
Organization and PprB-Dependent Control of the … specific/Bernard,2009.pdf · levels and nutrient...
13
JOURNAL OF BACTERIOLOGY, Mar. 2009, p. 1961–1973 Vol. 191, No. 6 0021-9193/09/$08.000 doi:10.1128/JB.01330-08 Copyright © 2009, American Society for Microbiology. All Rights Reserved. Organization and PprB-Dependent Control of the Pseudomonas aeruginosa tad Locus, Involved in Flp Pilus Biology ‡ Christophe S. Bernard, Christophe Bordi, Elise Termine, Alain Filloux,† and Sophie de Bentzmann* Membrane Complexes and Pathogenicity in Pseudomonas aeruginosa, CNRS LISM (UPR9027)—Aix-Marseille Universite ´, 31 Chemin Joseph Aiguier, 13402 Marseille Cedex 20, France Received 23 September 2008/Accepted 2 January 2009 Bacterial attachment to the substratum involves several cell surface organelles, including various types of pili. The Pseudomonas aeruginosa Tad machine assembles type IVb pili, which are required for adhesion to abiotic surfaces and to eukaryotic cells. Type IVb pili consist of a major subunit, the Flp pilin, processed by the FppA prepilin peptidase. In this study, we investigated the regulatory mechanism of the tad locus. We showed that the flp gene is expressed late in the stationary growth phase in aerobic conditions. We also showed that the tad locus was composed of five independent transcriptional units. We used transcriptional fusions to show that tad gene expression was positively controlled by the PprB response regulator. We subsequently showed that PprB bound to the promoter regions, directly controlling the expression of these genes. We then evaluated the contribution of two genes, tadF and rcpC, to type IVb pilus assembly. The deletion of these two genes had no effect on Flp production, pilus assembly, or Flp-mediated adhesion to abiotic surfaces in our conditions. However, our results suggest that the putative RcpC protein modifies the Flp pilin, thereby promoting Flp-dependent adhesion to eukaryotic cells. Pseudomonas aeruginosa is an opportunistic pathogen re- sponsible for severe infections in immunocompromised pa- tients and individuals suffering from cystic fibrosis. Post- genomic studies have focused attention on the ability of P. aeruginosa to bind to various surfaces. This binding involves combinations of type IVa (3) and type IVb (4) pili, flagella, and Cup fimbriae (26, 32) on the surface of the bacterium, the release of adhesive proteins via secretion systems (8), and the production of an exopolysaccharide matrix (9, 15, 22, 34). Most type IVb pilins have similar characteristics (3). The 15- to 30-residue leader peptide of the prepilin is processed downstream from a conserved hydrophobic residue, generating a mature pilin of 190 amino acids in length. The resulting exposed D region in assembled type IVb pili is large. The type IVb pilin of Vibrio cholerae, TcpA, assembles as a left-handed, three-start helix with six subunits per turn (3). These features result in type IVb pili being thicker than type IVa pili. The type IVb pilin family in- cludes a clearly monophyletic Flp (fimbrial low-molecular-weight protein) prepilin subfamily, initially described for Aggregatibacter (Actinobacillus) actinomycetemcomitans. Members of this subfam- ily have a unique set of features in common: a long leader pep- tide, a short mature pilin (50 to 80 amino acids), and an Flp motif consisting of 20 hydrophobic residues at the N terminus of the mature pilin, with adjacent glutamate and tyrosine residues at the center of this motif (19). The flp gene is linked to the tad (tight adhesion) locus, which encodes a macromolecular machine ded- icated to the assembly of adhesive pili of the type IVb Flp sub- family (17, 18, 25). Similar tad loci have been identified in archaea, actinobacteria, and gram-negative bacteria, usually as a single copy, but in up to four copies in some species (19, 31). Several conserved elements have been identified in Tad machines to date: the TadA trafficking NTPase (17, 24), the RcpA secretin, one or two inner membrane PilC homologues, TadB and TadC, the TadV/FppA prepilin peptidase (4, 30), the Flp pilin, and the TadE and TadF pseudopilins (30). These components are also shared by type IVa pilus machines (T4P) and type II secretion systems (T2S). However, the presence within the tad locus of several specific genes encoding proteins with no homologue in the T4P and T2S systems, such as RcpC, RcpB, TadZ, and TadG, suggests that Tad machines represent a new type of prokaryotic secretion system (31). The tad genes are usually organized linearly in a single direction, suggesting that they may constitute an operon. This is likely, as the tad locus of A. actinomycetemcomi- tans is transcribed as a polycistronic mRNA (10). Classical tad loci start with the flp gene, followed by the gene encoding the prepilin peptidase (tadV) and the remaining genes encoding the Tad ma- chine elements. The tad locus of P. aeruginosa has a distinctive genetic organization, with four or five putative transcriptional units operating in different directions: the flp gene, nine genes (rcpC-tadG) constituting the rcp-tad locus, the gene encoding the TadV/FppA prepilin peptidase, the gene encoding the response regulator PprB, and the tadF gene (4). The divergent orientations of the flp and rcp-tad genes and the requirement for flp overex- pression for efficient Flp pilus assembly (4) suggest that different regulatory processes are involved in Flp production, maturation, and assembly. Environmental conditions, such as O 2 and CO 2 * Corresponding author. Mailing address: Membrane Complexes and Pathogenicity in Pseudomonas aeruginosa, CNRS LISM (UPR9027)— Aix-Marseille Universite ´, 31 Chemin Joseph Aiguier, 13402 Marseille Cedex 20, France. Phone: 33-491164125. Fax: 33-491712124. E-mail: [email protected]. ‡ Supplemental material for this article may be found at http://jb .asm.org/. † Present address: Centre for Molecular Microbiology and Infec- tion, Division of Cell and Molecular Biology, Flowers Building, South Kensington Campus, Imperial College London, London SW7 2AZ, United Kingdom. Published ahead of print on 16 January 2009. 1961
Transcript of Organization and PprB-Dependent Control of the … specific/Bernard,2009.pdf · levels and nutrient...

JOURNAL OF BACTERIOLOGY, Mar. 2009, p. 1961–1973 Vol. 191, No. 60021-9193/09/$08.00�0 doi:10.1128/JB.01330-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Organization and PprB-Dependent Control of thePseudomonas aeruginosa tad Locus, Involved
in Flp Pilus Biology�‡Christophe S. Bernard, Christophe Bordi, Elise Termine, Alain Filloux,† and Sophie de Bentzmann*
Membrane Complexes and Pathogenicity in Pseudomonas aeruginosa, CNRS LISM (UPR9027)—Aix-Marseille Universite,31 Chemin Joseph Aiguier, 13402 Marseille Cedex 20, France
Received 23 September 2008/Accepted 2 January 2009
Bacterial attachment to the substratum involves several cell surface organelles, including various types ofpili. The Pseudomonas aeruginosa Tad machine assembles type IVb pili, which are required for adhesion toabiotic surfaces and to eukaryotic cells. Type IVb pili consist of a major subunit, the Flp pilin, processed bythe FppA prepilin peptidase. In this study, we investigated the regulatory mechanism of the tad locus. Weshowed that the flp gene is expressed late in the stationary growth phase in aerobic conditions. We also showedthat the tad locus was composed of five independent transcriptional units. We used transcriptional fusions toshow that tad gene expression was positively controlled by the PprB response regulator. We subsequentlyshowed that PprB bound to the promoter regions, directly controlling the expression of these genes. We thenevaluated the contribution of two genes, tadF and rcpC, to type IVb pilus assembly. The deletion of these twogenes had no effect on Flp production, pilus assembly, or Flp-mediated adhesion to abiotic surfaces in ourconditions. However, our results suggest that the putative RcpC protein modifies the Flp pilin, therebypromoting Flp-dependent adhesion to eukaryotic cells.
Pseudomonas aeruginosa is an opportunistic pathogen re-sponsible for severe infections in immunocompromised pa-tients and individuals suffering from cystic fibrosis. Post-genomic studies have focused attention on the ability of P.aeruginosa to bind to various surfaces. This binding involvescombinations of type IVa (3) and type IVb (4) pili, flagella, andCup fimbriae (26, 32) on the surface of the bacterium, therelease of adhesive proteins via secretion systems (8), and theproduction of an exopolysaccharide matrix (9, 15, 22, 34).
Most type IVb pilins have similar characteristics (3). The 15- to30-residue leader peptide of the prepilin is processed downstreamfrom a conserved hydrophobic residue, generating a mature pilinof �190 amino acids in length. The resulting exposed D region inassembled type IVb pili is large. The type IVb pilin of Vibriocholerae, TcpA, assembles as a left-handed, three-start helix withsix subunits per turn (3). These features result in type IVb pilibeing thicker than type IVa pili. The type IVb pilin family in-cludes a clearly monophyletic Flp (fimbrial low-molecular-weightprotein) prepilin subfamily, initially described for Aggregatibacter(Actinobacillus) actinomycetemcomitans. Members of this subfam-ily have a unique set of features in common: a long leader pep-tide, a short mature pilin (50 to 80 amino acids), and an Flp motif
consisting of 20 hydrophobic residues at the N terminus of themature pilin, with adjacent glutamate and tyrosine residues at thecenter of this motif (19). The flp gene is linked to the tad (tightadhesion) locus, which encodes a macromolecular machine ded-icated to the assembly of adhesive pili of the type IVb Flp sub-family (17, 18, 25). Similar tad loci have been identified inarchaea, actinobacteria, and gram-negative bacteria, usually as asingle copy, but in up to four copies in some species (19, 31).Several conserved elements have been identified in Tad machinesto date: the TadA trafficking NTPase (17, 24), the RcpA secretin,one or two inner membrane PilC homologues, TadB and TadC,the TadV/FppA prepilin peptidase (4, 30), the Flp pilin, and theTadE and TadF pseudopilins (30). These components are alsoshared by type IVa pilus machines (T4P) and type II secretionsystems (T2S). However, the presence within the tad locus ofseveral specific genes encoding proteins with no homologue in theT4P and T2S systems, such as RcpC, RcpB, TadZ, and TadG,suggests that Tad machines represent a new type of prokaryoticsecretion system (31). The tad genes are usually organized linearlyin a single direction, suggesting that they may constitute anoperon. This is likely, as the tad locus of A. actinomycetemcomi-tans is transcribed as a polycistronic mRNA (10). Classical tad locistart with the flp gene, followed by the gene encoding the prepilinpeptidase (tadV) and the remaining genes encoding the Tad ma-chine elements. The tad locus of P. aeruginosa has a distinctivegenetic organization, with four or five putative transcriptionalunits operating in different directions: the flp gene, nine genes(rcpC-tadG) constituting the rcp-tad locus, the gene encoding theTadV/FppA prepilin peptidase, the gene encoding the responseregulator PprB, and the tadF gene (4). The divergent orientationsof the flp and rcp-tad genes and the requirement for flp overex-pression for efficient Flp pilus assembly (4) suggest that differentregulatory processes are involved in Flp production, maturation,and assembly. Environmental conditions, such as O2 and CO2
* Corresponding author. Mailing address: Membrane Complexes andPathogenicity in Pseudomonas aeruginosa, CNRS LISM (UPR9027)—Aix-Marseille Universite, 31 Chemin Joseph Aiguier, 13402 MarseilleCedex 20, France. Phone: 33-491164125. Fax: 33-491712124. E-mail:[email protected].
