
GalNAc mimetics: from synthesis to potential inhibitors in ...

Developmental Biology 242, 58–73 (2002)doi:10.1006/dbio.2001.0530, available online at http://www.idealibrary.com on
Overexpression of the CT GalNAc Transferasein Skeletal Muscle Alters Myofiber Growth,Neuromuscular Structure, and Laminin Expression
Bing Xia,* Kwame Hoyte,* Anja Kammesheidt,* Tom Deerinck,†Mark Ellisman,† and Paul T. Martin* ,1
*Department of Neuroscience, Glycobiology Research and Training Center, and †NationalCenter for Microscopy, University of California, San Diego, School of Medicine,9500 Gilman Drive, La Jolla, California 92093-0691
Carbohydrates have been shown to mediate or modulate a number of important events in the development of the nervoussystem; however, there is little evidence that they participate directly in the development of synapses. One carbohydratestructure that is likely to be important in synaptic development of the neuromuscular junction is the CT carbohydrateantigen {GalNAcb1,4[NeuAca2,3]Galb1(-3GalNAc or -4GlcNAc)}. The synaptic localization of the CT antigen is due to thepresence of the terminal b1,4 GalNAc linkage, and such linkages are localized to the neuromuscular junction in manyspecies. Here we show that an enzyme that can create the synaptic CT structure, the CT GalNAc transferase, is alsoconfined to the neuromuscular junction in mice. Using transgenic mice, we show that overexpression of the CT GalNActransferase in extrasynaptic regions in skeletal myofibers caused as much as a 60% reduction in the diameter of adultmyofibers and an order of magnitude increase in satellite cells. Neuromuscular junctions of transgenic mice had severelyreduced numbers of secondary folds, Schwann cell processes were present in the synaptic cleft, and secondary folds wereoften misaligned with active zones. In addition, multiple presynaptic specializations occurred on individual myofibers. Inaddition, some normally synaptic proteins, including laminin a4, laminin a5, utrophin, and NCAM, were expressed alongextrasynaptic regions of myofibers. One of the muscle proteins that displayed increased glycosylation with the CT antigenin the transgenic mice was a-dystroglycan. These experiments provide the first in vivo evidence that a synapticcarbohydrate antigen has important roles in the development of the neuromuscular synapse and suggest that the CT antigenis involved in controlling the expression of synaptic molecules. © 2002 Elsevier Science
Key Words: synapse; GalNAc; neuromuscular junction; laminin; satellite cell; dystroglycan.
INTRODUCTION
Synapse formation requires the precise apposition of alarge number of proteins, both in time and in space (forreview, see Sanes and Lichtman, 1999). Since most proteinsthat reside within the synaptic cleft are glycoproteins, onemeans of discriminating synaptic proteins from their extra-synaptic counterparts would be the creation of a uniquesynaptic “glyco-environment.” Such a specialized glycanenvironment could be required for the precise alignmentand packing of the high density of proteins that occurs atsynapses, for unique synaptic cell adhesion, for synaptic
1 To whom correspondence should be addressed. Fax: (858) 822-
1021. E-mail: [email protected].58
targeting of proteins, or to create novel synaptic signalingmechanisms. A number of studies have implicated carbo-hydrates in other aspects of neural development, includingcell fate, neural migration, stem cell proliferation, myelina-tion, axon guidance, dendrite maturation, and neural sig-naling (for reviews, see Rutishauser and Landmesser, 1996;Yamaguchi, 2001; Muramatsu, 2000; Jessel et al., 1990;Margolis and Margolis, 1989). Given the importance ofcarbohydrates in these developmental events, it is likelythat they also have roles in synapse formation, although theevidence in support of this notion is scarce.
Several carbohydrate structures have been implicated inaspects of synapse formation. Changes in polysialic acid(PSA) levels have been implicated in estrogen-induced
changes in synaptic structure in the arcuate nucleus (Hoyk0012-1606/02 $35.00© 2002 Elsevier Science
All rights reserved.

59Overexpression of the CT GalNAc Transferase
et al., 2001), in environmentally induced morphologicalplasticity in the hypothalamic–neurohypophysial system(Theodosis et al., 1999), and in ectopic mossy fiber–pyramidal cell synapse formation in the hippocampus (Sekiand Rutishauser, 1998). In addition, PSA (Eckhardt et al.,2000), heparan sulfate (Lauri et al., 1999), and mannoseoligosaccharides (Luthi et al., 1994) can affect synapticplasticity paradigms such as long-term potentiation in thehippocampus. Mannose structures have been implicated inpresynaptic vesicle aggregation in sensory neurons in theleech (Tai and Zipser, 1998), and studies of mannose bind-ing lectins such as concanavalin A suggest that glycans maycontrol the specificity of certain synaptic connections inAplysia (Lin and Levitan, 1987). In many of these studies,however, it is not clear whether the effects observed aredirect effects of glycan (e.g., glycan binding to a receptor) orwhether they are a secondary consequence of more genericalterations in membrane structure or charge. Moreover, it isoften not clear in such studies whether the carbohydrate inquestion is uniquely present at synapses or whether suchlocalization is generally applicable to the synapses of morethan one species.
One of the keys to identifying glycans that are importantfor synapse formation is to identify those structures that areuniquely present at synapses and whose synaptic expres-sion has been phylogenetically conserved. Demonstrationof the conservation of synaptic expression in differentspecies is an important clue to function for carbohydratestructures, as carbohydrates are often receptors or corecep-tors for bacterial or viral pathogens (Gagneux and Varki,1999). As such, they are likely to be placed under intenseevolutionary pressure. The neuromuscular junction con-tains such a synaptic saccharide, terminal b-linkedN-acetyl-D-galactosamine or bGalNAc (Sanes and Cheney,1982; Scott et al., 1988). The synaptic localization ofbGalNAc in skeletal muscle is conserved in rodents, hu-mans, chicks, frogs, fish, and birds, and is highly concen-trated, though not exclusively so, at neuromuscular junc-tions in lamprey and Torpedo (Scott et al., 1988). SeveralbGalNAc-containing structures have been defined at themammalian neuromuscular junction (Martin et al.,1999). Among these are the CT carbohydrate antigens{GalNAcb1,4[NeuAca2,3]Galb1(-3GalNAc or -4GlcNAc)-R}, which are defined by the CT1 and CT2 monoclonalantibodies (Lefrancois and Bevan, 1985). In mice, thesestructures are referred to as the CT antigen for their originaldescription in activated cytotoxic T cells (Lefrancois andBevan, 1985). In humans, they correspond to the Cad or Sdablood group antigens (Conzelmann and Lefrancois, 1988).Cad and Sda are relatively rare blood groups in humans,although these structures are more commonly expressed inother human tissues (Dohi et al., 1996).
The enzyme responsible for making the synapticb1,4GalNAc linkage on the CT antigen is the CT GalNActransferase (Smith and Lowe, 1994). This enzyme requiresNeuAca2,3Galb1,-R (where R is usually 4GlcNAc or 3Gal-
NAc) as a substrate and modifies glycoproteins but gener-© 2002 Elsevier Science. A
ally not glycolipids (Smith and Lowe, 1994). While it hasnot been proven, the CT GalNAc transferase is most likelynot the same enzyme as the GalNAc O-phosphotransferaseactivity previously described at the neuromuscular junction(Scott et al., 1990). This enzyme migrates at a very differentmolecular weight than that of the CT GalNAc transferaseand appears to have an entirely different enzymatic activity(Balsamo et al., 1986). Here we describe the synaptic local-ization of the CT GalNAc transferase at the neuromuscularjunction. Using ectopic expression of the transferase in theskeletal muscles of transgenic mice, we also identify novelfunctions for this type of glycosylation in muscle andneuromuscular development and demonstrate its presenceon a-dystroglycan.
MATERIALS AND METHODS
Materials
Monoclonal antibody to desmin (DE-U-10) was obtained fromSigma. Monoclonal antibodies to a-sarcoglycan (Ad1/20A6),b-dystroglycan (43DAG1/8D5), utrophin (DRP3/20C5), and dystro-phin (Dy4/6D3) were obtained from Nova Castra. Antibody tolaminin a2 (4H8-2) was obtained from Alexis Biochemicals. Anadditional polyclonal antisera, DG67, was made against theC-terminal 15 amino acids of b-dystroglycan in our laboratory andpurified against an immunogenic peptide-coupled resin. Antibodyto laminin b1 (5A2) was a gift from Dale Abrahamson (Universityof Alabama, Birmingham). Antisera to ab-dystroglycan (OR12)and antibody to a-dystroglycan (IIH6) were gifts from Kevin Camp-bell (HHMI, University of Iowa, Iowa City). Antibody toa-dystroglycan (VIA4-1) was purchased from Upstate Biotechnol-ogy. Hybridomas producing antibodies against the CT antigen(CT1, CT2) were gifts from Leo Lefrancois (University of Connecti-cut, Storrs). Polyclonal antibodies to NCAM, laminin a4, laminina5, and laminin b2 were gifts from Joshua Sanes (WashingtonUniversity, St. Louis, MO). Polyclonal antibodies to the acetylcho-line receptor epsilon chain and c-met were obtained from SantaCruz Biotechnology. Polyclonal antibody to acetylcholinesterasewas a gift from Palmer Taylor [University of California, San Diego(UCSD)]. Polyclonal antibody to neuron-specific enolase was ob-tained from Chemicon. Antibody to synaptophysin was obtainedfrom Boehringer Mannheim. Monoclonal antibody to a-dystro-brevin (d32620) was obtained from BD Transduction Labs. Threeanti-peptide antibodies were raised in rabbit against peptide se-quences encoded in the CT GalNAc transferase. CT76 was madeagainst KVALQYFKNHLYCST, CT77 was made against MTSSVS-FASFRFPWLLK, and CT68 was made against KLRMKYKFQ-DAYNQKD. Staining and blotting with CT68 are shown in Fig. 1.These antibodies were affinity purified against the immunogenicpeptide conjugated to Affigel 10 (Bio-Rad) using standard low pHmethods (Harlow and Lane, 1988). All secondary antibodies werepurchased from Boehringer Mannheim or Zymed. Rhodamine-a-bungarotoxin was purchased from Molecular Probes. WFA (Wist-eria floribunda Agglutinin) agarose was purchased from EY labora-tories. Protein G agarose was purchased from BoehringerMannheim. Anti-mouse IgM agarose was purchased from Jackson
Laboratories.ll rights reserved.

