
PAPER 2 Cell Evolution
7
1 2 3 4 5 6 7 8 9 10 11 12 13 14 Glu Asp Asn Thr His Aromatic Trp Gln Ala Ser Pro Val Polar uncharged Ile Met Leu Non-polar hydrophobic Lys Phospholipids Arg Basic Phe Tyr Gly Cys Acidic RNA Mineral surface chemistry Vesicles Self-synthesizing membrane RNA-DNA DNA Evolutionary time Genomic analysis leaves no doubt that the three main kingdoms of life — eubacteria, archaea and eukaryotes — evolved from a common ancestor 1 . The idea that all life evolved from one source is perhaps the most profound consequence of the Darwin–Wallace model of evolution. With increasing amounts of DNA sequence information available, the search is on to find the minimum set of genes that existed in the last common ancestor (LCA) — the precursor cell(s) from which all living organisms subsequently evolved. The LCA must have been a rather sophisticated cell because it contained all the cellular machin- ery that is common to all present living forms, including a DNA-based information system 2,3 . From sequence analysis, the LCA is suggested to contain 250–600 genes 4,5 . Mycoplasma genitalium, the simplest known prokaryo te cell (which is nevertheless dependent on host-cell parasitism), requires 270 of its 380 genes for normal function, as revealed when all genes were individually knocked out 6 . So, the minimum conceivable cell needs a lot of genetic information. The Earth is ~4.56 billion years (Gyr) old, and the best current estimates argue that the first cells appeared by 3.0–3.3 Gyr at the latest 7 . How these first cells emerged is one of the biggest unsolved problems in biology. It seems undeniable that the LCA must have been preceded by a spontaneous generation of cells from abiotic precursor molecules. Even Darwin in his theory of evolution was obliged to accept the necessity of some kind of spontaneous generation. He brilliantly speculated (in a letter to Hooker) that in a “warm little pond” a particular chemistry involving “all sorts of ammonia and phosphoric salts, light, heat, electricity etc. was present so that a protein compound was chemically formed ready to undergo more complex changes on the path towards life” 8 . It seems likely that his (unpublished) hypothesis was not far from the mark. Modern theories were initiated by Oparin in 1924 (REF. 9) and by Haldane 10 , who first discussed the origin of membranes and speculated that an ‘oily film’ on the sur- face of sea water evolved into the lipid-rich cell membrane. Here, I attempt to build up a plausible scenario for how cells may have evolved — with an emphasis on the more difficult question of the origin of the cell membrane. A brief discussion of the (more actively pur- sued) question of the evolution of cytoplasm, including the nucleic acids, sets the stage for the main hypothesis. The evolution of the cytoplasm Cells are made up of nucleic acids, proteins, carbohydrates, lipids and many small molecules suspended in a particular ionic OPINION Cell evolution and the problem of membrane topology Gareth Griffiths Abstract | Cells somehow evolved from primordial chemistry and their emergence depended on the co-evolution of the cytoplasm, a genetic system and the cell membrane. It is widely believed that the cytoplasm evolved inside a primordial lipid vesicle, but here I argue that the earliest cytoplasm could have co-evolved to high complexity outside a vesicle on the membrane surface. An invagination of the membrane, aided by an early cytoskeletal system, may have formed the first cells — initially within p rimordial vesicles. Figure 1 | Biosynthetic pathways for amino acids, phospholipids and central metabolism. It has been proposed by Davis 22,26 and many others that primitive biochemical pathways exi sted before a genetic-based system. These include the reductive carbon cycle (the equivalent of the tricarboxylic acid (TCA) cycle working in reverse), the reductive pentose pathway and the central trunk (proposed to be a remnant of the formose cycle), which together make up the central biochemical pathway (CBP). Davis proposes that the emergence of genetically encoded amino acids correlates with the number of chemical reactions from the CBP that are required to generate each amino acid (evolutionary steps 1–14). Acidic amino acids (Glu and Asp) are close to the CBP (1–2 reactions), whereas aromatic residues (such as Trp) require up to 14 steps to be synthesized and may have appeared later. The hydrophobic amino acids that could associate with membranes required four steps. The synthesis of phospholipids requires 10 steps and, therefore, self-synthesizing membranes might only have arisen after this point. Davis provides detailed analysis to argue that the evolutionary appearance of the different amino acids correlates well with the emergence of their corresponding triplet codes; this implies co-evolution of biochemistry and the genetic code, an idea extensively championed by Wong 60 . Davis also identified an 11-amino acid sequence in the FtsZ–tubulin family that he mapped to his evolutionary stage 7.5, which could support a role for this protein in cellularization. Gly, Cys and Pro cannot easily be placed into any of the five categories shown. 1018 | DECEMBER 2007 | VOLUME 8 www.nature.com/reviews/molcellbio PERSPECTIVES © 2007 Nature Publishing Group
-
Upload
lorena-nacif-marcal -
Category
Documents
-
view
220 -
download
0
Transcript of PAPER 2 Cell Evolution

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 1/7
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Glu
Asp
Asn Thr His
Aromatic
Trp
Gln
Ala
Ser
Pro
Val
Polar uncharged
Ile
Met
Leu
Non-polarhydrophobic
Lys
Phospholipids
Arg
Basic
PheTyr
Gly
Cys
Acidic
RNA
Mineralsurfacechemistry
Vesicles Self-synthesizingmembrane
RNA-DNA DNAEvolutionary time
Genomic analysis leaves no doubt that thethree main kingdoms of life — eubacteria,archaea and eukaryotes — evolved froma common ancestor1. The idea that alllife evolved from one source is perhapsthe most profound consequence of theDarwin–Wallace model of evolution. With
increasing amounts of DNA sequenceinformation available, the search is on tofind the minimum set of genes that existedin the last common ancestor (LCA) — theprecursor cell(s) from which all livingorganisms subsequently evolved. The LCAmust have been a rather sophisticated cellbecause it contained all the cellular machin-ery that is common to all present livingforms, including a DNA-based informationsystem2,3. From sequence analysis, the LCAis suggested to contain 250–600 genes4,5. Mycoplasma genitalium, the simplest knownprokaryote cell (which is neverthelessdependent on host-cell parasitism), requires270 of its 380 genes for normal function, asrevealed when all genes were individually knocked out6. So, the minimum conceivablecell needs a lot of genetic information.
