
PAPER View Article Online
9
LncRNA MALAT1 aggravates MPP-induced neuronal injury by regulating miR-212 in SH-SY5Y cells Dahua Yuan,† ab Qun Wang,† a Nan Ding, b Pu Du, b Lingmei Peng, b Zhenpeng Duan b and Suyue Pan * a Parkinson's disease (PD) is the most common neurodegenerative disease and its incidence is rising. Long noncoding RNAs (lncRNAs) have been reported to have essential roles in development of PD. LncRNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is dysregulated in PD, while the role of MALAT1 and its mechanism in PD remain poorly understood. In this study, SH-SY5Y cells were exposed to 1-methyl-4-phenylpyridinium (MPP + ) to induce a PD model in vitro. Then we explored the effect of MALAT1 on cell viability, apoptosis and inflammatory response as well as its interaction with miR-212 in MPP + -treated SH-SY5Y cells. The results showed that MALAT1 was up-regulated in MPP + -treated SH- SY5Y cells compared with that in the normal group. Overexpression of MALAT1 exacerbated MPP + - induced neuronal injury, uncovered by inhibition of cell viability and increase of cell apoptosis as well as inflammatory cytokine expressions in SH-SY5Y cells. However, knockdown of MALAT1 exerted the opposite effect in MPP + -treated SH-SY5Y cells. Moreover, MALAT1 was bound to miR-212 and negatively regulated the miR-212 level. Furthermore, addition of miR-212 ablated the regulatory effect of MALAT1 on MPP + -induced neuronal injury, as indicated by restoration of cell viability and lower apoptotic rate along with inflammatory cytokine levels in SH-SY5Y cells. Therefore, we concluded that MALAT1 exacerbated MPP + -induced neuronal injury through regulating cell viability, apoptosis and inflammatory cytokines by sponging miR-212, providing a novel theoretical foundation for application of MALAT1 in PD. Introduction Parkinson's disease (PD) is a common neurological disorder characterized by neuronal injury threatening many elderly patients worldwide. 1 The available evidence has indicated that the inammatory response, oxidative stress and apoptosis are associated with development of PD. 2 Neuroinammation has been reported to contribute to neurodegeneration in many neurological diseases, including PD. 3 With the advance of research, many emerging and alternative therapies have been exploited in treatment of PD. 4 However, it is urgent to explore novel biomarkers of PD for effective strategies of treatment. Non-coding RNAs (ncRNAs), including long ncRNAs (lncRNAs) and microRNAs (miRNAs), have been suggested to have an important impact on the pathogenesis and progression of PD. 5 Emerging evidence has suggested that lncRNAs play essential roles in brain development, neuron function and neurodegenerative diseases. 6 For example, lncRNA small nucleolar RNA host gene 1 (SNHG1) has been reported to promote neuroinammation in PD by targeting the miR-7/NOD- like receptor protein 3 (NLRP3) pathway. 7 Moreover, lncRNA nuclear paraspeckle assembly transcript 1 (NEAT1) has been indicated to regulate autophagy in 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine (MPTP)-induced PD by regulating PTEN- induced kinase 1 (PINK1). 8 Furthermore, a recent study has reported various lncRNAs to be involved in the pathogenesis of neurodegenerative diseases, including lncRNA beta-site amyloid precursor protein-cleaving enzyme 1-antisense tran- script (BACE1-AS), human accelerated region 1 (HAR1) and metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). 9 Moreover, available evidence has indicated that the expressions of some lncRNAs are altered in PD patients, such as lncRNA-p21, MALAT1 and SNHG1. 10 Among those, MALAT1, as a well-known lncRNA, has been reported to be highly associated with numerous diseases and cancers. 11 In addition, earlier work has suggested that MALAT1 may participate in the progression of MPTP-induced PD by regulating a-synuclein protein expres- sion. 12 However, the role of MALAT1 and its potential mecha- nism in PD are not clearly established. Earlier research has indicated miRNAs as promising biomarkers for treatment of neurodegenerative diseases, including PD, through addressing central nervous system homeostasis. 13 a Department of Neurology, Nanfang Hospital, Southern Medical University, 1023-1063, Sha Tai Road, Baiyun District, 510515, Guangzhou, China. E-mail: [email protected]; Tel: +86-020-62787320 b Department of Neurology, The First People's Hospital of Foshan, Foshan, China † The two authors contributed to this work equally. Cite this: RSC Adv. , 2019, 9, 690 Received 9th November 2018 Accepted 13th December 2018 DOI: 10.1039/c8ra09260e rsc.li/rsc-advances 690 | RSC Adv., 2019, 9, 690–698 This journal is © The Royal Society of Chemistry 2019 RSC Advances PAPER Open Access Article. Published on 04 January 2019. Downloaded on 10/22/2021 1:17:09 PM. This article is licensed under a Creative Commons Attribution-NonCommercial 3.0 Unported Licence. View Article Online View Journal | View Issue RETRACTED
Transcript of PAPER View Article Online

RSC Advances
PAPER
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.