is 50
60 Xia et al.
HistologyFor hematoxylin–eosin, myosin ATPase, and immunofluores-
cence staining of thin sections, muscles were snap frozen in liquidnitrogen-cooled isopentane and sectioned at 6 mm on a cryostat. Forwhole-mount preparations, diaphragm or intercostal muscles weredissected and fixed in 2% paraformaldehyde with 0.1% TritonX-100. For confocal imaging, mice were anesthetized with 1 ml/kgnembutol, stretched, and perfused by intracardiac catheterizationwith buffer (135 mM NaCl, 14 mM NaHCO3, 1.2 mM Na2HPO4, 5mM KCl, 2 mM CaCl2, and 1 mM MgCl2) at 35°C, followed by 4%
FIG. 1. Characterization of transgenic mice expressing the CT Gacompared to wild type mice(Wt) for expression of the CT GalNAc trof its confinement to the neuromuscular junction, the CT GalNAcis highly expressed in Tg mice. Detection of a FLAG epitope tag oand not in other tissues. Sk, skin; Li, liver; Bl, bladder; Te, testes;cord; Ki, kidney; He, heart; Lu, lung; SM, skeletal muscle (D, diaphsmall compared to their littermates, but moved and fed normally. Creceptors (B) and antibodies to the CT GalNAc transferase (B9) showjunction. (C) Expression of the CT GalNAc transferase was absent fTg mice (D). The CT antigen was also absent from extrasynaptic ain Tg mice (F). (G) Immunoblotting of muscle cell protein lysatespossessed the CT antigen in skeletal muscle extracts. In the threethe CT GalNAc transferase. Bar in B9 is10 mm (for B, B9). Bar in D
formaldehyde in phosphate-buffered saline (PBS) for 5 min. Gas-
© 2002 Elsevier Science. A
trocnemious, diaphragm, and intercostal muscles were dissectedand fixed for an additional hour. For electron microscopy, micewere prepared as above, only they were perfused with 2% formal-dehyde and 2% gluteraldehyde in 0.15 M sodium cacodylate. Foranti-carbohydrate staining, sections were blocked in Tris-bufferedsaline (TBS; pH 7.4) in 3 mg/ml bovine serum albumin (BSA). CT2antibody (5 mg/ml) was precomplexed with goat anti-mouse IgM-FITC (2.5 mg/ml) for 1 h, then added with 50 nM rhodamine-a-bungarotoxin. All other antibodies were added at 1 mg/ml. Anti-peptide antibodies were added at 2 mg/ml in TBS with 3 mg/ml
transferase in skeletal muscle.(A) Three transgenic (Tg) lines wererase by Western blot using antibodies against the enzyme. Becausesferase is not easily detected in such experiments in Wt mice, butnsgenic protein showed it to be expressed only in skeletal muscletestine; St, stomach; Th, thymus; Sp, spleen; Br, brain; SC, spinal; G, gastrocnemius; I, intercostal), Pa, pancreas. Tg mice were veryining of adult muscles with a-bungarotoxin to label acetylcholinehat the transferase was highly concentrated at the neuromuscularextrasynaptic regions of Wt myofibers but was highly expressed inof Wt myofibers (E) and was present within and around myofibersan antibody to the CT carbohydrate showed that several proteinsnes studied, several protein bands were increased by expression of
mm (for C–F).
lNAcansfetran
n traIn, inragm
ostaed t
romreaswithTg li
BSA. Sections were washed in TBS and incubated with 10 mg/ml
ll rights reserved.

