestellt. Der Test für die Ermittlung des optimalen pH-Werts wir d folgendermassen aufgebaut:

29
Ansatz-Nr.: 1 2 3 4 5 6 7 8 9 10
0,75 ml Phosphatpuffer pH 3,6 4,5 5,0 5,2 5,2 5,4 5,8 6,5 7,0 7,8
0,75 ml 1% Stärkelösung + + + + + + + + + +
Temperaturangleich im 37 °C Wasserbad, Reagenzgläser mit Alu-Folie
verschliessen
0,5 ml optimal verdünnter
Rohextrakt bzw.
gekochter Rohextrakt
+ + + +
+
+ + + + +
10 min (genau) Amylolyse
Stop mit 2 ml DNSS-Reagens, Reagenzgläser mit Alu-Foli e verschliessen
5 min. (genau) ins kochende Wasserbad
Abkühlen auf Zimmertemperatur
Extinktion bei 540 nm gegen Ansatz Nr. 5 messen
Zur Auswertung wird ein Diagramm mit
�E gegen pH-Wert erstellt und der
optimale pH-Wert festgestellt.
Fragen zur Auswertung: 1. Warum kann man mit dem DNSS-Test den Stärkeabbau du rch Amylase quantifizieren ? 2. Warum wird für den Versuch zur Bestimmung des pH-Opti mums gerade eine Verdünnung des Rohextrakts gewählt, die im Teilversuch a ) eine Extinktion von 0,8 ergibt ? (Wozu dient der Teilversuch a) überhaupt ? ) 3. Was ist die Ursache für eine Änderung der Enzymaktivit ät bei einer Änderung des pH- Wertes ?

30
V. Die Steuerung der pflanzlichen Entwicklung durch Hormone und die Bewegungen von Pflanzen Steuerung der pflanzlichen Entwicklung durch Hormon e Die Entwicklung einer höheren Pflanze aus einer einzigen Zelle, der Zygote, zu einem komplizierten, vielzelligen System, das sich schliesslich fortzupflanzen vermag, besteht aus einem komplexen Zusammenspiel von Wa chstums- und Differenzierungsprozessen. Die heutige Entwicklungsphysiologie spricht von einer durch das genetische Material gesetzten Reaktionsnorm, inne rhalb derer äussere und innere Faktoren regulierend auf Wachstum und Diffe renzierung einwirken können. Die Steuerung der Entwicklung durch innere Fakto ren, die im Organismus selbst vorhanden sind, umfasst eine Regulation sowohl auf intra- als auch auf interzellulärer Ebene. Die interzelluläre Regulation, die eine Koordinierung der Regulationsprozesse in den verschiedenen Zellen und Teile n des Organismus bewirkt, erfolgt in Pflanzen über das Leitbahnsystem, wo bei die pflanzlichen Hormone (Phytohormone) eine dominierende Rolle spiel en. Der Ablauf der Entwicklungssteuerung durch die inneren Faktoren wird abe r letztlich von den Aussen- oder Umweltfaktoren bedingt, die somit einen ma ssgebenden Einfluss auf das Wachstum und die Differenzierung der Pflanze ausüben.
In diesem Praktikum sollen einige Beispiele der pflanzlich en Steuerung durch Phytohormone untersucht werden.
Wie alle Hormone sind auch die Phytohormone meist (wen n auch nicht immer) in geringen Mengen wirksame Stoffe, bei denen Bildungsort und Wirkungsort voneinander verschieden sind. Der Transport vom Bildung sort zum Wirkungsort erfolgt im Leitbahnsystem der Pflanze. In einer Reihe von Fällen allerdings werden die Phytohormone auch am Bildungsort selber wirksam. Phy tohormone zeigen meist ein sehr breites Wirkungsspektrum; ein und dasselbe Ph ytohormon vermag eine Vielzahl gänzlich verschiedener Prozesse zu beeinflussen. So ist z. B. Auxin (IES) an der Zell- und Internodienstreckung, der Anlag e von Seitenwurzeln, der apikalen Dominanz, der Verhinderung von Blatt- und Fr uchtfall und der Fruchtentwicklung beteiligt. Die fünf wichtigsten Gruppen von Phytohormonen sind: 1. die Indolderivate (Auxine), 2. die Gibberelline, 3. die Cytokinine, die in der Regel Wachstumsprozesse fördern , sowie 4. Abszisinsäure und 5. Ethylen, die eine vorwiegend hemmende Wirkung auf Wachstumsprozesse ausüben. Vom Konzentrationsverhältnis zwischen Abszisinsäure und Ethyl en einerseits und den anderen drei Phytohormonen hängt es ab, ob ein b estimmter Entwicklungsschritt ablaufen kann oder nicht.

31
Hormone kommen in der Pflanze nur in sehr geringen Men gen vor; so enthält z. B. eine Haferkoleoptile nur etwa 10 -10 Mol des Streckungshormons Auxin ( β -Indolylessigsäure). Für den Nachweis und vor allem für d ie quantitative Bestimmung ist deshalb eine aufwendige chemische Analytik vonnöten. Einfacher ist die Quantifizierung über einen sog. Biotest, dessen P rinzip darin besteht, dass man an lebenden, auf das Hormon ansprechenden Objekten eine spezifische Reaktion misst. Die Stärke der Reaktion wird dann anhand einer vorher aufgestellten Eichkurve (Reaktion auf das entsprechende Ho rmon in bekannter Konzentration) in Hormonkonzentration umgerechnet. Ein solcher Biotest bietet noch einen weiteren Vorteil: Man kann damit prüfen, o b synthetisch hergestellte Stoffe die gleiche Reaktion hervorrufen wie das pflanze neigene Hormon, und darüberhinaus auch noch, von welchen Faktoren die untersu chte Reaktion abhängt.
A. Einfluss von Auxin (IES) auf das Streckungswachs tum von Getreidekoleoptilen Als Testobjekte verwendet man verständlicherweise solche Pf lanzenteile, die auf das zu untersuchende Hormon mit einer möglichst starken Rea ktion ansprechen. Beim Auxin ist das klassische und auch heute noch meistverwen dete Objekt die Koleoptile (Keimscheide) von Gräsern (vorwiegend von A vena = Hafer) in etioliertem Zustand.
Bei diesen Koleoptilen wird das Auxin in der Spitze pr oduziert, wandert nach unten und löst dort ein starkes Streckungswachstum aus. Entfernt ma n die Spitze, dann reagiert das übrige Koleoptilgewebe wegen Mangels an eigenem Auxin sehr empfindlich auf exogenes (von aussen zugegebenes) Auxin. Das Ausmass des Streckungswachstums kann man auf zwei Arten messen : Man führt dem Koleoptilstumpf Auxin einseitig zu, löst damit an dieser Flanke Zellstreckung aus, was zu einer Krümmung in Richtung der e ntgegengesetzten Flanke führt. Die Grösse des Krümmungswinkels dient als Mass für die Auxinmenge (WENT'scher Krümmungstest). Dieser Test ist zwar die genaueste Methode, ist aber relativ schwierig auszuführen. Einfache r, aber etwas weniger empflindlich ist der Zylindertest, bei dem die Längenzunahme eines abgeschnittenen Koleoptilzylinders in einer Hormonlösung gemessen wird.
Indol-3-essigsäure (IES, Auxin)