The Earth is ~4.56 billion years (Gyr)old, and the best current estimates arguethat the first cells appeared by 3.0–3.3 Gyrat the latest7. How these first cells emergedis one of the biggest unsolved problems inbiology. It seems undeniable that the LCAmust have been preceded by a spontaneousgeneration of cells from abiotic precursor
molecules. Even Darwin in his theory of evolution was obliged to accept the necessity of some kind of spontaneous generation.He brilliantly speculated (in a letter toHooker) that in a “warm little pond” aparticular chemistry involving “all sorts of
ammonia and phosphoric salts, light, heat,electricity etc. was present so that a proteincompound was chemically formed ready to undergo more complex changes on thepath towards life”8. It seems likely that his(unpublished) hypothesis was not far fromthe mark. Modern theories were initiatedby Oparin in 1924 (REF. 9) and by Haldane10,who first discussed the origin of membranesand speculated that an ‘oily film’ on the sur-
face of sea water evolved into the lipid-richcell membrane.
Here, I attempt to build up a plausiblescenario for how cells may have evolved— with an emphasis on the more difficultquestion of the origin of the cell membrane.A brief discussion of the (more actively pur-sued) question of the evolution of cytoplasm,including the nucleic acids, sets the stage forthe main hypothesis.
The evolution of the cytoplasm
Cells are made up of nucleic acids, proteins,carbohydrates, lipids and many smallmolecules suspended in a particular ionic
O P IN IO N
Cell evolution and the problem of membrane topology
Gareth Griffiths
Abstract | Cells somehow evolved from primordial chemistry and their emergencedepended on the co-evolution of the cytoplasm, a genetic system and the cell
membrane. It is widely believed that the cytoplasm evolved inside a primordial lipid
vesicle, but here I argue that the earliest cytoplasm could have co-evolved to high
complexity outside a vesicle on the membrane surface. An invagination of the
membrane, aided by an early cytoskeletal system, may have formed the first cells
— initially within primordial vesicles.
Figure 1 | Biosynthetic pathways for amino acids, phospholipids and central metabolism.
It has been proposed by Davis22,26 and many others that primitive biochemical pathways existed before
a genetic-based system. These include the reductive carbon cycle (the equivalent of the tricarboxylic
acid (TCA) cycle working in reverse), the reductive pentose pathway and the central trunk (proposed
to be a remnant of the formose cycle), which together make up the central biochemical pathway (CBP).
Davis proposes that the emergence of genetically encoded amino acids correlates with the number
of chemical reactions from the CBP that are required to generate each amino acid (evolutionary steps1–14). Acidic amino acids (Glu and Asp) are close to the CBP (1–2 reactions), whereas aromatic residues
(such as Trp) require up to 14 steps to be synthesized and may have appeared later. The hydrophobic
amino acids that could associate with membranes required four steps. The synthesis of phospholipids
requires 10 steps and, therefore, self-synthesizing membranes might only have arisen after this point.
Davis provides detailed analysis to argue that the evolutionary appearance of the different amino acids
correlates well with the emergence of their corresponding triplet codes; this implies co-evolution of
biochemistry and the genetic code, an idea extensively championed by Wong60. Davis also identified
an 11-amino acid sequence in the FtsZ–tubulin family that he mapped to his evolutionary stage 7.5,
which could support a role for this protein in cellularization. Gly, Cys and Pro cannot easily be placed
into any of the five categories shown.
1018 | DECEMBER 2007 | VOLUME 8 www.nature.com/reviews/molcellbio
PERSPECTIVES
© 2007 Nature Publishing Group

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 2/7
Mineral surface
PP
P P P PP
–
–– – – –
–
+ + + + + + + + +Mineralsurface
Gel
ProtocytoplasmCytoplasm insidevesicle
Cytoplasm outsidevesicle
Lipid vesicle
a
c
b d
Liposome
Unstable permeablebilayer Less permeable bilayer ‘Stable’ bilayer
Single acyl chain lipids• Monoglycerides• Fatty acids• Carboxylic acids
Impermeable membrane• Electric potential• Proton gradient
Two acyl chainphospholipids
Autonomouscell divides
Transient breaksin the membrane
Obcell
Protocell
RNARibosome
Protein
Pore/channel
environment. Before cells emerged, howwere sufficient amounts of the necessary starting materials generated? Following thepioneering experiments of Miller9, extensivestudies described many chemical reactionsthat plausibly occurred under the presumedconditions of the early Earth. These reac-tions could synthesize many (but not all)amino acids and other key precursors, suchas purines, and precursors of liposomessuch as carboxylic acids11–13.
The key precursors of life could havebeen made on Earth or in outer space andcarried to Earth via meteorites, such asthe Murchison meteorite that landed inAustralia in 1969 (REF. 14). Meteorites, aswell as interplanetary dust particles, bringan extraordinary selection and amountof chemicals and biochemical precursors of life from space, including >90 differentamino acids (of which 19 are found inliving organisms) and, as discussed below,bilayer-forming lipids15,16.
The cytoplasms of most modern cellshave a similar chemical composition with areducing environment, neutral pH and anionic composition that is rich in K+, Cl– andMg2+ but low in Ca2+ and Na+. Perhaps thisuniversal composition reflects the environ-ment where the first cells evolved; if so, theenvironment where the earliest chemistry that preceded life occurred is likely tohave been in fresh water17,18 rather thanthe high-salt ocean environment proposedby others10,19. The latter is also difficult toreconcile with the need for primordial lipid vesicles, which are unstable at high saltconcentrations 20.
For a protocytoplasm to emerge, many schemes have been postulated that involvethe occurrence of chemical catalysis andautocatalytic reactions on the surface of positively charged minerals, such as clay or iron pyrites21–25. A useful ‘yardstick’for timing the main events leading tocellularization is a scheme proposed by
Davis22,26 (FIG. 1; see below). The evolutionof the cytoplasm is often proposed to haveoccurred in parallel with the emergence of aliposome system, which eventually becamethe delimiting membrane of the cell.