‡ Supplemental material for this article may be found at http://jb.asm.org/.
† Present address: Centre for Molecular Microbiology and Infec-tion, Division of Cell and Molecular Biology, Flowers Building, SouthKensington Campus, Imperial College London, London SW7 2AZ,United Kingdom.
� Published ahead of print on 16 January 2009.
1961

levels and nutrient availability, have been reported to affect Flpproduction in Aggregatibacter aphrophilus and A. actinomycetem-comitans (27, 31). In P. aeruginosa, global approaches have shownthat some tad genes are either regulated by quorum sensing (28,38, 39) or are under the control of the VqsR master regulator (16)or the MvaT transcriptional regulator (33). The fppA and tadFgenes are located between the pprA and pprB genes. These genesencode a classical two-component system (TCS) that was thoughtto act as a master regulator of quorum sensing (6, 35) until therecent identification of a secondary mutation in the LasR regu-lator in the transposon pprB mutant (6). Nevertheless, this TCShas been reported to play a key role in membrane permeabilityregulation and drug resistance in P. aeruginosa (40). The proxim-ity of this TCS to the rcp-tad locus suggests it may be involved inthe transcriptional regulation of type IVb pilus gene expression.
We determined the conditions under which the flp genewas expressed in P. aeruginosa and showed that flp geneexpression occurred late in the stationary growth phase inaerobic conditions. We determined the operon structure ofthe tad locus and assessed the potential regulatory role ofthe TCS PprAB. We demonstrated that the tad locus wasorganized into five transcriptional units and positively reg-ulated by the PprB response regulator. We also studied theproducts of several genes encoding components of the Tadmachine, including TadF, which is probably the only pseu-dopilin in this system, and RcpC, a putative protein from afamily of bacterial proteins containing two �-clip domainsand unique to Tad machines.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The strains and plasmids used in thisstudy are listed in Table 1. Strains were grown at 30°C or 37°C in L broth, in thepresence of 100 �M isopropyl-�-D-thiogalactopyranoside (IPTG). The Esche-richia coli TG1, MG1655, TOP10F�, and CC118(�pir) strains were used forstandard genetic manipulations. Recombinant plasmids were introduced into P.aeruginosa, using the conjugative properties of pRK2013. Transconjugants wereselected on Pseudomonas isolation agar (Difco Laboratories) supplemented withappropriate antibiotics. The following antibiotic concentrations were used: (i) forE. coli, ampicillin at 50 �g/ml, kanamycin at 25 �g/ml, tetracycline at 15 �g/ml,and streptomycin at 50 �g/ml, and (ii) for P. aeruginosa, carbenicillin at 250�g/ml, tetracycline at 200 �g/ml, and streptomycin at 2 mg/ml.
Construction of P. aeruginosa deletion mutants. PCR was used to generate a550-bp DNA fragment upstream (Up) from the pprB, rcpC, and tadF genes withthe DelPBUp5-DelPBUp3, DelRCUp5-DelRCUp3, and DelTFUp5-DelTFUp3oligonucleotide pairs, respectively (Table 2), and a 550-bp DNA fragment down-stream (Dn) from the pprB, rcpC, and tadF genes, using the DelPBDn5-DelPBDn3, DelRCDn5-DelPRCDn3, and DelTFDn5-DelTFDn3 oligonucleo-tide pairs, respectively (Table 2). The resulting DNA fragments were insertedseparately into the pCR2.1 vector. The Up and Dn DNA fragments for the pprB,rcpC, and tadF genes, bearing appropriate sites—SacI/KpnI and KpnI/BamHI—were digested and inserted into the suicide vector pEX18, previously digestedwith SacI/BamHI, via a three-partner procedure. The recombinant plasmidswere introduced into P. aeruginosa, and the deletion mutants were obtained bydouble selection on LB agar supplemented with Irgasan (25 �g/ml) and carben-icillin (250 �g/ml) at 37°C and NaCl-free LB agar containing 6% sucrose at 30°C(20).
Chromosomal transcriptional fusions. DNA fragments containing the puta-tive promoter regions between the flp gene and the rcp-tad locus (501 bp) and thefppA and the tadF genes (150 bp) were amplified by PCR, using the PromFlRUp-PromFlRDn and PromFpTUp-PromFpTDn oligonucleotide pairs, respectively(Table 2). The DNA fragments were inserted into pCR2.1, excised by digestionwith KpnI/XhoI or XhoI/HindIII, and inserted into the miniCTX-lacZ vector(which is a miniCTX1-derived vector [1]), generating the miniCTX-pflp-lacZ andminiCTX-prcpC-lacZ constructs, or they were inserted into the BamHI/XhoI orXhoI/KpnI sites of the miniCTX-lacZ vector, thus generating the miniCTX-pfppA-lacZ and miniCTX-ptadF-lacZ constructs. These plasmids were used to
generate chromosomal flp-lacZ, rcpC-lacZ, fppA-lacZ, and tadF-lacZ fusions inthe PAO1 strain, as previously described (11, 12). The expression of the varioustranscriptional fusions was monitored by assaying galactosidase activity in at leastthree independent experiments.
Expression of tad fusions in the E. coli heterologous host. Both plasmids (i.e.,miniCTX-pflp-lacZ, miniCTX-prcpC-lacZ, miniCTX-pfppA-lacZ, or miniCTX-ptadF-lacZ and pMMB67-HE or pMMBpprB vectors) were introduced into theMG1655 strain of E. coli by transformation. After double selection on agar platescontaining appropriate antibiotics, cells were cultured and induced by incubationwith 0.1 mM IPTG for 1 h. The expression of the various transcriptional fusionswas monitored by assaying galactosidase activity in three independent experi-ments.
Cloning procedures for the rcpC and pprB genes. A 1,411-bp DNA fragmentencompassing the putative promoter region of the rcpC gene upstream from thercp-tad locus and the rcpC gene itself was amplified by PCR with the PromFlRDnand RcpCDn oligonucleotides (Table 2). It was inserted into pCR2.1, excised byHindIII/XhoI digestion, and inserted into the miniCTX-lacZ vector, thus gen-erating the miniCTX-rcpC construct. This construction was used for cis comple-mentation experiments in the rcpC mutants to generate the PAO1�rcpCattB::rcpC and PAO1�pilA�fliC�rcpC attB::rcpC strains. In contrast to the pro-cedure described for lacZ fusions, the FRT cassette excision step was not per-formed, resulting in the generation of a strain with tetracycline resistance for themonitoring of complementation. The pprB gene was amplified by PCR, using thePprBUp and PprBDn oligonucleotides (Table 2). It was inserted into pCR2.1and then excised and inserted between the XbaI and SacI sites of the broad-host-range vector pMMB67-HE.
EMSAs. The His-tagged version of the PprB (PprB-6His) protein was producedin the E. coli M15 strain after induction with 0.5 mM IPTG for 4 h at 28°C and waspurified as recommended (40). Electrophoretic mobility shift assays (EMSAs) wereperformed as follows. The intergenic DNA regions encompassing the promoterregions of the five transcriptional units identified in the tad locus were amplified byPCR with the corresponding oligonucleotide pairs (PromFlRUp-PromFlRDn,PromFpTlUp-PromFpTlDn, and 4296-95U40–4296-95R), generating DNA frag-ments of 501, 561, and 560 bp for the three intergenic regions upstream from theflp-rcp, fppA-tadF, and pprB genes, respectively. The 360-bp DNA region corre-sponding to the promoter region of the rsmZ gene was amplified by PCR with thePromrsmZUp and PromrsmZDn oligonucleotides and was used as a negative con-trol. The mixture of PCR products obtained, consisting of the intergenic DNAregion of interest and the negative control (1:2) in a 50 mM Tris-HCl buffer, pH 8.2,containing 1 mM EDTA and 0.25 mM saccharose, was incubated for 30 min at roomtemperature with various amounts of purified PprB-6His protein, ranging from 0 to2.34 �M. The corresponding mixtures were run on native 12% acrylamide gels andstained with ethidium bromide.
Tris-glycine gel electrophoresis and Western blot analysis. Flp production wasanalyzed by electrophoresis in a 16.5% polyacrylamide Tris-glycine gel, as pre-viously described (4). Bacterial cells were cultured and mixed with loading buffer(the equivalent of 0.025 OD600 [optical density at 600 nm] units per �l). Thesamples were treated with benzonase (1 unit/�l) for 30 min at 37°C and boiledfor 10 min, and the proteins were separated by electrophoresis. Proteins wereblotted onto nitrocellulose membranes. Flp protein was immunodetected withthe polyclonal antibody against Flp, used at a dilution of 1:150. A peroxidase-conjugated goat anti-rabbit immunoglobulin G was used at a dilution of 1:5,000in Tris-buffered saline supplemented with 10% milk and 0.1% Tween for thedetection of primary antibody binding to the Flp protein.