61Overexpression of the CT GalNAc Transferase
goat anti-rabbit-FITC and 50 nM rhodamine-a-bungarotoxin. Anti-peptide antibody specificity was confirmed by blocking with excesspeptide. Slides were dried and mounted in glycerol with paraphe-nylenediamine to prevent quenching. For routine immunofluores-cence and color staining, slides were analyzed using a Nikon E800epifluorescence microscope.
Transgenic MiceThe mouse CT GalNAc transferase cDNA containing a FLAG
epitope tag at its N-terminus (Parkhomovskiy et al., 2000) wassubcloned into pBS-HSAvpA (Muscat and Kedes, 1987) at the PacIsite. pBS-HSAvpA is a plasmid that contains a portion of thehuman skeletal alpha actin promoter with intronic sequences, anSV40 splice acceptor site, and SV40 polyadenylation sequences.After purification by double banding on cesium chloride gradients,the plasmid was linearized by cutting with KpnI and PvuI, sepa-rated by agarose gel electrophoresis, and purified using Quiex IIresin (Qiagen). DNA was diluted and injected into fertilized oo-cytes of CB6 mice by the UCSD Cancer Center Mouse CoreFacility.
Eight lines of transgenic mice were made and analyzed. Geno-typing was done by RT-PCR and confirmed by Southern blottingand Western blotting (Fig. 1). Of the eight lines made, two did notbreed, two had low or uneven levels of expression, and four hadhigh and uniform levels of expression in all skeletal muscles and noexpression in other organs. Three of these lines, 2124 (Tg1), 2159(Tg2), and 2162 (Tg3), were used in this study. All phenotypesreported and all molecular analyses shown were found to be presentin multiple lines. Because of their small size, transgenic mice haddifficulty mating. Therefore, several lines were maintained bytransplant of transgenic ovaries into surrogate mothers.
ImmunoblottingWhole cell lysates of skeletal muscle were solubilized by homog-
enization in 75 mM Tris, pH 6.8, 4% SDS, 4 M urea, 20% glycerol,and 1% b-mercaptoethanol. Samples were diluted and proteinlevels were quantitated using Bio-Rad DC protein assay kit (Bio-Rad, Richmond, CA). Proteins were separated on either 6% or 12%SDS–PAGE and transferred to nitrocellulose. For anti-carbohydrateblotting, blots were blocked in Tris-buffered saline (pH 7.4) with0.1% Tween 20 and 3 mg/ml BSA. Blots were incubated withanti-CT2 antibody, washed in TBST, incubated in anti-mouseIgM-alkaline phosphatase, washed, and developed. For all otherantibodies except IIH6 and VIN4-1, blots were blocked in TBSTwith 5% nonfat dry milk and secondary antibodies were conjugatedto peroxidase and developed using ECL chemiluminescence. IIH6and VIN4-1 were used as previously described (Ervasti and Camp-bell, 1991).
Electron MicroscopyFixed muscles were postfixed with 2% OsO4, dehydrated, and
embedded. Thin sections (80 nm) were cut using a diamond knife(Diatome) and an Ultracut E ultramicrotome (Leica) and mountedon uncoated copper grids. Thin sections were imaged at 80 keVusing a 1200 EX electron microscope (JEOL).
Confocal MicroscopyFixed muscle was permeabilized in a solution containing 0.1%
Triton X-100, 1% normal goat serum, and 1% cold water fish
© 2002 Elsevier Science. A
gelatin (Sigma) in PBS for 30 min before incubation in anti-synaptophysin or anti-neuron-specific enolase for 18 h at 4°C.Tissue was then washed in buffer and incubated in donkey anti-mouse IgG-FITC conjugate or donkey anti-rabbit Cyan-conjugatemixed with 50 nM rhodamine-a-bungarotoxin in PBS for 1 h at 4°C.Muscles were rinsed in PBS and mounted in Gelvatol. Confocalmicroscopy was performed using an MRC-1024 system (Bio-Rad)attached to an Axiovert 35M (Zeiss AG) using a 340 1.3 n.a.objective. Individual images (1024 3 1024 pixels) were converted toPICT format and merged as pseudocolor RGB images using AdobePhotoshop (Adobe Systems).
Primary Muscle Cultures
Gastrocnemius muscles from 7-week-old wild type and trans-genic mice were dissected, minced, and digested for 30 min at37°C, 5% CO2 in dissociation medium containing 2% fetal calfserum, 1.0 mg/ml collagenase (Sigma), 1.5 mg/ml trypsin (IrvineScientific), and 2 mg/ml DNAseI (Boehringer Mannheim) inDMEM. The now-dissociated muscle tissue was then titurated 20to 30 times, spun for 5 min at 1500g, and resuspended in primaryculture media containing 20% fetal calf serum, 4% chick embryoextract (Gibco/BRL), 50 U/ml penicillin, and 50 mg/ml streptomy-cin in Ham’s F12 media supplemented with glutamine. Cells werepreplated on plastic for 20 min to remove fibroblasts. The mediawas removed and cells were titurated again before plating cells on0.4% gelatin-coated plastic dishes. For myotube fusion assays, cellswere cultured at high density and allowed to grow to confluence,after which they were induced to fuse into myotubes by placementin serum-poor media (DMEM with 2% horse serum, 50 U/mlpenicillin, and 50 mg/ml streptomycin) for 3 days. Cells were thenwashed and fixed in 2% paraformaldehyde, and myotube fusionwas quantitated by light microscopy by counting random 320fields of view. To quantitate growth rate, cells were cultured atlower density and kept in growth media. Cells were counted usinga hemocytometer during passaging at Days 0, 3, and 6.
Analysis of Muscles and Neuromuscular Junctions
Quantitation of central nuclei, myofiber diameter and variance,and nuclei/myotubes was done using hemotoxylin and eosin stain-ing of frozen sections using Sigma premade reagents according tothe manufacturer’s instructions. Data for gastrocnemius and ex-tensor digitorum longus muscles were pooled for statistics, but didnot vary significantly from one another. The coefficient of variance(standard deviation of diameter 3 1000/mean diameter) was alwaysbelow the 250 value expected of normal muscle. Similar resultswere also seen for diaphragm. Reductions of at least 60% inmyofiber diameter were seen for all three lines; however, Tg2162was reduced by 70% and this was significantly more than thereduction measured for Tg2159 or Tg2124 (P , 0.001). Fiber typingwas done by performing myosin ATPase staining at pH 4.6 (Ha-malainen and Pette, 1993) on fresh frozen sections. While type 1fiber levels were increased in the gastrocnemius, the level ofincrease was uneven and paralleled the medial–lateral variations infiber type reported in Frey et al. (2000). An example of the mostextreme concentration of type 1 fibers is shown in Fig. 2. Thelength of the diaphragm and endplate band and number of myofi-bers with multiple presynaptic specializations were analyzed usingwhole-mount stains in which postsynaptic areas were visualizedwith rhodamine-a-bungarotoxin. The percentage of satellite cells,
length of I- and A-bands, length of neuromuscular junction, num-ll rights reserved.
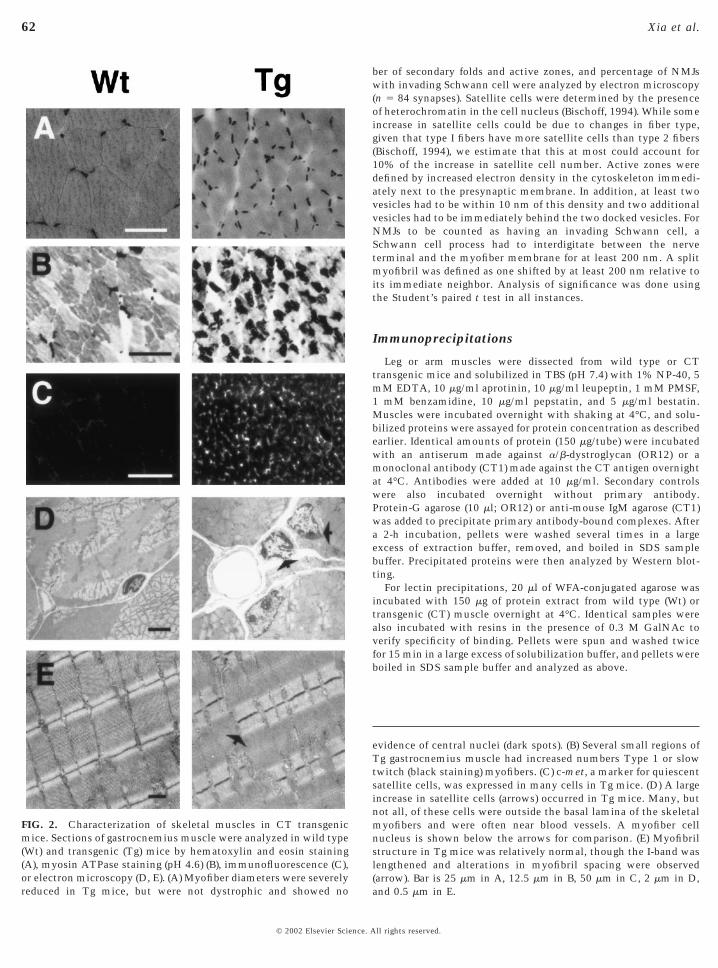
reduced in Tg mice, but were not dystrophic and showed no
62 Xia et al.
© 2002 Elsevier Science. A
ber of secondary folds and active zones, and percentage of NMJswith invading Schwann cell were analyzed by electron microscopy(n 5 84 synapses). Satellite cells were determined by the presenceof heterochromatin in the cell nucleus (Bischoff, 1994). While someincrease in satellite cells could be due to changes in fiber type,given that type I fibers have more satellite cells than type 2 fibers(Bischoff, 1994), we estimate that this at most could account for10% of the increase in satellite cell number. Active zones weredefined by increased electron density in the cytoskeleton immedi-ately next to the presynaptic membrane. In addition, at least twovesicles had to be within 10 nm of this density and two additionalvesicles had to be immediately behind the two docked vesicles. ForNMJs to be counted as having an invading Schwann cell, aSchwann cell process had to interdigitate between the nerveterminal and the myofiber membrane for at least 200 nm. A splitmyofibril was defined as one shifted by at least 200 nm relative toits immediate neighbor. Analysis of significance was done usingthe Student’s paired t test in all instances.
Immunoprecipitations
Leg or arm muscles were dissected from wild type or CTtransgenic mice and solubilized in TBS (pH 7.4) with 1% NP-40, 5mM EDTA, 10 mg/ml aprotinin, 10 mg/ml leupeptin, 1 mM PMSF,1 mM benzamidine, 10 mg/ml pepstatin, and 5 mg/ml bestatin.Muscles were incubated overnight with shaking at 4°C, and solu-bilized proteins were assayed for protein concentration as describedearlier. Identical amounts of protein (150 mg/tube) were incubatedwith an antiserum made against a/b-dystroglycan (OR12) or amonoclonal antibody (CT1) made against the CT antigen overnightat 4°C. Antibodies were added at 10 mg/ml. Secondary controlswere also incubated overnight without primary antibody.Protein-G agarose (10 ml; OR12) or anti-mouse IgM agarose (CT1)was added to precipitate primary antibody-bound complexes. Aftera 2-h incubation, pellets were washed several times in a largeexcess of extraction buffer, removed, and boiled in SDS samplebuffer. Precipitated proteins were then analyzed by Western blot-ting.
For lectin precipitations, 20 ml of WFA-conjugated agarose wasincubated with 150 mg of protein extract from wild type (Wt) ortransgenic (CT) muscle overnight at 4°C. Identical samples werealso incubated with resins in the presence of 0.3 M GalNAc toverify specificity of binding. Pellets were spun and washed twicefor 15 min in a large excess of solubilization buffer, and pellets wereboiled in SDS sample buffer and analyzed as above.
evidence of central nuclei (dark spots). (B) Several small regions ofTg gastrocnemius muscle had increased numbers Type 1 or slowtwitch (black staining) myofibers. (C) c-met, a marker for quiescentsatellite cells, was expressed in many cells in Tg mice. (D) A largeincrease in satellite cells (arrows) occurred in Tg mice. Many, butnot all, of these cells were outside the basal lamina of the skeletalmyofibers and were often near blood vessels. A myofiber cellnucleus is shown below the arrows for comparison. (E) Myofibrilstructure in Tg mice was relatively normal, though the I-band waslengthened and alterations in myofibril spacing were observed(arrow). Bar is 25 mm in A, 12.5 mm in B, 50 mm in C, 2 mm in D,
FIG. 2. Characterization of skeletal muscles in CT transgenicmice. Sections of gastrocnemius muscle were analyzed in wild type(Wt) and transgenic (Tg) mice by hematoxylin and eosin staining(A), myosin ATPase staining (pH 4.6) (B), immunofluorescence (C),or electron microscopy (D, E). (A) Myofiber diameters were severely
and 0.5 mm in E.
ll rights reserved.

