
Patulin Biosynthesis: The Role of Mixed-Function Oxidases in the Hydroxylation of m-Cresol
13
Eur. J. Biochem. 49,443-455 (1974) Patulin Biosynthesis : The Role of Mixed-Function Oxidases in the Hydroxylation of m-Cresol Gillian MURPHY, Giinter VOGEL, Gunther KRIPPAHL, and Feodor LYNEN Max-Planck-Institut fur Biochemie, Martinsried (Received May 27/August 28, 1974) Two hydroxylation reactions of m-cresol, an intermediate in the secondary biosynthesis of the antibiotic patulin, have been characterised in cell-free preparations of Penicillium patulum. The ability to convert m-cresol to 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol is located in separable fractions of the 105000 x g microsomal pellet. The activities had K, values for m-cresol of 80 pM and 5 pM, respectively. Both reactions require molecular oxygen and NADPH for activity. Inhibition of m-cresol hydroxylases by carbon monoxide is reversible by light with a photochemical action spectrum maximal at 450 nm. Carbon monoxide difference spectra of mycel extracts show the presence of absorbing pigments at 422 and 450-455 nm. The enzymes are not appreciably inhibited by cyanide but are inhibited by cytochrome c. rn-Cresol hydroxylation therefore appears to be catalysed by typical mixed-function oxidases involving an NADPH-dependent reductase and a pigment like cytochrome P-450. In the light of studies on the utilisation of radioactively labelled intermediates, the methyl hydrox- ylation of m-cresol to m-hydroxybenzyl alcohol probably represents an important reaction in the sequence leading to patulin formation. Ring hydroxylation of m-cresol to 2,5-dihydroxytoluene appears to be a side reaction functional at higher concentrations of m-cresol. Penicillium patulum synthesises the hemi-acetal lactone, patulin, from acetyl-CoA and malonyl-CoA via the polyketide 6-methylsalicylic acid [l, 21 and secretes it as a secondary end product. The first postulated routes for patulin biosynthesis were based upon the secondary metabolites identified by Bir- kinshaw et al. [3], Tanenbaum and Bassett [4], Bu'- Lock et a/. [5] and others and the establishment of a preferred pathway spans from the serial investigation of metabolite appearance in P.patulum cultures [6,7] to the administration of radioactively labelled de- rivatives and analysis of their utilisation [S]. The detection of m-cresol in continuous culture [5], the presence of significant levels of a specific 6-methyl- salicylic acid decarboxylase [9, lo], and the efficient conversion of m-[2H]cresol or m-['4C]cresol to patu- lin [8,11,12] suggest that rn-cresol is an important intermediate in the biosynthesis of patulin. m-Hydroxy- benzyl alcohol has also been established as major metabolic product [ 131, with efficient conversion to Enzymes. m-Cresol l-methylhydroxylase and m-cresol 2-hydroxylase, m-cresol, reduced NADP : oxygen oxido- reductases (EC 1.14.13.-). patulin [ll, 121. Incorporation of radioactively labelled m-cresol into 2,5-dihydroxytoluene was demonstrated by Scott and Yalpani [8] and by Bu'Lock et al. [7] in cultures treated with protein-synthesis inhibitors, although it has been observed that levels of 2,5-di- hydroxytoluene are generally much lower in culture media than the levels of other proposed metabolites [7,10]. Uptake of labelled 2,5-dihydroxytoluene by P.patulum was found to be slow [ll] and Scott et al. [I21 have recently shown that deuterated 2,5-di- hydroxytoluene is metabolised to deoxyepoxidon by cultures, with no significant incorporation into patu- lin. Under the conditions of P.patulum culture estab- lished in this laboratory, first m-cresol and then m-hydroxybenzyl alcohol were found to reach their maximum concentrations in the culture medium, after 6-methylsalicylic acid levels had reached a maximum [lo]. Preliminary studies have also shown that both m-hydroxybenzyl alcohol and 2,5-dihydroxytoluene are the specific products of m-cresol metabolism by cell-free preparations of P. patulum (Fig. 1). The pres- ent work was therefore undertaken to investigate and compare the nature of the enzymic activities in- Eur. J. Biochem. 49 (1974)
-
Upload
gillian-murphy -
Category
Documents
-
view
212 -
download
0
Transcript of Patulin Biosynthesis: The Role of Mixed-Function Oxidases in the Hydroxylation of m-Cresol

Eur. J. Biochem. 49,443-455 (1974)
Patulin Biosynthesis : The Role of Mixed-Function Oxidases in the Hydroxylation of m-Cresol Gillian MURPHY, Giinter VOGEL, Gunther KRIPPAHL, and Feodor LYNEN Max-Planck-Institut fur Biochemie, Martinsried
(Received May 27/August 28, 1974)
Two hydroxylation reactions of m-cresol, an intermediate in the secondary biosynthesis of the antibiotic patulin, have been characterised in cell-free preparations of Penicillium patulum. The ability to convert m-cresol to 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol is located in separable fractions of the 105000 x g microsomal pellet. The activities had K , values for m-cresol of 80 pM and 5 pM, respectively. Both reactions require molecular oxygen and NADPH for activity. Inhibition of m-cresol hydroxylases by carbon monoxide is reversible by light with a photochemical action spectrum maximal at 450 nm. Carbon monoxide difference spectra of mycel extracts show the presence of absorbing pigments at 422 and 450-455 nm.
The enzymes are not appreciably inhibited by cyanide but are inhibited by cytochrome c. rn-Cresol hydroxylation therefore appears to be catalysed by typical mixed-function oxidases involving an NADPH-dependent reductase and a pigment like cytochrome P-450.
In the light of studies on the utilisation of radioactively labelled intermediates, the methyl hydrox- ylation of m-cresol to m-hydroxybenzyl alcohol probably represents an important reaction in the sequence leading to patulin formation. Ring hydroxylation of m-cresol to 2,5-dihydroxytoluene appears to be a side reaction functional at higher concentrations of m-cresol.
Penicillium patulum synthesises the hemi-acetal lactone, patulin, from acetyl-CoA and malonyl-CoA via the polyketide 6-methylsalicylic acid [l, 21 and secretes it as a secondary end product. The first postulated routes for patulin biosynthesis were based upon the secondary metabolites identified by Bir- kinshaw et al. [3], Tanenbaum and Bassett [4], Bu'- Lock et a/. [5] and others and the establishment of a preferred pathway spans from the serial investigation of metabolite appearance in P.patulum cultures [6,7] to the administration of radioactively labelled de- rivatives and analysis of their utilisation [S]. The detection of m-cresol in continuous culture [5], the presence of significant levels of a specific 6-methyl- salicylic acid decarboxylase [9, lo], and the efficient conversion of m-[2H]cresol or m-['4C]cresol to patu- lin [8,11,12] suggest that rn-cresol is an important intermediate in the biosynthesis of patulin. m-Hydroxy- benzyl alcohol has also been established as major metabolic product [ 131, with efficient conversion to
Enzymes. m-Cresol l-methylhydroxylase and m-cresol 2-hydroxylase, m-cresol, reduced NADP : oxygen oxido- reductases (EC 1.14.13.-).