32
Ausführung: Im Dunkeln angekeimte Hafer- oder Weizenkörner werden a us der Anzuchtschale entnommen (am Korn anfassen, um die Koleoptile nicht zu sehr zu drücken ) und mit einer Rasierklinge die Koleoptilenspitze dekapitiert (von der Spitze her 2-3 mm abschneiden). Der Stumpf wird dann auf eine Glasplatte mit Raster (Millimeterpapier) gelegt und genau 10 mm hinter de r Schnittfläche mit der Rasierklinge eingeritzt. Durch mehrmaliges vorsichtiges Bie gen wird die Koleoptile vom Korn abgebrochen und von dem in ihr eingeschlossenen Primärblatt abgezogen. Je 15 dieser Zylinder werden in Erlenmeyerkolben in je weils 10 ml bidest Wasser und 10-2 M, 10-4 M und 10 -7 M IES-Lösung gebracht und auf der Schüttelmaschine über Nacht ständig in langsamer Bewegung gehalten. Am nächsten Tag wird der Zuwachs der einzelnen Zylinder auf den Glasplatten mögl ichst genau gemessen und in eine Tabelle eingetragen.
Auswertung:
Berechnen Sie die Mittelwerte und tragen Sie die Aux in-Konzentration in einem Diagramm gegen den Längenzuwachs auf.
Diskutieren Sie das Ergebnis unter folgenden Gesichtspunkte n:
- Beschreiben Sie kurz den Mechanismus des Wachstums des Pflanze ngewebes, aufgrund dessen die Längenzunahme stattfindet.
- Wachsen die Koleoptilen auch in bidest Wasser ? Warum ?
- Was für eine Kurve ergibt sich aus der Auftragung der Auxinkonzentration gegen den Längenzuwachs ? Kurze Begründung und Interpretation !
B) Induktion der αααα-Amylase-Synthese durch Gibberellinsäure in Getreidekörnern
a. Allgemeines Phytohormone (z. B. IES, Gibberellinsäure, Kinetin u. a.) haben multiple Wirkungen und führen zu einer Veränderung der genetischen Aktivit ät der betroffenen Zellen. Zur Erklärung geht man von der Voraussetzung aus, dass es i m Genom neben aktiven und inaktiven auch potentiell aktive und potent iell inaktive Gene gibt. Diese potentiell aktiven oder inaktiven Gene sollen z. B. dur ch Hormone aktiviert oder inaktiviert werden. Dabei wirkt das Hormon also nur als unspezifischer Auslöser, was die multiple Wirkung erklärt. Direkte Folge der Hor monwirkung muss dann die Neusynthese oder der Synthesestop von Messenger-RNA sein, was die Neusynthese oder die Hemmung der Synthese von Proteine n zur Folge hat.
b. Wirkungsweise des Phytohormons Gibberellinsäure (GA3) Die Wirkung eines Phytohormons lässt sich in eindrucksvoller Weise bei der Samenkeimung des Getreides demonstrieren. Die Samen der Gräser (die

33
Karyopsen) bestehen aus dem schon toten, aber stärkereichen Endosperm, das von der lebenden Aleuronschicht umgeben ist bzw. an eine r Seite an das Scutellum (Keimblatt) des Keimlings angrenzt. Bei der Keimung wird die Stärke gespalten; aus der dab ei freigesetzten Glucose und Maltose gewinnt der Embryo die zur Keimung und zum anfänglich Wachstum notwendige Energie sowie seine Baustoffe. Die Keimung wird also erst durch die Stärkespaltung mögl ich; hierfür sind besondere Enzyme, die Amylasen, notwendig, die aber vo r der Keimung keinen nennenswerten Abbau der Stärke verursachen dürfen. Der Samen enthält β -Amylase bereits im ungekeimten Zustand: Dieses Exoenzym al lein kann jedoch die Stärkekörner selbst nicht vollständig abbauen (vergl. Ver such V !). α-Amylase dagegen, die die Stärkekörner vollständig auflösen kann, wird erst bei der Keimung in der Aleuronschicht synthetisiert: Dies wird du rch das vom (gequollenen) Embryo abgesonderte Hormon Gibberellinsäure (GA3) ausgelöst (induziert).
Die Induktion der α-Amylase-Synthese ist für Gibberellinsäure spezifisch und ka nn als äusserst empflindlicher Biotest benützt werden. Neben der Amylase wird durch die Sekretion des Hormons auch die Synthese von Proteasen und Ribonucleasen induziert. Diese Enzyme wandern von der Aleuronschicht in die toten Endospermzellen und hydrolysieren dort das Reservemateri al (Abb. V).
Gibberellinsäure (GA3)
Abb. V: Mobilisierung des Reservematerials im Endosperm von Getreidekörnern durch die Induktion von hydrolytischen Enzymen durch Gibberellinsäure.

