
PARÁSITOS DE LA PERDIZ ROJA: IMPLICACIONES PARA SU ... · La perdiz roja (Alectoris rufa) es una...
130
PARÁSITOS DE LA PERDIZ ROJA: IMPLICACIONES PARA SU APORVECHAMIENTO CINEGÉTICO Y CONSERVACIÓN Diego Villanúa Inglada
Transcript of PARÁSITOS DE LA PERDIZ ROJA: IMPLICACIONES PARA SU ... · La perdiz roja (Alectoris rufa) es una...
PARÁSITOS DE LA PERDIZ ROJA: IMPLICACIONES PARA SU APORVECHAMIENTO
CINEGÉTICO Y CONSERVACIÓN
Diego Villanúa Inglada

PARÁSITOS DE LA PERDIZ ROJA: IMPLICACIONES PARA SU APORVECHAMIENTO CINEGÉTICO Y CONSERVACIÓN.
MEMORIA PRESENTADAS POR
Diego Villanúa Inglada
PARA OPTAR AL TÍTULO DE DOCTOR.
VºBº DE LOS DIRECTORES
DR. CHRISTIAN GORTÁZAR DRA. URSULA HÖFLE
UNIVERSIDAD DE CASTILLA-LA MANCHA
INSTITUTO DE INVESTIGACIÓN EN RECURSOS CINEGÉTICOS
(CSIC – UCLM – JCCM)
DEPARTAMENTEO DE CIENCIA Y TECONOLOGÍA AGORFORESTAL
Junio 2007

Para la realización de este trabajo se ha contado con la financiación del
Convenio entre el Principado de Asturias y el CSIC

AGRADECIMIENTOS
No es fácil aceptar que tu hijo te llene la casa de egagrópilas, plumas,
cangrejos, culebras, tortugas, ratoneros, lechuzas, picarazas, cardelinas..., que
en lugar de jugar al fútbol como el resto de zagales se pegue el día subiendo a
los árboles o con las redes japonesas de Paño, que te haga construir una
pajarera y una balseta para las tortugas...en fin, no es fácil. A pesar de todo se
le perdona porque, al fin y al cabo, a lo hijos hay que quererlos ¿no?. El límite
llega cuando, después de haber acabado la carrera y haber conseguido trabajo
de veterinario en una clínica de Huesca con horario de tienda y pijama verde,
pilla el crío y se te marcha a Ciudad Real, de becario precario a estudiar las
cagarrutas de las perdices..., eso es que no hay quien lo entienda. Pues no,
ellos sí lo entendieron y lo respetaron, y es más, en lugar de darme una colleja
para que se me fuesen los pájaros de la cabeza, me animaron a continuar por
este camino tan raro pero tan bonito que he escogido. Por eso, por haber
estado siempre ahí, dándome todo lo que tenían y más, incluidas muchas dosis
de paciencia, y haberme permitido optar por esta vida, tengo que estar
agradecido a mis padres. Gracias.
Tres cuartos de lo mismo para el resto de mi sufrida familia, en especial a
mi hermana Lidius, porque la dejé en Sangarrén siendo una crieta y cuando
volví ya se me había convertido en una mujerona. Por esas horas que le
correspondían a ella y que tuve que pasar en Zaragoza o en Ciudad Real, y
que nunca me ha echado en cara, también gracias. También para Javier por
aquello de “yaye vamo al pampo”, los primeros prismáticos y la primera guía.
En fin, a todos. La verdad es que la familia no se elige y es una suerte que te
toque una de la que te puedas sentir tan orgulloso. Gracias.
En la facultad, hay mucha gente a la que agradecer muchas cosas, Lluís
Luján me echó una importantísima primera mano para poder comenzar con
esto del bicho de monte en la Alfranca y en la sala de necropias. La gente de
AVAFES fue una segunda familia para mi, tantas cervezotas juntos, tantos
cursos... y que decir de Javier Lucientes, cuanto frío anillando en el carrizal de
Sangarrén, pobre buenazo que se dejó engañar por el que subscribe para que
le dirigiese una tesina con “las mosquetas” de los Monegros. Sin su apoyo ni se

me habría pasado por la cabeza llamar a las puertas de IREC, y esta Tesis
nunca habría visto la luz.
De la época del IREC, el primero debe estar Christian Gortázar; que se dejó
engañar y dio una oportunidad al coletas de Avafes que había hecho la tesina
con “papa Lucientes”. Es de agradecer la paciencia que ha demostrado tener
conmigo todo este tiempo, y la confianza en que al final saldrían las cosas (yo
dudé de mi capacidad en más de una ocasión). Más de lo mismo para Úrsula
Höefle, la cual, a pesar de andar siempre corriendo de aquí para allá, supo
sacar el tiempo necesario para enseñarme a ver algo más que tripas en la sala
de necropsias, a ser “Eduardo manos-pipetas” y por hacer inteligible mi
“inglés”. Con el resto del clan del submundo; Pela, Xuacu, Isabel, Chute, Paqui,
Vane, Moni, Eli, Mariclón, Oscarín, Manolín y Pablo me une mucho más que
una amistad, somos desde luego “hermanos de sangre” (y de bazos, de
riñones, de garrapatas...). Fuera de coñas, viajaré a otras partes, conoceré
mucha gente, pero veo difícil que en ningún sitio pueda llegar a sentir más “en
casa” que con vosotros, gracias. De los primos de ecología y de genética
mucho y bueno, muchos conteos de todo tipo de bicho viviente, muchos cafés
en los que se aprendía más que en algunas charlas de catedráticos, muchas
cañitas con tapa, esos marcapáginas finos de Rouconen, en fin, lo dicho,
mucho y bueno. Y mención especial para el clan del micro pollo con Sarva
Quillo Ariquitaung, Fabi el torero churriego, el Triquiñuelas terror de la noche
culiparda y sobre todo Loren, que tanto ha aportado al desarrollo de esta tesis.
Y si en Ciudad Real estuve bien, que no decir de Asturies. Cuanto que
agradecer a Maria, Emilio, Pilar y Pepe, que me acogieron como a uno más de
su familia y que mostraron paciencia infinita cada vez que, en lugar de cortarme
la coleta por llenarles la cuadra de vísceras de corzo, jabalí, ciervo, zorro u
otras lindezas, me esperaban con quesín de ese bueno de la peral, sidra
abundante y “sarandonga” para arreglar el mundo. Gracias.
Aunque me estoy alargando demasiado, no puedo dejar de nombrar a los
compañeros de Navarra; Isabel, Jóse, Javi, Alex, Enrique y el resto de gente de
Viveros, con los que continúo disfrutando de mi trabajo y de los que cada día
aprendo algo y a Lorenzo Bintanel, que puso el toque artístico con los dibujos
de esta tesis.

Y por último, a Esther, porque me quiere mucho, por que la quiero más y
porque ha sufrido como nadie la elaboración de esta tesis. Resulta difícil no
mandar a paseo a un novio al que sólo ves dos días al mes durante 3 años y
que además te regala fines de semana románticos necropsiando jabalíes en
Riglos, corzos en el alto La Campa o perdices de Miguelturra, pero ella se
mantuvo fiel a su ONG de “nadie sin novia” y me ha aguantado hasta hoy.
Espero que lo siga haciendo por mucho tiempo y que yo sepa recompensarla
por todo lo que me da.
Gracias a todos.

1
INDICE
I. ESTRUCTURA DE LA TESIS…………………………………. 3
II. INTRODUCCIÓN GENERAL………………..……………….. 4 II.1. Descripción y distribución de la especie…………………………… 4
II.2. Importancia ecológica y socioeconómica…..………………........... 6
II.3. Tendencia de sus poblaciones……………………………………… 8
II.4. Factores implicados en la disminución de las poblaciones………10
II.5. El papel de las enfermedades………………..………………………11
II.6. Enfermedades y parásitos de la perdiz roja……………………….. 13
II.7. Las repoblaciones como herramienta de gestión………………..... 22
II.8. Bibliografía……………..………………………………………………. 27
III. OBJETIVOS……………………………………………………………… 38
IV. CAPITULO 1…………………………………………………………….. 41
Las repoblaciones con aves de granja como focos de
introducción de nuevos parásitos en las poblaciones silvestres
de perdiz roja.
V. CAPITULO 2……………………………………………………………… 56
Variaciones en el resultado de los análisis coprológicos
preventivos actuales en función del tipo de heces analizadas y
de la hora de recogida de las mismas.
VI. CAPITULO 3…………………………………………………………….. 77
Efectividad de los tratamientos antiparasitarios como método
para prevenir la introducción de nematodos en el campo.

2
VII. CAPITULO 4……………………………………………...…………….. 86
Posible transmisión de nematodos propios perdices de granja
a especies amenazadas.
VIII. CAPITULO 5……………………………………………………...……. 91
Factores limitantes de la abundancia estival de perdiz roja en
Aragón. Posibles alternativas a las repoblaciones con aves de
granja.
IX. SÍNTESIS…...……………………………………………………….....… 114
X. CONCLUSIONES, PERSPECTIVAS Y REFLEXIONES….… 121

3
I. ESTRUCTURA DE LA TESIS
La presente tesis doctoral está dividida en introducción, objetivos, una serie
de capítulos (1-5) que constituyen el cuerpo de la tesis, una síntesis general y
las conclusiones. Se incluyen además unas breves perspectivas y reflexiones
al final del trabajo.
En la introducción se revisan los antecedentes y el contexto del tema. A
continuación se exponen los objetivos de la tesis, a desarrollar en los sucesivos
capítulos. Cada uno de estos capítulos reproduce el texto íntegro, en inglés, de
manuscritos enviados para su publicación en revistas científicas internacionales
(indicándose si es un manuscrito enviado, aceptado o ya publicado y su
referencia). Previamente a dicho texto en inglés, en cada capítulo se presenta
un resumen en castellano. Finalmente, en la síntesis general, se discuten los
resultados más relevantes de cada capítulo y se enuncian las principales
conclusiones, perspectivas de trabajo y reflexiones que se extraen de la
presente tesis.

4
II. INTRODUCCIÓN GENERAL
II.1. Descripción y distribución de la especie
La perdiz roja (Alectoris rufa) es una galliforme de entre 300 y 500 gramos,
de aspecto compacto y hábitos gregarios fuera de la temporada de
reproducción. Su coloración se compone de tonos castaños en el dorso,
obispillo y pecho gris azulado, vientre canela y plumas costales con un
característico diseño formado por una banda blanca, una negra y una marrón.
La cara es de color blanco y está enmarcada por un collar negro que se inicia
en el pico, atraviesa el ojo y se prolonga hasta la garganta formando un babero.
La parte inferior de este collar se prolonga por el lateral del cuello y parte del
pecho en forma de pequeñas listas negras no presentes en el resto de
especies del género alectoris. Tanto el pico como la carúncula ocular y las
patas son de un rojo vivo, que da nombre a la especie.
La gran plasticidad ecológica que presenta la perdiz roja le ha permitido
ocupar multitud de hábitats diferentes, desde el nivel del mar hasta los prados
alpinos por encima de los 2.000 metros (Mullarney et al., 2001). No obstante, el
hábitat ideal para la especie lo constituyen las llanuras cerealistas con
alternancia de otros cultivos, como viñas u olivares, y campos en barbecho. Los
paisajes mejores son aquellos en los que se haya mantenido la distribución en
parcelas de pequeño tamaño y la conservación de márgenes de vegetación
natural (Cheylan, 1976; Garcia et al., 1983; Lartiges y Mallet, 1983; Berger,
1984 y 1987; Gaudin y Ricci, 1987; Lucio y Purroy, 1987; Birkan, 1990; Lucio
1991; Reudet, 1992; Blanco-Aguiar et al., 2003). Estas peculiaridades se
cumplen en grandes áreas de la zona centro-sur de la Península Ibérica, donde
se alcanzan las mayores densidades para la especie (Blanco Aguiar et al.,
2003).
Dentro del género Alectoris se incluyen otras 6 especies más (A.magna, A.
melanocephala, A.philbyi, A. graeca, A. chukar, y A. barbara) distribuidas a lo
largo del paleártico occidental (Cramp y Simmons 1980, Figura 1). La
distribución natural de la perdiz roja abarca la mayor parte de la Península
Ibérica, sur de Francia y una pequeña franja del noroeste de Italia (Mullarney et

5
al., 2001). A esta distribución natural hay que añadir la población del sureste de
Inglaterra, originada a partir de unas primeras sueltas de perdices francesas en
1.770 (Tapper, 1992) y mantenida, probablemente, gracias al continuo aporte
de aves de granja (Tapper, 1999).
Tradicionalmente se han diferenciado tres subespecies de perdiz roja: A.
rufa rufa, A. r. hispanica y A. r. intercedens. La primera de ellas distribuida por
el sur de Francia y norte de Italia, así como en las poblaciones introducidas del
Reino Unido; la segunda, ocuparía la franja noroeste de la Península Ibérica,
constituida por Cataluña, Aragón, Navarra, País Vasco, Cantabria, Asturias,
Galicia, Castilla León y el norte de Extremadura; y la tercera y última
subespecie, sería la presente en el resto del área de distribución (McGowan,
1994; Díaz et al. 1996).
Figura 1. Distribución de las distintas especies del género Alectoris tomado de
Cramp y Simmons (1980).

6
II.2. Importancia ecológica y socioeconómica.
El mediano tamaño y la abundancia de esta especie, hacen que alrededor
de 40 especies de predadores la incluyan en su dieta (Yanes et al., 1998),
constituyendo, junto con el conejo de monte, la dieta de especies tan
amenazadas como el águila imperial (Aquila adalberti) (Delibes e Hiraldo, 1981;
Calderón, 1983). A este papel como presa de especies amenazadas hay que
añadir el papel que desempeña la perdiz como “especie paraguas” para los
ecosistemas agrícolas tradicionales, unos de los más amenazados en la
actualidad (De la Concha et al., 2006), ya que algunas de las mejoras del
hábitat realizadas por el colectivo de cazadores para la perdiz tienen un efecto
muy positivo sobre otras especies vinculadas a este tipo hábitat tales como el
sisón (Tetrax tetrax) o las poblaciones de aláudidos (De la Concha et al., 2006).
No obstante, este efecto positivo sobre otras especies puede cambiar de
sentido en aquellas zonas más intensamente manejadas ya que, dentro de las
actuaciones efectuadas se incluyen en ocasiones el control ilegal de los
depredadores o la suelta de perdices híbridas o portadoras de patógenos que
podrían incorporarse al medio (Millán et al., 2004 a y b; Villanúa et al., 2007).
Por último hay que considerar también la gran importancia que tiene por sí
misma la conservación de la perdiz roja al tratarse de una de las especies más
típicas y emblemáticas de los ambientes mediterráneos de la Península Ibérica,
punto de origen de la especie y casi reducto exclusivo de sus poblaciones
naturales (Cramp y Simmons, 1980).
A este indudable valor ecológico y conservacionista hay que añadir la gran
importancia económica y social que la caza de esta especie tiene en la
Península Ibérica. Esta práctica constituye una de las actividades económicas
más importantes en multitud de áreas rurales de nuestro país (APROCA, 1998;
Lucio, 1998; Bernabeu, 2000) y tiene un arraigo cultural y social único (Delibes
1963 y 1988). A modo de ejemplo cabe señalar que en la provincia de Ciudad
Real, se ha estimado que la caza de esta especie genera más de 200 millones
de euros anuales (Otero, 1995), si bien estos grandes ingresos no siempre son
declarados.

7
II.3. Tendencia de sus poblaciones.
Las poblaciones de perdiz roja parecen estar sufriendo una marcada
regresión en las últimas décadas (Cramp y Simmons, 1980). Este descenso ha
sido registrado tanto en su área de distribución natural en Francia (ONC, 1986),
Italia (Baratti et al., 2005) y Península Ibérica (Rueda et al., 1992; Borralho et
al., 1998; Lucio, 1998; Blanco Aguiar et al., 2003), como en la población
introducida en el Reino Unido (Aebischer y Potts, 1994). Esta tendencia, unida
a su limitado área de distribución, ha hecho que la perdiz roja esté considerada
actualmente como especie de estatus “Vulnerable” a nivel mundial (Aebischer y
Potts, 1994) y haya sido declarada SPEC 2 por Birdlife International (Tucker y
Heath, 1994).
Centrándonos en los trabajos acerca de la evolución de sus poblaciones en
la Península Ibérica, hay que hacer referencia al fuerte descenso registrado en
Castilla y León (Lucio, 1998). Para un periodo de menos de 20 años, se calculó
una disminución superior al 70 % en las tablas de caza, pasando de una media
de 12 perdices capturadas por kilómetro cuadrado a finales de los 1970 a tan
sólo 2 en los 1990 (Lucio, 1998). En el mismo sentido van los datos
demográficos más recientes, obtenidos mediante los censos del programa
SACRE, llevados a cabo por voluntarios de SEO/Birdlife en España. Estos
datos sugieren una reducción demográfica del 20 % entre 1996 y 2001 (Blanco
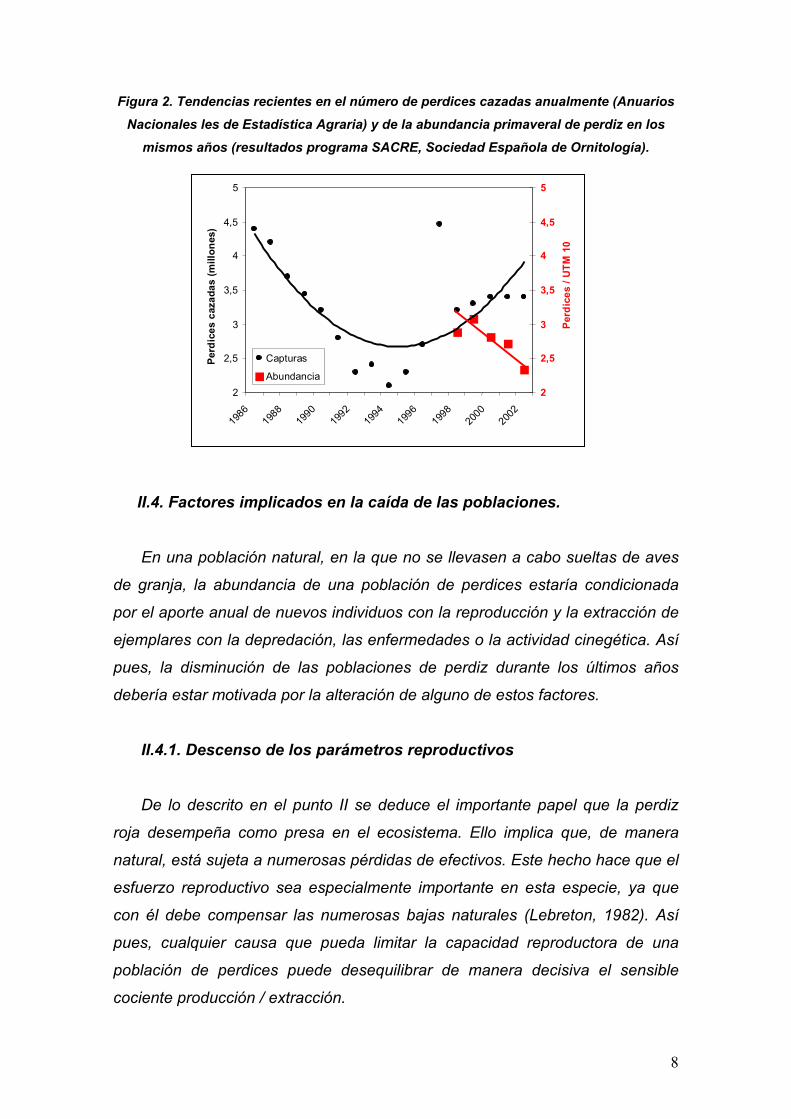
Aguiar et al., 2003; Figura 2). Paradójicamente, las estadísticas oficiales de
caza sugieren una notable recuperación en las capturas a partir de mediados
de los 90, que parecen tender a estabilizarse en torno a los 3.5 millones
anuales (Baragaño y Otero 2001; Figura 2).
No obstante, hay que tener en cuenta que anualmente se liberan al campo
varios millones de perdices criadas en granja, de manera que esta aparente
recuperación en las tablas de caza podría ser tan sólo el reflejo de tales
sueltas.
8
Figura 2. Tendencias recientes en el número de perdices cazadas anualmente (Anuarios Nacionales les de Estadística Agraria) y de la abundancia primaveral de perdiz en los
mismos años (resultados programa SACRE, Sociedad Española de Ornitología).
II.4. Factores implicados en la caída de las poblaciones.
En una población natural, en la que no se llevasen a cabo sueltas de aves
de granja, la abundancia de una población de perdices estaría condicionada
por el aporte anual de nuevos individuos con la reproducción y la extracción de
ejemplares con la depredación, las enfermedades o la actividad cinegética. Así
pues, la disminución de las poblaciones de perdiz durante los últimos años
debería estar motivada por la alteración de alguno de estos factores.
II.4.1. Descenso de los parámetros reproductivos
De lo descrito en el punto II se deduce el importante papel que la perdiz
roja desempeña como presa en el ecosistema. Ello implica que, de manera
natural, está sujeta a numerosas pérdidas de efectivos. Este hecho hace que el
esfuerzo reproductivo sea especialmente importante en esta especie, ya que
con él debe compensar las numerosas bajas naturales (Lebreton, 1982). Así
pues, cualquier causa que pueda limitar la capacidad reproductora de una
población de perdices puede desequilibrar de manera decisiva el sensible
cociente producción / extracción.
2
2,5
3
3,5
4
4,5
5
1986
1988
1990
1992
1994
1996
1998
2000
2002
Perd
ices
caz
adas
(mill
ones
)
2
2,5
3
3,5
4
4,5
5
Perd
ices
/ U
TM 1
0
Capturas
Abundancia2
2,5
3
3,5
4
4,5
5
1986
1988
1990
1992
1994
1996
1998
2000
2002
Perd
ices
caz
adas
(mill
ones
)
2
2,5
3
3,5
4
4,5
5
Perd
ices
/ U
TM 1
0
Capturas
Abundancia2
2,5
3
3,5
4
4,5
5
1986
1988
1990
1992
1994
1996
1998
2000
2002
Perd
ices
caz
adas
(mill
ones
)
2
2,5
3
3,5
4
4,5
5
Perd
ices
/ U
TM 1
0
Capturas
Abundancia2
2,5
3
3,5
4
4,5
5
1986
1988
1990
1992
1994
1996
1998
2000
2002
Perd
ices
caz
adas
(mill
ones
)
2
2,5
3
3,5
4
4,5
5
Perd
ices
/ U
TM 1
0
Capturas
Abundancia

9
Una de las causas de fallo reproductivo más frecuente en galliformes es la
depredación tanto de huevos y pollos, como de adultos incubando (Rands,
1988; Yanes et al., 1998). La perdiz roja no es una excepción, tal y como
confirman los trabajos de radio-seguimiento realizados en Francia por Ricci et
al. (1990) y Leonanrd y Reitz (1998). En estos estudios se confirma un alto
porcentaje de pérdida de nidadas (de un 61% y 59% respectivamente), debidas
principalmente a la depredación (>90%). Esta alta tasa de depredación tiene
mucho que ver con las preferencias de la especie a la hora de ubicar el nido.
Así, la perdiz roja selecciona para nidificar linderos, setos o bordes de cultivo
(Rands, 1986 a; Ricci et al., 1990) en lugar de las manchas más espesas de
matorral, donde su vulnerabilidad frente a la predación podría ser menor
(Carvalho y Borralho, 1998). Estas estructuras son peores a la hora de
asegurar el éxito reproductivo por varios motivos; por una parte, se trata de
estructuras lineales donde la tasa natural de depredación es mayor (Angelstam,
1986) y por otra, son formaciones susceptibles de ser modificadas a lo largo del
año por las prácticas agrícolas (Duarte y Vargas, 2002).
La estrecha relación existente entre productividad de la perdiz y agricultura
es considerada por multitud de autores como uno de los puntos clave del
descenso de las poblaciones (Potts 1980; Lartiges y Mallet, 1983; Rands, 1987;
Pepin y Blayac, 1990; Nadal et al., 1996; Crick et al., 1994; Aebischer y
Kavanagh, 1997; Lucio, 1998, Borralho et al., 2000; Gortázar et al., 2002a) ya
que, durante los últimos años, se ha llevado a cabo una transformación radical
de las prácticas agrícolas, coincidente en el tiempo con la caída de las
poblaciones de perdiz. Las concentraciones parcelarias efectuadas en gran
parte del territorio han simplificado enormemente el hábitat agrícola,
implantando el monocultivo y reduciendo los márgenes necesarios para la
ubicación de los nidos y para la protección necesaria frente a depredadores.
Además, los recientes avances tecnológicos han permitido reducir el tiempo
requerido para realizar las distintas labores, a la vez que se han desarrollado
variedades de cultivo de ciclo más corto. Con estas modificaciones, el impacto
sobre las especies ligadas al medio agrícola es mucho mayor, sobre todo
durante la cosecha, ya que, en el momento en que ésta se lleva a cabo,
muchas puestas están todavía sin eclosionar, lo que origina su pérdida por la
acción directa de las labores agrícolas (Crick et al., 1994; Green 1995).

10
II.4.2. Incremento de la pérdida directa de ejemplares
Pueden distinguirse tres causas por las cuales una población de perdices
puede “perder ejemplares”: la depredación, la caza y las enfermedades. Dentro
de la primera puede distinguirse entre la depredación sobre adultos y sobre
pollos, que no se iguala hasta los 2-3 meses de edad (Hudson y Rands, 1988).
En lo concerniente a factores que puedan afectar a la depredación sobre
pollos, cabe el papel que juega la disponibilidad de insectos, base de su dieta
en los primeros días de vida (Rueda et al., 1993). Teniendo esto en cuenta, es
lógico pensar que a menor disponibilidad de insectos, mayor será el esfuerzo
de búsqueda. Ello implicaría más desplazamientos y por lo tanto más
probabilidades de ser detectado por los depredadores. Esta hipótesis es
barajada por diversos autores que encuentran una relación significativa del
descenso del éxito reproductivo de la aves ligadas al medio agrícola con el uso
de insecticidas (Rands, 1986 b; Potts, 1986; Campbell et al., 1997; Brickle et
al., 2000) o con aquellas variaciones climáticas capaces también de disminuir
la disponibilidad puntual de insectos (Lucio, 1990).
Transcurrida esta primera etapa, las distintas polladas de perdiz se
agrupan formando bandos de mayor tamaño, que ofrecen una mayor
probabilidad de supervivencia ante el frío y la lluvia, y aumentan la capacidad
de vigilancia frente a depredadores (Putaala et al., 1995). A partir de este
momento la depredación pierde protagonismo como causa de muerte de la
perdiz, lugar que pasa a ocupar la caza (Duarte y Vargas, 2002).
La caza, entendida como la extracción racional de los excedentes de unos
recursos renovables que serían las poblaciones de fauna silvestre, no tendría
por qué suponer un riesgo para la conservación de dichas especies, ya que,
como se dice en su propia definición, sólo se extraerían los excedentes (Ley
4/1989, de 27 de marzo, de la Conservación de los Espacios Naturales y de la
Flora y Fauna Silvestres). Sin embargo, es conocido el caso de la paloma
migratoria (Ectoistes migratorius), la cual, a pesar de ser considerada como el
ave más abundante del mundo llegó a extinguirse en parte debido a su caza
indiscriminada (Dorst, 1971). Así pues, parece evidente que la actividad

11
cinegética puede ser capaz de mermar las poblaciones de las especies
cinegéticas si no se gestiona correctamente.
En el caso de la perdiz roja, numerosos autores han considerado la
sobreexplotación cinegética de la especie como uno de los principales
responsables del descenso de las poblaciones (Potts, 1986; Pepin y Blavac,
1990; Lucio y Purroy, 1992; Borralho et al., 1997). Un ejemplo cercano es el de
Portugal, donde, a raíz de la Revolución de los Claveles, la gran mayoría de la
superficie del país se convirtió en terreno libre de caza. Esta situación produjo
una clara sobrecaza de las poblaciones de esta especie, que la llevó a una
situación crítica (Borralho et al., 1997). Desde 1988 una nueva ley promovió el
establecimiento de cotos sociales o privados, así como incentivos para la
gestión y mantenimiento de caza sostenible en estos acotados. Desde dicha
fecha, se fueron estableciendo progresivamente los acotados, y en 1996
alrededor del 30 % de la superficie del país se encontraba ya dentro de esa
figura (Borralho et al., 1997). Esta nueva situación legal probablemente ha
permitido una recuperación parcial de las poblaciones portuguesas de perdiz
en los últimos años (Borrahlo et al., 1997 y 2000).
II.5.El papel de las enfermedades.
Hasta hace pocos años no se había prestado demasiada atención al papel
de las enfermedades en las especies silvestres. Sin embargo, éstas pueden
suponer un importante problema para la conservación y gestión de la fauna.
Basta con recordar el efecto que dos enfermedades víricas, la mixomatosis y la
enfermedad vírica hemorrágica, han tenido sobre las poblaciones de conejo de
la Península (Muñoz-Goyanes, 1960; Villafuerte et al., 1994); e, indirectamente,
sobre las de sus depredadores (Moreno y Villafuerte, 1995; Villafuerte et al.,
1998), para comprender que las enfermedades juegan un papel igual de
relevante que la disponibilidad de refugio y alimento o la predación.
En lo concerniente al papel de las enfermedades como limitantes de las
poblaciones de aves, destacan los trabajos realizados con el nematodo
parásito del ciego, Trichostrongylus tenuis, y el Lagópodo escocés (Lagopus
lagopus scoticus). Ya en 1963, Jenkins y colaboradores encontraban que los
lagópodos con mayor intensidad de infección por este parásito mostraban un
12
0
500
1000
1500
1987 1988 1989 1990 1991 1992 1993 1994 1995 1996
Lagó
podo
s ca
zado
s
Zona controlZona tratada
menor éxito reproductivo, aunque no llegaban a aclarar si el parásito era la
causa de esta menor tasa reproductiva o si, por el contrario, el aumento de la
intensidad de parasitación era tan sólo una consecuencia más de otro factor
desconocido. Posteriormente, Potts y colaboradores (1984), profundizaron en
el conocimiento de la relación existente entre T. tenuis y el Lagópodo escocés,
llegando a la conclusión de que dicho nematodo estaba íntimamente
relacionado con los ciclos de abundancia que mostraba esta tetraónida, aunque
nuevamente, sin aclarar la causalidad. Potts y colaboradores hicieron hincapié
en la necesidad de llevar a cabo trabajos de desparasitación experimental que
permitiesen clarificar el posible papel del nematodo T. tenuis en la dinámica
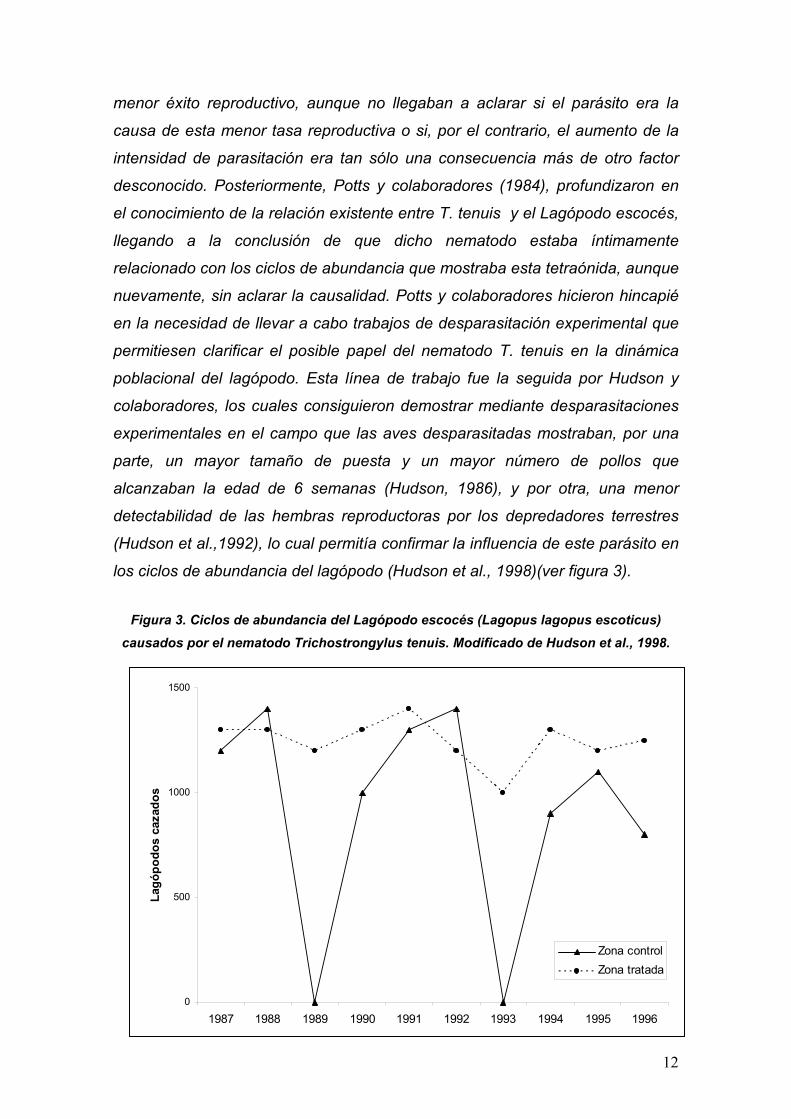
poblacional del lagópodo. Esta línea de trabajo fue la seguida por Hudson y
colaboradores, los cuales consiguieron demostrar mediante desparasitaciones
experimentales en el campo que las aves desparasitadas mostraban, por una
parte, un mayor tamaño de puesta y un mayor número de pollos que
alcanzaban la edad de 6 semanas (Hudson, 1986), y por otra, una menor
detectabilidad de las hembras reproductoras por los depredadores terrestres
(Hudson et al.,1992), lo cual permitía confirmar la influencia de este parásito en
los ciclos de abundancia del lagópodo (Hudson et al., 1998)(ver figura 3).
Figura 3. Ciclos de abundancia del Lagópodo escocés (Lagopus lagopus escoticus) causados por el nematodo Trichostrongylus tenuis. Modificado de Hudson et al., 1998.