patulin [ l l , 121. Incorporation of radioactively labelled m-cresol into 2,5-dihydroxytoluene was demonstrated by Scott and Yalpani [8] and by Bu'Lock et al. [7] in cultures treated with protein-synthesis inhibitors, although it has been observed that levels of 2,5-di- hydroxytoluene are generally much lower in culture media than the levels of other proposed metabolites [7,10]. Uptake of labelled 2,5-dihydroxytoluene by P.patulum was found to be slow [ l l ] and Scott et al. [I21 have recently shown that deuterated 2,5-di- hydroxytoluene is metabolised to deoxyepoxidon by cultures, with no significant incorporation into patu- lin. Under the conditions of P.patulum culture estab- lished in this laboratory, first m-cresol and then m-hydroxybenzyl alcohol were found to reach their maximum concentrations in the culture medium, after 6-methylsalicylic acid levels had reached a maximum [lo]. Preliminary studies have also shown that both m-hydroxybenzyl alcohol and 2,5-dihydroxytoluene are the specific products of m-cresol metabolism by cell-free preparations of P. patulum (Fig. 1). The pres- ent work was therefore undertaken to investigate and compare the nature of the enzymic activities in-
Eur. J. Biochem. 49 (1974)

444 Hydroxylation of m-Cresol during Patulin Biosynthesis
& - &::. - OH
m - Cresol 2 ,5 -D ihydroxy to luene ( I ) 6 - Methylsal icyl ic ac id
m - Hydroxybenzyl a lcohol (It) P a t u l i n
Fig. 1 . Metabolism of m-cresol during the biosynthesis of patulin in P. patulum
volved in the conversion of m-cresol to these two fermentor (New Brunswick, model FS-314) and grown hydroxylated derivatives, their location in the cell at 25 "C, aeration rate 6 l/min and stirring rate and possible role in the control of the patulin bio- 150 rev./min for the required time period. Severe synthetic pathway. foaming was prevented by the addition of Niax-
Polyol.
MATERIALS AND METHODS
Materials
Coenzymes and enzymes (Boehringer Mannheim GmbH, Mannheim, Germany), m-hydroxybenzyl alcohol (Th. Schuchardt, Miinchen), 2,5-dihydroxy- toluene (EGA-Chemie, D-7924 Steinheim) and patulin (F. Hoffmann-La Roche and Co.,AG., CH-4002 Basel) were all commercially available. Other chemicals were purchased from E. Merck (Darmstadt). [l-14C]Acetate and [2-14C]acetate were purchased from the Radio- chemical Centre (Amersham, England). Penicillium patulum CBS 384.48 cultures were obtained from the Centvaal Bureau vor Schimmelcultures (Baarn, Hol- land), Bacto agar and yeast extract from Difco Labs, beer wort from the Lowenbrauerei (Miinchen) and Niax-Polyol from Brenntag GmbH. (Miilheim).
Cultivation of P. patulum
Penicillium patulum was grown on agar slopes (1.5 g agar and 2 ml light, unhopped beer wort in 100 ml distilled water) at room temperature and was transferred to fresh medium every month. Spores (1 - 4 weeks old) from one vial transferred to 600 ml Czapek-Dox medium (glucose, 30 g; KH2P04, 2.5 g; K2HP04, 2.5g; NaNO,, 3 g ; MgSO,, 0.5g; KCl, 0.1 g; yeast extract, 3 g and distilled water to 11) in an 1.8 1 Fernbach flask and incubated at 30 "C for 36 h on a rotary shaker (New Brunswick, model G-53) at 100 rev./min. The contents of two such flasks were transferred to 10 1 of the same medium in a 14-1
Isolation and Identification of Metabolites
The medium from fermentor cultures, whole mycel incubations or mycel extract incubations (2- 5 ml) was acidified to approximately pH 1 with 6 N HC1, saturated with sodium chloride and extracted four times with peroxide-free ether (3 ml). The combined ether extracts washed with water (2 ml) and dried over anhydrous sodium sulphate. Dried extracts were evaporated to dryness prior to thin-layer chromatog- raphy or gas-liquid chromatography of derivatives.
Trimethylsilylation was effected with 0.3 ml reagent (trimethylchlorsilane, 1.8 ml ; hexamethylsilazane, 3.2 ml and water-free pyridine to 10 ml [14]). Gas- liquid chromatography separations were carried out in a Varian Aerograph 202 using stainless steel columns (3.175 mm, 3 m long) and helium carrier gas (25 ml/ min). The stationary phase (7.5% NPGS) was sup- ported on Chromosorb (80- 100 mesh). Using a temperature programme with a heating rate of 2 "C/ min and commencing at 120 "C for 10 min, retention times of 6.5, 18, 20, 25.5, 26.5 and 37 min were observed for m-cresol, 2,5-dihydroxytoluene, m-hy- droxybenzyl alcohol, gentisyl alcohol, 6-methylsali- cylic acid and patulin respectively. Estimations of the radioactive content of the separated compounds were made by combustion to 14C02 in a Packard model 325 CuO oven and passing over an anthracene flow- through cell (Tricarb scintillation spectrometer models 3101 and 2001).
Thin-layer chromatography separations of un- treated metabolites were carried out on pre-coated
Eur. J. Biochem. 4Y (1974)

G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 445
F 70 E
2 60
- 50 m 40
2 30
'f 20
.- . 3 0
c 0 ._ +
0 a L
- 0
0
@ 10
0
6- Methy lsa l icy l ic ac id
I I I I I I I
0 1 2 3 4 5 6 7 8 Mycel age (days)
Fig. 2. The incorporation of (I-'4C]acetate into metabolites of the patulin biosynthetic pathway. Mycel, after varying lengths of fermentor growth, were incubated in fresh medium containing [14C]acetate, as described in Materials and Meth- ods. 14C-labelled metabolites excreted into the medium after
silica gel F 254 plates (Merck, 0.2- 0.5-mm thickness) using chloroform - ethyl acetate - ether (5 : 2 : 1, by vol.) as solvent. On 0.2-mm aluminium foil plates RF values of 0.86, 0.72, 0.45 and 0.25 were obtained for m-cresol, 2,5-dihydroxytoluene m-hydroxybenzyl al- cohol and gentisyl alcohol, respectively. Scanning for radioactivity was carried out using a Berthold LB 2722 with a dual ratemeter integrator LB 242 K.
Bromine derivatives of 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol produced by m[14C]cresol hydroxylation and of biosynthesised m- ['4C]cresol were made by preliminary isolation of the labelled compounds by addition of 10 mg of authentic material and reaction in ethanolic solution with a brominating reagent (KBr, 1.5 g; Br,, 1 g and water to 10 ml [15]). Portions of the reaction mixture were chromato- graphed on silica gel (tribromoderivative of m-hy- droxybenzyl alcohol, RF 0,77 in benzene- methanol, 93 : 3, v/v; dibromoderivative of 2,5-dihydroxy- toluene, R, 0.2 in chloroform; tribromoderivative of m-cresol, RF 0.5 in chloroform).
(14C]Acetate Incorporation Mycel grown in submerged fermentor culture for
varying lengths of time, from 1 - 8 days, was filtered from the medium and washed with 100 mM potassium phosphate buffer, pH 6.5. Portions of mycel (200 mg) were incubated in 3 ml of medium containing 100 mM potassium phosphate, pH 6.5, glucose (0.2 g) and sodium [14C]acetate (0.1 pmol, 5 x lo6 counts/min) for 2 h at 30°C. The incubations were then filtered and the mycel washed with water (5 ml). The combined filtrate and washings were supplemented with 200 pg
2 h were extracted and analysed by radioactive gas-liquid chromatography. Note: the specific activity of 6-methyl- salicylic acid is one third greater than that of the other metabolites, formed after decarboxylation of the former
of the various metabolites as carrier before ether extraction, trimethylsilylation and gas-liquid chro- matography separations as described above.