34
Die Abbauprodukte der Reservestoffe werden als Malz bezei chnet. Da Gibberellinsäure also (indirekt) die Malzbildung induzie rt, gibt man in der Brauerei heute zusätzlich dieses Hormon zu keimenden Gerstenkörnern da zu, um die Malzausbeute zu erhöhen.
c. Nachweis der Wirkung von Gibberellinsäure
Der Nachweis der Induktion der α-Amylase-Synthese durch das Phytohormon Gibberellinsäure in Weizenkörnern basiert auf folgenden Überlegungen: a) Embryofreie Kornhälften können bei Quellung keine Gibberellinsäure zur Induktion der α-Amylase-Synthese produzieren. Künstliche Zufuhr von Gibberellinsäure zu solchen Kornhälften aber muss zur Indu ktion von α-Amylase-Synthese führen, genau so, wie dies in embryohaltigen Körnern oder Kornhälften stattfindet. b) Es ist zu beweisen, dass α-Amylase neu synthetisiert wird. Dies lässt sich dadurch zeigen, dass man als Kontrollproben Körner benüt zt, die mit einem Inhibitor der Proteinsynthese, z. B. Actidion oder Cyclo heximid, behandelt werden.
Versuchsdurchführung:
1. Schritt: Sterilierisung, Quellung und Inkubation der Weizenkörner Achtung: Dieser Schritt wird von den Assistenten durchgefü hrt. 30 Weizenkörner werden mit einer Rasierklinge äquatori al in zwei Hälften auf einer Glasplatte in eine embryohaltige und in eine embryofreie Hälfte zer schnitten. Jeweils 10 embryohaltige bzw. embryofreie Kornhälften: Ansatz 1) 10 embryohaltige Kornhälften 2) 10 embryofreie Kornhälften 3) 10 embryofreie Kornhälften 4) 10 embryofreie Kornhälften werden in bezeichnete 100 ml-Bechergläser eingefüllt: Jeder Ansatz wird 10 Min. lang mit 50 ml 0,5% Chinosol -Lösung oberflächensterilisiert. Dann wird die Chinosol-Lösung abdekantiert und jeder Ansatz 3 x mi t jeweils ca. 50 ml bidest. Wasser gewaschen. Nach Abdekantierung der letzten Waschwasserportion wird j eder Ansatz von Kornhälften noch mit ca. 20 ml der entsprechenden Inkubationslösung gewaschen: Ansatz 1) bidest Wasser 2) bidest Wasser 3) 10 -6 M GA3-Lösung (99 ml bidest Wasser + 1 ml 10 -4 M GA3-Lösung). 4) 10 -6 M GA3-Lösung (98 ml bidest Wasser + 1 ml 10 -4 M GA3-Lösung) + 1 ml Cycloheximid-Lösung (2 mg/ml) Dann werden die Ansätze in ca. 50 ml der entsprechenden Inkubationslösung bei 4°C im Kühlschrank über Nacht gequollen. Am nächsten Tag wird di e Quellungsflüssigkeit von den Kornhälften abdekantiert. 4 PVC-Petrischalen werden jeweils mit 8-lagigem Zellsto ff ausgekleidet und mit so viel der entsprechenden Inkubationslösung getränkt, dass die Flüssigkei t gerade noch in den tiefsten

35
Dellen des Papiers frei steht (das Papier soll also sehr n ass, aber nicht überflutet sein). Die 10 Kornhälften von jedem der 4 Ansätze werden dann so in die Petrischalen eingebracht, dass die Schnittflächen nach oben in die Luft ragen. Die verschlosse nen Petrischalen werden nun bis zum nächsten Tag bei 30°C im Brutschrank inkubiert. Hier beginnt die Praktikumsaufgabe:
2. Schritt: Extraktion der Kornhälften (Herstellung des Rohextrakts) Die 10 Kornhälften eines Ansatzes werden in einer Reibsch ale mit etwas Sand und 10 ml dest. Wasser zu einem möglichst homogenen Brei zerri eben. Das Mörsern wird mit 3 ml Wasser begonnen, und erst nach weitgehend er Zerstörung der Kornstruktur werden weitere 7 ml zugesetzt. Das Homogenat wird durch 4-fachen Mull filtriert (Mull auswinden!) und anschliessend 10 Mi n. lang in der Tischzentrifuge bei maximaler Geschwindigkeit abzentrifugiert. Der Überstand ist der jeweilige Rohextrakt und wird für die weiteren U ntersuchungen verwendet.
3. Schritt: Nachweis der Amylasen: Reaktion im Stär ke-Agar Stärkehaltige Agarplatten (2 % Agar, 1 % lösliche Stär ke, 0,1 % CaCl2 in PVC-Petrischalen) werden gestellt. Mit Hilfe eines Korkbohre rs werden 6 gleichmässig verteilte Löcher aus der Agarschicht ausgestanzt und auf de r Rückseite der Petrischale numeriert. In jedes Loch wird nun der Rohex trakt, α- oder β-Amylase (vom Assistenten folgendermassen vorbereitet: 10 µl Enzym vo n Boehringer in 10 ml K-Phosphatpuffer pH 5.8) nach dem folgenden Schema pipettiert (jeweils 150 µl):
Loch 1 Rohextrakt vom Ansatz 1) 2 " " " 2) 3 " " " 3) 4 " " " 4)
5 α-Amylase 6 β-Amylase Es wird eine derartige Platte hergestellt und im Brut schrank bei 30°C inkubiert. Nun können die im Rohextrakt gelösten Amylasen in die A garschicht hineindiffundieren und die im Agar deponierte Stärke abbauen. Am nächsten Versuchstag wird die Platte dem Brutschrank entnommen und angefärbt: Die Agarschicht wird mit Jod-Jod-Kalium-Lösung übergossen u nd 3 min stehengelassen. Dann wird die JJK-Lösung abdekantiert, die Agar-Oberfläche kurz mit Leitungswasser gespült und die Platte auf dem Overh eadprojektor betrachtet: Noch in dem Agar befindliche Stärke wird durch Einlager ung von Jod in die Stärkespirale gefärbt, wobei intakte Stärke dunkelviolet t und β-Grenzdextrin (nur β-Amylase) rosa erscheint. Da das eingelagerte Jod relativ schnell sublimiert, ist es wichtig, dass alle Farbnuancen gleich nach dem Anfärben t abellarisch notiert werden! Dabei soll auf das Ausmass und die Farbqualität der Entf ärbungszonen geachtet werden.