The emergence of membranes
In a water-based system, many lipids canself-assemble into bilayer-containing vesicles. Phospholipids are the main bilayer-forming lipids in bacteria and eukaryotes(assuming that the ether-lipid-basedmembranes in archaea are a later adapta-tion to extreme environments22). However,the simplest bilayer-forming lipids arelong-chain (>C9) fatty acids, carboxylicacids and monoglycerides18,20. When suchlipids were extracted with solvents fromthe Murchison meteorite, they formedbilayered vesicles in aqueous solution16.Deamer and colleagues proposed that vesicles containing lipids made up of singleacyl chains formed the first template for the
Figure 2 | Surface interactions and the membrane problem. a | A pure
lipid vesicle in contact with a protocytoplasmic milieu. The system provides
two catalytic surfaces, the positively charged mineral surface and the
negatively charged, hydrophobic lipid vesicle. b | Two proposed membrane
scenarios for cellularization during evolution. In the ‘inside-out’ or ‘leaky
liposome’ model (left) the cytoplasm evolved within the vesicle and the
topology of the membrane (outer leaflet outside) remains in place during
the evolutionary process. In the ‘cytoplasm outside’ model, the cytoplasm
co-evolves with the membrane by associating with the outer membrane
surface (which will later become the cytoplasmic surface). A gel-like fila-
mentous material may prevent the diffusion of protocytoplasmic compo-
nents36. c | The ‘cytoplasm inside the vesicle’ hypothesis requires that the
molecular precursors of life must have found a way to pass selectively
through the liposome barrier. Deamer proposed that the earliest vesicles
contained bilayer membranes made up of single acyl chain lipids that are
more permeable to many molecules, including ions. As the system evolved
to use the more complex two acyl chain phospholipids, the membrane
became more impermeable. Additional proposed mechanisms that over-
come the permeability barrier include osmotic forces, transient breaks in
the membrane caused by polymerized amino acids such as polyleucine,
and transient openings in the bilayer caused by temperature changes or
freeze-thaw cycles20. d | The obcell model of the ‘outside-in’ hypothesis of
cellularization, as proposed by Blobel38 and Cavalier-Smith39. The early
liposome system is postulated to induce cisternae to fuse with themselves
to form double-membrane ‘obcells’ (inside-out cells)39. The model necessi-
tates the loss of the outer of the two membranes to release a protocell that
has the correct topology (with the luminal domains of the membrane
proteins facing outwards).
P E R S P E C T I V E S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | DECEMBER 2007 | 1019
© 2007 Nature Publishing Group

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 3/7
Golgi
Spore
Nucleus
Mitochondrion
pindleole body
b Budding yeast
a Vaccinia virus
c Autophagy
Fusionwithlysosome
(Microtubule) (Actin)IMV EEV
TGN
Golgi complex
Plasma membrane
cell membrane and that more complex (twoacyl chain) phospholipids emerged later20 (FIG. 2). Phospholipids can be synthesizedwithout enzymes under plausible abioticconditions12,13,27–29, but it seems likely thatthese lipids became more important whenRNA ribozymes or ribosomes and proteinenzymes emerged that could synthesizethem (FIG. 1).
The evolution of cells
If we assume that an increasingly complexcytoplasm with protein-synthesizing ribo-somes, nucleotides, RNA and even DNA
emerged in contact with a system of lipo-somes, we face a crucial unresolved issue inunderstanding the origin of cells and the cellmembrane: on which side of the liposomesdid the first key reactions occur?
The scenario of life within the vesicle. Mostspecialists think that such a system couldonly have evolved within the primordial vesicle20,30–34 (FIG. 2b). How then could theprecursors of the key molecules of life havecrossed a bilayer that presents a significantbarrier to charged molecules? Although vesicles comprised of single acyl chain
lipid membranes are more permeable touncharged and charged molecules, theiroverall permeability is several orders of magnitude lower than phospholipid bilayers.Significantly, all bilayers are highly imper-meable to ions such as K+ (REFS 18,20).As membrane complexity increases, thebilayer becomes more impermeable. In this‘cytoplasm within the vesicle’ scenario, theemerging ribosomes inside the vesicle wouldeventually evolve the capacity to insertmembrane proteins from the inside withtheir extracellular domains facing outwards,as in modern cells (FIG. 2b,c).
Many elegant experiments have been con-ducted to try and reconstitute some aspectsof this hypothesis. For example, investigatorstrapped enzymes and whole transcription–
translation systems within vesicles and foundthat these became functionally active 20,33–35 (FIG. 2c). Such analyses are still a long way from reconstituting life in such vesicles, asconceded even by some of the strongestproponents of this hypothesis18.
The ‘outside the vesicle’ scenario. Giventhe difficulties with the above hypothesis,let us consider another scenario in whichthe cytoplasm evolved to a high degree of complexity outside the vesicles. A mineralsurface could provide an environmenton which lipid vesicles become attached,and these surfaces could provide theenvironment where some rudimentary
components of a complex protocytoplasm— for example, perhaps ATP, GTP, proteins,RNA and maybe even DNA — could evolve.Given the problem of these key precursorsdiffusing away from the site of action, it isattractive to consider the emergence of apolymer in the space around the vesiclesthat can form a hydrophilic gel (REF. 36). Thiscould initially be a simple polymer such aspolysugars or polyglutamic acid that mightlater be replaced by a cytoskeletal proteinpolymer such as actin (see below). Suchsystems of filaments could serve to attachthe vesicles to the mineral surface, captureprecursor molecules within their gel-likematrix or provide a scaffold for protectionas well as an additional catalytic surface foremerging biochemistry. Under these condi-tions, there would be three catalytic surfaces— the positively charged mineral surface24,25,the surface of the gel36 and the negatively charged lipid vesicle surface — that couldprovide a rich surface for many reactions37.The mineral surface could itself catalyse theassembly of lipid vesicles25. In this micro-environment, sandwiched between thesesurfaces, a sophisticated protocytoplasm
Box 1 | Double membrane compartments in modern cells
There are at least three examples in modern cells that are topologically similar to the ‘life outside the
vesicle’ model (see figure). a | During the cellular release of vaccinia virus, the transGolgi network
(TGN) cisternal domain engulfs the intracellular mature virus (IMV)56. This intracellular enveloped
virus is transported along microtubules and fuses with the plasma membrane. During fusion with the
plasma membrane, or shortly thereafter, the local polymerization of actin around the TGN-derived
membrane facilitates virus release into the extracellular space (extracellular enveloped virus (EEV))57.