63Overexpression of the CT GalNAc Transferase
RESULTS
Using affinity-purified anti-peptide antibodies madeagainst regions of the CT GalNAc transferase, we firstdemonstrated that this enzyme, like the carbohydrateantigens it creates (Martin et al., 1999), is localized tothe neuromuscular junction (Fig. 1). Expression of theCT GalNAc transferase was highly colocalized withrhodamine-a-bungarotoxin staining, which identifies thehigh density of acetylcholine receptors present in thepostsynaptic membrane at the neuromuscular junction. Inaddition, occasional staining was seen in small blood ves-sels and capillaries (data not shown), although no extrasyn-aptic expression was observed in myofibers. Thus, theexpression of the synaptic CT carbohydrate antigen inskeletal muscle is likely due to the synaptic localization ofthe CT GalNAc transferase, which is the enzyme thatcreates the synaptic b1,4GalNAc linkage (Smith and Lowe,1994).
To ascertain potential roles for the CT GalNAc trans-ferase in synaptic development, we created transgenic (CT)mice that would maintain extrasynaptic expression of theCT GalNAc transferase in the skeletal muscles of adultanimals. We characterized three transgenic lines, all ofwhich expressed the enzyme at high levels in all skeletalmuscles examined and did not express in other tissues (Fig.1A). This was done by driving CT GalNAc transferaseexpression using the human skeletal a actin promoter.Increased expression of the CT GalNAc transferase wasseen in most muscles by immunostaining (Figs. 1C and 1D)and all myofibers expressed the CT antigen in relativelyeven amounts (Figs. 1E and 1F). Intracellular aggregates ofCT staining could also be seen in some transgenic muscles(Fig. 1F). While it is impossible to identify all of theCT-containing glycoproteins using blotting techniques, weidentified several glycoprotein bands that immunoblottedin wild type muscle extracts with the anti-CT2 monoclonalantibody (Fig. 1G). Given the low expression of the CTGalNAc transferase in muscle and its presence in non-muscle structures such as blood vessels, it was possible thatthese were in fact not muscle-specific glycoproteins, de-spite the use of a muscle protein extract. Therefore, werepeated this analysis on three lines of transgenic micemade to overexpress the CT GalNAc transferase specifi-cally in skeletal muscles, with the hope that newly glyco-sylated bands would in fact be derived from the glycosyla-tion of muscle proteins. Several new CT-positive proteinbands were observed in whole muscle lysates from all threetransgenic lines (Fig. 1G). Bands at 35, 85, and 160 kDaappeared to be increased, though there were clearly othersas well. Similar results were seen with both the CT1 andthe CT2 antibody, both of which can be created by the CTGalNAc transferase (Smith and Lowe, 1994). These experi-ments demonstrate that transgenic mice maintained extra-synaptic expression of the normally synaptic CT antigenand that CT overexpression altered the glycosylation of
several muscle glycoproteins.© 2002 Elsevier Science. A
CT transgenic mice walked, fed normally, and lived anormal lifespan. The immediately obvious phenotype of thetransgenic mice was their small size (Fig. 1). Adult micewere reduced in weight by 40% compared to age- andsex-matched littermates (Table 1). Analysis of cross sec-tions of the gastrocnemius, extensor digitorum longus,intercostal, and diaphragm muscles demonstrated thatmyofiber diameters were reduced by 60% at 6 weeks of age(Fig. 2 and Table 1). This reduction in myofiber diameterwas not caused by a simple developmental delay, as 18-month-old transgenic mice still had a 40 6 1% reduction inmyofiber diameter (for gastrocnemius, P , 0.001). Only a7 6 1% decrease in relative myofiber diameter was seen atbirth, while a 41 6 4% change was evident by postnatal day14 (for gastrocnemius, P , 0.001 for both ages). Thus, mostof the loss in myofiber diameter occurred in the earlypostnatal period and persisted well into adulthood. Similarreductions in diameter were also observed in other muscles,including the trapezius, gluteus, tricep, quadricep, andtibialis anterior (data not shown).
While the muscles from the CT transgenic mice weresmall, this was not due to a failure of muscle formation ordegeneration. All major muscle groups were present. Thetotal number of myofibers per muscle was unchanged, aswas the percentage of cells with central nuclei (Table 1). Nosignificant change in the variability of fiber diameter wasseen (Table 1). The number of nuclei in total cross sectionsof the gastrocnemius muscle remained constant (Table 1).Thus, no significant differences in cell death, degeneration,or regeneration were apparent in transgenic muscles com-pared to normal animals. Transgenic muscles were morerounded and the percentage of type 1 (or slow) fibers wasincreased in small regions of the gastrocnemius muscle (Fig.2, Table 1). However, fiber typing of other muscles, includ-ing the diaphragm (Table 1), showed no change in fiber typecomposition. Muscle ultrastructure was assessed by elec-tron microscopy of longitudinal sections. All filamentstructures appeared normal, with two exceptions. First, theI-band length was increased in transgenic mice. This couldbe an artifact of fixation; however, the muscles were allstretched prior to fixation and the lengthening was observedin multiple animals. Thus, it is possible that some aspect ofmyofibril structure was altered. The second finding was anincrease in the number of split myofibrils (Fig. 2, Table 1).This is analogous to what is observed in mice lacking desmin(Lin et al., 1997), an intermediate filament protein that mayhelp connect myofibril units (Patel and Lieber, 1997).
The reduced myofiber growth in CT transgenic mice wasdue in part to the arrest of satellite cell fusion withmyotubes. During late embryonic and early postnatal de-velopment, satellite cells associate and often fuse withalready formed myofibers. In doing so, such cells contributeto the dramatic growth of myofibers that occurs in the earlypostnatal period (Bischoff, 1994; Seale et al., 2000). At birth,approximately 32% of the nuclei in a muscle section aresatellite cells, while in adults less than 5% remain and
serve as a reservoir of regenerative material (Albrook et al.,ll rights reserved.

64 Xia et al.
1971; Cardasis and Cooper, 1975; Schmalbruch and Hell-hammer, 1977). In CT transgenic mice, we estimated thatalmost 50% of the nuclei outside of myofibers were satel-lite cells. This was based on the observation of heterochro-matin in mononucleated cells using electron microscopy.We admit that this definition is hardly adequate; neverthe-less, it was trivial to identify transgenic muscles blindlyusing this criterion, and the change was quite striking.Expression of c-met, a marker for quiescent satellite cells,was dramatically increased in cells surrounding myofibersin transgenic mice, and electron micrographs showed manysatellite cells present both within and outside the basallamina surrounding myofibers (Fig. 2). The reduction inmyofiber diameter was similar to that seen in mice lackingPax7, a transcription factor required for the production ofmyogenic satellite cells (Seale et al., 2000).
To test further the idea that CT expression caused anincrease in satellite cells or an arrest in their fusion intomyotubes, we extracted satellite cells from muscles of adulttransgenic and wild type mice (Figs. 3A and 3B). Satellitecells were made using standard techniques with preplatingon tissue culture plastic to reduce intramuscular fibro-
TABLE 1Alterations in Synaptic and Myofiber Parameters Resulting from C
Parameter Muscles
Weight (Male)Weight (Female)Tibia length% Central nuclei G, EDLMyofiber diameter G, EDLCoeff. of diameter variance G, EDLMyofiber length DI-band length D, IA-band length D, I% Type 1 fibers G% Type 1 fibers D% Split myofibrils G, D, INuclei/myotube G% Satellite cell nuclei GNMJ length GNMJ length D% NMJ with invading SC GArea of endplate band DActive zones/NMJ GActive zones/NMJ DSecondary folds/NMJ GSecondary folds/NMJ D% Myofibers with multiple
presynaptic specializationsG
Note. Measures of myofiber and neuromuscular characteristicsgastrocnemius; D, diaphgram; EDL, extensor digitorum longus; I, istandard errors of the mean.
* P , 0.05, **P , 0.01, ***P , 0.001, NS, not significant.
blasts. Based on c-met staining of plated cells done imme-
© 2002 Elsevier Science. A
diately after extraction, this produced an average of 95%satellite cells. We counted the number of cells extractedfrom wild type and transgenic animals, normalized to theweight of muscle tissue used for extraction, and thencultured them using rich media optimized for growth ofsatellite cells in culture. At 3 and 6 days, we then recountedcells to determine relative growth rates (Fig. 3A). Immedi-ately after plating, there was an 8.5-fold increase (P , 0.02)in the number of satellite cells extracted from CT trans-genic mice compared to that from wild type animals. Thisdegree of increase was similar to the increase in satellitecells seen in vivo using electron microscopy (Table 1).Satellite cells taken from wild type mice grew rapidly, asexpected of satellite cells in culture. Wild type cells dis-played a linear growth rate (R 5 0.97) and had a doublingtime of about once per day.
Cells taken from CT transgenic muscles, by contrast,showed a negative linear growth rate (R 5 0.99), with abouthalf of the cells dying by 6 days. We can think of threescenarios to explain these data. First, CT expression couldcause a developmental defect in satellite cell growth. Sec-ond, CT expression could be toxic to satellite cells in
ntigen Overexpression in Skeletal Muscle
ild Type Transgenic P value
6 0.6 g 14 6 0.6 g ***6 0.6 g 14 6 0.2 g **6 0.4 mm 15 6 0.4 mm NS6 0.4 3.7 6 1 NS6 0.4 mm 12 6 0.2 mm ***160 188 NS
6 0.1 mm 4.2 6 0.2 mm ***6 40 nm 820 6 40 nm ***6 0.1 mm 1.3 6 0.1 mm NS6 1 18 6 3 **6 1 18 6 1 NS6 2 20 6 3 ***6 0.1 1.2 6 0.1 NS6 2 48 6 5 ***6 0.7 mm 1.6 6 0.1 mm **6 0.8 mm 4.4 6 0.6 mm **6 0 70 6 6 ***6 20 mm 1,400 6 60 mm ***6 1 3 6 1 *6 1 5 6 1 ***6 1 5 6 1 ***6 2 4 6 2 ***6 10 54 6 8 *
compared in 7-week-old wild type and CT transgenic mice. G,ostal; SC, Schwann cell; NMJ, neuromuscular junction. Errors are
T A
W
242115
2.929
3.43801.3
6183
1.22
4.87.6
0450
611152110
arenterc
culture; or third, satellite cells could be at such a high
ll rights reserved.