13
Estos trabajos zanjaron la clásica discusión sobre las enfermedades como
factor de mortalidad compensatorio (si no las hay, las aves morirán de otras
causas) o aditivo (en ausencia de enfermedades habría más aves). El
experimento puso de manifiesto la capacidad de un patógeno aparentemente
inofensivo para mermar una población de aves mediante la reducción de su
capacidad reproductiva y un aumento de la depredación de aves adultas.
Pero existen también casos en los que la propia enfermedad es capaz de
causar un número alto de bajas y suponer un factor limitante para la población.
A modo de ejemplo cabe citar varios casos recientes y ocurridos en la
Península Ibérica; el brote de Tricomoniasis (Trichomonas gallinae) descrito en
paloma torcaz (Columba palumbus) en el sur de la península, en el cual se
produjo la muerte de más de 2.000 aves (un 15% del total de la población) en
apenas unos días (Höfle et al., 2004) o el de viruela aviar (Avipoxvirus) en una
población de perdiz roja del sur de la Península Ibérica, donde encontraban
más de un 40% de aves afectadas (Gortázar et al., 2002b; Buenestado et al.,
2004).
Queda patente el hecho de que las enfermedades son uno de los factores a
tener en cuenta a la hora de analizar las causas de regresión de una especie.
II.6.Enfermedades y parásitos de la perdiz
A pesar de que las limitaciones logísticas y económicas dificultan el estudio
en profundidad de muchas de las enfermedades que afectan a la perdiz, lo cual
hace que la información publicada sea todavía escasa, a continuación se
tratará de resumir las principales afecciones descritas en esta especie.
II.6.1. Enfermedades de etiología vírica
Dentro de las afecciones de etiología vírica descritas en la perdiz roja
destaca sin duda el virus de la viruela aviar.
Causado por un Avipoxvirus (familia Poxviridae), se presenta en dos formas
clínicas; una forma cutánea, caracterizada por la aparición de nódulos
proliferativos de aspecto verrugoso sobre la piel desprovista de plumas, y una
forma diftérica, que consiste en la aparición de lesiones díftero-necróticas en la

14
mucosa de la boca y de las vías respiratorias y digestivas altas. La primera de
estas presentaciones suele provocar escasas bajas, pero si se generaliza o
deriva en la forma diftérica, puede dar lugar a una gran mortalidad.
La transmisión puede ser directa, por contacto con descamaciones,
secreciones o heces, a través del agua o por medio de dípteros, que pueden
actuar como “vectores” mecánicos. La entrada del virus tiene lugar a través de
pequeñas erosiones o heridas de la piel, mucosa oral o respiratoria donde se
multiplica, dando origen a las lesiones características.
Esta enfermedad ha sido descrito en Europa en numerosas especies de
aves incluída la perdiz roja, para la que se han descrito casos en España con
más de un 40% de aves afectadas (Gortázar et al., 2002b, Buenestado et al.
2004).
Existen otras muchas afecciones víricas descritas en la perdiz roja tales
como la enfermedad de New Castle, la enfermedad de Gumboro, la
laringotraqueítis infecciosa, la bronquitis infecciosa o la enfermedad de West
Nile, pero su frecuencia de presentación es mucho menor (ver Tabla 1).
Tabla 1. Principales afecciones causadas por virus descritas en la perdiz roja (*Enfermedad de declaración obligatoria).
Enfermedad Etiología Importancia Referencias
Viruela aviar
Avipoxvirus Descrito en perdices
silvestres en España. Altas mortalidades
Gortázar et al., 2002b Buenestado et al., 2004
New Castle Paramyxovirus tipo 1 Descrito en perdices. EDO*
Randi et al., 1985 OIE, 2007
Gumboro
Birnavirus
Plan de erradicación. Altas mortalidades. EDO*
Sacristán et al., 2005 OIE, 2007
Laringotraqueítis
Herpesvirus alfa Descrita en perdiz. EDO*
OIE, 2007
Cabeza Hinchada
Pneumovirus Descrito en perdices silvestres en España.
Pizarro y Elices 1997
Bronquitis infecciosa
Coronavirus Descrita en perdiz. EDO*
Cavanagh, 2005 OIE, 2007
Gripe aviar Virus Influenza Descrita en perdiz. EDO*
Humberd et al., 2006 OIE, 2007
West Nile
Flavivirus Zoonosis altamente transmisible. Recientemente descrita en A. chuka.r
Hayes et al., 2005. OIE, 2007 CDC, 2007

15
II.6.2. Enfermedades de etiología bacteriana.
Entre aquellas de etiología bacteriana, sobresaldría tanto por su
frecuencia como por gravedad la colibacilosis.
Esta enfermedad está producida por la enterobacteria Escherichia coli,
presente de manera natural en el intestino de animales sanos, pero capaz de
ocasionar graves mortalidades cuando prolifera en exceso o cuando aparecen
cepas especialmente virulentas.
Esta variable virulencia, así como las diferentes localizaciones de la
bacteria van a condicionar la patogenia, síntomas o lesiones detectados. Los
procesos que con mayor frecuencia van a afectar a las aves de cría o voladero
cursan de la forma siguiente:
Septicémica: Proceso rápido que conduce a la muerte del animal en un
tiempo muy breve. Las lesiones más comunes se observan en el hígado con
pequeños focos blanquecinos y músculos pectorales congestivos. Las canales
están en buen estado de carnes lo que sugiere la rapidez del proceso.
Digestiva: Se caracteriza por inapetencia, fiebre, plumas erizadas y
abatimiento. Los animales presentan una diarrea intensa, blanquecina o
negruzca, pastosa y mal oliente. Las lesiones se corresponden con una
enteritis catarral aguda o hemorrágica, con presencia ocasional de depósitos
de fibrina en hígado, sacos aéreos y pericardio.
Coligranuloma: Proceso de evolución lenta que solo afecta a las aves,
principalmente perdices y codornices. Clínicamente no se observa un cuadro
evidente, salvo adelgazamiento progresivo, disminución del consumo de
pienso, diarrea rebelde, y trastornos respiratorios que resultan en un goteo de
bajas continuo. Todo ello lleva a que esta forma clínica sea un hallazgo de
matadero o necropsia, en la que se observan nódulos blanco-amarillentos de
tamaño variable (0,1-5 cm. de diámetro) que se disponen en cuentas de collar y
tienen consistencia variable. Los coligranulomas se localizan en órganos como
intestino, molleja, ciego, pulmones, hígado, etc.
El contagio puede tener lugar a través del huevo, por vía respiratoria, por
aguas o alimentos contaminados por lo que la higiene y desinfección es
fundamental para evitar infecciones.

16
Esta afección es, sin lugar a dudas, la primera causa de bajas en las
granjas de perdiz roja en España donde se controla mediante el uso, a menudo
inadecuado, de antibióticos, lo cual da lugar a numerosas antibioresistencias
que dificultan su control y que obligan en ocasiones a realizar el vacío sanitario
de la explotación afectada.
A nivel de campo, solamente aparecen casos en aves procedentes de
sueltas para ojeos o caza intensiva, pero nunca en aves autóctonas.
Aunque menos frecuentes, existen otras dos enfermedades descritas en la
perdiz roja y que, por ser transmisibles al hombre, merecen ser consideradas.
Una sería la Salmonelosis, causada por S. typhimurium, S. pullorum y S.
gallinarum (Pennycot et al., 2002), especies todas transmisibles al hombre y
que producen graves diarreas en perdices de granja.
La otra zoonosis a tener en cuenta es la Tuberculosis aviar, producida por
Mycobacterium avium y descrita en perdices silvestres del sur de la Península
Ibérica (Millán et al., 2004c).
Otras enfermedades bacterianas descritas en la perdiz roja son la
pasterelosis, la clamidiosis, la micoplasmosis o el coriza contagioso pero su
importancia en esta especie es mucho menor (ver Tabla 2).
Tabla 2. Principales enfermedades causadas por bacterias descritas en la perdiz roja. (*Enfermedad de declaración obligatoria).
Enfermedad Etiología Importancia Referencias
Colibacilosis Escherichia coli. Frecuente en granjas.
Resistencias. Revilla et al., 2006 Johnson et al., 2003
Salmonelosis Salmonella typhimurium, S. pullorum y S. gallinarum
Zoonosis. Frecuente en granja.
Sharma et al., 1980
Tuberculosis aviar
Mycobacterim avium Zoonosis. Descrita en perdices silvestres.
Millán et al (2004c)
Clostridiosis
Clostridium perfringens Zoonosis. Descrita en galliformes silvestres.
Stuve et al. 1992
Clamidiosis
Chlamydia psittaci Zoonosis. Descrita en galliformes.EDO*.
Erbeck y Nunn 1999 OIE, 2007
Pasterelosis
Pasteurella multocida tipo A, serotipos 3, 5 y 7
Descrita en perdices. Frecuente en granja. Graves pérdidas.
Cucco et al., 2007
Micoplasmosis Mycoplasma gallisepticum, M. imitans, o M. synoviae
Descrita en perdices. EDO*.
Cookson and Shivaprasad 1994
OIE, 2007

17
II.6.3. Enfermedades de etiología parasitaria.
Dentro de las enfermedes producidas por parásitos destacaría por su
frecuencia de presentación la infección por cocciddios en aves de granja
(APROCA, 2004; Revilla et al., 2006). Se han descrito numerosas especies de
coccidio en la perdiz roja, todas ellas incluídas en el género Eimeria (Tabla 3).
La forma de presentación son diarreas de gravedad variable que pueden
originar graves pérdidas económicas (APROCA, 2004).
La transmisión se lleva a cabo por ingesta de ooquistes esporulados con la
comida o el agua (Melhorm, 1992) , motivo por el cual la presentación suele
limitarse a las aves alojadas en los parques de vuelo, ya que los ejemplares
reproductores se mantienen en jaulas elevadas que cortan los ciclos
parasitarios. Actualmente, la mayoría de las granjas suministran de manera
preventiva coccidostáticos rutinariamente en el pienso o el agua de bebida, lo
cual consigue disminuir las pérdidas económicas pero supone un handicap
cuando las aves son liberadas al campo, ya que con el cese de los tratamientos
se produce un incremento de la excreción de ooquistes con el consiguiente
riesgo tanto para las aves liberadas como para las silvestres de la zona en que
se efectue la suelta (Villanúa et al., 2006).
En el campo, las coccidiosis suelen aparecer con la llegada de las lluvias en
el inicio del otoño, aunque tan sólo originan patología en aquellas fincas
intensamente manejadas donde se produce un fuerte hacinamiento en torno a
comederos y bebederos (datos propios).
Otros protozoos que en ocasiones se identifican en la perdiz roja serían los
flagelados del género Trichomonas, Histomona meleagridis y varias especies
de hemoparásitos (Leukozytozoon sp., Haemoproteus sp.), aunque son mucho
menos frecuentes. Tabla 3. Principales especies de coccidios descritos en la perdiz roja.
Nematodo Referencias
Eimeria sp. Hernandez-Rodriguez et al., 1974; Cordero del Campillo et al., 1994. Eimeria coturnicis Romero, 1974; Cordero del Campillo et al., 1994. Eimeria gonzalezi Lizano y Romero, 1972; Cordero del Campillo et al., 1994 . Eimeria kofoidi Lizano y Romero, 1972; Cordero del Campillo et al., 1994 Eimeria legionensis Cordero del Campillo y Pla, 1965; Hernandez-Rodriguez et al., 1974 Eimeria padulensis Romero, 1974; Romero y Lizano, 1974; Cordero del Campillo et al., 1994. Eimeria phasiani Romero, 1974; Cordero del Campillo et al., 1994. Eimeria procera Romero, 1974; Reina et al., 1992 ; Cordero del Campillo et al., 1994.

18
El grupo de los nematodos es para el cual se han descrito más especies
parasitando a la perdiz (Tabla 4). La presencia de unas u otras está
condicionada por el sexo o edad de las aves parasitadas (Millán et al., 2004d),
diversos factores geográficos (Calvete et al., 2004), la densidad del perdices
(Calvete et al., 2004) o por el hecho de que se trate de aves silvestres o criadas
en granja (Millán et al., 2004 a y b).
En las aves de granja, las especies detectadas acostumbran a ser aquellas
de ciclos monoxenos (Millán et al 2004a) y dentro de estas, las diferentes
especies del género Heterakis (principalmente H. gallinarum) y los capilaridos
E. contortus y C. caudinflata, serían probablemente los más frecuentemente
identificados (datos propios). Las parasitaciones por Heterakis no suelen
mostrar clínica, a no ser que se acompañen de la infección por Histomona. Por
el contrario C. caundinflata produce graves diarreas con importantes
mortalidades y pérdidas económicas (APROCA, 2004).
En las poblaciones silvestres, las especies presentes suelen ser de ciclos
heteroxenos, es decir, que requieren un hospedador intermediario para
completar sus ciclos (Millán et al., 2004a). Aunque, como ya se comentó
anteriormente, la presencia de una u otra especie se ve influenciada por
numerosos factores (Calvete et al., 2004; Millán et al., 2004d), Cheilospirura
gruveli sería tal vez la más frecuente (ver referencias de tabla 2), aunque
resulta difícil saber si ocasiona patología y la importancia de esta.

19
Tabla 4. Principales nematodosis descritas en el género Alectoris. (A. rufa=r, A. graeca= g, A. chukar=c, A. barbara=b)
Nematodo Referencias
Acuaria sp. Masala, 1986b Acuaria gruweli Tarazona et al., 1979r Acuaria harmulosa Belleau y Leonard, 1978g Acuaria spinosa Akhumyan y Khanbegyan, 1982 g Allopoda spp. Masala, 1986 b Aonchoteca caudinflata Tarazona et al., 1979 r; Millán et al., 2004a r; Foronda et al., 2005 b Ascardia sp. Calvete et al., 2004r Ascaridia comprar Gozdev, 1956 g; Millán et al., 2004a r
Ascaridia galli Tibbits y Babero, 1969c; Reina et al., 1992 r; Millán et al., 2004a r; Foronda et al., 2005 b
Aviosperpens mosgovoyi Cordero del Campillo et al., 1994 r Baruscapillaria obsignata Foronda et al., 2005 b Baylisascaris larvae Sass y Gorgacz, 1978 g Capillaria spp. Govoni y Maestrini, 1979 r; Githkopoulos, 1984 c; Belleou y Leonard, 1991 g; Reina et al., 1992 r
Capillaria anatis Calvete et al., 2003r Capillaria caudinflata Calvete et al., 2004r Capillaria contorta Githkopoulos, 1984 c; Reina et al., 1992 r; Pizarro et al., 2000 r; Calvete et al., 2003r Capillaria obsignata Varela, 1974 r; Masala, 1986 b; Reina et al., 1992 r; Calvete et al., 2004r Capillaria phasianina Githkopoulos, 1984 c Cheilospirura gruvelli Gozdev, 1956 g; Varela, 1974 r; Calvete et al., 2003r; Calvete et al., 2004r; Millán et al., 2004a r Cheilospirura spinosa Koroglu y Tasan, 1996 g Cyrnea eurycerca Gozdev, 1956 g Cyrnea parroti Varela, 1974 r Cyrnea seurati Millán et al., 2004a r Eucoleus contortus Millán et al., 2004a r Eucoleus annulatus Foronda et al., 2005 b Eustrongyloides mergorum Cordero del Campillo et al., 1994 r Canguleterakis sp. Vasilev, 1992 c Canguleterakis altaica Gozdev, 1956 g; Vasilev, 1992 c Canguleterakis macroura Vasilev, 1992 c Canguleterakis tenuicaudata Tarazona et al., 1979 r; Belleau y Leonard, 1991 g;Vasilev, 1992 c Heterakis sp. Vasilev, 1992 c Heterakis dispar Varela, 1974 r; Reina et al., 1992 r; Koroglu y Tasan, 1996 g
Heterakis gallinarum Gozdev, 1956 g; Varela, 1974 r; Tarazona et al., 1979 r; Masala et al., 1986 b; Belleau y Leonard, 1991 g; Reina et al., 1992 r; Koroglu y Tasan, 1996 g; Calvete et al., 2003r; Calvete et al., 2004r; Millán et al., 2004ar; Foronda et al., 2005 b
Heterakis tenuicauda Tarazona et al., 1979 r; Calvete et al., 2003r; Calvete et al., 2004r; Millán et al., 2004a r Oxyspirura schulzi Gozdev, 1956 g Oxyspirura rijikovi Borgarenko, 1970 g Seudocyrnea colini Koroglu y Tasan, 1996 g Seudocyrnea eurycerca Koroglu y Tasan, 1996 g Streptocara crassicauda Vasilev, 1992 c Strongylida sp. Reina et al., 1992 r Subulura brumpti Gozdev, 1956 g Subulura coturnicis Akhumyan y Khanbegyan, 1982 g Subulura differens Koroglu y Tasan, 1996 g Subulura suctoria Varela, 1974 r; Tarazona et al., 1979 r Calvete et al., 2003r; Calvete et al., 2004r Tetrameres sp. Calvete et al., 2003r Tetrameres timopheevoi Gozdev, 1956 g
Trichostrongylus tenuis Tarazona et al., 1979 r; Masala et al., 1986 b; Calvete et al., 2003r; Calvete et al., 2004r; Millán et al., 2004a r

20
Con los cestodos sucede algo semejante a lo mencionado con los
nematodos, y diversos factores condicionan la presencia de las diferentes
especies (Calvete et al., 2004; Millán et al., 2004a, b y d).
El número de especies descritas en la perdiz es bastante elevado (Tabla 5)
y su presencia relativamente frecuente. Las especies más habitualtemente
encontradas serían tal vez Raillietina (R.) tetragona y Rhabdometra
nigropunctata en aves silvestres (Calvete et al., 2003; Millán et al., 2004a;
datos propios) y Skryabinia bolivari en aves de granja (Millán et al., 2004a;
datos propios). Aunque a menudo se encuentre aves parasitadas por estos
vermes sin patología aparente, las cestodosis pueden ocasionar un alto
número de bajas en aquellas granjas con higiene insuficiente o incluso en
fincas en las que se den altas concentraciones de aves en torno a bebederos y
comederos (Revilla et al., 2006). Tabla 5. Principales cestodosis descritas en el género Alectoris. (A. rufa=r, A. graeca= g, A. chukar=c, A. barbara=b)
Cestodo Referencias
Choanotaenia infundibulum Tarazona et al., 1979r; Masala et al.,1986 b; Belleauy Léonard, 1991 g;Vasiliev, 1992c; Cordero del Campillo et al., 1994r; Calvete et al., 2003r ; Calvete et al., 2004r; Millán et al., 2004ar;Foronda et al., 2005 b
Hymenolepis spp. Gozdev, 1956 g; Belleau y Léonard, 1991 g Hymenolepis carioca Masala et al., 1986 g Hymenolepis graeca Johri, 1960 g
Lyruterina nigropunctata Gozdev, 1956 g; Varela, 1974r; Tarazona et al., 1979 r; Calvete et al., 2004r; Foronda et al., 2005 b
Mesocestoides sp. Millán et al., 2003b r Metroliasthes sp. Masala et al., 1986 b Metroliasthes lucida Belleau y Leonard, 1991 g Paradicranotaenia anormales Tarazona et al., 1979 r Raillietina bolivari Calvete et al., 2003r Raillietina cesticillus Masala et al., 1986 b; Calvete et al., 2004r Raillietina circumvallata Gozdev, 1956 g Raillietina echinobothrida Masala et al., 1986 b; Calvete et al., 2004r Raillietina friedbergeri Akhumyan y Khanbegyan, 1982 g Raillietina graeca Gozdev, 1956 g Raillietina korkei Gozdev, 1956 g Raillietina micracantha Tarazona et al., 1979 r Raillietina skrjabini Akhumyan y Khanbegyan, 1982 g
Raillietina (R.) tetragona Tarazona et al., 1979 r; Masala et al.,1986 b; Cordero del Campillo et al., 1994 r; Koroglu y Tasan, 1996 g; Reina et al., 1999 r; Calvete et al., 2003r; Calvete et al., 2004r; Millán et al., 2004a r
Rhabdometra dogieli Akhumyan y Khanbegyan, 1982 g Rhabdometra nigropunctata Calvete, et al., 2003r
Skryabinia bolivari López-Neyra, 1947; Varela, 1974 r; Tarazona et al., 1979 r; Akhumyan y Khanbegyan, 1982 g; Illescas and Gómez, 1987 r; Cordero del Campillo et al., 1994 r; Calvete et al., 2004r; Millán et al., 2004a r
Tetrathyridium variable Gozdev, 1956 g Variolepis farciminosa Smith, 1986g

21
Por último hay que mencionar al grupo de los trematodos, del cual se han
descrito un menor número de especies parasitando a la perdiz (Tabla 6) pero
que, en determinadas zonas, pueden ser unos de los helmintos más
frecuentemente encontrados (Calvete et al., 2003). Tabla 6. Principales trematodosis descritas en el género Alectoris. (A. rufa=r, A. graeca= g, A. chukar=c, A. barbara=b)
Trematodo Referencias Brachylaemus sp. Tarazona et al., 1979 r; Millán et al., 2004a r
Brachylaima fuscatus Gozdev, 1956 g; Perrucci et al., 1997 r
Conspicuum alectoris Varela, 1974 r
Corrigia corrigia Gozdec, 1956 g
Corrigia skrjabini Akhumyan y Khanbegyan, 1982 g
Dicrocoelium sp. Cordero del Campillo et al., 1994 r; Calvete et al., 2003 r; Calvete et al., 2004r; Millán et al., 2004a r
Dicrocoelium petroni Vasilev, 1992 c
Hypoderaeum conoideum Vasilev, 1992 c
Postharmostomum gallinum Gozdev, 1956 g
Tamerlania zarudnyi Gozdev, 1956 g
22
II.7.Las repoblaciones como herramienta de gestión.
A pesar de la caída de las poblaciones de perdiz roja, la presión cinegética
que se ejerce sobre esta especie no ha descendido, sino que ha aumentado
espectacularmente durante los años 60 y 70 y de manera más suave pero
continua durante las últimas décadas (Vargas y Duarte, 2002). Además, el
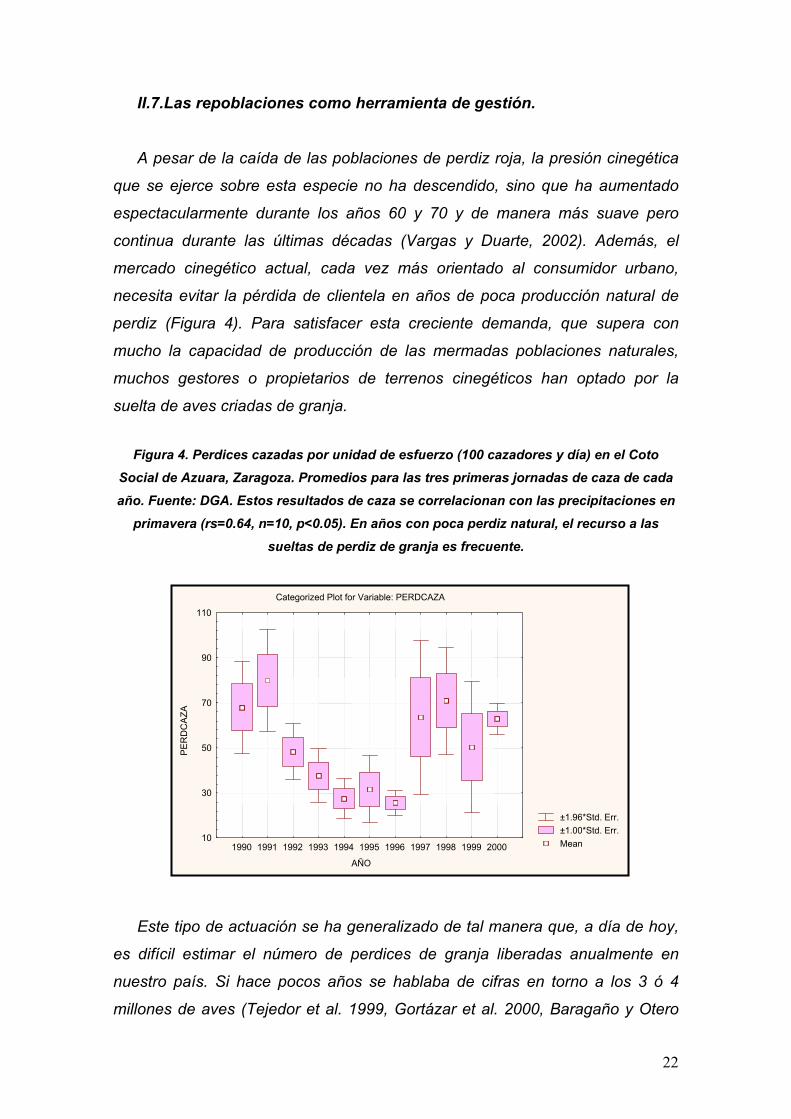
mercado cinegético actual, cada vez más orientado al consumidor urbano,
necesita evitar la pérdida de clientela en años de poca producción natural de
perdiz (Figura 4). Para satisfacer esta creciente demanda, que supera con
mucho la capacidad de producción de las mermadas poblaciones naturales,
muchos gestores o propietarios de terrenos cinegéticos han optado por la
suelta de aves criadas de granja.
Figura 4. Perdices cazadas por unidad de esfuerzo (100 cazadores y día) en el Coto Social de Azuara, Zaragoza. Promedios para las tres primeras jornadas de caza de cada
año. Fuente: DGA. Estos resultados de caza se correlacionan con las precipitaciones en primavera (rs=0.64, n=10, p<0.05). En años con poca perdiz natural, el recurso a las
sueltas de perdiz de granja es frecuente.
Este tipo de actuación se ha generalizado de tal manera que, a día de hoy,
es difícil estimar el número de perdices de granja liberadas anualmente en
nuestro país. Si hace pocos años se hablaba de cifras en torno a los 3 ó 4
millones de aves (Tejedor et al. 1999, Gortázar et al. 2000, Baragaño y Otero
±1.96*Std. Err.±1.00*Std. Err.Mean
Categorized Plot for Variable: PERDCAZA
AÑO
PE
RD
CA
ZA
10
30
50
70
90
110
1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000

23
2001, Millán et al 2003a), existen sospechas de que las cifras reales superen
los 10 millones de ejemplares por año. Si tenemos en cuenta los datos de las
estadísticas oficiales de caza que, como se comentó anteriormente, cifraban en
torno a los 3,5 millones las perdices capturadas anualmente (Baragaño y Otero
2001), podemos darnos cuenta de la importancia alcanzada por las sueltas de
aves de granja en la actualidad.
II.6.1. Viabilidad de las repoblaciones
En el uso de las perdices de granja cabe distinguir dos alternativas: (1) los
refuerzos de poblaciones naturales para aumentar la población estable
(“repoblaciones” o refuerzos poblacionales según la terminología de la UICN), y
(2) las “sueltas” para caza inmediata o refuerzos de caza, típicos de los cotos
comerciales o intensivos. Entre ambas cabe encontrar diversas situaciones
intermedias, como por ejemplo el refuerzo de poblaciones naturales en las
primeras jornadas a fin de aliviar la presión cinegética sobre las perdices
salvajes, tal como ocurre en algunos cotos del Valle del Ebro, o la “repoblación”
con perdices juveniles en verano para su adaptación al terreno de cara a los
ojeos de otoño, manejo frecuente en Castilla – La Mancha.
La viabilidad de las repoblaciones con aves de granja con fines cinegéticos
es en general muy baja, y normalmente incapaz de recuperar las poblaciones
silvestres (Hessler et al.,1970; Birkan, 1971; Leránoz y Castien, 1989; Dowell,
1992; Robertson, 1988; Brittas et al., 1992; Sodeikat et al., 1995; Capelo y
Pereira, 1996; Gortázar et al., 2000; Millán et al., 2002; Pérez et al., 2004;
Alonso et al., 2005). No obstante, la suelta masiva de perdices de granja,
acompañada de un manejo intensivo del hábitat incluyendo el
aprovisionamiento artificial con agua y alimento, y segruamente de un intenso
control de la predación con sus consiguientes conflictos, puede dar lugar a
poblaciones densas semi-naturales. Existen ejemplos de esta situación en
diversos terrenos de Castilla – La Mancha.
En la mayoría de los trabajos científicos en los que se ha evaluado la
supervivencia y las causas de muerte de las aves liberadas se ha llegado a la
conclusión de que el zorro es el principal responsable del fracaso de las
mismas (Papeschi et al., 1993; Gortázar et al., 2000; Millán et al. 2003).
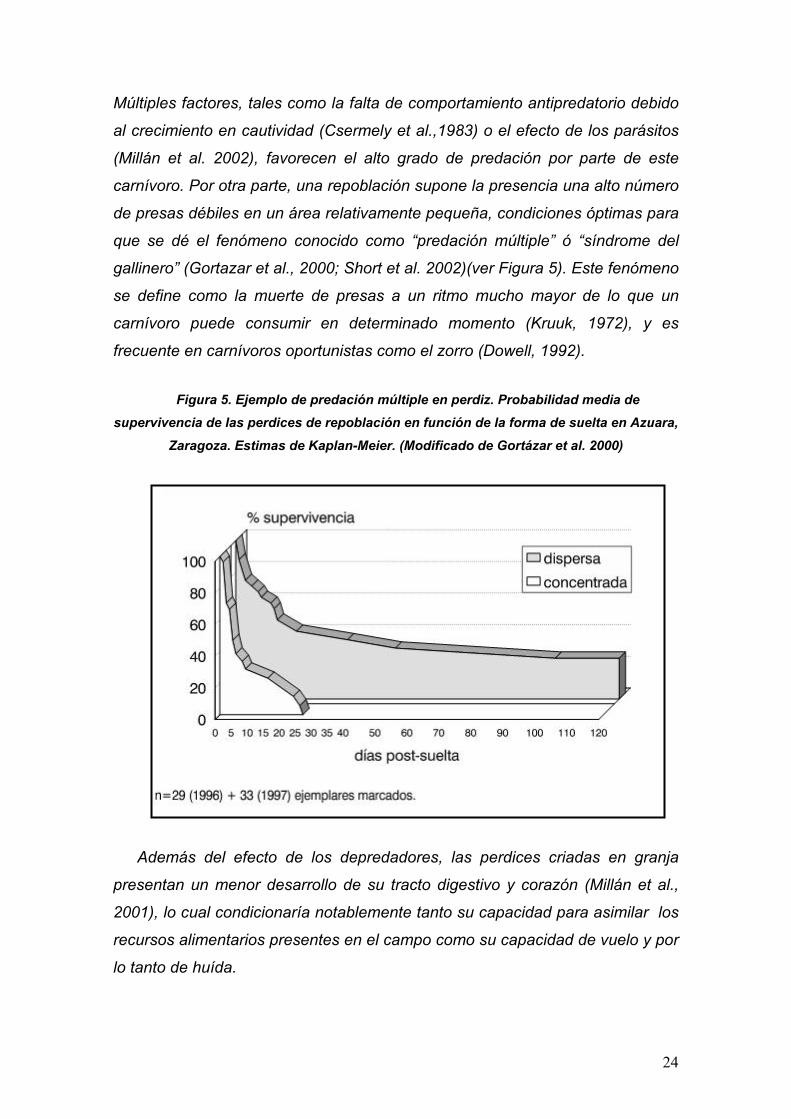
24
Múltiples factores, tales como la falta de comportamiento antipredatorio debido
al crecimiento en cautividad (Csermely et al.,1983) o el efecto de los parásitos
(Millán et al. 2002), favorecen el alto grado de predación por parte de este
carnívoro. Por otra parte, una repoblación supone la presencia una alto número
de presas débiles en un área relativamente pequeña, condiciones óptimas para
que se dé el fenómeno conocido como “predación múltiple” ó “síndrome del
gallinero” (Gortazar et al., 2000; Short et al. 2002)(ver Figura 5). Este fenómeno
se define como la muerte de presas a un ritmo mucho mayor de lo que un
carnívoro puede consumir en determinado momento (Kruuk, 1972), y es
frecuente en carnívoros oportunistas como el zorro (Dowell, 1992).
Figura 5. Ejemplo de predación múltiple en perdiz. Probabilidad media de supervivencia de las perdices de repoblación en función de la forma de suelta en Azuara,
Zaragoza. Estimas de Kaplan-Meier. (Modificado de Gortázar et al. 2000)
Además del efecto de los depredadores, las perdices criadas en granja
presentan un menor desarrollo de su tracto digestivo y corazón (Millán et al.,
2001), lo cual condicionaría notablemente tanto su capacidad para asimilar los
recursos alimentarios presentes en el campo como su capacidad de vuelo y por
lo tanto de huída.

25
Por último hay que tener en cuenta que las perdices en granja están
expuestas a una serie de parásitos que, como se ha demostrado en trabajos
previos, son diferentes a los presentes en las poblaciones naturales (Millán et
al., 2003, 2004). La liberación al campo de estas aves supone, por una parte, el
cese de los tratamientos sanitarios que controlaban a los patógenos propios de
la granja, y por otra, la exposición a nuevos patógenos con los que no habrían
tenido contacto previo. Si a esto añadimos la inmunodepresión causada por el
estrés de la suelta, parece lógico pensar que la capacidad de respuesta de las
aves de granja frente a las enfermedades va a ser muy baja, y que éste va a
ser un factor muy importante a la hora de mermar la supervivencia de las aves
liberadas (Villanúa et al., 2006).
II.6.2. Las hibridaciones.
A pesar de estar terminantemente prohibido por la ley, se han venido
empleando para repoblar híbridos entre perdiz roja y perdiz chukar (Alectoris
chukar), debido a su elevada productividad en cautividad (Padrós, 1991), y a
que en su segunda generación son prácticamente indistinguibles por rasgos
externos de la auténtica perdiz roja (Negro et al., 2001; Millán et al. 2001). Esta
práctica se ha generalizado de tal manera que a día de hoy resulta difícil
encontrar poblaciones silvestres puras y mucho menos granjas sin
contaminación genética (Dávila, 2005) lo cual supone un grave problema para
la conservación de la especie.
II.6.3. Riesgos sanitarios asociados.
Si, como se comentaba en el punto anterior, las diferencias existentes entre
los parásitos propios de granja y los del campo pueden suponer un riesgo para
la viabilidad de las repoblaciones, la liberación de aves parasitadas podría
suponer un riesgo para las poblaciones silvestres. De hecho, la posible
transmisión de enfermedades de los animales liberados a las poblaciones
silvestres es uno de los puntos más importantes a la hora de valorar la
idoneidad de un programa de repoblación (Viggers et al., 1993). Trabajos
previos llevados a cabo por Millán y colaboradores (2004 a y b) mostraron

26
como la parasitofauna de las perdices de granja era totalmente diferente de la
que aparecía en las poblaciones silvestres. Así, las aves de granja aparecían
parasitadas por especies monoxenas, es decir, de ciclo directo; mientras que
en las silvestres se identificaban parásitos heteroxenos, esto es, de ciclo
indirecto. Estas diferencias se explicarían por el hecho de que, en las
condiciones de granja, las perdices no tendrían acceso a los invertebrados
necesarios para cerrar los ciclos de los parásitos heteroxenos. A la inversa, la
menor agregación existente en el campo dificultaría la transmisión de las
especies monoxenas. Teniendo en cuenta estas diferencias, Millán et al., (2004
a y b) sugerían el grave riesgo que la liberación de aves parasitadas al campo
podría tener para las poblaciones silvestres, ya que estarían introduciendo
nuevos patógenos con los que no habrían tenido contacto previo y para los
cuales es de esperar que tuviesen una mayor susceptibilidad.
Un ejemplo lo constituye el caso de las sueltas de faisán (Phasianus
colchicus) y el descenso de las poblaciones naturales de perdiz pardilla (Perdix
perdix) en el Reino Unido. Los estudios llevados a cabo por Tompkins et al.
(1999, 2000, 2001) sugirieron que el nematodo intestinal Heterakis gallinarum
tenía un efecto muy negativo sobre la condición física de la perdiz pardilla,
mientras que resultaba prácticamente apatógeno para el faisán. Ello daría lugar
a una competencia aparente entre el faisán y la perdiz pardilla, mediada por
este parásito. Esta competencia mediada por un parásito podría ser
parcialmente responsable del importante descenso sufrido por las poblaciones
de pardilla del Reino Unido (Tompkins et al., 1999, 2000, 2001).
Si se trasladan estas hipótesis a nuestro caso, la liberación
masiva de perdices de granja podría, lejos de solucionar la
caída de las poblaciones naturales de la especie, estar
añadiendo un nuevo problema a su conservación.