36
Fragen fürs Protokoll:
1. Was sind die Endprodukte beim Stärkeabbau durch α-Amylase bzw β-Amylase? 2. Warum kann aus dem Versuch geschlossen werden, dass α-Amylase erst neu synthetisiert wird und β-Amylase bereits vorhanden ist ?
C. Gemeinsame Demonstrationsversuche: Bei den folgenden Versuchen 1 und 2. handelt es sich um gemeinsame Demon-strationsversuche. Dafür erhält jede Gruppe zwei Coleus-Pflanzen (Buntnessel) zur Betreuung und beginnt am ersten Versuchstag mit den Versuchen 1 und 2. An jedem darauffolgenden Versuchstag werden die Veränderu ngen mit Hilfe einer Skizze und den nötigen schriftlichen Notizen protokolliert und am Ende des Praktikums in einem eigenen Protokoll zusammengestel lt und interpretiert .
1. Erhaltung der Apikaldominanz durch Auxin Vom apikalen Ende des Hauptsprosses einer Pflanze gehen He mmwirkungen aus, welche das Austreiben der Seitenknospen unterdrücken ("api kale Dominanz"). Schneidet man den Hauptspross ab, so entwickeln sich aus den nächstgelegenen Seitenknospen Seitentriebe. Trägt man aber auf die Sch nittstelle Auxin (IES) auf, so bleibt das Wachstum der Seitenknospen weiterhin gehemm t. Offensichtlich ist das Auxin in irgendeiner Weise an der Apikaldominanz b eteiligt ist. Vermutlich wirkt IES aber nicht direkt, sondern induziert im Seiten knospenbereich die Bildung von Ethylen, das seinerseits das Austreiben dieser Knospen unterdrückt.
Ausführung: Die Sprosse von zwei 2- bis 3 Wochen alten Coleus-Pflanzen (Primärblätter bereits entwickelt) werden etwa 5 cm über dem Ansatz der Primärblätter dekapitiert. Bei einer dieser Pflanzen wird dann ein kl eines Reaktionsgefäss mit Wuchsstoffpaste (1 ml 10 -2 M IES vermischt mit 10 g Wollfett) auf die Schnittstel le (Stumpf) fest aufgesetzt. Da IES lichtempfindlich ist, mu ss das Gefäss gut mit Alufolie eingewickelt sein ! Die zweite Kontrollpflanze erhält ein Gefäss mit wuchsstofffreier Wollfettpaste (mit dest. Wasser statt IES- Lösung). Die Gefässe werden täglich von den Assistenten erneuert, wobei jewe ils vorher der Stumpf um etwa 5 mm gekürzt werden muss.
2. Vielfalt der Wirkung von Auxin auf eine Pflanze Wie alle Phytohormone übt Auxin eine multiple Wirkun g auf pflanzliche Entwicklungsprozesse aus. Neben der schon untersuchten Förderu ng des Streckungswachstums (Versuch 1) kann Auxin auch Wachstumskrümmung en und Wurzelbildung auslösen, und Blattfall verhindern. Wachst umskrümmungen kommen durch die Förderung eines lokalen Streckungswachstum s zustande.

37
Die Bildung von Adventiv- und Seitenwurzeln geht von bestimmten Zellteilungsnestern (sekundäre Meristeme) aus. Die zur Wur zelbildung führenden Zellteilungen werden von Auxin stimuliert, ein Effekt der in der Praxis zur Bewurzelung von Stecklingen angewandt wird. Der Blattfall wird mit der Ausbildung einer durch Zel lteilung entstehenden Trennungszone an der Basis eingeleitet und vollzieht sich nach enzymatischem Abbau der mechanisch stabilisierenden Zellwandkomponenten. Die genannten drei Wirkungen können an Coleus-Pflanzen demonstriert werden.
Ausführung: Verschiedene Teile von Coleus-Pflanzen werden mit Wuchsstoffpaste (IES-Wollfettpaste wie bei Demonstrationsversuch 1 bzw. wuchsstof ffreier Wollfettpaste (mit Wasser statt IES-Lösung) behandelt. a) Wachstumskrümmung: Junge Stengelteile (Internodien) werden mit IES-Past e bestrichen. b) Wurzelbildung: Basale Stengelteile (Interodien) werden mit IES-Past e bestrichen. c) Verhinderung des Blattfalls: Einige Blattspreiten werden abgeschnitten und IES- Paste bzw. IES-freie Paste (zur Kontrolle) au f die noch auf dem Stengel stehenden Blattstielstümpfe gestrichen.
Die Bewegungen von Pflanzen Abgesehen von den Ortsbewegungen freischwimmender Algen und von Zell-organellen, die man, sofern sie auf einen Reiz hin erf olgen, als Taxien bezeichnet, unterscheidet man bei festgewachsenen Pflanzen Bewegungen , deren Richtung von der Richtung des Reizes unabhängig ist, die Nastien, von den Bewegungen, deren Richtung in einem Zusammenhang mit der Reizrichtu ng steht, den Tropismen.
1. Nastien: Die Nastien sind in der Regel turgorabhängige Bewegun gen, bei denen infolge kurzfristigen Zusammenbruchs des Turgors einer Gewebepartie (Extensor) der volle Turgor des gegenüberliegenden Gewebes (Flexor) das Pflanzenorgan in eine Richtung biegt. Man hat herausgefunden, dass der Turgor zusammenbruch durch K+-Export aus den Extensorzellen hervorgerufen wird. Die Rückbewegung erfolgt durch Erholung des Turgors im Extensor aufgrund von K +-Aufnahme. Die Bewegung erfolgt in Gelenken oder Blattkissen, Pulv ini (Pulvinus) genannt, die eine besondere anatomische Struktur aufweisen. Als Beispie l ist die Struktur des Gelenks am Fiederblättchen von Albizzia in Abbildung V/2 dargestellt.

38
Abb V/2: Darstellung der Blattkissen (Pulvinus) bzw. Fl exor / Tensor an einem Albizzia Blatt HR = Hellrotes Licht, DR = Dunkelrotes Licht (Quelle : Lüttge U., Kluge & Bauer) Die Blätter vieler Dikotylen, z. B. die vieler Fabaceen, führen sog. Schlafbewegungen (Nyctinastie) durch. Dabei senkt sich der Blattstiel am Abend

