
Green Toehold Switches de-novo-Designed Regulators of Gene Expression. Cell 2014

On the biophysics and kinetics of toehold-mediatedDNA strand displacementNiranjan Srinivas1 Thomas E Ouldridge2 Petr Sulc2 Joseph M Schaeffer3
Bernard Yurke4 Ard A Louis2 Jonathan P K Doye5 and Erik Winfree136
1Computation and Neural Systems California Institute of Technology Pasadena CA 91125 USA 2RudolphPeierls Centre for Theoretical Physics Department of Physics University of Oxford Oxford OX1 3NP UK3Computer Science California Institute of Technology Pasadena CA 91125 USA 4Departments of Electricaland Computer Engineering Materials Science and Engineering Boise State University ID83725 USA 5Physicaland Theoretical Chemistry Laboratory Department of Chemistry University of Oxford Oxford OX1 3QZ UKand 6Bioengineering California Institute of Technology Pasadena CA 91125 USA
Received March 29 2013 Revised July 18 2013 Accepted August 14 2013
ABSTRACT
Dynamic DNA nanotechnology often uses toehold-mediated strand displacement for controllingreaction kinetics Although the dependence ofstrand displacement kinetics on toehold length hasbeen experimentally characterized and phenomeno-logically modeled detailed biophysical understand-ing has remained elusive Here we study stranddisplacement at multiple levels of detail using anintuitive model of a random walk on a 1D energylandscape a secondary structure kinetics modelwith single base-pair steps and a coarse-grainedmolecular model that incorporates 3D geometricand steric effects Further we experimentally inves-tigate the thermodynamics of three-way branch mi-gration Two factors explain the dependence ofstrand displacement kinetics on toehold length(i) the physical process by which a single step ofbranch migration occurs is significantly slowerthan the fraying of a single base pair and (ii) initiatingbranch migration incurs a thermodynamic penaltynot captured by state-of-the-art nearest neighbormodels of DNA due to the additional overhangit engenders at the junction Our findings are con-sistent with previously measured or inferred ratesfor hybridization fraying and branch migrationand they provide a biophysical explanation ofstrand displacement kinetics Our work pavesthe way for accurate modeling of strand displace-ment cascades which would facilitate the
simulation and construction of more complex mo-lecular systems
INTRODUCTION
Recent advances in DNA nanotechnology have enabledthe construction of 2D and 3D nanoscale structures (1ndash7)Nucleic acids have predictable double-helical structureand generally well-understood thermodynamic (8ndash11)and mechanical (12) properties which makes them excel-lent engineering materials In addition to static structuresdynamic nanoscale devices such as circuits (13ndash16) cata-lysts (1718) autonomous molecular motors (19ndash22) andreconfigurable nanostructures (1823ndash25) have been engin-eered using DNA Inspired by experimental advances the-oretical schemes have been proposed (2627) to engineerarbitrarily complex chemical dynamics using DNA If suc-cessful such efforts could enable dynamic DNA circuitsto actively control nanoscale devicesUnfortunately the biophysical understanding of key
kinetic phenomena remains underdeveloped relative toour knowledge of static properties limiting the develop-ment of dynamic DNA nanotechnology Here we studythe biophysical basis of a molecular mechanism calledtoehold-mediated strand displacement which is centralto many dynamic DNA devices built to date Toehold-mediated strand displacement enables control over thekinetics of molecular rearrangement allowing theengineer to program when and where specific steps takeplace in a molecular machineFigure 1 illustrates strand displacement using domain
notation A domain is a set of contiguous nucleotidesdesigned to be either fully bound or fully unbound in
To whom correspondence should be addressed Tel +1 626 395 6246 Fax +1 626 584 0630 Email winfreecaltecheduCorrespondence may also be addressed to Niranjan Srinivas Tel +1 626 395 6994 Email niranjancaltecheduCorrespondence may also be addressed to Thomas E Ouldridge Tel +44 1865 273970 Email touldridge1physicsoxacuk
The authors wish it to be known that in their opinion the first two authors should be regarded as joint First Authors
Nucleic Acids Research 2013 1ndash18doi101093nargkt801
The Author(s) 2013 Published by Oxford University PressThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (httpcreativecommonsorglicensesby-nc30) which permits non-commercial re-use distribution and reproduction in any medium provided the original work is properly cited For commercialre-use please contact journalspermissionsoupcom
Nucleic Acids Research Advance Access published September 9 2013 at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
at C
alifornia Institute of Technology on O
ctober 13 2013httpnaroxfordjournalsorg
Dow
nloaded from
at California Institute of T
echnology on October 13 2013
httpnaroxfordjournalsorgD
ownloaded from
stable configurations Our system initially comprises atwo-stranded complex (S) and a single-stranded lsquoinvaderrsquo(X) S consists of an lsquoincumbentrsquo strand (Y) bound to alsquosubstratersquo strand which has a single-stranded overhangcalled a lsquotoeholdrsquo The invader is fully WatsonndashCrickcomplementary to the substrate and may bind reversiblyto it using the toehold domain (h) This binding is revers-ible because the toehold may lsquofrayrsquo and eventually dissoci-ate (We use the term lsquofrayingrsquo to describe the disruptionof base pairs at the end of a duplex if all base pairs fraythe duplex melts or dissociates Conversely lsquozipperingrsquorefers to when a new base pair forms at the end of anexisting duplex) Once the toehold is bound the lsquoover-hangingrsquo branch migration domain (b) of the invadermay compete with the incumbent for binding with thesubstrate As the incumbent and invader exchange basepairs with the substrate the branch point of the three-stranded complex moves back and forth This lsquothree-waybranch migrationrsquo (henceforth lsquobranch migrationrsquo)process has previously been modeled as an unbiasedrandom walk (28ndash30) as each step causes no net changein base pairing Eventually the incumbent may dissociatecompleting strand displacement Overall displacement isthermodynamically driven forward by the net gain in basepairs due to the toeholdAlthough it is known that bimolecular rate constants
for strand displacement can vary a million-fold dependingon the length of the toehold little is known about theunderlying biophysics or how to generalize the results toother reaction conditions or molecular modifications Forexample what are the mechanisms and intermediate statesinvolved in a single step of branch migration How do thekinetics of strand displacement depend on the length ofthe branch migration domain or on the temperature andbuffer conditions How different is strand displacementin RNA and DNA What is the effect of sequencemismatches at different positions along the branch migra-tion domain To begin answering these questions a fun-damental biophysical understanding is necessary
However the commonly accepted view of stranddisplacement biophysics appears at odds with kinetic meas-urements (3031) Consider strand displacement with a1-base toehold and a 20-base branch migration domainLet us assume that the rate constant for the formationof the toehold base pair is on the order of 106Ms [Thisvalue for 1-nt association is within a factor of two ofexperimental values for 6- 10- 15- and 20-mer hybridiza-tion rates (3032) after accounting for the linear length-dependence for short oligonucleotides (33)] Once thetoehold has bound there are two possibilities (i) thetoehold base pair could dissociate leading to the dissoci-ation of the invader or (ii) the nearest base pair of thesubstrate-incumbent complex could fray allowing theinvader to compete to replace that base pair andcomplete the first step of branch migration Assuming asmay seem reasonable that the rate at which either base pairfrays is similar process (ii) should be approximately half asfast as process (i) This is because once the substrate-incumbent base pair frays there is a 50 chance of theinvader replacing the frayed base pair and a 50 chanceof returning to the initial step Once the first step of branchmigration is complete subsequent forward and backwardsteps are assumed to occur at the same rate Therefore theprobability of successfully completing the remaining stepsof branch migration before going back to the toehold-only-bound state is 120 from the gamblerrsquos ruin analysis [seeSection 142 of Feller (34)] The lifetime of the three-stranded intermediate should be independent of concentra-tion Therefore at low enough concentrations the overallreaction can be modeled as an instantaneous second-orderprocess as reported by Zhang and Winfree (30) Underthese conditions the overall effective rate constant (keff)is given by a hybridization rate constant for the toeholdmultiplied by a success probability of displacement oncethe toehold is bound Even though the time spent in thethree-stranded intermediate is small keff depends stronglyon the probability of displacement once boundkeff 106 eth1=3THORN eth1=20THORN frac14 16 104Ms for a one-base toehold This is over three orders of magnitudelarger than the 8Ms value measured by Zhang andWinfree (30) This is a large unexplained discrepancydespite the approximate nature of our calculation
We now summarize the experimental evidence for theexponential acceleration in keff with toehold length whichwas first reported by Yurke and Mills (31) Zhang andWinfree (30) further characterized this exponential accel-eration and confirmed that it saturates in the long-toeholdlimit Relevant data from both studies (Figure 2A) suggestthat the exponential acceleration in itself is not an artifactof particular sequences although details may well dependon the experimental system (sequences modifications forread-out etc) or conditions (salt temperature etc) Thekinetics of zero-toehold (lsquoblunt endrsquo) strand displacementwas investigated by Reynaldo et al (35) whose measure-ment of 36Ms at 30C is similar to the 14Ms at 25Creported by Zhang and Winfree (30)
Zhang and Winfree (30) built a phenomenologicalmodel for predicting keff from toehold sequence(Figure 2B) They modeled branch migration using twomacro-states I and J containing the first and second
A B
Figure 1 (A) Domain notation Arrows indicate 30-ends Asteriskindicates WatsonndashCrick complementarity (B) Toehold h mediates thedisplacement of the incumbent (Y) by the invader (X) Dots indicatebranch migration intermediates which are not shown