27
II.8. Bibliografía
1. Aebisher, N. J. y Potts, G. R. (1994). Red–legged partridge. In: Tucker, G.
M. y Heath, M. F., Birds in Europe. Their Conservation Status. Birdlife
Conservation Series nº3, Birdlife International, Cambridge, UK.
2. Aebischer, N. y Kavanagh, B. (1997). Grey partridge. In Hagemeijer,
W.J.M., Balir, M.J. (eds.), The EBCC Atlas of European Breeding Birds, their
Distribution and Abundance. TyAD Poyser, London, pp. 212-213.
3. Akhumyan, K.S. y Khanbegyan, R.A. (1982). The helmint fauna of wild
galliforms in Armenia (Coturnix coturnix, Alectoris graeca, Perdix perdix,
Lyrulus mlokosiewiczi and Tetraogallus caspius). Zoologischeskii Sbornik,
Akademiya Nauk Armyanskoi SSR, Institut Zoologii (Fauna parasitov
Zhivotnykh I vyzyvaemye imi zabolevaniya) 18: 9-45.
4. Alonso, M.E., Perez, J.A., Gaudioso, V.R., Diez, C. y Prieto, R. (2005).
Study of survival, dispersal and home range of autumn-released red-legged
partridges (Alectoris rufa). British Poultry Science, 46(4): 401-6
5. APROCA (1998). La caza en la provincia de Ciudad Real. Ciudad Real.
6. APROCA (2004). Jornadas de sobre la cría de especies cinegéticas.
Madrid, Noviembre 2004.
7. Baratti, M., Ammannati, M., Magnelli, C. y Dessi-Fulgheri, F. (2005).
Introgression of chukar genes into a reintroduced red-legged partridge
(Alectoris rufa) population in central Italy. Animal Genetics, 36(1): 29-35
8. Belleau, E. y Léonard, P. (1991). Le parasitisme digesttif chez la perdrix
bartavelle (Alectoris graeca saxatilis), le lagopede alpin (Lapogus mutus),
letetras-lyre (Tetras tetrix), dans le departement des Autes-Alpes. Gibier
Faune Sauvage, 8:161-174.
9. Birkan, M. (1990). La perdrix rouge. Brochures techniques O.N.C., Paris. 36
pp.
10. Blanco-Aguiar, J.A., Virgós, E. y Villafuerte, R. (2003). Perdiz Roja
(Alectoris rufa). En: Martí R. and Del Moral. J. C. (Eds), Atlas de las aves
reproductoras de España. Dirección General de Conservación de la
Naturaleza y Sociedad Española de Ornitología, Madrid, Spain, pp. 212-213

28
11. Borgarenko,I.F. (1970). Trematodes of Rallidae in Tadzhikistan. Izvestiya
Akademii Nauk Tadzhikskoi SSR, Biologiya (Ahboroti Akademijai Fanhoi
Rss Tocikiston) 2: 47-55.
12. Borralho, R., Rego, F. y Vaz Pinto, P. (1997). Demographic trends of red-
legged partridges (Alectoris rufa) in southern Portugal after implementation
of management actions. Gibier Faune Sauvage, 14: 585-599.
13. Borralho, R., Rito, A., Rego, F., Simoes, H. y Vaz-Pinto, P. (1998).
Summer distribution of Red-legged partridge (Alectoris rufa) in relation to
water availability on Mediterranean farmland. Ibis, 140: 620-625.
14. Borralho, R., Stoate, C. y Araujo, M. (2000). Factors affecting the
distribution of Red-legged Partridges Alectoris rufa in an agricultural
landscape of southern Portugal. Bird Study, 47: 304-310
15. Brickle, N.W., Harper, D.G.C., Aebischer, N.J. y Cockayne, S.H. (2000).
Effects of agricultural intensification on the breeding success of corn
buntings Milaria calandra. Journal of Applied Ecology, 37: 742-755.
16. Buenestado, F., Gortázar, C., Millán, J., Höfle, U. y Villafuerte, R. (2004).
Descriptive study o fan avian pox outbreak in wild red-legged partridges
(Alectoris rufa) in Spain. Epidemiology and Infection, 132: 369-374.
17. Calnek, B.W. (1991). Diseases of Poultry. Iowa State University Press.
Iowa. EEUU.
18. Campbell, L.H., Avery, M.I., Donald, P., Evans, A.D., Green, R.E. y
Wilson, J.D. (1997). A review of the indirect effects of pesticides on birds.
JNCC Report No. 227. Joint Nature Conservation Committee, Peterborough,
UK.
19. Capelo, M. y Castro Pereira, D. (1996). Sobrevivência e dispersão de
perdizes (Alectoris rufa L.) largadas em duas operações de repovoamento
cinegético. Revista Florestal, 9: 245-253.
20. Carvalho, J. y Borralho, R. (1998). Productividade e sucesso reproductivo
de duas populaçoes de perdiz-vermelha (Alectoris rufa) em diferentes
habitats. Silva Lusitanica, 6(2): 215-226.
21. Cavanagh, D. (2005).Coronaviruses in poultry and other birds. Avian
Pathology,34(6): 439-448
22. CDC (2007). Http://www.cdc.gov/ncidod/dvbid/westnile/birdspecies.htm

29
23. Cookson, K.C. y Shivaprasad, H.L. (1994). Mycoplasma gallisepticum
infection in chukar partridges, pheasants, and peafowl. Avian Disease,
38(4): 914-921.
24. Cordero del Campillo, M. y Pla, M (1965). Sobre las coccidiosis de las
perdices, condescripción de Eimeria legionensis n.sp. parásita de Alectoris
rufa y una clave para su diferencdiación. Anales de la Facultad de
Veterinaria de León, 11: 107.
25. Cordero del Campillo, M., Castañón, L. y Reguera, A. (1994). Índice-
catálogo de zooparásitos ibéricos. Secretariado de Publicaciones,
Universidad de León. León. Spain.
26. Cramp, S. y Simmons, K.E.L. (1980). The Birds of the Western Palearctic.
Vol. II. Oxford University Press. Oxford.
27. Crick, H.Q.P., Dudley, C., Evans, A.D. y Smith, K.W. (1994). Causes of
nest failure among buntings in the UK. Bird Study, 41: 88-94
28. Cucco, M., Guasco, B., Malacarne, G. y Ottonelli, R. (2007). Effects of
beta-carotene on adult immune condition and antibacterial activity in the
eggs of the Grey Partridge, Perdix perdix. Comp Biochem Physiol A Mol
Integr Physiol. 147(4): 1038-1046.
29. Dávila, J.A. (2005). La problemática de la hibridación de la perdiz roja
(Alecotris rufa) con perdiz chukar (Alectoris chukar) en las poblaciones
silvestres Españolas. I Jornadas Internacionales sobre la Genética de la
Perdiz Roja, Zaragoza.
30. De la Concha, I., Hernández, C y Pinilla, J. (2006). Medidas beneficiosas
para las aves financiables a traves del nuevo reglamento de desarrollo rural.
Sugerencias para su diseño y aplicación en NATURA 2000. SEO/Birdlife.
Madrid. España.196 pp.
31. Díaz, M., Asensio, B. y Tellería, J.L. (1996). Aves Ibéricas I. No
paseriformes. J.M.Reyero Ed., Madrid. 303pp.
32. Dorst, J. (1971). Les oiseaux dans leer milieu. Reencontré. Lusanne.
33. Duarte, J. y Vargas, J.M. (2002). Los sumideros de perdiz roja a lo largo
del ciclo anual. En Lucio, J.A. y Sáenz de Buruaga, M. (eds). Aportaciones a
la gestión sostenible de la caza. FEDENCA-EEC: 63-80.

30
34. Erbeck, D.H. y Nunn, S.A.(1999). Chlamydiosis in pen-raised bobwhite
quail (Colinus virginianus) and chukar partridge (Alectoris chukar) with high
mortality. Avian Disease, 43(4): 798-803.
35. Foronda, P., Casanova, J.C., Figeruelo, E., Abreu, N. y Feliz, C. (2005).
The helminto fauna of the barbary partridge, Alectoris barbara, in Tenerife,
Canary Islands. Journal of Helminthology, 79: 133-138.
36. Githkopoulos, P.R. (1984). Capillaria phasianina in pheasants (Phasianus
colchicus mongolicus) and partridges (Alectoris chukar). Hellenike
Kteniatrike, 27: 8-13.
37. Gortázar, C., Villafuerte, R., y Martín, M. (2000) Success of traditional
restocking of red-legged partridge for hunting purposes in areas of low
density of northeast Spain Aragón. Zeitschrift für Jagdwissenschaft, 46: 23-
30.
38. Gortázar C., Villafuerte R., Escudero M.A. y Marco J. (2002a). Post-
breeding densities of the red-legged partridge (Alectoris rufa) in
agrosystems: a large.scale study in Aragón, Northeastern Spain. Zeitschrift
für Jagdwissenschaft, European Journal of Wildlife Research, 48: 94-101.
39. Gortázar, C., Millán, J., Hofle, U., Buenestado, F.J., Villafuerte, R. y
Kaleta, E.F. (2002b). Pathology of avian pox in wild red-legged partridges
(Alectoris rufa) in Spain. Annals of the New York Academy of Science,969:
354-7
40. Govoni, S. y Maestrini, N. (1979).Considerazioni sulle forme morbose
dellávifauna allevata intesivamente a scopo venatoria, riscontrate presso
Hist. Patología Av Bologna periodo 1.8.1976-31.8.1978. Clinica Veterinaria,
102: 309-314.
41. Gozdev, E.V. (1956). Parasitic worms of Alectoris graeca in south-eastern
Kazakhastan. Trudi Instituta Zoologii Akademiya Nauk Kazakhskoi SSR, 5:
61-76.
42. Green, R.E. (1995). The decline of the corncrake Crex crex in Britain
continues. Bird Study, 42: 66-75.
43. Hayes, E.B., Sejvar, J.J., Zaki, S.R., Lanciotti, R.S., Bode, A.V. y
Campbell, G.L. (2005). Virology, pathology, and clinical manifestations of
West Nile Virus disease. Emerging Infection Disease,11: 1174-1179.

31
44. Hernández-Rodríguez, S., Martínez-Gómez, F. y Calero-Carretero, R.
(1074). Life cycle of coccidia of Spanish partridges Preceedings.of the II
Congreso Internacional de Parasitología, secc. A-6, 2(1): 95.
45. Hoefle, U., Gortázar, C., Ortíz, J.A. y Knispel, B. (2004). Outbreak of
Trichomoniasis in a woodpigeon (Columba palumbus) wintering roost.
European Journal of Wildlife Research, 50: 73-77.
46. Holsboer-Buogo, C., Bacciarini, L., Robert, N., Bodmer, T. y Nicolet, J.
(1997). Vorkommen von Mycobacterium genavense bei Vögeln. Schewiz
Arch Tierh, 139: 397-402.
47. Hudson, P.J. (1986). The Effect of a parasitic nematode on breeding
production of Red Grouse. Journal of Animal Ecology, 55: 85-92.
48. Hudson, P. y Rands, M. (1988). Ecology and management of gamebirds.
BSP Professional Books, Oxford, 263 pp.
49. Hudson, P., Dobson, A.P. y Newborn, D. (1992) Do parasites make prey
vulnerable to preation? Red grouse and parasites. Journal of Animal
Ecology, 61: 681-692.
50. Hudson, P.J., Dobson, A.P. y Newborn, B. (1998) Prevention of
population cycles by parasite removal. Science, 282: 2256-2258.
51. Humberd, J., Guan, Y. y Webster, R.G. (2006). Comparison of the
Replication of Influenza A Viruses in Chinese Ring-Necked Pheasants and
Chukar Partridges. Journal of Virology, 80(5): 2151–2161
52. Jenkins, D., Watson, A., y Miller, G.R. (1963). Population studies on red
grouse (Lagopus lagopus scoticus) in north-east Scotland. Journal of Animal
Ecology, 32: 317-376.
53. Johnson, T.J., Skyberg, J. yNolan, L.K. (2003). Multiple Antimicrobial
Resistance Region of a Putative Virulence Plasmid from an Escherichia coli
Isolate Incriminated in Avian Colibacillosis Avian Diseases, 48(2): 351–360.
54. Koroglu, E. y Tasan, E. (1996). Distribution of helmintos in qualis (Coturnix
coturnix) and partridges (Alectoris graeca) in Elazig and Tunceli areas. Türk
Veterinerlikve Hayvancilik Dergisi, 20: 241-249.
55. Lartiges, A. y Mallet, C. (1983). Conséquences sur le petit gibier de
l'évolution del'agriculture française. Bulletin technique d'information, 377-
378, 103-117.

32
56. Lebreton, P. (1982). Quelques remerques d`ordre écologique et biologique
formulées à propos des Gallinacées européens. Alauda, 50 (4): 260-277.
57. Lizano, J. y Romero, J. (1972). Contribución al estudio de las
coccidiopatías de Alectoris rufa. Descripción de Eimeria gonzalezi n.sp. y
redescripción de E. kofoidi. Revista Ibérica de Parasitología, 32: 95.
58. López Ontiveros, A. y García Verdugo, F. J. (1991). Geografía de la caza
en España. Agricultura y Sociedad, 58: 81-112.
59. Lucio, J.A. (1990). Influencia de las condiciones climáticas en la
productividad de la perdiz roja (Alectoris rufa). ARDEOLA, 37: 207-218.
60. Lucio, A.J. y Purroy, F.J. (1992). Caza y conservación de aves en España.
ARDEOLA, 39(2): 85-98.
61. Lucio, A.J. (1998). Recuperación y gestión de la perdiz roja en España. Pp.
63-92, en La perdiz roja. FEDENCA, Madrid.
62. Masala, S., Garippa, G. y Leoni, A. (1986). Indagine conoscitiva
sullélmintofauna Della pernice sarda (Alecotris barbara). Parasitologia, 28:
282-283.
63. McGowan, P.J.K. (1994). Family Phasianidae (Pheasants and partridges)
En del Hoyo, J. Elliot, A. y Sargatal, J. (Eds). Handbook of the birds of the
world. Vol2.New World vultures to guineafowl. Lynx ed. Barcelona
64. Melhorn, H., Düwell, D., y Raether, W. (1992). Atlas de Parasitología
Veterinaria. GRASS ediciones, Spain.
65. Metra Seis. (1985). Turismo cinegético en España. Ministerio de
Transportes, Turismo y Comunicaciones, Secretaría General de Turismo,
Madrid. 276 pp
66. Millán,J., Gortázar,C. y Villafuerte, R (2001). Marked differences in the
splanchnometry of farm-bred and wild red-legged partridges (Alectoris rufa
L.). Poultry Science, 80: 972-976,
67. Millán, J., Gortázar, C., Tizzani, P. y Buenestado F.J. (2002) Do helmints
increase the vulnerability of released pheasants to fox predation?. Journal of
Helmintology, 76: 225-229.
68. Millán J. Gortázar C., Buenestado F.J., Rodriguez P., Tortosa F.S. y
Villafuerte, R. (2003a). Effects of a fiber-rich diet on physiology and survival
of farm-reared red-legged partridges (Alectoris rufa). Journal of Comparative
Biochemistry and Physiology, 134: 85-91.

33
69. Millán, J., Gortázar,C. and Casanova, J.C. (2003b). First occurrence of
Mesocestoides sp. in a bird, the red-legged partridge, Alectoris rufa, in
Spain. Parasitology Research, 90: 80-81.
70. Millán,J., Gortázar,C. y Villafuerte, R (2004a). A comparison of the helmint
faunas of wild and faro-reared red-legged partridges. Journal of Wildlife
Management, 68(3): 701-707.
71. Millan, J., Gortázar, C., Martin-Mateo, M.P. y Villafuerte, R. (2004b).
Comparative survey of the ectoparasite fauna of wild and farm-reared red-
legged partridges (Alectoris rufa), with an ecological study in wild
populations. Parasitology Research, 93(1): 79-85.
72. Millan, J., Gortázar, C., y Villafuerte, R. (2004c). Mycobaterium avium
disease in wild red-legged partridges (Alectoris rufa). European Journal of
Wildlife Research, 50: 97-99
73. Millan, J., Gortázar, C., y Villafuerte, R. (2004d). Ecology of nematodo
parasitism in red-legged partridge (Alectoris rufa) in Spain. Helminthologia,
41(1): 22-27.
74. Moreno, S. y Villafuerte, R. (1995). Tradicional management of scrubland
for the conservation of rabbits Oryctolagus cuniculus and their predators in
Doñana National Park, Spain. Biological Conservation, 73: 81-85.
75. Mullarney, K., Svensson, L., Zetterstrom, D y Grant, P.J. (2001). Guia de
aves. La guía de campo de aves de España y de Europa más completa.
Ediciones Omega S.A.. Barcelona.400 pp.
76. Muñoz-Goyanes, G. (1960). Anverso y reverso de la mixomatosis. Madrid:
Dirección General de Monets, Caza y Pesca Fluvial.
77. Nadal, J., Nadal, J. y Rodríguez-Teijerio, J.D. (1996). Red-legged
partridge (Alectoris rufa) age and sex ratios in declining populations in
Huesca (Spain) applied to management. Rev. Ecol. (Terre Vie), 51: 243-
257.
78. Negro, J.J., Torres, M.J. y Godoy, J.A. (2001). RAPD análisis for detection
and eradicarion of hibrid partridges (Alectoris rufa x Alectoris graecca) in
Spain. Biological Conservation, 98: 19-24.
79. OIE (2007). http://www.oie.int/esp/maladies/es_classification2007
80. OFFICE NATIONAL DE LA CHASSE (1986). La Perdix rouge. Notes
techniques. Bull. No. 106. ONC, París.

34
81. Otero, C. (1995). Control de depredadores en la gestión integrada de un
territorio. En: Depredación, caza y vida silvestre, pp. 151-180. Fundación
“La Caixa”/Aedos, Barcelona.
82. Padrós, J. (1991). Situación actual del sector. Presente y Futuro. En
Fundación la Caixa (Ed.): La perdiz roja, pp 7-10.
83. Pennycott, T.W., Cinderey, R.N., Park, A., Mather, H.A., y Foster, G.
(2002). Salmonella enterica subspecies enterica serotype Typhimurium and
Escherichia coli O86 in wild birds at two garden sites in south-west Scotland.
Veterinary Record, 151: 563-567.
84. Pepin, D. y Blayac, J. (1990). Impacts d´un amenagement de la garrigue et
de l’instauration d´un plan de chasse sur la démographie de la perdix rouge
(Alectoris rufa) en milieu méditerranéen. Gibier Faune Sauvage, Game Wild,
7: 145-158.
85. Potts, G.R. (1980). The effects of modern agriculture, nest predation and
game management on the population ecology of partridges (Perdix perdix
and Alectoris rufa). Ecological Research, 2: 2-79.
86. Potts, G.R., Tapper, S.C. y Hudson, P.J. (1984). Population fluctuations in
red grouse: analysis of bag records and a simulation model. Journal of
Animal Ecology, 53: 21-36.
87. Potts, G.R. (1986). The partridge, pesticide, predation and conservation.
London: Collins.
88. Pérez, J.A., Gaudioso, V.R., Alonso, M.E., Olmedo, J.A., Díez, C. y
Bartolomé, D. (2004). Use of raditracking techniques to study a summer
repopulation with red-legged partridge (Alectoris rufa) chicks. Poultry
Science, 83: 882-888.
89. Pizarro, M., Villegas, P., Rodríguez, A., González, M. y Flores, J.M.
(2000). Capillaria contorta parasitism in red-legged partridge under farm
conditions in Spain: histiopathology of the upper digestive system. World´s
Poultry Science Journal, 56: 159-166.
90. Putaala, A., Hohtola, E. y Hissa, R. (1995). The effect of group size on
metabolism in huddling grey partridge (Perdix perdix). Journal of
Comparative Biochemistry and Physiology, 111(2): 243-247.
91. Randi, E., Chiricolo, M., Spagnesi, M., Ghedini, I., Savigni, G.,
Giovannini, A. y Franceschi, C. (1985). Antibody response in partridge

35
(Perdix perdix). Effect of sex and age on the immune response to sheep red
blood cells (SRBC), newcastle disease virus (NDV) and (Buck 19).
Developmental & Comparative Immunology, 9(4): 679-690
92. Rands, M.R.W. (1986 a). Effect of hedgerow characteristics on partridge
breeding densities. Journal of Applied Ecology, 23: 479-487.
93. Rands, M.R.W. (1986 b). The survival of gamebirds (Galliforms) chicks in
realtion to the pesticida use on cereals. Ibis, 128: 57-64.
94. Rands, M.R.W. (1987). Hedgerow management for the conservation of
partridges Perdix perdix and Alectoris rufa. Biological Conservation, 40: 127-
139.
95. Rands, M.R.W. (1988). The efffect of nest predation on grey partridge
(Perdix perdix) and red-llegged partridge (Alectoris rufa). Ornis
Scandinavica, 19: 35-40.
96. Reina, D., Habela, M., Serrano, F., Nieto, C.G., Breña, M., Pérez, E. y
Navarrete, I. (1992). Contribución al conocimiento de la parasitofauna de
los animales silvestres y de vida libre en la provincia de Cáceres (España).
Pp.407-428 in Hernández Rodríguez, S. (Ed.) in memoriam Prof. Dr.
Francisco de Paula Martínez Gómez.
97. Revilla, M., Pérez, E., Arnal, M.C., Villanúa, D., Fernández de Luco, D. y
Gortázar, C. (2007). Principales patologías de la perdiz roja (Alectoris rufa).
Porceedings of the 24émes Rencontres du G.E.E.F.S.M., Junio 2006,
Reserva Nacional de Caza de los Puertos de Tortosa y Beceite. España
98. Romero, J. (1974). Coccidiopatías de interes veterinario en España. II
Reunión Nacional de Centros de investigación Ganadera del CSIC,
Córdoba, España.
99. Romero, J. y Lizano, J. (1974). Eimeria padulensis n.sp. Protozoo-
Eimeriidaae, parásita de Alecotris rufa. Revista Ibérica de Parasitología, 34:
29.
100. Rueda, M.J., Baragaño, J.R., Notario, A. y Castresana, I. (1993).
Estudio de la alimentación natural de los pollos de perdiz roja (Alectoris
rufa). Ecologia, 7: 429-454.
101. Sacristán, J.P. y Sagardía, J. (2005). La enfermedad de Gumboro (y
II). Incidencia en España. Mundo Ganadero, XVI (176): 36-38

36
102. Sass, B. y Gorgacz, E.J. (1978). Cerebral nematodiasis in a chukar
partridge. Journal of the American Veterinary Medical Asociation, 173: 1248-
1249.
103. Sharma, V.D., Sethi, M.S., Singh, S.P. (1980). The occurrence of
salmonellae in free-flying-avifauna: isolation and antibiogram. International
Journal of Zoonoses,7(1): 54-57.
104. Stuve, G., Hofshagen, M. y Holt, G. (1992). Necrotizing lesions in the
intestine, gizzard, and liver in captive capercaillies (Tetrao urogallus)
associated with Clostridium perfringens. Journal of Wildlife Disease, 28(4):
598-602.
105. Tarazona, J.M., Sanz-Pastor, A. y De la Camara, R. (1979). Helmintos
y helmintosis de la perdiz roja (Alectoris rufa). Anales del Instituto Nacional
de Investigaciones Agrarias, Higiene y Sanidad Animal, 4: 55-68.,;
106. Tapper, S.C. (1992). Game heritage. An ecological review from shooting
and game keeping records. Joint Nature Conservation Committee. Game
Conservancy.
107. Tapper, S.C., Potts G.R. y Brockless M.H. (1996). The effecte of an
experimental reduction in predation pressure on the breeeding success and
population density of grey partridges Perdix perdix. Journal of Applied
Ecology, 33: 965,978.
108. Tibbits, F.D. y Babero, B.B. (1969). Ascaridia galli from the chukar
partridge, Alectoris chukar, in Nevada. Journal of Parasitology, 55: 1252.
109. Tompkins, D.M., Dickson, G. y Hudson, P.J. (1999). Parasite-mediated
competition between pheasant and grey partridge: a preliminary
investigation. Oecologia, 119: 378-382
110. Tompkins, D.M., Greenman, J.V., Robertson, P.A. y Hudson, P.J.
(2000). The role of shared parasites in the exclusion of wildlife host:
Heterakis gallinarum in the ring-necked pheasant and the grey partridge.
Journal of Animal Ecology, 69: 829-840.
111. Tompkins, D.M., Greenman, J.V. y Hudson P.J. (2001). Differential
impact of shared nematode parasite on two gamebird host: implications for
apparent competition. Parasitology, 122: 187-193.
112. Tucker, G.M. y Heath M.F. (1994). Birds in Europe. Their conservation
status. Conservation series Nº 3. Bird Life International, Cambirdge.

37
113. Vargas, J.M. y Duarte, J. (2002). Dos modelos discrepantes de gestión
de la perdiz roja en España. En Lucio, J.A. y Sáenz de Buruaga, M. (eds).
Aportaciones a la gestión sostenible de la caza. FEDENCA-EEC: 101-126.
114. Varela, M.C. (1974). Some ecological and epidemiological aspects of
helminto fauna of the red-legged partridge (Alectoris rufa) in te forest
perimeter of Contenda. Thesis, Universidade Técnica de Lisboa, Escuela
Superior de Medicina Veterinaria. Lisboa.
115. Vasiliev, I. (1992). Helminths in Thracian rock partridge (Alectoris
chukar) in Bulgaria. Helminthologia, 29: 117-120.
116. Viggers, K.L., Lindenmayer, D.B. y Spratt, D.M. (1993). The
importance of disease in reintroduction programmes. Wildlife Research, 20:
687-698.
117. Villanúa D., Acevedo P., Höfle U., Rodríguez, O. y Gortázar C (2006).
Changes in parasite transmission stage excretion after pheasant release.
Journal of Helminthology, 80: 1-7.
118. Villafuerte, R., Calvete, C., Gortázar, C. y Moreno, S. (1994). First
epizootic of rabbit hemorrhagic disease in free living populations of
Oryctolagus cuniculus at Doñana National Park, Spain. Journal of Wildlife
Diseases, 30(2): 176-179.
119. Villafuerte, R., Viñuela, J. y Blanco, J.C. (1998). Extensive predator
persecution caused by population crash in a game species: the case of red
kites and rabbits in Spain. Biological Conservation, 84: 181-188.
120. Yanes, M., Herranz, J., de la Puente, J., Tognoni, C y Suárez, F.
(1998). Efectos de los predadores sobre la caza menor y evaluación de
sistemas selectivos para regular los niveles de predación. Consejería de
Agricultura y Medio Ambiente. Junta de Comunidades de Castilla-La
Mancha.

38
III. OBJETIVOS La presente Tesis Doctoral ha sido concebida bajo un enfoque
eminentemente práctico, tratando de responder a las dudas de carácter
sanitario que se le plantearían a cualquier persona interesada en la
conservación de la perdiz roja y que barajase la posibilidad de recurrir a las
sueltas de aves de granja como herramienta para recuperar las poblaciones
naturales.
La primera pregunta sería probablemente la siguiente: ¿Qué riesgos
sanitarios podría tener la liberación de las perdices criadas en cautividad?
Dado que una tesis anterior demostró marcadas diferencias entre las
parasitocenosis de las perdices de granja y las perdices natrales (Millán 2004 a
y b), mediante el estudio parasitológico de las perdices cazadas en fincas con
distintos modelos de gestión, se trata de constatar o desmentir la introducción
de nuevos parásitos en el medio mediante las sueltas de perdiz de granja. Este
objetivo se desarrolla en el capitulo 1.
En caso de demostrase que realmente las aves liberadas pueden introducir
nuevos parásitos en el campo, una de las posibles soluciones sería la de la de
liberar aves que no estuviesen infectadas. Para ello deberíamos contar con un
método seguro que nos permitiese identificar a las aves parasitadas, lo cual
nos plantearía la siguiente pregunta: ¿Son los controles llevados a cabo
actualmente en las granjas de perdiz roja suficientemente eficaces como para
evitar esta introducción de parásitos? Para responder a esta pregunta se ha
realizado una experiencia controlada donde se comparan los resultados de los
análisis coprológicos realizados a las mismas aves a distintas horas del día y
con distintos tipos de heces, con el fin de detectar los posibles fallos del
método de control seguido en la actualidad. Este objetivo se desarrolla en el
capitulo 2.
Si los controles actuales fuesen insuficientes para garantizar que las aves
liberadas no son portadoras de parásitos, el siguiente paso sería el de plantear
el tratamiento de todas las aves antes de la suelta para así asegurar que
lleguen al campo desparasitadas. Esta actuación plantearía la siguiente
pregunta: ¿Son los tratamientos antiparasitarios actuales suficientemente
eficaces como para evitar la introducción de parásitos en el campo a través de

39
sueltas de perdiz roja? Nuevamente se ha diseñado un experimento controlado
con el que poder evaluar la efectividad del Albendazol, uno de los
antihelmínticos más frecuentes en las granjas de perdiz, como tratamiento
frente a Aonchoteca caudinflata y Heterakis gallinarum, dos de los nematodos
más frecuentes en las perdices de granja pero generalmente ausentes en las
poblaciones silvestres. Este objetivo se desarrolla en el capitulo 3.
Como en numerosos acotados la perdiz roja cohabita con otras especies de
aves, algunas de ellas en grave peligro de extinción, habría que plantearse
analizar también el efecto que las suletas de pediz de granja podría tener sobre
las mismas. Así pués, la pregunta que se nos plantearía sería la siguiente:
¿Podrían las sueltas de perdices tener consecuencias negativas sobre el
estado sanitario de otras especies silvestres? Mediante el caso práctico de un
sisón (Tetrax tetrax) capturado en una zona donde anualmente se liberan miles
de perdices de granja, se trata de demostrar el riesgo que estas repoblaciones
podrían tener para otras especies. Este objetivo se desarrolla en el capitulo 4.
Por último, y tras haber obtenido unos resultados que desaconsejasen la
repoblación con perdiz de granja, cabría preguntarse ¿Es posible recuperar la
perdiz roja sin sueltas de granja? Para responder a esta cuestión se analizan
los datos obtenidos en un seguimiento de las abundancias de perdiz en
Aragón, tratando de identificar los factores que podrían estar causando el
descenso de las poblaciones, con el fin de poder atajar el problema desde su
inicio, sin tener que recurrir a la liberación de aves criadas en granja. Este
objetivo se desarrolla en el capitulo 5.

40
Así pues, los objetivos de la presente tesis se podrían resumir en las
siguientes preguntas:
IV.1. ¿Son realmente las repoblaciones con aves de granja un foco
de introducción de nuevos parásitos en las poblaciones silvestres
de perdiz roja?
IV.2. ¿Son los controles llevados a cabo actualmente en las granjas
de perdiz roja suficientemente eficaces para evitar la introducción
de parásitos?
IV.3. ¿Son los tratamientos antiparasitarios actuales
suficientemente eficaces en la perdiz roja como para evitar la
introducción de parásitos en el campo?
IV.4. ¿Podrían las sueltas de perdices tener consecuencias
negativas sobre el estado sanitario de las especies amenazadas?
IV.5. ¿Existen otras alternativas para recuperar la perdiz roja?

41
IV. CAPITULO 1
Las repoblaciones con aves de granja como fuentes de
introducción de parásitos en las poblaciones silvestres de
perdiz roja.
Sanitary risks of red-legged partridge releases: introduction of parasites.
European Journal of Wildlife Research. Villanúa, D., Pérez-Rodríguez, L.,
Casas, F., Alzaga, V., Acevedo, P. and Gortázar, C. (Aceptado).