65Overexpression of the CT GalNAc Transferase
density prior to extraction that they have already differen-tiated to some extent and have consequently dropped out ofthe cell cycle. We tend to favor the third scenario, assatellite cells in the adult animal were densely packed andlikely contacted each other (e.g., see Fig. 2D). To testwhether satellite cells were competent to fuse into myo-tubes, we cultured satellite cells from the gastrocnemiusmuscle of CT transgenic and wild type mice at high density(Fig. 3B). When induced to fuse by growth in serum-poormedia, transgenic cells formed myotubes at levels similar
FIG. 3. Analysis of satellite cell growth and fusion in culture.Muscles from wild type (Wt) and CT transgenic mice (CT) weredissected and primary cultures of satellite cells were grown inculture. (A) Upon initial culturing, there was an 8.5-fold increase inthe number of satellite cells per weight of dissected muscle takenfrom transgenic mice compared to that from wild type mice (P ,0.02). Satellite cells from wild type mice grew at a linear rate (R 50.97) over the course of 6 days, multiplying roughly every day. Bycontrast, satellite cells from CT mice did not grow and decreased innumber linearly over the course of 6 days (R 5 0.99). (B) Satellitecells from wild type (Wt) and CT transgenic mice (CT) werecultured at high density. Cells were induced to fuse into myotubesby placement in serum-poor media for 3 days. The number ofmyotubes per random 320 field of view was counted. Wt and CTcells fused equally well (P 5 0.2).
to those seen with wild type cells (P 5 0.2). Therefore, the
© 2002 Elsevier Science. A
failure of postnatal muscle growth in CT transgenic micewas not due to a general inability of satellite cells to formmyotubes.
Neuromuscular junctions of CT transgenic mice dis-played abnormal structure by a number of measures (Fig. 4,Table 1). First, the size of individual junctions was reduced,as would be expected given the smaller size of the myofi-bers. This was apparent both by electron (Figs. 4C and 4D)and confocal (Figs. 4E and 4F) microscopy. The length ofneuromuscular junctions was reduced by 67 6 7% in thegastrocnemius muscle and by 43 6 5% in the diaphragm(P , 0.01 for both). Secondary folds in the muscle mem-brane were reduced and in some cases were completelyabsent. Electron micrographs of transgenic muscles (Figs.4D and 4G) showed that secondary folds were not onlyreduced but were often not aligned with active zones andsometimes not even with the nerve terminal (Fig. 4D).Some synapses appeared more normal, however; overall,there was a 55 6 8% reduction in folds in the gastrocne-mius (P , 0.001) and a 67 6 9% reduction in folds in thediaphragm (P , 0.001). The width of the synaptic cleft wasnormal, but Schwann cells that normally cap the nerveterminal often extended processes between the nerve andmuscle. Fully two thirds of transgenic neuromuscular junc-tions examined had some Schwann cell process betweentheir nerve terminal and the postsynaptic membrane (Table1). Nerve terminals had active zones (though reduced innumber; Table 1) with docked secretory vesicles and hadnormal vesicle distribution, with most secretory vesiclesbeing present on the half of the nerve terminal closest to themyofiber. In addition, colabeling of postsynaptic mem-brane, motor axons, and nerve terminals was done tofurther characterize neuromuscular development (Figs. 4E,4F, 4H, and 4I). Several differences were observed. First,individual synapses had disjointed expression of acetylcho-line receptors (AChRs) that lacked the contiguous pretzel-like pattern of normal junctions (Figs. 4E and 4F). Second,the innervation pattern of the muscle was fundamentallydifferent. In many muscles, including diaphragm and inter-costal muscles, axons migrate in a direction perpendicularto that of their target myofibers. This creates an “endplateband” of innervation near the middle muscle. In CT trans-genic mice, the distance over which innervation occurredtripled (Fig. 4B and Table 1), and axons sometimes sproutedand migrated beyond neuromuscular junctions (Figs. 4Hand 4I). Third, many myofibers had more than one presyn-aptic specialization (Figs. 4H and 4I). This contrasts withthe one synapse per muscle situation found at most adultneuromuscular junctions (see Sanes and Lichtman, 1999).
Given the large changes observed in neuromuscularstructure, we were interested to determine whether any ofthe molecules implicated in neuromuscular developmentby genetic studies would have altered protein expressionpatterns in the CT transgenic mice. Two nerve-derivedproteins, agrin and ARIA, have been shown to be importantfor the organization of the postsynaptic membrane. Mice
lacking neural agrin fail to properly localize acetylcholinell rights reserved.

66 Xia et al.
receptors under the nerve during development (Gautum etal., 1996; Burgess et al., 1999), and mice that are heterozy-gous for a form of ARIA have a 50% reduction in acetyl-choline receptor mRNA levels (Sandrock et al., 1997).Likewise, several muscle-derived synaptic chains of lami-
FIG. 4. Characterization of neuromuscular innervation in CT tranwith rhodamine-a-bungarotoxin to label acetylcholine receptors (microscopy (C, D, G, J) or confocal microscopy (E, F, H, I), w(synaptophysin, green) and postsynaptic membrane (acetylcholinegreatly expanded in Tg mice (B) compared to control animals (Aindicated by arrows in C), while Tg muscles had few and sometimeswere often present on Tg muscles (D, J) and multiple synapses wjunctions displayed the typical pretzel-shaped pattern of postsynmembranes (F) and displayed abnormal axonal migration (arrows inbut secondary folds were very reduced and processes from Schwannthe synaptic cleft. Bar in A is 500 mm (for A, B), 7.5 mm (for E, F), ain J.
nin, in particular laminin b2 and laminin a4, have impor-
© 2002 Elsevier Science. A
tant roles in presynaptic development. Mice lacking lami-nin b2 have deficits in active zone and secondary foldformation as well as Schwann cell invasion into the synap-tic cleft (Noakes et al., 1995; Patton et al., 1998). Micelacking laminin a4 have normal numbers of active zones
ic mice. Diaphragm muscle was analyzed by whole-mount staining, or sections of gastrocnemius muscle were analyzed by electronaxons (neuron-specific enolase, blue in E, F), nerve terminals
tors, red) were colabeled. The endplate band of the diaphragm wasividual NMJs in Wt mice had numerous secondary folds (somecondary folds (D). In addition, multiple presynaptic specializations
resent on the same muscle (numbers in H, I). Wt neuromuscularmembrane (E), while Tg junctions had disjointed postsynaptic
). Active zones were still present at Tg synapses (arrowheads in G),(arrow in G), often extended between the nerve and muscle within
mm (for H, I). Bar is 0.5 mm in C (for C, D), 0.2 mm in G, and 1 mm
sgenA, B)here
recep). Indno seere papticH, Icellsnd 5
and secondary folds, but do not properly organize them
ll rights reserved.

67Overexpression of the CT GalNAc Transferase
across the synaptic cleft (Patton et al., 2001). Mice lackinglaminin a5 die as embryos (Miner et al., 1998), although cellculture studies suggest that laminin a5 can alter presynap-tic development and be responsible for some of the pheno-types observed in laminin b2-deficient mice (Patton et al.,1997). Mice lacking a particular laminin chain often replaceit with another isoform (Martin et al., 1996; Patton et al.,1998); thus, it is often difficult to discriminate whether agiven phenotype is caused by the loss of one chain or theectopic expression of another. Mice lacking NCAM havealso been implicated in pre- and postsynaptic development.Mice overexpressing NCAM in skeletal muscles showsprouting of Schwann cells and axons (Walsh et al., 2000),and NCAM-deficient mice have slightly decreased neuro-muscular size and deficits in paired-pulse facilitation (Mos-coso et al., 1998; Rafuse et al., 2000). In addition, muscleslacking dystroglycan have abnormal neuromuscular struc-tures (Cote et al., 1999). a-Dystroglycan can bind to lami-nins and agrins, while b-dystroglycan can bind to cytoplas-mic proteins, including dystrophin and utrophin (forreview, see Henry and Campbell, 1999).
As we had expressed the CT antigen in skeletal muscle,we focused on the expression of synaptic laminin chainsand their binding proteins (Fig. 5). Laminin a4 and laminina5 were expressed around myofibers in transgenic mice, aswas utrophin, a synaptic homolog of dystrophin (Ohlen-dieck et al., 1991) (Fig. 5). In addition, expression of the celladhesion molecule NCAM, a normally synaptic protein(Couvalt and Sanes, 1986), and the intermediate filamentprotein desmin, were increased (Fig. 5). Other moleculesthat are normally expressed along extrasynaptic regions ofmyofibers, including laminin b1, b-dystroglycan, a-sarco-glycan, dystrophin (all Fig. 5), and dystrobrevin (not shown),were unchanged in their expression pattern. Expression oflaminin b2 and acetylcholinesterase, both normally synap-tic molecules (McMahan et al., 1978; Hunter et al., 1989),remained synaptic (Fig. 5). Increased protein levels could bemeasured for laminin a4, laminin a5, NCAM, and utrophin(Fig. 6), consistent with their increased expression aroundmyofibers. Expression of c-met was also increased, consis-tent with the increase in satellite cells. Several additionalprotein bands were evident for desmin and dystrobrevin(Fig. 6). It is not clear whether these represent proteolyticproducts or novel isoforms. For example, the new dystro-brevin band seen in transgenic extracts migrates at theexpected molecular weight of dystrobrevin 3 and/or 4(Peters et al., 1998). An additional dystrobrevin band wasalso seen in muscle extracts of two of the three CTtransgenic lines at 22 kDa, which would be consistent withthe molecular weight of dystrobrevin 5 (data not shown).Other developmental events, such as expression of theepsilon chain of the acetylcholine receptor (Fig. 6), theelimination of M-cadherin expression along myofibers, andthe concentration of synapsin and SNAP 25 in nerve termi-nals (data not shown), occurred normally. Thus, ectopic
expression of the CT antigen altered the expression of a© 2002 Elsevier Science. A
group of normally synaptic molecules but did not appear tosimply arrest all development.
Given the rather complex group of phenotypes, bothcellular and molecular, in the CT transgenic mice, it was ofinterest to begin to identify the glycoproteins that hadincreased expression of the CT antigen in transgenicmuscles. In immunoblotting for proteins in extracts oftransgenic muscle, we noticed that very developed immu-noblots for a-dystroglycan had several proteolytic products(85 and 35 kDa; Fig. 7) that comigrated with bands identi-fied by anti-CT antibodies (Fig. 1). To test whethera-dystroglycan was glycosylated with the CT antigen, weextracted skeletal muscle tissue from wild type and CTtransgenic muscle using nonionic detergent and subjected itto precipitation with anti-dystroglycan antisera, anti-CTantibody, or WFA, a bGalNAc-binding lectin (Fig. 7). Pre-cipitated proteins were then blotted with antibodies todystroglycan or the CT carbohydrate. Two protein bands(160 and 35 kDa) were precipitated by both CT1 anddystroglycan antibodies that could be recognized by anti-bodies to the CT antigen and to a-dystroglycan (Fig. 7A).These molecular weights correspond to the weight of ma-ture a-dystroglycan (160 kDa) and a presumed proteolyticfragment (35 kDa).
Since anti-carbohydrate IgM antibodies are difficult touse in such experiments, we also precipitated identicalamounts of protein from wild type and transgenic muscleextracts using WFA, a bGalNAc-specific binding lectin,conjugated to agarose (Fig. 7B). A significant amount ofmature (160 kDa) a-dystroglycan could be precipitated fromCT muscle, while none was precipitated from wild typemuscle. a-Dystroglycan precipitated from CT muscle wasalso recognized by anti-CT antibody. In addition,b-dystroglycan, which binds tightly to the a-chain (Ervastiand Campbell, 1991), was also precipitated from CT muscleusing this lectin. Addition of 0.3 M GalNAc during theprecipitation blocked a-dystroglycan binding to WFA, dem-onstrating that lectin binding was GalNAc specific. Noproteolytic fragments of a-dystroglycan were precipitatedto a significant degree in these experiments. Because the CTglycans are likely present in the mucin region of thea-dystroglycan protein, where many copies of these struc-tures would be highly packed together, the binding ofa-dystroglycan to IgM antibodies and WFA, both of whichare large multivalent binding proteins, is likely to becomplex. Nevertheless, these data strongly suggest thata-dystroglycan can be glycosylated with the CT antigen inthe skeletal muscles of CT transgenic mice.
DISCUSSION
The results presented suggest four novel roles for the CTcarbohydrate antigen in muscle growth and neuromusculardevelopment. First, the CT antigen alters the expression ofa group of synaptic proteins, including laminin a4, laminin
a5, utrophin, and NCAM (Figs. 5 and 6). All of thesell rights reserved.