View Article OnlineView Journal | View Issue
LncRNA MALAT1
aDepartment of Neurology, Nanfang H
1023-1063, Sha Tai Road, Baiyun Distric
[email protected]; Tel: +86-020-62787320bDepartment of Neurology, The First People'
† The two authors contributed to this wo
Cite this: RSC Adv., 2019, 9, 690
Received 9th November 2018Accepted 13th December 2018
DOI: 10.1039/c8ra09260e
rsc.li/rsc-advances
690 | RSC Adv., 2019, 9, 690–698
aggravates MPP-inducedneuronal injury by regulating miR-212 in SH-SY5Ycells
Dahua Yuan,†ab Qun Wang,†a Nan Ding,b Pu Du,b Lingmei Peng,b Zhenpeng Duanb
and Suyue Pan *a
Parkinson's disease (PD) is the most common neurodegenerative disease and its incidence is rising. Long
noncoding RNAs (lncRNAs) have been reported to have essential roles in development of PD. LncRNA
metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is dysregulated in PD, while the role of
MALAT1 and its mechanism in PD remain poorly understood. In this study, SH-SY5Y cells were exposed
to 1-methyl-4-phenylpyridinium (MPP+) to induce a PD model in vitro. Then we explored the effect of
MALAT1 on cell viability, apoptosis and inflammatory response as well as its interaction with miR-212 in
MPP+-treated SH-SY5Y cells. The results showed that MALAT1 was up-regulated in MPP+-treated SH-
SY5Y cells compared with that in the normal group. Overexpression of MALAT1 exacerbated MPP+-
induced neuronal injury, uncovered by inhibition of cell viability and increase of cell apoptosis as well as
inflammatory cytokine expressions in SH-SY5Y cells. However, knockdown of MALAT1 exerted the
opposite effect in MPP+-treated SH-SY5Y cells. Moreover, MALAT1 was bound to miR-212 and negatively
regulated the miR-212 level. Furthermore, addition of miR-212 ablated the regulatory effect of MALAT1
on MPP+-induced neuronal injury, as indicated by restoration of cell viability and lower apoptotic rate
along with inflammatory cytokine levels in SH-SY5Y cells. Therefore, we concluded that MALAT1
exacerbated MPP+-induced neuronal injury through regulating cell viability, apoptosis and inflammatory
cytokines by sponging miR-212, providing a novel theoretical foundation for application of MALAT1 in PD.
ACTED
Introduction
Parkinson's disease (PD) is a common neurological disordercharacterized by neuronal injury threatening many elderlypatients worldwide.1 The available evidence has indicated thatthe inammatory response, oxidative stress and apoptosis areassociated with development of PD.2 Neuroinammation hasbeen reported to contribute to neurodegeneration in manyneurological diseases, including PD.3 With the advance ofresearch, many emerging and alternative therapies have beenexploited in treatment of PD.4 However, it is urgent to explorenovel biomarkers of PD for effective strategies of treatment.
Non-coding RNAs (ncRNAs), including long ncRNAs(lncRNAs) and microRNAs (miRNAs), have been suggested tohave an important impact on the pathogenesis and progressionof PD.5 Emerging evidence has suggested that lncRNAs playessential roles in brain development, neuron function andneurodegenerative diseases.6 For example, lncRNA smallRETR
ospital, Southern Medical University,
t, 510515, Guangzhou, China. E-mail:
s Hospital of Foshan, Foshan, China
rk equally.
nucleolar RNA host gene 1 (SNHG1) has been reported topromote neuroinammation in PD by targeting themiR-7/NOD-like receptor protein 3 (NLRP3) pathway.7 Moreover, lncRNAnuclear paraspeckle assembly transcript 1 (NEAT1) has beenindicated to regulate autophagy in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced PD by regulating PTEN-induced kinase 1 (PINK1).8 Furthermore, a recent study hasreported various lncRNAs to be involved in the pathogenesis ofneurodegenerative diseases, including lncRNA beta-siteamyloid precursor protein-cleaving enzyme 1-antisense tran-script (BACE1-AS), human accelerated region 1 (HAR1) andmetastasis-associated lung adenocarcinoma transcript 1(MALAT1).9 Moreover, available evidence has indicated that theexpressions of some lncRNAs are altered in PD patients, such aslncRNA-p21, MALAT1 and SNHG1.10 Among those, MALAT1, asa well-known lncRNA, has been reported to be highly associatedwith numerous diseases and cancers.11 In addition, earlier workhas suggested that MALAT1 may participate in the progressionof MPTP-induced PD by regulating a-synuclein protein expres-sion.12 However, the role of MALAT1 and its potential mecha-nism in PD are not clearly established.
Earlier research has indicated miRNAs as promisingbiomarkers for treatment of neurodegenerative diseases, includingPD, through addressing central nervous system homeostasis.13
This journal is © The Royal Society of Chemistry 2019

Paper RSC Advances
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
miR-212, as a novel miRNA, has been suggested to be necessary forthe development and function of neurons.14 Moreover, miR-212 isreported to be expressed in cerebrospinal uid in PD patients.15
Additionally, such work has indicated that miR-212 can alleviate 1-methyl-4-phenylpyridinium (MPP+)-induced neuronal damage inSH-SY5Y cells by regulating Kruppel-like factor 4 (KLF4).16
However, little is known about the exact role of miR-212 in PD.Intriguingly, bioinformatics analysis provides the putative bindingsites ofmiR-212 andMALAT1. However, there is no direct evidencein support of this prediction. Hence, we hypothesized thatMALAT1 might regulate the progression of PD by regulating miR-212. In the present study, we used MPP+-treated SH-SY5Y cells asa PD model in vitro to explore the roles of MALAT1 in neuronalinjury (cell viability, apoptosis and inammation) and probed theinteraction between MALAT1 and miR-212.