39
und richtet sich morgens wieder auf. Ähnliches beobachtet man bei den primären und sekundären Fiedern vieler Caesalpiniaceen, z. B. Albizzia, und Mimosaceen, z. B. Mimose: Sie schliessen sich in der Dunkelperiode und öffnen sich b ei Tag. An den Fiederblättern von Albizzia lässt sich verhältnismässig einfach zeigen, dass die Bewegung vom Phytochromsystem gesteuert wird. Manche Pflanzen besitzen eine sogenannte endogene Rhythm ik, die man beobachten kann, wenn man sie nach einem Licht-Dunkel-Wech sel ins Dauerdunkel oder Dauerlicht bringt. Die rhythmischen Sch lafbewegungen gehen weiter, allerdings flacht die Amplitude allmählich ab und die Phase wird nicht mehr genau eingehalten. Man muss annehmen, dass in diesen Pfl anzen ein Steuerungssystem oszilliert, das durch den natürlichen Licht -Dunkel-Wechsel immer wieder neu angestossen und hinsichtlich der Phase ju stiert wird.
Aufgabe: Untersuchung der Regulation der photonasti schen Blattbewegung bei Albizia durch Phytochrom Die Regulation der photonastischen Blattbewegung bei Albizia erfolgt durch das Phytochrom. Das Phytochrom kann in zwei unterschiedlichen, relativ stabilen Formen vorliegen, die reversibel ineinander überführ bar sind: Als physiologisch aktive P730-Form nach Bestrahlung mit hellrotem Licht und als physi ologisch inaktive P660-Form nach Bestrahlung mit dunkelrotem Licht. Werden die Pflanzen mit Tageslicht oder hellrotem Licht bestrahlt und anschliessend ins Dunkle gebracht, so schliessen sich die Fiedern . Bestrahlt man jedoch die Blätter unmittelbar vor dem Verdunkeln 4 m in. lang mit dunkelrotem Licht, so bleiben die Fiedern im Dunkeln geöffnet. Die ser Effekt des dunkelroten Lichts kann mit hellrotem revertiert werden. Daraus kann man ableiten, dass sich die Fiedern schliessen, wenn das System zu Beginn der Dunke lphase hauptsächlich in der Form P 730 vorliegt, und dass sie geöffnet bleiben, wenn zu Beginn der Dunkelphase P660 dominiert. Die Schlafbewegungen lassen sich allerdings mit dem Phytochromsystem allein nicht erklären , denn P730 (das durch Bestrahlen mit Hellrot- oder Tageslicht erzeugt wird) w irkt nur, wenn die Blätter ins Dunkle gebracht werden. Man muss also annehmen, dass zum Sch liessen der Fiedern zwei Bedingungen erfüllt sein müssen:
1. Eine ausreichende Konzentration P 730 und 2. einen weiteren Mechanismus, der in der Pflanze im Ta geslicht den P730 Effekt überdeckt und erst bei Dunkelheit die Wirkung zeigen lässt (man hat mittlerweile diesen Lichtrezeptor identifiziert, es handelt sich um ein en Blaulichtrezeptor). Als Nachweis einer Regulation durch Phytochrom dient die Revertierbarkeit eines z. B. durch Hellrotbestrahlung ausgelösten Effekts durch Be lichtung mit dunkelrotem Licht und vice versa. Deshalb muss zu den einfa ch bestrahlten Ansätzen auch ein solcher mit Nachbelichtung durch die inakt ivierende bzw. aktivierende Lichtqualität erfolgen: z. B. Belichtung m it Hellrot, Nachbelichtung mit Dunkelrot.
Junge Topfpflanzen von Albizzia, die vorher im Langtag gehalten wurden, werden im Praktikumsraum vor Beginn des Praktikums etwa 3 Std. la ng dunkel gestellt. Im Praktikum werden sie dann den folgenden Belichtungen un terworfen:

40
Pflanze Nr. Behandlung 1 4 min. Hellrot 2 4 min. Dunkelrot 3 4 min. Hellrot + 4 min. Dunkelrot 4 4 min. Dunkelrot + 4 min. Hellrot 5 Tageslicht 6 Dunkel
Die Pflanzen 5 und 6 sind Kontrollpflanzen. Der Effekt wird nach 30 und 60 Minuten notiert.
2. Tropismen Die Tropismen sind Bewegungen, die entweder zum Reiz hi n (positiver Tropismus) oder vom Reiz weg (negativer Tropismus) gerichtet sind o der senkrecht zum Reiz verlaufen (Dia- oder Plagio-Tropismus). Tropismen sind praktisch ausschliesslich Wachstumsbewegungen. Verstärktes oder reduziertes Wachstum au f einer Seite des Pflanzenorgans führt nach dem Bimetallprinzip zur ein seitigen Krümmung. Dieses unterschiedlich starke Wachstum ist möglicherweise eine Folge ungleicher Wuchsstoff-Konzentrationen (z. B. IES) in den beiden Fla nken. Die verbreitetsten Tropismen sind Phototropismus und Gravitropismus, welche zu einer besseren Ausnutzung des belichteten Raums und einer besseren Veranke rung der Pflanze im Substrat führen. Besonders günstige Objekte für das St udium dieser Tropismen sind Keimlinge. Belichtet man einen Keimling einseitig mit Weiss-, Blau- oder UV-Licht, so krümmt er sich nach kurzer Zeit dem Licht zu (positi ver Phototropismus). Beim Gravitropismus reagiert der Spross negativ, die Wu rzel positiv auf den Reiz Schwerkraft, d.h. der Spross wächst senkrecht gegen die Richt ung, die Wurzel in Richtung der Schwerkraft. Wie der Phototropismus ist auch der Gravitropismus ein Induktionsphänomen, d.h. dass unter Umständen einige Min uten der Induktion (z. B. Horizontallage eines gestreckten Keimlings) ausreichen, um die gravitropische Aufkrümmung des Sprosses bzw. Abwärtskrümmung der Wurzel zu i nduzieren. Reaktionsort ist die Zone unterhalb des Meristems, also di e Streckungs- und Differenzierungszone. Der Ort der Reizaufnahme ist, wie beim Phototropismus, ein Meristem (Sprossscheitel, Wurzelhaube). Man weiss, dass sich bei einer gravitropischen Reizung des Meristems die Richtung des Drucks von Zellorgane llen (u.a. Stärkekörner) auf ihre Unterlage, z. B. auf das ER, ver lagert. Dies könnte dazu führen, dass allein aufgrund räumlicher Behinderung da s Zellwandwachstum an den Flanken des Organs verschieden schnell abläuft, doch spi elen sicher auch komplexere Vorgänge eine Rolle, z. B. Ca 2+-Freisetzung, Wuchsstoffe etc. Als reizauslösende Organellen kommen, vor allem im Wurzelber eich, Stärkekörner in der Wurzelhaube in Frage, die sogenannte Statolithenst ärke, die auf schüsselförmigen Ausbildungen des ER liegen.
Aufgabe: Untersuchung der Kinetik der gravitropisch en Aufkrümmung eines Sonnenblumenkeimlings Der Versuch wird in einem dunklen Nebenraum des Praktikum ssaales durchgeführt. Ein Sonnenblumenkeimling wird mit seinem Anzuchttöpfchen mit Hilfe einer Schelle so an einem Stativ fixiert, dass der Spross waage recht liegt. Über die