As proposed in the evolutionary schemes presented in FIG. 3, actin and microtubules also interact
here with membranes. b | During sporulation in the budding yeast Saccharomyces cerevisiae, a
double-membrane prospore cisterna is formed that originates from post-Golgi vesicles that fuse
around the forming (haploid) spore (only one spore is shown but up to four may be made). The Golgi
vesicles aggregate on the surface of the nucleus (pink) at the spindle pole body to enclose the spore
within two membranes58. c | The formation of autophagic vacuoles is proposed to occur by the
formation of a double-membrane structure59. A cytoplasmic cisternal structure (of unknown origin
but suspected to originate from the endoplasmic reticulum) wraps around cytoplasmic components
such as a mitochondrion to form a double-membrane vesicle. The outer membrane fuses with a late
endocytic compartment, whereas the inner membrane is expected to be lysed by the hydrolytic
conditions of the lysosomal lumen. In parts a andc, the purple ball and stick structure represents a
membrane-spanning protein, with the ball representing the luminal domain.
P E R S P E C T I V E S
1020 | DECEMBER 2007 | VOLUME 8 www.nature.com/reviews/molcellbio
© 2007 Nature Publishing Group
8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 4/7
LamellipodiaCell motility
Endocytosis
PhagocytosisMacropinocytosis
Fusion
Exocytosis
Cytokinesis
Filopodia
Microtubule
+ Motor + Motor
Membranenucleationof actin +
+
+
–
–
–
Comet
Cytokinesis
MreB (actin homologue)
FtsA (actin homologue)
FtsZ (tubulin homologue)
Myosin
might have emerged that interacted firstwith the outer surface of the vesicles. Smallmolecules may indeed have crossed spon-taneously into the vesicle lumen and I willrevisit this issue below.
If ribosomes could make hydrophobicpolypeptides, these would interact with the vesicle outer surface and, in time, the systemwould evolve the capacity to insert mem-brane proteins that spanned the membrane,
with their extracellular domains facing theinside of the vesicle38. To develop normalcell-membrane topology, the vesicle wouldhave to invert upon itself and fuse to form adouble-layered vesicle (FIG. 2d). The outer of the two vesicles would then need to lyse torelease the cell. This clever idea by Blobel38 is rarely cited by origin-of-life specialistsbut has been extensively championed by Cavalier-Smith39, who conceived the idea
of an ‘obcell’ or ‘inside-out cell’ (FIG. 2d).Whereas the ‘life within the vesicle’ model(FIG. 2c) has no obvious mechanistic prece-dents in modern cells, there are examplesfrom present-day cells in which a cisternalwrapping process occurs that is topologi-cally similar to the ‘cytoplasm outside the vesicle’ model (BOX 1). Below, I arguethat this (admittedly complex) inversionoccurred by numerous steps and dependedon the earliest cytoskeletal filaments.
Functions of the cytoskeleton
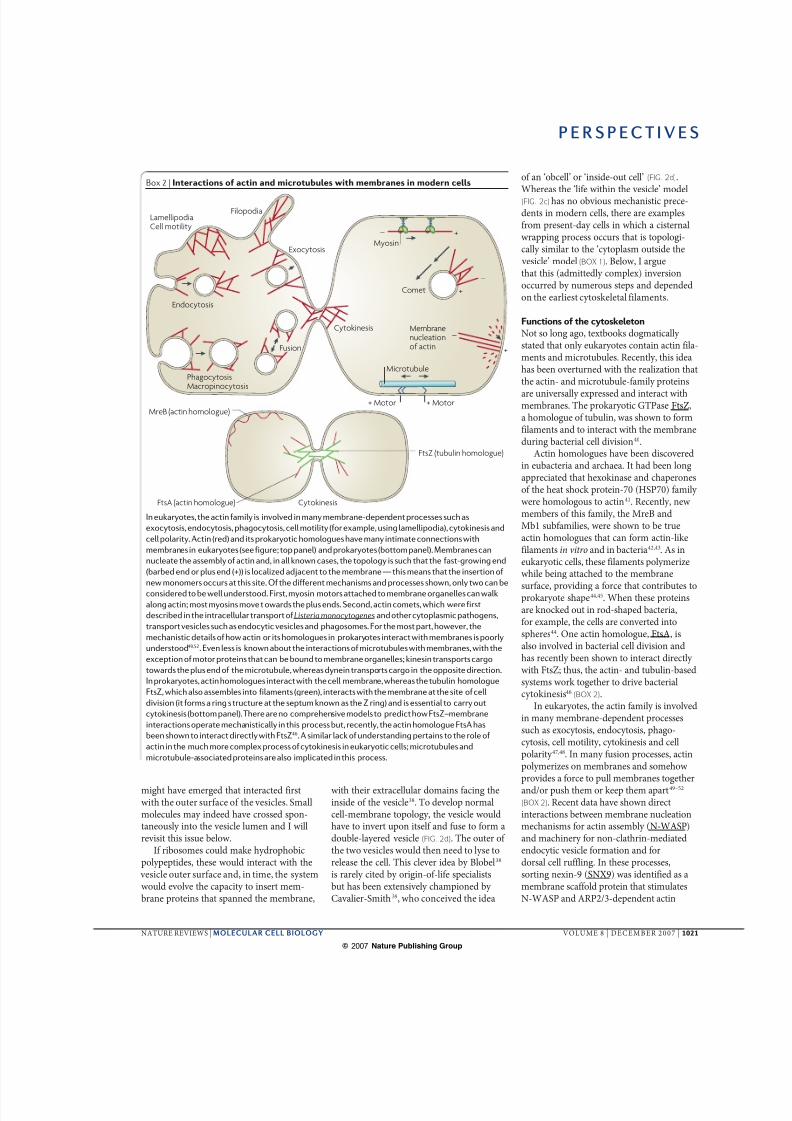
Not so long ago, textbooks dogmatically stated that only eukaryotes contain actin fila-ments and microtubules. Recently, this ideahas been overturned with the realization thatthe actin- and microtubule-family proteins
are universally expressed and interact withmembranes. The prokaryotic GTPase FtsZ,a homologue of tubulin, was shown to formfilaments and to interact with the membraneduring bacterial cell division40.
