
Peer-reviewed | Manuscript received: February 13, 2013 ... · cranial nerves (VII., IX., X.)...
8
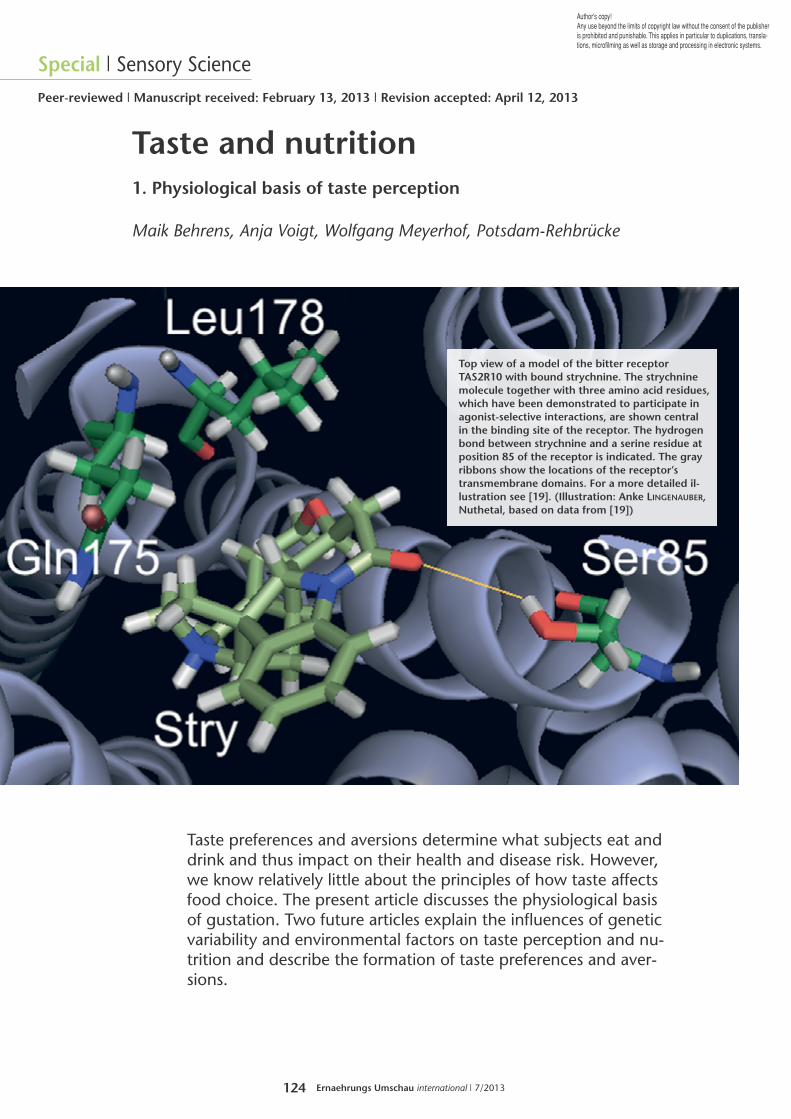
124 Ernaehrungs Umschau international | 7/2013 Special | Sensory Science Peer-reviewed | Manuscript received: February 13, 2013 | Revision accepted: April 12, 2013 Author's copy! Any use beyond the limits of copyright law without the consent of the publisher is prohibited and punishable. This applies in particular to duplications, transla- tions, microfilming as well as storage and processing in electronic systems. Taste and nutrition 1. Physiological basis of taste perception Maik Behrens, Anja Voigt, Wolfgang Meyerhof, Potsdam-Rehbrücke Top view of a model of the bitter receptor TAS2R10 with bound strychnine. The strychnine molecule together with three amino acid residues, which have been demonstrated to participate in agonist-selective interactions, are shown central in the binding site of the receptor. The hydrogen bond between strychnine and a serine residue at position 85 of the receptor is indicated. The gray ribbons show the locations of the receptor’s transmembrane domains. For a more detailed il- lustration see [19]. (Illustration: Anke LINGENAUBER, Nuthetal, based on data from [19]) Taste preferences and aversions determine what subjects eat and drink and thus impact on their health and disease risk. However, we know relatively little about the principles of how taste affects food choice. The present article discusses the physiological basis of gustation. Two future articles explain the influences of genetic variability and environmental factors on taste perception and nu- trition and describe the formation of taste preferences and aver- sions.
-
Upload
vuongnguyet -
Category
Documents
-
view
214 -
download
0
Transcript of Peer-reviewed | Manuscript received: February 13, 2013 ... · cranial nerves (VII., IX., X.)...