42
IV. CAPITULO 1
Las repoblaciones con aves de granja como fuentes de
introducción de parásitos en las poblaciones silvestres de
perdiz roja.
Sanitary risks of red-legged partridge releases: introduction of parasites.
European Journal of Wildlife Research. Villanúa, D., Pérez-Rodríguez, L.,
Casas, F., Alzaga, V., Acevedo, P. and Gortázar, C. (Aceptado).
RESUMEN
Se ha estudiado la parasitofauna y la condición física de 99 perdices rojas
(Alectoris rufa) cazadas en Ciudad Real (zona centro de España). Cuarenta y
seis de ellas procedían de dos fincas de caza donde anualmente se lleva a
cabo la suelta de un importante número de perdices de granja. Las cincuenta y
tres restantes fueron muestreadas en fincas naturales en las que no se realizan
repoblaciones con aves de granja y de localización próxima a las fincas con
suelta.
Se identificaron cuatro nematodos (Heterakis gallinarum, Aonchoteca
caudinflata, Eucoleus contortus y Cheilospirura gruveli) y dos cestodos
(Raillietina (R.) tetragona y Skryabinia bolivari). Las fincas con sueltas de perdiz
de granja presentaron una mayor diversidad de especies de parásitos, con
mayores prevalencias e intensidades de parasitación para todos los helmintos
encontrados. Tres de estas especies son típicas de aves criadas en granja y
dos de ellas, A. caundinflata y S. bolivari, fueron encontradas parasitando aves
adultas, lo cual pone de manifiesto la introducción de estos helmintos en la
población natural reproductora. Las aves muestreadas en las fincas naturales
mostraron una condición física mejor que la de las de la zona con sueltas, pero
esta relación aparentemente no estaba relacionada con la parasitación.
Los resultados obtenidos sugieren que la suelta de aves de granja, práctica
cada vez más común en los cotos de caza españoles, puede suponer un riesgo
para la conservación de la poblaciones naturales, ya que se estarían
introduciendo nuevos patógenos al medio. Sin embargo, los resultados
sugieren también que, a día de hoy, el simple cese de las sueltas podría tal vez

43
resultar suficiente para evitar la incorporación permanente de dichos patógenos
al medio.
ABSTRACT
We studied the helminth community and body condition of 99 hunter
harvested red-legged partridges (Alectoris rufa) from Ciudad Real (Central
Spain). Forty six were sampled in two game estates where an important number
of farm-reared red-legged partridges are released yearly. The remaining 53
were obtained from natural wild populations adjacent to one of the estates with
releases.
Four nematode species (Heterakis gallinarum, Aonchoteca caudinflata,
Eucoleus contortus and Cheilospirura gruveli) and two cestode species
(Raillietina (R.) tetragona and Skryabinia bolivari) were identified. The managed
areas showed higher parasite diversity, with higher prevalences and intensities
for all helminths found. Three of these species were typical of farm-bred
partridges and two of these, A. caundinflata and S. bolivari, were found
parasitizing adult partridges. This suggests introduction of these helminths into
the breeding population of managed states. The birds sampled in the non-
managed estates showed a better body condition, but no relation with parasite
infection was found.
Our results suggest that the release of farm reared red-legged partridges, a
strategy that is becoming a common practice in Spanish hunting areas, poses
risk to wild populations because of introducing parasites. However, these
results also suggest that simply stopping releases may be a good way to
remove locally those parasites from populations.
KEYWORDS: Alectoris rufa, condition, helminths, restocking, Spain
INTRODUCTION
The red-legged partridge (Alectoris rufa) is the most abundant game bird in
the Iberian Peninsula, and its hunting is one of the most important economical
and social activities in central Spain (Bernabeu 2000). During the last decades,
natural populations of this game bird have declined in most of its distribution
range (Aebischer and Potts 1994). In Spain, only declines have been

44
documented (Gortázar et al. 2002, and references therein). The strategy of
many hunting estate managers to overcome this decline has been releasing
farm reared birds.
Previous studies evidenced differences between the parasites found in
farmed and those found in wild red-legged partridges (e.g. Millán et al. 2004a).
Regarding helminths, monoxenous species were more abundant in farms and
heteroxenous parasites in natural populations (Millán et al. 2004b, 2004c).
These differences can negatively affect autochthonous populations in the
managed areas, due to the probably limited previous contact with these
pathogens. In fact, potential disease transmission from released animals to wild
populations is one of the most important points to be considered in restocking
programs (Viggers et al. 1993). As an example, the annual release of farmed
ring necked pheasants (Phasianus colchicus) in the United Kingdom is believed
to maintain or even increase Heterakis gallinarum burdens in the wild pheasant
population (Draycott et al. 2000), which in turn could be one of the factors
involved in the grey partridge Perdix perdix decline (Tompkins et al. 2001).
However, the survival of released red-legged partridges is low (see Birkan
1977; Capelo and Pereira 1996; Gortázar et al. 2000; Duarte and Vargas 2004),
and this could be limiting pathogen transmission between farmed and wild birds.
Thus, the aim of the present study was to confirm if parasites could be
effectively introduced into the field by farm reared red-legged partridge
releases.
MATERIAL AND METHODS
Study area
The study was performed in four game estates, ranging from 1009 Ha. to
3145 Ha., located in Ciudad Real (Central Spain, 30T, X419903/Y 4313925).
Habitat is characterized by undulated farmland of wheat and barley crops with
olive trees and vineyard patches. Most cereals are cultured by a traditional two-
year rotation system.
The red-legged partridge is the most important game species in these
hunting areas, although other game species like the Iberian hare (Lepus
granatensis) or the feral pigeon (Columba livia) are present. The red fox (Vulpes
vulpes) is the most abundant predator, but feral cats (Felis catus) and dogs

45
(Canis familiaris), montagu´s harrier (Circus pygargus), marsh harrier (Circus
aeruginosus), buzzards (Buteo buteo) and golden eagle (Aquila chrysaetos) can
prey on red-legged partridges. Moreover, the study area holds important
populations of several species of steppe birds, such as great bustard (Otis
tarda), little bustard (Tetrax tetrax), pin-tailed sandgrouse (Pterocles orientalis),
or stone curlew (Burhinus oedicnemus).
Two of the sampled game estates followed an intensive management model
with more than 2,000 farm reared partridges released yearly. The other two
consisted of wild populations where no restocking with captive-bred game-birds
was performed in the last ten years, and that were adjacent to one of the
managed states.
Study animals
A total of 99 hunter-harvested red-legged partridges were sampled, 46 from
the managed areas and 53 from the wild populations. All birds were weighed
using a 1000 g Pesola® precision balance (±5 grams) and left tarsus length was
measured using a caliper (±0.01 mm). All measures were taken by the same
person (LPR). Sex and age were determined in the field according to the
morphological description of Sáenz de Buruaga et al. (2001), but were
confirmed by examination of the gonads and the bursa of Fabricius. We tried to
obtain a sex- and age-ratio next to 50% in our sample.
Partridge body condition was estimated using two different measurements:
the pectoral muscle thickness (PMT), measured using a portable ultrasonic
meter (Krautkramer USM 22 device) (Pérez-Rodríguez et al. 2006) and the
residuals of the regression of the body mass on the cube of tarsus length
(RBMTL)(Andersson 1992).
Laboratory methods
The digestive tract was opened longitudinally, and the content was collected
for parasite isolation. The digestive tract was soaked overnight in water to allow
the liberation of any parasite that could be attached to the mucosa. The liver
was cut into 3 mm slices and put into water to allow the visual inspection for
trematodes. The samples obtained were examined by means of a
stereomicroscope and the detected worms were counted and stored in 70 %
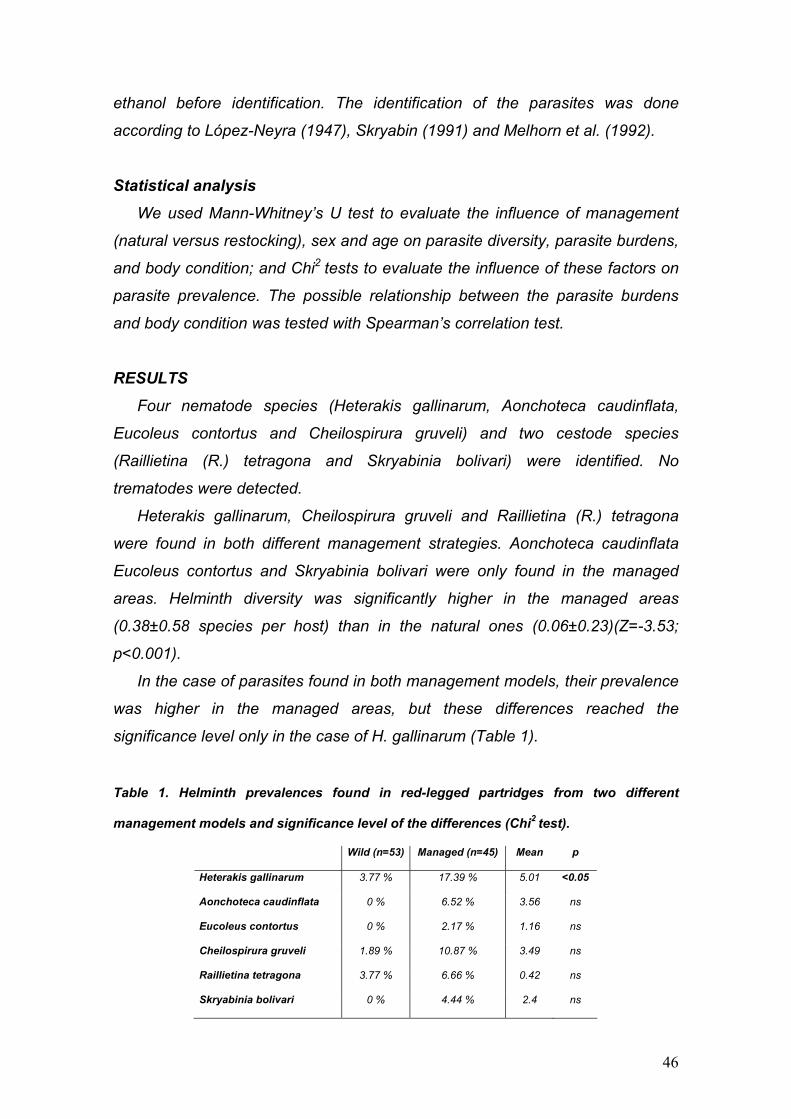
46
ethanol before identification. The identification of the parasites was done
according to López-Neyra (1947), Skryabin (1991) and Melhorn et al. (1992).
Statistical analysis
We used Mann-Whitney’s U test to evaluate the influence of management
(natural versus restocking), sex and age on parasite diversity, parasite burdens,
and body condition; and Chi2 tests to evaluate the influence of these factors on
parasite prevalence. The possible relationship between the parasite burdens
and body condition was tested with Spearman’s correlation test.
RESULTS
Four nematode species (Heterakis gallinarum, Aonchoteca caudinflata,
Eucoleus contortus and Cheilospirura gruveli) and two cestode species
(Raillietina (R.) tetragona and Skryabinia bolivari) were identified. No
trematodes were detected.
Heterakis gallinarum, Cheilospirura gruveli and Raillietina (R.) tetragona
were found in both different management strategies. Aonchoteca caudinflata
Eucoleus contortus and Skryabinia bolivari were only found in the managed
areas. Helminth diversity was significantly higher in the managed areas
(0.38±0.58 species per host) than in the natural ones (0.06±0.23)(Z=-3.53;
p<0.001).
In the case of parasites found in both management models, their prevalence
was higher in the managed areas, but these differences reached the
significance level only in the case of H. gallinarum (Table 1).
Table 1. Helminth prevalences found in red-legged partridges from two different
management models and significance level of the differences (Chi2 test).
Wild (n=53) Managed (n=45) Mean p
Heterakis gallinarum 3.77 % 17.39 % 5.01 <0.05
Aonchoteca caudinflata 0 % 6.52 % 3.56 ns
Eucoleus contortus 0 % 2.17 % 1.16 ns
Cheilospirura gruveli 1.89 % 10.87 % 3.49 ns
Raillietina tetragona 3.77 % 6.66 % 0.42 ns
Skryabinia bolivari 0 % 4.44 % 2.4 ns
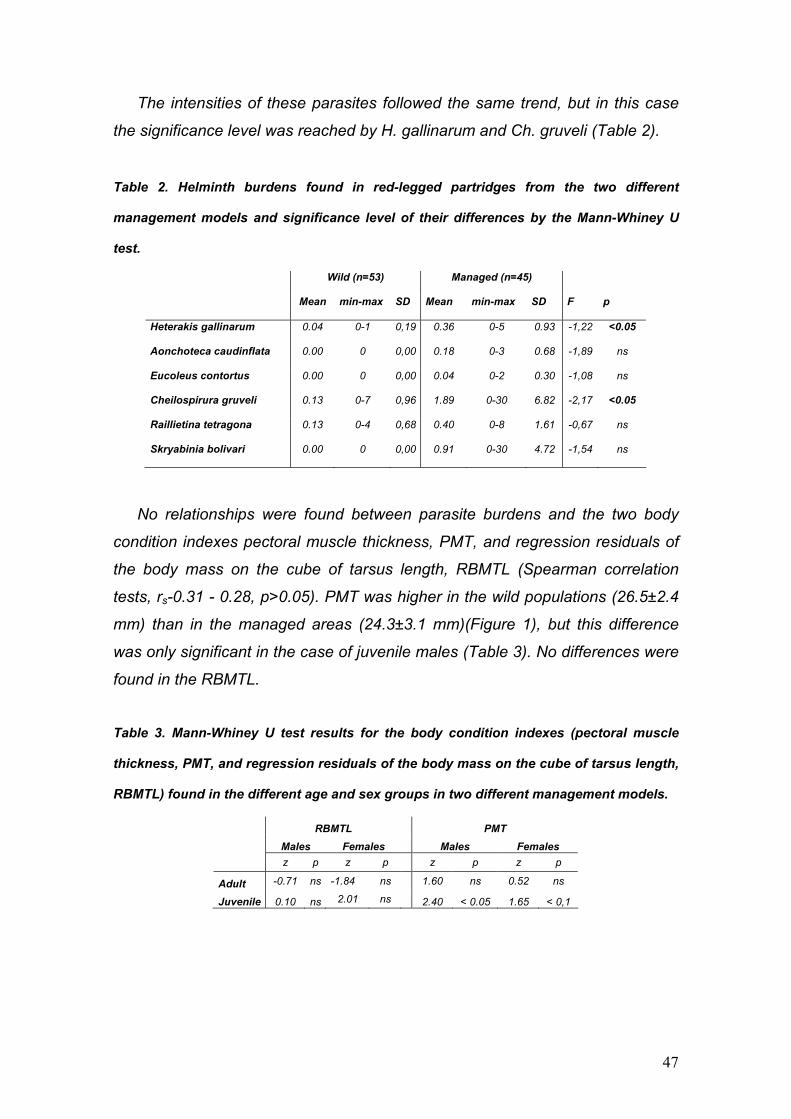
47
The intensities of these parasites followed the same trend, but in this case
the significance level was reached by H. gallinarum and Ch. gruveli (Table 2).
Table 2. Helminth burdens found in red-legged partridges from the two different
management models and significance level of their differences by the Mann-Whiney U
test.
Wild (n=53) Managed (n=45)
Mean min-max SD Mean min-max SD F p
Heterakis gallinarum 0.04 0-1 0,19 0.36 0-5 0.93 -1,22 <0.05
Aonchoteca caudinflata 0.00 0 0,00 0.18 0-3 0.68 -1,89 ns
Eucoleus contortus 0.00 0 0,00 0.04 0-2 0.30 -1,08 ns
Cheilospirura gruveli 0.13 0-7 0,96 1.89 0-30 6.82 -2,17 <0.05
Raillietina tetragona 0.13 0-4 0,68 0.40 0-8 1.61 -0,67 ns
Skryabinia bolivari 0.00 0 0,00 0.91 0-30 4.72 -1,54 ns
No relationships were found between parasite burdens and the two body
condition indexes pectoral muscle thickness, PMT, and regression residuals of
the body mass on the cube of tarsus length, RBMTL (Spearman correlation
tests, rs-0.31 - 0.28, p>0.05). PMT was higher in the wild populations (26.5±2.4
mm) than in the managed areas (24.3±3.1 mm)(Figure 1), but this difference
was only significant in the case of juvenile males (Table 3). No differences were
found in the RBMTL.
Table 3. Mann-Whiney U test results for the body condition indexes (pectoral muscle
thickness, PMT, and regression residuals of the body mass on the cube of tarsus length,
RBMTL) found in the different age and sex groups in two different management models.
RBMTL PMT Males Females Males Females
z p z p z p z p
Adult -0.71 ns -1.84 ns 1.60 ns 0.52 ns
Juvenile 0.10 ns 2.01 ns 2.40 < 0.05 1.65 < 0,1
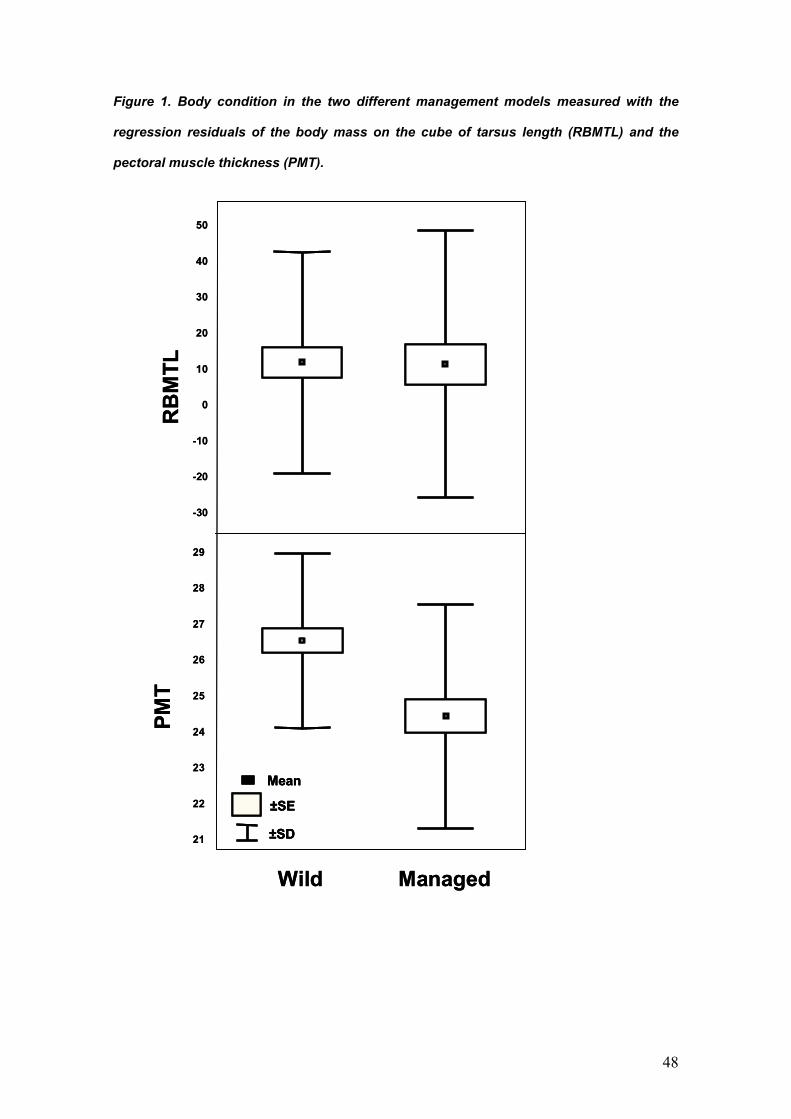
48
Figure 1. Body condition in the two different management models measured with the
regression residuals of the body mass on the cube of tarsus length (RBMTL) and the
pectoral muscle thickness (PMT).
-30
-20
-10
0
10
20
30
40
50
RB
MTL
Managed
21
22
23
24
25
26
27
28
29
PMT
Wild
Mean
±SE
±SD
-30
-20
-10
0
10
20
30
40
50
RB
MTL
Managed
21
22
23
24
25
26
27
28
29
PMT
Wild
Mean
±SE
±SD
Mean
±SE
±SD

49
DISCUSSION
All parasite species found in our study have been previously described
parasitizing the red-legged partridge (López-Neyra 1947; Carvalho-Varela and
Ferradeira 1997; Cordero del Campillo and Rojo 1999; Calvete et al. 2003;
Millán et al. 2004a, 2004b).
The species richness detected in this study (6 species in the managed area
and only 3 in the natural one) is lower than those reported in two previous large-
scale studies (13 different species by Calvete et al. 2003; or 14 different species
by Millán et al. 2004a), but similar to the richness reported by Millán et al.
(2004c) in a small study area in southern Spain (4 species). However,
prevalences and abundances are markedly lower than those reported in these
studies.
Millán et al. (2004a, 2004b) found that the parasite community in wild
populations of red-legged partridges was different from that of farm-bred birds.
They suggested that new parasites could be introduced into the field with the
release of farm-reared birds. Three species identified in our study, Heterakis
gallinarum, Cheilospirura gruveli and Raillietina (R.) tetragona, are usually
found in natural populations (López-Neyra 1947; Tarazona et al. 1979; Millán et
al. 2004b, 2004c). However, the other three ones (Eucoleus contortus,
Aonchoteca caundinflata and Skryabinia bolivari), are more usually found in
farm-reared birds (see Millán et al. 2004a). These were found in our study
parasitizing birds sampled in the managed areas only.
Furthermore, two of these species were found in adult birds in our study.
Partridges released for hunting are usually juveniles. Thus, the presence of
these parasites in adult birds suggests: a) that there had been a transmission
from farmed to wild birds; b) that we found birds released in the previous year
that had kept the infection for a whole year; or c) that some adult birds were
released. Releasing of farm-bred birds had occurred two months before the
samples for this study were taken. This implies that the partridges parasitized
by these helminths must be birds that survived in the field for two months, or
even wild birds. Any of these hypotheses are consistent with the introduction of
these parasites into the field through releases of their farm-bred hosts.
The transmission of parasites from released partridges to wild birds could be
facilitated by an increased parasite excretion after releasing, as experimentally

50
shown in the case of the pheasant (Villanúa et al. 2006a). This in turn could be
due to the absence of antiparasite treatment and the stress immunodepression
that follows releasing into an unknown habitat.
The introduction of new pathogens into a wild population can be an
important conservation problem because natural populations could have lower
resistance to a pathogen to which they have had no previous contact (Newton
1998). In our study area, some endangered steppe birds share habitat with the
red-legged partridge. So, the introduction of new non-specific parasites
supposes an additional problem for their conservation (Villanúa et al. 2007a).
The detection of higher parasite burdens in the managed areas in this study
could be an indicator of the low effectiveness of the preventive measures to
reduce parasites in the released animals. Current preventive measures applied
prior to the release of farm-reared game birds (if they exist) are based on: i) a
coprological analysis performed in faeces collected in the aviary soil, and ii)
antiparasite treatment following the same protocol that is used in the case of
poultry (Villanúa et al. 2007b). The first point of this protocol has the problem
that there are a lot of factors such as host reproductive status (Ruíz de Ybañez
et al. 2004), weather (Vicente et al. 2005), season (Kumba et al. 2003), random
day-to-day variations (Giver et al. 2000), phase of the parasitic infection (Giver
et al. 2000), or hour of sampling (Villanúa et al. 2006b), that can modify
propagule excretion, and are not always considered. In addition, the
effectiveness of antiparasite protocols in game species is insufficient to prevent
introducing parasites into the field as recently shown in the case of Albendazole
treatment against Aonchoteca caudinflata and H. gallinarum in red-legged
partridges (Villanúa et al. 2007b).
If the preventive protocols are not enough to eliminate the parasites from the
released birds, the other option taken is antiparasite treatments administered in
the field. Previous studies evaluated the effect of administering anthelminthic
drugs in the field together with the supplementary food (Woodburn et al. 2002;
Draycott and Sage 2005). These authors found that the birds given anthelmintic
treatment and supplementary food had significantly lower worm burdens and
increased their productivity. Nevertheless, some authors think that treatment of
free-living birds is not practical (Cole 1999) and the pros and cons of the

51
administration of drugs to wild birds have to be considered carefully, taking
ethical and public health concerns into account (Höfle et al. 2004).
On the other hand, we must remark that the two sampled game states with
wild partridges were adjacent to one of the managed states. Thus, our results
indicate that wild partridge populations have lower parasite diversity, prevalence
and burdens that adjacent populations where thousands of farm-bred partridges
are released yearly, that is, that the transmission of parasites form farmed birds
to wild birds is relatively limited to the hunting lands were releases are
performed, but may not affect so much neighbour populations. This may be due
to natural factors (poor dispersal ability of partridges in general, or of farmed
birds in particular) or to management actions (maintenance of released
partridges within the boundaries of game states by pursuing them, or intense
hunting removing most released partridges before they may disperse). In any
case, this result suggests that when releases are interrupted in a given hunting
land, the problem of parasite transmission may be largely avoided. In fact, this
may be the main reason explaining why when sampling wild and released
populations in Spain we still find differences in parasite communities, albeit
millions of partridges have been released during last decades all over Spain.
As we have shown, lower body condition found in juvenile partridges from
the managed areas was not apparently related with parasites. It can be
explained by two different causes: a) a deficient muscle development caused by
the farm-rearing (in case a significant part of the juvenile birds sampled in these
areas were farm-reared partridges); or b) the effect of other pathogens such as
bacteria or viruses, that may equally be introduced along with released birds.
However, more specific studies are needed to test this hypothesis.
In conclusion, the present study supports the initial hypothesis that hunting
estates with partridge releases are introducing helminth parasites into the
ecosystem. Thus, improved controls are necessary to prevent associated risks
for sustainable game management and conservation.
ACKNOWLEDGEMENTS
The authors thank S. J. Luna, E. Biescas, M.P. Martín and E. Chico for their
assistance in the field, and P. Talavera for help in the lab. This is a contribution
to the joint project CSIC/Principado de Asturias, and to CYCIT projects MCYT-

52
REN200307851/GLO and CGL2004-02568/BOS. Lorenzo Pérez-Rodríguez
had a FPU grant from the Spanish Ministerio de Educación y Ciencia and
Fabián Casas from Junta de Comunidades de Castilla-La Mancha.
REFERENCES
1. Aebischer, N.J., & Potts, G.R. (1994) Red–legged partridge. In: Tucker
GM, Heath MF (eds) Birds in Europe. Their Conservation Status. Birdlife
Conservation Series nº3, Birdlife International, Cambridge, UK.
2. Andersson, S. (1992) Female preference for long tails in lekking Jackson´s
widowbirds: experimental evidence. Journal of Animal Behaviour, 43: 379-
388
3. Bernabeu, R.L. (2000) Evaluación económica de la caza en Castilla-La
Mancha, Ph.D. Thesis. Castilla-La Mancha University, Spain
4. Birkan, M. (1977) Analyse des tableaux de chasse de perdrix. Courbes
d´eclosion, structure et dynamique des populations, plan de chasse. In
Pesson P, Birkan M (eds) Ecologie du petit gibier et aménagement des
chasses. Gauthier Villars: 55-77
5. Capelo, M. & Castro Pereira, D. (1996) Sobrevivência e dispersão de
perdizes (Alectoris rufa L.) largadas em duas operações de repovoamento
cinegético. Revista Florestal, 9: 245- 253
6. Carvalho-Varela, M. & Ferradeira, C. (1997) Parasitas e paraitoses da
perdiz-vermelha (Alectoris rufa) na Peninsula Ibérica. Algunas
consideraçoes sobre a clinica parasitaria da sua criaçaoem cativeiro.
Colección de Monográficas, Junta Extremadura, Cáceres, España. I
Congreso Internacional de Medio Ambiente y Veterinaria
7. Calvete, C., Estrada, R., Lucientes, J., Estrada, A. & Telletxeta, I. (2003)
Correlates of Helminth community in the red-legged partridge (Alectoris rufa)
in Spain. Parasitology, 89(3): 445-51
8. Cole, R.A. (1999) Trichomoniasis. pp. 201-206. In: Fried M, Frason JC (eds)
Field Manual of Wildlife Diseases. U.S. Department of Interior. National
Geological Survey.
9. Cordero del Campillo, M. & Rojo, F.A. (1999) Parasitología Veterinaria.
McGraw-Hill-Interamericana, Madrid

53
10. Draycott, R.A.H. & Sage, R.B. (2005) Parasite control and breeding
success in pheasants (Phasianus colchicus). In: Pohlmeyer K (ed) Extended
Abstracts of the XXVIIth Congress of the International Union of Game
Biologists, Hannover, pp 76-77
11. Duarte, J. & Vargas, J.M. (2004) Field interbreeding of released Farm-
reared red-legged partridges (Alectoris rufa) with wild ones. Game and
Wildlife Science, 21: 55-61
12. Give,r H., de Vlas, S.J., Johansen, M.V., Christensen, N.O. & Nansen, P.
(2000) Schistosoma japonicum: day to day variation in excretion and
hatchability of parasite eggs in the domestic pig Suis suis. Expl Parasitology,
95: 8-18
13. Gortázar, C., Villafuerte, R. & Martín, M. (2000) Success of traditional
restocking of red-legged partridge for hunting purposes in areas of low
density of northeast Spain Aragón. Zeitschrift für Jagdwissenschaft, 46: 23-
30
14. Gortázar, C., Villafuerte, R., Escudero, M.A. & Marco, J. (2002) Post-
breeding densities of the red-legged partridge (Alectoris rufa) in
agrosystems: a large scale study in Aragón, Northeastern Spain. Zeitschrift
für Jagdwissenschaft, 48: 94-101
15. Höfle, U., Gortazar, C., Ortiz, J.A. & Knispel, B. (2004) Outbreak of
trichomoniasis in a woodpigeon wintering roost. European Journal of Wildlife
Research, 50: 73-77
16. Kumba, F.F., Katjivena, H., Kauta, G. & Lutaaya, E. (2003) Seasonal
evolution of faecal egg output by gastrointestinal worms in goats on
communal farms in eastern Namibia. Ondestepoort Journal of Veterinay
Research, 70(4): 265-271
17. López-Neyra, C.R. (1947) Helmintos de los vertebrados ibéricos. Consejo
Superior de Investigaciones Científicas, Patronato “Santiago Ramón y
Cajal”, Instituto Nacional de Parasitología, Granada, Spain
18. Melhorn, H., Düwell, D. & Raether, W. (1992) Atlas de Parasitología
Veterinaria. GRASS ediciones, Spain
19. Millan, J., Gortazar, C., Martin-Mateo, M.P. & Villafuerte, R. (2004a)
Comparative survey of the ectoparasite fauna of wild and farm-reared red-

54
legged partridges (Alectoris rufa), with an ecological study in wild
populations. Parasitology Research, 93 (1): 605-611
20. Millán, J., Gortázar, C. & Villafuerte, R. (2004b) A comparison of the
helminth faunas of wild and farm-reared red-legged partridges. Journal of
Wildlife Management, 68(3): 701-707
21. Millán, J., Gortázar, C. & Villafuerte, R. (2004c) Ecology of nematode
parasitism in red-legged partridges (Alectoris rufa) in Spain. Helminthologia,
41(1): 33-37
22. Newton, I. (1998) Population Limitation in Birds. Academic Press, London
23. Pérez-Rodríguez, L., Blas, J., Viñuela, J., Marchant, T.A. & Bortolotti,
G.R. (2006) Condition and androgen levels: are condition-dependent and
testosterone-mediated traits two sides of the same coin? Journal of Animal
Behaviour, 72: 97-103
24. Ruíz de Ybánñez, M.R., Goyena, M., Abaigar, T., Garijo, M,, Martínez-
Carrasco, C., Espeso, G., Cano, M. & Ortiz, J.M. (2004) Periparturient
increase in faecal egg counts in a captive population of mohor Gazella
(Gazella dama mhorr). The Journal of Veterinary Record, 154: 49-52
25. Sáenz de Buruaga, M., Lucio, A. & Purroy, F.J. (2001) Reconocimiento de
sexo y edad en especies cinegéticas. EDILESA eds, León
26. Skryabin, K.I. (1991) Key to parasitic nematodes. E.J.Brill Publishing,
Leiden, The Netherlands
27. Tarazona, J.M., Sanz–Pastor, A. & de la Cámara, R. (1979) Helmintos y
helmintiosis de la perdiz roja (Alectoris rufa). Anales del INIA, Higiene y
Sanidad Animal, 4: 55-68
28. Tompkins, D.M., Dickson, G. & Hudson, P.J. (1999) Parasite-mediated
competition between pheasant and grey partridge: a preliminary
investigation. Oecologia, 119: 378-382
29. Tompkins, D.M., Greenman, J.V. & Hudson, P.J. (2001) Differential
impact of shared nematode parasite on two gamebird host: implications for
apparent competition. Parasitology, 122: 187-193
30. Vicente, J., Fierro, Y. & Gortázar, C. (2005) Seasonal dynamics of the
fecal excretion of Elaphostrongylus cervi (Nematoda, Metastrongyloidea)
first-stage larvae in Iberian red deer (Cervus elaphus hispanicus) from
southern Spain. Parasitology Research, 95(1): 60-4

55
31. Viggers, K.L. & Lindenmayer, D.B., Spratt, D.M. (1993) The importance of
disease in reintroduction programmes. Wildlife Research, 20: 687-698
32. Villanúa, D., Acevedo, P., Höfle, U., Rodríguez, O. & Gortazar, C.
(2006a) Changes in parasite transmission stage excretion after pheasant
release. Journal of Helminthology, 80(3): 313-318
33. Villanúa, D., Pérez-Rodríguez, L., Gortázar, C., Höfle, U. & Viñuela, J.
(2006b) Avoiding bias in parasite excretion estimates: the effect of sampling
time and type of faeces. Parasitology, 133: 251-259
34. Villanúa, D., Casas, F., Viñuela, J., Gortázar, C., García de la Morena,
E.L. & Morales, M.B. (2007a) First occurence of Eucoleus contortus in a
little bustard Tetrax tetrax. A negative effect of red-legged partridge Alectoris
rufa releases on steppe bird conservation?. Ibis (in press) doi:
10.1111/j.1474-919x.2006.00620.x
35. Villanúa, D., Pérez-Rodríguez, L., Rodríguez, O., Viñuela, J. & Gortazar,
C. (2007b) How effective is pre-release nematode control in farm reared
red-legged partridges (Alectoris rufa)?. Journal of Helminthology, 81(1): 101-
3
36. Woodburn, M., Sage, R.B. & Carroll, J.P. (2002) The efficacy of a
technique to control parasitic worm burden in pheasants (Phaisanus
colchicus) in the wild. Zeitschrift für Jagdwissenschaft, 48: 364-372

56
V. CAPITULO 2
Variaciones en el resultado de los análisis coprológicos
preventivos actuales en función del tipo de heces analizadas y
la hora de recogida de las mismas.
Avoiding bias in parasite excretion estimates: the effect of sampling time and
type of faeces. Villanúa, D., Pérez-Rodríguez, L, Gortázar, C., Höfle, U. &
Viñuela, J. (2006). Parasitology,133: 251-259

57
V. CAPITULO 2
Variaciones en el resultado de los análisis coprológicos
preventivos actuales en función del tipo de heces analizadas y
la hora de recogida de las mismas.
Avoiding bias in parasite excretion estimates: the effect of sampling time and
type of faeces. Villanúa, D., Pérez-Rodríguez, L, Gortázar, C., Höfle, U. &
Viñuela, J. (2006). Parasitology,133: 251-259
RESUMEN
El creciente interés de los ecólogos en las relaciones parásito-hospedador
hace que hayan proliferado los estudios en los que se recurre a diferentes
métodos con los que estimar el grado de parasitación. En el caso de los
parásitos intestinales, el método más utilizado es la cuantificación de
propágulos parasitarios en heces mediante técnicas coprológicas. Sin
embargo, la excreción parasitaria puede verse influenciada por numeros
factores tanto endógenos como exógenos. La identificación de estos factores
es imprescindible para obtener resultados correctos.
En el presente estudio, se ha analizado el efecto de la hora de recogida de
las heces en el resultado del análisis coprológico cuantitativo para dos
parásitos intestinales, Aonchoteca caudinflata y Eimeria sp., presentes en la
perdiz roja (Alectoris rufa). Además, se evaluaron también las posibles
diferencias existentes en el conteo de parásitos en heces corrientes y en heces
procedentes de los ciegos.
En Octubre de 2004, ocho perdices rojas fueron separadas durante un brote
de capilariasis en una población de granja. Tras confirmar la infección por
capilarias en todas las aves y por coccidios en seis de ellas, se inició un
protocolo de recogida de heces de manera individual en cuatro momentos del
día y durante un periodo de tiempo de varios días.
Se pudo detectar una importante variación en los valores del análisis
coprológico de ambos parásitos en función de la hora de recogida, con un claro
y constante incremento de la excreción conforme avanzaba el día. El día de
recogida influyó de manera significativa en los dos casos, así como su

58
interacción con el individuo en el caso de la excreción de capilaria. Además, los
propágulos de capilaria fueron más abundantes en las heces de intestino,
mientras que en el caso de los coccidios el patrón fue el opuesto, con mayor
recuento de ooquistes en las heces de ciego.
Nuestros resultados muestran que la hora del día en que se recoge la
muestra de heces afecta drásticamente al resultado. De ahí que sea necesario
estandarizar la hora de recogida de la muestra para evitar errores. De manera
similar, en el caso de especies con ciegos muy desarrollados, se debe tener en
cuenta el tipo de heces recogidas (a poder ser del tramo donde el parásito sea
más abundante), con el fin de obtener los resultados correctos. Por último, en
este trabajo se pone de manifiesto la necesidad de profundizar en el
conocimiento acerca del sistema parástio-hospedador, con el objetivo de
desarrollar un protocolo de muestras adecuado que permita obtener datos más
fiables.
ABSTRACT
The interest of ecologists on host-parasite relationships has lead to an
increasing number of studies involving indirect methods of parasite burden
estimation. With regards to intestinal parasites, the most widely employed
method is the quantification of parasite propagules in faeces. However, parasite
excretion in faeces may be subjected to a certain degree of variation due to
endogenous or exogenous factors. The identification of those influencing factors
is required to obtain reliable results.
In this paper we analyse the effect of the hour of the day at which samples
are collected in propagule counts of two intestinal parasites infecting the red-
legged partridge: the capillarid nematode Aonchoteca caudinflata and coccidia
of the genus Eimeria. Also, we tests whether there are differences in propagule
counts between caecal and intestinal faeces.
In October 2004, eight red-legged partridges were isolated during an
outbreak of capillariasis in a captive population. After confirming the capillarid
infection in these eigth individuals and coccidian infection in six out of them,
individual faecal samples were daily collected at four different hours during
several days.