Actin homologues have been discoveredin eubacteria and archaea. It had been longappreciated that hexokinase and chaperonesof the heat shock protein-70 (HSP70) family were homologous to actin41. Recently, newmembers of this family, the MreB andMb1 subfamilies, were shown to be trueactin homologues that can form actin-likefilaments in vitro and in bacteria42,43. As ineukaryotic cells, these filaments polymerizewhile being attached to the membrane
surface, providing a force that contributes toprokaryote shape44,45. When these proteinsare knocked out in rod-shaped bacteria,for example, the cells are converted intospheres44. One actin homologue, FtsA, isalso involved in bacterial cell division andhas recently been shown to interact directly with FtsZ; thus, the actin- and tubulin-basedsystems work together to drive bacterialcytokinesis46 (BOX 2).
In eukaryotes, the actin family is involvedin many membrane-dependent processessuch as exocytosis, endocytosis, phago-cytosis, cell motility, cytokinesis and cellpolarity 47,48. In many fusion processes, actinpolymerizes on membranes and somehowprovides a force to pull membranes togetherand/or push them or keep them apart49–52 (BOX 2). Recent data have shown directinteractions between membrane nucleationmechanisms for actin assembly (N-WASP)and machinery for non-clathrin-mediatedendocytic vesicle formation and fordorsal cell ruffling. In these processes,sorting nexin-9 (SNX9) was identified as amembrane scaffold protein that stimulatesN-WASP and ARP2/3-dependent actin
Box 2 | Interactions of actin and microtubules with membranes in modern cells
In eukaryotes, the actin family is involved in many membrane-dependent processes such as
exocytosis, endocytosis, phagocytosis, cell motility (for example, using lamellipodia), cytokinesis and
cell polarity. Actin (red) and its prokaryotic homologues have many intimate connections with
membranes in eukaryotes (see figure; top panel) and prokaryotes (bottom panel). Membranes can
nucleate the assembly of actin and, in all known cases, the topology is such that the fast-growing end
(barbed end or plus end (+)) is localized adjacent to the membrane — this means that the insertion of
new monomers occurs at this site. Of the different mechanisms and processes shown, only two can be
considered to be well understood. First, myosin motors attached to membrane organelles can walk
along actin; most myosins move towards the plus ends. Second, actin comets, which were first
described in the intracellular transport of Listeria monocytogenes and other cytoplasmic pathogens,
transport vesicles such as endocytic vesicles and phagosomes. For the most part, however, the
mechanistic details of how actin or its homologues in prokaryotes interact with membranes is poorly
understood49,52. Even less is known about the interactions of microtubules with membranes, with the
exception of motor proteins that can be bound to membrane organelles; kinesin transports cargo
towards the plus end of the microtubule, whereas dynein transports cargo in the opposite direction.
In prokaryotes, actin homologues interact with the cell membrane, whereas the tubulin homologue
FtsZ, which also assembles into filaments (green), interacts with the membrane at the site of cell
division (it forms a ring structure at the septum known as the Z ring) and is essential to carry out
cytokinesis (bottom panel). There are no comprehensive models to predict how FtsZ–membrane
interactions operate mechanistically in this process but, recently, the actin homologue FtsA has
been shown to interact directly with FtsZ46. A similar lack of understanding pertains to the role of
actin in the much more complex process of cytokinesis in eukaryotic cells; microtubules and
microtubule-associated proteins are also implicated in this process.
P E R S P E C T I V E S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | DECEMBER 2007 | 1021
© 2007 Nature Publishing Group

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 5/7
+ + + + + + + + +
Pre-cytoplasmicenvironment
Microtubule/FtsZ
Actin
Lumen
RibosomeExtracellular space
External milieu
Protocell
a
de
cb
Luminalfusion
Cytoplasmicfusion
Inverted vesicle
Extracellular space
Cytokinesis
Autonomous cells
Channel
assembly, especially when it is allowed tooligomerize on phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P
2)-enriched
membrane domains in vitro. SNX9 containsa BAR domain that is known to facilitatemembrane bending53. Such studies startto resolve the long-elusive molecular linksbetween actin, its assembly and membranefunctions. Besides its role in cytoplasmicfusion processes (where the cytoplasmicleaflets of membranes first interact), arecent study shows that N-WASP-basedactin polymerization is also essential fortwo apposing plasma membrane luminaldomains to fuse completely duringDrosophila melanogaster myoblast fusion54.Thus, actin facilitates both cytoplasmic andluminal fusion events (FIG. 3).
Homologues of actin and tubulin havetherefore been identified in all kingdomsof life. The ATPase actin and the GTPasetubulin–FtsZ protein family may haveappeared before the cell became surrounded
by a membrane and, if so, could havefunctioned in cellularization. These sugges-tions are supported by an analysis by Davis,who argued that the order in which codedsynthesis of the different amino acids andlipids emerged during evolution correlateswith the number of reactions needed for theirsynthesis from an already evolved biochemi-cal system, which included the tricarboxylicacid (TCA) cycle and the pentose path-way 22,26 (FIG. 1). These reaction systems areuniversally involved in the synthesis of all 20common amino acids used by modern cells.Davis26 identified a conserved 11-residuesequence in the FtsZ–tubulin family that hemapped to his evolutionary stage 7.5, a stagehe classified as occurring before the systemcould self-synthesize membrane phospho-lipids (stage 10; FIG. 1). Because microtubulesand FtsZ, and especially actin, have intricateinteractions with modern membranes,I propose a speculative model for the role of their precursors in cellularization.
The cytoskeleton in cellularization?