124 Ernaehrungs Umschau international | 7/2013
Special | Sensory Science Peer-reviewed | Manuscript received: February 13, 2013 | Revision accepted: April 12, 2013
Author's copy!Any use beyond the limits of copyright law without the consent of the publisheris prohibited and punishable. This applies in particular to duplications, transla-tions, microfilming as well as storage and processing in electronic systems.
Taste and nutrition1. Physiological basis of taste perception
Maik Behrens, Anja Voigt, Wolfgang Meyerhof, Potsdam-Rehbrücke
Top view of a model of the bitter receptorTAS2R10 with bound strychnine. The strychninemolecule together with three amino acid residues,which have been demonstrated to participate inagonist-selective interactions, are shown centralin the binding site of the receptor. The hydrogenbond between strychnine and a serine residue atposition 85 of the receptor is indicated. The grayribbons show the locations of the receptor’stransmembrane domains. For a more detailed il-lustration see [19]. (Illustration: Anke LINGENAUBER,Nuthetal, based on data from [19])
Taste preferences and aversions determine what subjects eat anddrink and thus impact on their health and disease risk. How ever,we know relatively little about the principles of how taste affectsfood choice. The present article discusses the physiological basisof gustation. Two future articles explain the influences of geneticvariability and environmental factors on taste perception and nu-trition and describe the formation of taste preferences and aver-sions.

�
Ernaehrungs Umschau international | 7/2013 125
Functional morphology ofthe peripheral taste system
FlavorAt least tree of our senses contributeto the perception of food. The senseof smell detects the scent of a mealwhich, through sniffing, reaches theolfactory mucosa via the nostrils orthe nasopharynx. Touch and pain re-port about the texture and tempera-ture of food and the presence of irri-tants that elicit hot, pungent, astrin-gent, metallic, burning, tingling orelectrical sensations. Taste, in thetrue sense of the word, is restrictedto the five basic tastes or taste qual-ities, sweet, sour, salty, bitter, andumami (savoury). The brain uses allthese types of impressions to con-struct complex flavors. In the fol-lowing the word taste merely refersto the five basic taste qualities.
Taste organs
Taste budsThe five basic tastes are elicited bysubstances that are recognized by thetaste buds. These principle taste or-gans are onion-shaped assemblies of~ 50–100 mostly elongated cells.With their apical tips numerous ofthese cells contact a depression in theoral epithelium, referred to as tastepore, the contact site between tastemolecules and their receptive struc-tures.
Taste papillaeThe majority of oral taste buds re-sides in the lingual taste papillae(� Figure 1A). We distinguish fungi-
form, foliate, and vallate papillaebased on their shape, location andnumber of hosted taste buds. About300 small, slightly elevated fungi-form papillae are distributed aboutthe anterior two thirds of thetongue. In humans, they contain upto five taste buds [1]. The foliatepapillae are found at the posterioredges of the tongue and form up tofive fissions which contain severalhundred taste buds [2]. The posteriortongue displays on average nine val-late papillae which are arranged in aV-shaped manner. They possessmost of the lingual taste buds aseach of them contains several hun-dred of these cell assemblies. Furthertaste buds are embedded in the oralepithelium of the palate, epiglottis,pharynx and larynx.
Chemosensory cellsThe chemosensory cells of taste budsdiffer in shape and function (� Figure1B) [2,3]. They are referred to as sec-ondary sensory cells to indicate thefact that they are specialized epithe-
lial cells and not neurons. The con-ventional cell typing that is based onmorphological criteria is currentlybeing replaced by functional para -meters which led recently to the dis-covery of a fundamental principle ingustation, i. e., the existence of ge-netically determined, segregatedpopulations of chemosensory cellsfor the 5 basic tastes [3]. This meansthe percept of a basic taste is equi-valent to the excitation of the cog-nate population of oral chemosen-sory cells. This principle explains theexistence of only few taste qualitiesas well as the high discriminatorypower across taste qualities and thelow discriminatory power with tastequalities. The molecular receptiveranges of the five taste cell popula-tions for taste compounds are de-fined and separated by the specialtaste receptor molecules they ex-press. The bitter off-taste of saccha-rin is thus a result from its ability topotently activate the sweet sensingcells and, at the same time, toweakly excite the sensory cells dedi-
SummaryEach taste quality is represented by a specific population of oralchemosensory cells. They are equipped with special receptor moleculeswhich determine the molecular receptive ranges of the sensory cells.Whereas the receptors for salty and sour are, at present, not extensivelycharacterized, a plethora of data exists for sweet, umami and bitter re-ceptors. Although sweet and bitter receptors recognize a broad rangeof compounds, two different strategies are effective to achieve this goal.In case of the bitter receptors numerous genes have evolved, whereasonly two genes code for subunits of the sweet taste receptor. This re-ceptor, however, possesses multiple binding sites to recognize a diversityof sweet substances.