Preparation of m-('4C]Cresol
Mycel grown in submerged fermentor culture for 48 h (Fig. 2) was filtered from the medium and washed with 100 mM potassium phosphate buffer, pH 6.5 and 400-mg portions were incubated in 10 ml of medium containing 100 mM potassium phosphate buffer, pH 6.5,l mM sodium [l-14C]acetate or [2-14C]- acetate (lo9 counts/min) and m-cresol (20 pmol) for 2 h at 30 "C, shaking at 100 rev./min.
The incubation was filtered and the mycel washed with water (1 ml) and acetate ( 4 x 2 ml), the total filtrate and washings were then acidified to pH 1 and saturated with NaCl prior to extraction with ether (3x6ml) . The ether extract was washed with H 2 0 (3 x 5 ml) and dried over anhydrous sodium sulphate. The volume of ether was reduced by rotary evaporation and the concentrated extract spotted over 8 cm of an 0.5-mm-thick silica gel plate prior to chromatography in the solvent system described. The area corresponding to m-cresol was located by ultraviolet fluorescence and by scanning for radioactivity, scraped off and eluted with ether. The ether solution was rotary evaporated to near dryness and the m-[14C]cresol stored at - 15 "C in the dark in methanol.
Purity of the product m-[1,3,5-14C,]cresol or m-[l -Me,2,4,6-'4C4]creso1 was established by gas- liquid chromatography of the trimethylsilyl derivative and thin-layer chromatography both as m-cresol and
Eur. J. Biochem. 49 (1974)

446 Hydroxylation of rn-Cresol during Patulin Biosynthesis
its bromine derivative. In each case the single sym- metrical radioactive peak had the same mobility as authentic m-cresol. m-Cresol content was measured by the absorption at 270 nm, assuming that at pH 7, F = 1400 M-' x cm-' and was adjusted to a specific activity of around 150000 counts/min-' x pmol-'.
Mycel Preparations
Lyophilised mycel, harvested and washed after 48 h of fermentor growth [lo], was broken down by shaking for 90 s in a glass-bead cell homogeniser [17] (dry mycel2.5 g; glass beads, 30 g and buffer, 35 ml), cooled by liquid carbon dioxide. m-Cresol hydroxyl- ases were found to be most stable when prepared in 100 mM potassium phosphate, pH 7.5 containing 10 % (v/v) glycerol and this buffer was used wherever pos- sible. For the sucrose density gradient experiments extracts were prepared in 250 mM sucrose containing 10 mM potassium phosphate, pH 7.5 and in experi- ments involving metal ions, 250 mM sucrose buffered by 20 mM Tris-HC1, pH 7.5 was used.
Extracts were separated from the glass beads and large cell wall fragments by filtering through a linen cloth and were then centrifuged for 15 min at 15 000 x g . The 15 000 x g supernatant was frequently used directly but a particulate fraction was also prepared by further centrifugation at 105000 x g for 30 min. The resulting pellet was resuspended in 4 - 5 ml of the same buffer and contained the majority of the m-cresol hydroxylating activity, although some vari- ation in the distribution of activity between fractions was observed. This was probably due to variations in batches of mycel and the degree of cell breakdown achieved in the cell homogeniser.
Sucrose density gradient centrifugation was carried out using 15000 x g supernatants (3.5 ml; approxi- mately 40 mg protein) layered onto a 9-ml gradient of sucrose (15- 35 %, w/v, containing 10 mM potas- sium phosphate, pH 7.5) with a 40% sucrose base (1 ml) in 14.2-ml cellulose nitrate tubes. Centrifugation was for 1 h at 200000xg (Beckman SW 40 rotor, Spinco model L2 65B). Fractions (1 ml) were collected and assayed for incorporation of m- ['4C]cresol into 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol.
Protein determinations were carried out by the biuret method [17] after precipitation with trichloro- acetic acid. Serum albumin was used as a standard of reference.
Assay of Enzyme Activities
Incubations were carried out in duplicate at 25 "C for 2 - 5 min in 100 mM potassium phosphate buffer,
pH 7.5 or 100 mM Tris-HC1, pH 7.5, generally using 0.5 mM NADPH and 0.2 mM rn-cresol(50000 counts/ min).
Blank values were estimated as the incorporation of m- [14C]cresol into 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol in the absence of added NADPH (< 100 counts/min). In all experiments re- actions were initiated by the addition of enzyme and stopped by the addition of 100 pl of 4 N HC1 to give an approximate pH of 1. The acidified solutions was saturated with NaCl, 1.5 pmol of 2,5-dihydroxy- toluene and m-hydroxybenzyl alcohol were added and extracted with ether (3 x 3 ml). The combined ether extracts were evaporated to dryness in a stream nitrogen and the residue dried by the addition of ethyl acetate (0.2 ml) and re-evaporation. Extracts in a small volume of ether, were then spotted on thin-layer chromatography plates which were developed as before. Regions corresponding to 2,5-dihydroxy- toluene and m-hydroxybenzyl alcohol were detected by their ultraviolet fluorescence and were scraped off into vials containing methanol (2 ml). Toluene scintil- lation fluid (10 ml; PPO, 7 g; POPOP, 0.5 g; toluene, 11) was added prior to measurement in a liquid scintillation spectrometer (Packard model 2405).
Light Reversal of Carbon Monoxide Inhibition
Studies on the inhibition of m-cresol hydroxylation by carbon monoxide were carried out using Warburg manometers. Mixtures of carbon monoxide, oxygen and nitrogen were prepared in a gas burette at atmospheric pressure and passed for 5 min (20 ml/) min) into the shaking Warburg flasks containing the standard incubations medium at 25 "C. Reaction was initiated by tipping the flasks (enzyme in side arm) and stopped by quick cooling and the addition of 0.1 ml of 4 N HCl. In the case of flasks under 100% N,, alkaline pyrogallol (0.4 ml) was included in the flask centre well to absorb traces of oxygen remaining in the system.
Initially, an apparatus consisting of a 250-watt projector lamp. concentrating lens, mirror and CuS04 (1 %, w/v, 5 cm thick) was used to observe light reversal of carbon monoxide inhibition of m-cresol hydroxylation. For further, more specific, investigation of the action of light of specific wavelengths, the method and apparatus of Warburg [18,19] were used with a 900-watt xenon high-pressure lamp as the light source. Flasks were incubated singly in the presence of light of the same intensity but of specific wavelengths varying between 400 and 510 nm, as produced by interference filters. Duplicate incubations were then carried out in the reverse direction, i.e. from 510-
Eur. J. Biochem. 49 (1974)

G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 447
400 nm to cut down errors due to slow inactivation of the enzyme preparation during longer-term experi- ments.
Difference Spectra
Supernatant extracts (15 000 x g) of mycel were diluted with 100 mM potassium phosphate, pH 7.5 containing 10 % (v/v) glycerol and NADPH (0.5 mM) was added. Difference spectra were recorded in a Cary model 14 spectrophotometer by establishing the re- duced versus reduced baseline, bubbling carbon mon- oxide through the sample cuvette for 1 min and re- recording the spectrum (390- 500 nm). A few crystals of sodium dithionite were then added to each cuvette and the spectrum again determined.