2 Nucleic Acids Research 2013
half of the isoenergetic branch migration intermediatesrespectively which are connected by a simple transitionwith rate constant kb (see Supplementary Figure S1 andSupplementary Section S1 for more details) Their modelfits the data but it is hard to physically interpret the tran-sition between I and J Branch migration is generallythought of as a random walk through many isoenergeticstates rather than a single reversible transitioncharacterized by a first-order rate constant Without amore nuanced understanding of the process it is unclearwhether the fitted value of kb frac14 10s can be justified onmore fundamental biophysical grounds
In this work we model branch migration at a moredetailed level that explicitly includes intermediatesthereby highlighting important thermodynamic andkinetic features of the process that are not evident fromthe phenomenological approach
First we analyze a 1D (single-pathway) model oftoehold-mediated strand displacement called the intuitiveenergy landscape (IEL) model Systematically exploringthe parameter space of this simple model suggests thatsome combination of two factors could explain the de-pendence of strand displacement rate on toehold length(i) the branch migration process is not isoenergetic andcontains a free energy penalty for intermediate statesand (ii) branch migration is slow relative to fraying ofthe toehold
To verify that these factors are not already implicitfeatures of more detailed models of DNA secondary struc-ture thermodynamics we simulated the experiments ofZhang and Winfree (30) using a secondary structurekinetics (SSK) simulator called Multistrand (36)Multistrand incorporates extensive thermodynamic infor-mationmdashfrom state-of-the-art nearest-neighbor (NN)thermodynamic models of DNA secondary structure(8ndash11)mdashcombined with a minimally parameterized ratemodel Multistrand predicts that keff would increase by afactor of 1038 as toehold length increases from 0 to 15 incontrast to the experimentally observed factor of 1065This quantitative discrepancy confirms that factors(i) and (ii) suggested by the IEL analysis are not alreadyimplicitly incorporated in more detailed models of DNAsecondary structure thermodynamics
We then experimentally investigate possibility (i) sug-gested by the IEL by measuring the relative stability of
complexes that mimic the geometric structure of branchmigration intermediates Our experiments provideevidence in support of a free energy penalty for branchmigration intermediates that is not predicted by the NNmodels of DNAFinally we use a recently proposed coarse-grained
molecular model of DNA (oxDNA) (37ndash39) which incorp-orates more physical detail including geometric and stericeffects In addition to correctly predicting the length-dependence of toehold-mediated strand displacementrates oxDNA independently predicts the thermodynamicpenalty for branch migration intermediates suggesting itarises from local steric effects between single-strandedoverhangs around the branch point Further oxDNApredicts that branch migration is slower than fraying ofthe toehold as it is geometrically more complex and neces-sarily involves more thermodynamically unfavorable steps
MATERIALS METHODS AND RESULTS
Intuitive Energy Landscape model
Through a simple approximate calculation we argued thatthe current view of strand displacement biophysics is atodds with kinetic measurements (3031) We now performa more rigorous intuitive analysis by building a simplesingle-pathway model called the IEL model The IEL issimple enough that its kinetic predictions may be analyt-ically or numerically calculated for a given parameteriza-tion We systematically explore the IELrsquos parameter spaceto obtain biophysical intuition and identify key thermo-dynamic and kinetic features essential for predictions tomatch experimental data
State spaceThe IEL considers an invading strand and a substrate-incumbent complex contained in a virtual box of volumeV in solution which defines a concentration u of onemolecule per volume V The state space of the IEL isillustrated in Figure 3 State A corresponds to theinvader being unattached to the substrate-incumbentduplex State B represents the formation of a first basepair within the toehold For each additional toeholdbase pair that zips up we define a new state with stateC indicating the fully formed toehold From here eachstep of branch migration involves replacing an incum-bent-substrate base pair with an invader-substrate basepair We describe the stepping between these intermediatesusing a simple model in which the system must passthough a single effective transition state of raised freeenergy so each complete branch migration step is shownas a single tooth of the lsquosawtoothrsquo pattern between states Cand D This unknown effective transition state could bepotentially as simple as a frayed substrate-incumbent basepair but could also be more complex The final stageof successful displacement involves the dissociation ofthe incumbent (state E) followed by the formation ofthe final base pair between invader and substrate(state F) Subtleties relating to the zero-toehold case arediscussed in Supplementary Section S2 (see Supple-mentary Figure S2)
A B
Figure 2 (A) Dependence of keff on toehold length measured byYurke and Mills (31) (at 20C 1M Na+) and Zhang and Winfree(30) (at 25C 125mM Mg++) Each curve is from a differenttoehold sequence (B) The phenomenological model of Zhang andWinfree (30) kd is assumed to be large relative to other rate constants
Nucleic Acids Research 2013 3
Energy modelThe IEL models the free energy of the virtual box (Gbox)relative to state A Gbox is defined as in Figure 3 Initialbinding (state A to B) incurs a free energy penalty ofGinit frac14 Gvolume+Gassoc 995+190 kcalmol (11)(at concentration u frac14 50 nM) due to the reduction in theentropy of the box caused by lost translational and orien-tational degrees of freedom Gassoc is the free energy costof association at a standard concentration of u0 frac14 1 Mand Gvolume frac14 RT ln u0=ueth THORN is a correction for the actualconcentration where R is the universal gas constant and Tis the temperature in Kelvin Figure 3 shows the first basepair of the toehold adjacent to the helix where it interactsfavorably with the adjacent duplex end For simplicity weneglect sequence-dependent interaction strengths and usethe average value Gbp 17 kcalmol for this and allother base pairs as taken from the NN model (8ndash10) ofDNA secondary structure thermodynamics Formation ofeach successive base pair in the toehold therefore contrib-utes Gbp Our effective transition states which are localfree energy maxima along the sawtooth are raised Gs
above the branch migration intermediates with fullypaired substrate As the physical details of branch migra-tion steps are not well-understood the effective lsquosawtoothamplitudersquo (Gs) is an adjustable parameter ChangingGs allows branch migration and toehold melting ratesto be independently modulated for Gs gt jGbpj branchmigration is slow relative to frayingWe also introduce a final parameter a plateau height
Gp which captures how the free energy of branch mi-gration intermediates could vary with the structure of thebranch migration junction In particular there is an asym-metry between state C and all other intermediates ofbranch migration only one single-stranded overhang ispresent at the junction in state C whereas an overhangprotrudes from both sides for all other intermediatesWe thus introduce a parameter Gp that accounts for apossible free energy penalty due to the additional
overhang which could conceivably arise from entropicor electrostatic effects Henceforth IEL (GsGp)denotes a sawtooth amplitude of Gs and a plateauheight of Gp (both in kcalmol)
In an intuitive model like the IEL one could consider arange of alternative perturbations to the current biophys-ical understanding of strand displacement Gs and Gphowever parameterize in a simple way aspects of thebranch migration process that are known to be poorlycharacterized Furthermore as we show later experimentsand more detailed modeling provide physical justificationfor the effects that these parameters represent
Rate modelThe system may undergo a transition to either of itsneighboring states on the 1D landscape To ensure thatstochastic simulations will eventually converge to thethermodynamic (Boltzmann) equilibrium over the statestransition rates must satisfy detailed balance ie
kijkjifrac14 e
Gbox ethjTHORNGbox ethiTHORN
RT eth1THORN
Here kij is the transition rate from i to j Equation (1) onlyfixes the relative rates in principle for each pair ofadjacent states i and j an independent scaling factorcould be chosen Consequently the number of candidaterate models is enormous In the spirit of minimal param-eterization to avoid over-fitting the data we use only twoindependent scaling factors kbi for all bimolecular andkuni for all unimolecular transitions
For unimolecular transitions (all except A B andD E in Figure 3) we use a model in which all energet-ically downhill steps have a constant rate kuni This is asensible first approximation given that these steps involveforming a base pair at the end of a duplex or steppingdown from the transition states of branch migrationwhich may resemble forming a base pair at the junctionIn other words for GboxethiTHORN gt GboxethjTHORN
kij frac14 kuni eth2THORN
and therefore
kji frac14 kuni e
Gbox ethiTHORNGbox ethjTHORN
RT eth3THORN
This model is a continuous-time variant of theMetropolis (40) scheme for calculating thermodynamicaverages
For bimolecular transitions (A B or D E) theIEL assumes that complexes join at the constant ratekbi u and calculates the reverse rate by detailedbalance If i to j is a join step
kij frac14 kbi u frac14 kbi e
GvolumeRT u0 eth4THORN
and
kji frac14 kbi e
Gboxethi THORNGboxeth j THORN+GvolumeRT u0 eth5THORN
We choose kbi to be 3 106Ms based on the hybrid-ization rate constant fitted by Zhang and Winfree (30)For simplicity we choose kuni such that the dissociation
Figure 3 Free energy landscape of the IEL at 25C for a six basetoehold States AndashF and the sawtooth amplitude (Gs) and plateauheight (Gp) parameters are described in the text Gs frac14 26 kcalmol and Gp frac14 12 kcalmol are used for illustration
4 Nucleic Acids Research 2013