In the taste buds of the tongue sensory cells form assemblies of about100 cells, which process and integrate taste information with metabolicneeds. Sensory afferent nerves transfer gustatory information from themouth to the brainstem to evoke stereotyped innate attraction or aver-sion and to prepare the body for digestion. The activity of nerve cells inspecial areas of the cerebral cortex represents the basic tastes and gen-erates complex flavours by integrating information about taste, smelland texture of food.
Keywords: taste, gustation, physiology, sweet, salty, sour, bitter, umami
Citation: Behrens M, Voigt A, Meyerhof W(2013) Taste and nutrition. 1.Physiological basis of taste per-ception. Ernaehrungs Umschau in-ternational 60(7): 124–131
This article is available online:DOI: 10.4455/eu.3013.024

126 Ernaehrungs Umschau international | 7/2013
Special | Sensory Science
cated to bitter. Taste cells are veryshort-lived and replaced by cells thatdevelop from spherical progenitorcells that reside at the base of tastebuds [4].
Detection and transmission oftaste stimuli
Stimulation of all types of taste re-ceptor cells leads to the excitation ofafferent sensory fibers in a mannerdependent on the neurotransmitterATP. This has been demonstrated inmice lacking a special type of ATP re-ceptor [5]. Noteworthy, only the re-ceptor cells dedicated to sour formconventional chemical synapses, i.e.,specialized contacts between twocells that propagate the excitationfrom cell to cell. However, it remainsunknown if sour sensing cellssynaptically release ATP [2]. The cellsfor sweet, bitter and umami stimulido not form synapses. However,they have specializations where af-
ferent fibers come particularly close.It is assumed that these are the sitesof ATP release. The use of the sameneurotransmitter by all of the 5 re-ceptor cell populations raises the im-portant question of how the braincorrectly interprets the taste quali-ties. The most attractive hypothesisto date assumes that the released ATPis confined to quality specific microdomains in the taste buds [2], yettheir existence remains to be demon-strated.In addition to ATP, several othertransmitter substances along withtheir corresponding receptors havebeen identified in taste buds. Al-though not in all cases the specificrole of these additional neurotrans-mitters was demonstrated unam-biguously, evidence is mounting thatthey serve functions in the commu-nication between sensory cells. Thereleased ATP for example, not onlyelicits action potentials in the affer-ent nerve fibers, but also stimulates,
via a different type of receptor, itsown release from sensory cells [6].Furthermore, adenosine resultingfrom the enzymatic degradation ofATP, stimulates selectively the re-sponsiveness of sensory cells devotedto the recognition of sweet stimuli[6]. This clearly indicates that tastebuds are in fact not just structureswhich detect and transmit taste stim-uli, but rather complex sensory or-gans that integrate taste informationalready in the oral cavity [2]. Thisconcept is underscored by the factthat taste sensitivity is dynamicallymodulated to fit acute metabolic re-quirements. For example the counter-regulatory roles of the hunger andsatiety hormones endocannabinoidand leptin, respectively, modify thesensitivity of sweet taste receptorcells in opposite directions [7].
Neuroanatomy
Taste information originating fromthe oral cavity is transmitted to thebrainstem via three cranial nerves(� Figure 1C) [8]. Contacts with othernerve cells residing in this brain re-gion mediate muscle contractions co-ordinating tongue movement, chew-ing and swallowing to facilitate theingestion of digestible food items andthe disgorging of potentially harm-ful substances. Furthermore, taste in-formation prepares the alimentarytract in advance for the digestion ofthe food [9]. The brainstem wirestaste information via the midbrain toa specialized area within the cere-brum, the gustatory cortex. Here, theconscious perception of the basictaste qualities is reflected by nerve ac-tivity patterns. Together with infor-mation about the smell and texturalproperties of the consumed food acomplex flavor percept is generated.Flavor recognition, in turn, is indis-pensable for the development of foodpreferences and aversions. Furtherbrain areas involved in the process-ing of taste information include re-gions that regulate intake of nutri-
Figure 1: Anatomy of the taste system (A) Schematic of the location and morphology of the three types of tastepapillae. The taste buds are embedded in the mucosa of protrusions or in-vaginations of taste papillae. (B) The onion-shaped appearance of tastebuds is caused by numerous elongated cells. Note that for each of the fivebasic taste qualities a separate cell population exists. (C) Branches of threecranial nerves (VII., IX., X.) innervate the taste buds and transmit taste in-formation to the Nucleus tractus solitarius (NTS), a part of the brainstem.The cell bodies of the three nerves are located in ganglia outside the brain.The NTS relays taste information via the thalamus (TH) to the gustatorycortex (GC). (Illustration: Jonas TÖLE, Nuthetal)
fungiform papilla
oral cavity
sweetumamisaltysourbitter
supporting cellprecursor cell
nerve fibers
taste receptorcells
epithelium
connectivetissue
foliate papilla
vallate papilla
A
B NTS
TH
GC
VII.