Materials and methodsCell culture and treatment
Human neuroblastoma SH-SY5Y cells were obtained from theAmerican Tissue Culture Collection (ATCC, Manassas, VA, USA)and cultured using Roswell Park Memorial Institute (RMPI)-1640 culture medium (Gibco, Carlsbad, CA, USA) containing10% fetal bovine serum (FBS, Gibco) and 1% penicillin–strep-tomycin (Invitrogen, Carlsbad, CA, USA) at 37 �C in 5% CO2
during the study. To establish a PDmodel in vitro, SH-SY5Y cellswere exposed to various concentrations (0, 0.25, 0.5, 1 and 2mM) of MPP+ (Sigma, St. Louis, MO, USA) for 24 h or 1 mM ofMPP+ for different treatment times (0, 6, 12, 24 and 48 h). Toexplore the regulatory mechanism of MALAT1, transfected SH-SY5Y cells were treated with 1 mM MPP+ for 24 h.
The MALAT1-overexpression vector (MALAT1), empty vector(pcDNA), siRNA for MALAT1 (si-MALAT1), negative control (si-NC), miR-212 mimic (miR-212), miR-NC, miR-212 inhibitor(anti-miR-212) and anti-miR-NC were obtained from Gene-pharma (Shanghai, China). Transient transfection with theoligonucleotides or plasmids in SH-SY5Y cells was conductedusing Lipofectamine 2000 (Invitrogen).
R
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was isolated from treated SH-SY5Y cells using Trizolreagent (Invitrogen) following the manufacturer's instructions.Aer being quantied using a NanoDrop Spectrophotometer(NanoDrop, Wilmington, DE, USA), 500 ng of total RNA was usedfor complementary DNA (cDNA) synthesis by a TaqMan ReverseTranscription Kit or TaqMan microRNA Reverse TranscriptionKit (Applied Biosystems, Foster City, CA, USA). Subsequently,cDNA was diluted and used for qRT-PCR using an SYBR greendetection kit (Toyobo, Tokyo, Japan) following the amplicationinstructions in an ABI 7500 real time PCR system (Applied Bio-systems). The relative expressions of MALAT1 and miR-212 wereevaluated with the 2�DDCt method using b-actin or U6 small RNAas housekeeping genes, respectively. All primers were obtainedfrom Sangon Biotech (Shanghai, China): MALAT1 (forward, 50-AGCGGAAGAACGAATGTAAC-30; reverse, 50-GAACAGAAGGAA-GAGCCAAG-30), b-actin (forward, 50-TGAGCGCGGCTACAGCTT-
RET
This journal is © The Royal Society of Chemistry 2019
30; reverse, 50-TCCTTAATGTC ACGCACGATTT-30), miR-212(forward, 50-CCCTCTGGGACATCTTTGACG-30; reverse, 50-TGCTCCGCCTCCCCTGCGTCTC-30), U6 (forward, 50-GCTTCGGCA GCACATATACTAAAAT-30; reverse, 50-CGCTTCAC-GAATTTGCGTGTCAT-30).
Cell viability
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide(MTT) assay was performed to analyze cell viability. SH-SY5Ycells were seeded into 96-well plates at a density of 1 � 104
cells per well. At the termination time, MTT (Sigma) was addedinto the cells and incubated for 4 h at 37 �C. Subsequently,dimethyl sulfoxide (DMSO, Sigma) was administrated to thecells to dissolve formazan. The absorbance was measured at570 nm with a microplate reader (Bio-Rad, Hercules, CA, USA).ED
Cell apoptosis
Cell apoptosis was measured by ow cytometry through anAnnexin V-FITC/propidium iodide (PI) apoptosis detection kit(Sigma) according to the manufacturer's protocols. Briey,treated SH-SY5Y cells were collected and incubated withAnnexin V-FITC and PI for 20 min in the dark aer beingwashed with PBS. The positive cells were examined using a owcytometer (BD Biosciences, Franklin Lakes, NJ, USA).CT
Enzyme linked immunosorbent assay (ELISA)Aer treatments, the cell culture medium was collected from the24-well plates and the levels of interleukin-1b (IL-1b), IL-6 andtumor necrosis factor-a (TNF-a) were quantied using the cor-responding ELISA Kit (Invitrogen) referring to themanufacturer'sinstructions. The intensity of color was assayed with a microplatereader at 450 nm with reference wavelength at 620 nm.
A
Luciferase assaysThe online soware starBase was used to predict the putativebinding sites of miR-212 and MALAT1. The 30 untranslatedregions (30-UTR) sequences of MALAT1-containing wild-type ormutant binding sites of miR-212 were amplied and cloned intopmirGlO luciferase reporter vector (Promega, Madison, WI,USA) to generate the wild-type plasmids (MALAT1-WT) ormutant-type plasmids (MALAT1-MUT), respectively. WT or MUTluciferase reporter plasmids and miR-212 or miR-NC were co-transfected into SH-SY5Y cells using Lipofectamine 2000according to the manufacturer's protocols. Then the lysed cellswere subjected to luciferase activity analysis using a Dual-Luciferase Assay Kit (Promega) aer transfection for 48 h.
RNA pull-down assay
RNA pull-down analysis was conducted to probe the interactionbetween MALAT1 and miR-212 using an RNA-Protein Pull DownKit (Thermo Fisher, Wilmington, DE, USA). miR-NC, miR-212 ormiR-212-MUT without complementary sites of MALAT1 waslabelled with biotin and transfected into SH-SY5Y cells. The celllysates were incubated with streptavidin agarose beads (Invitrogen)
RSC Adv., 2019, 9, 690–698 | 691

RSC Advances Paper
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
for 2 h. Aer being eluted with biotin elution buffer, the complexwas used for measurement of MALAT1 abundance by qRT-PCR.