rate of the last base pair of the toehold (state B to A) isapproximately equal to the fraying rate for every otherbase pair of the toehold (steps from state C toward B)This yields kuni frac14 75 107s (see Supplementary SectionS2 for details) This choice of kuni at the very leastensures that two somewhat similar processes havesimilar rates
Analytic and numerical calculationsWe use an analytic formula for calculating absorptionprobabilities for a 1D random walk with absorbingboundaries (41) to calculate keff as a function of toeholdlength h for various values Gs and Gp (Figure 4A)(For details see Supplementary Section S2) To quantifythe extent of control provided by toeholds we define
Ah2 h1 frac14 log10ethkeffethh1THORNTHORN log10ethkeffethh2THORNTHORN eth6THORN
to be the orders of magnitude acceleration in keff astoehold length increases from h1 to h2 We will be mostinterested in A150 which corresponds to the differencebetween the leak rate and the maximal rate measured inZhang and Winfree (30) [We choose 15 rather than 1because the length-dependence of hybridization rates(33) suggests that experimental values for Ah0 may notbe bounded but for our purposes there is insignificantchange past length 15] The experimental results ofZhang and Winfree (30) can be matched by the IEL butonly with surprisingly large values of Gs and GpIEL(26 0) with a plausible value of Gs frac14 26 kcalmol (15 base pair stacks) and an a priori choice ofGp frac14 0 kcalmol predicts A150 frac14 31 This is 34orders of magnitude smaller than the experimentallyobserved value of 65 Increasing either Gs or Gp
serves to increase the predicted A150 as shown by thecontour plot in Figure 4B The slope of contour linessuggests that Gs+p frac14
defGs+Gp is the key quantity
Indeed a scatter plot of the dependence on Gs+p
(Figure 4C) using all the data in Figure 4B producesalmost no vertical spread Gs+p frac14 73 kcalmol matchesthe experimentally observed value of A150 frac14 65
Other perturbations of the IEL model were unable tomatch the experiments For example we considered analternative to the Metropolis method for setting unimol-ecular rates the Kawasaki (42) method which scales bothuphill and downhill transition rates based on the corres-ponding change in Gbox After rescaling kuni so thatdissociation of the last toehold base pair still occurs atthe same rate as fraying the Kawasaki method predicts avalue for A150 within 10 of the Metropolis method (seeSupplementary Figure S3) Going further and consideringthat kbi and kuni may not be chosen ideally we first notethat logically a uniform change to both rates will canceland thus have no effect on A150 In contrast IEL predic-tions do depend on the ratio kuni=kbi which substantiallyaffects the probabilities that initial contacts lead to success-ful zippering of the toehold and to successful branch mi-gration before dissociation (see Supplementary Figure S4and Supplementary Section S2) However even implaus-ibly low values of kuni=kbi cannot account for the datawithout a large value of Gs+pThese features of the IEL can be understood through
simple analytical approximations Conceptually we cansplit the strand displacement process into an attachmentstep (AB) followed by success or failure of zipping upthe toehold followed by either dissociation of the toeholdor successful displacement We first treat the case of longtoeholds for which in the IEL model keff saturates at
keffeth1THORN kbi pzip eth7THORN
with pzip frac14 kuni=ethkuni+kbiTHORN and frac14 eethjGbpjGassocTHORN=RTu0giving the approximate probability that after making thefirst base pair the invader does not dissociate and theremaining toehold bases zip up (For long toeholds suc-cessful displacement is guaranteed once the toehold isformed) For shorter toeholds that do not saturate keffthe probability of toehold dissociation before branchmigration dominates and we can derive
keffethhTHORN kuni2b e
h jGbp jGs+pGassoc
RT u0 eth8THORN
A B C
Figure 4 (A) Predictions of IEL (Gs Gp) for different values of the sawtooth amplitude Gs and plateau height Gp Gs+p needs to be as highas 73 kcalmol (gt4 base-pair stacks) to match experiment (30) (B) Contour plot of orders of magnitude acceleration (A150) as a function of(Gs Gp) for Gs 2 frac12073 kcalmol and Gp 2 frac12073 kcalmol (C) Scatter plot of A150 versus Gs+p using all the data in (B) The points plottedshow almost no vertical spread meaning that all points lie on a line This indicates that Gs+p is the predictive quantity Inset IEL (13 0) and IEL(50 50) for a 1-base toehold The bimolecular initial binding step which is identical in both landscapes is marked in orange to highlight thecontrast between the landscapes
Nucleic Acids Research 2013 5
for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

stable configurations Our system initially comprises atwo-stranded complex (S) and a single-stranded lsquoinvaderrsquo(X) S consists of an lsquoincumbentrsquo strand (Y) bound to alsquosubstratersquo strand which has a single-stranded overhangcalled a lsquotoeholdrsquo The invader is fully WatsonndashCrickcomplementary to the substrate and may bind reversiblyto it using the toehold domain (h) This binding is revers-ible because the toehold may lsquofrayrsquo and eventually dissoci-ate (We use the term lsquofrayingrsquo to describe the disruptionof base pairs at the end of a duplex if all base pairs fraythe duplex melts or dissociates Conversely lsquozipperingrsquorefers to when a new base pair forms at the end of anexisting duplex) Once the toehold is bound the lsquoover-hangingrsquo branch migration domain (b) of the invadermay compete with the incumbent for binding with thesubstrate As the incumbent and invader exchange basepairs with the substrate the branch point of the three-stranded complex moves back and forth This lsquothree-waybranch migrationrsquo (henceforth lsquobranch migrationrsquo)process has previously been modeled as an unbiasedrandom walk (28ndash30) as each step causes no net changein base pairing Eventually the incumbent may dissociatecompleting strand displacement Overall displacement isthermodynamically driven forward by the net gain in basepairs due to the toeholdAlthough it is known that bimolecular rate constants
for strand displacement can vary a million-fold dependingon the length of the toehold little is known about theunderlying biophysics or how to generalize the results toother reaction conditions or molecular modifications Forexample what are the mechanisms and intermediate statesinvolved in a single step of branch migration How do thekinetics of strand displacement depend on the length ofthe branch migration domain or on the temperature andbuffer conditions How different is strand displacementin RNA and DNA What is the effect of sequencemismatches at different positions along the branch migra-tion domain To begin answering these questions a fun-damental biophysical understanding is necessary
However the commonly accepted view of stranddisplacement biophysics appears at odds with kinetic meas-urements (3031) Consider strand displacement with a1-base toehold and a 20-base branch migration domainLet us assume that the rate constant for the formationof the toehold base pair is on the order of 106Ms [Thisvalue for 1-nt association is within a factor of two ofexperimental values for 6- 10- 15- and 20-mer hybridiza-tion rates (3032) after accounting for the linear length-dependence for short oligonucleotides (33)] Once thetoehold has bound there are two possibilities (i) thetoehold base pair could dissociate leading to the dissoci-ation of the invader or (ii) the nearest base pair of thesubstrate-incumbent complex could fray allowing theinvader to compete to replace that base pair andcomplete the first step of branch migration Assuming asmay seem reasonable that the rate at which either base pairfrays is similar process (ii) should be approximately half asfast as process (i) This is because once the substrate-incumbent base pair frays there is a 50 chance of theinvader replacing the frayed base pair and a 50 chanceof returning to the initial step Once the first step of branchmigration is complete subsequent forward and backwardsteps are assumed to occur at the same rate Therefore theprobability of successfully completing the remaining stepsof branch migration before going back to the toehold-only-bound state is 120 from the gamblerrsquos ruin analysis [seeSection 142 of Feller (34)] The lifetime of the three-stranded intermediate should be independent of concentra-tion Therefore at low enough concentrations the overallreaction can be modeled as an instantaneous second-orderprocess as reported by Zhang and Winfree (30) Underthese conditions the overall effective rate constant (keff)is given by a hybridization rate constant for the toeholdmultiplied by a success probability of displacement oncethe toehold is bound Even though the time spent in thethree-stranded intermediate is small keff depends stronglyon the probability of displacement once boundkeff 106 eth1=3THORN eth1=20THORN frac14 16 104Ms for a one-base toehold This is over three orders of magnitudelarger than the 8Ms value measured by Zhang andWinfree (30) This is a large unexplained discrepancydespite the approximate nature of our calculation
We now summarize the experimental evidence for theexponential acceleration in keff with toehold length whichwas first reported by Yurke and Mills (31) Zhang andWinfree (30) further characterized this exponential accel-eration and confirmed that it saturates in the long-toeholdlimit Relevant data from both studies (Figure 2A) suggestthat the exponential acceleration in itself is not an artifactof particular sequences although details may well dependon the experimental system (sequences modifications forread-out etc) or conditions (salt temperature etc) Thekinetics of zero-toehold (lsquoblunt endrsquo) strand displacementwas investigated by Reynaldo et al (35) whose measure-ment of 36Ms at 30C is similar to the 14Ms at 25Creported by Zhang and Winfree (30)
Zhang and Winfree (30) built a phenomenologicalmodel for predicting keff from toehold sequence(Figure 2B) They modeled branch migration using twomacro-states I and J containing the first and second
A B
Figure 1 (A) Domain notation Arrows indicate 30-ends Asteriskindicates WatsonndashCrick complementarity (B) Toehold h mediates thedisplacement of the incumbent (Y) by the invader (X) Dots indicatebranch migration intermediates which are not shown
2 Nucleic Acids Research 2013