IX.
X.
gustatorynerves
C

�
Ernaehrungs Umschau international | 7/2013 127
ents and liquids as well as rewardsystems involved in the generation ofaddictive behaviors [1].
The receptors for sweet,umami, and bitter tasteperception
The tasks for sweet and umami tastereceptors on the one hand, and thatof bitter taste receptors on the otherhand couldn’t be more different.Whereas sweet and umami receptorsare tailored for the detection of thebuilding blocks for the universalmacronutrients, carbohydrates andproteins, bitter taste receptors are re-quired to respond to a large varietyof potentially harmful toxic food in-gredients. For these very differenttasks nature has chosen its perhapsmost versatile tools, the G protein-coupled receptors (GPCRs). Numer-ous hormone and neurotransmitterreceptors, but also the visual pig-ment, rhodopsin, belong to this classof molecules.
TAS1Rs
The family of TAS1R (taste receptor,type 1) genes is responsible for rec-ognizing the taste qualities sweetand umami (for a recent review arti-cle see [10]). Based on their aminoacid sequence and structural simi-larities the three members of thisgene family, TAS1R1, -R2 and –R3,belong to the class C of GPCRs. Thisreceptor class is characterized bylong, amino terminally located ex-tracellular domains. They form astructure resembling the traps of theplant Venus flytrap (“venus flytrapmotif”). The carboxy terminal partof these proteins with its 7 trans-membrane domains that are con-nected via 3 intra- and 3 extracellu-lar loops, however, takes the shapeof a typical GPCR. Between theamino terminal Venus flytrap motifand the transmembrane region anadditional, cystein-rich domain ispresent.
A functional TAS1-receptor consistsof two different subunits. Whereasthe TAS1R1 and TAS1R3 subunitsassemble to form the umami recep-tor [11], the sweet taste receptorconsists of the TAS1R2 and TAS1R3subunits [12]. Hence, both receptorsshare the TAS1R3 subunit and pos-sess one specific subunit each. On acellular level this principle is evidentby the observed co-expression of theTAS1R3 gene with either of theTAS1R1 or TAS1R2 genes, but astrict separation of TAS1R1 andTAS1R2 gene expression.
The sweet taste receptor TAS1R2/3is the universal sensor for all sub-stances eliciting a sweet taste percep-tion. Given the large number ofsweet tasting substances, this is anenormous task to fulfill. On top ofthe prototypical sweet tastants suchas mono- and disaccharides (� Table1), numerous natural and syntheticcompounds activate this receptor.Perhaps one of the most importantfindings of recent research concern-ing the sweet taste receptor is thecharacterization of individual bind-ing sites of various sweet tastants in
Table 1: Human taste receptors and some of their activators TAS1R/TAS2R = tast receptor, type 1/2
Receptor Agonists
umamiTAS1R1/R3 L-glutamic acid, enhanced by 5’-ribonucleoside phosphate
sweetTAS1R2/R3 Mono-/disaccharides,
plant sweet proteins (e.g. brazzein, thaumatin), artificialsweeteners (e.g. saccharin, aspartame), natural sweeteners(e. g. steviosides)
bitterTAS2R1 Humulones
TAS2R3 Chloroquine
TAS2R4 Colchicin, L-tryptophane, D-tryptophane
TAS2R5 1,10-Phenanthrolin
TAS2R7 caffeine
TAS2R8 Chloramphenicol
TAS2R9 Ofloxazin
TAS2R10 Parthenolide, denatonium benzoate, strychnine
TAS2R13 Diphenidol, denatonium benzoate
TAS2R14 picrotoxinin, aristolochic acid, caffeine, absinthin
TAS2R16 D-(-)-salicin, D-arbutin, amygdalin
TAS2R20 Cromolyn
TAS2R30 Denatonium benzoate, Picrotoxinin
TAS2R31 Aristolochic acid, saccharin, acesulfame K
TAS2R38 Phenylthiocarbamide (PTC), 6-n-propylthiouracil (PROP)
TAS2R39 Quinine, thiamin, amarogentin
TAS2R40 Humulones, quinine
TAS2R43 Aristolochic acid, saccharin, acesulfame K
TAS2R46 strychnine, absinthin, parthenolide
TAS2R50 Amarogentin, andrographolide

128 Ernaehrungs Umschau international | 7/2013
Special | Sensory Science