RESULTS
Incorporation of (l4C]Acetate into Patulin Metabolites
The sequential incorporation of [14C]acetate into the proposed metabolites of the patulin biosynthetic pathway was observed using P.patulum mycel har- vested after increasing times of fermentor growth. Fig. 2 shows a typical pattern of metabolite labelling observed by incubation of such mycel with [I4]-
acetate and glucose and gas-liquid chromatography analysis of the detectable 14C-labelled compounds subsequently released into the medium. After 6-methyl- salicylic acid, m-cresol was the first maximally-labelled metabolite detectable, followed by peak activity in m-hydroxybenzyl alcohol and a low, but maximal incorporation into 2,5-dihydroxytoluene. Gentisyl alcohol was maximally labelled after a further 24 h, whilst the end product, patulin, did not reach a maxi- mum for another 3 - 4 days. The exact time of maximal incorporation was found to vary with the mycel batch but the same sequence of incorporation was observed. As in the gas-liquid chromatography analysis of the original fermentor medium [lo], no evidence for the presence of gentisaldehyde was obtained. The results show that all the metabolites from 6-methylsalicylic acid to patulin are being synthesised after 1 day of fermentor growth, albeit in various amounts, in- dicating that, at the beginning of the iodophase, all the enzymes necessary for patulin biosynthesis are already present. With increasing time the amount of the individual enzymes, or their activity must be varied to yield the pattern of metabolite labelling observed. The preparation of labelled patulin metabolites from [14C]acetate and the study of individual enzymic reactions was subsequently carried out using mycel of an appropriate age, as indicated by these results.
Characterisation of m-Cresol Hydroxjilases
Incubation of 48-h mycel extracts with m-[14C]- cresol in the presence of NADPH for 30-60 min yielded three hydroxylation products which were shown to be 2,5-dihydroxytoluene, m-hydroxybenzyl alcohol and gentisyl alcohol. Identification was based on the identical mobility of the labelled reaction products and authentic unlabelled material on thin- layer chromatography and the identical retention times of the trimethylsilylated derivatives with au- thentic derivatives during gas-liquid chromatography in several systems. Shorter-term incubation for be- tween 2 and 5 min only yielded detectable amounts of labelled 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol. Preparation of the bromine derivatives of the reaction products and thin-layer chromatography also gave compounds with the same mobility as the bromine derivatives of the authentic compounds.
Using the assay conditions described throughout this paper it was found that incorporation of m-[14C]- cresol by raw mycel extracts, 15 000 x g supernatant preparations and the 105 000 x g microsomal fraction was linear with respect to enzyme concentration. Activity was found to be linear only over a short time period (Fig. 3), according to the stability of the mycel preparation, and assay times were adjusted accordingly. Most raw e)ttracts produced 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol in approximately equal amounts directly after preparation, activities of up to 0.8 pmol x min-’ x g lyophilised mycel-’ being ob- served. However, the ability to incorporate m- [‘“CI- cresol into m-hydroxybenzyl alcohol declined very rapidly and was far more sensitive to preparation conditions than the 2,5-dihydroxytoluene-producing activity. Both activities were extremely unstable at all times. It is therefore probable that the observed activity in vitro is not indicative of that in vivo.
Centrifugation studies on raw mycel extracts in- dicated that both the 2,5-dihydroxytoluene-producing and the m-hydroxybenzyl-alcohol-producing activities were located in the 15 000 x g supernatant fraction, although in some cases a slight loss of activity was observed during this procedure, not totally accountable for by the activity detectable in the pellet fraction. High-speed centrifugation of 15 000 x g supernatant (105000 xg) yielded a pellet fraction with up to 70% of the 2,5-dihydroxytoluene-producing activity and 65 % of the m-hydroxybenzyl-alcohol-producing ac- tivity. The amount of nz-cresol-hydroxylating activity remaining in the 105 000 x g supernatant varied from 3 - 15 % but was generally very low. Recombination of the pellet and supernatant material gave no increase in total m-cresol-hydroxylating activity.
Eur. J. Biochem. 49 (1974)

448 Hydroxylation of m-Cresol during Patulin Biosynthesis
0 10 20 30 40 50 60 Time (rnin)
0 5 10 Time (m in )
Fig. 3. The effect of time on the incorporation of m-['4C]cresol into 2,s-dihydroxytoluene and m-hydroxybenzyl alcohol by preparations of P. patulum. Incubations were carried out for varying lengths of time using (A) 50 pl raw mycel extract; (B) 100 it1 15000 x g supernatant or (C) 50 pl of a microsomal
preparation in a total volume of 2 ml containing 100 mM potassium phosphate buffer, pH 7.5, 0.5 mM NADPH and 0.2 mM m-['4C]cresol at 25 "C. (0) 2,5-Dihydroxytoluene; (0) m-hydroxybenzyl alcohol
600 - c .- E
2 500 . 3 0 u - g 400 a, = 0
V
- L. 300 2 I
x" 200 e
0 100
n Lz x
L" N
- 0 L 0 " m -
F
0 ' I I I 10 A ! ; A A 10 12 14 Gradient volume (m l ) I I
I
15 Sucrose (%)
35 - Fig. 4. Sucrose density gradient centrlfugation oj P. patulum mycel extract to separate the 2,s-dihydroxytoluene-producing and m-hydroxybenzyl-alcohol-producing activities. Super- natant extracts were centrifuged on sucrose density gradients as described under Materials and Methods. Fractions (1 ml) were assayed for their ability to incorporate m-[14C]cresol into 2,5-dihydroxytoluene (0) and m-hydroxybenzyl alcohol (0)
On the assumption that the 105 000 x g pellet frac- tion was equivalent to a similarly prepared microsomal fraction from mammalian tissues, preliminary studies on alternative methods of microsome preparation [20] were made with respect to the recovery of m-cresol hydroxylases. The use of 8 mM CaClz on 15 000 x g supernatant preparations precipitated a fraction with 35 "/, of the original 2,5-dihydroxytoluene-producing activity but only 5 % of the m-hydroxybenzyl-alcohol- producing activity, leaving 10 % and 3 %, respectively,
in the supernatant. 8 mM MgCl, yielded a similar result. It was also observed that the fraction pre- cipitated by the use of 5 % (w/v) poly(ethyleneglyco1) 6000 yielded up to 80% of the 2,5-dihydroxytoluene- producing activity and 10 % of the m-hydroxybenzyl- alcohol-producing activity. These investigations in- dicate that the rn-cresol-hydroxylating activities of P. patulum preparations are membrane-bound to a fraction not unlike the microsomal fraction described for mammalian tissues. Due to the great instability of these activities, however, only conventionai centri- fugation methods yield a membrane fraction with sufficient activity to enable further studies of the system. It was shown that centrifugation of material up to 250000 x g did not yield any further m-cresol- hydroxylating activity in the pellet fraction.