My model of cellularization starts with inter-actions of the protocytoplasm with the outersurface of the initially pure lipid liposomes(FIG. 3a). In the model, I propose that proteins(made by ribosomes present in the proto-cytoplasm) evolved hydrophobic domainsthat allowed them to interact with liposomes.Later, membrane-spanning proteins, includ-ing channels and pumps, were inserted. Theproton and other pumps formed chemicaland electrical gradients and synthesized ATPon the cytoplasmic side of the membrane.The inside of the vesicle could then developa different composition to the outside andmight later have become the extracellularspace. Thus, in this model, even sophisticatedmembrane functions such as proton andelectrochemical gradients emerged beforecellularization.
Actin and/or tubulin ancestors theninteracted with the outer surface of the mem-brane and facilitated membrane bending.
Figure 3 | The outside-in model of cellularization. Shown is a possible evo-
lutionary mechanism by which the outside-in model (FIG. 2d) may have
occurred. a | An evolutionary stage exists in which a complex protocytoplasm
has a genetic code that is RNA- or even DNA-based, ribosomes for synthe-
sizing proteins and a relatively advanced biochemistry. The system has evolved
the capacity to insert membrane proteins such that the future extracellular or
luminal domains are inside the liposome. In parallel, an actin- and tubulin-
based cytoskeleton evolved the capacity to interact with the liposome surface.
b | The emergence of the cytoplasmic fusion machinery (orange) allows lipo-
somes to fuse together and bend membranes.c | The fusion machinery could
collaborate with the cytoskeleton to form an inwards-budding vesicle. This
budding process, which is similar to modern endocytosis but has the opposite
topology, allows the genetic material (not shown) and the protocytoplasm to
enter into a vesicle within a larger vesicle — the protocell. Channels and
transporters that allow ions to cross the membrane would be important for
maintaining the ionic homeostasis of the protocytoplasm inside the protocells
and in the extracellular space. d | The emergence of luminal fusion mecha-
nisms allows the process of fission out of the parental vesicle to occur.
This machinery also allows the protocells to fuse together within the extra-
cellular space. e | The cytoplasmic fusion machinery evolves into the process
of cytokinesis, which allows protocells to divide in a regulated fashion such
that each daughter cell contains everything it needs to metabolize and repli-
cate. The protocells are eventually released when the outer membrane lyses.
These are now independent living forms that are capable of self-replication.
P E R S P E C T I V E S
1022 | DECEMBER 2007 | VOLUME 8 www.nature.com/reviews/molcellbio
© 2007 Nature Publishing Group

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 6/7
A cytoplasmic machinery evolved that fusedthe vesicles together (FIG. 3b). A coordinatedeffort between the cytoskeleton and thecytoplasmic fusion machinery then allowedapposing membrane lipid coalescence andthe inwards pinching of vesicles to formprotocells (FIG. 3c). These ‘inverted’ vesicleswithin the larger vesicle enclose the proto-cytoplasm and the genetic material (DNA orRNA); it is implicitly assumed that this evolv-ing genetic material is intimately associatedwith the cytoplasmic surface of the vesicle(not shown in FIG. 3).
The formation of these vesicles istopologically equivalent to the budding of vesicles into the lumen of specialized endo-cytic organelles — multivesicular bodies— in modern eukaryotic cells. Additional
machinery must have evolved that allowedthe luminal domains of the protocell mem-branes to fuse together. The same machinery could allow cells to bleb off a membrane vesicle by fission, a process equivalent toblebbing in modern cells. The protocellscould undergo selection by content mix-ing driven by fusion with themselves andwith newly made protocells bringing innew components from the external milieu(FIG. 3d).
At a later stage, the cytoplasmic fusionmachinery facilitates the separation of daughter protocells in a regulated fashion(cytokinesis) (FIG. 3e). The final stage is lysisof the first outer membrane to release inde-
pendent cells. These are expected to haveprobably several hundred DNA-encodedgenes in order for the minimal cell functionsto be permitted.
The extracellular compartment (FIG. 3c) would offer several advantages to theemerging cells bathing within this space.The evolution of mechanisms to transferglycoconjugates bound to lipids or proteinsonto and beyond the luminal surface of theboundary membrane of protocells couldprovide a reserve energy source for cellsif sugar hydrolases were also secreted intothis space. A high-viscosity environmenthere could also protect the enclosed cellsfrom extreme environmental changesin the external environment. Ion channels inthe boundary membrane and the protocellmembrane could concentrate some ions,such as Ca2+ and protons, which would bestored in the extracellular buffering com-partment and injected into the protocells asrequired. It is conceivable that many of themechanisms proposed for permeation of themembrane in the ‘inside the vesicle’ scenario(FIG. 2c) could be operational in the ‘outsidethe vesicle’ model (FIG. 3). However, instead
of molecules entering the future cytoplasm,in the latter model, the transport of mole-cules into the lumen of the vesicles wouldallow the future extracellular space to evolve.
The stage of protocells within a vesiclemight, perhaps, have existed for a relatively long time. The evolutionary split betweeneukaryotes and prokaryotes55 could also havedeveloped in this system, protected by twomembranes.
Conclusions and perspectives
I have outlined a plausible scenario forthe co-evolution of the cytoplasm and themembrane based on existing cell biologicalprinciples. I have discussed the two principaltheories with respect to the evolution of themembrane and suggested that the ‘cytoplasm
outside’ model (FIG. 3) is more plausible thanthe more favoured ‘cytoplasm inside the vesicle’ model. If the inside-out (or obcell)model is correct, it seems reasonable to sug-gest that it needed the active participation of a cytoskeleton to ‘invert’ the topology of themembrane system and initiate the cellular-ization process. The model predicts that theactin and tubulin family preceded the LCAand that they are probably as universal as the vacuolar ATPase family.
Experimentally, one could ask how aliposome system with a subset of recon-stituted membrane proteins would behavewith cytosolic extracts and factors such asglucose, GTP and ATP. If the cytoplasmic
domain of these membrane proteins all facedoutwards, some aspects of the ‘life outsidethe vesicle’ model presented here could betested. Alternatively, if they had the oppositeorientation, they could perhaps be usedto test some aspects of the ‘life within the vesicle’ scenario.