different parts of this receptor (� Fi-gure 2). A key finding for the suc-cessful identification of these bindingsites has been the observation ofspecies-specific differences in sweettaste perception. Whereas many ar-tificial sweet compounds activatedthe human sweet taste receptor, thecorresponding rodent receptors donot respond to many of these sub-stances [12,13]. Through the smartcombination of subunits from dif-ferent species or parts thereof andsubsequent functional in vitro exper-iments it was possible to locate thesebinding sites. These experimentsdemonstrated that the sweet taste re-ceptor possesses several binding sitesfor sweet compounds distributedover different parts of the moleculeand, moreover, provided a convinc-ing explanation for the apparent dis-crepancy between the number anddiversity of sweet compounds andthe existence of only a single sensorfor these compounds. The identifiedbinding sites are located in the Venusflytrap domains of both subunits,the transmembrane region of theTAS1R3 as well as within the cys-tein-rich domain.
In contrast to the sweet taste recep-tor, for the umami receptor,
TAS1R1/3, much fewer activatingmolecules have been identified.Whereas the human umami recep-tor is specifically tailored for therecognition of L-glutamate, the cor-responding rodent receptor is acti-vated by additional L-amino acids[11]. A common hallmark of thehuman as well as the rodent umamireceptor is a strong, ~30-fold en-hancement of its sensitivity by ino-sine monophosphate (IMP) and other5-ribonucleotides such as GMP.
TAS2Rs
There are countless of structurallydiverse bitter compounds present innature. Many of these possess con-siderable pharmacological activitiesand hence could be harmful if in-gested. The perception of all thesepotentially toxic substances is medi-ated by the bitter taste receptors ofthe TAS2R (taste receptor, type 2)gene family [14,15]. Humans pos-sess 25 of these receptors, whereasother mammals can have more (e.g.mouse with 35) or fewer (e.g. horseswith 19) TAS2R genes.Bitter taste receptor genes in the oralcavity are expressed in a separatepopulation of taste sensors, the bit-ter taste receptor cells. Since each of
these bitter taste receptor cells ex-press on average only 4-11 differentTAS2R genes, the population ofhuman bitter taste receptor cells ishighly heterogeneous [16]. In con-trast to the already describedTAS1Rs, TAS2Rs exhibit only veryshort extracellular amino termini(� Figure 2) and, due to their lowamino acid sequence relationshipwith other GPCRs, are difficult to begrouped into existing GPCR-subfam-ilies.
One of the most burning questionsin bitter taste research has been, howa rather small number of TAS2Rsmay possibly suffice to detect hun-dreds of diverse bitter compounds?The enormous progress made in theidentification of activators for the 25human TAS2Rs (a process called “de-orphanization” because previously“orphan” receptors become associ-ated with cognate activators) hasprovided important results to an-swer this question (� Table 1).
In the course of these experiments,which resulted in the successful de-orphanization of, so far, 21 of the 25human TAS2Rs, it was shown thatthe breadth of agonist spectra ofthese receptors deviate considerably.Some receptors, the “generalist” re-ceptors, exhibit an extremely broadagonist spectrum, the “specialist” re-ceptors respond only to few ago-nists, whereas the majority of recep-tors showed an intermediate breadthof tuning [17]. The three mostbroadly tuned receptors, TAS2R10, -R14, and –R46, each respond toabout one-third of all tested bittercompounds and their combined ago-nist spectra are sufficient for therecognition of already half of all bit-ter substances. On the other extreme,the receptors TAS2R3, -R5, -R13, -R20, and -R50 are activated by onlyvery few bitter substances. Intri-guingly, even the most broadly tunedTAS2Rs possess only a single ligandbinding pocket (� cover illustration),which accommodates all the diverse
Figure 2: Schematic of sweet, umami, and bitter receptor structures The extracellular Venus flytrap motifs of the TAS1R are shown. Thecystein-rich domains of TAS1Rs are highlighted as bold lines. Trans-membrane helices are shown as cylinders traversing the plasmamembrane. Identified binding sites for activators are indicated bygray spheres. (Illustration: Authors’ own schematic)TAS1R/TAS2R = taste receptor, Type 1/2
TAS1R1 TAS1R3 TAS1R2 TAS1R3 TAS2R
sweet receptor umami receptor bitter receptor

�
Ernaehrungs Umschau international | 7/2013 129
bitter compounds via different con-tact points between receptor and ag-onists [18].