Sucrose density gradient centrifugation of 15 000 x g mycel extract supernatants at 300 000 x g for 1 h separated the particles bearing the 2,5-di- hydroxytoluene-producing activity from those capable of incorporating m- ['4C]cresol into m-hydroxybenzyl alcohol, the latter sedimenting much more slowly in the gradient (Fig. 4). The activities were, however, not separable in a CsCl,, sucrose gradient designed for the separation of rough and smooth micro- somes [21]. Both 15000xg supernatant and micro- soma1 preparations from the mycel extracts were used to compare the requirement of the two hydroxylation reactions for the substrate m-cresol. Both activities showed typical Michaelis-Menten kinetics (Fig. 5) and were not inhibited by up to 1 mM m-cresol. The m-hydroxybenzyl-alcohol-producing activity gave an approximate K, for m-cresol of 5 pM whereas a K, of 80 pM was calculated for the 2,5-dihydroxytoluene-
Eur. J . Biochem. 49 (1974)
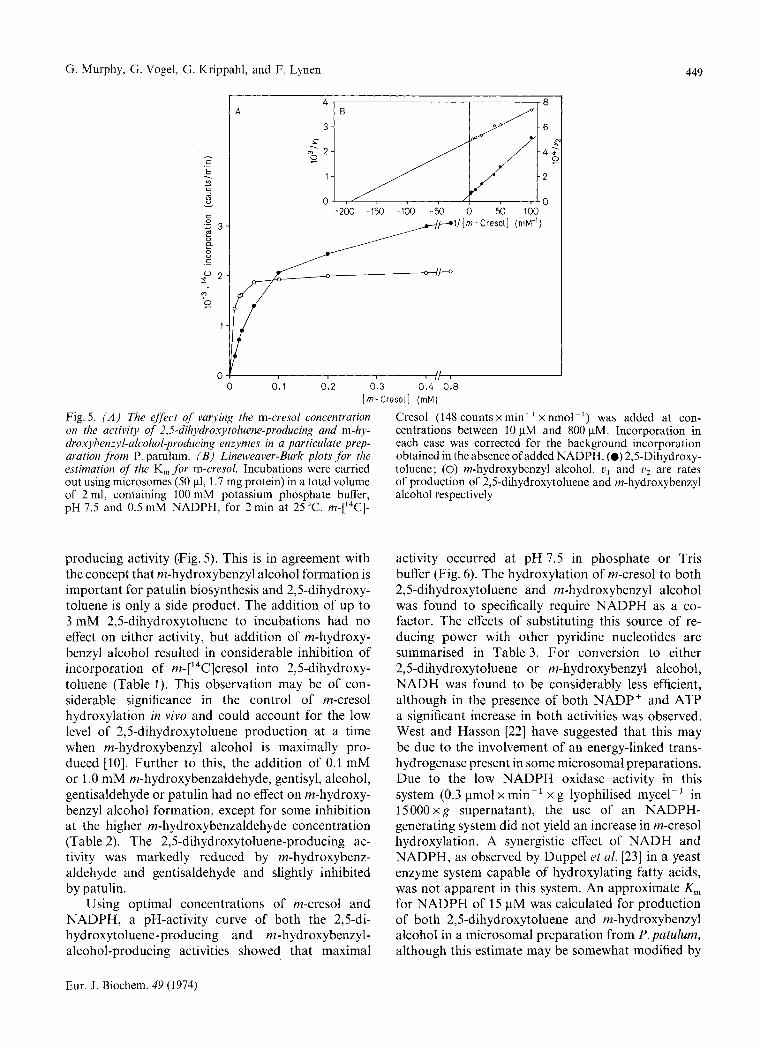
G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 449
Fig. 5. ( A ) The effect of varying the m-cresol concentration on the activity of 2,5-dil~ydroxytoluene-producing and m-hy- droxyhenzyl-alcohol-producing enzymes in a particulate prep- aration from P. patulum. ( B ) Lineweaver-Burk plots for the estimation of the K, for m-cresol. Incubations were carried out using microsomes ( S O pl, 1.7 mg protein) in a total volume of 2 ml, containing 100 mM potassium phosphate buffer, pH 7.5 and 0.5 mM NADPH, for 2 min at 25 "C. m-['4C]-
producing activity (Fig. 5). This is in agreement with the concept that m-hydroxybenzyl alcohol formation is important for patulin biosynthesis and 2,5-dihydroxy- toluene is only a side product. The addition of up to 3 mM 2,5-dihydroxytoluene to incubations had no effect on either activity, but addition of m-hydroxy- benzyl alcohol resulted in considerable inhibition of incorporation of m-['4C]cresol into 2,5-dihydroxy- toluene (Table 1). This observation may be of con- siderable significance in the control of m-cresol hydroxylation in vivo and could account for the low level of 2,5-dihydroxytoluene production at a time when m-hydroxybenzyl alcohol is maximally pro- duced [lo]. Further to this, the addition of 0.1 mM or 1 .0 mM m-hydroxybenzaldehyde, gentisyl, alcohol, gentisaldehyde or patulin had no effect on m-hydroxy- benzyl alcohol formation, except for some inhibition at the higher m-hydroxybenzaldehyde concentration (Table 2). The 2,5-dihydroxytoluene-producing ac- tivity was markedly reduced by m-hydroxybenz- aldehyde and gentisaldehyde and slightly inhibited by patulin.
Using optimal concentrations of m-cresol and NADPH, a pH-activity curve of both the 2,5-di- hydroxytoluene-producing and m-hydroxybenzyl- alcohol-producing activities showed that maximal
Cresol (148 counts x min-' x nmol-') was added at con- centrations between 10 pM and 800 pM. Incorporation in each case was corrected for the background incorporation obtained in the absence of added NADPH. (0) 2,s-Dihydroxy- toluene; (0) m-hydroxybenzyl alcohol. u, and l j 2 are rates of production of 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol respectively
activity occurred at pH 7.5 in phosphate or Tris buffer (Fig. 6). The hydroxylation of m-cresol to both 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol was found to specifically require NADPH as a co- factor. The effects of substituting this source of re- ducing power with other pyridine nucleotides are summarised in Table 3. For conversion to either 2,5-dihydroxytoluene or m-hydroxybenzyl alcohol, NADH was found to be considerably less efficient, although in the presence of both NADP' and ATP a significant increase in both activities was observed. West and Hasson [22] have suggested that this may be due to the involvement of an energy-linked trans- hydrogenase present in some microsomal preparations. Due to the low NADPH oxidase activity in this system (0.3 pmol x min-' x g lyophilised mycel-' in 15000xg supernatant), the use of an NADPH- generating system did not yield an increase in m-cresol hydroxylation. A synergistic effect of NADH and NADPH, as observed by Duppel et al. [23] in a yeast enzyme system capable of hydroxylating fatty acids, was not apparent in this system. An approximate K,,, for NADPH of 15 pM was calculated for production of both 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol in a microsomal preparation from P. patulum, although this estimate may be somewhat modified by
Eur. J. Biochem. 49 (1974)

450 Hydroxylation of m-Cresol during Patulin Biosynthesis
Table 1. The effect of the presence of excess products on the incorporation of m-['4C]cresol into 2,5-dihydroxytoluene ( I ) and m-hydroxybenzyl alcohol (11) Incubations were carried out using 100 p1 of a mycell5000 x g supernatant fraction as described under Materials and Methods. The results are expressed as a percentage of the incorporation into the hydroxylation products of rn-cresol, 2,5-dihydroxytoluene (I) and rn-hydroxybenzyl alcohol (11) under the optimal assay conditions with no added product
Added product (concn) Relative incorporation into :
I I1
None - 100 100 I (3.0) 106 99 I1 (0.1) 58 103 I1 (1.0) 19.5 108 111 (5.0) 7.0 93
Table 2. The effect of other proposed patulin pathway meta- bolites on the incorporation of m-['4C]cresol into 2,s-dihydroxy- toluene ( I ) and m-hydroxybenzyl alcohol ( I I ) Incubations were carried out as described in the text at two concentrations of various compounds shown to be of significance in the biosynthesis of patulin, consequent to rn-cresol hydroxylation. The results are expressed as a per- centage of the incorporation into the hydroxylation products of m-cresol, 2,s-dihydroxytoluene (I) and m-hydroxybenzyl alcohol (II), under optimal assay conditions
~ ~~ ~
Added metabolite (concn) Relative incorporation into :
I I1
None -
m-H ydroxybenz- aldehyde (0.1)
(1 .0) Gentisyl alcohol (0.1)
(1.0) Gentisaldehyde (0.1)
(1 .0) Patulin (0.1)
(1.0) (3.0)
100 100
42 97.5 6 43
104 104 100 102 86.5 105 32 97.5 97.5 98.5 95.5 100 63 101
the low background oxidation of the reduced pyridine nucleotide.