RNA immunoprecipitation (RIP)
SH-SY5Y cells were transfected with miR-212 or miR-NC andthen Argonaute 2 (Ago2) RNA immunoprecipitation (RIP) wasused to probe the link between miR-212 and MALAT1 usinga Magna RIP Kit (Millipore, Billerica, MA, USA) following themanufacturer's protocol. In brief, transfected SH-SY5Y cellswere lysed and then added to magnetic beads (Thermo Fisher)bound with anti-Ago2 or IgG. Following washing with PBS, theRNA in the bead complexes was isolated and detected byagarose electrophoresis and qRT-PCR.
Statistical analysis
All data were presented as the mean � standard deviation (SD)from three independent experiments. The statistical differenceswere evaluated by Student's t test or one-way ANOVA using SPSS18.0 soware (SPSS, Inc., Chicago, IL, USA). Statistical signi-cance was regarded as *p < 0.05, **p < 0.01 or ***p < 0.001.
ResultsMALAT1 expression was enhanced in MPP+-treated SH-SY5Ycells
To investigate the potential roles of MALAT1 in PD, theexpression of MALAT1 was rst measured in MPP+-treated SH-SY5Y cells. MPP+ is a positively charged neurotoxin and itschemical structure is shown in Fig. 1A. SH-SY5Y cells weretreated with 0.25, 0.5, 1 and 2 mM of MPP+ for 24 h. The resultsshowed that the exposure to MPP+ led to a progressive increaseof the MALAT1 level in SH-SY5Y cells in a concentrationdependent manner (Fig. 1B). Additionally, SH-SY5Y cells wereexposed to 1 mM of MPP+ for 6, 12, 24 and 48 h. Elevatedabundance of MALAT1 was displayed in the MPP+-treated SH-SY5Y cells in a time dependent manner (Fig. 1C). Hence, SH-SY5Y cells were treated with 1 mM of MPP+ for 24 h in furtherexperiments.
R
Fig. 1 MALAT1 expression was enhanced in MPP+-treated SH-SY5Y cells.measured by qRT-PCR in SH-SY5Y cells after treatment with different concby qRT-PCR in 1 mM MPP+-treated SH-SY5Y cells at different treatment t
692 | RSC Adv., 2019, 9, 690–698
RET
MALAT1 exacerbated MPP+-mediated regulatory effect on cellviability and apoptosis in SH-SY5Y cellsTo explore whether MALAT1 regulated neuronal injury in PD,cell viability and apoptosis were detected in SH-SY5Y cellstransfected with pcDNA, MALAT1, si-NC or si-MALAT1 aertreatment withMPP+. The abundance of MALAT1 was effectivelyenhanced in SH-SY5Y cells transfected with MALAT1 over-expression vector, whereas its expression was inhibited in cellstransfected with si-MALAT1, compared with their correspond-ing controls (Fig. 2A). Moreover, treatment with MPP+ inhibitedcell viability, which was exacerbated by overexpression ofMALAT1 and attenuated by knockdown of MALAT1 in SH-SY5Ycells compared with their corresponding controls (Fig. 2B). Inaddition, exposure to MPP+ promoted cell apoptosis, andoverexpression of MALAT1 exacerbated MPP+-inducedapoptosis, while knockdown of MALAT1 alleviated the pro-apoptosis effect of MPP+ in SH-SY5Y cells compared with theircorresponding controls (Fig. 2C and D).
MALAT1 exacerbated MPP+-induced expressions ofinammatory cytokines in SH-SY5Y cells
Seeing that the inammatory response is also associated withneuronal injury in PD, the levels of inammatory cytokines weredetected in SH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MALAT1 aer treatment with MPP+. The resultsshowed that treatment with MPP+ resulted in a signicantenhancement in the levels of IL-1b (Fig. 3A), IL-6 (Fig. 3B) andTNF-a (Fig. 3C) in SH-SY5Y cells. Moreover, the pro-inammatoryeffect of MPP+ was exacerbated by accumulation of MALAT1 andwas counteracted by interference of MALAT1 (Fig. 3).
MALAT1 was bound to miR-212 in SH-SY5Y cells
To investigate the underlying mechanism by which MALAT1causes neuronal injury in PD, starBase soware was used toexplore potential miRNAs modulated by MALAT1 as a competingendogenous RNA (ceRNA). Bioinformatics analysis predicted theputative binding sites of miR-212 and MALAT1, suggesting thepossibility of MALAT1 sponging miR-212 (Fig. 4A). To validate theprediction of the interaction between MALAT1 and miR-212,
ACTED
(A) The chemical structure of MPP+. (B) The expression of MALAT1 wasentrations of MPP+ for 24 h. (C) The abundance of MALAT1 was detectedimes. *p < 0.05, **p < 0.01 and ***p < 0.001.
This journal is © The Royal Society of Chemistry 2019

Fig. 2 MALAT1 exacerbated MPP+-mediated regulatory effect on cell viability and apoptosis in SH-SY5Y cells. (A) The abundance of MALAT1 wasdetected by qRT-PCR in SH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MALAT1. (B) Cell viability was measured by MTT in SH-SY5Ycells transfected with pcDNA, MALAT1, si-NC or si-MALAT1 after treatment with MPP+. (C and D) Cell apoptosis was examined by flow cytometryin SH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MALAT1 after treatment with MPP+. **p < 0.01 and ***p < 0.001.