TEM. Aliquots of bacteria were collected for transmission electron microscopy(TEM). The bacterial pellet was obtained by centrifugation and resuspended in 0.15M NaCl in 10 mM Tris, pH 7.8 (Tris-NaCl). A drop of the bacterial suspension wasplaced on Formvar- and carbon-coated copper grids and left for approximately 5min. Grids were then fixed by incubation with 4% paraformaldehyde for 5 min andrinsed twice with Tris-NaCl, for 5 min each. Grids were then incubated with 5%bovine serum albumin (BSA) in Tris-NaCl for 10 min and then for 45 min with theFlp antiserum at a dilution of 1:150 in 0.5% BSA in Tris-NaCl. Grids were finallyincubated for 30 min with 10 nm colloidal gold-conjugated protein A in 0.5% BSAin Tris-NaCl. Grids were washed several times in Tris-NaCl and then in water, andthey were then immersed in a drop of 1% uranyl acetate for 1 min. Grids wereexamined in a JEOL 1200EX TEM operating at 80.0 kV.
The lengths of Flp pili observed in bacteria with different genetic backgroundswere determined by measurements of 30 pictures for each strain, systematicallyacquired from at least three independent experiments. Statistical analysis wasbased on unpaired t tests (GraphPad Prism 4 software).
Biofilm formation and confocal microscopy. Bacteria were grown in M63 me-dium supplemented with 0.4% L-arginine and 1 mM MgSO4, at 30°C, withoutshaking. Coverslips were partially immersed in the bacterial cultures in Falcon tubes
1962 BERNARD ET AL. J. BACTERIOL.

and were then processed for confocal microscopy. Before observation, coverslipswere fixed by incubation with 4% paraformaldehyde and stained by incubation with4�,6�-diamidino-2-phenylindole (DAPI) for 15 min. Slides were observed in anOlympus FV-1000 microscope, using a laser diode for excitation at 405 nm. Imageswere captured and processed with Fluoview and Amarys software.
Assay of adhesion to the surface of bronchial epithelial cells. 16HBE14o�human bronchial epithelial cells were incubated in 24-well microplates contain-ing coverslips in minimal essential medium supplemented with 10% fetal calf
serum and antibiotics for 24 h at 37°C, under an atmosphere containing 5% CO2.Four hours before infection, the medium was replaced by serum- and antibiotic-free medium. The epithelial cells were infected with bacteria at a multiplicity ofinfection of 30, for a period of 4 h at 37°C. The samples were rinsed twice withphosphate-buffered saline, fixed with 4% formaldehyde, stained with 0.1% crys-tal violet for 5 min, washed twice with water, dried, mounted, and sealed withEukitt mounting medium, and observed with an Axioscop 40 microscope (Zeiss).The bacteria adhering to epithelial cells (30 epithelial cells/assay) were counted
TABLE 1. Strains and plasmids used in this study
Strain or plasmid Relevant characteristicsa Source
StrainsE. coli
TG1 supE �(lac-proAB) thi hsdR�5 (F� traD36 rpoA�B� lacIqZ�M15) Lab collectionMG1655 K �lacZ pcnB::Kmr Lab collectionTOP10F� F� (lacIq Tn10 Tcr) mrcA �(mrr-hsdRMS-mcrBC) �80
lacZ�M15 �lacX74 recA1Invitrogen
CC118(�pir) Host strain for pKNG101 replication; �(ara-leu) araD �lacX74galE galK phoA20 thi-1 rpsE rpoB argE(Am) recA1 Rfr (�pir)
Lab collection
M15 lac ara gal mtl 40
P. aeruginosaPAO1 WT Lab collectionPAO1�flp PAO1 deletion mutant for the flp gene 4PAO1�pprB PAO1 deletion mutant for the pprB gene This studyPAO1�tadF PAO1 deletion mutant for the tadF gene This studyPAO1�rcpC PAO1 deletion mutant for the rcpC gene This studyPAO1�pilA�fliC PAO1 deletion mutant for the pilA, fliC genes 4PAO1�pilA�fliC�flp PAO1 deletion mutant for the pilA, fliC, flp genes This studyPAO1�pilA�fliC�rcpC PAO1 deletion mutant for the pilA, fliC, rcpC genes This studyPAO1�pilA�fliC�tadF PAO1 deletion mutant for the pilA, fliC, tadF genes This studyPAO1�rcpC attB::rcpC PAO1�rcpC strain cis complemented with WT rcpC gene
inserted at attB sitesThis study
PAO1�pilA�fliC�rcpC attB::rcpC PAO1�pilA�fliC�rcpC strain cis complemented with WT rcpCgene inserted at attB sites
This study
PAO1/flp-lacZ PAO1 strain with flp-lacZ fusion inserted at attB sites This studyPAO1/rcpC-lacZ PAO1 strain with rcpC-lacZ fusion inserted at attB sites This studyPAO1/fppA-lacZ PAO1 strain with fppA-lacZ fusion inserted at attB sites This studyPAO1/tadF-lacZ PAO1 strain with tadF-lacZ fusion inserted at attB sites This studyPAO1�pprB/flp-lacZ PAO1�pprB strain with flp-lacZ fusion inserted at attB sites This studyPAO1�pprB/rcptad-lacZ PAO1�pprB strain with rcpC-lacZ fusion inserted at attB sites This studyPAO1�pprB/fppA-lacZ PAO1�pprB strain with fppA-lacZ fusion inserted at attB sites This studyPAO1�pprB/tadF-lacZ PAO1�pprB strain with tadF-lacZ fusion inserted at attB sites This study
PlasmidspCR2.1 TA cloning vector for PCR products; lacZ� ColE1 f1 ori Apr
KmrInvitrogen
pMMB67-HE Broad-host-range vector; IncQ Ptac lacZ� Apr Lab collectionpEX18 Apr oriT� sacB�; gene replacement vector with MCS from
pUC18Lab collection
miniCTX-lacZ Tcr lacZ�; self-proficient integration vector with tet, V-FRT-attPMCS, ori, int, and oriT
12
pMMBpprB pprB gene cloned in pMMB67-HE, Apr This studypYW024 pprB gene cloned in pQE31 vector with a His tag, Apr 40miniCTX-pflp-lacZ Promoter region of flp gene cloned in miniCTX-lacZ at KpnI/
XhoI sitesThis study
miniCTX-prcpC-lacZ Promoter region of rcpC locus cloned in miniCTX-lacZ at XhoI/HindIII sites
This study
miniCTX-pfppA-lacZ Promoter region of fppA gene cloned into miniCTX-lacZ atXhoI/BamHI sites
This study
miniCTX-ptadF-lacZ Promoter region of tadF locus cloned into miniCTX-lacZ atXhoI/KpnI sites
This study
miniCTX-rcpC rcpC gene and its promoter region cloned into miniCTX-lacZ atXhoI/HindIII sites
This study
pEX18-DelpprB pprB gene mutator cloned into pEX18; Apr sacB� This studypEX18-DelrcpC rcpC gene mutator cloned into pEX18; Apr sacB� This studypEX18-DeltadF tadF gene mutator cloned into pEX18; Apr sacB� This studypRK2013 ColE1 ori tra� mob� Kmr Lab collection
a Apr, ampicillin resistance; Kmr, kanamycin resistance; Tcr, tetracycline resistance; Rfr, rifampin resistance; MCS, multiple-cloning site.
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1963

in randomly chosen microscopic fields from three independent assays. The dataobtained for each genetic background were compared, in a one-way analysis ofvariance and through unpaired t tests (GraphPad Prism 4 software).
Isolation of RNA and RT-PCR. The PAO1 strain, into which pMMBpprB wasintroduced, was cultured with 0.1 mM IPTG for 6 h at 37°C. Total cellular RNA
was isolated, using the PureYield RNA Midiprep system (Promega). Reversetranscription (RT)-PCR was carried out with the Access RT-PCR system (Pro-mega), according to the manufacturer’s instructions, but with the addition of 6%dimethyl sulfoxide to the RT-PCR mixture. We used a total of 1.8 �g of RNA orgenomic DNA. RT-PCR was carried out with gene-specific primers overlapping
TABLE 2. Oligonucleotides used for mutation engineering and gene cloning
Gene, mutation, or primer Additional designation Oligonucleotide (5�33�)a
pprBDelPBUp5 TATATAGAGCTCCCGTCTGGCTGAAAAGCATGGCCDelPBUp3 TATATAGGTACCTGACAGGCGCGATGGCCCGGDelPBDn5 TATATAGGTACCCATGGTCATACGCTCCATTTGDelPBDn3 TATATAGGATCCAACGTTTGCGCGTCTCCAAC
rcpCDelRCUp5 TATATAGAGCTCTTCTTCGTCCGCCAGGAATGCDelRCUp3 TATATAGGTACCCATGGGCAGACCCCTGCTATGDelRCDn5 TATATAGGTACCGGCAGCGCCATGACCCATGADelRCDn3 TATATAGGATCCACGAACGAGGTGCTGGCCTG
tadFDelTFUp5 TATATAGAGCTCCCATGGCGAATATAAAAGGGAATGDelTFUp3 TATATAGGTACCCATGAGCTGACATCCCCGATCDelTFDn5 TATATAGGTACCCAGACGCTGAAGGCGAGTGCCDelTFDn3 TATATAGGATCCATCTCCTGGACCGCCTCCGC
lacZ fusionsPromFlRUp CACCGCACCGAGCAGCAGCAGPromFlRDn TTCTTCGTCCGCCAGGAATGCPromFpTUp ACAAGGCCAGTGCTAATTGCPromFpTDn CGGAAAGCTTTTCCGAGCAT
ComplementationspprB
PprBUp TAAGCAAATGGAGCGTATGACCPprBDn GCGCCTGTCAGTGCACCAC
rcpCRcpCUp or PromFlRDn TTCTTCGTCCGCCAGGAATGCRcpCDn CCGGTGCTCCGATGCATATG
Operon structureb
1. rcpC-rcpA genes Primer 20; 4305-04U AGCAAGGACGAAGAACTCTACCPrimer 21; 4305-04R AAGCCGCGACGATCCAGCA
2. rcpA-tadZ genes Primer 22; 4304-03U TCGACAAGTTTCCCTGGCTGPrimer 23; 4304-03R CCGATACCAATCCCTCGAC
3. tadZ-tadA genes Primer 24; 4303-02U GAGCGCTACCTGCCCAATGTPrimer 25; 4303-02R AGTTGATGGCGCGAGCCGTA
4. tadA-tadB genes Primer 26; 4302-01U CTGATGATTTGCGCCGCGCTPrimer 27; 4302-01R CCAGTGACTGCTGCGCGG
5. tadB-tadC genes Primer 28; 4301-00U GCTCATCCACGAGCGCGAPrimer 29; 4301-00R ACCGAGATCAGGTACAGGGT
6. tadC-tadD genes Primer 30; 4300-4299U CTGCGGCGTGCTACGGCAPrimer 31; 4300-4299R GATCAGCGCCCTTTCCTCG
7. tadD-PA4298 genes Primer 32; 4299-98U AGCTGGATGCCCGGGACATPrimer 33; 4299-98R TAGCGCTGGTAGGCACGCT
8. PA4298-tadG genes Primer 34; 4298-97U AGCGTGCCTACCAGCGCTAPrimer 35; 4298-97R CGCGCTGGCTATGAAACTGC
9. tadG gene Primer 36; 4297-96U GAAAGCCGACGGTTCCGATCPrimer 37; 4297-96R1 CTAGATCAGCAGTTCCGCCC
10. tadG-pprB genes Primer 38; 4297-96R2 CACCCAGTTGGCGCTGGC11. pprB-fppA genes Primer 39; 4296-95U GGTTTCTGGTAGTAGTCGGC
Primer 40; 4296-95R GCCTTCGAAGGGCACCGTCA12. tadF-pprA genes Primer 41; 4294-93U CCAGGGACGAGTTGCTCGCA
Primer 42; 4294-93R CTGCATCGGCTGGAAACGAC
PromFpTlUp GGCTGAGGCAGACGCCTATAAPromFpTlDn GCCGGCATCCAGTCCAGGPromrsmZUp CCGGAATTCCCTTAGACCCACTGAAGACCPromrsmZDn GGGGTACCATCCTTCGGGGTTGCGTGTTCC
a Nucleotides corresponding to restriction sites are underlined.b Junction regions (1 to 12) between adjacent genes of the tad locus were amplified with primers 20 to 42.