59
Hour of the day exerted a very strong effect in propagule counts, excretion
of both types of parasites showing a clear and constant increase from dawn to
dusk. Day of sampling affected significantly in both cases and interacted
significantly with the individual in the case of capillarid eggs. Also, capillarid
eggs were more abundant in intestinal than in caecal faeces, whereas the
inverse pattern was found for coccidian oocysts.
Our results show that hour of the day at which samples are collected can
drastically affect parasite excretion estimates. Standardization of the hour of
sample collection or statistical control of this variable is therefore
recommendable to prevent this bias. Similarly, in the case of bird species with
long caeca, consistent collection of one type of faeces (ideally that one where
propagules are most abundant) may avoid important errors in parasite burden
estimates.
Finally, this study stresses the importance of gathering more information
about the studied host-parasite system in order to develop adequate sampling
methods leading to obtain more reliable data.
KEYWORDS: Alectoris rufa, Aonchoteca caudinflata, Eimeria, nematoda,
coccidia
INTRODUCTION
In the last two decades the interest of behavioral ecologists in the effect of
parasites on host fitness has increased. Parasites may affect host body
condition (Hall 1985; Gulland 1995; Delahay et al 1995), survival probabilities
(Vorísek et al. 1998; Murray et al. 1997), some reproductive parameters (Newey
&Thirgood 2004; Albon et al. 2002) or population dynamics (Holmes 1982;
Hudson et al. 1998; Tompkins et al. 2001). Therefore, parasites seem to be an
important factor in host life-history and may exert a strong selective force on
host evolution (Clayton & Moore 1997).
This relatively recent focus on host-parasite interaction has lead to an
increasing number of ecological empirical studies in which parasite load is
related to different measures of host fitness. It uses to be relatively easy to
determine the prevalence and intensity of ectoparasite infection as they can be
directly counted or estimated by exploring the skin, hair or feathers of the

60
individual. In contrast, determining the level of infection by endoparasites is
more difficult, especially when the study requires the individual to remain in
perfect conditions after sampling. Regarding intestinal endoparasites, the most
common approach to solve this problem is to estimate prevalence and intensity
of the infection by the analysis of faecal samples and the quantification of
parasite propagules, typically eggs or larvae (Gordon & Whitlock 1939; Shaw &
Moss 1989; Guyatt & Bundy 1993). However, propagule counts in faeces may
be subjected to a great within-individual variation due to factors like host
reproductive status (Ruiz de Ybañez et al. 2004), weather (Rickard &
Zimmerman 1992; Vicente et al. 2005), season of the year (Shaw & Moss 1989;
Theodoropoulos et al. 1998; Kumba et al. 2003), random day-to-day variations
(Yu et al. 1998; Giver et al., 2000; Utzinger et al. 2001), phase of the parasitic
infection (Giver et al. 2000; Cordero del Campillo et al. 1999) or hour of the day
(Boughton 1933; Brawner & Hill 1999). For example, Brawner & Hill (1999)
showed that a single factor such as hour of the day at which samples are
collected can drastically alter the results of assessment of coccidian prevalence
and individual parasite burden in the house finch (Carpodacus mexicanus).
Therefore, accurate determination of the effect of these variables on
propagule excretion is essential for a correct assessment of parasitic infection
and for a successful testing of the hypothesis studied. Unfortunately, despite the
remarked relevance of knowing the effects of these factors, no accurate
information is available for host-parasite systems with little medical or veterinary
importance, that are precisely the more studied by behavioural and evolutionary
biologists.
In this paper we analyse the day-to-day and daily variation in the excretion
of parasite propagules in red-legged partridges (Alectoris rufa L.) naturally
infected with two different intestinal parasites: coccidia of the genus Eimeria and
the capillarid nematode Aonchoteca caudinflata. Eimeria coccidia exhibit both
sexual and asexual phases in their life cycle. The asexual phase may occur in
the intestinal epithelium or as a diffuse infection in several other organs. The
sexual phase takes place only in the epithelium of the intestine or intestinal
caeca (depending of the species) and results in the production of oocysts that
are elimininated in the faeces of the host (Cordero del Campillo et al. 1999). A.
caundinflata is an heteroxenous parasite of the small intestine of gallinaceus

61
and anatid birds (Anderson 2000). Female worms lay eggs that mature after
ingestion by earthworms (Allobophora caliginosa, Eisenia foetida), the
necessary intermediate hosts.
In many avian taxa, like Galliforms and Anseriforms, intestinal caeca are
specially elongated. In these species, caecal faeces represent a significant
proportion of the total amount of faeces produced and can be easily
distinguished from intestinal faeces by their appearance. In this paper we also
analyse the differences in propagule content of both kinds of faeces in the red-
legged partridge. We finally discuss the implications of our results in the design
of and data analysis of empirical studies involving parasite estimation from
faecal counts.
MATERIAL AND METHODS
Data collection took place during October 2004 in the experimental red-
legged partridge farm of the IREC in Ciudad Real (Spain). During late summer-
autumn of that year there was an outbreak of capillariasis in one of the outdoor
aviaries of the farm. As soon as the parasite was identified and before starting
any medication, a sample of 15 birds of the affected aviary was isolated in
individual outdoor elevated cages and individual samples of feces were
collected. The coprological analysis showed that eight of the birds (six females
and two males) were infected by capillarids and these were subsequently
employed for the experiment. Also, six of them were infected by coccidia,
allowing the combined analysis of the daily variation of excretion of both
parasites in the same sample of birds.
During the days 5th, 6th, 12th, 13th, 14th, 15th and 16th of October individual
faecal samples were collected from all birds (except from one of them the day
14th due to an error of sampling) at 08:00, 12:00, 16:00 and 19: 30 hours.
Faecal samples were obtained by placing a large piece of fresh paper on the
ground just below each one of the cages slightly before the designated times
The first droppings produced by each bird during the following 30 minutes were
collected and stored in Eppendorf tubes. Most of the faeces obtained were
intestinal faeces, which is the most common type of faeces produced by
Galliforms. However, when caecal faeces where also found at sampling times,
they were collected separately allowing a posterior comparison. Caecal faeces

62
can be easily and unambiguously distinguished from intestinal faeces by their
colour (dark brown in the former, green-pale brown in the later), texture (soft
and homogeneous in the former, granular in the later) and by the much more
intense odour of caecal faeces.
Quantitative analysis of the parasite propagules (oocysts in the case of
coccidia, eggs in the case of capillarids) present in each faecal sample were
carried out by flotation in a known volume of saturated SO4Zn solution and
counting in MacMaster chambers (Melhorn et al. 1992). The concentration of
oocysts and capillarid eggs was finally referred to the weight of the faecal
sample analysed, obtaining the number of propagules per gram of faeces.
It should be noted that no any capillarid nematode can be identified up to the
species level just by observing its eggs. Therefore, at the end of the experiment
birds were sacrificed as a part of another experiment and adult worms were
identified according to Skryabin (1991), confirming the presence of Aonchoteca
caudinflata.
Statistical analysis
We employed General Linear Mixed Models (GLMMs) for the analysis of the
effect of sampling time on oocyst and capillarid egg counts. As parasite
propagule counts were normally distributed after log10+1 transformation, we
used normal distribution of errors and an identity link function (GLMMs macro of
SAS, Littell et al. 1996). Capillarid eggs or coccidian oocysts (after the cited
transformation) were introduced as dependent variables in the models. Day and
hour of the day (nested in day) were introduced as categorical factors and the
individual was considered in the model as a random variable. As day-to-day
variations in propagule excretion may be caused by endogenous factors, the
effect of this variable could differ for each bird. Therefore, the interaction
between day and individual was also entered in the model as a fixed factor.
Propagule excretion may vary in the time due to changes in the degree of
infection of the individual. Therefore, to minimize the effect of this on our
analysis of day-to-day variation we restricted our analysis to data from the five
consecutive days (12 th to 16 th).
However, as the analysis revealed an effect of the day in the models for
capillarids and coccidia (see results) two further GLMMs were performed in

63
order to assess whether there was a common tendency during the five
consecutive days previously analysed. In both models the number of
propagules (either capillarid eggs or coccidian oocysts) per gram of faeces
(after log10+1 transformation) excreted at 19: 30 hours of each day was entered
as dependent variable whereas day -as a continuous variable- and the
interaction between day and individual were entered as fixed factors. The
individual was again entered as a random factor.
To test for differences between caecal and intestinal faeces, propagule
content of caecal and intestinal faecal samples collected at the same hour of
the same day from each bird were compared by means of Wilcoxon Matched
Pairs Tests (raw data were employed in this analysis because the larger
number of zero values in this data set made impossible data normalization).
RESULTS
Effect of the time of the day and day-to-day variation on propagule excretion
GLIMMs results for capillarid eggs and coccidian oocysts are shown in Table
1. In the case of capillarid egg excretion the hour of the day at which samples
where collected was the most important factor, explaining the 42.17 % of the
deviance of the model.
Capillarid egg shedding followed the same temporal pattern in all birds,
showing a clear and constant increase from dawn to dusk that was repeated
during all the days of sampling (Fig. 1). In fact, the amount of capillarid eggs in
faeces was on average 27.4±14.2 times higher in samples collected at 19: 30
than in those samples collected at 8:00 from the same birds.
Day of sampling also exerted a significant effect (explaining a 7.6% of the
deviance). As can be observed in Fig. 1, in some individuals (e.g. bird 54, bird
59) the line signalling the daily pattern for some days was almost five times
higher than for other days, showing a strong day-to-day variation. In contrast,
such day-to-day variation was minimal in other individuals (e.g. bird 61, bird 72).
Consequently, the deviance explained in the final model was greater for the
interaction between day and individual (27.5%) than for day as a single factor,
suggesting that day-to day-variation in capillarid egg excretion followed a
particular pattern for each bird. To assess whether such effect of the day of
sampling indicated a common tendency during the five consecutive days of
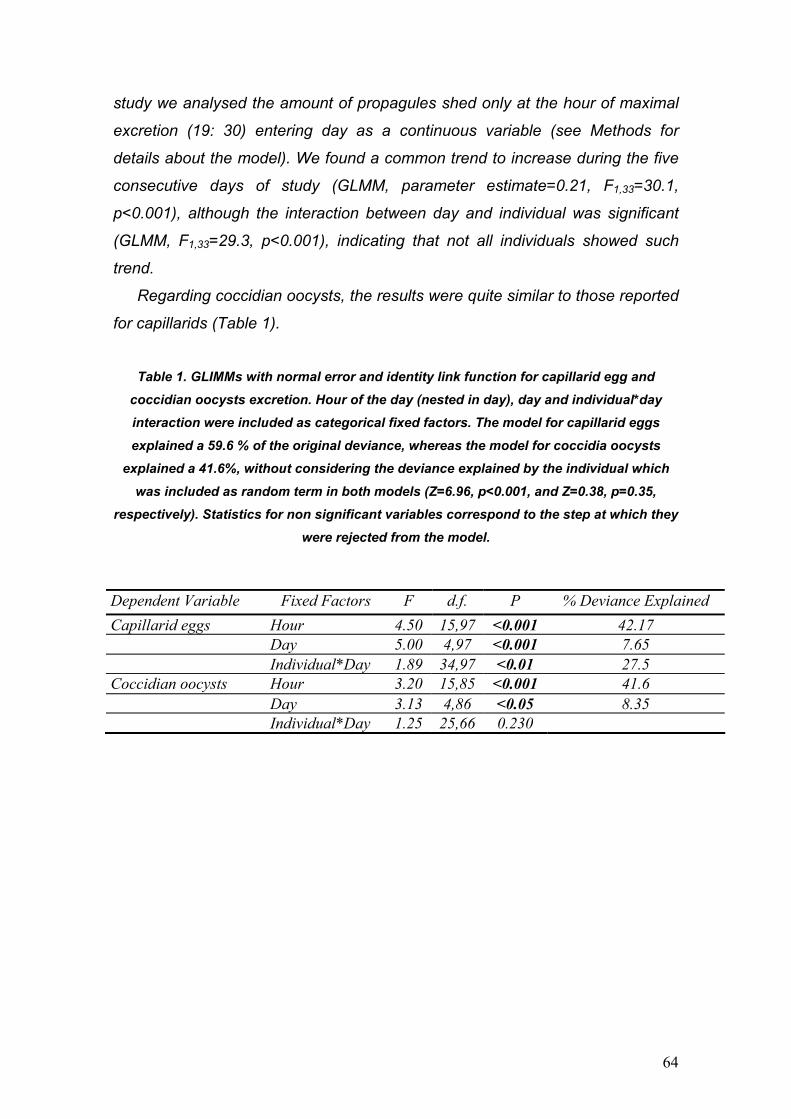
64
study we analysed the amount of propagules shed only at the hour of maximal
excretion (19: 30) entering day as a continuous variable (see Methods for
details about the model). We found a common trend to increase during the five
consecutive days of study (GLMM, parameter estimate=0.21, F1,33=30.1,
p<0.001), although the interaction between day and individual was significant
(GLMM, F1,33=29.3, p<0.001), indicating that not all individuals showed such
trend.
Regarding coccidian oocysts, the results were quite similar to those reported
for capillarids (Table 1).
Table 1. GLIMMs with normal error and identity link function for capillarid egg and
coccidian oocysts excretion. Hour of the day (nested in day), day and individual*day interaction were included as categorical fixed factors. The model for capillarid eggs
explained a 59.6 % of the original deviance, whereas the model for coccidia oocysts explained a 41.6%, without considering the deviance explained by the individual which
was included as random term in both models (Z=6.96, p<0.001, and Z=0.38, p=0.35, respectively). Statistics for non significant variables correspond to the step at which they
were rejected from the model.
Dependent Variable Fixed Factors F d.f. P % Deviance Explained Capillarid eggs Hour 4.50 15,97 <0.001 42.17 Day 5.00 4,97 <0.001 7.65 Individual*Day 1.89 34,97 <0.01 27.5 Coccidian oocysts Hour 3.20 15,85 <0.001 41.6 Day 3.13 4,86 <0.05 8.35 Individual*Day 1.25 25,66 0.230

65
Figure 1 Number of capillarid eggs per gram of faeces from eight red legged partridges collected at four different hours of the day and during five consecutive days. Dashed
lines show daily patterns whereas solid lines indicate mean values. Error bars have been eliminated for clarity.
Bird 72
0
900
1800
2700
8:00 12:00 16:00 19:30
Bird 71
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 69
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 59
0
9000
18000
27000
8:00 12:00 16:00 19:30
Bird 57
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 62
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 61
0
3000
6000
9000
8:00 12:00 16:00 19:30
Hour of sampling
Cap
illar
ideg
gspe
rgr
amof
faec
es
Bird 72
0
900
1800
2700
8:00 12:00 16:00 19:30
Bird 71
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 69
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 59
0
9000
18000
27000
8:00 12:00 16:00 19:30
Bird 57
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 62
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 61
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 72
0
900
1800
2700
8:00 12:00 16:00 19:30
Bird 71
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 69
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 59
0
9000
18000
27000
8:00 12:00 16:00 19:30
Bird 57
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 62
0
3000
6000
9000
8:00 12:00 16:00 19:30
Bird 61
0
3000
6000
9000
8:00 12:00 16:00 19:30
Hour of sampling
Cap
illar
ideg
gspe
rgr
amof
faec
es
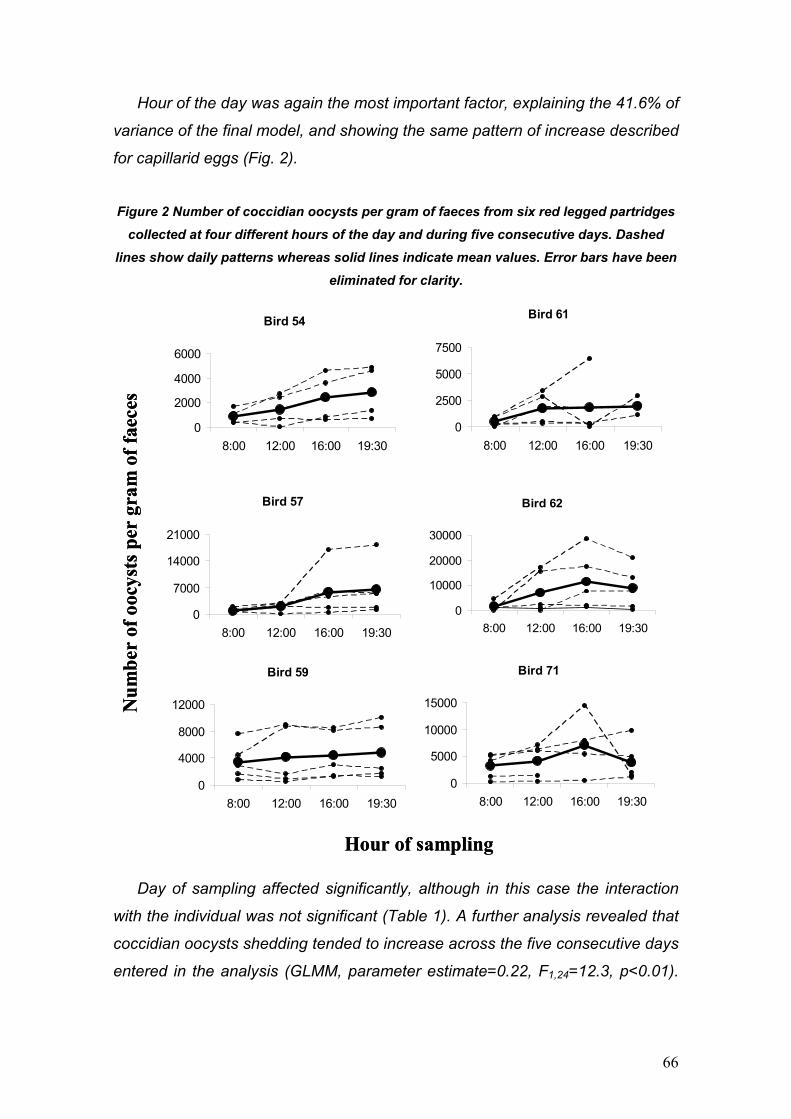
66
Hour of the day was again the most important factor, explaining the 41.6% of
variance of the final model, and showing the same pattern of increase described
for capillarid eggs (Fig. 2).
Figure 2 Number of coccidian oocysts per gram of faeces from six red legged partridges
collected at four different hours of the day and during five consecutive days. Dashed lines show daily patterns whereas solid lines indicate mean values. Error bars have been
eliminated for clarity.
Bird 71
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 57
0
7000
14000
21000
8:00 12:00 16:00 19:30
Bird 59
0
4000
8000
12000
8:00 12:00 16:00 19:30
Bird 62
0
10000
20000
30000
8:00 12:00 16:00 19:30
Bird 61
0
2500
5000
7500
8:00 12:00 16:00 19:30
Num
ber
ofoo
cyst
sper
gram
offa
eces
Hour of sampling
Bird 71
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 57
0
7000
14000
21000
8:00 12:00 16:00 19:30
Bird 59
0
4000
8000
12000
8:00 12:00 16:00 19:30
Bird 62
0
10000
20000
30000
8:00 12:00 16:00 19:30
Bird 61
0
2500
5000
7500
8:00 12:00 16:00 19:30
Bird 71
0
5000
10000
15000
8:00 12:00 16:00 19:30
Bird 54
0
2000
4000
6000
8:00 12:00 16:00 19:30
Bird 57
0
7000
14000
21000
8:00 12:00 16:00 19:30
Bird 59
0
4000
8000
12000
8:00 12:00 16:00 19:30
Bird 62
0
10000
20000
30000
8:00 12:00 16:00 19:30
Bird 61
0
2500
5000
7500
8:00 12:00 16:00 19:30
Num
ber
ofoo
cyst
sper
gram
offa
eces
Hour of sampling
Day of sampling affected significantly, although in this case the interaction
with the individual was not significant (Table 1). A further analysis revealed that
coccidian oocysts shedding tended to increase across the five consecutive days
entered in the analysis (GLMM, parameter estimate=0.22, F1,24=12.3, p<0.01).
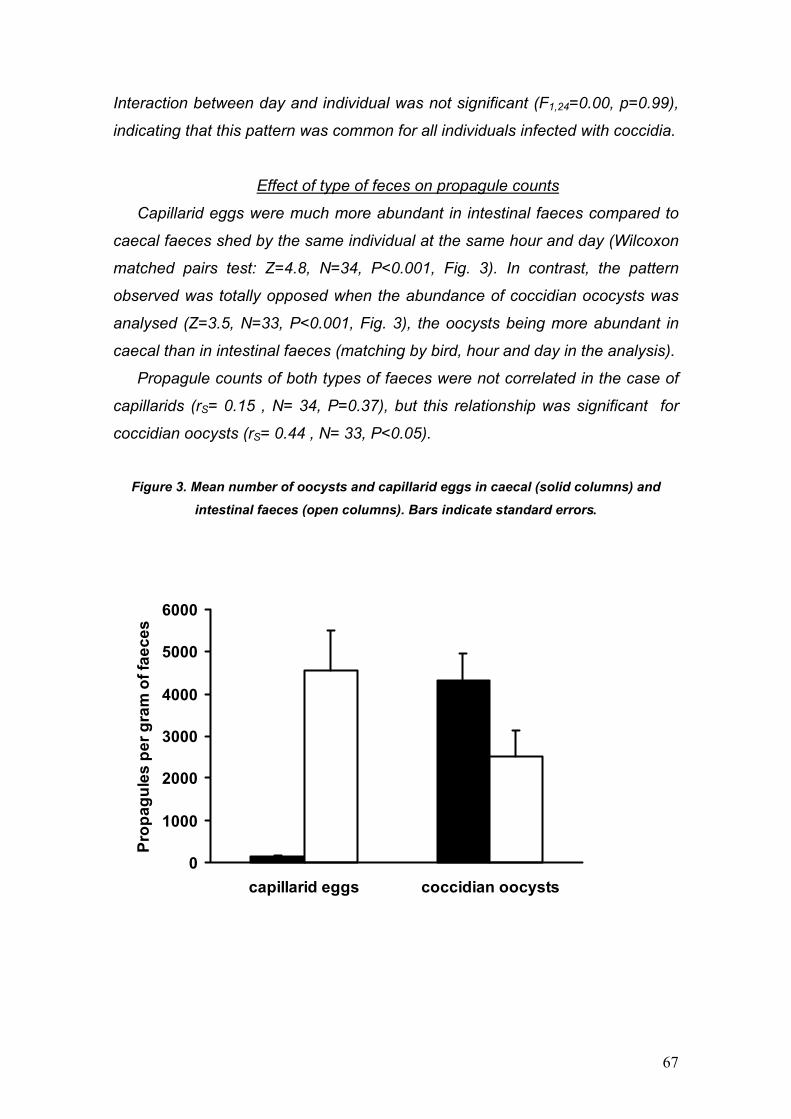
67
Interaction between day and individual was not significant (F1,24=0.00, p=0.99),
indicating that this pattern was common for all individuals infected with coccidia.
Effect of type of feces on propagule counts
Capillarid eggs were much more abundant in intestinal faeces compared to
caecal faeces shed by the same individual at the same hour and day (Wilcoxon
matched pairs test: Z=4.8, N=34, P<0.001, Fig. 3). In contrast, the pattern
observed was totally opposed when the abundance of coccidian ococysts was
analysed (Z=3.5, N=33, P<0.001, Fig. 3), the oocysts being more abundant in
caecal than in intestinal faeces (matching by bird, hour and day in the analysis).
Propagule counts of both types of faeces were not correlated in the case of
capillarids (rS= 0.15 , N= 34, P=0.37), but this relationship was significant for
coccidian oocysts (rS= 0.44 , N= 33, P<0.05).
Figure 3. Mean number of oocysts and capillarid eggs in caecal (solid columns) and
intestinal faeces (open columns). Bars indicate standard errors.
0
1000
2000
3000
4000
5000
6000
capillarid eggs coccidian oocysts
Prop
agul
es p
er g
ram
of f
aece
s

68
DISCUSION
Our results indicate that the hour of the day may exert a very strong effect
on parasite propagule excretion. This point was confirmed in two different
parasite species coming from two extremely different taxa (Nematoda and
Protozoa) infecting the same bird host species (the red legged partridge). In
both cases propagule shedding increased as the day progressed, reaching
maximal values in the late afternoon. Scarce attention has been paid in the
literature to the existence of this kind of consistent daily trends in parasite
propagule excretion, especially for birds or host-parasite systems with little or
no relevance in human health or economy. For example, Giver et. al. (2000)
found no consistent pattern across individuals when comparing morning and
afternoon egg counts in faeces of domestic pigs artificially infected with the
trematode Schistosoma japonicum. In contrast, Boughton (1933) and Brawner
& Hill (1999) described a diurnal periodicity in the eliminiation of oocysts of the
genus Isospora in two avian passerine species (house sparrows and house
finches, respectively) consistent with the pattern found in this study. However,
the diurnal trend found for coccidia in our case does not seem to be as
pronounced as that found by Brawner & Hill (1999). This may be due to intrinsic
differences between host-parasite systems, or to the fact that the intensity of
coccidian infection in our experimental birds was not as high as those studied
by Brawner & Hill (1999). However, to the authors´ knowledge, this is the first
report of the existence of a consistent within-day pattern of excretion in a
nematode parasite of a bird species.
Although the hour was by far the most influencing factor in both parasites,
we also found an effect of the day of sampling. In particular, we found a
tendency to increase parasite propagule excretion during the course of the
study which was stronger for coccidian oocysts than for capillarid eggs. In our
study, samples were collected during five consecutive days just to exclude the
effect of any seasonal variation in propagule excretion as described in other
studies (e.g.: Shaw & Moss 1998; Theodoropoulos et al. 1998; Vicente et al.
2005). It is widely known that propagule excretion varies throughout the course
of a parasitic infection (Cordero del Campillo et al. 1999). Birds employed in this
study were naturally infected during an outbreak of capillariasis that affected our
captive population. As a result, birds may have been in different phases of the

69
parasitic infection when sampled, explaining why some birds showed a
tendency to increase the amount of capillarid eggs excreted whereas others did
not show this pattern. Unlike capillarids, infection by coccidia seemed to be
more benign, as revealed by low number of oocysts counted and the absence
of individuals showing clinical signs compatible with coccidiosis in the preceding
and following months. Probably this difference in the type and degree of
infection may explain the absence of interaction between day and individual in
the case of coccidia, although it may not be coherent with the fact that an
increasing between-day pattern was found in both parasites. One further
possible explanation is that repeated sample collection (that required the
investigator to go under the cages four times each day) may have stressed the
individuals during the five days of study. It is known that physiological stress
may lead to immunosuppression (Sapolsky 1992; Besedowsky & del Rey
1996), leading parasites to take advantage to the host and increase their
reproductive rate and propagule production. This stress-induced effect on
parasite propagule production may be relatively quick, even less than a week
both for nematodes and coccidia, as reflected by a recent study in ring-necked
pheasants (Phasianus colchicus)(Villanúa et al. 2006).
Apart from the effect of time of sampling, we found important differences in
propagule counts between types of faeces. In particular, capillarid eggs were
much more abundant in intestinal faeces than in caecal faeces, whereas the
pattern was the opposite for coccidian oocysts. This tendency was expected
attending to the part of the intestinal tract where the adult reproductive forms of
each parasite live. Adults of the capillarid Aonchoteca caudinflata live in the
intestine and therefore females release the eggs in the intestinal content. In
contrast, the oocysts found in our captive red-legged population belong to two
different species, Eimeria tenella and Eimeria colchici, both with sexual phases
that replicate in the epithelium of the caeca or in the caeca and final portion of
the intestine (the latter)(Cordero del Campillo & Rojo 1999). As a result, the
total number of oocysts counted (oocysts of both species are indistinguishable
unless they are sporulated, which was not the case in our samples) was higher
in caecal than in intestinal faeces.
In the last years the interest of behavioural and evolutionary ecologists on
parasites has increased due to their possible effects on host fitness and

70
evolution (Clayton & Moore 1997). Apart from ectoparasites, intestinal parasites
are the most commonly studied ones as possible factors affecting several
components of host fitness. Interestingly, both coccidia (Hillgarth 1990;
Buchholz 1995; Vorísek et al. 1998) and nematodes (Hudson et al. 1998;
Murray 2002; Newey & Thirgood 2004) are the most commonly endoparasite
taxa employed in such ecological studies, and most of those surveys that do not
involve culling the animal require indirect parasite load estimates based on
counts of the number of propagules in faeces. In parasites from these two taxa
we have found a marked diurnal periodicity in propagule excretion. However,
despite that the hour of the day at which samples are collected may be of vital
relevance, this variable is never controlled for in any study.
The great magnitude of within individual variation in propagule excretion
during the day may covey serious errors in non-invasive parasite load
estimates. Experiments in captivity may easily control for this effect by sampling
all the birds approximately at the same hour, preferentially in the late afternoon,
when propagule shedding is maximal and therefore differences between
individuals are maximised. However, such kind of data standardization is
difficult in the wild. Moreover, most of wild birds are trapped in the morning
hours, when propagule shedding is minimal, thus diminishing the power to
detect differences in intensities of parasite excretion between individuals. As
parasite estimates from birds sampled at morning and late afternoon hours may
not be comparable, the best way to solve this problem in studies in wild is to
control statistically for the hour of sampling in all the analysis by including it as a
covariable. Either the methodological standardization or the statistical control of
the effect of this variable may prevent from fatal errors and will lead to sounder
results. Similarly, studies where faecal samples are collected from the
environment instead of directly from the animal should take into account this
source of variability. Collection of fresh drops exclusively and during the same
hour of the day may be a good option, although the effect of weather in faecal
drying time should be considered. Alternatively, sample collection from places
where individuals are only during a determinate and known period of the day
(roosts, for example) may allow to increase the reliability of the results obtained.
With regards to the differences in parasite counts between types of faeces,
the main implication of our findings is that researchers should pay attention to

71
the life cycle of the parasite to develop the most appropriate protocol for sample
collection in species with long caeca like Galliforms or Anseriforms. In our
particular case, we have shown that parasite counts coming from intestine or
caeca are not comparable for this two parasite species. The ideal procedure
should be to count each parasite only in that type of faeces where it is more
abundant. However, caecal faeces are only a minor proportion of the total
amount of faeces produced by the bird. Therefore, it is very difficult to have the
opportunity to choose what type to collect, especially in the wild, and the
researcher will have to be satisfied with the type of sample excreted by the bird
when trapped (usually a intestinal drop). In our case, consistent collection of
intestinal faeces is suitable for capillarids, but seems to be acceptable for
coccidia too, as caecal and intestinal counts were correlated in the latter.
Although in the two parasites studied here the hour of the day and the type
of faeces were by far the most important factors affecting individual variation in
propagule counts, other possible factors may also affect. As said above, we
found an effect of the day of sampling that varied between individuals in the
case of capillarids. Other authors have found a day-to-day variation in
propagule counts that seems to be due to variations in propagule production by
the parasite or to aggregation of the propagules in the feces, leading to
sampling errors (e.g. Yu et al. 1998; Giver et al. 2000; Utzinger et al. 2001).
Also, great differences in propagule excretion may occur along the course of
infection (Giver et al. 2000; Cordero del Campillo et al. 1999). Moreover, in
some parasite species, fecal propagule counts may not necessarily reflect the
number of adult parasites present in the body of the individual (Welch et al.
1991). Density-dependent constraints may also alter the relationship between
parasite burden and faecal counts (Anderson & Shad 1985; Tompkins &
Hudson 1999). Hence, the use of propagule counts to indirectly estimate
parasite burden should ideally be addressed in any host-parasite system before
being employed.
It is remarkable that we found identical diurnal trends in two extremely
different taxa of parasites. Although previous studies in other coccidian species
showed the same pattern (Boughton 1933; Brawner & Hill 1999), more research
involving different parasite and host taxa is required to confirm whether such
kind of diurnal variation is common across host-parasite systems. This study

72
remarks the point made by other authors (McLennan & Brooks 1991; Zuk 1992;
Brawner & Hill 1999) about the necessity of gathering more information about
the studied host-parasite system studied. A survey to identify the best method
for assessing parasite prevalence and burdens is recommended as a previous
step for any host-parasite study (e.g.: Seivwright et al. 2004; Brawner & Hill
1999). A better understanding of the dynamics of infection as regards for
example fluctuations in propagule number within and between days or between
types of feces may lead to obtain more reliable data and give the opportunity of
a more throughout understanding.
ACKNOWLEDGEMENTS
We thank Elisa Pérez, Salvador Jesús Luna and Paqui Talavera for
assistance during sample collection and analysis and Jesús Martínez-Padilla for
statistical advice. Financial support was provided by the Research Project PAI-
02-006 of the Junta de Comunidades de Castilla-La Mancha and by the
agreement between CSIC and Principado de Asturias. Lorenzo Pérez-
Rodríguez was supported by a FPU grant from the Ministerio de Educación y
Ciencia.
REFERENCES
1. Albon, S.D., Stien, A., Irvine, R.J., Ropstad, R. & Halvorsen, O. (2002)
The role of parasites in the dynamic of a reindeer population. Proceding of
the Royal Society of London, Series B. Biological ciences, 269: 1625-1632.
2. Anderson, R. M. & Schad, G. A. (1985) Hookworm burdens and faecal egg
counts: an analysis of the biological basis of variation. Transactions of the
Royal Society of Tropical Medicine and Hygiene, 79: 812-825.
3. Anderson, R.C. (2000). Nematode parasite of the vertebrates: their
development and transmision. Second edition. CAB International,
Wallingford, United Kingdom.
4. Besedovsky, H.O. & del Rey, A. (1996) Immune-neuro-endocrine
interactions: facts and hypotheses. Endocrine reviews, 17: 64–97
5. Boughton, D. C. (1933) Diurnal gametic periodicity in Avian Isospora.
American Journal of Hygiene, 18: 161-184.