Paper RSC Advances
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
ACTED
luciferase activity analysis, RNA pull-down assay and RIP wereestablished in SH-SY5Y cells. The results indicated that over-expression of miR-212 led to a signicant reduction of luciferaseactivity in SH-SY5Y cells transfected with MALAT1-WT comparedwith treatment with miR-NC, whereas the efficacy of miR-212 waslost in response to MALAT1-MUT (Fig. 4B). Moreover, biotinylatedTR
Fig. 3 MALAT1 exacerbated MPP+-induced production of inflammatory cin SH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MALAT1 aSH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MALAT1 afELISA in SH-SY5Y cells transfected with pcDNA, MALAT1, si-NC or si-MA
This journal is © The Royal Society of Chemistry 2019
RE
miR-212 indicated a higher capacity for enrichment of MALAT1compared with the bio-NC group, while biotinylated miR-212-MUT showed little efficacy (Fig. 4C). In addition, agarose electro-phoresis indicated the enrichment of MALAT1 and miR-212 inextracts produced by RIP assay, and accumulation of miR-212enhanced the abundance of MALAT1 in products enriched in
ytokines in SH-SY5Y cells. (A) The level of IL-1bwas measured by ELISAfter treatment of MPP+. (B) The level of IL-6 was detected by ELISA inter exposure to MPP+. (C) The expression of TNF-a was examined byLAT1 after stimulation of MPP+. **p < 0.01 and ***p < 0.001.
RSC Adv., 2019, 9, 690–698 | 693

Fig. 4 MALAT1 was bound tomiR-212 in SH-SY5Y cells. (A) The putative binding sites of miR-212 andMALAT1 were predicted by starBase online.(B) Luciferase activity was measured in SH-SY5Y cells co-transfected with MALAT1-WT or MALAT1-MUT and miR-212 or miR-NC. (C) RNA pull-down assay was conducted in SH-SY5Y cells transfected with bio-NC, bio-miR-212-WT or bio-miR-212-MUT and the level of MALAT1 wasmeasured by qRT-PCR. (D) Ago2 RIP assay was performed in SH-SY5Y cells transfected with miR-212 or miR-NC and the abundance of MALAT1was detected by agarose electrophoresis and qRT-PCR. (E) The expression of miR-212 was measured by qRT-PCR in SH-SY5Y cells transfectedwith pcDNA, MALAT1, si-NC or si-MALAT1. ***p < 0.001.
RSC Advances Paper
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
ETRACTE
D
anti-Ago2, while IgG showed little efficacy of enrichment (Fig. 4D).Additionally, the expression of miR-212 was detected in SH-SY5Ycells transfected with pcDNA, MALAT1, si-NC or si-MALAT1. Theresults showed that accumulation of MALAT1 inhibited the miR-212 level, while abrogation of MALAT1 increased miR-212expression in SH-SY5Y cells (Fig. 4E).R
694 | RSC Adv., 2019, 9, 690–698
Overexpression of miR-212 reversed the regulatory effect ofMALAT1 on cell viability and apoptosis in MPP+-treated SH-SY5Y cells
To evaluate whether the regulatory effect of MALAT1 wasmediated by miR-212, SH-SY5Y cells were transfected withMALAT1 + miR-NC, MALAT1 + miR-212, si-MALAT1 + anti-
This journal is © The Royal Society of Chemistry 2019

Fig. 5 Overexpression of miR-212 reversed the regulatory effect of MALAT1 on cell viability and apoptosis in MPP+-treated SH-SY5Y cells. (A)Cell viability wasmeasured byMTT in SH-SY5Y cells transfectedwithMALAT1 +miR-NCorMALAT1 +miR-212 after treatment withMPP+. (B) Cellapoptosis was detected by flow cytometry in SH-SY5Y cells transfected with MALAT1 + miR-NC or MALAT1 + miR-212 after treatment withMPP+. (C) Cell viability was measured by MTT in SH-SY5Y cells transfected with si-MALAT1 + anti-miR-NC or si-MALAT1 + anti-miR-212 afterstimulation of MPP+. (D) Cell apoptosis was detected by flow cytometry in SH-SY5Y cells transfected with si-MALAT1 + anti-miR-NC or si-MALAT1 + anti-miR-212 after exposure to MPP+. **p < 0.01 and ***p < 0.001.
Paper RSC Advances
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
ACTED
miR-NC or si-MALAT1 + anti-miR-212 prior to MPP+ treatment.The results showed that overexpression of miR-212 reversedthe MALAT1-mediated inhibition of viability in MPP+-inducedSH-SY5Y cells (Fig. 5A). Moreover, the MALAT1-mediatedincrease of apoptosis was ablated by restoration of miR-212in MPP+-induced SH-SY5Y cells (Fig. 5B). In addition, theabrogation of miR-212 counteracted the effect of inhibition ofMALAT1 on cell viability in MPP+-induced SH-SY5Y cells(Fig. 5C). Furthermore, the inhibitory effect of MALAT1knockdown onMPP+-induced apoptosis was attenuated byexhaustion of miR-212 (Fig. 5D).