1964 BERNARD ET AL. J. BACTERIOL.

junction regions between adjacent genes (listed in Table 2 and in Fig. 1A), witha T1 thermocycler (Biometra), using the following protocol: reverse transcriptionfor 45 min at 45°C, inactivation of reverse transcriptase by incubation at 94°C for2 min, followed by 45 cycles of PCR amplification with heating at 94°C for 30 s,60°C for 1 min, and 68°C for 2 min. We checked that RNA preparations were notcontaminated with DNA, by carrying out the same experiment without addingthe reverse transcriptase. The expected sizes of the amplicons for the variouspairs of primers were as follows: (1) 20-21, 460 bp; (2) 22-23, 480 bp; (3) 24-25,460 bp; (4) 26-27, 370 bp; (5) 28-29, 540 bp; (6) 30-31, 460 bp; (7) 32-33, 390 bp;
(8) 34-35, 450 bp; (9) 36-37, 300 bp (positive control); (10) 36-38, 390 bp(negative control); (11) 39-40, 560 bp; and (12) 41-42, 590 bp.
RESULTS
Insight into the genetic organization of the P. aeruginosa tadlocus. The P. aeruginosa tad locus is characterized by the di-vergent transcription of the flp gene, encoding the prepilin Flp,
FIG. 1. (A) Genetic organization of the P. aeruginosa tad locus and locations of the primers (see Table 2) used for genetic manipulations.Arrows indicate the five transcriptional units identified (I to V). Known functions of tad genes are specified below. Shown are the primersused for RT-PCR manipulations (indicated in bold above the locus), leading to amplification of the fragments (dotted arrows and circlednumbers from 1 to 12, which are indicated under the locus for all other manipulations). RR, response regulator; HK, histidine kinase.(B) Schematic representation of the domains identified by SMART prediction in the sensor PprA and in the response regulator PprB.Lengths of peptide sequences are indicated on the left (aa, amino acids). REC, receiver domain; HTH, helix-turn-helix domain. (C) Thepolycistronic transcription of tad genes within the locus was studied using primers (indicated below the circled numbers) designed to amplifyregions (numbered 1 to 12) spanning gene junctions with genomic DNA (upper panel), RNA (middle panel), or cDNA (lower panel)obtained by reverse transcription of extracted mRNA. The molecular masses of the marker bands (in bp) in each gel are indicated on theleft.
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1965

and the rcp-tad locus, containing genes encoding the assemblymachine, including the secretin RcpA, the ATPase TadA, andthe two PilC homologues, TadB and TadC (Fig. 1A). Wepreviously identified and characterized the product of the fppAgene: the prepilin peptidase of the Flp assembly machine (4).The PA4294 open reading frame (ORF), for which no putativehomologue has been identified, is adjacent to the fppA genebut is transcribed in the opposite direction. However, carefulexamination revealed that this ORF had another putative startcodon, 42 nucleotides downstream from that identified at http://www.pseudomonas.com. The ORF transcribed from this startcodon encodes a putative TadF protein homologue, a 154-residue pseudopilin containing the conserved GAVXIEF se-quence, the putative cleavage site being located after the Gresidue (30). The fppA and tadF genes are flanked by thePA4293 and PA4296 genes encoding the histidine kinase PprAand its cognate response regulator PprB, respectively (Fig.1A). This TCS regulates membrane permeability and confersaminoglycoside sensitivity (40). The classical sensor PprA is a922-amino-acid protein with no predicted transmembrane do-main. However, this protein does have one GAF domain (adomain present in phytochromes and cGMP-specific phos-phodiesterases, in Anabaena adenylate cyclases, and in E. coliFhlA protein), two PAS domains (a domain present in theperiod circadian protein [Per], the Ah receptor nuclear trans-locator protein [Arnt], and the single-minded protein [Sim])separated by three PAC (PAS-associated C-terminal) do-mains, and one transmitter domain (H) (Fig. 1B) (35). Theresponse regulator PprB, a member of the LuxR family (6), isa 275-amino-acid protein with a receiver domain and a DNA-binding, helix-turn-helix domain (Fig. 1B).
The tad locus is organized into five transcriptional units. Insilico analysis carried out with Operon Finding 2.1 software athttp://www.pseudomonas.com predicted cotranscription of thercpC to tadG genes. We thus investigated whether the rcpC-tadG, fppA-pprB, and tadF-pprA loci were transcribed as poly-cistronic mRNA, by using oligonucleotides designed to amplifyregions spanning gene junctions (numbered 1 to 12; Fig. 1) oncDNA obtained by the reverse transcription of extractedmRNA (Fig. 1C, lower panel). Control experiments were car-ried out on genomic DNA (Fig. 1C, upper panel) and RNA(Fig. 1C, middle panel). Cotranscribed genes give a signal ofthe same size in the cDNA and DNA panels. As shown in Fig.1C, this was the case for rcpC-rcpA (lane 1), rcpA-tadZ (lane 2),tadZ-tadA (lane 3), tadA-tadB (lane 4), tadB-tadC (lane 5),tadC-tadD (lane 6), tadD-PA4298 (lane 7), PA4298-tadG (lane8), and tadF-pprA (lane 12). A positive internal control withinthe tadG gene (lane 9) and a negative control between the twodivergent genes tadG and pprB (lane 10) were included. Ourresults demonstrate the transcription of the rcpC-tadG locus asa polycistronic mRNA, constituting transcriptional unit II. ThetadF and pprA genes were also cotranscribed, constituting tran-scriptional unit V, with transcriptional unit I consisting of theflp gene. In contrast, two independently transcribed genes gen-erated a signal in the DNA panel but not in the cDNA panel.This was the case for the fppA and pprB genes (lane 11), whichthus form independent transcriptional units, referred to as IVand III, respectively. The lack of an amplification signal (mid-dle panel) in the absence of reverse transcriptase confirmedthat RNA samples were not contaminated by genomic DNA.
Overall, these results demonstrate that the P. aeruginosa tadlocus is organized into five transcriptional units (I to V;Fig. 1A).
The Flp pilin is produced late in the stationary growthphase. In a previous study, we found that Flp was producedlate in the growth phase, but this was not strictly reproduciblebetween cultures (4). We inserted the putative promoter re-gion upstream from the flp gene into the miniCTX-lacZ vector,generating miniCTX-pflp-lacZ. This construct was introducedinto the chromosome of the PAO1 strain, to generate thePAO1/flp-lacZ strain. Expression peaked when the bacteriahad been cultured at 37°C in L broth with vigorous shaking,and the bacteria were then subcultured under aerobic condi-tions, with vigorous shaking, at 30°C. Under these conditions,expression of the flp-lacZ fusion was maximal in the late sta-tionary growth phase (Fig. 2A). Indeed, immunodetection ofthe Flp protein with specific anti-Flp antibodies showed thatFlp was not produced during the exponential growth phase,was barely detectable at early stationary phase, and was pro-duced in significant amounts during the late stationary growthphase (Fig. 2B). We then investigated whether Flp subunitsproduced under these conditions assembled into type IVb pili.Using anti-Flp antibodies, we showed by immunogold labelingand TEM that Flp pili were formed at the cell surface (Fig.2C). The appearance and number (one per cell) of the typeIVb pili assembled when Flp was produced from a gene in-serted into the chromosome (Fig. 2C) were identical to thoseobserved when Flp was overproduced from a replicative plas-mid (4; also data not shown). Thus, the Flp pilin was producedunder aerobic conditions, late in the growth phase, and wasassembled into a single type IVb pilus.