73
6. Brawner, W. R. III & Hill, G. E. (1999) Temporal variation in shedding of
coccidian oocysts: implications for sexual-selection studies. Canadian
Journal of Zoology, 77: 347-350.
7. Buchholz, R. (1995) Female choice, parasite load and ornamentation in
wild turkeys. Animal Behaviour, 50: 384-387.
8. Clayton, D. L. & Moore, J (1997) Host-parasite evolution: General
principles and avian models. Oxford University Press, Oxford.
9. Cordero del Campillo, M. & Rojo, F. A. (1999) Parasitología Veterinaria.
McGraw-Hill-Interamericana, Madrid.
10. Delahay, R.J., Speakman, J.R. & Moss, R. (1995) The energetic
consequences of parasitism: effect of a developing infection of
Trichostrongylus tennuis on Red Grouse (Lagopus lagopus scoticus) energy
balance, body weight and condition. Parasitology, 110: 473-482.
11. Giver, H., de Vlas, S. J., Johansen, M. V., Christensen, N. O. & Nansen,
P. (2000). Schistosoma japonicum: Day to day variation in excretion and
hatchability of parasite eggs in the domestic pig, Suis suis. Experimental
Parasitology, 95: 8-18.
12. Gordon, H. McL. & Whitlock, H. V. (1939) A new technique for counting
nematode eggs in sheep faeces. Journal of the Council for Scientific and
Industrial Research (Australia), 12: 50-52.
13. Gulland, F. M. D. (1995) Impact of Infectious Diseases on Wild Animals.
Ecology of Infectious Diseases in Natural Populations. (Eds B. T. Grenfell &
A. P. Dobson), pp. 20-51. Cambridge University Press, Cambridge.
14. Guyatt, H. L. & Bundy, D. A. P. (1993) Estimation of intestinal nematode
prevalence: influence of parasite matting patterns. Parasitology, 107: 99-
106.
15. Hall, A. (1985) Nutritional aspects of parasitic infection. Progress in food
and nutrition science, 9: 227-256.
16. Hillgarth, N. (1990) Parasites and female choice in the ring-necked
pheasant. American Zoologist, 30: 227-233.
17. Holmes, J.C. (1982) Impact of infectious disease agents in the population
growth and geographical distribution of animals. Population Biology of
Infectious Diseases (Eds M.A. &.R.M.M.R), pp. 37-51. Springer-Verlag,
Berlin.

74
18. Hudson, P.J., Dobson, A.P. & Newborn, B. (1998) Prevention of
population cycles by parasite removal. Science, 282: 2256-2258.
19. Kumba, F.F., Katjivena, H., Kauta, G. & Lutaaya, E. (2003) Seasonal
evolution of faecal egg output by gastrointestinal worms in goats on
communal farms in eastern Namibia. The Onderstepoort Journal of
Veterinary Research, 70(4): 265-71.
20. Littell, R. C., Milliken, G. A., Stroup, W. W. & Wolfinger, R. D. (1996) SAS
system for mixed models. SAS Institute. Cary. New York.
21. McLennan, D. A., & Brooks, D. R. (1991) Parasites and sexual selection: a
macroevolutionary perspective. The Quarterly Review of Biology, 66: 255-
286.
22. Melhorn, H., Düwell, D. & Raether, W. (1992) Atlas de Parasitología
Veterinaria. GRASS, Spain
23. Murray, D.L. (2002) Differential body condition and vulnerability to predation
of snowshoe hares. Journal of Animal Ecology, 71: 614-625.
24. Murray, D,L,, Cary, J.R. & Keith, L.B. (1997). Interactive effects of
sublethal nematodes and nutritional status on snowshoe hare vulnerability to
predation. Journal of Animal Ecology, 66: 264
25. Newey, S. & Thirgood, S. (2004) Parasite-mediated reduction in fecundity
of mountain hares. Proceedings of the Royal Society of London. Series B.
Biological sciences 271: 413-415.
26. Rickard, L.G. & Zimmerman, G.L. (1992) The epizootiology of
gastrointestinal nematodesof sheep, cattle and deer. Journal of
Helminthology, 69: 357-362.
27. Ruiz de Ybáñez, M.R., Goyena, M., Abrigar, T., Garito, M.M., Martínez-
Carrasco, C., Espeso, G., Cano, M. & Ortiz, J.M. (2004) Periparturient
increase in faecal egg counts in a captive population of mohor Gazella
(Gazella dama mhorr). Veterinary Record: 154, 49-52.
28. Sapolsky, R. M. (1992) Neuroendrocrinology of the stress response.
Behavioural endrocrinology. (Eds. Becker, J.B., Breedlove, S.M. & Crews,
D.). pp 287–324. MIT Press, Cambridge.
29. Seivwright, L. J., Redpath, S. M., Mougeot, F., Watt, L., & Hudson, P. J.
(2004) Faecal egg counts provide reliable measure of Trichostrongylus

75
tenuis intensities in free-living red grouse Lagopus lagopus scoticus. Journal
of Helminthology, 78: 69-76.
30. Shaw, J. L. & Moss, R. (1989) The role of parasite fecundity and longevity
in the success of Trichostrongylus tenuis in low density red grouse
populations. Parasitology, 99: 253-258.
31. Skryabin, K. I. (1991) Key to Parasitic Nematodes. E. J. Brill Publishing
Company. New Delhi.
32. Theodoropoulos, G., Koutsotolis, K., Nikolaou, E., Kalogiannis, D. &
Petrakos, G. (1998) Seasonal variations of gastrointestinal nematodes of
sheep in the region of Joannina, Greece. International Journal for
Parasitology, 28: 1287-1292.
33. Tompkins, D.M., Dobson, A.P., Arneberg, P., Begon, M.E., Cattadori,
I.M., Greenman, J.V., Heesterbeek, H., Hudson, P.J., Newborn, B.,
Pugliese, A., Rizzoli, A.P., Rosa, R., Rosso, F. & Wilson, K. (2001).
Parasites and host population dynamics. Oxford University Press, Oxford.
34. Tompkins, D.M. & Hudson, P.J. (1999) Regulation of nematode fecundity
in the ring-necked pheasant (Phasianus colchicus): not just density
dependence. Parasitology, 118: 417-423.
35. Vicente, J., Fierro, Y. & Gortázar, C. (2005) Seasonal dynamics of the
fecal excretion of Elaphostrongylus cervi (Nematoda, Metastrongyloidea)
first-stage larvae in Iberian red deer (Cervus elaphus hispanicus) from
southern Spain. Parasitology Researchearch, 95(1): 60-4.
36. Villanúa, D., Acevedo, P., Höfle, U., Rodríguez, O. & Gortázar, C. (2006).
Changes in transmission stage excretion after pheasant release. Journal of
Helminthology, 80: 1-7.
37. Vorísek, P., Votýpka, J., Zvára, K. & Svobodova, M. (1998).
Heteroxenous coccidia increase the predation risk of parasited rodents.
Parasitology, 117: 521-524
38. Welch, D.A., Pybus, M.J., Samuel, W.M. & Wilke, C.J., (1991) Reliability
of fecal examination for detecting infections of meningeal worm in elk.
Wildlife Society Bulleting, 19: 326–331.
39. Yu, J.M., de Vlas, S.J., Yuan, H.C. & Gryseels, B. (1998) Variations in
fecal Schistosoma japonicum eggs counts. The American journal of tropical
medicine and hygiene, 59(3): 370-375.

76
40. Zuk, M. (1992) The role of parasites in sexual selection: current evidence
and future directions. Advances In Study Behaviour., 21: 39-68.

77
VI. CAPITULO 3
Efectividad de los tratamientos antiparasitarios actuales para
prevenir la introducción de nematodos en el campo.
How effective is pre-release nematode control in farm reared red-legged
partridges (Alectoris rufa)?. Villanúa, D., Pérez-Rodríguez, L, Rodríguez,
O., Viñuela, J. & Gortázar C (2007). Journal of Helmithology (in press).

78
VI. CAPITULO 3
Efectividad de los tratamientos antiparasitarios actuales para
prevenir la introducción de nematodos en el campo.
How effective is pre-release nematode control in farm reared red-legged
partridges (Alectoris rufa)?. Villanúa, D., Pérez-Rodríguez, L, Rodríguez,
O., Viñuela, J. & Gortázar C (2007). Journal of Helmithology (in press).
RESUMEN
La cría de la perdiz roja en granja lleva asociado a un alto grado de
parasitación capaz de disminuir la productividad de la propia granja y la
supervivencia de las aves una vez liberadas, además de suponer un riesgo
para las poblaciones naturales. En el presente trabajo se ha evaluado la
efectividad de albendazol (V.O. 20 mg/kg) en perdices de granja infectadas de
manera natural con Aonchoteca caudinflata y Heterakis gallinarum.
Las aves tratadas mostraron una clara mejora de su condición física y se
redujo la excreción de propágulos parasitarios y el porcentaje de hembras
grávidas de A. caudinflata, pero no se consiguieron eliminar los parásitos
adultos. La efectividad encontrada fue de un 36,8 % en A. caudinflata y de un
17,1 % en H. gallinarum.
Estos resultados indican que el tratamiento antihelmíntico usado
habitualmente en las granjas de perdiz de nuestro país no es lo suficientemente
efectivo como para evitar la introducción de nuevos parásitos en el campo a
través de las sueltas de perdiz roja.
ABSTRACT
Game bird farming is associated with higher parasitation levels that diminish
farm productivity, reduce survival after releasing, and may pose a health risk for
natural populations. We evaluated the efficacy of albendazole (orally, 20 mg/kg)
in farmed red-legged partridges naturally infected with Aonchoteca caudinflata
and Heterakis gallinarum.

79
Treated birds improved their body condition, reduced nematode-egg
deposition and the proportion of gravid A. caudinflata females, but not the worm
burden. We found a 36.8% and a 17.1% of effectiveness on A. caudinflata and
H. gallinarum, respectively.
These results indicate that the anthelmintic treatment used normally in
Spanish partridge farms is not effective enough to avoid the introduction of
parasites into the field after release.
KEYWORDS: Albenzadole, Game birds, Helminths, Treatment.
INTRODUCTION
The red-legged partridge Alectoris rufa is one of the most important game
birds in Spain with more than 5 million individuals hunted yearly (Millán et al.,
2004). Natural populations have declined considerably during the past decade,
leading to an important increase in partridge releases to supplement stocks for
shooting (Millán et al., 2003).
The release of farmed animals can involve a risk for wild populations due to
the possible introduction of new parasites into the field (Tompkins et al. 2002;
Millán et al., 2004).
This study evaluates the efficacy of albendazole, one of the most widely
used anthelmintics in Spanish partridge farms (APROCA, 2004), to prevent the
introduction into the field of two nematodes, Aonchoteca caudinflata (Holger-
Madsen, 1954) and Heterakis gallinarum (Scrank, 1788), parasites that are
exclusively found in farm-reared partridges, but not in wild ones (Millán et al.,
2004).
MATERIAL AND METHODS
The study was carried out on 16 month old red-legged partridges during a
natural capillarid outbreak in a partridge farm in Ciudad Real, central Spain. In
September 2004, eight randomly selected birds coming from the infected
outdoor pen were isolated in individual elevated wire-cages.

80
Faecal samples were obtained on day 0 at dusk (time of the day of maximal
nematode egg excretion, Villanúa et al. in press) for a quantitative coprological
analysis in MacMaster chambers as described by Melhorn et al. (1992), which
confirmed the infection of all birds with capillarid nematodes (later identified as
Aonchoteca caudinflata).
Five randomly selected individuals were treated orally with 20 mg/kg of
albendazole (Albendavet®, Divasa Farmavic, S.A.) on day 1 and day 15,
according to the manufacturer´s instructions. The same volume of water was
given as a control to the other three infected birds. Fifteen days after the second
administration all birds were humanely sacrified and necropsied.
The necropsy included a second coprological analysis and the examination
of the digestive tract searching for parasites using a stereomicroscope.
The efficacy (E) of albendazole treatment on worm burden was calculated
as follows after Reina et al.(2000):
Geometric mean of control animals-Geometric mean of treated animalsE= x 100Geometric mean of control animals
The nematode sex-ratio (SR) and fertility (F) were calculated as follows after
Kassai (1998):
Nº female nematodesSR=Nº total nematodes
Nº gravid females F=Nº total females
Body condition of partridges was estimated using three different methods:
an arbitrary score of the degree of fat deposition (1 to 5), the pectoral angle as a
measure of the width of pectoral muscles after Millán et al. (2003), and the
residuals of the regression of body weight on tarsus length after Andersson
(1992).
The effect of Albendazole treatment on propagule excretion, number of adult
worms and partridge body condition was analysed by means of non-parametric
statistics (Wilcoxon matched pairs tests and Mann-Whitney U tests), as
recommended by Kassai (1998) for studies with natural infections. Differences
in nematode sex ratio and fertility were analysed using Chi2 tests.
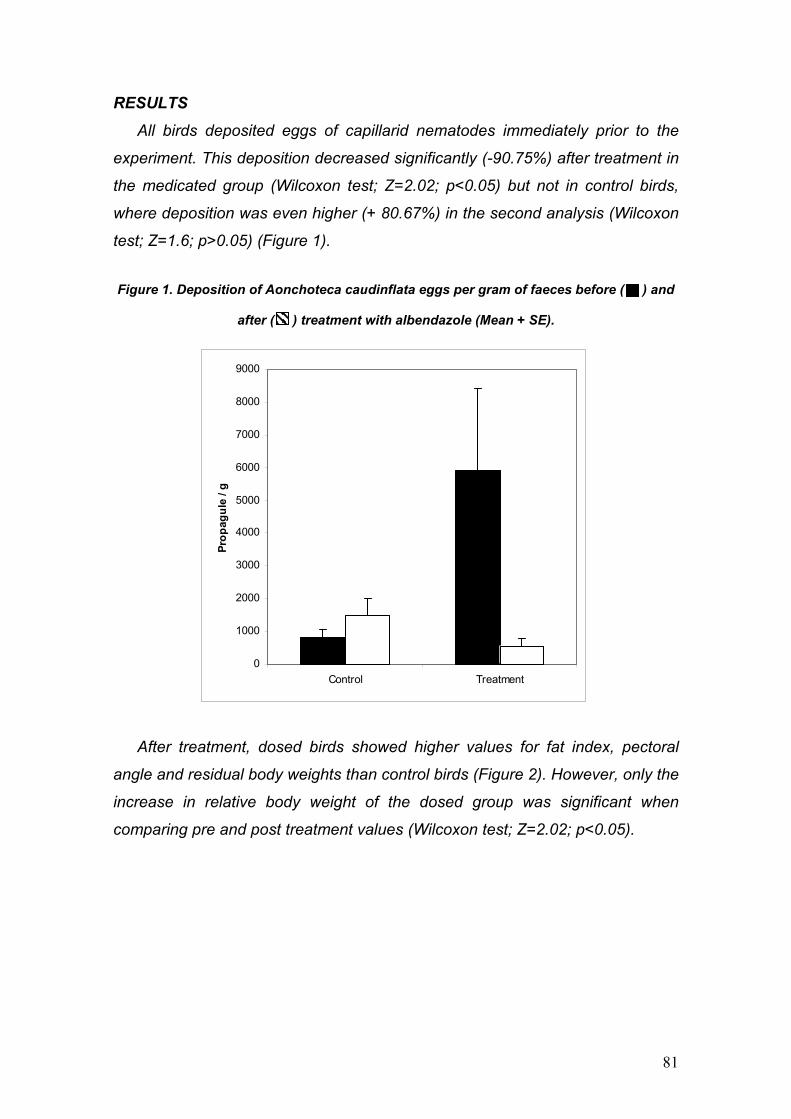
81
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Control Treatment
Prop
agul
e / g
RESULTS
All birds deposited eggs of capillarid nematodes immediately prior to the
experiment. This deposition decreased significantly (-90.75%) after treatment in
the medicated group (Wilcoxon test; Z=2.02; p<0.05) but not in control birds,
where deposition was even higher (+ 80.67%) in the second analysis (Wilcoxon
test; Z=1.6; p>0.05) (Figure 1).
Figure 1. Deposition of Aonchoteca caudinflata eggs per gram of faeces before ( ) and
after ( ) treatment with albendazole (Mean + SE).
After treatment, dosed birds showed higher values for fat index, pectoral
angle and residual body weights than control birds (Figure 2). However, only the
increase in relative body weight of the dosed group was significant when
comparing pre and post treatment values (Wilcoxon test; Z=2.02; p<0.05).
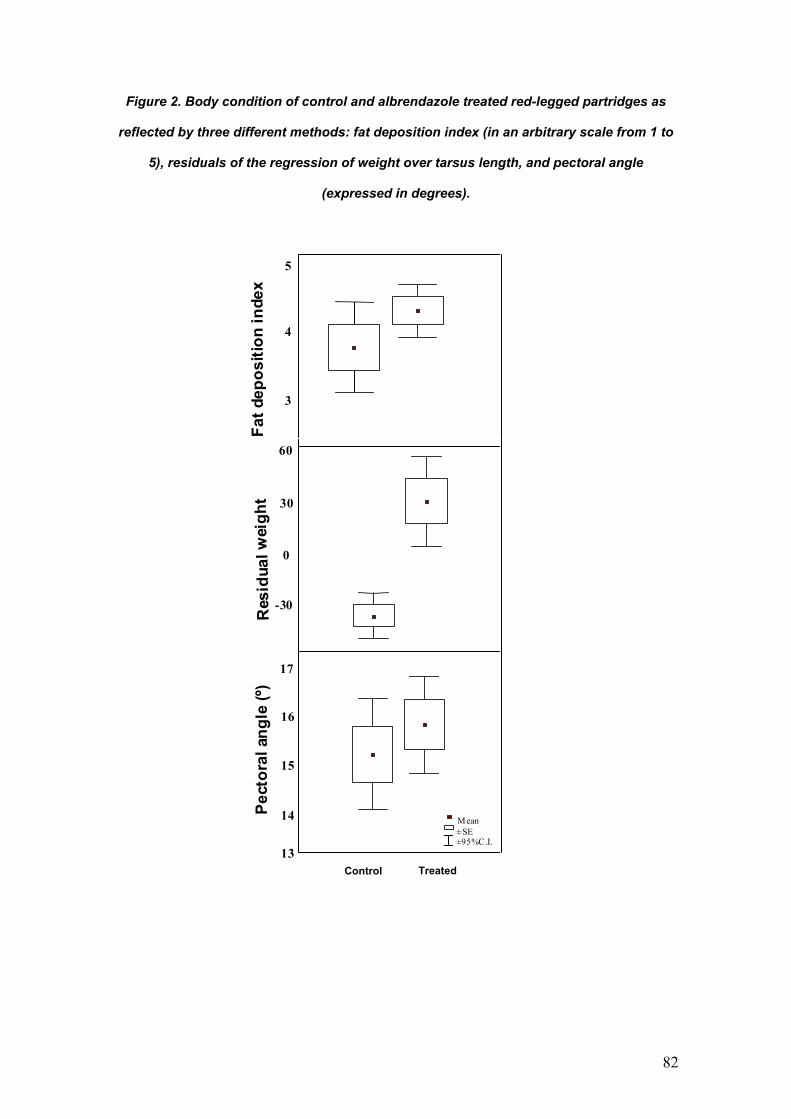
82
Figure 2. Body condition of control and albrendazole treated red-legged partridges as
reflected by three different methods: fat deposition index (in an arbitrary scale from 1 to
5), residuals of the regression of weight over tarsus length, and pectoral angle
(expressed in degrees).
Control Treated
Fat d
epos
ition
inde
xPe
ctor
al a
ngle
(º)
Res
idua
l wei
ght
Mean ±SE ±95%C.I.
5
4
3
-30
0
30
60
13
14
15
16
17
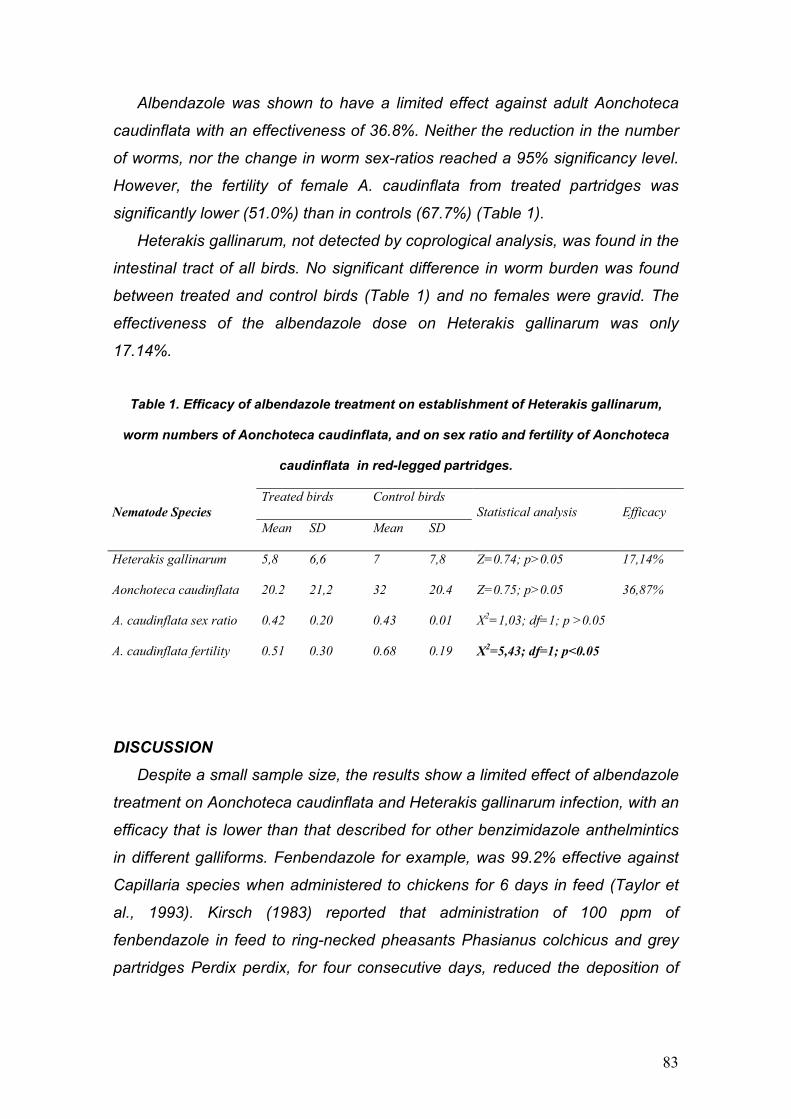
83
Albendazole was shown to have a limited effect against adult Aonchoteca
caudinflata with an effectiveness of 36.8%. Neither the reduction in the number
of worms, nor the change in worm sex-ratios reached a 95% significancy level.
However, the fertility of female A. caudinflata from treated partridges was
significantly lower (51.0%) than in controls (67.7%) (Table 1).
Heterakis gallinarum, not detected by coprological analysis, was found in the
intestinal tract of all birds. No significant difference in worm burden was found
between treated and control birds (Table 1) and no females were gravid. The
effectiveness of the albendazole dose on Heterakis gallinarum was only
17.14%.
Table 1. Efficacy of albendazole treatment on establishment of Heterakis gallinarum,
worm numbers of Aonchoteca caudinflata, and on sex ratio and fertility of Aonchoteca
caudinflata in red-legged partridges.
Treated birds Control birds Nematode Species
Mean SD Mean SD Statistical analysis Efficacy
Heterakis gallinarum 5,8 6,6 7 7,8 Z=0.74; p>0.05 17,14%
Aonchoteca caudinflata 20.2 21,2 32 20.4 Z=0.75; p>0.05 36,87%
A. caudinflata sex ratio 0.42 0.20 0.43 0.01 X2=1,03; df=1; p >0.05
A. caudinflata fertility 0.51 0.30 0.68 0.19 X2=5,43; df=1; p<0.05
DISCUSSION
Despite a small sample size, the results show a limited effect of albendazole
treatment on Aonchoteca caudinflata and Heterakis gallinarum infection, with an
efficacy that is lower than that described for other benzimidazole anthelmintics
in different galliforms. Fenbendazole for example, was 99.2% effective against
Capillaria species when administered to chickens for 6 days in feed (Taylor et
al., 1993). Kirsch (1983) reported that administration of 100 ppm of
fenbendazole in feed to ring-necked pheasants Phasianus colchicus and grey
partridges Perdix perdix, for four consecutive days, reduced the deposition of

84
Heterakis gallinarum and Capillaria obstignata eggs and the number of adult
worms by more than 90%.
In the present study, a similar reduction in Capillaria egg counts was
obtained, but not in worm burden.This suggests that albendazole has some
limitant effect on the reproduction of nematodes, but it does not eliminate the
infection. This may be due to an insuffcient dosage. In the present study the
dose administred was that recommended for chickens by the manufacturer, so
future studies should confirm whether a dose increase would improve the
efficacy of the treatment.
In conclusion, the study shows that the treatment currently used by game
bird farmers in Spain may help to improve body condition of farm-reared
partridges, but is not enough to avoid the introduction of parasites into the field
after release. Alternative drugs, higher doses, or other administration methods
must be evaluated.
This study also highlights that coprological analyses alone are not enough to
ensure the absence of a given nematode species in farm-reared birds.
Sanitary screenings of farm-reared birds should preferably include some
complete necropsies, for example of casualties, rather than rely exclusively on
non-invasive sampling.
ACKNOWLEDGEMENTS
This contributes to the agreement IREC - Principado de Asturias. We
acknowledge Galiana facilities (FGUCLM) and thank E. Pérez and F. Talavera
for their help. L. Pérez-Rodríguez has a FPU grant and O. Rodríguez a Torres
Quevedo contract partly supported by MEC. Two referees improved the ms.
REFERENCES
1. Andersson, S.(1992) Female preference for long tails in lekking Jackson´s
widowbirds: experimental evidence. Animal Behaviour, 43: 379-388.
2. APROCA (2004) Jornadas de sobre la cría de especies cinegéticas. Madrid,
Noviembre 2004.
3. Kassai, T. (1998) Helmintologia Veterinaria. Ed. Acribia. Zaragoza, España
259 pp.

85
4. Kirsch, R. (1983) Treatment of Nematodiasis in Poultry and Game birds
with Febendazole. Avian Diseases, 28(2): 311-318.
5. Melhorn, H., Düwell, D., & Raether, W. (1992) Atlas de Parasitología
Veterinaria. GRASS ediciones, Spain.
6. Millán, J., Gortázar, C., Buenestado, F.J., Rodriguez, P., Tortosa, F.S. &
Villafuerte, R. (2003) Effects of a fiber-rich diet on physiology and survival
of farm-reared red-legged partridges (Alectoris rufa). Journal of Comparative
Biochemistry and Physiology, 134: 85-91.
7. Millán, J., Gortázar, C. & Villafuerte, R. (2004). A comparison of the
helminth faunas of wild and farm-reared red-legged partridges. Journal of
Wildlife Management ,68(3): 701-707.
8. Reina, D., Anderson, L., Habela, M., Weatherley, A. J., Navarrete, I.
(2000) Efficacy of doramectin against naturally acquired nematode infection
in Iberian swine. Veterinary Parasitology, 89: 139-147.
9. Taylor, S.M., Kenny, J., Houston, A. & Hewitt, S.A. (1993) Efficacy,
pharmacokinetics and effects on eggs-laying and hatchability of two dose
rates of in-feed febendazole for the treatment of Capillaria species infection
in chickens. The Veterinary Record, 20: 519-521.
10. Tompkins, D.M., Parish, D.M.B., and Hudson, P.J. (2002) Parasite-
mediated competition among red-legged partridges and other lowland
gamebirds. Journal of Wildlife Management, 66: 445-450.
11. Villanúa, D., Pérez-Rodríguez, L., Gortázar, C., Höfle, U. & Viñuela, J.
(2006). Avoiding bias in parasite excretion estimates: the effect of sampling
time and type of faeces. Parasitology, 133: 251-259

86
VII. CAPITULO 4
Posible transmisión de nematodos propios de perdices de granja a
especies amenazadas.
First occurence of Eucoleus contortus in a little bustard Tetrax tetrax. A
negative effect of red-legged partridge Alectoris rufa releases on steppe bird
conservation?. Villanúa, D., Casas, F., Viñuela, J., Gortázar, C., García de
la Morena, E.L. & Morales, M.B. (2007). Ibis, 149: 405-406

87
VII. CAPITULO 4
Posible transmisión de nematodos propios de perdices de granja a
especies amenazadas.
First occurence of Eucoleus contortus in a little bustard Tetrax tetrax. A
negative effect of red-legged partridge Alectoris rufa releases on steppe bird
conservation?. Villanúa, D., Casas, F., Viñuela, J., Gortázar, C., García de
la Morena, E.L. & Morales, M.B. (2007). Ibis, 149: 405-406
RESUMEN
Cinco adultos de Eucoleus contortus fueron encontrados en el buche de un
macho de sisón, Tetrax tetrax, procedente de Ciudad Real (España, UTM
0421739 4311458).
En la zona en la que se recogió el animal se llevan a cabo munerosas
sueltas cinegéticas de perdiz roja (Alectoris rufa) criadas en granja.
El nematodo Eucoleus contortus nunca había sido descrito en sisones ni
avutardas (Otididae), pero es un parásito común en perdices de granja. Por
ello, este hallazgo pone de manifiesto la importancia de realizar estudios
parasicológicos comparativos en aquellas zonas sometidas a repoblaciones
para la caza.
ABSTRACT
Five adult forms of Eucoleus contortus were found in the crop of a male little
bustard, Tetrax tetrax, from Ciudad Real (Spain, UTM 0421739 4311458).
This area is subject of many releases of farm bred red-legged partridges
(Alectoris rufa) for hunting.
Eucoleus contortus had never been reported in bustards (Otididae), but is a
common parasite of farmed gamebirds. Hence, this finding highlights the
interest of compared parasitological studies in areas managed for hunting.
KEYWORDS: Eucoleus contortus; Game management; Tetrax tetrax; Partridge
releases; Bird conservation

88
Little bustard (Tetrax tetrax), populations are rapidly declining in most
European countries (BirdLife International 2004). The cereal steppes of central
Spain are the most important wintering quarters for the little bustard, with more
than 90% of the west European population (De Juana and Martínez 2001,
García de la Morena et al. 2004). These steppe habitats are also important
partridge hunting areas in Spain.
During the last decades, natural populations of the red-legged partridge
(Alectoris rufa), have declined considerably in their historical range and more
than 4 million farm-reared partridges are released yearly in autumn to
compensate this decline. This has already raisen concerns regarding the
possible introduction of new parasites into natural populations (Millán et al.
2004a, 2004b).
In winter 2005, 14 little bustards (5 males and 9 females) were captured with
cannon nets in Miguelturra (Ciudad Real, Central Spain, UTM 0421739
4311458) for radio-tagging. The capture area was close (less than 5 km) to an
important hunting estate where partridge releases are common (about 3000
partridges released yearly).
One adult male died due to traumatism during the capture and was
necropsied. In the parasitological examination, adult forms of a capillarid
nematode were found in the crop. After further examination under a
stereomicroscope, these were found to represent adults of Eucoleus contortus
Creplin, 1839 (3 males, 2 gravid females) according to Anderson (2000).
The body condition of the necropsied bird, estimated as the ratio of body
weight on cube tarsus length (0,0018g/mm3), was lower than the ones found in
other 15 adult males captured the same season (0,0025 ± 0,0004; mean ±
standard desviation ).
No E. contortus have been found in any of 4 little bustards, nor in any of 17
great bustards (Otis tarda), both species sampled in central Spain and
necropsied in our laboratory.
In contrast, E. contortus is present in 7.7% of the red-legged partridges from
a neighboring hunting estate where releases of farm-bred gamebirds take place
(the authors, in prep.).
Like other monoxenous nematodes, Eucoleus contortus is almost
exclusively found in farmed gamebirds (Millán et al. 2004b) and had, to the best

89
of our knowledge, never been found in members of the Otididae family (Cordero
del Campillo et al. 1994).
E. contortus can affect the host´s body condition (Bosch et al. 2000), and
make their hosts more vulnerable to predation (Millán et al. 2002).
Our results suggest that the release of farm reared gamebirds can
eventually introduce new pathogens to wild populations of different species,
many of which are of conservation concern, as it is the case in the little bustard .
As hypothesized by Tompkins et al., (2001), if these parasites are able to find a
new host, they can become an additional problem for its conservation.
ACKNOWLEDGEMENTS
This is a contribution to the agreement between IREC and Principado de
Asturias and to the CICYT Project CGL2004-06147-C02/BOS. We wish to thank
I. Hervás, R. Agudo, R. Mateo, M. Martínez and S. Luna for their assistance in
the field. F. Casas has a JCCM grant and E. L. García de la Morena was
funded by the Ministry of Education’s FPU Program. This experiment was made
following European, National and University of Castilla – La Mancha Ethics
Committee regulations.
REFERENCES
1. Anderson, R.C. (2000). Nematode parasites of vertebrales: their
development and transmission. CABI Publishing, Wallingford.
2. Birdlife International (2004). Birds in Europe II: Population Estimates,
Trends and Conservation Status. BirdLife International, Cambridge.
3. Bosch, M., Torres, J. & Figuerola, J. (2000). A helminth community in
breeding Yellow-legged Gulls (Larus cachinnans): pattern of association and
its effect on host fitness. Canadian Journal of Zoology, 78: 777-786.
4. Cordero del Campillo, M., Castañón, L. & Reguera, A. (1994). Índice-
catálogo de zooparásitos ibéricos. Universidad de León, León.
5. De Juana, E. & Martínez, C. (2001). European Union Species Action Plan
For Little Bustard (Tetrax tetrax). In Schäffer, N & Gallo-Orsi, U. (eds.),
European Union Action Plans for eight priority birds species. Office for
Official Publications of the European Communities, Luxembourg (pp. 1-17).

90
6. García de la Morena, E.L., De Juana, E., Martínez, C., Morales, M.B. &
Suárez, F. (2004). Sisón Común, Tetrax tetrax. In Madroño, A, González, C.
& Atienza, J.C. (eds.), Libro Rojo de las Aves de España. Dirección General
para la Biodiversidad-SEO/Birdlife. Madrid (pp. 202-207).
7. Millán, J., Gortázar, C., Tizzani, P. & Buenestado, F.J. (2002). Do
helmints increase the vulnerability of released pheasants to fox predation?.
Journal of Helminthology, 76: 225-229.
8. Millán, J., Gortázar, C., Martín-Mateo, M.P. & Villafuerte, R. (2004a).
Comparative survey of the ectoparasite fauna of wild and farm-reared red-
legged partridges (Alectoris rufa), with an ecological study in wild
populations. Parasitology Research , 93(1): 605-611.
9. Millán, J., Gortázar, C. & Villafuerte, R. (2004b). A comparison of the
helminth faunas of wild and farm-reared red-legged partridges. Journal of
Wildlife Management 68(3): 701-707.
10. Tompkins, D.M., Greenman, J.V., Hudson, P.J. (2001). Differential impact
of shared nematode parasite on two gamebird hosts: implications for
apparent competition. Parasitology, 122: 187-193.