41
Koleoptilen wird vorsichtig ein grosser Plastiktrichter ge stülpt, der mit feuchtem Zellstoff ausgekleidet ist, um eine möglichst hohe Luftfe uchtigkeit um die Keimblätter herum zu gewährleisten. Hinter den Keimli ng wird eine Glasplatte aufrecht stehend fixiert, auf deren Aussenseite mit Tesaf ilm ein Transparentpapier geklebt wird. Durch eine Lichtquelle hinter dem Keimli ng wird auf dem Papier ein Schattenbild des Keimlings erzeugt, das mit einem weichen Bleistift 2 Stunden lang in Abständen von 30 min nachgezeichnet wird. Die L ichtquelle soll nur für die Zeit der Zeichnung des Schattenbildes angeschaltet sein. Tragen Sie den Krümmungswinkel des oberen Sprossteils des Keimlings gegen die Zeit auf.
Fragen und Überlegungen: 1) Bezüglich der Bewegungsmechanismen unterscheidet man zwi schen Turgor- und Wachstumsbewegungen: Wie ist der Mechanismus des Wachstums einer Pflanzenzelle? 2) Woher weiss man, dass Photosynthese nicht für den Photo tropismus verantwortlich ist? 3) Welchen Nutzen hat die Pflanze von Photo- und Gravi tropismus? 4. An welchem Ort erfolgt die Reizaufnahme bei Photo- bzw. Gravitropismus ? 5. Wozu ist die hohe Luftfeuchtigkeit beim Gravitropismu s-Versuch nötig ?

42
VI. Die Dissimilation Die Dissimilation dient der Freisetzung der zur Aufrechte rhaltung des Stoffwechsels notwendigen Energie. Sie besteht im wesent lichen aus dem exergonen Abbau energiereicher Verbindungen und stell t den Gegensatz zu den energieverbrauchenden Aufbauvorgängen der Assimilation dar. Je nachdem, ob Dissimilation in Gegenwart oder Abwesenheit von Sauerst off abläuft, spricht man von Atmung bzw. Gärung.
a. Atmung Insgesamt gesehen, beginnt die Dissimilation mit der Zer legung polymerer Substanzen (Reservestoffe) in ihre Monomere, d. h. der Mobilisierung (z. B. von Stärke oder Glykogen in Glucose; von Fett in Fettsäuren und Glycerin; von Protein in Aminosäuren). Sodann werden diese in spezifischen Reakt ionsfolgen (Glykolyse; β-Oxidation der Fettsäuren; Desaminierung von Aminosäur en) weiter zerlegt, bis schliesslich alle Abbauwege - unter aeroben B edingungen - im Tricarbonsäurezyklus zusammenmünden. In diesem wird der Koh lenstoff (als CO2) vom Wasserstoff (hauptsächlich auf NAD übertragen) getren nt. In der Atmungskette schliesslich wird der Wasserstoff unter ATP-Gewi nn auf Sauerstoff übertragen, wobei die zum Verlauf der Dissimilation no twendigen Wasserstoffakzeptoren (hauptsächlich wie oben erwähnt NAD) regeneriert werden. Bei der aeroben Dissimilation werden also CO 2 und H2O freigesetzt und O2 verbraucht. Der vollständige Abbau des Ausgangsmaterials bei der Atmung zu energiearmen Produkten (CO2 und H2O) ist durch einen hohen Energiegewinn gekennzeichnet (z. B. 36 Mol ATP/Mol Glucose).
Atmung: C6H12O6 + 6 O2 → 6 CO2 + 6 H2O (∆ G0’ = -686 kcal)
b. Gärung Unter anaeroben Bedingungen werden die Dissimilationssu bstrate ab der Stufe der Monosaccharide unvollständig abgebaut, wobei relati v energiereiche Endprodukte (z. B. Milchsäure, Ethanol) entstehen. Infol gedessen ist die Energieausbeute der Gärung weit geringer als die der Atmung (z. B. 2 Mol ATP / Mol Glucose). Um gleich viel ATP zu gewinnen wie bei de r Atmung, muss also bei der Gärung wesentlich mehr Substrat umgesetzt werden. Unter anaeroben Bedingungen verläuft die Dissimilatio n von Glucose zunächst über die Reaktionsfolge der Glykolyse, wobei pro Molekül Glucose 2 Moleküle ATP gewonnen und 2 Moleküle NAD + verbraucht werden. Um letzteres zu regenerieren, wird entweder das entstandene Pyruvat zu Lactat reduzier t (Milchsäuregärung):
C6H12O6 → 2 CH3-CHOH-COOH (∆G0’ = -28 kcal)
oder der im Anschluss an die Glykolyse entstehende Acetalde hyd (Pyruvat-Decarboxylase-Reaktion) zu Ethanol hydriert (alkoholische Gärung):