half of the isoenergetic branch migration intermediatesrespectively which are connected by a simple transitionwith rate constant kb (see Supplementary Figure S1 andSupplementary Section S1 for more details) Their modelfits the data but it is hard to physically interpret the tran-sition between I and J Branch migration is generallythought of as a random walk through many isoenergeticstates rather than a single reversible transitioncharacterized by a first-order rate constant Without amore nuanced understanding of the process it is unclearwhether the fitted value of kb frac14 10s can be justified onmore fundamental biophysical grounds
In this work we model branch migration at a moredetailed level that explicitly includes intermediatesthereby highlighting important thermodynamic andkinetic features of the process that are not evident fromthe phenomenological approach
First we analyze a 1D (single-pathway) model oftoehold-mediated strand displacement called the intuitiveenergy landscape (IEL) model Systematically exploringthe parameter space of this simple model suggests thatsome combination of two factors could explain the de-pendence of strand displacement rate on toehold length(i) the branch migration process is not isoenergetic andcontains a free energy penalty for intermediate statesand (ii) branch migration is slow relative to fraying ofthe toehold
To verify that these factors are not already implicitfeatures of more detailed models of DNA secondary struc-ture thermodynamics we simulated the experiments ofZhang and Winfree (30) using a secondary structurekinetics (SSK) simulator called Multistrand (36)Multistrand incorporates extensive thermodynamic infor-mationmdashfrom state-of-the-art nearest-neighbor (NN)thermodynamic models of DNA secondary structure(8ndash11)mdashcombined with a minimally parameterized ratemodel Multistrand predicts that keff would increase by afactor of 1038 as toehold length increases from 0 to 15 incontrast to the experimentally observed factor of 1065This quantitative discrepancy confirms that factors(i) and (ii) suggested by the IEL analysis are not alreadyimplicitly incorporated in more detailed models of DNAsecondary structure thermodynamics
We then experimentally investigate possibility (i) sug-gested by the IEL by measuring the relative stability of
complexes that mimic the geometric structure of branchmigration intermediates Our experiments provideevidence in support of a free energy penalty for branchmigration intermediates that is not predicted by the NNmodels of DNAFinally we use a recently proposed coarse-grained
molecular model of DNA (oxDNA) (37ndash39) which incorp-orates more physical detail including geometric and stericeffects In addition to correctly predicting the length-dependence of toehold-mediated strand displacementrates oxDNA independently predicts the thermodynamicpenalty for branch migration intermediates suggesting itarises from local steric effects between single-strandedoverhangs around the branch point Further oxDNApredicts that branch migration is slower than fraying ofthe toehold as it is geometrically more complex and neces-sarily involves more thermodynamically unfavorable steps
MATERIALS METHODS AND RESULTS
Intuitive Energy Landscape model
Through a simple approximate calculation we argued thatthe current view of strand displacement biophysics is atodds with kinetic measurements (3031) We now performa more rigorous intuitive analysis by building a simplesingle-pathway model called the IEL model The IEL issimple enough that its kinetic predictions may be analyt-ically or numerically calculated for a given parameteriza-tion We systematically explore the IELrsquos parameter spaceto obtain biophysical intuition and identify key thermo-dynamic and kinetic features essential for predictions tomatch experimental data
State spaceThe IEL considers an invading strand and a substrate-incumbent complex contained in a virtual box of volumeV in solution which defines a concentration u of onemolecule per volume V The state space of the IEL isillustrated in Figure 3 State A corresponds to theinvader being unattached to the substrate-incumbentduplex State B represents the formation of a first basepair within the toehold For each additional toeholdbase pair that zips up we define a new state with stateC indicating the fully formed toehold From here eachstep of branch migration involves replacing an incum-bent-substrate base pair with an invader-substrate basepair We describe the stepping between these intermediatesusing a simple model in which the system must passthough a single effective transition state of raised freeenergy so each complete branch migration step is shownas a single tooth of the lsquosawtoothrsquo pattern between states Cand D This unknown effective transition state could bepotentially as simple as a frayed substrate-incumbent basepair but could also be more complex The final stageof successful displacement involves the dissociation ofthe incumbent (state E) followed by the formation ofthe final base pair between invader and substrate(state F) Subtleties relating to the zero-toehold case arediscussed in Supplementary Section S2 (see Supple-mentary Figure S2)
A B
Figure 2 (A) Dependence of keff on toehold length measured byYurke and Mills (31) (at 20C 1M Na+) and Zhang and Winfree(30) (at 25C 125mM Mg++) Each curve is from a differenttoehold sequence (B) The phenomenological model of Zhang andWinfree (30) kd is assumed to be large relative to other rate constants
Nucleic Acids Research 2013 3
Energy modelThe IEL models the free energy of the virtual box (Gbox)relative to state A Gbox is defined as in Figure 3 Initialbinding (state A to B) incurs a free energy penalty ofGinit frac14 Gvolume+Gassoc 995+190 kcalmol (11)(at concentration u frac14 50 nM) due to the reduction in theentropy of the box caused by lost translational and orien-tational degrees of freedom Gassoc is the free energy costof association at a standard concentration of u0 frac14 1 Mand Gvolume frac14 RT ln u0=ueth THORN is a correction for the actualconcentration where R is the universal gas constant and Tis the temperature in Kelvin Figure 3 shows the first basepair of the toehold adjacent to the helix where it interactsfavorably with the adjacent duplex end For simplicity weneglect sequence-dependent interaction strengths and usethe average value Gbp 17 kcalmol for this and allother base pairs as taken from the NN model (8ndash10) ofDNA secondary structure thermodynamics Formation ofeach successive base pair in the toehold therefore contrib-utes Gbp Our effective transition states which are localfree energy maxima along the sawtooth are raised Gs
above the branch migration intermediates with fullypaired substrate As the physical details of branch migra-tion steps are not well-understood the effective lsquosawtoothamplitudersquo (Gs) is an adjustable parameter ChangingGs allows branch migration and toehold melting ratesto be independently modulated for Gs gt jGbpj branchmigration is slow relative to frayingWe also introduce a final parameter a plateau height
Gp which captures how the free energy of branch mi-gration intermediates could vary with the structure of thebranch migration junction In particular there is an asym-metry between state C and all other intermediates ofbranch migration only one single-stranded overhang ispresent at the junction in state C whereas an overhangprotrudes from both sides for all other intermediatesWe thus introduce a parameter Gp that accounts for apossible free energy penalty due to the additional
overhang which could conceivably arise from entropicor electrostatic effects Henceforth IEL (GsGp)denotes a sawtooth amplitude of Gs and a plateauheight of Gp (both in kcalmol)
In an intuitive model like the IEL one could consider arange of alternative perturbations to the current biophys-ical understanding of strand displacement Gs and Gphowever parameterize in a simple way aspects of thebranch migration process that are known to be poorlycharacterized Furthermore as we show later experimentsand more detailed modeling provide physical justificationfor the effects that these parameters represent
Rate modelThe system may undergo a transition to either of itsneighboring states on the 1D landscape To ensure thatstochastic simulations will eventually converge to thethermodynamic (Boltzmann) equilibrium over the statestransition rates must satisfy detailed balance ie
kijkjifrac14 e
Gbox ethjTHORNGbox ethiTHORN
RT eth1THORN
Here kij is the transition rate from i to j Equation (1) onlyfixes the relative rates in principle for each pair ofadjacent states i and j an independent scaling factorcould be chosen Consequently the number of candidaterate models is enormous In the spirit of minimal param-eterization to avoid over-fitting the data we use only twoindependent scaling factors kbi for all bimolecular andkuni for all unimolecular transitions
For unimolecular transitions (all except A B andD E in Figure 3) we use a model in which all energet-ically downhill steps have a constant rate kuni This is asensible first approximation given that these steps involveforming a base pair at the end of a duplex or steppingdown from the transition states of branch migrationwhich may resemble forming a base pair at the junctionIn other words for GboxethiTHORN gt GboxethjTHORN
kij frac14 kuni eth2THORN
and therefore
kji frac14 kuni e
Gbox ethiTHORNGbox ethjTHORN
RT eth3THORN
This model is a continuous-time variant of theMetropolis (40) scheme for calculating thermodynamicaverages
For bimolecular transitions (A B or D E) theIEL assumes that complexes join at the constant ratekbi u and calculates the reverse rate by detailedbalance If i to j is a join step
kij frac14 kbi u frac14 kbi e
GvolumeRT u0 eth4THORN
and
kji frac14 kbi e
Gboxethi THORNGboxeth j THORN+GvolumeRT u0 eth5THORN
We choose kbi to be 3 106Ms based on the hybrid-ization rate constant fitted by Zhang and Winfree (30)For simplicity we choose kuni such that the dissociation
Figure 3 Free energy landscape of the IEL at 25C for a six basetoehold States AndashF and the sawtooth amplitude (Gs) and plateauheight (Gp) parameters are described in the text Gs frac14 26 kcalmol and Gp frac14 12 kcalmol are used for illustration
4 Nucleic Acids Research 2013
rate of the last base pair of the toehold (state B to A) isapproximately equal to the fraying rate for every otherbase pair of the toehold (steps from state C toward B)This yields kuni frac14 75 107s (see Supplementary SectionS2 for details) This choice of kuni at the very leastensures that two somewhat similar processes havesimilar rates
Analytic and numerical calculationsWe use an analytic formula for calculating absorptionprobabilities for a 1D random walk with absorbingboundaries (41) to calculate keff as a function of toeholdlength h for various values Gs and Gp (Figure 4A)(For details see Supplementary Section S2) To quantifythe extent of control provided by toeholds we define