91
VIII. CAPITULO 5
Factores limitantes de la abundancia estival de perdiz roja en Aragón.
Posibles alternativas a las repoblaciones con aves de granja.
Factors affecting summer densities of the red-legged partridge (Alectoris
rufa). Bird Study. Villanúa, D., Acevedo, P., Escudero, M.A., Marco, J. and
Gortázar, C. (En evaluación).

92
VIII. CAPITULO 5
Factores limitantes de la abundancia estival de perdiz roja en Aragón.
Posibles alternativas a las repoblaciones con aves de granja.
Factors affecting summer densities of the red-legged partridge (Alectoris
rufa). Ibis. Villanúa, D., Acevedo, P., Escudero, M.A., Marco, J. and
Gortázar, C. (En evaluación).
RESUMEN
La perdiz roja (Alectoris rufa) es el ave de caza más importante en la
Península Ibérica. A lo largo de los últimos años, el colectivo de cazadores ha
percibido un descenso de sus poblaciones en la práctica totalidad de su área
de distribución. Diversos factores, tales como la depredación, la presión
cinegética, la meteorología o la alteración del hábitat han sido considerados
habitualemente como posibles causas de este descenso.
En el presente trabajo se ha estimado la densidad de perdices tras la
reproducción (siguiendo el método de Kelker) en 36 acotados (con una media
de 5 transectos por acotado repetidos anualmente) desde 1998 hasta 2004. Se
ha testado el efecto de factores meteorológicos, de hábitat, de depredación
(abundancia de carnívoros) y presión cinegética sobre las variaciones en la
abundancia de perdiz roja.
Los resultados obtenidos muestran que los factores meteorológicos son los
más limitantes para esta especie. En concreto, la temperatura máxima y el
número de días de lluvia fueron los parámetros más significativamente
relacionados con la abundancia de perdices. Además de estos dos factores,
varias características del hábitat mostraron una influencia significativa sobre las
abundancias de perdiz. Así, las mayores abundancias se encontraron en los
acotados con un alto grado de mosaico de cultivos, en parcelas irregulares y
con alta cantidad de linderos. No se detectó relación entre la presión cinegética
o la depredación y la abundancia de perdices en esa época del año.
ABSTRACT
The red-legged partridge (Alectoris rufa) is the most important game bird in
the Iberian peninsula. During the last years, hunters perceive the decline of

93
partridge populations in practically all its distribution area. Factors such as
predation, hunting pressure, meteorology, and habitat structure are known to
affect game bird populations.
We estimated postreproductive partridge densities following Kelker’s method
in 36 hunting estates (an average of 5 transects per hunting estate). Transects
were carried out yearly from 1998 to 2004, and the influence of different
meteorological factors, habitat characteristics, predation (carnivore abundance)
and hunting pressure was analysed.
Our results suggest that meteorological conditions were the most important
factor limiting partridge abundance, but also in determining inter-annual
variations. Partridge densities were significantly limited by the maximum
temperature and by the number of days of hail. Some habitat characteristics do
benefit the partridge densities. The highest ones were found in areas with high
patch density of irregular mosaic cultures. No apparent relationship between
hunting pressure and predation and partridge abundance was observed.
KEY WORDS: Agricultural landscape, Density, Game birds, Fox predation,
Hunting, Meteorology.
INTRODUCTION
Red-legged partridge (Alectoris rufa) hunting is a important economic and
social activity in the rural areas of the Iberian peninsula (Delibes 1988, Lucio
1998, Bernabeu 2000). In addition, this species is, along with the wild rabbit
(Oryctolagus cuniculus), one of the most important preys for endangered
Mediterranean predators such as the Iberian imperial eagle (Aquila adalberti)
(Delibes & Hiraldo 1981, Calderón 1983).
During the last years, populations of this game bird have declined seriously
in almost all its distribution area (Cramp & Simons 1980, Potts 1980, Aebischer
& Potts 1994, Baratti et al 2005), including the Iberian peninsula (Nadal et al.
1996, Borralho et al. 1998, Lucio 1998, Carvalho et al. 1998, Duarte 1998,
Blanco Aguiar et al 2003, SEO/BirdLife 2004). The causes of this decline are
not clear, but agriculture intensification (Potts 1980; Rands, 1987; Pepin y
Blayac, 1990; Crick et al. 1994, Green 1995, Nadal et al., 1996; Aebischer y
Kavanagh, 1997; Lucio, 1998; Gortázar et al., 2002), hunting pressure (Potts,

94
1986; Pepin y Blavac, 1990; Lucio y Purroy, 1992; Borralho et al 1997),
predation (Ricci et al. 1990, Rands 1988, Yanes et al. 1998) or meteorological
factors (Lucio 1990) are some of the most frequently suspected causes of this
decline.
There are many papers on the influence of habitat, climate, and predation on
partridge densities in the Iberian peninsula. However, some of theme are short
term surveys (Lucio 1990, Nadal et al. 1996, Borralho et al. 1998, Fortuna
2002), and others were focused on small study areas or on specific habitats
(Lucio 1990; Nadal et al. 1996, Borralho et al. 1998, Fortuna 2002). Gortázar et
al. (2002) gave the first large-scale data on summer red-legged partridge
abundance, based on the data of the first year (1998) of regular partridge
census in Aragón, north-eastern Spain. These authors suggested that the lower
partridge densities found in Aragón, as compared to those in Central and
Southern Spain, can be explained by climatic and habitat-related factors, but
also by management differences.
The aim of our present study is to evaluate how habitat, hunting pressure,
fox predation and meteorology affect summer red-legged partridge densities
during a 7-year series (from 1998 to 2004) in a wide range of habitats in north-
eastern Spain.
MATERIAL AND METHODS
Study area
The study area (40º01'13''N to 42º23'27''N in latitude and from
03º53'22''W to 06º13'14''W in longitude) is located in the region of Aragón, north
east of the Iberian peninsula (Figure 1). Aragón is an autonomous region of
47,669 km2. It includes the central part of the Spanish Pyrenean mountains
(more than 3000 m a.s.l.), the southeastern part of the Iberian mountain chain
and the Central Ebro Valley (less than 300 m a.s.l.). This gradient explains the
presence of both big game species (includes Rupicapra pyrenaica, Capra
pyrenaica, Sus scrofa, Cervus elaphus and Capreolus capreolus), and small
game species (including Alectoris rufa, Lepus europaeus, L. granatensis and
Oryctolagus cuniculus).
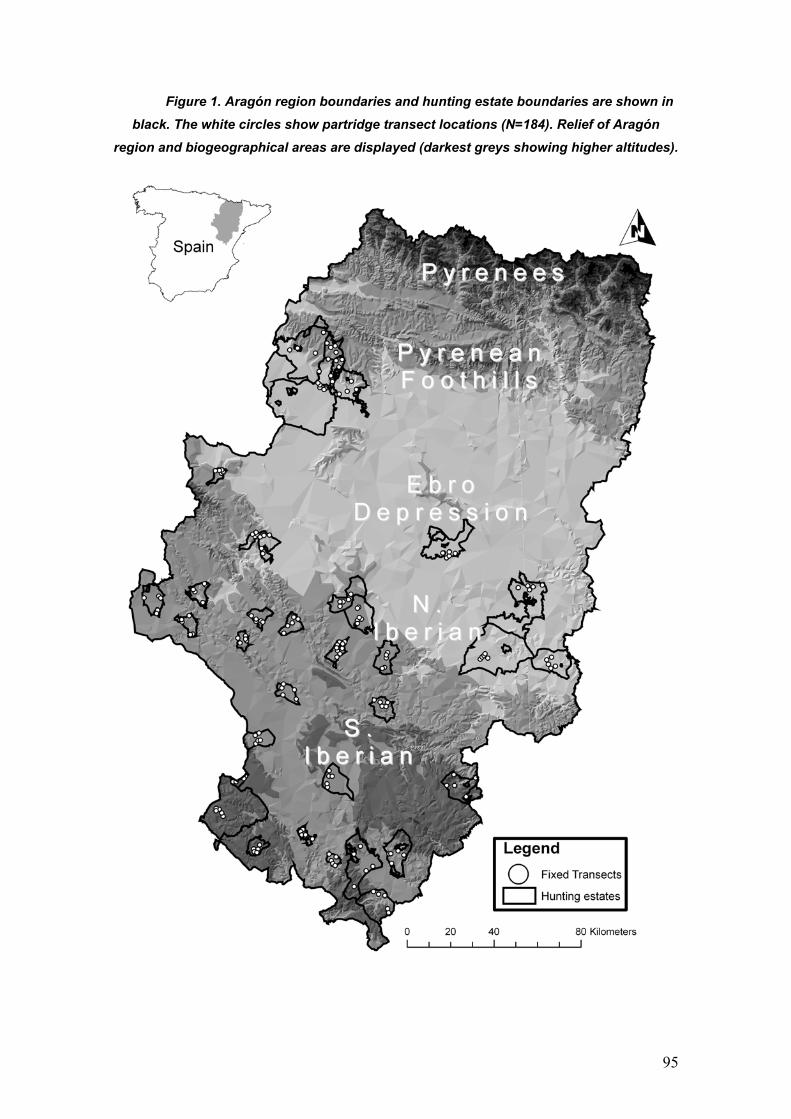
95
Figure 1. Aragón region boundaries and hunting estate boundaries are shown in black. The white circles show partridge transect locations (N=184). Relief of Aragón
region and biogeographical areas are displayed (darkest greys showing higher altitudes).

96
Aragón has a diverse raptor and carnivore community, but the red fox is
clearly the most important predator for small game species (e.g. Villafuerte et al.
1996, Gortázar 1997, Gortázar et al. 2000).
The climate is Mediterranean, more continental as it gets away from the
sea, with an Atlantic influence in the western Pyrenees. Wood and scrublands
cover 13,518 km2. Fifty percent of the 1.2 million inhabitants concentrate in the
largest city, Zaragoza. Rural abandonment has been especially severe during
the industrial development of the sixties, allowing the natural vegetation to
recover (Bielza de Ory 1993).
Within this area, 5 biogeographical areas can be distinguished (Gortázar et
al. 2002), see Figure 1: (1) the Pyrenees (yearly mean temperature (T) lower
than 12ºC, annual rainfall (R) 800-1700 mm), (2) the Pyrenean foothills (T: 12-
14 ºC, R: 500-800 mm), (3) the Ebro depression (T: 13-15 ºC, R: 250-500 mm),
(4) the northern Iberian mountains (T: 10-14 ºC, R: 400-700 mm) and (5) the
southern Iberian mountains (T: 8-11 ºC, R: 400-800 mm). The last 4 areas are
occupied by the red-legged partridge and were included in our research.
Hunting in Aragón has not the commercial character typical from the south
of Spain, and the main part of the 2,091 hunting estates present in the region
are managed directly by the local associations. This social character makes
game bird releases rare. For this reason, hunting estates included in our
research have only natural populations of red-legged partridges, with no
restocking with captive-bred game birds in the last ten years.
Partridge densities
Partridge densities (individuals/100 ha) were estimated in 36 different
hunting estates throughout the study area (Figure 1) using the censuses
included in the wildlife monitoring scheme (Aragón Government). Censuses
were carried out in August and September because the aim of this monitoring
plan is to know the hunting species abundance just before the hunting season,
in October. In each hunting estate, 5 fixed transects were placed selecting open
areas of high visibility. Transects were georeferenced (by means of GPS,
Garmin eTrex Vista™) on the Forest Map of Aragón (vector map produced by
Aragón Government, scaled 1: 50,000). The censuses were repeated during 7
consecutive years (from 1998 to 2004) for a total of 1,112 transects (552

97
km/year). As described Gortázar et al. (2002), each fixed transect was 3,000 m
long and 100 m wide (30 ha), and was censused by 3 observers, thus the
transect width for each observer was 34 m. The short distance between
observers, and the selection of rather open habitats, should reduce the bias due
to visibility (see Bibby et al. 1992), and made us confident of having little
underestimation. We assumed that all birds within the transect width were
detected. Censuses began shortly after dawn until 3 to 5 hours later and were
done by foot. In order to reduce variability, only days without wind, rain or other
unfavourable meteorological circumstances were selected.
Partridge density for each transect was estimated with Kelker´s method
(see Burnham et al. 1980), using the formula:
6· ·10nD
L W −=
where D is the partridge density per 100 ha, n is the number of partridges
seen inside the 100 m counting strip, and 6· ·10L W − is the surface of the counting
strip in 100 ha units (i.e. 3000·100·10-6= 0.3). Annual partridge density for each
hunting estate was calculated as the average of partridge densities obtained in the
fixed transects (generally 5 per hunting estate).
Weather conditions
Meteorological data were taken from 24 official meteorological stations of
the National Institute of Meteorology located across the study area from 1998 to
2002. To select the most representative parameters, a preliminary correlation
analysis was made with the different meteorological data. After this first
selection, we used the following indicators:
• Rainfall: winter rainfall accumulation, number of days of hail and number of
days of storm from April to September, number of days with precipitation
higher than 300 mm in the same period.
• Temperature: average of the maximum temperatures per month, average of
the minimum temperatures per month, number of days with maximum
temperature higher than 25 ºC and number of days with minimum
temperature lower than –5 ºC.
These factors characterized meteorologically each hunting estate.

98
Vegetation and orography characteristics
To characterize the vegetation and orography of each hunting estate
(sampling area) we used Geographic Information Systems.
We defined 10 different classes of vegetation (patch types) using the Forest
Map of Aragón (scaled 1: 50,000): oak and pine woodlands, oak savannah
habitats (“dehesas”), short scrub, tall scrub, cultures mosaic, sparse pines,
cereal cultures, woody cultures, pastures, and others (vegetation classes that
rarely occurred in the study area).
We used the following indices:
• Patch Density (PD): the total number of patches per unit area (number of
patches/m2). We considered the number of patches of each vegetation class
(vegetation-class level) and/or the total number of patches per sampling unit
(landscape level) to calculate the PD index.
• Average Size of Patches (ASP) of each vegetation class (m2).
• Average Edge length of Patches (AEP) of each vegetation class (m).
• Average Edge length per unit Area (AEA) of each vegetation class (m-1).
The orography characteristics were obtained from a digital elevation model
with a spatial resolution (pixel width) of 100 m (SRTM, European Environment
Agency 2000). The average altitude and slope of each sampling unit were
considered to evaluate the influence of the orography on red-legged partridge
abundance.
Fox population trends
The red fox (Vulpes vulpes) is considered one of the most important
partridge predators (e.g. Tapper et al. 1982, Calderón 1983). Hence, the fox
population trend for each estate was included in the analysis. Red fox
abundance was estimated using the spotlight counts also included in the wildlife
monitoring scheme. These counts were carried out yearly in winter on fixed
routes that cross open habitats far from forests or human-inhabited places. The
same two rangers carried out all surveys of a given locality using a 4-wheel
drive vehicle with a handheld 100-watt spotlight, which light was swept round in
a semi-circle (Barnes & Tapper 1984) (see Williams et al 2007 for details).

99
From 1998 to 2004, each transect was censused an average of 8 times.
Every fixed route was about 30 km long, and the average speed was 20 km/h.
Data were converted into abundance per 100 km (100*KA) to get a relative
abundance index. These values were also used to calculate a fox trend index
(percentage of a common base value). This base value was the average of the
different yearly abundances (see Crawford 1991 for a discussion on the
calculation of indexes in monitoring schemes). Thus, the population trend
indexes do reflect the % abundance in relation to the average for the whole 7-
year period, and are independent from the detectability.
Hunting pressure
The low human population density typical from the rural areas in Aragón and
the social characteristics of hunting in this region, originates a lower hunting
pressure as compared to other regions (Gortázar et al. 2002). In the present
study, we use the number of hunters per hectar as an estimator of the hunting
pressure in each hunting state.
Statistical analysis
Linear regression analyses were used to calculate partridge population
trends. Moreover, we designed a 2 step procedure to assess the effect of the
environmental variables on red-legged partridge density.
First, the bivariate association between the hypothesized factors and
partridge summer abundances were analysed by means of Spearman’s
correlations. The variables which captured the effect of any set of highly
correlated variables on the outcome were selected for inclusion in the models
(to avoid multicollinearity). For categorical variables, Kruskall-Wallis analysis
was used for simple assessment of associations.
As second step, we conducted a generalized linear mixed model (GLIMMIX)
and used partridge densities per 100 hectar as response variable (n=213). We
considered a Poisson error distribution and a logarithmic link function (Wilson &
Grenfell 1997). The models were reduced to their simplest form by eliminating
in a backward stepwise procedure any explanatory variables that failed to
explain significant variation in the response (Crawley 1993). We controlled for
the hunting estate as random factor.
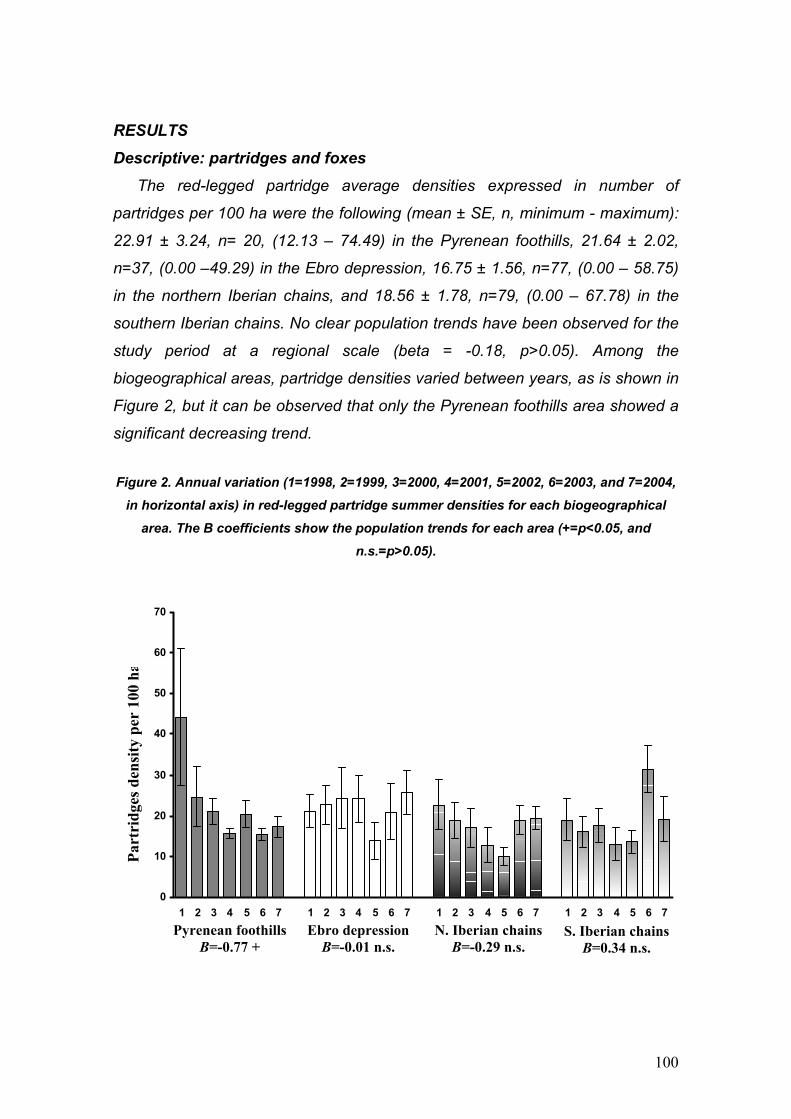
100
RESULTS
Descriptive: partridges and foxes
The red-legged partridge average densities expressed in number of
partridges per 100 ha were the following (mean ± SE, n, minimum - maximum):
22.91 ± 3.24, n= 20, (12.13 – 74.49) in the Pyrenean foothills, 21.64 ± 2.02,
n=37, (0.00 –49.29) in the Ebro depression, 16.75 ± 1.56, n=77, (0.00 – 58.75)
in the northern Iberian chains, and 18.56 ± 1.78, n=79, (0.00 – 67.78) in the
southern Iberian chains. No clear population trends have been observed for the
study period at a regional scale (beta = -0.18, p>0.05). Among the
biogeographical areas, partridge densities varied between years, as is shown in
Figure 2, but it can be observed that only the Pyrenean foothills area showed a
significant decreasing trend.
Figure 2. Annual variation (1=1998, 2=1999, 3=2000, 4=2001, 5=2002, 6=2003, and 7=2004, in horizontal axis) in red-legged partridge summer densities for each biogeographical
area. The B coefficients show the population trends for each area (+=p<0.05, and n.s.=p>0.05).
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 1 2 3 4 5 6 7 1 2 3 4 5 6 7 1 2 3 4 5 6 7
Part
ridg
es d
ensit
y pe
r 10
0 ha
Pyrenean foothills B=-0.77 +
Ebro depression B=-0.01 n.s.
N. Iberian chainsB=-0.29 n.s.
S. Iberian chains B=0.34 n.s.
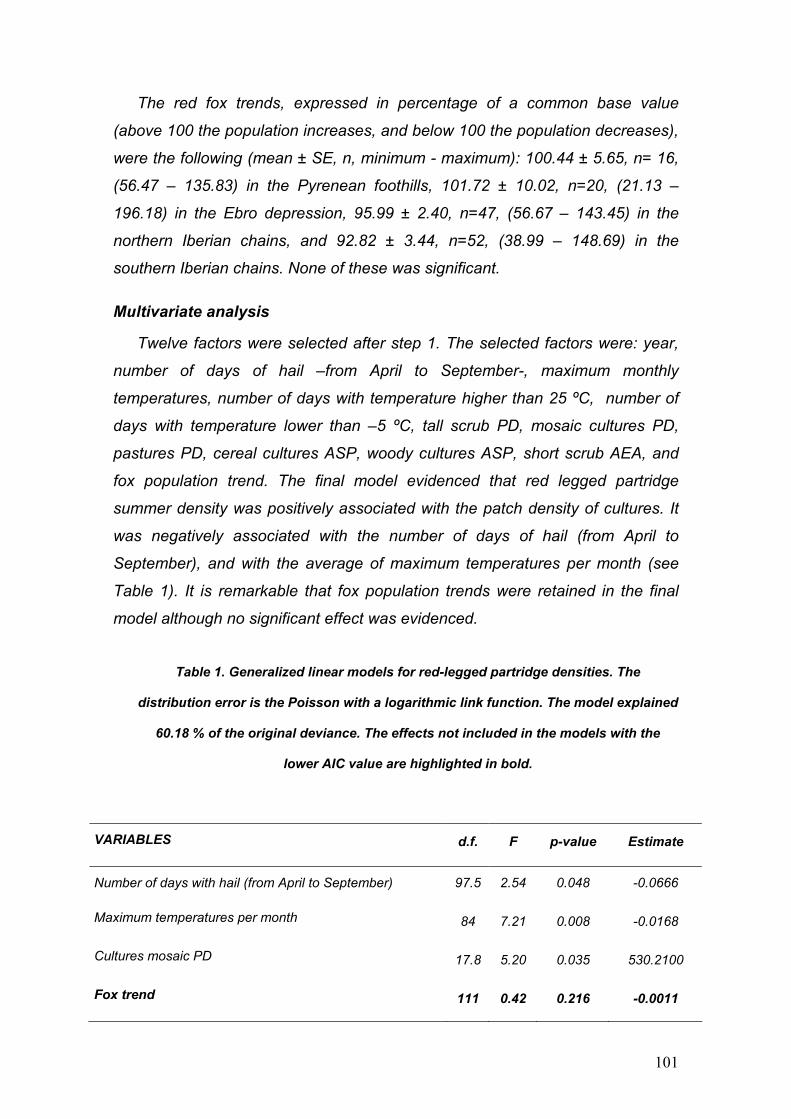
101
The red fox trends, expressed in percentage of a common base value
(above 100 the population increases, and below 100 the population decreases),
were the following (mean ± SE, n, minimum - maximum): 100.44 ± 5.65, n= 16,
(56.47 – 135.83) in the Pyrenean foothills, 101.72 ± 10.02, n=20, (21.13 –
196.18) in the Ebro depression, 95.99 ± 2.40, n=47, (56.67 – 143.45) in the
northern Iberian chains, and 92.82 ± 3.44, n=52, (38.99 – 148.69) in the
southern Iberian chains. None of these was significant.
Multivariate analysis
Twelve factors were selected after step 1. The selected factors were: year,
number of days of hail –from April to September-, maximum monthly
temperatures, number of days with temperature higher than 25 ºC, number of
days with temperature lower than –5 ºC, tall scrub PD, mosaic cultures PD,
pastures PD, cereal cultures ASP, woody cultures ASP, short scrub AEA, and
fox population trend. The final model evidenced that red legged partridge
summer density was positively associated with the patch density of cultures. It
was negatively associated with the number of days of hail (from April to
September), and with the average of maximum temperatures per month (see
Table 1). It is remarkable that fox population trends were retained in the final
model although no significant effect was evidenced.
Table 1. Generalized linear models for red-legged partridge densities. The
distribution error is the Poisson with a logarithmic link function. The model explained
60.18 % of the original deviance. The effects not included in the models with the
lower AIC value are highlighted in bold.
VARIABLES d.f. F p-value Estimate
Number of days with hail (from April to September) 97.5 2.54 0.048 -0.0666
Maximum temperatures per month 84 7.21 0.008 -0.0168
Cultures mosaic PD 17.8 5.20 0.035 530.2100
Fox trend 111 0.42 0.216 -0.0011

102
DISCUSSION
This paper presents an analysis of red-legged partridge population densities
over seven years in many localities in north-eastern Spain, combined with a
look at several factors that may affect these densities. Truly the only conclusive
statement this study makes is that in the whole region of Aragón (about 10% of
the Spanish mainland), partridge population densities are not significantly
decreasing, which is a very considerable finding in itself.
Partridge densities
A wide range of red-legged partridge density values is described in the
literature. The highest densities, over 400 partridges per 100 ha, were found in
intensively managed hunting areas from south-central Spain (Nadal 1998).
However, in some natural populations without this kind of management,
partridge densities were around 50 to 100 partridges per 100 ha (e.g. in French
scrublands, Pepin & Blayac 1990; and in Spanish olive groves, Duarte 1998).
Our results showed much lower partridge densities, only similar to those
obtained in released populations (Carvalho et al. 1998, Mereggi & Mazzoni
della Stella 2004), and in areas with low partridge densities (e.g. in Portuguese
agricultural lands, Borralho et al. 1996). The lower densities found in Aragón as
compared to other areas were documented previously by Gortázar et al. (2002).
We used the same fixed transects than the previous study. They attributed the
low densities obtained to two possibilities, the use by different authors of a wide
number of field methods to estimate partridge densities, and a true effect of
environmental constraints.
No clear population trends were observed for the study period at a regional
scale. This is in contrast with the hunters’ perceptions and with previous authors
(e.g. Carvalho et al. 1998). Among the biogeographical areas, partridge
densities varied between years but clear population trends were not observed,
and even a slight increase occurred in the southern Iberian chains.
The lowest partridge densities were obtained in the Iberian chains, which
showed the most stable population trends during the study period (see Figure
2). In the northern Iberian chains some areas have a well-preserved habitat and
a favourable climate. In contrast, some of the higher zones in the south Iberian
chains have a lack of cereal crops due to rural abandonment and most areas

103
are above 1,000 m a.s.l., with frequent frost that may limit the reproductive
success of the partridges (Slagsvold & Grasaas 1979). The highest densities
were obtained in the Pyrenean foothills and Ebro depression populations, the
trend being slightly decreasing in the 7 studied years. These differences could
be do to the intensification of agricultural practices in some areas of the
depression (mainly in the 1980’s), while traditional agriculture still dominates in
the mountain areas (Gortázar et al. 2002).
Influence of meteorological factors
Our results suggest that meteorological conditions are the main factors
affecting the summer abundance of red-legged partridges, even over the habitat
quality or predation. Most authors think that climatic conditions are key factors
limiting the galliform densities (Slagsvold & Grasaas 1979, Hermes et al. 1983,
Green 1984, Lucio 1990, Panek 1992, 2005), the weather conditions causing
only annual fluctuations (Panek 2005). The latter author suggested that the
weather conditions were an additional reason for the inter-annual density
changes, and consequently partridge populations oscillated around their
equilibrium level due to density-dependent parameters. We found that extreme
temperatures and hail were negatively related with partridge densities. This can
be explained in different ways:
i) Decreasing the chick survival: chick survival is a fundamental factor in
galliform population dynamics (Potts 1986), and the differences in this
parameter may produce the year-to-year changes in population densities (Blank
et al. 1967). Weather characteristics can influence the partridge chick, affecting
the quality and availability of food to both chicks and adults. Some studies have
shown that changes in temperature limit the number of chicks per pair in natural
populations of grey partridge, Perdix perdix, (Panek 1992) and red-legged
partridge (Lucio 1990). The influence of temperature on the number of chicks
per pair was shown experimentally (Hermes et al. 1983). These authors found
two explanations for this influence. First, excessively low temperatures in winter
may motivate a decrease in insect abundance (Panek 1992) that is positively
correlated with chick survival rates (Green 1984, Lucio 1991, Panek 1992).
Second, the relationship found between maximum temperature and partridge

104
densities can be related with chick death-rate during the harvest period. High
spring temperatures can bring forward the development of vegetation (Lucio
1990), but this is associated with bringing forward the harvest period. Hence, it
can increase the chick death-rate by agricultural machinery (Ricci 1985, Lucio
1990, Nadal et al. 1996, Meriggi & Mazzoni della Stella 2004).
Additionally, extremely high summer temperatures may increase the death
of partridges due to miopathy-like syndromes, that are reported in other lowland
partridge habitats in Spain (Höfle et al. 2004).
ii) Decreasing the productivity: it is well known that the climate influences a
bird’s fertility and productivity. Slagsvold & Grasaas (1979) working with
capercaille (Tetrao urogallus) found a positive relationship between the early
defrosting and autumnal densities. The food critical period can influence the
nest size (Lack 1968). The percentage of successfully hatched eggs may also
depend on factors such as temperature, storms, hail and rainfall (Green 1984).
iii) Increasing the predation of adults: the activity of partridges decreases
with low temperature and wet weather (Green 1984). This can make the birds
more vulnerable to predators, and can cause a loss of condition, again
increasing the vulnerability.
Normally, these demographic parameters do control the partridge population
dynamics. This is well known in grey partridge populations. The grey partridge
decrease in Great Britain between the 1960s and the 1990s was a result of a
decrease of both brood-production rate and chick survival rate (Potts 1986,
Potts & Aebischer 1994). Nevertheless, the grey partridge decrease in France
in 1990s was mainly an outcome of a decline in the survival rate of adults birds
(Bro et al. 2000, Reitz 2000). The population decrease of grey partridges in
Poland showed that it was connected with the drop of reproductive success,
including both the brood-production rate and the chick-survival rate, as well as
with the decline of the annual survival rate of adult birds (Panek 2005).

105
Habitat structure
Our results showed that cultures in mosaic were the habitat type with most
influence on red-legged partridge densities. This type of habitat is associated to
traditional agriculture schemes. These traditional schemes were based on the
distribution of cultures in small patches where the hedges were a key landscape
structure feature.
The development of agriculture was characterized by the increase of plot
dimension, hedge destruction, monocultures and development of harvesting
mechanization (Myers et al. 2000). Intensified agricultural practises and habitat
changes are believed to be key factors in explaining the decline of many
agricultural bird species (Chamberlain et al. 2000, Siriwardena et al. 2000).
Fuller et al. (1995) argued that the decrease in area of semi-natural grasslands
has reduced their capacity for farmland birds. Pärt & Söderström (1999) argued
that the combined effects of good foraging habitat close to safe nesting sites are
critical factors for breeding farmland birds. The negative effect of modern
agriculture on partridge populations was suggested before by several authors
(Ricci 1985, Lucio1991, Nadal et al. 1996, Fortuna 2002, Meriggi & Mazzoni
della Stella 2004). The grouping of agricultural plots that has taken place during
the last decades has already been indicated as a constraint for red-legged
partridge density in the north-west of Spain (Lucio 1991). Hedge reduction also
has a negative effect on partridge density in Spain (Fortuna 2002). Hedges are
areas with high availability of food, nesting-sites, shade and cover. The last two
characteristics can also be supplied by other types of vegetation, such as
vineyards or olive trees (Borralho et al. 1998). In our study, vineyards and olive
trees (noted woody cultures) were not related with the highest partridge
densities. These habitat types give shade and protective cover and are rich in
hedge-like microhabitats, but are intensively managed with agrochemicals and
machinery.
Hunting pressure
The over hunting pressure are considered for a lot of authors one of the most
important causes of of the Red-legged partridge decrease (Potts, 1986; Pepin y
Blavac, 1990; Lucio y Purroy, 1992; Borralho et al., 1997). Our findings in this
study are in contrast with this result, but is with the limited data on hunting

106
pressure available, the true effect of hunting on partridge populations may be
hidden. In addition, hunting occurs in autumn and winter and censusing was
done in late summer. If hunting is really less important than other factors, this
also implies that hunting regulations alone (e.g. lowering the hunting quotas)
may not significantly improve partridge densities in Aragón, in contrast with the
case of the rabbit in the same area (Williams et al. 2007).
Fox predation
The red fox is the most abundant wild rabbit predator in Aragón (Gortázar
1997), and rabbit populations were decreasing in practically all its distribution
area due to the additive effect of myxomatosis and RHD (e.g. Villafuerte et al.
1995, Gortázar 1997). Thus, one cause of the decline in the red legged
partridge populations could have been the switch of rabbit hunters and
predators towards alternative prey species following the decline in rabbit
populations (Gortázar et al. 2002). A few studies have shown the red fox or
generalist carnivores and raptors to be a significant cause of mortality in
partridge populations (e.g. Tapper et al. 1996, Bro et al. 2001, Panek 2005).
Through a predator control experiment, Tapper et al. (1996) showed that grey
partridge population densities were reduced by the combined effects of
predation by several farmland bird predators, including red fox. Panek (2005)
suggested that to improve the situation of grey partridges, efforts should be
focused on the reduction of predators, mainly red fox populations. Moreover,
the environmental changes may have increased predator impact (Reynolds &
Tapper 1996).
Much to our surprise, fox predation effects were not clearly evidenced in our
study. The first explanation for this apparent lack of relationship is that foxes
were counted in transects that were close to, but not exactly at the partridge
census transects. Moreover, no fox data were available for 12 of 36 sites. But it
may also be explained by the low fox abundances according to our surveys and
to similar data already reported from the region (Gortázar et al. 2000, 2002).
From a regional perspective, our results did not show any correlation
between partridge population densities and red fox population trends. While
trends do not directly measure predation pressure, as was done in other
studies, it would be expected that if fox predation was a main cause of partridge

107
death for a majority of the localities in our study then there would be a negative
correlation between the two species’ population parameters. This finding agrees
with similar results obtained in analysing fox and rabbit trends in Aragón
(Williams et al. 2007).
Conclusions
We found that the meteorological circumstances were the most important
factors that limited summer partridge densities. Hence, the climatic
characteristics of a territory are, in our opinion, the main factor that determines
their environmental potentiality as far as the red-legged partridge summer
abundance is concerned.
Within a given bioclimatic area, some habitat characteristics such as small
plot size and high boundary proportion do also benefit the partridge densities.
One of the main conservation challenges in Mediterranean areas is the loss of
this habitat type caused by the intensification of agricultural practices.
According to the premise that red-legged partridge exploitation should be
sustainable, populations with low densities generate questions regarding the
management of the hedges in the landscape, regeneration of agricultural
mosaics to improve partridge densities.
ACKNOWLEDGEMENTS
Partridge censusing is part of the game management actions of Asesoría de
Caza y Pesca, Dirección General de Medio Natural, Gobierno de Aragón.
Special thanks to Emilio Escudero, Adela García and Gaspar León who
coordinate the scheme. This work would never have been done without the
enthusiastic collaborations of Aragón’s Game Rangers. Joaquín Vicente helped
us with the statistical analysis. D Villanúa and P Acevedo were supported by the
joint project CSIC/Principado de Asturias.
REFERENCES
1. Aebischer, N. & Kavanagh, B. (1997). Grey partridge. In Hagemeijer,
W.J.M., Balir, M.J. (eds.), The EBCC Atlas of European Breeding Birds, their
Distribution and Abundance. T&AD Poyser, London, pp. 212-213.