43
C6H12O6 → 2 C 2H5OH + 2 CO2 (∆G0’ = -62 kcal) Während die Milchsäuregärung hauptsächlich von Bakterien oder tierischem Gewebe vollzogen wird, findet man bei Hefepilzen und pflanzlichen Geweben unter anaeroben Bedingungen fast immer die alkoholische Gärun g. Sie wird in vielen biotechnologischen Prozessen eingesetzt, wobei es entweder a uf das entstehende CO2 (Backvorgang), den entstehenden Alkohol (Weinherstellun g) oder beide Produkte (Bierbereitung) ankommt.
c. Gaswechselmessungen bei Atmung und Gärung Ohne die Zelle zu beschädigen, sind Aussagen über Vorlieg en und Ausmass der Atmung bzw. Gärung nur möglich, wenn man eine Messung d er daran beteiligten gasförmigen Produkte vornimmt. So gibt das Ausmass des aus einem Reaktionsvolumen verschwindenden Sauerstoffs sowie die Men ge des gebildeten Kohlendioxids für viele das Stoffwechselgeschehen der Dissi milation betreffende Fragen wichtige Hinweise. Durch die folgenden zwei Versu che sollen einfache quantitative Nachweise des Sauerstoffverbrauchs bei der A tmung und der CO2-Entwicklung bei der Gärung vorgestellt werden. (Diese a nschaulichen Methoden vernachlässigen jedoch mehrere Faktoren, die zur exakten Q uantifizierung des Vorgangs notwendig wären. So wären z. B. die Konstanz u nd genaue Bestimmung des Volumens des Reaktionsraums und gleichbleibende Temper atur unerlässlich für die genaue quantitative Bestimmung des Gaswechsels.)
Volumetrischer Nachweis des O 2-Verbrauchs und der CO 2-Produktion
1. Volumetrischer Nachweis des Sauerstoffverbrauchs bei der Atmung Der Sauerstoffverbrauch durch atmende Pflanzenteile lässt sich durch die Abnahme des Gasvolumens in einem geschlossenen Raum von kon stanter Temperatur messen, wenn man das an die Atmosphäre gleich zeitig abgegebene CO2 entfernt. Ausführung: Zur Vorbereitung des Versuchs muss die Apparatur aufgebau t werden. Zuerst wird das Wasserbad aus Styropor mit kaltem Leitungswasser gefüll t und die Stative auf die richtige Höhe eingestellt. Ausserdem wird Aluminium folie zugeschnitten, mit der später die Erlenmeyerkolben umhüllt werden. Dann wird in zweien der drei 250 ml Erlenmeyerkolben ein kleines Gläschen auf dem Boden befestigt. Diese beiden Erlenmeyerkolben werden mit je 5 g Weizenkeimlingen vo rsichtig bestückt (lange Pinzette). Dabei ist darauf zu achten, dass sich kein Keimli ng in dem Einsatzgläschen befindet. Nun werden die am Boden befestigten Gläschen vorsichtig mit Hilfe einer Pipette gefüllt (Peläusballs): Eines mit 5 ml 10N KOH, das ande re mit 5 ml H 2O, als Parallelansatz. Der dritte Erlenmeyerkolben enthält nu r 5 ml Wasser (und keine Weizenkeimlinge) und dient als Thermobarometer. Nun werden die mit Keimlingen und KOH bzw. H 2O beschickten Erlenmeyerkolben vorsichtig mit Alufolie umhüllt und alle drei mit Gum mistopfen verschlossen, durch
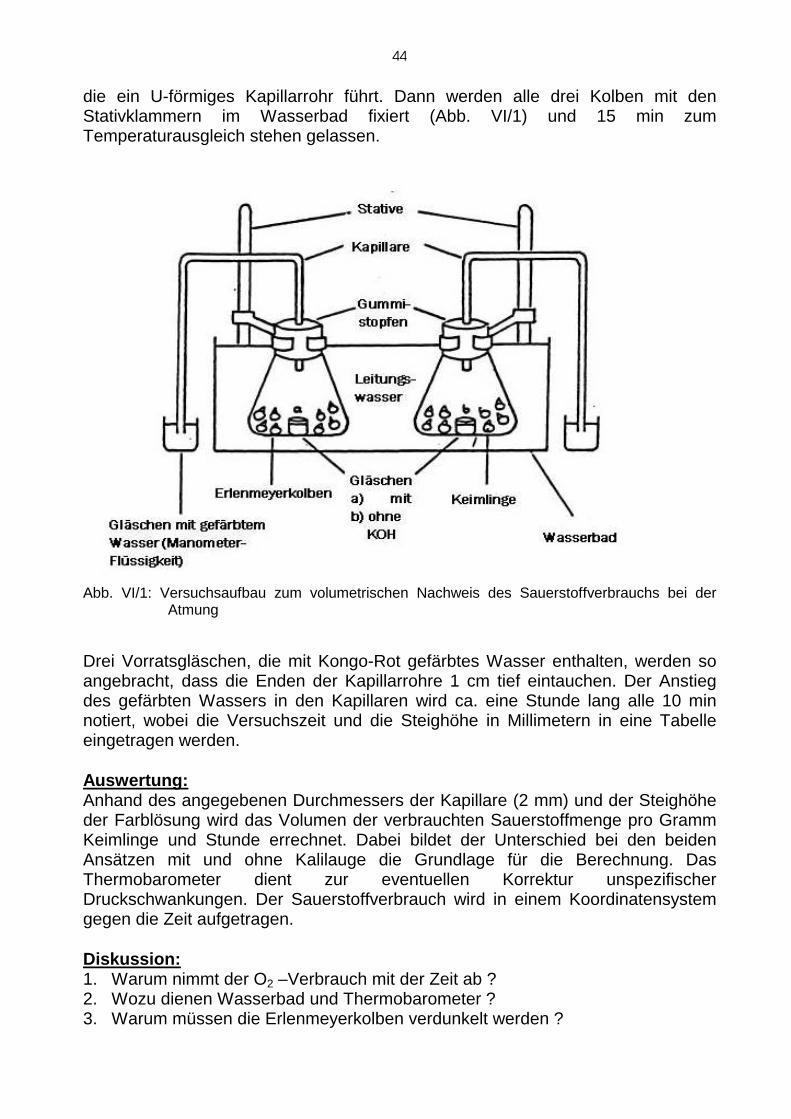
44
die ein U-förmiges Kapillarrohr führt. Dann werden a lle drei Kolben mit den Stativklammern im Wasserbad fixiert (Abb. VI/1) und 15 min zum Temperaturausgleich stehen gelassen.
Abb. VI/1: Versuchsaufbau zum volumetrischen Nachweis des S auerstoffverbrauchs bei der Atmung
Drei Vorratsgläschen, die mit Kongo-Rot gefärbtes Wasser enthalten, werden so angebracht, dass die Enden der Kapillarrohre 1 cm tief eintauchen. Der Anstieg des gefärbten Wassers in den Kapillaren wird ca. eine St unde lang alle 10 min notiert, wobei die Versuchszeit und die Steighöhe in Mi llimetern in eine Tabelle eingetragen werden. Auswertung: Anhand des angegebenen Durchmessers der Kapillare (2 mm) und der Steighöhe der Farblösung wird das Volumen der verbrauchten Sauer stoffmenge pro Gramm Keimlinge und Stunde errechnet. Dabei bildet der Unt erschied bei den beiden Ansätzen mit und ohne Kalilauge die Grundlage für die Berechnung. Das Thermobarometer dient zur eventuellen Korrektur unspezi fischer Druckschwankungen. Der Sauerstoffverbrauch wird in einem K oordinatensystem gegen die Zeit aufgetragen. Diskussion: 1. Warum nimmt der O 2 –Verbrauch mit der Zeit ab ? 2. Wozu dienen Wasserbad und Thermobarometer ? 3. Warum müssen die Erlenmeyerkolben verdunkelt werden ?