Incorporation of rn- ['4C]cresol into 2,5-dihydroxy- toluene and m-hydroxybenzyl alcohol was also found to be dependent on the presence of oxygen. Incuba- tions carried out in an atmosphere of oxygen-free nitrogen showed negligible hydroxylating activity with respect to incubations in air or oxygen-nitrogen
0 1 6.5 7.0 7.5 8.0 8.5
PH Fig. 6. pH-activity curves for u particulute preparation of m-cresol hydroxylases. Microsomes (50 pl; 1.4 mg protein) were incubated for 2 min at 25 "C in the presence of potassium phosphate and Tris-HCI buffers of varying pH with 0.2 mM m-[14C]cresol and 0.5 mM NADPH. (w) Incorporation into m-hydroxybenzyl alcohol; ( O d ) incorporation into 2,s-dihydroxytoluene
mixtures (Table 4). Oxygen (100%) gave no significant increase in activity when substituted for air in the gas phase.
The observed requirements for both NADPH and molecular oxygen for the hydroxylation of m-cresol suggested that the reactions are catalysed by mixed-function oxidases with the involvement of an electron donor and an electron acceptor, respec- tively [24]. Preliminary experiments were carried out to characterise the possible type of mixed-function oxi- dation involved in these two hydroxylation reactions. As summarised in Table 5, removal of metal ions by EDTA or sodium diethyldithiocarbamate had no effect on activity, ferrous ions had a small inhibitory effect, but ferric ions were less effective. Tetrahydro- folic acid and ascorbate were not active as electron donors in this system and catechol had no effect either in catalytic or in greater quantities as observed in the case of polyphenol oxidase [25]. Bubbling carbon monoxide through the incubation mixture for 1 min resulted in 100% inhibition of all rn-cresol hydroxyl- ation activity, implicating the involvement of a factor similar to cytochrome P-450 in the activation of oxygen during the reaction. This was supported by the observation that m-cresol hydroxylation to form 2,5-dihydroxytoluene was only slightly inhibited by cyanide ions at a very high concentration whilst the rn-hydroxybenzyl alcohol-forming activity was totally unaffected, in contrast to many cyanide-sensitive mixed-function oxidases which not involve cytochrome
Eur. J. Biochem. 49 (1974)

G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 451
Table 3. Effect of pyridine nucleotides on m-('4C/cresol hydroxylation by cell-free preparations of P. patulum Reactions were carried out in a total volume of 2 ml in the assay system described under Materials and Methods using 100 pI of 15 000 x g supernatant of a mycel extract. Incorpora- tion in the absence of any added cofactor was taken as a blank. Results are expressed as a percentage of the final corrected incorporation of radioactivity into 2,5-dihydroxytoluene (I) and in-hydroxybenzyl alcohol (11) under optimal conditions
Pyridine nucleotide (concn) Relative incorporation into :
I I1
NADPH NADH NADH
NADH + ATP
+ ATP + NADP'
+ Glc-6-P + Glc-6-P dehydro- genase
NADP'
NADPH + NADH
3 00 14
13
37
90
103
Table 4. Effect of varying gas mixtures on the ability* of P. patulum extracts to hydroxylate m-('4C/cresol Reactions were carried out in a Warburg apparatus containing the standard incubation medium in the main compartment and 100 pl of 15000 x g supernatant extract in the side arm. The effects of different gas mixtures are expressed as a per- centage of the incorporation of m-['4C]cresol into 2,5-di- hydroxytoluene (I) and in-hydroxybenzyl alcohol (11) in the presence of air, taken as 100 %
Gas phase Relative incorporation into:
I IT
100 - 95 5 - 100
6 3 100 99 102 97
P-450. Cytochrome c (70 enzymically reducible), however, caused a considerable fall in hydroxylation activity, suggesting that NADPH-cytochrome c re- ductase present in the microsomal fraction can act as a competitor to NADPH-cytochrome P-450 reductase for electrons from NADPH. The former enzyme has been shown to act as an NADPH cytochrome P-450 reductase [26] and may be identical to it [27,28]. Inhibition by cytochrome c was also partially reversed
Table 5. The action of various ejfectors of mixed-function oxidases on the hydroxylation of m-cresol by P. patulum extracts Incubations were carried out using particulate preparations (50 pl; 1.4- 1.8 mg protein) in the standard assay system
described in the text. Results are expressed as a percentage of the incorporation of m-[14C]cresol under optimal con- ditions at 0.5 mM NADPH into 2,5-dihydroxytoluene (I) and m-hydroxybenzyl alcohol (11) and are the average of three separate determinations. 0.5 mM NADPH was present unless indicated otherwise
Added effector (concn) Relative incorporation into :
% - EDTA (Et),NCS,Na" (Et)2NCS2Na a
Fez +
Fez+ Fez +
Fe3 +
Fe3 +
Fe3 +
H,folate H,folate Ascorbate Ascorbate Catechol Catechol Catechol Catechol Carbon monoxide Cytochrome c KCN KCN Cytochrome c
+ KCN
Mg2+
100 112 96 80
114 87 54
1 96 80 24 0
97 103
0 100
2 95 2 0
45 86 27
64
100 107 102 92
113 96 54 0
96 95 69 0
95 100
0 99
1 98 1 0
39 107 102
89
a Sodium diethyldithiocarbamate. Tetrahydrofolate. No NADPH present. Saturating concentration.
by cyanide ions, probably due to the inhibition of electron transport through the NADPH-cytochrome c reductase pathway.
Table 6 shows the effects of varying amounts of carbon monoxide relative to oxygen in the gas phase of incubations on the production of 2,5-dihydroxy- toluene and m-hydroxybenzyl alcohol. The results indicate a competition between carbon monoxide and oxygen for reaction with a component, such as cytochrome P-450, of the rn-cresol-hydroxylating system.