Moreover, it has been demonstratedthat the ligand binding pockets aretailored to accommodate numerouscompounds at the expense of possi-bly higher affinities for each of thesingle substances [19]. Two of the 25human TAS2Rs exhibit pronouncedselectivity for distinct large classes ofbitter compounds. One of them isthe TAS2R16, which responds al-most exclusively to stimulation withβ-D-glucopyranosides, the other re-ceptor, the TAS2R38, is activatedonly by molecules possessing iso -thiocyanate or thioamide groups. Allother receptors exhibit intermediatetuning properties.
In summary, the deorphanizationprocess allowed to clarify the ques-tion of how so few receptors can pos-sibly be responsible to facilitate recog-nition of so many different bittercompounds. In fact, the question cannow be turned around and one mayask why do humans possess so manyreceptors with, in part, clearly over-lapping agonist spectra? The recentidentification of naturally occurringbitter taste receptor inhibitors mayprovide an explanation for this phe-nomenon [20]. These inhibitors arepresent in the same plants synthesiz-ing typical bitter compounds, how-ever, unlike agonists they block bitterreceptor responses instead of activat-ing them. The presence of multiple re-ceptors with overlapping agonist in-teraction patterns may prevent thecomplete inhibition of bitter tasterecognition by such substance andthus, prevent the accidental ingestionof those plants with possibly fatalconsequences.
Intracellular signaling cascades ofTAS1Rs and TAS2RsDespite of the obvious differences instructure, number, activation mech-anisms and their strict separation indifferent cell populations among
sweet and umami receptors on theone hand, and bitter receptors on theother hand, the signal transductionmechanism is similar (� Figure 3).Activation of the receptors is trans-mitted via a specific heterotrimericguanine nucleotide binding proteinonto the enzyme phospholipase C 2.The resulting inositol-1,4,5-tris -phosphate stimulates the type 3 IP3-
receptor in the endoplasmic reticu-lum causing elevation of intracellu-lar calcium levels. The calcium-in-duced opening of the ion channelTRPM5 initiates a kation currentthrough the plasma membrane, re-sulting in the depolarization of tastereceptor cells and the concurrent re-lease of the neurotransmitter ATPthrough pannexin-hemichannels.
Figure 3: Flow diagram of the cellular signal transduction cascade intaste receptor cellsThe single components are shown following the sequence of theiractivation. Abbreviations: IP3-receptor, inositol-1,4,5-trisphosphatereceptor; TRPM5, transient receptor potential cation channel sub-family M, member 5; ATP, adenosine trisphosphate (Authors’ owndiagram).
sweet-, umami-, bitter receptor
heterotrimeric G ProteinGα-gustducin, Gβ3(1), Gy13
phospholipase Cβ2
inositol-1,4,5-trisphosphate
IP3-receptor
calcium ions
TRPM5 channel
cell depolarization
pannexin hemichannel
neurotransmitter ATP

130 Ernaehrungs Umschau international | 7/2013
Special | Sensory Science
Sour transduction
Acids are the principle stimuli ofsour taste. Since strong inorganicacids such as hydrochloric acid dis-sociate completely in protons andanions, we have to assume that pro-tons themselves serve as sour stim-ulus. But also weak organic acidstaste sour. Paradoxically, at constantpH the weak acetic acid appears to besourer than the strong hydrochloricacid [6]. This led to the assumptionthat, in addition to protons, theundissociated organic acids serve asadequate sour stimuli [21]. In theprotonated state organic acids are ca-pable of permeating plasma mem-branes. In the cytosol they dissociateinto protons and anions which leadsto intracellular acidification.
Receptor candidates for sour have re-peatedly been reported about in theliterature. To date, none of themmatches all of the necessary criteriathat a true sour receptor should ful-fill. Despite this lack of knowledgewe have to assume that at least twotransduction pathways are involvedin the recognition of acids [2].Plasma membrane proton channelsenable the influx of protons into thecytosol of sour-detecting receptorcells in the taste bud. Together withthose protons released from the or-ganic acids in the cytosol they acti-vate intracellular target sites. Even-tually this leads to calcium influx inthe sour-detecting sensory cells andrelease of neurotransmitter sub-stances. This hypothesis is also sup-ported by the observation that alltaste cells are accessible to organicacids and show the drop in pH butonly in the sour receptor cells lowpH is coupled to calcium influx andneurotransmitter release [6].