Ah2 h1 frac14 log10ethkeffethh1THORNTHORN log10ethkeffethh2THORNTHORN eth6THORN
to be the orders of magnitude acceleration in keff astoehold length increases from h1 to h2 We will be mostinterested in A150 which corresponds to the differencebetween the leak rate and the maximal rate measured inZhang and Winfree (30) [We choose 15 rather than 1because the length-dependence of hybridization rates(33) suggests that experimental values for Ah0 may notbe bounded but for our purposes there is insignificantchange past length 15] The experimental results ofZhang and Winfree (30) can be matched by the IEL butonly with surprisingly large values of Gs and GpIEL(26 0) with a plausible value of Gs frac14 26 kcalmol (15 base pair stacks) and an a priori choice ofGp frac14 0 kcalmol predicts A150 frac14 31 This is 34orders of magnitude smaller than the experimentallyobserved value of 65 Increasing either Gs or Gp
serves to increase the predicted A150 as shown by thecontour plot in Figure 4B The slope of contour linessuggests that Gs+p frac14
defGs+Gp is the key quantity
Indeed a scatter plot of the dependence on Gs+p
(Figure 4C) using all the data in Figure 4B producesalmost no vertical spread Gs+p frac14 73 kcalmol matchesthe experimentally observed value of A150 frac14 65
Other perturbations of the IEL model were unable tomatch the experiments For example we considered analternative to the Metropolis method for setting unimol-ecular rates the Kawasaki (42) method which scales bothuphill and downhill transition rates based on the corres-ponding change in Gbox After rescaling kuni so thatdissociation of the last toehold base pair still occurs atthe same rate as fraying the Kawasaki method predicts avalue for A150 within 10 of the Metropolis method (seeSupplementary Figure S3) Going further and consideringthat kbi and kuni may not be chosen ideally we first notethat logically a uniform change to both rates will canceland thus have no effect on A150 In contrast IEL predic-tions do depend on the ratio kuni=kbi which substantiallyaffects the probabilities that initial contacts lead to success-ful zippering of the toehold and to successful branch mi-gration before dissociation (see Supplementary Figure S4and Supplementary Section S2) However even implaus-ibly low values of kuni=kbi cannot account for the datawithout a large value of Gs+pThese features of the IEL can be understood through
simple analytical approximations Conceptually we cansplit the strand displacement process into an attachmentstep (AB) followed by success or failure of zipping upthe toehold followed by either dissociation of the toeholdor successful displacement We first treat the case of longtoeholds for which in the IEL model keff saturates at
keffeth1THORN kbi pzip eth7THORN
with pzip frac14 kuni=ethkuni+kbiTHORN and frac14 eethjGbpjGassocTHORN=RTu0giving the approximate probability that after making thefirst base pair the invader does not dissociate and theremaining toehold bases zip up (For long toeholds suc-cessful displacement is guaranteed once the toehold isformed) For shorter toeholds that do not saturate keffthe probability of toehold dissociation before branchmigration dominates and we can derive
keffethhTHORN kuni2b e
h jGbp jGs+pGassoc
RT u0 eth8THORN
A B C
Figure 4 (A) Predictions of IEL (Gs Gp) for different values of the sawtooth amplitude Gs and plateau height Gp Gs+p needs to be as highas 73 kcalmol (gt4 base-pair stacks) to match experiment (30) (B) Contour plot of orders of magnitude acceleration (A150) as a function of(Gs Gp) for Gs 2 frac12073 kcalmol and Gp 2 frac12073 kcalmol (C) Scatter plot of A150 versus Gs+p using all the data in (B) The points plottedshow almost no vertical spread meaning that all points lie on a line This indicates that Gs+p is the predictive quantity Inset IEL (13 0) and IEL(50 50) for a 1-base toehold The bimolecular initial binding step which is identical in both landscapes is marked in orange to highlight thecontrast between the landscapes
Nucleic Acids Research 2013 5
for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

half of the isoenergetic branch migration intermediatesrespectively which are connected by a simple transitionwith rate constant kb (see Supplementary Figure S1 andSupplementary Section S1 for more details) Their modelfits the data but it is hard to physically interpret the tran-sition between I and J Branch migration is generallythought of as a random walk through many isoenergeticstates rather than a single reversible transitioncharacterized by a first-order rate constant Without amore nuanced understanding of the process it is unclearwhether the fitted value of kb frac14 10s can be justified onmore fundamental biophysical grounds
In this work we model branch migration at a moredetailed level that explicitly includes intermediatesthereby highlighting important thermodynamic andkinetic features of the process that are not evident fromthe phenomenological approach
First we analyze a 1D (single-pathway) model oftoehold-mediated strand displacement called the intuitiveenergy landscape (IEL) model Systematically exploringthe parameter space of this simple model suggests thatsome combination of two factors could explain the de-pendence of strand displacement rate on toehold length(i) the branch migration process is not isoenergetic andcontains a free energy penalty for intermediate statesand (ii) branch migration is slow relative to fraying ofthe toehold
To verify that these factors are not already implicitfeatures of more detailed models of DNA secondary struc-ture thermodynamics we simulated the experiments ofZhang and Winfree (30) using a secondary structurekinetics (SSK) simulator called Multistrand (36)Multistrand incorporates extensive thermodynamic infor-mationmdashfrom state-of-the-art nearest-neighbor (NN)thermodynamic models of DNA secondary structure(8ndash11)mdashcombined with a minimally parameterized ratemodel Multistrand predicts that keff would increase by afactor of 1038 as toehold length increases from 0 to 15 incontrast to the experimentally observed factor of 1065This quantitative discrepancy confirms that factors(i) and (ii) suggested by the IEL analysis are not alreadyimplicitly incorporated in more detailed models of DNAsecondary structure thermodynamics
We then experimentally investigate possibility (i) sug-gested by the IEL by measuring the relative stability of
complexes that mimic the geometric structure of branchmigration intermediates Our experiments provideevidence in support of a free energy penalty for branchmigration intermediates that is not predicted by the NNmodels of DNAFinally we use a recently proposed coarse-grained
molecular model of DNA (oxDNA) (37ndash39) which incorp-orates more physical detail including geometric and stericeffects In addition to correctly predicting the length-dependence of toehold-mediated strand displacementrates oxDNA independently predicts the thermodynamicpenalty for branch migration intermediates suggesting itarises from local steric effects between single-strandedoverhangs around the branch point Further oxDNApredicts that branch migration is slower than fraying ofthe toehold as it is geometrically more complex and neces-sarily involves more thermodynamically unfavorable steps
MATERIALS METHODS AND RESULTS
Intuitive Energy Landscape model
Through a simple approximate calculation we argued thatthe current view of strand displacement biophysics is atodds with kinetic measurements (3031) We now performa more rigorous intuitive analysis by building a simplesingle-pathway model called the IEL model The IEL issimple enough that its kinetic predictions may be analyt-ically or numerically calculated for a given parameteriza-tion We systematically explore the IELrsquos parameter spaceto obtain biophysical intuition and identify key thermo-dynamic and kinetic features essential for predictions tomatch experimental data
State spaceThe IEL considers an invading strand and a substrate-incumbent complex contained in a virtual box of volumeV in solution which defines a concentration u of onemolecule per volume V The state space of the IEL isillustrated in Figure 3 State A corresponds to theinvader being unattached to the substrate-incumbentduplex State B represents the formation of a first basepair within the toehold For each additional toeholdbase pair that zips up we define a new state with stateC indicating the fully formed toehold From here eachstep of branch migration involves replacing an incum-bent-substrate base pair with an invader-substrate basepair We describe the stepping between these intermediatesusing a simple model in which the system must passthough a single effective transition state of raised freeenergy so each complete branch migration step is shownas a single tooth of the lsquosawtoothrsquo pattern between states Cand D This unknown effective transition state could bepotentially as simple as a frayed substrate-incumbent basepair but could also be more complex The final stageof successful displacement involves the dissociation ofthe incumbent (state E) followed by the formation ofthe final base pair between invader and substrate(state F) Subtleties relating to the zero-toehold case arediscussed in Supplementary Section S2 (see Supple-mentary Figure S2)
A B
Figure 2 (A) Dependence of keff on toehold length measured byYurke and Mills (31) (at 20C 1M Na+) and Zhang and Winfree(30) (at 25C 125mM Mg++) Each curve is from a differenttoehold sequence (B) The phenomenological model of Zhang andWinfree (30) kd is assumed to be large relative to other rate constants
Nucleic Acids Research 2013 3
Energy modelThe IEL models the free energy of the virtual box (Gbox)relative to state A Gbox is defined as in Figure 3 Initialbinding (state A to B) incurs a free energy penalty ofGinit frac14 Gvolume+Gassoc 995+190 kcalmol (11)(at concentration u frac14 50 nM) due to the reduction in theentropy of the box caused by lost translational and orien-tational degrees of freedom Gassoc is the free energy costof association at a standard concentration of u0 frac14 1 Mand Gvolume frac14 RT ln u0=ueth THORN is a correction for the actualconcentration where R is the universal gas constant and Tis the temperature in Kelvin Figure 3 shows the first basepair of the toehold adjacent to the helix where it interactsfavorably with the adjacent duplex end For simplicity weneglect sequence-dependent interaction strengths and usethe average value Gbp 17 kcalmol for this and allother base pairs as taken from the NN model (8ndash10) ofDNA secondary structure thermodynamics Formation ofeach successive base pair in the toehold therefore contrib-utes Gbp Our effective transition states which are localfree energy maxima along the sawtooth are raised Gs