68 Xia et al.
FIG. 5. Extrasynaptic expression of normally synaptic proteins in CT transgenic mice. Cross sections of gastrocnemius muscle in wildtype (Wt) and transgenic (Tg) muscles were stained with antibodies to laminin a2, laminin a4, laminin a5, laminin b2, laminin b1, NCAM,acetylcholinesterase (AChE), b-dystroglycan, a-sarcoglycan, utrophin, dystrophin, and desmin. Increased extrasynaptic expression oflaminin a4, laminin a5, and utrophin was seen around myofibers in Tg mice, and increased expressions of desmin, both intracellular andextracellular, and NCAM were seen. Therefore, CT carbohydrate expression caused the ectopic expression of a group of normally synapticproteins. Normal expression of laminin a5 in capillaries and blood vessels is shown in Wt as is expression of laminin b2 in blood vessels.Acetylcholinesterase was expressed only at neuromuscular junctions (examples shown by arrows) in both Tg and Wt animals, whilelaminin a2, laminin b1, dystrophin, b-dystroglycan, and a-sarcoglycan, which are normally expressed around myofibers, were unchanged.
NCAM is normally confined to the neuromuscular junction in adult animals (arrow shows NMJ in Wt). Bar is 50 mm for all images.© 2002 Elsevier Science. All rights reserved.

holin
69Overexpression of the CT GalNAc Transferase
proteins have effects of their own on neuromuscular struc-ture or function (Noakes et al., 1995; Patton et al., 1997,1998, 2001; Moscoso et al., 1998; Rafuse et al., 2000; Walshet al., 2000). While we cannot rule out that some of theseproteins are directly glycosylated with the CT antigen,some of the proteins affected by CT expression do notappear to comigrate with glycoproteins bearing the carbo-hydrate (Figs. 1 and 6). Thus, it is likely that at least someof these proteins are not targeted by direct glycosylationwith the CT carbohydrate. Second, CT expression alterspostnatal myofiber growth (Fig. 2 and Table 1). This appearsto be due in part to a failure of satellite fusion withpreexisting myofibers (Figs. 2 and 3). Satellite cells havebeen reported to be concentrated about 20-fold in areassurrounding the neuromuscular junction (Wokke et al.,1989). If such an accumulation involves signals from thesynaptic matrix or membrane, the increased extracellularexpression of such signals in the CT transgenic mice mayincrease satellite cell numbers in extrasynaptic areas. It isalso possible that the increased expression of synapticlaminins or NCAM may inhibit satellite cell fusion intomyotubes and, in doing so, increase their number. Third,the CT carbohydrate alters neuromuscular structure (Fig. 4and Table 1). The loss of secondary folds in the muscle
FIG. 6. Increased expression of synaptic proteins in CT transgenic(20 mg) were compared in wild type (Wt) and transgenic (Tg) mice. Alaminin a4 (200 kDa), laminin a5 (;400 kDa), utrophin (;400 kDaalso increased for desmin (53 kDa) and dystrobrevin (90 and 65lower-molecular-weight bands (;50 kDa for desmin and 44 kDakDa), laminin b1 (;220 kDa), and the epsilon chain of the acetylc
membrane, the invasion into the synaptic cleft of the
© 2002 Elsevier Science. A
Schwann cell processes, and the misalignment of activezones and secondary folds are similar to findings in micelacking laminin b2 (Noakes et al., 1995; Patton et al., 1998),laminin a2 (Ellisman, 1981), or laminin a4 (Patton et al.,2001). Therefore, as CT transgenic mice display alteredexpression of several laminin chains, redistribution of lami-nins a4 and a5 may contribute to these phenotypes. Finally,the CT carbohydrate alters synaptic topography (Fig. 4). Thepattern of neuromuscular junctions was altered, both onindividual myofibers and on the muscle as a whole. Sprout-ing of axons is also seen in transgenic mice that overexpressNCAM (Walsh et al., 2000). As CT transgenic mice alsooverexpress NCAM in extrasynaptic regions, NCAM wouldappear to be a logical candidate to mediate these pheno-types.
The finding that a-dystroglycan can be glycosylated withthe CT antigen in muscle cell extracts (Fig. 7) points to apotential mechanism that would explain the misexpres-sion of both laminin chains and utrophin, as dystroglycanbinds to these proteins. a-Dystroglycan isolated fromskeletal muscle contains a unique carbohydrate struc-ture, NeuAca2,3Galb1,4GlcNAcb1,2Mana-O-Ser, on itsmucin domain (Sasaki et al., 1998). The terminalNeuAca2,3Galb1,4GlcNAc portion of this structure should
e. Western blots of equivalent amounts of total muscle cell proteinnts of total protein were increased for c-met (50 kDa, N-terminal),NCAM (160–180 kDa) in CT transgenic mice. Protein levels were), but in both cases this was due to the presence of additionalstrobrevin). Levels of dystrophin (;400 kDa), b-dystroglycan (43e receptor (AChR, ;60 kDa) were unchanged.
micmou
), andkDa
for dy
be a good substrate for the CT GalNAc transferase (Smith
ll rights reserved.

70 Xia et al.
and Lowe, 1994). a-Dystroglycan can bind extrasynapticlaminin chains, such as laminin a2 and the embryonicallyexpressed laminin a1 (Talts et al., 1999), and can bind withapparently weaker affinity to laminin a4 (Talts et al., 2000).Its transmembrane binding partner, b-dystroglycan, like-wise can bind both the extrasynaptic cytoplasmic protein
FIG. 7. Glycosylation of a-dystroglycan with the CT antigen in skextract from wild type (Wt) and CT transgenic (CT) showed that smuscles. Nonionic detergent extracts of skeletal muscle from CT(CT1) or with an anti-serum against dystroglycan (OR12). Antibanti-mouse IgM-agarose (for CT1), or extracts were incubated withWestern blotting using antibodies to a-dystroglycan (IIH6) or the Cto a-dystroglycan and the CT antigen when precipitated with eithfor the CT1 precipitation and subsequent immunoblot (asterisk). (B)were precipitated by WFA agarose, a lectin that binds bGalNAc. Wthat could be identified by antibodies to the CT antigen (CT1) and aexperiments that was identified by antibodies to b-dystroglycanproteins was precipitated from an identical amount of protein extrbe blocked by addition of excess GalNAc (1GalNAc).
dystrophin (Ervasti and Campbell, 1991) and its synaptic
© 2002 Elsevier Science. A
counterpart, utrophin (James et al., 2000). In adult animals,utrophin (Ohlendieck et al., 1991) and laminins a4 and a5(Patton et al., 1997) are normally present only at theneuromuscular junction, while a- and b-dystroglycan arepresent both at the synapse and in extrasynaptic regions(Ohlendieck et al., 1991). Therefore, it is highly likely that
al muscles of CT transgenic mice. (A) Immunoblots of muscle celll fragments of a-dystroglycan (aDG) were increased in transgenicgenic mice were precipitated with an antibody to the CT antigenbound complexes were precipitated by Protein-G (for OR12) orns alone (control, C). Precipitated proteins were then subjected toigen (CT1). Bands at 160 and 35 kDa were identified by antibodiestibody. Secondary antibody alone recognized the IgM heavy chainionic detergent extracts of wild type (Wt) or transgenic (CT) muscleprecipitated a 160-kDa mucinlike protein from transgenic muscletroglycan (VIN4-1). A 43-kDa band was also coprecipitated in these7), but it did not coblot with anti-CT antibodies. None of these
from wild type muscle. Binding of a-dystroglycan to WFA could
eleteveratransody-resi
T anter anNonFA
-dys(DG6acted
there is something about dystroglycan at the neuromuscu-
ll rights reserved.