Salt taste transduction
We continuously loose electrolytesthrough excretion. In this contextsalty taste can be considered a con-trol element ensuring electrolytehomeostasis. The recognition of saltand its intrinsic pleasant taste at lowto moderate concentrations pro-motes salt uptake compensating thecontinued loss.
Salty taste is mostly elicited by NaCl,table salt, but other sodium saltselicit a very similar taste. The tasteof other mineral salts does not com-pare to the taste of NaCl, it is clearlydifferent.
In rodents two salt tastes have beendescribed. One of them is pleasantand specifically elicited by sodiumions. This taste is induced by lowstimulus concentrations and blockedby the diuretic drug amiloride. Theother is repulsive, depends on highsodium concentration, is also elicitedby various other mineral salts in-cluding calcium, magnesium, am-monium and potassium salts, andinsensitive to amiloride. The obser-vation that rodents are unable to dis-tinguish NaCl from KCl in the pres-ence of amiloride illustrates the du-alism of salty taste [22].
A well-known target for amiloride inthe kidney is ENaC, the epithelialsodium channel which is composedof two alpha, a beta and a gammasubunit. The ENaC is highly selectivefor sodium ions and mediates the re-covery of sodium from the primaryurine. These findings led to the as-sumption that ENaC in oral taste tis-sues is involved in salt transduction.Various circumstantial evidence sup-ported this assumption, yet onlyvery recent direct evidence demon-strated that α-ENac is part of the salttransduction machinery [23]. How-ever, the precise channel composition
remains unknown. The amiloride-insensitive salt taste involves othertransduction mechanisms. Appar-ently, high-salt-reception recruitssour and bitter transduction path-ways [24] which impressively ex-plains the unpleasant sensation thatis evoked by high sodium concentra-tions and other mineral salts.
Comparatively, little is known aboutsalt taste in humans. It is true thatthe ENaC subunits are present inhuman taste buds but salt taste inhumans is amiloride insensitive [25]challenging a potential role for ENaCin human salt transduction. Perhapsthe insensitivity of human salt tasteto amiloride can be explained by adifferent channel composition. Hu-mans possess an additional ENaCgene which is absent in rodents andwhich encodes the δ-subunit. Delta-ENaC is also present in human tastebuds and can functionally replacethe α-subunit. However, δ-ENaC isfar less sensitive to amiloride thanthe α-subunit [26] which could ex-plain the amiloride-insensitivity ofhuman salty taste. However, futureresearch has to clarify this and otheropen questions regarding sour andsalty transduction.
Outlook
We have seen that important con-cepts and basic principles in tasteperception have been worked outquite recently. However, new know-ledge raises new questions. Thus, wehave to register unexpected largegaps in our knowledge about sourand salty taste and about funda-mental principles of gustatory infor-mation transmission from mouth tobrain. Moreover, taste receptors haverecently been found in numerous ex-traoral sites where they probablyexert hitherto unknown functions.

Ernaehrungs Umschau international | 7/2013 131
Numerous laboratories world-wide aim at filling the gaps inknowledge. In particular, eluci-dating the molecular and cellu-lar basis of gustatory transmis-sion and processing in the brainis indispensable for understand-ing of how taste impacts on in-gestive behavior. The to-do-listfor taste researcher is long andcomplex and hopefully can beworked off quick in order tosolve public health issues in nu-trition some of which will betargeted in the next articles.
AcknowledgementThe authors thank Jonas TÖLE
(Nuthetal) for preparing fig-ure 1 and Anke LINGENAUBER
(Nuthetal) for the preparationof the cover illustration basedon data from [19].