Gareth Griffiths is at the
European Molecular Biology Laboratory,
Heidelberg, Germany.
e-mail:
doi:10.1038/nrm2287
Published online 31 October 2007
1. Doolittle, R. F. Searching for the common ancestor.
Res. Microbiol. 151, 85–89 (2000).
2. Poole, A., Jeffares, D. & Penny, D. Early evolution:
prokaryotes, the new kids on the block. Bioessays 21,
880–889 (1999).3. Reichard, P. The evolution of ribonucleotide reduction.
Trends Biochem. Sci. 22, 81–85 (1997).
4. Gil, R., Silva, F. J., Pereto, J. & Moya, A.
Determination of the core of a minimal bacterial gene
set. Microbiol. Mol. Biol. Rev. 68, 518–537 (2004).
5. Koonin, E. V. Comparative genomics, minimal gene-
sets and the last universal common ancestor. Nature
Rev. Microbiol. 1, 127–136 (2003).
6. Glass, J. I. et al. Essential genes of a minimal bacterium.
Proc. Natl Acad. Sci. USA 103, 425–430 (2006).
7. Brasier, M., McLoughlin, N., Green, O. & Wacey, D.
A fresh look at the fossil evidence for early Archaean
cellular life. Philos. Trans. R. Soc. Lond. B Biol. Sci.
361, 887–902 (2006).
8. Kutschera, U. & Niklas, K. J. The modern theory of
biological evolution: an expanded synthesis.
Naturwissenschaften 91, 255–276 (2004).
9. Miller, S. L., Schopf, J. W. & Lazcano, A. Oparin’s
“Origin of Life”: sixty years later. J. Mol. Evol. 44,
351–353 (1997).
10. Haldane, J. The origin of life. Rationalist Annual 3,
148–153 (1929).
11. Ferris, J. P. Prebiotic synthesis: problems and
challenges. Cold Spring Harb. Symp. Quant. Biol. 52,
29–35 (1987).
12. Oro, J., Miller, S. L. & Lazcano, A. The origin and early
evolution of life on Earth. Annu. Rev. Earth Planet. Sci.
18, 317–356 (1990).
13. Pohorille, A. & Wilson, M. Molecular dynamics studies
of simple membrane–water interfaces: structure and
functions in the beginnings of cellular life. Orig. Life
Evol. Biosph. 25, 21–46 (1995).
14. Kvenvolden, K. et al. Evidence for extraterrestrial
amino-acids and hydrocarbons in the Murchison
meteorite. Nature 228, 923–926 (1970).
15. Oro, J., Mills, T. & Lazcano, A. Comets and the
formation of biochemical compounds on the primitive
Earth — a review. Orig. Life Evol. Biosph. 21,
267–277 (1992).
16. Deamer, D. W. & Pashley, R. M. Amphiphilic
components of the Murchison carbonaceous
chondrite: surface properties and membraneformation. Orig. Life Evol. Biosph. 19, 21–38
(1989).
17. Griffith, R. W. Freshwater or marine origin of the
vertebrates? Comp. Biochem. Physiol. A 87,
523–531 (1987).
18. Monnard, P. A. & Deamer, D. W. Membrane self-
assembly processes: steps toward the first cellular life.
Anat. Rec. 268, 196–207 (2002).
19. Martin, W. & Russell, M. J. On the origins of cells:
a hypothesis for the evolutionary transitions from
abiotic geochemistry to chemoautotrophic
prokaryotes, and from prokaryotes to nucleated cells.
Philos. Trans. R. Soc. Lond. B Biol. Sci. 358, 59–85
(2003).
20. Deamer, D. W. & Dworkin, J. P. Chemistry and physics
of primitive membranes. Top. Curr. Chem. 259, 1–27
(2005).
21. Bernal, J. The Origin of Life (Weidenfeld and
Nicholson, London, 1967).
22. Davis, B. Evolution of the genetic code. Prog. Biophys.
Mol. Biol. 72, 157–243 (1999).
23. Maden, B. E. No soup for starters? Autotrophy and
the origins of metabolism.Trends Biochem. Sci. 20,
337–341 (1995).
24. Wachtershauser, G. Groundworks for an evolutionary
biochemistry: the iron-sulphur world. Prog. Biophys.
Mol. Biol. 58, 85–201 (1992).
25. Hanczyc, M. M., Mansy, S. S. & Szostak, J. W. Mineral
surface directed membrane assembly. Orig. Life Evol.
Biosph. 37, 67–82 (2007).
26. Davis, B. Molecular evolution before the origin of
species. Prog. Biophys. Mol. Biol. 79, 77–133 (2002).
27. Deamer, D. W. & Oro, J. Role of lipids in prebiotic
structures. Biosystems 12, 167–175 (1980).
28. Hargreaves, W. R., Mulvihill, S. J. & Deamer, D. W.
Synthesis of phospholipids and membranes in
prebiotic conditions. Nature 266, 78–80 (1977).
29. Rao M, E. M., Oro J. Synthesis of phosphatidylcholine
under possible primitive earth conditions. J. Mol. Evol.
18, 196–202 (1982).
30. Baeza, I. et al. Liposomes with polyribonucleotides as
model of precellular systems.Orig. Life Evol. Biosph.
17, 321–331 (1987).
31. Morowitz, H. Beginnings of Cellular Life: Metabolism
Recapitulates Biogenesis (Yale Univ. Press, New
Haven, 1992).
32. Luisi, P. L., Ferri, F. & Stano, P. Approaches to semi-
synthetic minimal cells: a review. Naturwissenschaften
93, 1–13 (2006).33. Monnard, P. A., Oberholzer, T. & Luisi, P. Entrapment
of nucleic acids in liposomes. Biochim. Biophys. Acta
1329, 39–50 (1997).
34. Yu, W. et al. Synthesis of functional protein in
liposome. J. Biosci. Bioeng. 92, 590–593 (2001).
35. Ishikawa, K., Sato, K., Shima, Y., Urabe, I. & Yomo, T.
Expression of a cascading genetic network within
liposomes. FEBS Lett. 576, 387–390 (2004).