The PprB response regulator controls tad gene expression.Based on the particular location of the pprA and pprB geneswithin the tad locus and on the cotranscription of the pprA andtadF genes, we further investigated the effect of the PprA-PprBTCS on expression of the transcriptional units correspondingto genes encoding the Tad machine (I, II, IV, and V). Weconstructed four different reporter PAO1 strains, correspond-ing to the various transcriptional units defined above and des-ignated as flp-lacZ (I), rcpC-lacZ (II), fppA-lacZ (IV), andtadF-lacZ (V). We monitored relative expression levels in thepresence and absence of inducible pprB overexpression. Over-production of the PprB regulator led to early expression of theflp-lacZ fusion (I) in the growth phase (Fig. 3A). The overpro-duction of PprB increased the level of expression of the rcpC-lacZ fusion (II) to levels up to 20 times higher than those in thecontrol strain (Fig. 3B). Moreover, PprB seems to be the mainregulator controlling flp expression, as expression of the flp-lacZ fusion was very weak throughout the growth phase in apprB mutant and was restored to parental levels when the pprBgene was introduced in trans (Fig. 3C). Similarly, expression ofthe rcpC-lacZ fusion was very weak in the pprB deletion mu-tant, reaching no more than 50 Miller units, corresponding tothe basal level of expression for the empty vector (miniCTX-lacZ integrated into the chromosome at attB sites but withouta promoter region; data not shown), but was restored to pa-rental levels by introducing the pprB gene in trans (Fig. 3C).The rcpC-lacZ fusion had a level of activity in the wild-type(WT) PAO1 background only 1/50 that of the flp-lacZ fusion(compare the scales in Fig. 3). However, this level of expres-
1966 BERNARD ET AL. J. BACTERIOL.

sion was sufficient for the assembly of a single pilus at the cellsurface. Expression of the fppA gene was also essentially de-pendent on the PprB response regulator, as the level of ex-pression of the fppA-lacZ fusion (IV) in the pprB mutant wasonly one-sixth that in the WT strain (Fig. 3C). The expressionof the fppA-lacZ fusion was only partially restored by introduc-ing the pprB gene in trans. The tadF-lacZ fusion (V) was muchless strongly expressed in the WT strain than the other lacZ
fusions and was therefore slightly affected in the pprB mutant(Fig. 3C). However, the expression of this fusion was inducedby the overproduction of PprB in the pprB mutant. Thus, thePprB response regulator of the PprA-PprB TCS positively con-trols the expression of these four transcriptional units of thetad locus, although the PprB-dependent activation of transcrip-tional unit V was detectable only under conditions of pprBoverexpression. To validate this conclusion, the PprB-depen-
FIG. 2. (A) Expression of the chromosomal flp-lacZ fusion, monitored in the PAO1 strain. Data are expressed in Miller units and correspondto the mean values (with error bars) obtained from three independent experiments. The corresponding growth curve (dotted curve) for the straincarrying the flp-lacZ chromosomal fusion is presented. (B) Detection of Flp pilin production in whole-cell extracts from the PAO1 strain at varioustime points (2, 4, 7, 12, and 24 h, referred to on the growth curve in panel A as points 1, 2, 3, 4, and 5, respectively) during growth in L broth at30°C, with vigorous shaking. Signal specificity was checked by Flp detection in the PAO1�flp strain collected at points 1 and 5 of growth. Theproteins were separated by electrophoresis in a 16.5% polyacrylamide Tris-glycine gel, transferred onto nitrocellulose, and detected with anantibody against Flp. The numbers on the right indicate the sizes of the molecular mass standards (in kDa). (C) Flp type IVb pilus (arrow) on thesurface of P. aeruginosa cells following Flp production from a chromosomal gene (left panel; magnification, 50,000) and the absence of a labeledstructure at the surface of the PAO1�flp strain (right panel; magnification, 50,000). Size markers correspond to 0.7 �m.
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1967
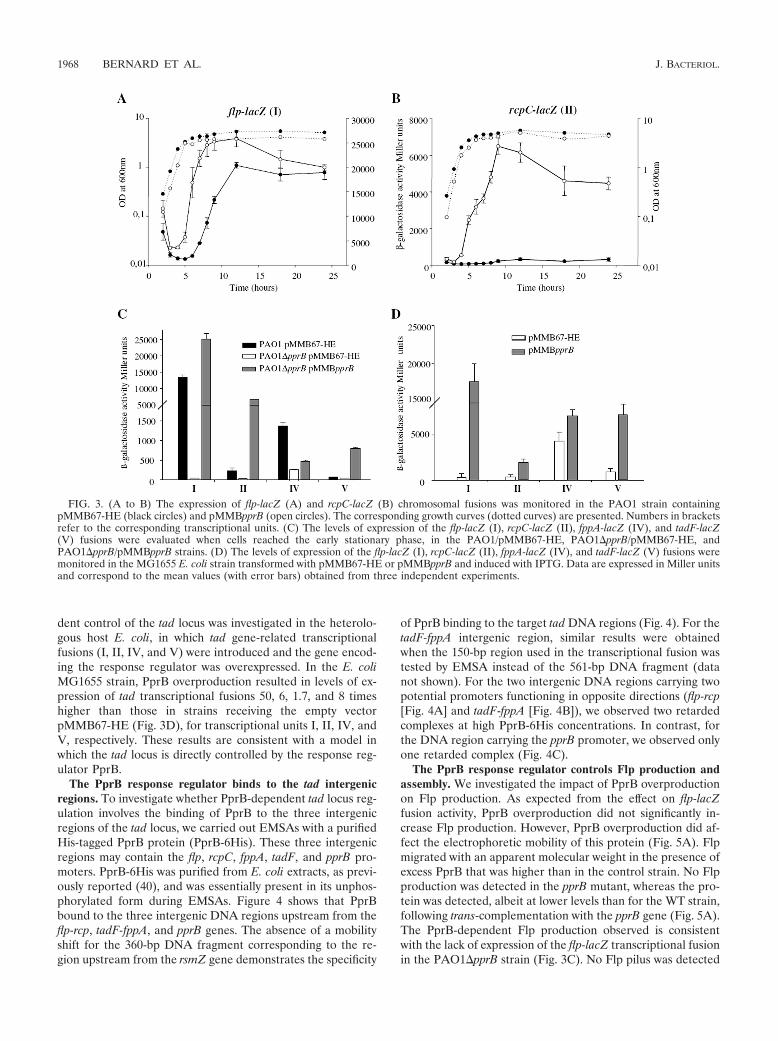
dent control of the tad locus was investigated in the heterolo-gous host E. coli, in which tad gene-related transcriptionalfusions (I, II, IV, and V) were introduced and the gene encod-ing the response regulator was overexpressed. In the E. coliMG1655 strain, PprB overproduction resulted in levels of ex-pression of tad transcriptional fusions 50, 6, 1.7, and 8 timeshigher than those in strains receiving the empty vectorpMMB67-HE (Fig. 3D), for transcriptional units I, II, IV, andV, respectively. These results are consistent with a model inwhich the tad locus is directly controlled by the response reg-ulator PprB.
The PprB response regulator binds to the tad intergenicregions. To investigate whether PprB-dependent tad locus reg-ulation involves the binding of PprB to the three intergenicregions of the tad locus, we carried out EMSAs with a purifiedHis-tagged PprB protein (PprB-6His). These three intergenicregions may contain the flp, rcpC, fppA, tadF, and pprB pro-moters. PprB-6His was purified from E. coli extracts, as previ-ously reported (40), and was essentially present in its unphos-phorylated form during EMSAs. Figure 4 shows that PprBbound to the three intergenic DNA regions upstream from theflp-rcp, tadF-fppA, and pprB genes. The absence of a mobilityshift for the 360-bp DNA fragment corresponding to the re-gion upstream from the rsmZ gene demonstrates the specificity
of PprB binding to the target tad DNA regions (Fig. 4). For thetadF-fppA intergenic region, similar results were obtainedwhen the 150-bp region used in the transcriptional fusion wastested by EMSA instead of the 561-bp DNA fragment (datanot shown). For the two intergenic DNA regions carrying twopotential promoters functioning in opposite directions (flp-rcp[Fig. 4A] and tadF-fppA [Fig. 4B]), we observed two retardedcomplexes at high PprB-6His concentrations. In contrast, forthe DNA region carrying the pprB promoter, we observed onlyone retarded complex (Fig. 4C).
The PprB response regulator controls Flp production andassembly. We investigated the impact of PprB overproductionon Flp production. As expected from the effect on flp-lacZfusion activity, PprB overproduction did not significantly in-crease Flp production. However, PprB overproduction did af-fect the electrophoretic mobility of this protein (Fig. 5A). Flpmigrated with an apparent molecular weight in the presence ofexcess PprB that was higher than in the control strain. No Flpproduction was detected in the pprB mutant, whereas the pro-tein was detected, albeit at lower levels than for the WT strain,following trans-complementation with the pprB gene (Fig. 5A).The PprB-dependent Flp production observed is consistentwith the lack of expression of the flp-lacZ transcriptional fusionin the PAO1�pprB strain (Fig. 3C). No Flp pilus was detected
FIG. 3. (A to B) The expression of flp-lacZ (A) and rcpC-lacZ (B) chromosomal fusions was monitored in the PAO1 strain containingpMMB67-HE (black circles) and pMMBpprB (open circles). The corresponding growth curves (dotted curves) are presented. Numbers in bracketsrefer to the corresponding transcriptional units. (C) The levels of expression of the flp-lacZ (I), rcpC-lacZ (II), fppA-lacZ (IV), and tadF-lacZ(V) fusions were evaluated when cells reached the early stationary phase, in the PAO1/pMMB67-HE, PAO1�pprB/pMMB67-HE, andPAO1�pprB/pMMBpprB strains. (D) The levels of expression of the flp-lacZ (I), rcpC-lacZ (II), fppA-lacZ (IV), and tadF-lacZ (V) fusions weremonitored in the MG1655 E. coli strain transformed with pMMB67-HE or pMMBpprB and induced with IPTG. Data are expressed in Miller unitsand correspond to the mean values (with error bars) obtained from three independent experiments.
1968 BERNARD ET AL. J. BACTERIOL.

in the pprB mutant, but Flp pili were recovered at the cellsurfaces of pprB mutant cells transformed with pMMBpprB(Fig. 5B). However, these pili were significantly shorter (0.56 �0.14 �m) than WT Flp pili (2.25 � 0.25 �m) (P � 0.0001;Fig. 5C).