1. Smith DV, Boughter JD, Jr. Neurochemistry ofthe Gustatory System. In: Lajtha A, JohnsonDA (ed). Handbook of Neurochemistry and Mo-lecular Neurobiology. Springer, New York(2007), S. 109–135
2. Roper SD (2007) Signal transduction and in-formation processing in mammalian tastebuds. Pflugers Arch 454: 759–776
3. Chandrashekar J, Hoon MA, Ryba NJ et al.(2006) The receptors and cells for mammaliantaste. Nature 444: 288–294
4. Beidler LM, Smallman RL (1965) Renewal ofcells within taste buds. J Cell Biol 27: 263–272
5. Finger TE, Danilova V, Barrows J et al. (2005)ATP signaling is crucial for communicationfrom taste buds to gustatory nerves. Science310: 1495–1499
6. Roper SD (2013) Taste buds as peripheralchemosensory processors. Semin Cell Dev Biol24: 71–79
7. Jyotaki M, Shigemura N, Ninomiya Y (2010)Modulation of sweet taste sensitivity by orexi-genic and anorexigenic factors. Endocr J 57:467–475
8. Hamilton RB, Norgren R (1984) Central projec-tions of gustatory nerves in the rat. J CompNeurol 222: 560–577
9. Spector AC (2000) Linking gustatory neurobi-ology to behavior in vertebrates. Neurosci Biobe-hav Rev 24: 391–416
10. Behrens M, Meyerhof W, Hellfritsch C et al.(2011) Sweet and umami taste: natural prod-ucts, their chemosensory targets, and beyond.Angew Chem Int Ed Engl 50: 2220–2242
11. Nelson G, Chandrashekar J, Hoon MA et al.(2002) An amino-acid taste receptor. Nature416: 199–202
12. Nelson G, Hoon MA, Chandrashekar J et al.(2001) Mammalian sweet taste receptors. Cell106: 381–390
13. Li X, Staszewski L, Xu H et al. (2002) Humanreceptors for sweet and umami taste. Proc NatlAcad Sci U S A 99: 4692–4696
14. Adler E, Hoon MA, Mueller KL et al. (2000) Anovel family of mammalian taste receptors. Cell100: 693–702
15. Chandrashekar J, Mueller KL, Hoon MA et al.(2000) T2Rs function as bitter taste receptors.Cell 100: 703–711
16. Behrens M, Foerster S, Staehler F et al. (2007)Gustatory expression pattern of the humanTAS2R bitter receptor gene family reveals a het-erogenous population of bitter responsive tastereceptor cells. J Neurosci 27: 12630–12640
17. Meyerhof W, Batram C, Kuhn C et al. (2010)The molecular receptive ranges of human TAS2Rbitter taste receptors. Chem Senses 35: 157–170
18. Brockhoff A, Behrens M, Niv MY et al. (2010)Structural requirements of bitter taste receptoractivation. Proc Natl Acad Sci U S A 107:11110–11115
19. Born S, Levit A, Niv MY et al. (2012) TheHuman Bitter Taste Receptor TAS2R10 Is Tai-lored to Accommodate Numerous Diverse Lig-ands. J Neurosci 33: 201–213
20. Brockhoff A, Behrens M, Roudnitzky N et al.(2011) Receptor agonism and antagonism of di-etary bitter compounds. J Neurosci 31: 14775–14782
21. Lyall V, Alam RI, Phan DQ et al. (2001) De-crease in rat taste receptor cell intracellular pHis the proximate stimulus in sour taste trans-duction. Am J Physiol Cell Physiol 281: C1005–C1013
22. Eylam S, Spector AC (2005) Taste discrimina-tion between NaCl and KCl is disrupted byamiloride in inbred mice with amiloride-insen-sitive chorda tympani nerves. Am J PhysiolRegul Integr Comp Physiol 288: R1361–R1368
23. Chandrashekar J, Kuhn C, Oka Y et al. (2010)The cells and peripheral representation ofsodium taste in mice. Nature 464: 297–301
24. Oka Y, Butnaru M, von Buchholtz L et al.(2013) High salt recruits aversive taste path-ways. Nature 494: 472-475
25. Staehler F, Riedel K, Demgensky S et al. (2008)A Role of the Epithelial Sodium Channel inHuman Salt Taste Transduction? Chem Percept1(1): 78–90
26. Halpern BP (1998) Amiloride and vertebrategustatory responses to NaCl. Neurosci BiobehavRev 23: 5–47
References
DOI: 10.4455/eu.2013.024
Dr. Maik Behrens1
Dr. Anja Voigt2
Prof. Dr. Wolfgang Meyerhof3
German Institute of Human Nutrition(DIfE) Potsdam-Rehbrücke,Arthur-Scheunert-Allee 114–116, 14558 Nuthetal, Germany;
1E-Mail: [email protected]: [email protected]: [email protected]
Conflict of InterestsMaik BEHRENS has filed patents and pa-tent applications on human bitter taste.
Anja VOIGT declares no conflict of interestin accordance with the guidelines of theInternational Committee of Medical Jour-nal Editors.
Prof. Dr. Wolfgang MEYERHOF filed severalpatents and patent applications on bitterreceptors. Parts of his research are fun-ded by industrial partners.


