108
2. Aebischer, N.J. & Potts, G.R. (1994). Red–legged partridge. In: Tucker, G.
M. and Heath, M. F., Birds in Europe. Their Conservation Status. Birdlife
Conservation Series nº3, Birdlife International, Cambridge, UK.
3. Baratti, M., Ammannati, M., Magnelli, C. & Dessi-Fulgheri, F. (2005).
Introgression of chukar genes into a reintroduced red-legged partridge
(Alectoris rufa) population in central Italy. Animal Genetics. 36(1): 29-35.
4. Barnes, R.F.W. & Tapper, S. (1984). A method for counting hares by
spotlight. Journal of Zoology of London, 206: 273-276.
5. Bernabeu, R.L. (2000). Evaluación económica de la caza en Castilla-La
Mancha, (PhD Thesis, Universidad de Castilla-La Mancha, España).
6. Bibby, C.J., Budgess, N.D. & Hill, D.A. (1992). Bird census techniques.
London : Academic Press.
7. Bielza de Ory, V. (1993). Atlas geográfico-temático de Aragón. Diputación
General de Aragón, Zaragoza.
8. Blank, T.H., Southwood, T.R.E. & Cross, D.R. (1967). The ecology of the
partridges. I. Outline of population processes with particular reference to
chick mortality and nest densitiy. Journal of Animal Ecology, 36: 549-556.
9. Borralho, R., Rego, F. & Vaz-Pinto, P. (1996). Is driven transect sampling
suitable for estimating Red-legged Partridge Alectoris rufa densities? Wildlife
Biology, 2: 259-268.
10. Borralho, R., Rego, F. & Vaz Pinto, P. (1997). Demographic trends of red-
legged partridges (Alectoris rufa) in southern Portugal after implementation
of management actions. Gibier Faune Sauvage, 14: 585-599.
11. Borralho, R., Rito, A., Rego, F., Simoes, H. & Vaz-Pinto, P. (1998).
Summer distribution of Red-legged partridge (Alectoris rufa) in relation to
water availability on Mediterranean farmland. Ibis, 140: 620-625.
12. Blanco-Aguiar, J.C., Virgós, E., Villafuerte, R. (2003) Perdiz roja,.
Alectoris rufa. In: Martí R, del Moral JC (eds) Atlas de las aves.
reproductoras de España.
13. Blank, T.H., Southwood, TRE, & Cross, DJ (1967) The ecology of the
partridge. I . Outline of population processes with particular reference to
chick mortality and nest density. Journal of Animal Ecology, 36: 549-556.

109
14. Brickle, N.W., Harper, D.G.C., Aebischer, N.J. & Cockayne, S.H. (2000).
Effects of agricultural intensification on the breeding success of corn
buntings Milaria calandra. Journal of Applied Ecology 37: 742-755.
15. Bro, E., Sarrazin, F., Clobert, J. & Reitz, F. (2000). Demography and
decline of the grey partridge Perdix perdix in France. Journal of Applied
Ecology 37: 432-448.
16. Bro, E., Reitz, F., Clobert, J., Migot, P. & Massot, M. (2001). Diagnosing
the environmental causes of the decline in grey partridge Perdix perdix
survival in France. Ibis. 143: 120-132.
17. Burnham, K.P., Anderson, D.R. & Laake, J.L. (1980). Estimation of
density from line transect sampling of biological populations. Wildlife
Monographs, 72: 1-202.
18. Calderón, J. (1983). La perdiz roja, Alectoris rufa (L.). Aspectos
morfológicos, taxonómicos y biológicos. (PhD Thesis Universidad
Coplutense de Madrid, España)
19. Campbell, L.H., Avery, M.I., Donald, P., Evans, A.D., Green, R.E. &
Wilson, J.D. (1997). A review of the indirect effects of pesticides on birds.
JNCC Report No. 227. Joint Nature Conservation Committee, Peterborough,
UK.
20. Carvalho, J., Castro-Pereira, D., Capelo, M. & Borralho, R. (1998). Red-
legged partridge (Alectoris rufa) restocking programs: Their success and
implications on the breeding population. Gibier Faune Sauvage, Game Wild.
15: 465-474.
21. Chamberlain, D.E., Fuller, R.J., Bunce, R.G.H., Duckworth, J.C. &
Shrubb, M. (2000). Changes in the abundance of farmland birds in relation
to the timing of agricultural intensification in England Wales. Journal of
Applied Ecology 37: 771-788.
22. Cramp, S. & Simmons, K.E.L. (Eds.)(1980). The Birds of the Western
Palearctic. Vol. II. Oxford University Press. Oxford.
23. Crawford, T.J. (1991). The calculation of index numbers from wildlife
monitoring data. Chap. 12 in Monitoring for Conservation and Ecology. (B.
Goldsmith, ed.). Chapman and Hall, London.
24. Crawley, M.J. (1993). GLIM for Ecologists. Blackwell.- London, UK.

110
25. Crick, H.Q.P., Dudley, C., Evans, A.D. & Smith, K.W. (1994). Causes of
nest failure among buntings in the UK. Bird Study. 41: 88-94.
26. Delibes, M. (1988). La caza de la perdiz roja en España. Destino (eds.).
Barcelona.
27. Delibes M. & Hiraldo, F. (1981). The rabbit as prey in the Iberian
Mediterranean ecosystems. Proceedings of the World Lagomorph
Conference (eds. K. Myers and C.D. MacInnes pp. 614-622. University of
Güelph and Wildlife Research, Ministry of Natural Resources, Ontario).
28. Duarte, J. (1998). La perdiz roja (Alectoris rufa) en el olivar: métodos de
estimación demográfica. Tesina. Universidad de Málaga.
29. Fortuna, M.A. (2002). Selección de hábitat de la perdiz roja (Alectoris rufa)
en periodo reproductor en relación con las características del paisaje de un
agrosistema de la mancha (España). ARDEOLA, 49: 59-66.
30. Fuller, R.J., Gregory, R.D., Gibbons, D.W., Marchant, J.H., Wilson, J.D.,
Baillie, S.R. & Carter, N. (1995). Population declines and range
contractions among lowland farmland birds in Britain. Conservation Biology,
9: 1425-1441.
31. Gortázar, C. (1997) Relative abundance wild rabbit (Oryctolagus cuniculus)
and red fox (Vulpes vulpes) after rabbit haemorrhagic disease (rhd) in the
central Ebro bassin in northeastern spain. Zeitschrift für Jagdwissenschaft
43: 259-265.
32. Gortázar, C., Villafuerte, R. & Martín, M. (2000). Success of traditional
restocking of red-legged partridge for hunting purposes in areas of low
density of northeast Spain Aragón. Zeitschrift für Jagdwissenschaft, 46: 23-
30.
33. Gortázar, C., Villafuerte, R., Escudero, M.A. & Marco, J. (2002). Post-
breeding densities of the red-legged partridge (Alectoris rufa) in
agrosystems: a large.scale study in Aragón, Northeastern Spain. Zeitschrift
für Jagdwissenschaft , 48: 94-101.
34. Green, R.E. (1984). The feeding ecology and survival of partridge chicks
(Alectoris rufa and Perdix perdix) on arable faroland in East Anglia. Journal
of Applied Ecology, 21: 817-830.
35. Green, R.E. (1995). The decline of the corncrake Crex crex in Britain
continues. Bird Study, 42: 66-75.

111
36. Hermes, J., Woodard, A., Vohra, P. & Snyder, R. (1983). The effect of
ambiental temperature and energy level on reproduciton in Red-legged
partridges. Poultry Science, 62: 1160-1168.
37. Höfle, U., Millán, J., Gortázar, C., Buenestado, F.J., Marco, I. &
Villafuerte, R. (2004). Self-injury and capture myopathy in net-captured
juvenile red-legged partridge with necklace radiotags. Wildlife Society
Bulletin 32: 344-350.
38. Lack, D. (1968). Ecological adaptations for breeding in birds.Methnen Ed.
London.
39. Lucio, A.J. (1990). Influencia de las condiciones climáticas en la
productividad de la perdiz roja (Alectoris rufa). ARDEOLA 37: 207-218.
40. Lucio, A.J. (1991). Selección de hábitat de la perdiz roja (Alectoris rufa) en
matorrales supramediterráneos del NW de la cuenca del Duero.
Aplicaciones a la gestión del hábitat cinegético. Ecología 5: 337-353.
41. Lucio A.J. (1998). Perdiz roja. In: F.J. Purroy (ed.) Atlas de las aves de
España. Barcelona: Lynx.
42. Lucio, A.J. & Purroy, F.J. (1992). Caza y conservación de aves en
España. ARDEOLA, 39(2): 85-98.
43. Mereggi, S. & Mazzoni della Stella, R. (2004). Dynamics of a reintroduced
population of red-legged partridges Alectoris rufa in central Italy. Wildlife
Biology, 9: 1-9.
44. Myers N., Mittermeier R.A., Mittermeier C.G., da Fonseca G.A.B. & Kent
J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403: 853-
958.
45. Nadal, J., Nadal, J. & Rodríguez-Teijerio, J.D. (1996). Red-legged
partridge (Alectoris rufa) age and sex ratios in declining populations in
Huesca (Spain) applied to management. Rev. Ecol. (Terre Vie) 51: 243-257.
46. Nadal, J. (1998). La bioecología de la perdiz roja. In: Federación Española
de Caza (eds.). La perdiz roja. Madrid. Federación Española de Caza, pp.
33-45.
47. Panek, M. (1992). The effect of environmental factors on survival of Grey
Partridge (Perdix perdix) chicks in Poland During 1987-1989. Journal of
Applied Ecology, 29: 745-750.

112
48. Panek, M. (2005). Demography of grey partridges Perdix perdix in Poland in
the years 1991-2004: reasons of population decline. European Journal of
Wildlife Research, 51: 14-18.
49. Pärt, T. & Söderström, B. (1999). The effects of management regimes and
location in landscape on the conservation of farmland birds breeding in
semi-natural pastures. Biological Conservation, 90: 113-123.
50. Pepin, D. & Blayac, J. (1990). Impacts d´un amenagement de la garrigue et
de l’instauration d´un plan de chasse sur la démographie de la perdix rouge
(Alectoris rufa) en milieu méditerranéen. Gibier Faune Sauvage, Game Wild.
7: 145-158.
51. Potts, G.R. (1980). The effects of modern agriculture, nest predation and
game management on the population ecology of partridges (Perdix perdix
and Alectoris rufa). Ecology Research, 2: 2-79.
52. Potts, G.R. (1986). The partridge, pesticide, predation and conservation.
London: Collins.
53. Potts, G.R. & Aebischer, N.J. (1994). Population dynamics of the grey
partridge Perdix perdix 1793-1993: monitoring, modelling and management.
Ibis, 137: S29-S37.
54. Rands, M.R.W. (1987). The effect of nest site selection on nest predation in
grey partridge Perdix perdix and red-legged partridge Alectoris rufa. Ornis
Scandinavica, 19: 35-40.
55. Reitz, F. (2000). The status of partridges in North-Central France. In :
Faragó S (ed.). Proccedings of an international symposium on partridges,
quails and pheasants in the Western Paleartic and Neartic. Hungarian Small
Game Bulletin, 5: 151-163.
56. Reynolds, J.C. & Tapper, S.C. (1996). Control of mammalian predators in
game management and conservation. Mammal Review, 26: 127-155.
57. Ricci, J.C. (1985). Utilisation de quelques ressources du milieu parles
nichées de perdix rouge (Alectoris rufa) dans un agrosystème de type
polyculture élevage. Gibier Faune Sauvage, 2: 15-38.
58. SEO/BirdLife (2004). Resultados del Programa de Seguimiento SACRE.
Informe 2004.

113
59. Siriwardena, G.M., Crick, H.Q.P., Baillie, S.R. & Wilson, J.D. (2000).
Agricultural land-use and spatial distribution of granivorous lowland
farmland birds. Ecography, 23: 702-719.
60. Slagsvold, T. & Grasaas, T. (1979). Autumn population size of the
Capercaille in relation to weather. Ornis Scandinavica, 10: 37-41.
61. Tapper, S., Green, R.E. & Rands M.R.W. (1982). Effect of mammalian
predators on partridges populations. Mammal Review, 12: 159-167.
62. Tapper, S.C., Potts, G.R. & Brockless, M.H. (1996). The effect of a
experimental reduction in predator pressure on the breeding success and
population density of grey partridges Perdix perdix. Journal of Applied
Ecology, 33: 965-978.
63. Villafuerte, R., Calvete, C., Blanco, J.C. & Lucientes, J. (1995). Incidence
of viral hemorrhagic disease in wild rabbit populations in Spain. Mammalia,
59: 651-659.
64. Villafuerte, R., Fernández de Luco, D., Gortázar, C. & Blanco, J.C.
(1996). Effect on red fox litter size and diet after rabbit haemorrhagic
disease in northeastern Spain. Journal of Zoology, 240: 764-767.
65. Williams, D., Acevedo, P., Gortázar, C., Escudero, M., Labarta, J.,
Marco, J. & Villafuerte, R. (2007). Hunting for answers: rabbit (Oryctolagus
cuniculus) population trends in northeastern Spain European Journal of
Wildlife Research, 53 (1): 19-28.
66. Wilson, K. & Grenfell, B.T. (1997). Generalized linear modelling for
parasitologists. Parasitology Today, 13: 33-38.

114
IX. SÍNTESIS GENERAL
Este apartado pretende ser, como su propio nombre indica, una síntesis de
los resultados más relevantes obtenidos durante la realización de la presente
Tesis Doctoral, discutiendo especialmente sus posibles implicaciones en la
conservación y gestión de la perdiz roja.
En el marco de esta tesis se han desarrollado estudios observacionales,
experimentos en perdices de granja, y se han tratado datos procedentes de un
programa de monitorización. La combinación de estas tres fuentes de
información ha permitido avanzar en el conocimiento práctico de cara a la
gestión y conservación de la perdiz roja en España.
La introducción de patógenos. (Capítulos 1 y 4)
Uno de los resultados más relevantes de la presente tesis es sin duda el
hallazgo en las fincas con suelta de tres especies de parásito (los nematodos
Eucoleus contortus y Aonchoteca caundinflata y el cestodo Skyabinia bolivari),
típicas de aves de granja y no descritas anteriormente en perdices silvestres
(Millán et al., 2004).
La presencia de estos parásitos en aves cazadas podría deberse en
principio a dos situaciones; o bien que estuviesen parasitando aves de granja
que portasen los parásitos antes de la suelta o que se tratase de aves
silvestres que se hubiesen infectado con los parásitos traídos por las perdices
liberadas.
La distinción entre aves de granja y silvestres puede resultar, a pesar de las
diferencias esplacnométricas existentes entre unas y otras (Millán et al 2001),
sumamente difícil si las aves liberadas no van convenientemente anilladas. Por
este motivo se hace necesario analizar las consecuencias que podría tener una
y otra posibilidad.
En caso de tratarse de aves de granja, estas llevarían en el campo más de
dos meses en el momento de la toma de muestras, tiempo más que suficiente
para transmitir los nuevos parásitos a sus congéneres silvestres. Esta
transmisión se vería además favorecida por el incremento de la excreción de
propágulos parasitarios que sigue a la suelta, motivado por el estrés que

115
supone para las aves la liberación a un medio desconocido y al cese de los
tratamientos (Villanúa et al 2006).
La otra posibilidad sería que se tratase de aves silvestres que se hubiesen
infectado tras la llegada de los nuevos parásitos con la suelta de aves de
granja. A favor de esta segunda hipótesis estaría el hecho de que menos de un
20 % de las perdices de granja liberadas sobreviven dos meses en el campo
(Birkan, 1971; Capelo y Pereira, 1996; Gortázar et al., 2000), que la
supervivencia se supone todavía menor en aves parasitadas por el nematodo
Eucoleus contortus, ya que éste hace a las galliformes más detectables por el
zorro (Millán et al 2002), y que dos de estos parásitos fueron encontrados en
aves adultas, cuando las sueltas se llevan a cabo con individuos juveniles.
Parece pues más probable que las aves infectadas por estos parásitos
propios de granja fuesen aves silvestres a las que les hubiesen transmitido los
patógenos las aves liberadas con fines cinegéticos. Este hecho tiene
importancia, ya que precisamente Eucoleus es un parásito con importante
efecto patógeno sobre su hospedador. Por otra parte, habiendo confirmado que
las sueltas de perdiz de granja dan lugar a la introducción de helmintos, surge
la duda sobre qué otros patógenos, por ejemplo virus o bacterias, estarán sido
introducidos de la misma forma.
Otra posible prueba de la introducción de parásitos sería el hallazgo del
nematodo Eucoleus contortus parasitando un macho de sisón (Tetrax tetrax),
procedente de una finca en la que se realizan sueltas de perdices con fines
cinegéticos. Nunca antes se había detectado este parásito en esta especie ni a
ninguna otra propia de los ambientes esteparios, mientras que resulta muy
frecuente en perdices en cautividad y se ha encontrado en ejemplares de
granja cazados en esta misma finca.
Para comprender la magnitud del problema que podría suponer la
introducción de nuevos patógenos en el campo se puede recurrir a los trabajos
de Tompkins et al (1999, 2000, 2001) comentados en la introducción a esta
tesis. Estos autores demuestran, basándose en estudios experimentales
(Tompkins et al 1999 y 2001) y en modelos matemáticos (Tompkins et al 2000),
que la introducción del nematodo Heterakis gallinarum con las sueltas de faisán
vulgar con fines cinegéticos podrían haber causado el descenso sufrido por la
perdiz pardilla en Inglaterra.

116
Trasladando este ejemplo al caso español, la suelta de perdices de granja,
con la consiguiente exposición de las poblaciones naturales de perdiz (u otras
aves como el sisón) a los nuevos parásitos introducidos, podría tener graves
consecuencias desde e punto de vista de conservación de las mismas,.
Como nota de esperanza cabe señalar el hecho de que el hallazgo de estos
parásitos propios de granja se ha limitado a aquellas zonas en las que se
realizan repoblaciones, no habiéndose encontrado nunca en fincas de perdiz
salvaje limítrofes a dichas fincas con suelta. Parece pues que la capacidad de
dispersión de estos patógenos en el campo está limitada a una superficie
pequeña, lo que permitiría a cada gestor controlar la introducción de estos
patógenos a sus poblaciones independientemente de lo que se haga en las
fincas vecinas. Obviamente, este extremo no debe tomarse como excusa para
relajar los controles sanitarios, y no tiene por qué ser extensivo a la eventual
difusión de patógenos de etiología vírica o bacteriana.
La insuficiente profilaxis actual. (Capítulos 2 y 3)
La cría de perdices en cautividad se inició en nuestro país hace apenas 30
años (Millán et al., 2004) y su relevancia dentro del mundo veterinario-
ganadero es todavía poco importante, motivos por los cuales la investigación
de las industrias farmacológicas en este campo ha sido muy escasa. Fuera de
nuestras fronteras existe algún trabajo acerca de la efectividad de tratamientos
antiparasitarios en aves cinegéticas de granja (Kirsch, 1983), pero se trata de
experiencias con otras especies (faisán y perdiz pardilla) y son relativamente
antiguas (casi 25 años). Este vacío de conocimientos científicos acerca de los
tratamientos en perdiz roja ha forzado a los propietarios y veterinarios a copiar
los protocolos de tratamiento de la industria avícola que, como se ha puesto de
manifiesto en el capitulo 3 de la presente tesis, no tienen por qué ser los
adecuados.
Esta tesis ha contribuido a comprobar que el tratamiento con Albendazol,
que habitualmente se usa para tratar las infecciones por nemátodos en las
granjas de perdiz, puede disminuir la excreción de propágulos parasitarios,
pero no es capaz de acabar con la infección. Desde un punto de vista de
producción en granja tal vez pudiese ser suficiente, ya que este tratamiento

117
disminuirá los síntomas y minimizará las bajas, pero desde luego no basta
cuando lo que se busca es conseguir aves con plenas garantías sanitarias de
cara a su liberaración en el medio natural.
La misma situación de falta de conocimientos específicos que hemos
comentado para el caso de los tratamientos es la que nos encontramos al
revisar los protocolos diagnósticos utilizados. Los resultados expuestos en el
capítulo 2 ponen de manifiesto cómo factores tan simples como la hora de
recogida de las muestras o el tipo de heces analizadas, son capaces de hacer
que se diagnostique como sana a una perdiz intensamente parasitada.
Mediante un experimento controlado, se pudo detectar un aumento muy
significativo en la excreción de E. contortus y Eimeria sp. a lo largo del día,
hasta el punto de que en las heces de aves que a las 8:00 de la mañana
estaban libres de excreción, doce horas más tarde se cuantificaban más de
15.000 propágulos por gramo para el caso de Eucoleus contortus o 30.000
ooquistes por gramo para el caso de Eimeria sp. Una situación semejante
sucedía al comparar los resultados de los análisis coprológicos realizados
sobre heces corrientes o sobre heces de ciegos, con una ausencia casi total de
propágulos de Eucoleus contortus en las de ciego y una presencia mucho
menor de ooquistes de Eimeria sp. en las heces corrientes. Estos errores
serán probablemente poco importantes en los sistemas de producción avícola
industrial, donde las aves reproductoras están permanentemente alojadas en
jaulas elevadas que cortan los ciclos parasitarios y los pollos van a matadero
con apenas 2 meses de vida, pero en el caso de las perdices criadas en granja
para su liberación en el campo el problema es muy grave, y debe ser por tanto
enfocado desde un punto de vista totalmente diferente.
En definitiva, teniendo en cuenta la insuficiente eficacia de los tratamientos
actuales y la variable fiabilidad en la detección de las aves parasitadas, a día
de hoy parece desaconsejable realizar repoblaciones con perdices de granja, al
menos si lo que se busca es mejorar el estado de las poblaciones naturales.
Existen otras alternativas. (Capitulo 5)
El análisis de los datos de abundancia obtenidos dentro del plan de
monitorización de Aragón ha puesto de manifiesto que las poblaciones de

118
perdiz de esta comunidad autónoma no están sufriendo el declive registrado (o
percibido) en otras zonas. Esto tiene implicaciones en gestión cinegética, ya
que implica que las poblaciones de perdiz pueden mantenerse (bien es cierto
que a densidades modestas) en ausencia de sueltas masivas y permitiéndo
una extracción cinegética aceptable. El análisis paralelo de las tendencias
poblacionales de predadores, particularmente del zorro, sugiere además que
las poblaciones vulpinas resultan igualmente estables, reduciendo así los
argumentos a favor de un mayor control de este depredador.
Así pues, parece lógico pensar que para conservar la perdiz roja en el
centro y sur, se deberían tal vez imitar algunos aspectos del modelo de gestión
de Aragón, donde la suelta de perdices es poco frecuente, el control de
predadores es limitado, y la administración cuenta con un excelente esquema
de monitorización de las poblaciones de especies cinegéticas.
En este mismo estudio se ha podido comprobar que, si bien los factores
climáticos son el principal factor limitante de las abundancias de perdiz, existen
una serie de elementos del hábitat que son de vital importancia para el
mantenimiento de las poblaciones de perdiz roja y que son susceptibles de ser
fomentados. El más significativo resultó ser el parámetro que cuantificaba el
mosaico de cultivos, de manera que las mayores abundancias de perdiz se
encontraban en aquellas zonas en las que no se había perdido todavía la
agricultura tradicional, con alternancia de cereales con otros cultivos como el
olivo, la viña o los espárragos.
A lo largo de los últimos años, este tipo de paisaje ha sido sustituído en
gran parte de la Península Ibérica por los monicultivos de cereal derivados de
concentraciones parcelarias y un mal enfoque de la Política Agraria
Comunitaria (PAC)(Díaz et al. 2006). Parece lógico asumir pues que esta
importante pérdida de calidad del hábitat sea una de las causas del declive de
la perdiz roja, pero no lo es menos pensar que, si se consiguiese restaurar este
paisaje, las poblaciones de perdiz podrían también recuperarse. A día de hoy
nos encontramos en plena renovación de los Programas de Desarrollo Rural
(PDR) debido a la aplicación del Reglamento (CE) nº 1698/2005 para el
periodo 2007-2013, motivo por el cual es el momento idóneo para, a traves de
subvenciones, tratar de recuperar este mosaico de cultivos y con el las aves

119
ligadas este medio, entre las que se encontraría la perdiz roja (De la Concha et
al., 2006).
Bibliografía
1. Birkan, M. (1990). La perdrix rouge. Brochures techniques O.N.C., Paris. 36
pp.
2. Capelo, M. y Castro Pereira, D. (1996). Sobrevivência e dispersão de
perdizes (Alectoris rufa L.) largadas em duas operações de repovoamento
cinegético. Revista Forestal, 9: 245- 253.
3. De la Concha, I., Hernández, C y Pinilla, J. (2006). Medidas beneficiosas
para las aves financiables a traves del nuevo reglamento de desarrollo rural.
Sugerencias para su diseño y aplicación en NATURA 2000. SEO/Birdlife.
Madrid. España.196 pp.
4. Díaz, M., Baquero, R.A., Carricondo, A., Fernández, F., García, J. y Yela,
J.L. (2006). Bases ecológicas para la definición de las prácticas agrarias
compatibles con las Directivas de Aves y de Hábitats. Convenio Ministerio
MEdio Ambiente-Universidad de Castilla la Mancha. Informe Inédito.
5. Gortázar, C., Villafuerte, R., y Martín, M. (2000) Success of traditional
restocking of red-legged partridge for hunting purposes in areas of low
density of northeast Spain Aragón. Zeitschrift für Jagdwissenschaft, 46: 23-
30.
6. Kirsch, R. (1983) Treatment of Nematodiasis in Poultry and Game birds
with Febendazole. Avian Diseases, 28(2): 311-318.
7. Millán, J., Gortazar, C., and Villafuerte, R. (2001) Marked differences in
the splanchnometry of farm-bred and wild red-legged partridges (Alectoris
rufa L.). Poultry Science, 80: 972-976.
8. Millán, J., Gortázar, C., Tizzani, P. and Buenestado F.J. (2002) Do
helmints increase the vulnerability of released pheasants to fox predation?.
Journal of Helmintology, 76: 225-229.

120
9. Millán, J., Gortázar, C. & Villafuerte, R. (2004) A comparison of the
helminth faunas of wild and farm-reared red-legged partridges. Journal of
Wildlife Management, 68(3): 701-707
10. Tompkins, D.M., Dickson, G. y Hudson, P.J. (1999). Parasite-mediated
competition between pheasant and grey partridge: a preliminary
investigation. Oecologia, 119: 378-382
11. Tompkins, D.M., Greenman, J.V., Robertson, P.A. y Hudson, P.J. (2000).
The role of shared parasites in the exclusion of wildlife host: Heterakis
gallinarum in the ring-necked pheasant and the grey partridge. Journal of
Animal Ecology, 69: 829-840.
12. Tompkins, D.M., Greenman, J.V. y Hudson P.J. (2001). Differential impact
of shared nematode parasite on two gamebird host: implications for
apparent competition. Parasitology, 122: 187-193.
13. Villanúa, D., Acevedo, P., Höfle, U., Rodríguez, O. & Gortázar, C. (2006).
Changes in transmission stage excretion after pheasant release. Journal of
Helminthology, 80: 1-7.

121
X. CONCLUSIONES, PERSPECTIVAS Y RELFLEXIONES
Conclusiones
1. La liberación con fines cinegéticos de perdices criadas en granja supone la
introducción de nuevos patógenos en el medio natural. Esto podría afectar
negativamente a las poblaciones silvestres de perdiz roja, así como a otras
aves que compartan su hábitat., No obstante, y al menos en lo que se refiere a
helmintos como Eucoleus contortus, Aonchoteca caundinflata y Skryabinia
bolivari, la capacidad de diseminación de estos patógenos en las poblaciones
silvestres de perdiz roja parece limitada.
2. La excreción de propágulos parasitarios varía a lo largo del día y con el tipo
de heces. En consecuencia, los protocolos actuales de detección de aves
parasitadas y cuantificación de la excreción de parásitos, basados en
exámenes coprológicos, no ofrecen suficientes garantías, hasta el punto de que
aves altamente parasitadas pueden pasar por sanas. Tanto el tipo de
excremento a muestrear como el horario de muestreo deberán estandarizarse.
3. El tratamiento con albendazol (a dosis de 20 mg/kg V.O.), habitualmente
utilizado contra las infecciones por nemátodos en las granjas de perdiz roja,
reduce significativamente la excreción de propágulos de los parásitos
Aonchoteca caundinflata y Heterakis gallinarum. Sin embargo, el tratamiento no
logra la eliminación completa de los helmintos adultos. Por consiguiente, este
tratamiento resulta insuficiente para prevenir la eventual introducción de estos
parásitos en el medio natural.

122
4. Las poblaciones de perdiz roja donde no se efectúan sueltas parecen no
estar sufriendo el acusado descenso detectado en las zonas más manejadas.
En estas poblaciones naturales, los principales factores limitantes de la
abundancia estival de perdiz son la temperatura máxima mensual, el número
de días de lluvia entre abril y septiembre y la abundancia de cultivos en
mosaico. La abundancia de depredadores y la presión cinegética del año
anterior parecen comparativamente menos relevantes.
Perspectivas y reflexiones
A lo largo del desarrollo de la presente tesis han ido surgiendo dudas y se
han puesto de manifiesto vacíos de conocimiento que sería interesante abarcar
en futuros trabajos. Por ello creemos acertado añadir la coletilla de
“reflexiones”.
• Mejora de los tratamientos profilácticos.
A lo largo del desarrollo de la presente tesis se ha podido comprobar cómo
existen numerosos trabajos científicos en los que se recurre a las
desparasitaciones como parte del estudio, tratando de evaluar el efecto que la
eliminación de los parásitos tendría sobre el éxito reproductor, la condición
física o cualquier otro parámetro. Por contra, apenas hay unos pocos estudios
en los que se evalúe la eficacia del fármaco utilizado o las posibles limitaciones
de los métodos diagnósticos utilizados. Como ya hemos mencionado
anteriormente, este vacío hace que los protocolos profilácticos utilizados en
fauna silvestre sean inadecuados. Así pues, parece muy importante que una
parte del esfuerzo investigador en fauna silvestre se centre en tratar de
encontrar protocolos de tratamiento mejores a los actuales puesto que es poco

123
probable que la industria farmacológica lo haga, y la falta de estos protocolos
puede suponer un grave problema de conservación.
• Evaluación del papel de las repoblaciones como focos de introducción
de otros patógenos, tales como bacterias o virus.
Por motivos económicos y de tiempo, esta tesis tuvo que centrarse en el
estudio de los parásitos, pero parece lógico pensar que otros agentes
patógenos, como bacterias o virus, puedan estar siendo también introducidos
en el campo con la suelta de aves de granja. Teniendo en cuenta que dentro de
los agentes infecciosos que afectan a las perdices en granja se encuentran
varias enfermedades de declaración obligatoria (Bronquitis infecciosa aviar,
enfermedad de Gumboro, clamidiosis aviar, enfermedad de Marek, enfermedad
de Newcastle, influenza aviar, laringotraqueítis infecciosa, micoplasmosis aviar,
tifosis aviar…) para las cuales existe además plan de erradicación en España,
parece lógico pensar que la importancia que tendría la introducción de estas en
el campo sería grandísima.
• Divulgación de los trabajos científicos entre los colectivos implicados.
En muchos casos, cuando un gestor de caza se plantea soltar perdices de
granja en la finca o coto que gestiona, lo hace porque piensa que es una buena
actuación y que con esa suelta está contribuyendo a la conservación de la
especie. Esa idea está, por desgracia, muy generalizada en el colectivo de
gestores y cazadores, y no hay otra causa que la amplia publicidad que se le
da dentro de la prensa del sector. Basta con ojear cualquier revista relacionada
con la caza para comprobar cuan numerosos son los artículos en los que se
alaban las extraordinarias tasas de supervivencia y capacidad de vuelo de las
perdices de tal o cual criador. Sin embargo, apenas aparecen unos pocos
artículos en los que se cuestione la idoneidad de estas actuaciones o en los
que se analice científicamente la viabilidad de las sueltas. Por el contrario,
cuando revisamos la bibliografía científica acerca del tema ocurre lo contrario;
encontramos numerosos trabajos en los que se demuestra la escasa
supervivencia de las aves criadas en granja y los graves riesgos que estas

124
actuaciones pueden suponer para las poblaciones naturales, pero no
encontramos ni un solo trabajo en el que se demuestre lo contrario. Así pues,
una de las primeras actuaciones que el colectivo científico debería llevar a cabo
para conservar la perdiz roja sería divulgar sus trabajos en medios accesibles
al colectivo de cazadores, que son quienes deciden sobre las sueltas en su
acotado.
• Evaluación de medidas de gestión alternativas a las sueltas.
Se ha puesto de manifiesto el papel clave del modelo de agricultura
tradicional con alternancia de cultivos sobre la abundancia de perdiz roja, pero
todavía no existen trabajos en los que se actúe experimentalmente sobre los
hábitats tratando de mejorar su potencialidad para la perdiz roja y otras
especies ligadas a los medios agrarios. Resulta triste que, debido a este vacio
de conocimiento científico, la administración apenas sepa qué medidas exigir
para el cumplimiento de la “Directiva Aves” (79/409/CEE del Consejo, de 2 de
abril de 1979) y a la “Directiva Hábitats” (92/43/CEE del Consejo, de 21 de
mayo de 1992) requisito necesario para el cobro de las ayudas europeas. Así
pues, sería muy necesario analizar científicamente la idoneidad de distintas
actuaciones, tales como el mantenimiento de bandas sin cultivar, el uso de
cultivos alternativos o la modificación de alguna práctica agrícola, de cara a
tratar de incluirlas, bien dentro de las medidas obligatorias de “condicionalidad
de la PAC” o bien dentro de las medidas opcionales y subvencionadas por el
plan de desarrollo rural (PDR).

