Electrophoretic migration of the Flp pilin is modified in anRcpC-dependent manner. As this PprB-dependent electro-phoretic mobility shift might potentially result from a defect inthe FppA-dependent processing of the Flp pilin (4), we inves-tigated whether Flp pili assembled at the surface of PprB-overproducing bacteria. The PAO1/pMMBpprB and PAO1/pMMB67-HE strains did have similar numbers and lengths oftype IVb pili at their surfaces, as shown by TEM (Fig. 5B). Asassembly of the Flp pilin subunit into a pilus requires prepilinpeptidase-dependent processing (4), the assembly of Flp pili atthe bacterial cell surface demonstrates that Flp pilins are cor-rectly processed under conditions of PprB overproduction andthat the electrophoretic mobility shift observed is not due to anFppA-dependent change in Flp maturation.
We therefore investigated the possible involvement of thercp-tad genes in the observed Flp electrophoretic mobility shift.As the functions of RcpA, TadA, TadB, TadC, TadF, and
FppA have been clearly defined, we investigated the role of thefirst gene of the rcpC-tadG operon, rcpC, which encodes aputative protein of unknown function. Overproduction of thePprB response regulator in the �rcpC background abolished
FIG. 5. (A) Flp production in the PAO1/pMMB67-HE (lane 1),PAO1/pMMBpprB (lane 2), PAO1�pprB/pMMB67-HE (lane 3), andPAO1�pprB/pMMBpprB (lane 4) strains. Cells at an OD600 of 0.5 wereinduced by incubation with 0.1 mM IPTG. The numbers on the rightindicate the sizes of the molecular mass standards (in kDa). (B) Theassembly of Flp type IVb pili (arrow) at the surface of the bacteria waschecked by TEM coupled with immunogold labeling, using the anti-Flpantibody in the PAO1/pMMB67-HE, PAO1/pMMBpprB, PAO1�pprB/pMMB67-HE, and PAO1�pprB/pMMBpprB strains (magnification, 50,000). The size bar indicates 0.7 �m. (C) Lengths of Flp pili werequantified for 30 images for each strain acquired systematically from atleast three independent experiments in the WT PAO1 and the pprBmutant trans-complemented with the pprB gene. Unpaired t tests wereused for the comparison of results; ***, P � 0.0001.
FIG. 4. EMSAs performed with the purified PprB-6His protein, atconcentrations of 0 to 2.34 �M, and the intergenic DNA regionsidentified in the P. aeruginosa tad locus that were amplified by PCR togenerate DNA fragments of 501, 561, and 560 bp for the regionsupstream from the flp-rcp (A), tadF-fppA (B), and pprB (C) genes. Tworetarded complexes (** and *) were clearly identified at high PprB-6His concentrations for the flp-rcp (A) and tadF-fppA (B) intergenicregions, whereas only one retarded complex was observed for the pprBintergenic region. No shift was observed for the 360-bp DNA fragmentcorresponding to the DNA region upstream from the rsmZ gene.Molecular size markers were run on each gel and are indicated on theright (in bp).
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1969

the Flp electrophoretic mobility shift (Fig. 6A) with respect tothe WT strain (Fig. 6A). This shift was fully restored by intro-ducing the rcpC gene into the chromosome at the attB sites(Fig. 6A). Thus, the RcpC putative protein is involved in themodification of the Flp protein, leading to the observed Flpelectrophoretic mobility shift. Further electron microscopystudies of Flp pilus assembly showed that deletion of the rcpCgene had no effect on Flp assembly at the surface of thebacteria (Fig. 6B). These results suggest that the rcpC gene
product may be involved in a modification that does not affectFlp stability and pilus assembly.
The putative RcpC protein regulates Flp pilus-mediatedattachment to eukaryotic cells. We previously showed that Flpoverproduction led to biofilm formation in a P. aeruginosastrain (PAO1�pilA�fliC) devoid of both type IVa pili andflagella (4). We investigated the role of the putative RcpCprotein in the Flp pilus-dependent biofilm phenotype, by cre-ating an rcpC mutation in the PAO1�pilA�fliC strain. Strainscollected after culture under conditions of optimal Flp produc-tion from the chromosomal gene, and for which pili werealready assembled at the surface, were used for inoculation instatic conditions, as described in Materials and Methods. ThePAO1�pilA�fliC parental strain formed clusters of cells after10 h (Fig. 6C). These clusters began to grow after 6 h (data notshown). At this time point, the parental PAO1 strain hadalready formed a continuous layer with a three-dimensionalstructure. The flp mutant adhered only as sparse, isolated bacte-ria, confirming the strong involvement of Flp pili in bacterium-surface attachment and in the bacterium-bacterium contact andaggregation observed in a previous study (4). The rcpC mutantdisplayed bacterial cell clustering similar to that of the parentalstrain (Fig. 6C), suggesting that RcpC-dependent Flp modifi-cation had no significant effect under these conditions.
We therefore investigated whether the rcpC gene productcontrolled the Flp-dependent adhesion to respiratory epithe-lial cells observed in a previous study (4). In the absence of Flppili, P. aeruginosa cells adhered only weakly to eukaryotic cells.The rcpC mutant displayed a significantly (P � 0.001) lower (bya factor of 3) level of adhesion to eukaryotic cells than theparental isogenic strain (Fig. 7). The parental phenotype wasfully restored by introducing the rcpC gene at the attB sites(Fig. 7B; P � 0.0001). This finding suggests that the putativeRcpC protein influences Flp type IVb pilus-host receptor in-teractions.
The putative TadF pseudopilin is not required for Flp pro-duction, type IVb pilus assembly, and adhesion. The TadFpseudopilin is essential for Flp pilus assembly in A. actinomy-cetemcomitans. This gene is weakly expressed in P. aeruginosa(Fig. 3C). The effect of tadF mutation was tested for Flpproduction, type IVb pilus assembly, and adhesion (see Fig. S1in the supplemental material). The tadF mutant produced Flpprotein in similar amounts to the PAO1 parental strain, andthe assembly of Flp type IVb pili was also similar in the tadFand the WT strains. The introduction of a tadF mutation intothe PAO1�pilA�fliC strain did not affect biofilm formation onglass slides or Flp-dependent adhesion to respiratory epithelialcells. These results demonstrate that, under the conditionsused, the P. aeruginosa TadF putative pseudopilin is dispens-able for Flp pilus biogenesis and Flp-dependent adhesion phe-notypes.
DISCUSSION
P. aeruginosa is the only bacterium known to be able toassemble both type IVa and type IVb pili. Type IVb pili of theFlp subfamily consist of the major pilin, Flp, assembledthrough the concerted action of a dedicated machine, the Tadmachine. We optimized the culture conditions allowing repro-ducible production of Flp. This made it possible (i) to dissect
FIG. 6. (A) Flp production in the PAO1/pMMBpprB (lane 1),PAO1�rcpC/pMMBpprB (lane 2), PAO1�rcpC attB::rcpC/pMMBpprB(lane 3), PAO1/pMMB67-HE (lane 4), and PAO1�rcpC/pMMB67-HE(lane 5) strains. Cells at an OD600 of 0.5 were induced by incubation with0.1 mM IPTG. The numbers on the right are the sizes of the molecularmass standards (in kDa). (B) Assembly of Flp type IVb pili (arrow) at thebacterial cell surface in the PAO1/pMMBpprB, PAO1�rcpC/pMMB67-HE, PAO1�rcpC/pMMBpprB, and PAO1�rcpC attB::rcpC/pMMBpprBstrains (magnification, 30,000 and 40,000). The size bars correspondto 1 �m and 0.5 �m, respectively. (C) Biofilm formation at the air-liquidinterface of glass slides immersed in culture medium was analyzed byconfocal laser scanning microscopy observation after DAPI staining.Stacked images and corresponding extracted z images (z slices of 200 nm)and their respective xy and xz planes at 180 magnification were obtainedfor the PAO1, PAO1�pilA�fliC, PAO1�pilA�fliC�flp, and PAO1�pilA�fliC�rcpC strains.
1970 BERNARD ET AL. J. BACTERIOL.

the mechanism regulating the tad locus and (ii) to obtain in-sight into the function of putative products encoded by severalgenes of this locus. In contrast to the results reported here forP. aeruginosa, anaerobic growth conditions favor the produc-tion of Flp pili in A. actinomycetemcomitans (27), whereas highCO2 levels ensure reproducible Flp pilus production in A.aphrophilus (31). The P. aeruginosa PprA sensor of the PprABTCS possesses PAS domains, which are involved in sensingenvironmental signals, such as oxygen concentration, redoxpotential, or light. The environment-dependent modulation oftype IVb pilin production seems to be a common feature,having also been reported in A. aphrophilus, A. actinomycetem-comitans (31), and Caulobacter crescentus (36). In C. crescentus,the histidine kinase PleC must be present at the cell pole forspatial PilA (the pilin subunit) accumulation and assembly(37). In C. crescentus, the global cell cycle regulator, CtrA, andthe GcrA master regulator control the concerted expression ofdifferent pilus genes over time during the bacterial cell cycle(13, 29), after the initiation of replication and flagellum bio-genesis (21).