71Overexpression of the CT GalNAc Transferase
lar junction that defines it as a synaptic form of the protein.The presence of the synaptic CT glycan on a-dystroglycanin CT transgenic muscles suggests that the CT antigen isthe synaptic modifier of a-dystroglycan that defines it as asynaptic protein. As the neuromuscular junction representssuch a small amount of the myofiber membrane, it may notbe possible to identify CT-containing a-dystroglycan inwild type mice. Indeed, we have not yet been able to do so.Nevertheless, the presence of CT on a-dystroglycan in CTtransgenic muscle suggests that it is normally present ona-dystroglycan at the synapse, and that this may give itunique properties in binding to or localizing synaptic lami-nins and utrophin. Overexpression of “synaptic” dystrogly-can in CT transgenic mice would then allow expression ofutrophin and laminins a4 and a5 along myofibers wherethis normally would not occur.
While the molecular mechanism for the developmentaleffects shown here has yet to be determined, the dramaticalterations in muscle growth and neuromuscular structureexhibited by animals overexpressing the CT antigen sug-gests that it is important for neuromuscular development.We are hopeful that similar transgenic approaches may beused to identify functions for other synaptic glycans whereno function has been described in vivo. The CT antigen, forexample, is also expressed on the presynaptic membrane atthe neuromuscular junction (Martin et al., 1999), and nofunction has yet been described for it there. Likewise,terminal b3-linked GalNAc structures, including globoside(Martin et al., 1999) and GalNAc phosphate structures(Scott et al., 1999), appear to be expressed at the mamma-lian neuromuscular junction. Therefore, other synapticGalNAc forms may be present on other proteins and orga-nize different developmental events. Finally, high mannosestructures are localized at excitatory synapses in the brain(Matus et al., 1973; Gurd and Fu, 1982; Clark et al., 1998),and transgenic approaches may be useful in elucidatingtheir role in synapse formation as well.
ACKNOWLEDGMENTS
We thank Leo LeFrancois (University of Connecticut), JohnLowe (HHMI, University of Michigan), Jeff Chamberlin (HHMI,University of Michigan), Kevin Campbell (HHMI, University ofIowa), Dale Abrahamson (University of Alabama, Birmingham),Palmer Taylor (University of California, San Diego), and JoshuaSanes (Washington University) for gifts of reagents; and ChristineKim and Vianney Jayasinha for technical support. This work wasfunded by NIH Grant RR 04050 (to M.E.) and by March of Dimes,NARSAD, Muscular Dystrophy Association, and NIH (NS37214)grants (to P.T.M.). A.K. was sponsored in part by a postdoctoralfellowship from NINDS.
REFERENCES
Allbrook, D. B., Han, M. F., and Hellmuth, A. E. (1971). Populationof muscle satellite cells in relation to age and mitotic activity.
Pathology 3, 223–243.© 2002 Elsevier Science. A
Balsamo, J., Pratt, R. S., and Lilien, J. (1986). Chick neural retinaN-acetylgalactosaminyltransferase/acceptor complex: Catalysisinvolves transfer of N-acetylgalactosamine phosphate to endog-enous acceptors. Biochemistry 25, 5402–5407.
Bischoff, R. (1994). Satellite cells. In “Myology: Basic and Clinical”(A. Engel and C. Franzini-Armstrong, Eds.), Vol. 1. McGraw Hill,New York.
Cardasis, C. A., and Cooper, G. W. (1975). An analysis of nuclearnumbers in individual muscle fibers during differentiation andgrowth: A satellite cell-muscle fiber growth unit. J. Exp. Zool.191, 347–358.
Clark, R. A., Gurd, J. W., Bissoon, N., Tricaud, N., Molnar, E.,Zamze, S. E., Dwek, R. A., McIlhinney, R. A., and Wing, D. R.(1998). Identification of lectin-purified neural glycoproteins, GPs180, 116, and 110, with NMDA and AMPA receptor subunits:Conservation of glycosylation at the synapse. J. Neurochem. 70,2594–2605.
Conzelmann, A., and Lefrancois, L. (1988). Monoclonal antibodiesspecific for T cell-associated carbohydrate determinants reactwith human blood group antigens CAD and SDA. J. Exp. Med.167, 119–131.
Cote, P. D., Moukhles, H., Lindenbaum, M., and Carbonetto, S.(1999). Chimaeric mice deficient in dystroglycans develop mus-cular dystrophy and have disrupted myoneural synapses. Nat.Genet. 23, 388–342.
Covault, J., and Sanes, J. R. (1986). Distribution of N-CAM insynaptic and extrasynaptic portions of developing and adultskeletal muscle. J. Cell Biol. 102, 716–730.
Dohi, T., Yuyama, Y., Natori, Y., Smith, P. L., Lowe, J. B., andOshima, M. (1996). Detection of N-acetylgalactosaminyl-transferase mRNA which determines expression of Sda bloodgroup carbohydrate structure in human gastrointestinal mucosaand cancer. Int. J. Cancer 67, 626–631.
Eckhardt, M., Bukalo, O., Chazal, G., Wang, L., Goridis, C.,Schachner, M., Gerardy-Schahn, R., Cremer, H., and Dityatev, A.(2000). Mice deficient in the polysialyltansferase ST8SiaIV/PST-1 allow discrimination of the roles of neural cell adhesionmolecule protein and polysialic acid in neural development andsynaptic plasticity. J. Neurosci. 20, 5234–5244.
Ellisman, M. H. (1981). The membrane morphology of the neuro-muscular junction, sarcolemma, sarcoplasmic reticulum andtransverse tubule system in murine muscular dystrophy studiedby freeze-fracture electron microscopy. Brain Res. 214, 261–273.
Ervasti, J. M., and Campbell, K. P. (1991). Membrane organizationof the dystrophin-glycoprotein complex. Cell 66, 1121–1131.
Frey, D., Schneider, C., Xu, L., Borg, J., Spooren, W., and Caroni, P.(2000). Early and selective loss of neuromuscular synapse sub-types with low sprouting competence in motoneuron diseases.J. Neurosci. 20, 2534–2542.
Gagneux, P., and Varki, A. (1999). Evolutionary considerations inrelating oligosaccharide diversity to biological function. Glyco-biology 9, 747–755.
Gurd, J. W., and Fu, S. C. (1982). Concanavalin A receptorsassociated with rat brain synaptic junctions are high mannose-type oligosaccharides. J. Neurochem. 39, 719–725.
Hamalainen, N., and Pette, D. (1993). The histochemical profiles offast fiber types IIB, IID, and IIA in skeletal muscles of mouse, rat,and rabbit. J. Histochem. Cytochem. 41, 733–743.
Harlow, E., and Lane, D. (1988). “Antibodies: A LaboratoryManual.” Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, NY.ll rights reserved.