Irradiation of such incubations with a strong light filtered through a copper sulphate solution (trans-
Eur. J. Biochem. 49 (1974)

452 Hydroxyhtion of m-Cresol during Patulin Biosynthesis
Table 6. The inhibitory effect of carbon monoxide on the hydroxylation of’ m-[’4C]cresol by P. patulum extracts and its reversal by light Dark and light reactions were carried out in the presence of various ratios of carbon monoxide to oxygen in the gas phase of Warburg manometers, as described in the text, using 15000xg supernatant extracts of mycel in a 2-ml incubation at optimal substrate concentration. Results are expressed as a percentage of the incorporation measured with air as the gas phase (taken as 100 %) into 2,5-dihydroxytoluene (I) and m-hydroxybenzyl alcohol (11)
Gas phase CO/02 Relative incorporation in -
dark light
co 0, I I1 I I1 ~~
x ”/,
5 2.5 2: 1 26 15 55 41 5 5 1 : l 35 38 12 65
10 5 2: 1 21 21 61 58 20 5 4: 1 13 25 56 45
400 420 440 460 480 500 Wavelength, X (nm)
Fig. I . Photochemicul action spectrum for the light reversal of cuvbon monoxide inhibition of m-cresol hydroxylation to 2,5- dihydroxytoluene and m-hydroxybenzyl alcohol. Incubations were carried out using 15000xg supernatants from mycel extracts (100 pl) in the standard assay system and in a gas phase of 10% carbon monoxide, 5% oxygen and 85% nitrogen. The ratio of light sensitivity a t a given wavelength, L;., to the light sensitivity a t 450 nm is plotted as a function of wavelength (see text). Each point represents the average of two determinations, carried out in duplicate. (0) 2,5-Di- hydroxytoluene ; (0) m-hydroxybenzyl alcohol
mitting between 380 and 520 nm) caused significant reversal of the carbon monoxide inhibition (Table 6). Subsequent to this a photoactivation spectrum for the oxidation of m-cresol to 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol was measured as an estimate
of the wavelength of maximal absorbance of the carbon-monoxide-binding pigment participating in the reaction. Fig.7 shows the effect of various wave- lengths of light of the same intensity on the reversal of carbon monoxide inhibition of m-cresol hydroxyl- ation. The maximum reversibility occurred with light of around 450 nm, consistent with the principle of a CO-binding pigment like cytochrome P-450.
An attempt was made to measure the amount of CO-binding pigment present by the difference spec- trum method of Omura and Sat0 [29], comparing the spectra of 15000 x g mycel extract supernatants in the presence and absence of carbon monoxide after reduction by NADPH or by sodium dithionite. Two peaks were observed (Fig.8) at 422nm and 450- 455 nm in the presence of NADPH, both being slightly enhanced by sodium dithionite addition. The peak at 450 - 450 nm was generally very much smaller than that at 422 nm and, in some cases, was not detectable due to difficulties caused by interference by the high protein concentrations employed. It was therefore not possible to make any accurate estimates of the total of cytochrome P-450 content of the extracts. High-speed centrifugation at 15 000 x g supernatants and estimation of carbon monoxide difference spectra on the 105 000 x g supernatant and resuspended pellet material indicated that the carbon-monoxide-binding pigment at 422 nm was soluble in nature. No 450-nm absorbing material was detectable in either fraction. The 422-nm absorbing pigment could be analogous to the solubilised cytochrome P-450 occurring in many mammalian microsomal preparations [29,30].
Several preliminary studies were made in an attempt to release the m-cresol-hydroxylating activities or material like cytochrome P-450 from the P.patulum microsomal fraction. Procedures used by various groups to release cytochrome P-450 from animal microsomes were tried, including sodium deoxycholate treatment [30] and sonication followed by sodium cholate digestion [31] but total loss of activity and carbon monoxide binding of the cytochrome P-450 pigment occurred. NADPH-cytochrome c reductase activity was also monitored as it has been implicated in the mechanism of cytochrome P-450 oxidation, due to its similarity to NADPH-cytochrome P-450 reductase. Only 30% of the total activity detectable in the original 15000 x g mycel supernatant was recoverable from the 105000xg pellet and 10% of the initial activity was released into the supernatant on digestion of the microsomal fraction with sodium deoxycholate, whilst no activity was measurable in the remaining precipitate.
It is possible that the loss of enzymic activity observed during such procedures reflects the need for some degree of membrane integrity for the activity
Eur. J. Biochem. 49 (1914)

G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 453
Wavelength ( n r n ) Fig. 8. Difference spectra of P. patulum extracts. These were determined as described in Materials and Methods and re- present the difference in absorption of a mycel supernatant preparation (0.95 ml; 12.3 mg protein in a total volume of
of m-cresol hydroxylases. Alternatively, the cyto- chrome P-450 involved may be extremely labile to such procedures and this renders the enzymes in- operative.
DISCUSSION
The conversion of m-cresol to the hydroxylated products, 2,5-dihydroxytoluene and m-hydroxybenzyl alcohol, is catalysed by particulate enzymes which show the characteristic requirements of mammalian microsomal mixed-function oxidases for molecular oxygen and a reduced pyridine nucleotide, the most efficient being NADPH. The participation of a pigment like cytochrome P-450 in these hydroxylation reactions, as in many other mono-oxygenases, was indicated on observing their inhibition by carbon monoxide and reactivation by light, the most effective wavelength being 450 nm.
In common with other mixed-function oxidases dependent on cytochrome P-450, cyanide ions do not interfere with electron transport in the m-cresol hydroxylatory system, but cytochrome c inhibits ac- tivity by initiating NADPH cytochrome c reductase activity and the removal of reducing equivalents from NADPH. Difference spectra studies of P. patulum cell- free extracts indicated a small, but significant ab- sorption by a CO-binding pigment at 450-455 nm, but this component was extremely labile and not accurately measurable. The function of cytochrome
2 ml) before and after treatment with carbon monoxide in the presence of NADPH ( 0 4 ) or sodium dithionite (O----a) as a reducing agent. Reduced plus carbon mon- oxide minus reduced spectrum)
P-450 appears to be in the oxygen-activating step of both mammalian [32] and bacterial [33] mixed-func- tion oxidation reactions. Acting in conjunction with an NADPH-dependent reductase and phospholipid, cytochrome P-450 has been shown to confer specific- ity towards individual substrates of a mixed-function oxygenase system [34] and several distinct forms have been isolated [35,36].
Similarly, the 2,5-dihydroxytoluene-producing and m-hydroxybenzyl-alcohol-producing activities of P. patulum could be catalysed by enzymes differing in the type of cytochrome P-540 involved, each with a specific affinity for m-cresol (K, values in Fig. 5 ) and associated with different membrane fractions (Fig. 4). The K, for NADPH was identical for both hydroxyl- ases, as would be the case if a common NADPH- dependent reductase was involved.
Forrester and Gaucher [37] have discussed in detail the possibilities for the regulation of the patulin bio- synthetic pathway in the light of the knowledge to date of metabolite flow and individual enzyme activity. Studies on coarse control, that is regulation of the total amount of enzyme present, have indicated the probability that the enzymes converting m-cresol to patulin are sequentially induced by newly appearing metabolites [5,7] at the onset of the fungal iodophase. Fine control of specific enzymic activities of the path- way has yet be studied in any detail and necessitates the isolation and characterisation of individual en- zymes. The difficulty of obtaining stable preparations has precluded rapid progress in this area. Forrester
Eur. J. Biochem. 49 (1974)

454 Hydroxylation of m-Cresol during Patulin Biosynthesis
and Gaucher [37] have proposed a role for m-hydroxy- benzyl alcohol dehydrogenase as a rate-determining step in the major biosynthetic pathway to patulin. The product of the reaction, m-hydroxybenzaldehyde, has been shown to be a more efficient precursor of patulin than gentisaldehyde [ll, 121. However, this would not account for the observation that under certain culture conditions gentisyl alcohol and patulin are alternative metabolites[38] or the high levels of [14C]acetate incorporation into gentisyl alcohol that we have observed, unless two routes are operative together or separately under different growth con- ditions.