TR
Addition of miR-212 attenuated MALAT1-promoted secretionof inammatory cytokines in MPP+-treated SH-SY5Y cells
To identify whether miR-212 was required for MALAT1-mediated inammatory damage in PD, the secretions ofinammatory cytokines were measured in MPP+-treated SH-SY5Y cells. The data revealed that the facilitating role ofMALAT1 in MPP+-induced expressions of IL-1b (Fig. 6A), IL-6(Fig. 6C) and TNF-a (Fig. 6E) was ablated by addition of miR-212 in SH-SY5Y cells compared with the correspondingcontrol. Moreover, the inhibitory effect of MALAT1 interferenceon MPP+-mediated productions of IL-1b (Fig. 6B), IL-6 (Fig. 6D)
RE
This journal is © The Royal Society of Chemistry 2019
and TNF-a (Fig. 6F) was recovered by knockdown of miR-212 inSH-SY5Y cells.
Discussion
PD is a common neurodegenerative disorder which wreakshavoc on human health worldwide.17 To date, a number ofinvestigators have reported that the inammatory response isassociated with neurodegeneration in PD.18 Pro-inammatorycytokines including IL-1b, IL-6 and TNF-a have been indicatedto aggravate inammatory injury in PD.19 SH-SY5Y cells havebeen popular in PD models in vitro because of the interaction ofthese neurons with dopamine.20 Moreover, MPP+, as a neuro-toxin, has been widely used to establish in vitro PD cell models.21
In the present study, we also developed MPP+-treated SH-SY5Ycells as a PD model in vitro. We provided the rst insightsinto the roles of MALAT1 in inammatory injury in PD and theinteraction between MALAT1 and miR-212.
The available evidence has indicated that lncRNAs haveessential roles in the development and pathological processesof PD through regulating chromatin, protein and some RNAs.22
MALAT1 has been reported to be up-regulated in PD, and itsknockdown was found to protect cell viability and inhibit
RSC Adv., 2019, 9, 690–698 | 695
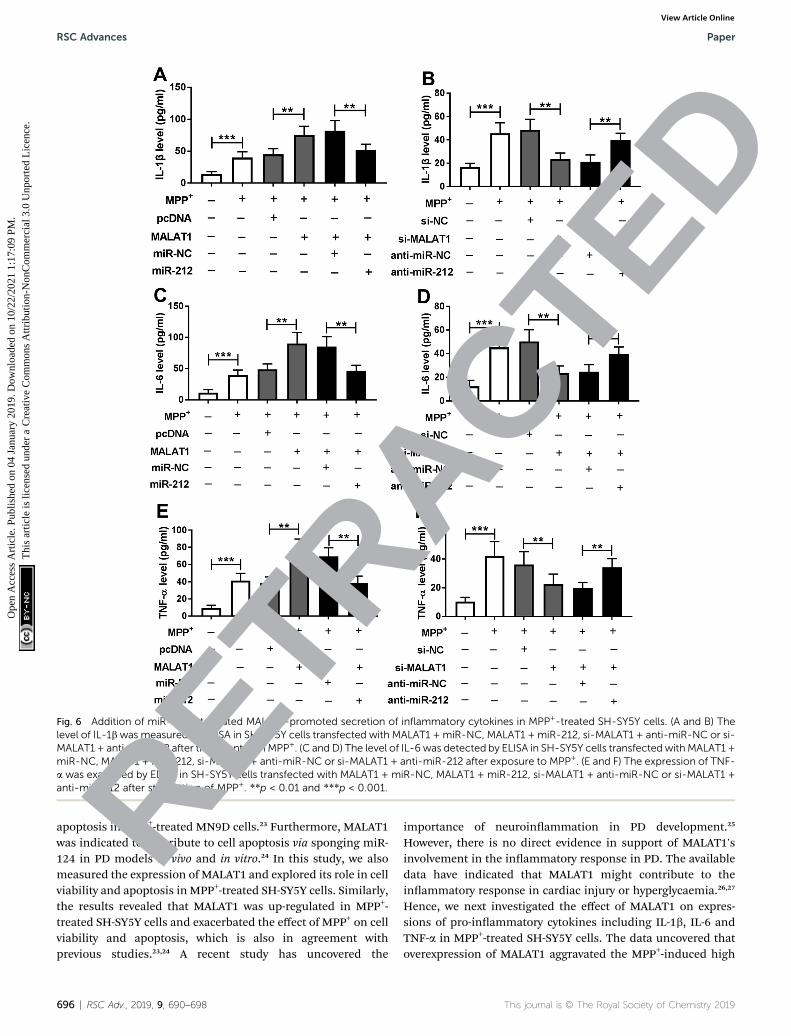
Fig. 6 Addition of miR-212 attenuated MALAT1-promoted secretion of inflammatory cytokines in MPP+-treated SH-SY5Y cells. (A and B) Thelevel of IL-1bwas measured by ELISA in SH-SY5Y cells transfected with MALAT1 + miR-NC, MALAT1 + miR-212, si-MALAT1 + anti-miR-NC or si-MALAT1 + anti-miR-212 after treatment withMPP+. (C and D) The level of IL-6 was detected by ELISA in SH-SY5Y cells transfected withMALAT1 +miR-NC, MALAT1 + miR-212, si-MALAT1 + anti-miR-NC or si-MALAT1 + anti-miR-212 after exposure to MPP+. (E and F) The expression of TNF-a was examined by ELISA in SH-SY5Y cells transfected with MALAT1 + miR-NC, MALAT1 + miR-212, si-MALAT1 + anti-miR-NC or si-MALAT1 +anti-miR-212 after stimulation of MPP+. **p < 0.01 and ***p < 0.001.