72 Xia et al.
Henry, M. D., and Campbell, K. P. (1999). Dystroglycan inside andout. Curr. Opin. Cell Biol. 11, 602–607.
Hoyk, Z. A., Parducz, A., and Theodosis, D. T. (2001). The highlysialylated isoform of the neural cell adhesion molecule is re-quired for estradiol-induced morphological synaptic plasticity inthe adult arcuate nucleus. Eur. J. Neurosci. 13, 649–656.
Hunter, D. D., Shah, V., Merlie, J. P., and Sanes, J. R. (1989). Alaminin-like adhesive protein concentrated in the synaptic cleftof the neuromuscular junction. Nature 338, 229–234.
James, M., Nuttall, A., Iisley, J. L., Ottersbach, K., et al. (2000).Adhesion-dependent tyrosine phosphorylation of beta-dystro-glycan regulates its interaction with utrophin. J. Cell Sci. 113,1717–1726.
Jessel, T. M., Hynes, M. A., and Dodd, J. (1990). Carbohydrates andcarbohydrate-binding proteins in the nervous system. Annu.Rev. Neurosci. 13, 227–255.
Lauri, S. E., Kaukinen, S., Kinnunen, T., Ylinen, A., Imai, S., Kaila,K., and Rauvala, H. (1999). Regulatory role and molecular inter-actions of a cell-surface heparan sulfate proteoglycan (N-syndecan) in hippocampal long-term potentiation. J. Neurosci.15, 1226–1235.
Lefrancois, L., and Bevan, M. J. (1985). Functional modifications ofcytotoxic T-lymphocyte T200 glycoprotein recognized by mono-clonal antibodies. Nature 314, 449–452.
Li, Z., Mericskay, M., Agbulut, O., Butler-Browne, G., Carlsson, L.,Thornell, L. E., Babinet, C., and Paulin, D. (1997). Desmin isessential for the tensile strength and integrity of myofibrils butnot for myogenic commitment, differentiation, and fusion ofskeletal muscle. J. Cell Biol. 139, 129–144.
Lin, S. S., and Levitan, I. B. (1987). Concanavalin A alters synapticspecificity between cultured Aplysia neurons. Science 237, 648–650.
Luthi, A., Laurent, J.-P., Figurov, A., Muller, D., and Schachner, M.(1994). Hippocampal long-term potentiation and neural celladhesion molecules L1 and NCAM. Nature 372, 777–779.
Margolis, R. U., and Margolis, R. K. (1989). “Neurobiology ofGlycoconjugates.” Academic Press, New York.
Martin, P. T., Kaufman, S. J., Kramer, R. H., and Sanes, J. R. (1996).Synaptic integrins in developing, adult, and mutant muscle:Selective association of alpha1, alpha7A, and alpha7B integrinswith the neuromuscular junction. Dev. Biol. 174, 125–139.
Martin, P. T., Scott, L. J., Porter, B. E., and Sanes, J. R. (1999).Distinct structures and functions of related pre- and postsynapticcarbohydrates at the mammalian neuromuscular junction. Mol.Cell. Neurosci. 13, 105–118.
Miner, J. H., Cunningham, J., and Sanes, J. R. (1998). Roles forlaminin in embryogenesis: Exencephaly, syndactyly, and placen-topathy in mice lacking the laminin alpha 5 chain. J. Cell Biol.143, 1713–1723.
Moscoso, L. M., Cremer, H., and Sanes, J. R. (1998). Organizationand reorganization of neuromuscular junctions in mice lackingneural cell adhesion molecule, tenascin-C, or fibroblast growthfactor-5. J. Neurosci. 18, 1465–1477.
Muramatsu, T. (2000). Essential roles of carbohydrate signals indevelopment, immune response, and tissue functions as revealedby gene targeting. J. Biochem. 127, 171–176.
Muscat, G. E., and Kedes, L. (1987). Multiple 59-flanking regions ofthe human alpha-skeletal actin gene synergistically modulatemuscle-specific expression. Mol. Cell Biol. 7, 4089–4099.
Matus, A., De Petris, S., and Raff, M. C. (1973). Mobility ofconcanavalin A receptors in myelin and synaptic membranes.
Nature 244, 278–280.© 2002 Elsevier Science. A
McMahan, U. J., Sanes, J. R., and Marshall, J. M. (1978). Cholines-terase is associated with the basal lamina at the neuromuscularjunction. Nature 271, 172–174.
Noakes, P. G., Gautam, M., Mudd, J., Sanes, J. R., and Merlie, J. P.(1995). Aberrant differentiation of neuromuscular junctions inmice lacking s-laminin/laminin beta 2. Nature 374, 258–262.
Ohlendieck, K., Ervasti, J. M., Matsumura, K., Kahl, S. D., Leveille,C. J., and Campbell, K. P. (1991). Dystrophin-related protein islocalized to neuromuscular junctions of adult skeletal muscle.Neuron 7, 499–508.
Parkhomovskiy, N., Kammesheidt, A., and Martin, P. T. (2000).N-acetyllactosamine and the CT carbohydrate antigen mediateagrin-dependent activation of MuSK and acetylcholine receptorclustering in skeletal muscle. Mol. Cell. Neurosci. 15, 380–397.
Patel, T. J., and Lieber, R. L. (1997). Force transmission in skeletalmuscle: From actomyosin to external tendons. Exerc. Sport Sci.Rev. 25, 321–363.
Patton, B. L., Chiu, A. Y., and Sanes, J. R. (1998). Synaptic lamininprevents glial entry into the synaptic cleft. Nature 393, 698–701.
Patton, B. L., Connolly, A. M., Martin, P. T., Cunningham, J. M.,Mehta, S., Pestronk, A., Miner, J. H., and Sanes, J. R. (1999).Distribution of ten laminin chains in dystrophic and regenerat-ing muscles. Neuromuscul. Disord. 9, 423–433.
Patton, B. L., Cunningham, J. M., Thyboll, J., Kortesmaa, J.,Westerblad, H., Edstrom, L., Tryggvason K., and Sanes, J. R.(2001). Properly formed but improperly localized synaptic spe-cializations in the absence of laminin alpha 4. Nat. Neurosci. 4,597–604.
Patton, B. L., Miner, J. H., Chiu, A. Y., and Sanes, J. R. (1997).Distribution and function of laminins in the neuromuscularsystem of developing, adult, and mutant mice. J. Cell Biol. 139,1507–1521.
Peters, M. F., Sadoulet-Puccia, H. M., Grady, M., Kramarcy, N. R.,Kunkel, L. M., Sanes, J. R., Sealock, R., and Froehner, S. C. (1998).Differential membrane localization and intermolecular associa-tions of a-dystrobrevin isoforms in skeletal muscle. J. Cell Biol.142, 1269–1278.
Rafuse, V. F., Polo-Parada, L., and Landmessser, L. (2000). Struc-tural and functional alterations in neuromuscular junctions inNCAM-deficient mice. J. Neurosci. 20, 6529–6539.
Rutishauser, U., and Landmesser, L. (1996). Polysialic acid in thevertebrate nervous system: Promoter of plasticity in cell–cellinteractions. Trends Neurosci. 19, 422–427.
Sandrock, A. W., Dryer, S. E., Rosen, K. M., Gozani, S. N., Kramer,R., Theill, L. E., and Fischbach, G. D. (1997). Maintenance ofacetylcholine receptor number by neuregulins at the neuromus-cular junction in vivo. Science 276, 599–603.
Sanes, J. R., and Cheney, J. M. (1982). Lectin binding reveals asynapse-specific carbohydrate in skeletal muscle. Nature 300,646–647.
Sasaki, T., Yamada, H., Matsumura, K., Shimizu, T., Kobata, A.,and Endo, T. (1998). Detection of O-mannosyl glycans in rabbitskeletal muscle alpha-dystroglycan. Biochim. Biophys. Acta1425, 599–606.
Schmalbruch, H., and Hellhammer, U. (1977). The number ofnuclei in adult rat muscles with special reference to satellitecells. Anat. Rec. 189, 169–175.
Scott, L. J., Bacou, F., and Sanes, J. R. (1988). A synapse-specificcarbohydrate at the neuromuscular junction: Association withboth acetylcholinesterase and a glycolipid. J. Neurosci. 8, 932–
944.ll rights reserved.

73Overexpression of the CT GalNAc Transferase
Scott, L. J., Balsamo, J., Sanes, J. R., and Lilien, J. (1990). Synapticlocalization and neural regulation of an N-acetylgalactosaminyltransferase in skeletal muscle. J. Neurosci. 10, 346–350.
Seale, P., Sabourin, L. A., Girgis-Gabardo, A., Mansouri, A., Gruss,P., and Rudnicki, M. A. (2000). Pax7 is required for the specifi-cation of myogenic satellite cells. Cell 102, 777–786.
Seki, T., and Rutishauser, U. (1998). Removal of polysialic acid-neural cell adhesion molecule induces aberrant mossy fiberinnervation and ectopic synaptogenesis in the hippocampus.J. Neurosci. 18, 3757–3766.
Smith, P. L., and Lowe, J. B. (1994). Molecular cloning of a murineN-acetylgalactosamine transferase cDNA that determines ex-pression of the T lymphocyte-specific CT oligosaccharide differ-entiation antigen. J. Biol. Chem. 269, 15162–15171.
Tai, M.-H., and Zipser, B. (1998). Mannose-specific recognitionmediates two aspects of synaptic growth of leech sensory affer-ents: Collateral branching and proliferation of synaptic vesicleclusters. Dev. Biol. 201, 154–166.
Talts, J. F., Andac, Z., Gohring, W., Brancaccio, A., and Timpl, R.(1999). Binding of the G domains of laminin alpha1 and alpha2chains and perlecan to heparin, sulfatides, alpha-dystroglycanand several extracellular matrix proteins. EMBO J. 18, 863–870.
Talts, J. F., Sasaki, T., Miosge, N., Gohring, W., Mann, K., Mayne,
R., and Timpl, R. (2000). Structural and functional analysis of the© 2002 Elsevier Science. A
recombinant G domain of the laminin alpha 4 chain and itsproteolytic processing in tissues. J. Biol. Chem. 275, 35192–35199.
Theodosis, D. T., Bonhomme, R., Vitiello, S., Rougon, G., andPoulain, D. A. (1999). Cell surface expression of polysialic acid onNCAM is a prerequisite for activity-dependent morphologicalneuronal and glial plasticity. J. Neurosci. 19, 10228–10236.
Walsh, F. S., Hobbs, C., Wells, D. J., Slater, C. R., and Fazeli, S.(2000). Ectopic expression of N-CAM in skeletal muscle oftransgenic mice results in terminal sprouting of the neuromus-cular junction and altered structure but not function. Mol. Cell.Neurosci. 15, 241–261.
Wokke, J. H., Van den Oord, C. J., Leppink, G. J., and Jennekens,F. G. (1989). Perisynaptic satellite cells in human externalintercostal muscle: A quantitative and qualitative study. Anat.Rec. 223, 174–180.
Yamaguchi, Y. (2001). Heparan sulfate proteoglycans in the ner-vous system: Their diverse roles in neurogenesis, axon guidance,and synaptogenesis. Semin. Cell Dev. Biol. 12, 99–106.
Received for publication July 29, 2001Revised October 27, 2001
Accepted October 27, 2001
Published online January 2, 2002ll rights reserved.



![Laminin-332 and Integrins: Signaling Platform for …Laminin-332 and Integrins 31 laminin globular (LG) subdomains (LG1-5) [2]. The latter is the major interaction sites for cell surface](https://static.fdocuments.net/doc/165x107/5f712e9e3f945d798f112220/laminin-332-and-integrins-signaling-platform-for-laminin-332-and-integrins-31-laminin.jpg)