In silico analyses showed that the genetic organization of thetad locus of P. aeruginosa is more complex than that of othertad loci (19), suggesting major genomic rearrangements orgene shuffling. The P. aeruginosa tad locus is also simpler thanmost of the tad loci identified to date. It lacks the second flpgene, the rcpB gene, and the second tadE pseudopilin gene.However, functional Flp pili are assembled in the absence of
these genes in P. aeruginosa (no orthologues were identified inthe P. aeruginosa genome, even outside the tad locus). Unlikeother tad loci, which are transcribed as a polycistronic mRNA(10), we have shown that the P. aeruginosa tad locus is tran-scribed as five independent transcriptional units (I, flp; II,rcpC-tadG; III, pprB; IV, fppA; and V, tadF-pprA). The pres-ence of the pprAB genes, encoding a classical TCS, within thelocus suggests that this system is involved in regulating the tadcluster. Indeed, our results demonstrate that transcriptionalunits I, II, and IV are regulated by the PprB response regulatorand that PprB binds to the putative promoters of the fivetranscriptional units of the tad locus. The tadF-pprA operon(V) is weakly expressed, but its expression is stimulated byPprB overproduction. As tadF is cotranscribed with pprA, thepositive effect of the PprB response regulator on the geneencoding its cognate sensor, PprA, creates a positive loop thatmay contribute to the sequential and hierarchical regulation ofa subset of genes, depending on the affinity of the responseregulator for different DNA-binding regions, as shown for theBvgS-BvgA TCS of Bordetella pertussis (5). The regulation ofthe transcriptional units IV and V seems to be more compli-cated than that of transcriptional units I and II. The presenceof regulatory elements (two promoters, each with at least onePprB-binding site) on a short DNA region between transcrip-tional units IV and V suggested that the PprB-dependent tran-scription of one gene might influence the transcription of theother. In the absence of PprB, both these genes are turned off.At a low (physiological) PprB concentration, the fppA gene wasexpressed, whereas the tadF gene was not. At a high PprBconcentration (pprB overexpression), the activity of the fppApromoter was repressed, whereas the tadF promoter wasswitched on. This hypothesis requires the existence of twoindependent PprB-binding sites. Alternatively, given the in-complete complementation of the pprB mutant for fppA-lacZexpression, it should also be borne in mind that the pprBdeletion generated may have had an unexpected polar effect onthe neighboring genes, including fppA in particular, despite theremoval of the full nucleotide sequence of the gene, with theretention of only the start and stop codons, in our deletionstrategy. The partial complementation of the pprB mutant forthe phenotypes tested suggests that additional regulatory ele-ments are lacking. Lastly, the binding of PprB to its own pro-moter region is consistent with a model involving autoregula-tion.
The PprA-PprB TCS was first identified as controlling cellpermeability and aminoglycoside sensitivity in the P. aerugi-nosa PAK strain (40). In this previous study, the pprA mutantgrew more slowly, had an altered membrane protein profile,and produced more OprF porin than the WT strain. However,our pprB deletion mutant grew as well as the WT strain (datanot shown), suggesting that the growth defect of the PAK pprAmutant may be linked to a regulatory network involving PprAbut not PprB, despite the demonstration of a functional linkbetween PprA and PprB (40). The overproduction of full-length PprA did not affect flp expression, production, or as-sembly (data not shown). This suggests that either PprB-de-pendent activation of the tad locus is independent of PprA orthe overproduced full-length PprA is not active. Alternatively,genetic organization and regulation may differ between thePAO1 and PAK strains. Differences in genetic organization
FIG. 7. (A) Representative views of adhesion to bronchial epithelialcells incubated with the PAO1�pilA�fliC, PAO1�pilA�fliC�flp,PAO1�pilA�fliC�rcpC, and PAO1�pilA�fliC�rcpC attB::rcpC strains.(B) The numbers of bacteria adhering to bronchial epithelial cells wereevaluated in three independent assays. Adherent bacteria were quantifiedfor the various genetic backgrounds (PAO1�pilA�fliC, PAO1�pilA�fliC�flp, PAO1�pilA�fliC�rcpC, and PAO1�pilA�fliC�rcpC attB::rcpCstrains), on epithelial cells (30 epithelial cells/assay), in randomly chosenmicroscopic fields. Comparisons were made by one-way analysis of vari-ance and unpaired t tests. ***, P � 0.0001; **, P � 0.001.
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1971

were ruled out by our observation that the organization of thetad locus was identical in PAK and PAO1 (data not shown).Transcriptomic analyses of a PAO1 pprB mutant showed thatthis gene positively regulated several operons, with levels ofvirulence factor secretion and of swimming or swarming mo-tility being much lower in the mutant than in the WT strain (6).This result was recently attributed to a second mutation in theLasR regulator in the pprB mutant used, rather than to PprBitself (6). In our hands, in-frame deletion of the pprB gene hadno effect on proteolytic activity, the secretion of extracellularcompounds, twitching, or swarming or swimming motility (seeFig. S2 in the supplemental material). Our fusions were notaffected in a pprA mutant (data not shown), indicating that thefunctional relationships between PprA and PprB in controlover the tad locus may not be simple.
We also investigated the involvement of several other genesfrom this genetic locus in type IVb pilus assembly or function.A gene encoding a pilin-like protein similar to the TadF pseu-dopilin of A. actinomycetemcomitans was identified in the P.aeruginosa tad locus. In contrast, the tad locus of A. actinomy-cetemcomitans contains two genes, tadE and tadF, encodingputative pseudopilins. These two proteins are processed in aTadV-dependent manner (30). In contrast to our results, atadF mutant of A. actinomycetemcomitans was shown to pro-duce less Flp pilin (30), to be unable to assemble Flp pili (17,41), and not to adhere to inert surfaces (17). In our laboratoryconditions, the putative TadF protein had no effect on P.aeruginosa Flp biology. However, we cannot rule out the possi-bility of another gene in the P. aeruginosa genome complementingTadF function or that the P. aeruginosa tadF gene product playssome role, as suggested by observations that the overproductionof PprB in a pprB mutant leads to short Flp pili and induces thetadF-lacZ fusion. The putative TadF protein may control Flppilus length in P. aeruginosa, as previously reported for the pseu-dopilin XcpX controlling the length of the pseudopilus assembledby the type II secretion machinery (7).
We then targeted the rcpC gene, because (i) this gene ispresent only in type IVb loci dedicated to Flp pilus assemblyand (ii) this gene has no known function (31). The rcpC geneproduct of P. aeruginosa has a signal peptide (predicted bySignalP 3.0 software; data not shown), suggesting that it maybe located in the outer membrane or periplasm. The putativeRcpC protein has been localized to the A. actinomycetemcomi-tans outer membrane, although it is also detected in the innermembrane (2). The additional protein of the machine, TadD,a putative lipoprotein (signal peptidase [II] cleavage site be-tween amino acids 16 and 17, as predicted by LipoP software;data not shown), is probably located in the outer membrane.The putative RcpC protein, together with the putative lipopro-tein TadD and the secretin RcpA, may therefore constitute anouter membrane complex unique to the Flp type IVb pilusmachine in P. aeruginosa. Flp protein stability and Flp pilusassembly were not altered in the rcpC mutant. However, weobserved an RcpC-dependent electrophoretic mobility shift ofthe P. aeruginosa Flp pilin upon PprB overproduction. Weinterpret this result as indicating that only a small proportionof the thousands of copies of Flp subunits assembled in a WTbackground is modified in an RcpC-dependent manner andthat this is not sufficient for detection of the change in elec-trophoretic mobility. Upon PprB overproduction, levels of the
putative RcpC protein—but not of Flp—increased, and themodified Flp protein was thus the major form observed. ThisRcpC-dependent electrophoretic mobility shift of Flp pilin haspreviously been reported in A. actinomycetemcomitans (23). InP. aeruginosa, the rcpC mutation had no effect on biofilm for-mation but decreased adhesion to epithelial cells. These find-ings suggest that the RcpC putative protein may be multifunc-tional, with independent roles in Flp modification andadhesion. Furthermore, we cannot rule out the possible role ofanother factor from epithelial cells during contact, renderingthe Flp pili of rpcC bacteria more sensitive to shearing. Alter-natively, the putative RcpC protein may influence Flp pilinquality by possible posttranslational modification of the pilinsubunit before its assembly. It has been suggested that thisphenomenon is due to incomplete or partial Flp pilin glycosy-lation in an A. actinomycetemcomitans rcpC mutant (30). TheRcpC-like proteins FlgA and CpaB belong to a family of bac-terial proteins containing two �-clip domains, probably locatedin the periplasm, which are thought to bind sugar moieties(14). Interactions between the RcpC �-clip domains and mod-ified Flp pilin to facilitate extrusion through the secretin ring(31) are not consistent with the assembly of the Flp pilus in anrcpC mutant, at least in P. aeruginosa. In this attractive hypoth-esis, the “unmodified” Flp pilus in the rcpC mutant fails to bindto eukaryotic receptors, whereas the modified version of thisprotein in the WT promotes adhesion to such receptors. How-ever, it remains unknown whether the adhesion phenotype ofrcpC cells is linked to Flp pilin modification.
Our results demonstrate that P. aeruginosa is able to producetype IVb Flp pili with a simplified version of the Tad machine,the expression of which depends on the PprB response regu-lator of the PprAB TCS.
ACKNOWLEDGMENTS
We thank A. Bernadac and D. Byrne for technical assistance withthe microscopy platform and the protein production platform of theInstitut de Microbiologie de la Mediterrannee (Marseille, France) andM. Ansaldi’s team (G. Panis and L. Jacquet) from UPR9043 (Mar-seille, France) for their advice on RT-PCR experiments. We thankShougouang Sin (University of Florida, Gainesville, FL), who kindlyprovided the E. coli strain containing the plasmid with the His-taggedversion of the PprB response regulator. We are particularly grateful toEric Cascales (UPR9027) for his invaluable scientific insight.
The work of S.D.B. and A.F. is supported by the French CysticFibrosis Foundation (VLM), the Bettencourt-Schueller Foundation,and CNRS institutional grants. C.S.B. is supported by the FrenchCystic Fibrosis Foundation (VLM). A.F. is supported by the BritishRoyal Society.
REFERENCES
1. Becher, A., and H. P. Schweizer. 2000. Integration-proficient Pseudomonasaeruginosa vectors for isolation of single-copy chromosomal lacZ and luxgene fusions. BioTechniques 29:948–950, 952.
2. Clock, S. A., P. J. Planet, B. A. Perez, and D. H. Figurski. 2008. Outermembrane components of the Tad (tight adherence) secreton of Aggregati-bacter actinomycetemcomitans J. Bacteriol. 190:980–990.
3. Craig, L., M. E. Pique, and J. A. Tainer. 2004. Type IV pilus structure andbacterial pathogenicity. Nat. Rev. Microbiol. 2:363–378.