above the branch migration intermediates with fullypaired substrate As the physical details of branch migra-tion steps are not well-understood the effective lsquosawtoothamplitudersquo (Gs) is an adjustable parameter ChangingGs allows branch migration and toehold melting ratesto be independently modulated for Gs gt jGbpj branchmigration is slow relative to frayingWe also introduce a final parameter a plateau height
Gp which captures how the free energy of branch mi-gration intermediates could vary with the structure of thebranch migration junction In particular there is an asym-metry between state C and all other intermediates ofbranch migration only one single-stranded overhang ispresent at the junction in state C whereas an overhangprotrudes from both sides for all other intermediatesWe thus introduce a parameter Gp that accounts for apossible free energy penalty due to the additional
overhang which could conceivably arise from entropicor electrostatic effects Henceforth IEL (GsGp)denotes a sawtooth amplitude of Gs and a plateauheight of Gp (both in kcalmol)
In an intuitive model like the IEL one could consider arange of alternative perturbations to the current biophys-ical understanding of strand displacement Gs and Gphowever parameterize in a simple way aspects of thebranch migration process that are known to be poorlycharacterized Furthermore as we show later experimentsand more detailed modeling provide physical justificationfor the effects that these parameters represent
Rate modelThe system may undergo a transition to either of itsneighboring states on the 1D landscape To ensure thatstochastic simulations will eventually converge to thethermodynamic (Boltzmann) equilibrium over the statestransition rates must satisfy detailed balance ie
kijkjifrac14 e
Gbox ethjTHORNGbox ethiTHORN
RT eth1THORN
Here kij is the transition rate from i to j Equation (1) onlyfixes the relative rates in principle for each pair ofadjacent states i and j an independent scaling factorcould be chosen Consequently the number of candidaterate models is enormous In the spirit of minimal param-eterization to avoid over-fitting the data we use only twoindependent scaling factors kbi for all bimolecular andkuni for all unimolecular transitions
For unimolecular transitions (all except A B andD E in Figure 3) we use a model in which all energet-ically downhill steps have a constant rate kuni This is asensible first approximation given that these steps involveforming a base pair at the end of a duplex or steppingdown from the transition states of branch migrationwhich may resemble forming a base pair at the junctionIn other words for GboxethiTHORN gt GboxethjTHORN
kij frac14 kuni eth2THORN
and therefore
kji frac14 kuni e
Gbox ethiTHORNGbox ethjTHORN
RT eth3THORN
This model is a continuous-time variant of theMetropolis (40) scheme for calculating thermodynamicaverages
For bimolecular transitions (A B or D E) theIEL assumes that complexes join at the constant ratekbi u and calculates the reverse rate by detailedbalance If i to j is a join step
kij frac14 kbi u frac14 kbi e
GvolumeRT u0 eth4THORN
and
kji frac14 kbi e
Gboxethi THORNGboxeth j THORN+GvolumeRT u0 eth5THORN
We choose kbi to be 3 106Ms based on the hybrid-ization rate constant fitted by Zhang and Winfree (30)For simplicity we choose kuni such that the dissociation
Figure 3 Free energy landscape of the IEL at 25C for a six basetoehold States AndashF and the sawtooth amplitude (Gs) and plateauheight (Gp) parameters are described in the text Gs frac14 26 kcalmol and Gp frac14 12 kcalmol are used for illustration
4 Nucleic Acids Research 2013
rate of the last base pair of the toehold (state B to A) isapproximately equal to the fraying rate for every otherbase pair of the toehold (steps from state C toward B)This yields kuni frac14 75 107s (see Supplementary SectionS2 for details) This choice of kuni at the very leastensures that two somewhat similar processes havesimilar rates
Analytic and numerical calculationsWe use an analytic formula for calculating absorptionprobabilities for a 1D random walk with absorbingboundaries (41) to calculate keff as a function of toeholdlength h for various values Gs and Gp (Figure 4A)(For details see Supplementary Section S2) To quantifythe extent of control provided by toeholds we define
Ah2 h1 frac14 log10ethkeffethh1THORNTHORN log10ethkeffethh2THORNTHORN eth6THORN
to be the orders of magnitude acceleration in keff astoehold length increases from h1 to h2 We will be mostinterested in A150 which corresponds to the differencebetween the leak rate and the maximal rate measured inZhang and Winfree (30) [We choose 15 rather than 1because the length-dependence of hybridization rates(33) suggests that experimental values for Ah0 may notbe bounded but for our purposes there is insignificantchange past length 15] The experimental results ofZhang and Winfree (30) can be matched by the IEL butonly with surprisingly large values of Gs and GpIEL(26 0) with a plausible value of Gs frac14 26 kcalmol (15 base pair stacks) and an a priori choice ofGp frac14 0 kcalmol predicts A150 frac14 31 This is 34orders of magnitude smaller than the experimentallyobserved value of 65 Increasing either Gs or Gp
serves to increase the predicted A150 as shown by thecontour plot in Figure 4B The slope of contour linessuggests that Gs+p frac14
defGs+Gp is the key quantity
Indeed a scatter plot of the dependence on Gs+p
(Figure 4C) using all the data in Figure 4B producesalmost no vertical spread Gs+p frac14 73 kcalmol matchesthe experimentally observed value of A150 frac14 65
Other perturbations of the IEL model were unable tomatch the experiments For example we considered analternative to the Metropolis method for setting unimol-ecular rates the Kawasaki (42) method which scales bothuphill and downhill transition rates based on the corres-ponding change in Gbox After rescaling kuni so thatdissociation of the last toehold base pair still occurs atthe same rate as fraying the Kawasaki method predicts avalue for A150 within 10 of the Metropolis method (seeSupplementary Figure S3) Going further and consideringthat kbi and kuni may not be chosen ideally we first notethat logically a uniform change to both rates will canceland thus have no effect on A150 In contrast IEL predic-tions do depend on the ratio kuni=kbi which substantiallyaffects the probabilities that initial contacts lead to success-ful zippering of the toehold and to successful branch mi-gration before dissociation (see Supplementary Figure S4and Supplementary Section S2) However even implaus-ibly low values of kuni=kbi cannot account for the datawithout a large value of Gs+pThese features of the IEL can be understood through
simple analytical approximations Conceptually we cansplit the strand displacement process into an attachmentstep (AB) followed by success or failure of zipping upthe toehold followed by either dissociation of the toeholdor successful displacement We first treat the case of longtoeholds for which in the IEL model keff saturates at
keffeth1THORN kbi pzip eth7THORN
with pzip frac14 kuni=ethkuni+kbiTHORN and frac14 eethjGbpjGassocTHORN=RTu0giving the approximate probability that after making thefirst base pair the invader does not dissociate and theremaining toehold bases zip up (For long toeholds suc-cessful displacement is guaranteed once the toehold isformed) For shorter toeholds that do not saturate keffthe probability of toehold dissociation before branchmigration dominates and we can derive
keffethhTHORN kuni2b e
h jGbp jGs+pGassoc
RT u0 eth8THORN
A B C
Figure 4 (A) Predictions of IEL (Gs Gp) for different values of the sawtooth amplitude Gs and plateau height Gp Gs+p needs to be as highas 73 kcalmol (gt4 base-pair stacks) to match experiment (30) (B) Contour plot of orders of magnitude acceleration (A150) as a function of(Gs Gp) for Gs 2 frac12073 kcalmol and Gp 2 frac12073 kcalmol (C) Scatter plot of A150 versus Gs+p using all the data in (B) The points plottedshow almost no vertical spread meaning that all points lie on a line This indicates that Gs+p is the predictive quantity Inset IEL (13 0) and IEL(50 50) for a 1-base toehold The bimolecular initial binding step which is identical in both landscapes is marked in orange to highlight thecontrast between the landscapes
Nucleic Acids Research 2013 5
for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

Energy modelThe IEL models the free energy of the virtual box (Gbox)relative to state A Gbox is defined as in Figure 3 Initialbinding (state A to B) incurs a free energy penalty ofGinit frac14 Gvolume+Gassoc 995+190 kcalmol (11)(at concentration u frac14 50 nM) due to the reduction in theentropy of the box caused by lost translational and orien-tational degrees of freedom Gassoc is the free energy costof association at a standard concentration of u0 frac14 1 Mand Gvolume frac14 RT ln u0=ueth THORN is a correction for the actualconcentration where R is the universal gas constant and Tis the temperature in Kelvin Figure 3 shows the first basepair of the toehold adjacent to the helix where it interactsfavorably with the adjacent duplex end For simplicity weneglect sequence-dependent interaction strengths and usethe average value Gbp 17 kcalmol for this and allother base pairs as taken from the NN model (8ndash10) ofDNA secondary structure thermodynamics Formation ofeach successive base pair in the toehold therefore contrib-utes Gbp Our effective transition states which are localfree energy maxima along the sawtooth are raised Gs
above the branch migration intermediates with fullypaired substrate As the physical details of branch migra-tion steps are not well-understood the effective lsquosawtoothamplitudersquo (Gs) is an adjustable parameter ChangingGs allows branch migration and toehold melting ratesto be independently modulated for Gs gt jGbpj branchmigration is slow relative to frayingWe also introduce a final parameter a plateau height
Gp which captures how the free energy of branch mi-gration intermediates could vary with the structure of thebranch migration junction In particular there is an asym-metry between state C and all other intermediates ofbranch migration only one single-stranded overhang ispresent at the junction in state C whereas an overhangprotrudes from both sides for all other intermediatesWe thus introduce a parameter Gp that accounts for apossible free energy penalty due to the additional