RSC Advances Paper
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
ETRACTE
D
apoptosis in MPP+-treated MN9D cells.23 Furthermore, MALAT1was indicated to contribute to cell apoptosis via sponging miR-124 in PD models in vivo and in vitro.24 In this study, we alsomeasured the expression of MALAT1 and explored its role in cellviability and apoptosis in MPP+-treated SH-SY5Y cells. Similarly,the results revealed that MALAT1 was up-regulated in MPP+-treated SH-SY5Y cells and exacerbated the effect of MPP+ on cellviability and apoptosis, which is also in agreement withprevious studies.23,24 A recent study has uncovered theR
696 | RSC Adv., 2019, 9, 690–698
importance of neuroinammation in PD development.25
However, there is no direct evidence in support of MALAT1'sinvolvement in the inammatory response in PD. The availabledata have indicated that MALAT1 might contribute to theinammatory response in cardiac injury or hyperglycaemia.26,27
Hence, we next investigated the effect of MALAT1 on expres-sions of pro-inammatory cytokines including IL-1b, IL-6 andTNF-a in MPP+-treated SH-SY5Y cells. The data uncovered thatoverexpression of MALAT1 aggravated the MPP+-induced high
This journal is © The Royal Society of Chemistry 2019

Paper RSC Advances
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
levels of IL-1b, IL-6 and TNF-a in SH-SY5Y cells. However, thepotential mechanism that underlies MALAT1's regulation of cellviability, apoptosis and inammatory response remains poorlyunderstood. LncRNAs have been shown to act as miRNAsponges to regulate the abundances and activities of boundmiRNAs.28 Several such reports have revealed MALAT1 asa ceRNA of some miRNAs in PD models in vitro. For instance,the MALAT1/miR-129 axis regulated a PD-like phenotype bycontrolling apoptosis of neurons.29 Moreover, the MALAT1/miR-205-5p axis mediated MPP+-induced apoptosis in a PD cellmodel.23 Furthermore, miR-124 was also reported to be boundto MALAT1, which regulated cell apoptosis in MPP+-treated SH-SY5Y cells.24 Hence, the novel concept of miRNA spongingpromises to contribute to a better understanding of theunderlying mechanism. Here we validated miR-212 as a targetof MALAT1 in SH-SY5Y cells for the rst time, by luciferaseactivity, RNA-pull down and RIP assays.
miR-212, as a novel mi-RNA, has been reported to be asso-ciated with neurogenesis and neuroinammation in mice.30
Furthermore, miR-212 was suggested to support neural viabilityin neurodegenerative disorders like Alzheimer's disease.31
Notably, it has been indicated that miR-212 attenuated MPP+-induced neuronal damage in SH-SY5Y cells, as indicated by anincrease of cell viability and decrease of apoptosis and inam-matory response.16 These ndings suggested that miR-212might serve as a protective biomarker during neuronal injury.To establish whether miR-212 played a protective role in PDprogression, we explored the roles of miR-212 in MPP+-treatedSH-SY5Y cells. The restoration of miR-212 weakened the regu-latory effect of MALAT1 on MPP+-mediated neuronal injury inSH-SY5Y cells, which indicated that MALAT1 facilitated PDprogression in our MPP+-induced model in vitro by spongingmiR-212. This novel signaling network of MALAT1/miR-212 mayprovide a promising theoretical foundation for application ofMALAT1 in PD.
In addition, earlier studies demonstrated that miR-212 dis-played an inhibitive effect in neurological damage by targetingKLF4 or sirtuin 2 (SIRT2).16,32 Moreover, the Wnt/b-catenin andmitogen-activated protein kinase (MAPK) pathways have beenreported to play important roles in PD.33,34 Notably, a number ofinvestigators have reported the interaction between MALAT1and these signaling processes in many conditions.35,36 However,none of these were observed in this study. Thus, it is necessaryto explore possible target mRNAs of miR-212 and establish a PDmodel in vivo in future. Furthermore, the potential signalingpathway should be explored in further studies.
Conclusion
In conclusion, MALAT1 expression was enhanced in MPP+-treated SH-SY5Y cells and its up-regulation exacerbated MPP+-induced reduction of cell viability and enhancement of cellapoptosis as well as inammatory injury in SH-SY5Y cells.Moreover, MALAT1 can be regarded as a decoy of miR-212.Furthermore, addition of miR-212 alleviated the promotiveeffect of MALAT1 on MPP+-induced neuronal injury in SH-SY5Ycells. Collectively, our study suggested that MALAT1
RETR
This journal is © The Royal Society of Chemistry 2019
exacerbated MPP+-induced neuronal injury by sponging miR-212 in SH-SY5Y cells, indicating the potential role of MALAT1as a biomarker for PD treatment.
Conflicts of interest
The authors have no conicts of interest to declare.
References
1 L. Kalia and A. Lang, Lancet, 2015, 386, 896–912.2 G. Andican, D. Konukoglu, M. Bozluolcay, K. Bayulkem,S. Firtiına and G. Burcak, Acta Neurol. Belg., 2012, 112,155–159.
3 R. Ransohoff, Science, 2016, 353, 777–783.4 A. Kabra, R. Sharma, R. Kabra and U. Baghel, Curr. Pharm.Des., 2018, 24(22), 2573–2582.
5 M. Majidinia, A. Mihanfar, R. Rahbarghazi, A. Nourazarian,B. Bagca and Ç. Avci, Mol. Biol. Rep., 2016, 43, 1193–1204.
6 P. Wu, X. Zuo, H. Deng, X. Liu, L. Liu and A. Ji, Brain Res.Bull., 2013, 97, 69–80.
7 B. Cao, T. Wang, Q. Qu, T. Kang and Q. Yang, Neuroscience,2018, 388, 118–127.
8 W. Yan, Z. Chen, J. Chen and H. Chen, Biochem. Biophys. Res.Commun., 2018, 496, 1019–1024.
9 P. Riva, A. Ratti and M. Venturin, Curr. Alzheimer Res., 2016,13(11), 1219–1231.
10 T. Kraus, M. Haider, J. Spanner, M. Steinmaurer, V. Dietingerand H. Kretzschmar, Mol. Neurobiol., 2017, 54, 2869–2877.