A possible control is indicated by our observation that m-hydroxybenzyl alcohol inhibits 2,5-dihydroxy- toluene production, reflected by the low levels of 2,5-dihydroxytoluene found in the fermentor medium. The fact that the two m-cresol-hydroxylating activities are found in association with different membrane fractions, separable by sucrose density centrifugation, suggests two separate enzymes. m-Cresol hydroxyl- ation to 2,5-dihydroxytoluene could simply be a side reaction of an enzyme mainly concerned with the hydroxylation of m-hydroxybenzyl alcohol to form gentisyl alcohol ; hence the presence of m-hydroxy- benzyl alcohol would initiate the main activity, crucial to the function of the patulin pathway, resulting in a slow down in 2,5-dihydroxytoluene formation. End- product, allosteric inhibition of an enzyme or enzymes on the pathway would represent an important means of rapid control of metabolism but no effect of other pathway metabolites on m-hydroxybenzyl production was observable.
Further understanding of the pathways for the biosynthesis of patulin and establishment of the major synthetic route will be possible only when all the individual enzymes involved have been fully charac- terised in terms of their activity with respect to their specific substrates at steady state concentrations in vivo and the action of various effectors, including other pathway metabolites. The more chemical studies of the main flux of metabolites through the patulin path- way should provide an important indicator of the sequence and importance of reactions involved in the total process.
The problems involved in the evaluation of patulin biosynthesis are common to all studies of secondary metabolism. A large number of metabolites and en- zymic activities can be observed, only a few of which are involved in one, or perhaps two major biosynthetic routes. Many of the metabolites are in equilibrium with true intermediates or are the result of the non- specificity of the enzymes in the system, catalysing reactions with substrates of slightly varying structure and creating small side routes to the end product.
True intermediates may also be membrane-bound or protein-bound and never detectable, thus obscuring the correct metabolic pathway [39].
G. Murphy is the recipient of a Royal Society NATO science fellowship.
REFERENCES 1.
2. 3.
4.
5.
6.
7.
8.
9.
10. 13.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24. 25.
26.
Bu’Lock, J. D. & Ryan,A. J. (1958) Proc. Chem. Soc.
Lynen, F.&Tada, M. (1961). Angew. Chem. 73,513-519. Birkinshaw, J. H., Bracken, A. & Raistrick, H. (1943)
Biochem. J . 37, 726-728. Tanenbaum, S. W. & Bassett, E. W. (1958) Biochim. Bio-
phys. Acta, 28,21- 31. Bu’Lock, J. D., Hamilton, D., Hulme, M. A,, Powell,
J . A,, Smalley, J. A,, Shepherd, D. & Smith, G. N. (1965) Can. J . Microbiol. I l , 765.
Bassett, E. W. & Tanenbaum, S. W. (1958) Biochim. Bio- phys. Acta, 28, 247-260.
Bu’Lock, J. D., Shepherd, D. & Winstanley, D. J. (1969) Can. J . Microbiol. 15, 279-285.
Scott, A. I. & Yalpani, M. (1967) Chem. Commun. 0, 945 - 947.
Light, R. J. (1969) Biochim. Biophys. Acta, 191, 430- 438.
Vogel, G. (1971) Ph. D. Thesis, University of Miinchen. Forrester, P. I. & Gaucher, G. M. (1972) Biochemistry,
Scott, A. I., Zamir, L., Phillips, G. T. & Yalpani, M.
Rebstock, M. C. (1964). Arch. Biochem. Biophys. 104,
Pierce, A. E. (1970) Silylation of Organic Compounds, Pierce Chemical Co., Rockford.
Shriner, R. L., Fuson, R. C. & Curtin, D. Y. (1956) The Systematic Identification of Organic Compounds, pp. 264-265, J. Wiley and Sons, New York.
Merkenschlager, M., Schlossmann, K. & Kurz, W. (1957) Biochem. Z. 329,332- 340.
Gornall, A. J., Bardawill, C. J. & David, M. M. (1949) J. Biol. Chem. 177,751 - 766.
Warburg, 0. (1949) Heavy Metal Prosthetic Groups and Enzyme Action, Ch. XI1 and XIII, Oxford University Press, London.
Rosenthal, 0. & Cooper, P. Y . (1967) Methods Enzymol.
Kamath, S . A. & Naraydn, K. A. (1972) Anal. Biochem.
Bergstrand, A. & Dallner, G. (1969) Anal. Biochem. 29,
West, C. A. & Hasson, P. E. (1973) Arch. Biochem. Bio-
Duppel, W., Lebeault, J. M. & Coon, M. J. (1973) Eur.
Hayaishi, 0. (1969) Annu. Rev. Biochem. 38, 21-44. Malmstrom, B. G. & Rydh , L. (1968) in Biological
Oxidations (Singer, T. P., ed.) pp. 415-438, Inter- science Pub]., J . Wiley and Sons, New York.
Sato, R., Omura, T. & Nishibayashi, H. (1965) in Oxi- dases and Related Redox Systems (King, T. E., Mason, H. S. & Morrison, M., eds) p. 861, J. Wiley and Sons, New York.
186-216.
11, 1102-1107.
(1973) Bioorg. Chem. 2, 124- 139.
156-159.
10, 616-629.
48, 53-61.
351 - 356.
phys. 155,258 - 269.
J . Biochem. 36, 583 - 592.
Eur. J. Biochem. 49 (1974)

G. Murphy, G. Vogel, G. Krippahl, and F. Lynen 455
27. Omura, T., Sanders, E., Estabrook, R. W., Cooper, D. Y. & Rosenthal, 0. (1966) Arch. Biochem. Biophys.
28. Masters, B. S. S., Bacon, J., Taylor, W. E., Isaacson, E. L. & LoSpalluto, J. (1971) J . Biol. Chem. 246, 4143- 4150.
29. Omura, T. & Sato, R. (1964) .I. Biol. Clzem. 239, 2370
30. Lu, A. Y. H. & Coon, M. J. (1968) J. Biol. Chem. 243,
31. Autor, A. P., Kaschnitz, R. M., Heidema, J. K. & Coon, M. J. (1972) Mol. Pharmacol. 9, 93-104.
32. Coon, M. J., van der Hoeven, T. A,, Kaschnitz, R. M. & Strobel, H. W. (1973) Ann. N.Y. Acad. Sci. 212, 449 - 457.
117,660-673.
- 2378.
1331 - 1334.
33. Gunsalus, I.C., Tyson,C.A., Tsai, R. & Lipscomb, J.D. (1971) CIiem.-Biol. Interactions, 4, 75-78.
34. Lu, A. Y. H., Kuntzmann, R., West, S., Jacobson, M. & Conney, A. H. (1972) J . Biol. Chem. 247, 1727- 1734.
35. Whysner, J. A,, Ramseyer, J., Kazmi, G. M. & Harding, B. W. (1969) Biochem. Biophys. Res. Commun. 36,
36. Comai, K. & Gaylor, J. L. (1973) J . Biol. Chem. 248,
37. Forrester, P. I. & Gaucher, G. M. (1972) Biochemistry,
38. Brack, A. (1947) Helv. Chim. Acta, 30, 1 - 14. 39. Turner, W. B. (1971) Fungal Metabolites, pp. 20-23,
795-801.
4947-4955.
11, 1108-1114.
Academic Press, London.
G. Murphy, G. Krippahl, and F. Lynen, Max-Planck-Institut fur Biochemie, D-8033 Martinsried, Am Klopferspitz, Federal Republic of Germany G. Vogel’s present address : Max-Planck-Institut fur Biologie, D-7400 Tubingen, CorrensstraBe 38, Federal Republic of Germany
Eur. J. Biochem. 49 (1974)


