overhang which could conceivably arise from entropicor electrostatic effects Henceforth IEL (GsGp)denotes a sawtooth amplitude of Gs and a plateauheight of Gp (both in kcalmol)
In an intuitive model like the IEL one could consider arange of alternative perturbations to the current biophys-ical understanding of strand displacement Gs and Gphowever parameterize in a simple way aspects of thebranch migration process that are known to be poorlycharacterized Furthermore as we show later experimentsand more detailed modeling provide physical justificationfor the effects that these parameters represent
Rate modelThe system may undergo a transition to either of itsneighboring states on the 1D landscape To ensure thatstochastic simulations will eventually converge to thethermodynamic (Boltzmann) equilibrium over the statestransition rates must satisfy detailed balance ie
kijkjifrac14 e
Gbox ethjTHORNGbox ethiTHORN
RT eth1THORN
Here kij is the transition rate from i to j Equation (1) onlyfixes the relative rates in principle for each pair ofadjacent states i and j an independent scaling factorcould be chosen Consequently the number of candidaterate models is enormous In the spirit of minimal param-eterization to avoid over-fitting the data we use only twoindependent scaling factors kbi for all bimolecular andkuni for all unimolecular transitions
For unimolecular transitions (all except A B andD E in Figure 3) we use a model in which all energet-ically downhill steps have a constant rate kuni This is asensible first approximation given that these steps involveforming a base pair at the end of a duplex or steppingdown from the transition states of branch migrationwhich may resemble forming a base pair at the junctionIn other words for GboxethiTHORN gt GboxethjTHORN
kij frac14 kuni eth2THORN
and therefore
kji frac14 kuni e
Gbox ethiTHORNGbox ethjTHORN
RT eth3THORN
This model is a continuous-time variant of theMetropolis (40) scheme for calculating thermodynamicaverages
For bimolecular transitions (A B or D E) theIEL assumes that complexes join at the constant ratekbi u and calculates the reverse rate by detailedbalance If i to j is a join step
kij frac14 kbi u frac14 kbi e
GvolumeRT u0 eth4THORN
and
kji frac14 kbi e
Gboxethi THORNGboxeth j THORN+GvolumeRT u0 eth5THORN
We choose kbi to be 3 106Ms based on the hybrid-ization rate constant fitted by Zhang and Winfree (30)For simplicity we choose kuni such that the dissociation
Figure 3 Free energy landscape of the IEL at 25C for a six basetoehold States AndashF and the sawtooth amplitude (Gs) and plateauheight (Gp) parameters are described in the text Gs frac14 26 kcalmol and Gp frac14 12 kcalmol are used for illustration
4 Nucleic Acids Research 2013
rate of the last base pair of the toehold (state B to A) isapproximately equal to the fraying rate for every otherbase pair of the toehold (steps from state C toward B)This yields kuni frac14 75 107s (see Supplementary SectionS2 for details) This choice of kuni at the very leastensures that two somewhat similar processes havesimilar rates
Analytic and numerical calculationsWe use an analytic formula for calculating absorptionprobabilities for a 1D random walk with absorbingboundaries (41) to calculate keff as a function of toeholdlength h for various values Gs and Gp (Figure 4A)(For details see Supplementary Section S2) To quantifythe extent of control provided by toeholds we define
Ah2 h1 frac14 log10ethkeffethh1THORNTHORN log10ethkeffethh2THORNTHORN eth6THORN
to be the orders of magnitude acceleration in keff astoehold length increases from h1 to h2 We will be mostinterested in A150 which corresponds to the differencebetween the leak rate and the maximal rate measured inZhang and Winfree (30) [We choose 15 rather than 1because the length-dependence of hybridization rates(33) suggests that experimental values for Ah0 may notbe bounded but for our purposes there is insignificantchange past length 15] The experimental results ofZhang and Winfree (30) can be matched by the IEL butonly with surprisingly large values of Gs and GpIEL(26 0) with a plausible value of Gs frac14 26 kcalmol (15 base pair stacks) and an a priori choice ofGp frac14 0 kcalmol predicts A150 frac14 31 This is 34orders of magnitude smaller than the experimentallyobserved value of 65 Increasing either Gs or Gp
serves to increase the predicted A150 as shown by thecontour plot in Figure 4B The slope of contour linessuggests that Gs+p frac14
defGs+Gp is the key quantity
Indeed a scatter plot of the dependence on Gs+p
(Figure 4C) using all the data in Figure 4B producesalmost no vertical spread Gs+p frac14 73 kcalmol matchesthe experimentally observed value of A150 frac14 65
Other perturbations of the IEL model were unable tomatch the experiments For example we considered analternative to the Metropolis method for setting unimol-ecular rates the Kawasaki (42) method which scales bothuphill and downhill transition rates based on the corres-ponding change in Gbox After rescaling kuni so thatdissociation of the last toehold base pair still occurs atthe same rate as fraying the Kawasaki method predicts avalue for A150 within 10 of the Metropolis method (seeSupplementary Figure S3) Going further and consideringthat kbi and kuni may not be chosen ideally we first notethat logically a uniform change to both rates will canceland thus have no effect on A150 In contrast IEL predic-tions do depend on the ratio kuni=kbi which substantiallyaffects the probabilities that initial contacts lead to success-ful zippering of the toehold and to successful branch mi-gration before dissociation (see Supplementary Figure S4and Supplementary Section S2) However even implaus-ibly low values of kuni=kbi cannot account for the datawithout a large value of Gs+pThese features of the IEL can be understood through
simple analytical approximations Conceptually we cansplit the strand displacement process into an attachmentstep (AB) followed by success or failure of zipping upthe toehold followed by either dissociation of the toeholdor successful displacement We first treat the case of longtoeholds for which in the IEL model keff saturates at
keffeth1THORN kbi pzip eth7THORN
with pzip frac14 kuni=ethkuni+kbiTHORN and frac14 eethjGbpjGassocTHORN=RTu0giving the approximate probability that after making thefirst base pair the invader does not dissociate and theremaining toehold bases zip up (For long toeholds suc-cessful displacement is guaranteed once the toehold isformed) For shorter toeholds that do not saturate keffthe probability of toehold dissociation before branchmigration dominates and we can derive
keffethhTHORN kuni2b e
h jGbp jGs+pGassoc
RT u0 eth8THORN
A B C
Figure 4 (A) Predictions of IEL (Gs Gp) for different values of the sawtooth amplitude Gs and plateau height Gp Gs+p needs to be as highas 73 kcalmol (gt4 base-pair stacks) to match experiment (30) (B) Contour plot of orders of magnitude acceleration (A150) as a function of(Gs Gp) for Gs 2 frac12073 kcalmol and Gp 2 frac12073 kcalmol (C) Scatter plot of A150 versus Gs+p using all the data in (B) The points plottedshow almost no vertical spread meaning that all points lie on a line This indicates that Gs+p is the predictive quantity Inset IEL (13 0) and IEL(50 50) for a 1-base toehold The bimolecular initial binding step which is identical in both landscapes is marked in orange to highlight thecontrast between the landscapes
Nucleic Acids Research 2013 5
for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013
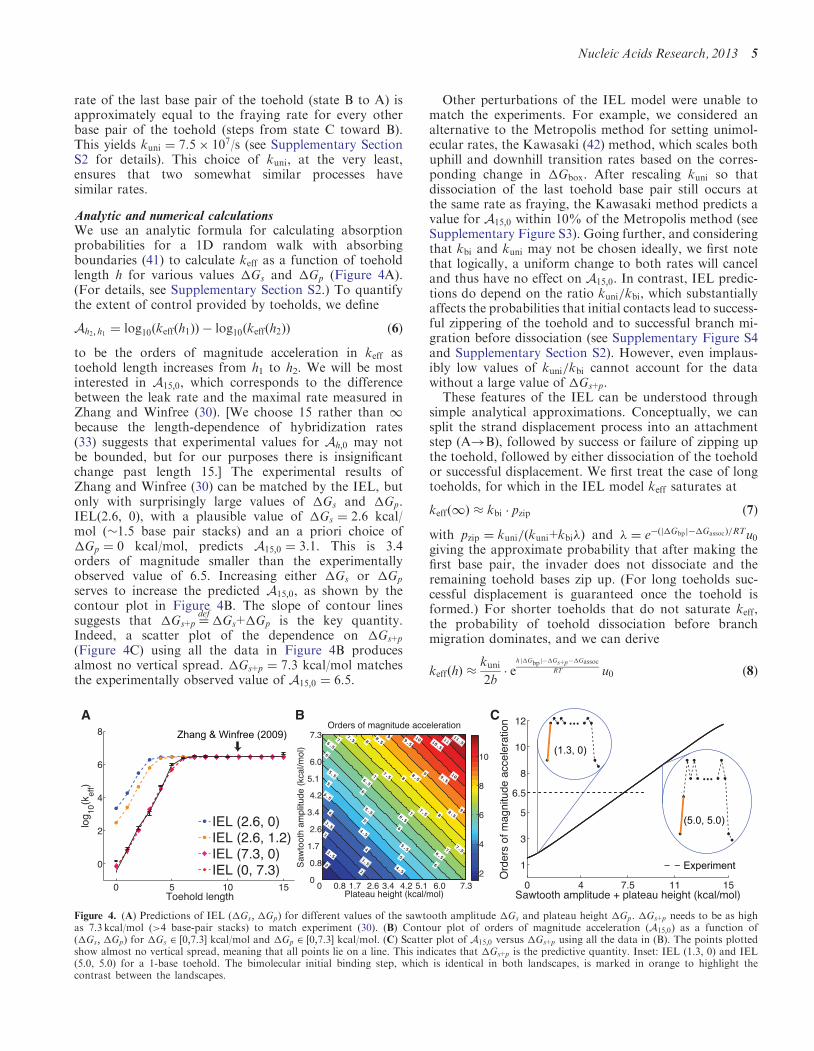
rate of the last base pair of the toehold (state B to A) isapproximately equal to the fraying rate for every otherbase pair of the toehold (steps from state C toward B)This yields kuni frac14 75 107s (see Supplementary SectionS2 for details) This choice of kuni at the very leastensures that two somewhat similar processes havesimilar rates
Analytic and numerical calculationsWe use an analytic formula for calculating absorptionprobabilities for a 1D random walk with absorbingboundaries (41) to calculate keff as a function of toeholdlength h for various values Gs and Gp (Figure 4A)(For details see Supplementary Section S2) To quantifythe extent of control provided by toeholds we define
Ah2 h1 frac14 log10ethkeffethh1THORNTHORN log10ethkeffethh2THORNTHORN eth6THORN
to be the orders of magnitude acceleration in keff astoehold length increases from h1 to h2 We will be mostinterested in A150 which corresponds to the differencebetween the leak rate and the maximal rate measured inZhang and Winfree (30) [We choose 15 rather than 1because the length-dependence of hybridization rates(33) suggests that experimental values for Ah0 may notbe bounded but for our purposes there is insignificantchange past length 15] The experimental results ofZhang and Winfree (30) can be matched by the IEL butonly with surprisingly large values of Gs and GpIEL(26 0) with a plausible value of Gs frac14 26 kcalmol (15 base pair stacks) and an a priori choice ofGp frac14 0 kcalmol predicts A150 frac14 31 This is 34orders of magnitude smaller than the experimentallyobserved value of 65 Increasing either Gs or Gp
serves to increase the predicted A150 as shown by thecontour plot in Figure 4B The slope of contour linessuggests that Gs+p frac14
defGs+Gp is the key quantity
Indeed a scatter plot of the dependence on Gs+p
(Figure 4C) using all the data in Figure 4B producesalmost no vertical spread Gs+p frac14 73 kcalmol matchesthe experimentally observed value of A150 frac14 65
Other perturbations of the IEL model were unable tomatch the experiments For example we considered analternative to the Metropolis method for setting unimol-ecular rates the Kawasaki (42) method which scales bothuphill and downhill transition rates based on the corres-ponding change in Gbox After rescaling kuni so thatdissociation of the last toehold base pair still occurs atthe same rate as fraying the Kawasaki method predicts avalue for A150 within 10 of the Metropolis method (seeSupplementary Figure S3) Going further and consideringthat kbi and kuni may not be chosen ideally we first notethat logically a uniform change to both rates will canceland thus have no effect on A150 In contrast IEL predic-tions do depend on the ratio kuni=kbi which substantiallyaffects the probabilities that initial contacts lead to success-ful zippering of the toehold and to successful branch mi-gration before dissociation (see Supplementary Figure S4and Supplementary Section S2) However even implaus-ibly low values of kuni=kbi cannot account for the datawithout a large value of Gs+pThese features of the IEL can be understood through