4. de Bentzmann, S., M. Aurouze, G. Ball, and A. Filloux. 2006. FppA, a novelPseudomonas aeruginosa prepilin peptidase involved in assembly of type IVbpili. J. Bacteriol. 188:4851–4860.
5. Deora, R., H. J. Bootsma, J. F. Miller, and P. A. Cotter. 2001. Diversity in theBordetella virulence regulon: transcriptional control of a Bvg-intermediatephase gene. Mol. Microbiol. 40:669–683.
6. Dong, Y. H., X. F. Zhang, H. M. Soo, E. P. Greenberg, and L. H. Zhang. 2005.The two-component response regulator PprB modulates quorum-sensing
1972 BERNARD ET AL. J. BACTERIOL.

signal production and global gene expression in Pseudomonas aeruginosa.Mol. Microbiol. 56:1287–1301. (Retraction, 69:780, 2008.)
7. Durand, E., G. Michel, R. Voulhoux, J. Kurner, A. Bernadac, and A. Filloux.2005. XcpX controls biogenesis of the Pseudomonas aeruginosa XcpT-contain-ing pseudopilus. J. Biol. Chem. 280:31378–31389.
8. Filloux, A., S. Bleves, P. van Ulsen, and J. Tommassen. 2004. Protein secretionmechanisms in Pseudomonas, p. 749–791. In J.-L. Ramos (ed.), Pseudomonas:genomics, life style and molecular architecture, vol. 1. Kluwer Academic/PlenumPublishers, New York, NY.
9. Friedman, L., and R. Kolter. 2004. Two genetic loci produce distinct carbo-hydrate-rich structural components of the Pseudomonas aeruginosa biofilmmatrix. J. Bacteriol. 186:4457–4465.
10. Haase, E. M., J. O. Stream, and F. A. Scannapieco. 2003. Transcriptionalanalysis of the 5� terminus of the flp fimbrial gene cluster from Actinobacillusactinomycetemcomitans. Microbiology 149:205–215.
11. Hoang, T. T., R. R. Karkhoff-Schweizer, A. J. Kutchma, and H. P. Schweizer.1998. A broad-host-range Flp-FRT recombination system for site-specificexcision of chromosomally located DNA sequences: application for isolationof unmarked Pseudomonas aeruginosa mutants. Gene 212:77–86.
12. Hoang, T. T., R. R. Karkhoff-Schweizer, A. J. Kutchma, and H. P. Schweizer.2000. Integration proficient plasmids for Pseudomonas aeruginosa: site-spe-cific integration and use for engineering of reporter and expression strains.Plasmid 43:59–72.
13. Holtzendorff, J., D. Hung, P. Brende, A. Reisenauer, P. H. Viollier, H. H.McAdams, and L. Shapiro. 2004. Oscillating global regulators control thegenetic circuit driving a bacterial cell cycle. Science 304:983–987.
14. Iyer, L. M., and L. Aravind. 2004. The emergence of catalytic and structuraldiversity within the beta-clip fold. Proteins 55:977–991.
15. Jackson, K. D., M. Starkey, S. Kremer, M. R. Parsek, and D. J. Wozniak.2004. Identification of psl, a locus encoding a potential exopolysaccharidethat is essential for Pseudomonas aeruginosa PAO1 biofilm formation. J.Bacteriol. 186:4466–4475.
16. Juhas, M., L. Wiehlmann, P. Salunkhe, J. Lauber, J. Buer, and B. Tummler.2005. GeneChip expression analysis of the VqsR regulon of Pseudomonasaeruginosa TB. FEMS Microbiol. Lett. 242:287–295.
17. Kachlany, S. C., P. J. Planet, M. K. Bhattacharjee, E. Kollia, R. Desalle,D. H. Fine, and D. H. Figurski. 2000. Nonspecific adherence by Actinoba-cillus actinomycetemcomitans requires genes widespread in bacteria andarchaea. J. Bacteriol. 182:6169–6176.
18. Kachlany, S. C., P. J. Planet, R. Desalle, D. H. Fine, and D. H. Figurski.2001. Genes for tight adherence of Actinobacillus actinomycetemcomitans:from plaque to plague to pond scum. Trends Microbiol. 9:429–437.
19. Kachlany, S. C., P. J. Planet, R. Desalle, D. H. Fine, D. H. Figurski, and J. B.Kaplan. 2001. flp-1, the first representative of a new pilin gene subfamily, isrequired for non-specific adherence of Actinobacillus actinomycetemcomi-tans. Mol. Microbiol. 40:542–554.
20. Kaniga, K., and J. Davison. 1991. Transposon vectors for stable chromo-somal integration of cloned genes in rhizosphere bacteria. Gene 100:201–205.
21. Laub, M. T., H. H. McAdams, T. Feldblyum, C. M. Fraser, and L. Shapiro.2000. Global analysis of the genetic network controlling a bacterial cell cycle.Science 290:2144–2148.
22. Matsukawa, M., and E. P. Greenberg. 2004. Putative exopolysaccharidesynthesis genes influence Pseudomonas aeruginosa biofilm development. J.Bacteriol. 186:4449–4456.
23. Perez, B. A., P. J. Planet, S. C. Kachlany, M. Tomich, D. H. Fine, and D. H.Figurski. 2006. Genetic analysis of the requirement for flp-2, tadV, and rcpBin Actinobacillus actinomycetemcomitans biofilm formation. J. Bacteriol. 188:6361–6375.
24. Planet, P. J., S. C. Kachlany, R. Desalle, and D. H. Figurski. 2001. Phylogenyof genes for secretion NTPases: identification of the widespread tadA sub-family and development of a diagnostic key for gene classification. Proc.Natl. Acad. Sci. USA 98:2503–2508.
25. Planet, P. J., S. C. Kachlany, D. H. Fine, R. Desalle, and D. H. Figurski.2003. The widespread colonization island of Actinobacillus actinomycetem-comitans. Nat. Genet. 34:193–198.
26. Ruer, S., S. Stender, A. Filloux, and S. de Bentzmann. 2007. Assembly offimbrial structures in Pseudomonas aeruginosa: functionality and specificityof chaperone-usher machineries. J. Bacteriol. 189:3547–3555.
27. Scannapieco, F. A., S. J. Millar, H. S. Reynolds, J. J. Zambon, and M. J.Levine. 1987. Effect of anaerobiosis on the surface ultrastructure and surfaceproteins of Actinobacillus actinomycetemcomitans (Haemophilus actinomyce-temcomitans). Infect. Immun. 55:2320–2323.
28. Schuster, M., C. P. Lostroh, T. Ogi, and E. P. Greenberg. 2003. Identifica-tion, timing, and signal specificity of Pseudomonas aeruginosa quorum-con-trolled genes: a transcriptome analysis. J. Bacteriol. 185:2066–2079.
29. Skerker, J. M., and L. Shapiro. 2000. Identification and cell cycle control ofa novel pilus system in Caulobacter crescentus. EMBO J. 19:3223–3234.
30. Tomich, M., D. H. Fine, and D. H. Figurski. 2006. The TadV protein ofActinobacillus actinomycetemcomitans is a novel aspartic acid prepilin pep-tidase required for maturation of the Flp1 pilin and TadE and TadF pseu-dopilins. J. Bacteriol. 188:6899–6914.
31. Tomich, M., P. J. Planet, and D. H. Figurski. 2007. The tad locus: postcardsfrom the widespread colonization island. Nat. Rev. Microbiol. 5:363–375.
32. Vallet, I., J. W. Olson, S. Lory, A. Lazdunski, and A. Filloux. 2001. Thechaperone/usher pathway of Pseudomonas aeruginosa: identification of fim-brial gene clusters (cup) and their involvement in biofilm formation. Proc.Natl. Acad. Sci. USA 98:6911–6916.
33. Vallet, I., S. P. Diggle, R. E. Stacey, M. Camara, I. Ventre, S. Lory, A.Lazdunski, P. Williams, and A. Filloux. 2004. Biofilm formation in Pseudo-monas aeruginosa: fimbrial cup gene clusters are controlled by the transcrip-tional regulator MvaT. J. Bacteriol. 186:2880–2890.
34. Vasseur, P., I. Vallet-Gely, C. Soscia, S. Genin, and A. Filloux. 2005. The pelgenes of the Pseudomonas aeruginosa PAK strain are involved at early andlate stages of biofilm formation. Microbiology 151:985–997.
35. Ventre, I., A. Filloux, and A. Lazdunski. 2004. Two-component signal trans-duction systems: a key to the adaptative potential of Pseudomonas aerugi-nosa, p. 257–288. In J.-L. Ramos (ed.), Pseudomonas: virulence and generegulation, vol. 2. Kluwer Academic/Plenum Publishers, New York, NY.
36. Viollier, P. H., N. Sternheim, and L. Shapiro. 2002. A dynamically localizedhistidine kinase controls the asymmetric distribution of polar pili proteins.EMBO J. 21:4420–4428.
37. Viollier, P. H., N. Sternheim, and L. Shapiro. 2002. Identification of alocalization factor for the polar positioning of bacterial structural and reg-ulatory proteins. Proc. Natl. Acad. Sci. USA 99:13831–13836.
38. Wagner, V. E., D. Bushnell, L. Passador, A. I. Brooks, and B. H. Iglewski.2003. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regu-lons: effects of growth phase and environment. J. Bacteriol. 185:2080–2095.
39. Wagner, V. E., R. J. Gillis, and B. H. Iglewski. 2004. Transcriptome analysisof quorum-sensing regulation and virulence factor expression in Pseudomo-nas aeruginosa. Vaccine 22:S15–S20.
40. Wang, Y., U. Ha, L. Zeng, and S. Jin. 2003. Regulation of membranepermeability by a two-component regulatory system in Pseudomonas aerugi-nosa. Antimicrob. Agents Chemother. 47:95–101.
41. Wang, Y., and C. Chen. 2005. Mutation analysis of the flp operon in Actino-bacillus actinomycetemcomitans. Gene 351:61–71.
VOL. 191, 2009 P. AERUGINOSA Tad MACHINE 1973


