45
4. Wozu dient die KOH ?
2. Volumetrische Bestimmung der CO 2-Produktion bei der Vergärung von Zuckern durch Hefe
Volumetrische Bestimmung des bei der alkoholischen Gärung freigesetzten Kohlendioxids kann im Gärungssaccharometer nach Einhorn du rchgeführt werden (Abb. VI/2).
Abb. VI/2: Gärröhrchen nach Einhorn (Quelle: A. Wild) Der Zuckergehalt pflanzlicher Extrakte kann durch Gärung und Messung des dabei gebildeten CO 2 quantitativ nachgewiesen werden. Parallel dazu wird di e Verfügbarkeit verschiedener Kohlenhydrate untersucht. Die Bestimmung der durch Gärung entstandenen CO 2-Produktion mit Hilfe des Gärungssaccharometers ermöglicht keine quantitative Aussage, aber da die einzelnen Bestimmungen unter annähernd gleichen
Bedingungen durchgeführt werden, sind die Ergebnisse un tereinander vergleichbar. Ausführung: Von den verschiedenen gestellten Extrakten bzw. Zucker- un d Stärkelösungen werden jeweils 7 ml mit 7 ml einer Hefesuspension (1g/1 5 ml Wasser: vorher kräftig schütteln um die Hefe gleichmässig zu verteilen!) in Reagenzgläsern vermischt und in Gärröhrchen eingefüllt. Darauf achten, dass im langen Schenkel des Saccharometers keine Luftblase zurückbleibt! Die Röhrchen werden bei 30°C in einem Brutschrank stehen gelassen. In Abständen von 15 Minuten wird das Volumen des gebildet en CO2 an Hand der am langen Schenkel des Saccharometers angebrachten Markierung 1,5 bis 2 Stunden lang abgelesen und in einer Tabelle protokolliert. (G asbläschen an der Gefässwandung vorher durch leichtes Klopfen lösen!) Untersuchungslösungen: a) Pflanzenextrakt: Gekochter und filtrierter Extrakt vo n 10 g Apfel in 10 ml dest. Wasser b) Zucker: 5%-ige Lösungen von Glukose, Fruktose, Galaktose, Saccharose, Lactose, Maltose und Mannit (ein Zuckeralkohol) c) Stärke: 1%-ige Lösung. Auswertung: Das Volumen an gebildetem CO2 pro 1 g Substanz wird berechnet und dann in einem Koordinatensystem gegen die Zeit eingetragen.

46
Diskussion: 1. Welche der getesteten Substanzen wird am besten von H efen vergoren? 2. Warum werden manche Substanzen überhaupt nicht vergo ren? 3. Warum kann keine quantitative Aussage gemacht werden? 4. Warum wird der Apfelextrakt bezogen auf 1 Gramm be sser vergoren als die reinen Zucker ? 5. Was für ein Gas entsteht bei diesem Versuch?

47
VII. Anhang
A. Übersicht über Kurstage und Versuche Aufgabe Gruppe
Kurstag 1
Kurstag 2
Kurstag 3
Kurstag 4
Kurstag 5
Kurstag 6
1. Wasserhaushalt A F E D C B 2. Mineralstoffe B A F E D C 3. Photosynthese & Assimilattransport
C B A F E D
4. Enzyme D C B A F E 5. Hormone und Bewegungen
E D C B A F
6. Dissimilation F E D C B A
B. Literatur: Larcher W. (1994) Ökophysiologie der Pflanzen 5. Auflage, 394 Seiten Verlag Eugen Ulmer Stuttgart Lüttge U., Kluge M. %Bauer G. (1999) Botanik 3. (veränderte) Auflage, 625 Seiten (+21), Wiley - VCH Verlag, Munk K. (2001) Grundstudium Biologie, Botanik Spektrum, Akademischer Verlag, Heidelberg Richter G. (1996) Biochemie der Pflanzen 522 Seiten (+11) Georg Thieme Vlg., Stuttgart Schopfer D. (1976) Experimente zur Pflanzenphysiologi e Nachdruck der 1. Auflage von1970, 418 Seiten Springer Verlag, Berlin Taiz L. & Zeiger E. (2000) Physiologie der Pflanzen 774 Seiten (+25) Spektrum, Akademischer Verlag, Heidelb erg Wild A. (1999) Pflanzenphysiologische Versuche in der Sch ule 1. Auflage, 413 Seiten (+17) Quelle und Mayer Vlg

48
C. Pipettierübungen
Pipettierübung und Eichung einer automatischen Mikr oliter-Pipette
Gebrauch von Mikroliter-Pipetten Um mit möglichst kleinen Probemengen auszukommen und auch um teure Enzyme und Substrate zu sparen, bemüht man sich, das Reakti onsvolumen in der Küvette klein zu halten (0.5 - 1.0 ml). Dies erfordert eine zuverlässige Handhabung um Mikroliter-Mengen genau zu pipettieren. Gebrauchsanleitung für Mikroliter-Pipetten.
A) Spitze aufsetzen. Druckknopf bis zum 1. Anschlag drück en. B) Spitze ca. 3 mm in die zu entnehmende Flüssigkeit e intauchen und Druckknopf langsam zurückgleiten (nicht zurückschnappen) lassen. Gefü llte Spitze beim Herausnehmen an der Wandung abstreifen. C) Pipettenspitze an die Wand des Gefässes, in welches pip ettiert werden soll, anlegen und Knopf langsam bis zum 1. Anschlag drücken. D) Nach ca. 1 sec. Knopf bis zum Endanschlag drücken und in dieser Position die Spitze an der Wand abstreifen und aus dem Gefäss z iehen, Knopf langsam zurückgleiten lassen, Spitze wechseln. Oft pipettiert man auch auf einen Kunststoffspatel (Pl ümper) und rührt dann die pipettierte Flüssigkeit z. B. in eine Küvet te ein. Wenn einzelne Lösungen wiederholt pipettiert werden, verwendet ma n die gleiche Spitze, die dann zwischendurch in einer Spitzenbank (Position bezeichnen) aufgehoben wird.
Die Pipetten dürfen nur im Ständer aufbewahrt werde n, nie hinlegen. Weitere verwendete Geräte: Analysenwaage Eichung der automatischen Pipette: Dieser Versuch gibt Ihnen Gelegenheit, den Umgang mit einer automatischen Pipette zu üben und die Genauigkeit Ihres Pipettierens (und der Pipette) zu bestimmen. An der Analysenwaage (Bedienung durch Assisten ten erklären

49
lassen!) wird in einer 50 µl Pipette die Menge Wasser a usgewogen, deren Volumen mit der Pipette abgemessen wird. Es werden 10 Messungen durchgeführt. Nach jeder Ablesung die Waage arretieren und erneut 50 µl Wasser zugeben. Auswertung:
Bilden Sie den arithmetischen Mittelwert x_
:
�=
=n
jx
nx j1
1
x j
= einzelne Messergebnisse
n = Anzahl der Messungen
ferner die Standardabweichung σ ( = mittlerer Fehler der Einzelmessung)
� �����
� −=−
=n
jxx jn 1
²_
1
1σ
Die prozentuale Abweichung wird durch Variationskoeffizie nten v ausgedrückt
x_
σν =
Standardabweichung und der daraus abgeleitete Variati onskoeffizient sind ein Mass für die Präzision und Reproduzierbarkeit der Messung.


