11 M. Zhao, S. Wang, Q. Li, Q. Ji, P. Guo and X. Liu, Oncol. Lett.,2018, 16, 19–26.
12 Q. Zhang, Z. Wang, J. Zhang, Y. Duan, G. Li and D. Zheng,Biomed. Pharmacother., 2016, 83, 153–159.
13 D. Li, Y. Li, Y. Li, X. Zhu, X. Du, M. Zhou, W. Li and H. Deng,China Med. J., 2018, 131, 2216–2225.
14 A. Wanet, A. Tacheny, T. Arnould and P. Renard, NucleicAcids Res., 2012, 40, 4742–4753.
15 K. Burgos, I. Malenica, R. Metpally, A. Courtright, B. Rakela,T. Beach, H. Shill, C. Adler, M. Sabbagh, S. Villa, W. Tembe,D. Craig and K. Van Keuren-Jensen, PLoS One, 2014, 9,e94839.
16 Y. Song, Y. Liu and X. Chen, Yonsei Med. J., 2018, 59, 416–424.
17 S. Przedborski, Nat. Rev. Neurosci., 2017, 18, 251–259.18 V. Calabrese, A. Santoro, D. Monti, R. Crupi, R. Di Paola,
S. Latteri, S. Cuzzocrea, M. Zappia, J. Giordano,E. Calabrese and C. Franceschi, Free Radical Biol. Med.,2018, 115, 80–91.
19 K. Kaur, J. Gill, P. Bansal and R. Deshmukh, J. Neurol. Sci.,2017, 381, 308–314.
20 H. Xie, L. Hu and G. Li, China Med. J., 2010, 123, 1086–1092.21 D. Hare, P. Adlard, P. Doble and D. Finkelstein, Metallomics,
2013, 5, 91–109.22 D. Wang, P. Fu, C. Yao, L. Zhu, T. Hou, J. Chen, Y. Lu, D. Liu
and L. Zhu, Mol. Ther.–Nucleic Acids, 2018, 10, 269–276.23 Q. Chen, X. Huang and R. Li, Am. J. Transl. Res., 2018, 10,
563–572.
ACTED
RSC Adv., 2019, 9, 690–698 | 697

RSC Advances Paper
Ope
n A
cces
s A
rtic
le. P
ublis
hed
on 0
4 Ja
nuar
y 20
19. D
ownl
oade
d on
10/
22/2
021
1:17
:09
PM.
Thi
s ar
ticle
is li
cens
ed u
nder
a C
reat
ive
Com
mon
s A
ttrib
utio
n-N
onC
omm
erci
al 3
.0 U
npor
ted
Lic
ence
.View Article Online
E
24 W. Liu, Q. Zhang, J. Zhang, W. Pan, J. Zhao and Y. Xu, CellBiosci., 2017, 7, 19.
25 R. Niranjan, Neurochem. Int., 2018, 120, 13–20.26 H. Chen, X. Wang, X. Yan, X. Cheng, X. He andW. Zheng, Int.
Immunopharmacol., 2018, 55, 69–76.27 P. Puthanveetil, S. Chen, B. Feng, A. Gautam and
S. Chakrabarti, J. Cell. Mol. Med., 2015, 19, 1418–1425.28 L. Salmena, L. Poliseno, Y. Tay, L. Kats and P. Pandol, Cell,
2011, 146, 353–358.29 D. Xia, R. Sui and Z. Zhang, J. Cell. Biochem., 2018, DOI:
10.1002/jcb.27769.30 S. Kempf, A. Casciati, S. Buratovic, D. Janik, C. von Toerne,
M. Ueffing, F. Neff, S. Moertl, B. Stenerlow, A. Saran,M. Atkinson, P. Eriksson, S. Pazzaglia and S. Tapio, Mol.Neurodegener., 2014, 9, 57.
698 | RSC Adv., 2019, 9, 690–698
RETR
31 Y. Wang, T. Veremeyko, A. Wong, R. El Fatimy, Z. Wei, W. Caiand A. Krichevsky, Neurobiol. Aging, 2017, 51, 156–166.
32 S. Sun, X. Han, X. Li, Q. Song, M. Lu, M. Jia, J. Ding andG. Hu, Front. Mol. Neurosci., 2018, 11, 381.
33 B. Marchetti, Int. J. Mol. Sci., 2018, 19, E3743.34 J. Yang, M. Jia, X. Zhang and P. Wang, Phytother. Res., 2018,
DOI: 10.1002/ptr.6221.35 C. Guo, X. Wang, L. P. Chen, M. Li, M. LI, Y. H. Hu,
W. H. Ding and X. Wang, Eur Rev. Med. Pharmacol. Sci.,2018, 22, 3703–3712.
36 Y. Li, Y. D. Liu, S. L. Chen, X. Cehn, D. S. Ye, X. Y. Zhou, J. Zheand J. Zhang,Mol. Hum. Reprod., 2018, DOI: 10.1093/molehr/gay045.
D
This journal is © The Royal Society of Chemistry 2019
ACT


