36. Trevors, J. T. & Pollack, G. H. Hypothesis: the origin of
life in a hydrogel environment. Prog. Biophys. Mol.
Biol. 89, 1–8 (2005).
37. Segre, D., Ben-Eli, D., Deamer, D. W. & Lancet, D.
The lipid world. Orig. Life Evol. Biosph. 31, 119–145
(2001).
P E R S P E C T I V E S
NATURE REVIEWS | MOLECULAR CELL BIOLOGY VOLUME 8 | DECEMBER 2007 | 1023
© 2007 Nature Publishing Group

8/6/2019 PAPER 2 Cell Evolution
http://slidepdf.com/reader/full/paper-2-cell-evolution 7/7
38. Blobel, G. Intracellular protein topogenesis.
Proc. Natl Acad. Sci. USA 77, 1496–1500
(1980).
39. Cavalier-Smith, T. Obcells as proto-organisms:
membrane heredity, lithophosphorylation, and the
origins of the genetic code, the first cells, and
photosynthesis. J. Mol. Evol. 53, 555–595
(2001).
40. Lowe, J., van den Ent, F. & Amos, L. A. Molecules of
the bacterial cytoskeleton. Annu. Rev. Biophys.
Biomol. Struct. 33, 177–198 (2004).
41. Bork, P., Sander, C. & Valencia, A. An ATPase domain
common to prokaryotic cell cycle proteins, sugar
kinases, actin, and Hsp70 heat shock proteins.
Proc. Natl Acad. Sci. USA 89, 7290–7294 (1992).
42. Egelman, E. H. Actin’s prokaryotic homologs.
Curr. Opin. Struct. Biol. 13, 244–248 (2003).
43. Li, J. Y. & Wu, C. F. Perspectives on the origin of
microfilaments, microtubules, the relevant chaperonin
system and cytoskeletal motors — a commentary on
the spirochaete origin of flagella. Cell Res. 13,
219–227 (2003).
44. Carballido-Lopez, R. & Errington, J. A dynamic
bacterial cytoskeleton. Trends Cell Biol. 13, 577–583
(2003).
45. Doolittle, R. F. & York, A. L. Bacterial actins? An
evolutionary perspective. Bioessays 24, 293–296
(2002).46. Erickson, H. P. Evolution of the cytoskeleton.
Bioessays 29, 668–677 (2007).
47. DeRosier, D. J. & Tilney, L. G. F-actin bundles are
derivatives of microvilli: what does this tell us about
how bundles might form? J. Cell Biol. 148, 1–6
(2000).
48. Janmey, P. A. & Lindberg, U. Cytoskeletal regulation:
rich in lipids. Nature Rev. Mol. Cell Biol. 5, 658–666
(2004).
49. Eitzen, G. Actin remodeling to facilitate membrane
fusion. Biochim. Biophys. Acta 1641, 175–181
(2003).
50. Kjeken, R. et al. Fusion between phagosomes, early
and late endosomes: a role for actin in fusion between
late, but not early endocytic organelles. Mol. Biol. Cell
15, 345–358 (2004).
51. Kaksonen, M., Toret, C. P. & Drubin, D. G.
Harnessing actin dynamics for clathrin-mediated
endocytosis. Nature Rev. Mol. Cell Biol. 7, 404–414
(2006).
52. Soldati, T. & Schliwa, M. Powering membrane traffic in
endocytosis and recycling. Nature Rev. Mol. Cell Biol.
7, 897–908 (2006).
53. Yarar, D., Waterman-Storer, C. M. & Schmid, S. L.
SNX9 couples actin assembly to phosphoinositide
signals and is required for membrane remodeling
during endocytosis. Dev. Cell 13, 43–56 (2007).
54. Massarwa, R., Carmon, S., Shilo, B. Z. &
Schejter, E. D. WIP/WASp-based actin-polymerization
machinery is essential for myoblast fusion in
Drosophila. Dev. Cell 12, 557–569 (2007).
55. Kandler, O. [Festival lecture. The position of
microorganisms in the global phylogenetic system of
three domains]. Mycoses 37 (Suppl. 1), 13–27 (1994).
56. Schmelz, M. et al. Assembly of vaccinia virus: the
second wrapping cisterna is derived from the trans
Golgi network. J. Virol. 68, 130–147 (1994).
57. Rietdorf, J. et al. Kinesin-dependent movement on
microtubules precedes actin-based motility of vaccinia
virus. Nature Cell Biol. 3, 992–1000 (2001).
58. Moreno-Borchart, A. C. & Knop, M. Prospore
membrane formation: how budding yeast gets shaped
in meiosis. Microbiol. Res. 158, 83–90 (2003).
59. Kim, J. & Klionsky, D. J. Autophagy,
cytoplasm-to-vacuole targeting pathway, and
pexophagy in yeast and mammalian cells. Annu. Rev.
Biochem. 69, 303–342 (2000).
60. Wong, J. T. Coevolution theory of the genetic code atage thirty. Bioessays 27, 416–425 (2005).
AcknowledgementsI thank T. Gibson, M. Knop, S. Trachtenberg, D. Lancet,
G. van Meer, L. Mayorga and J. Reich for their comments
and discussion, and C. Bleck for preparing the figures.
DATABASESEntrez Genome Project: http://www.ncbi.nlm.nih.gov/sites/
entrez?db=genomeprj
Listeria monocytogenes | Mycoplasma genitalium
UniProtKB: http://beta.uniprot.org/
FtsA | FtsZ | N-WASP| SNX9
FURTHER INFORMATIONGareth Griffiths’s homepage:
http://www-db.embl.de/jss/EmblGroupsOrg/g_79.html
ALL LINKS ARE ACTIVE IN THE ONLINE PDF
P E R S P E C T I V E S
1024 | DECEMBER 2007 | VOLUME 8 www.nature.com/reviews/molcellbio
© 2007 Nature Publishing Group









![[Paper] Darwinian Design TheMemetic Evolution](https://static.fdocuments.net/doc/165x107/577cdac71a28ab9e78a68269/paper-darwinian-design-thememetic-evolution.jpg)