simple analytical approximations Conceptually we cansplit the strand displacement process into an attachmentstep (AB) followed by success or failure of zipping upthe toehold followed by either dissociation of the toeholdor successful displacement We first treat the case of longtoeholds for which in the IEL model keff saturates at
keffeth1THORN kbi pzip eth7THORN
with pzip frac14 kuni=ethkuni+kbiTHORN and frac14 eethjGbpjGassocTHORN=RTu0giving the approximate probability that after making thefirst base pair the invader does not dissociate and theremaining toehold bases zip up (For long toeholds suc-cessful displacement is guaranteed once the toehold isformed) For shorter toeholds that do not saturate keffthe probability of toehold dissociation before branchmigration dominates and we can derive
keffethhTHORN kuni2b e
h jGbp jGs+pGassoc
RT u0 eth8THORN
A B C
Figure 4 (A) Predictions of IEL (Gs Gp) for different values of the sawtooth amplitude Gs and plateau height Gp Gs+p needs to be as highas 73 kcalmol (gt4 base-pair stacks) to match experiment (30) (B) Contour plot of orders of magnitude acceleration (A150) as a function of(Gs Gp) for Gs 2 frac12073 kcalmol and Gp 2 frac12073 kcalmol (C) Scatter plot of A150 versus Gs+p using all the data in (B) The points plottedshow almost no vertical spread meaning that all points lie on a line This indicates that Gs+p is the predictive quantity Inset IEL (13 0) and IEL(50 50) for a 1-base toehold The bimolecular initial binding step which is identical in both landscapes is marked in orange to highlight thecontrast between the landscapes
Nucleic Acids Research 2013 5
for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

for h 6frac14 0 and there being two ends where branch migra-tion could start twice the given value for h=0We clearly see that the slope in Figure 4A is governed
by Gbp while the role of b (Supplementary Figure S5)and Gs+p in determining
A150 log10keffeth1THORN
keffeth0THORN 09+log10
2b eGs+p=RT
kuni=kbi+
eth9THORN
is immediate from their influence on keffeth0THORN The moresubtle effect of changing kuni=kbi results partly fromeffects on pzip a slightly more accurate approximationcan explain the behavior seen in Supplementary FigureS4 Details on this and other derivations are given inSupplementary Section S2
Matching both strand displacement and branchmigration ratesThe IEL analysis suggests that we can reconcile kineticmodels of displacement at the base pair level with the ex-perimental measurements of Zhang and Winfree (30) byincorporating a thermodynamic penalty for initiatingbranch migration (Gp) andor slowing down the rate ofbranch migration through a relatively large sawtoothamplitude (Gs) associated with each step of branchmigration Although Gs+p is constrained to be at least73 kcalmol to match the data the individual contribu-tions of Gs and Gp are not determined by the analysisTo compare the inferences of the IEL analysis with
experimental measurements we need to account for theincrease in initial binding rate (AB) with toehold lengthas observed for short oligonucleotide hybridization (33)(For simplicity the IEL assumes that the initial bindingrate is independent of toehold length) To this end we alsoexplored a variant of the IEL called the AugmentedEnergy Landscape (AEL) model that includes the linearincrease in the formation rate of the first base pair (seeSupplementary Section S3) For the AEL we find that asmaller value of Gs+p frac14 56 kcalmol is required owingto the contribution of the binding rate to the overall ac-celeration A good fit to the experimental data shownin Figure 4 is obtained for kbi frac14 33 105Ms andkuni frac14 82 106s (Supplementary Table S1)
We have not yet presented evidence to suggest that theplateau height Gp is non-zero However if Gp frac14 0 asawtooth amplitude of 73 kcalmol (56 for the AEL)would be required to account for the data which impliesan average branch migration step time of 30 ms ( 16ms for the AEL) This is much slower than experimentallyinferred step times on the order of 12ndash20 ms (2829)Therefore simultaneously matching both measuredbranch migration and strand displacement rates requiresa significant thermodynamic penalty to initiating branchmigration
The IEL analysis raises two important questions Firstcould the necessary values of Gs and Gp representfeatures missing in the IELrsquos simplified thermodynamiclandscape that are implicitly already present in moredetailed models If this is not the case are Gs andGp purely phenomenological parameters used to fit thedata or do they represent real physical effects that arisefrom the molecular properties of DNA
Secondary Structure Kinetics model
As a first step toward answering these questions we use aSSK simulator called Multistrand (36) to study strand dis-placement as a random walk on a more complex energylandscape the NN secondary structure model (8ndash11) thatincorporates a wealth of existing thermodynamic know-ledge Multistrand extends the Kinfold simulator (43)from single-strand landscapes to landscapes for multipleinteracting nucleic acid molecules Code implementing theMultistrand model is available for public download (seeSupplementary Section S4 for details)
State spaceMultistrand considers a set of strands in a virtual box ofvolume V in solution which defines a concentration u ofone molecule per volume V The state space consists of allpossible sets of WatsonndashCrick base pairs (such as states inFigure 5A) with two restrictions (i) no base can havemore than one pairing interaction and (ii) secondary struc-tures containing pseudo knots are not allowed Structureswithout pseudo knots are tree-like and have nested basepairing (11) The size of Multistrandrsquos state space growsexponentially in the number of bases (11)
A B
Figure 5 (A) Example states and elementary steps in Multistrand (36) a SSK simulator States illustrated are each adjacent to state i as they differfrom i by only one base pair Transition rates are chosen to obey detailed balance Dots and arrows at the top indicate other possible elementarysteps from state i (not shown) (B) Multistrand predictions of experimentally measured (30) strand displacement rates as a function of toehold lengthExperimental data points and error bars are from Zhang and Winfree (30) the fitted line is their phenomenological model Standard errors forMultistrand simulations are under 1 (not shown)
6 Nucleic Acids Research 2013
Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

Energy modelLet GboxethiTHORN be the free energy of the virtual box in state irelative to a completely unstructured state with no basepairs GboxethiTHORN is the sum of free energies of each isolatedcomplex c GethcTHORN in state i The free energy of eachcomplex is estimated using the NN model (8ndash10) whichhas been extended to multiple interacting nucleic acidstrands (11) NN parameters were measured (910) in1M Na+ which is roughly thermodynamically equivalentto the 125mM Mg++ used by Zhang and Winfree (30)according to salt corrections (4445) to the NN model TheNN model assumes that the free energy contribution ofeach base pair is dependent only on the identity and orien-tation (50 or 30) of its nearest neighbors on either side andcalculates GethcTHORN by summing contributions from eachsub-structure (lsquolooprsquo) closed by a base-paired sectionwhere
GethcTHORN frac14 ethL 1THORNGinit+X
loop2 c
GethloopTHORN eth10THORN
L is the number of strands in complex c and Ginit frac14
Gassoc+Gvolume is like in the IEL the free energy costof bringing two separate strands together Stabilizing con-tributions to GethloopTHORN mainly arise from base-pair stacksand destabilizing contributions arise from the entropiccost of closing loops At duplex ends in either interior orexterior loops the first overhanging nucleotide contributesa lsquodanglersquo energy term (46) When two duplex ends abutas at a nick the duplexes are known to lsquocoaxially stackrsquoonto each other (47ndash50) thereby stabilizing the structurethis interaction is not explicitly incorporated intoMultistrand although in some cases dangle energy termspartially account for it Multistrandrsquos energy model isidentical to that used in NUPACK (51) and is thussimilar to those in Vienna RNA (52) and Mfold (53)
Rate modelMultistrand allows transitions between states i and j ifthey differ by a single base pair The rate models wehave explored for Multistrand are identical to the IELexcept for scaling factors Like the IEL Multistrandrsquos pre-dictions are not particularly sensitive to the choicebetween standard unimolecular rate models The predictedorders of magnitude acceleration in keff between toeholds0 and 15 differ by less than 3 between Metropolis andKawasaki (Supplementary Figure S6B) For Metropoliskbi frac14 126 106Ms and kuni frac14 44 108s werecalibrated (36) by fitting Multistrand simulations to ex-perimentally measured DNAndashDNA hybridization (32)and zippering (33) rates respectively Given the ratemodel energy model and current state the choice andtiming of the next transition is determined using aGillespie algorithm (54) Multistrand allows any initialfirst pair of nucleotides to interact each at the standardbimolecular rate resulting in an increase in hybridizationrates with increasing toehold lengths
In principle each i to j transition could have bothforward and reverse rates scaled arbitrarily but equallythus obtaining a distinct kinetic model that still satisfiesdetailed balance However doing so would amount to
treating the microscopic barriers that impede transitionsbetween the states of the NN model as different in everycase In the absence of additional information justifyingsuch differences the Metropolis approach constitutes aminimally parameterized rate model in which microscopicbarriers associated with downhill processes (which typic-ally involve base-pair formation) are assumed to beuniform and their effects are incorporated in the valueof kuni As such the Metropolis approach allows us toask whether the free-energy landscape given by the NNmodel is sufficiently accurate and precise that it capturesthe factors that determine relative kinetics and that finerdetails are only important in setting an overall rateconstant kuni
Comparing Multistrand predictions with dataWe simulated the lsquoaverage strength toeholdrsquo experimentsof Zhang and Winfree (30) and calculated keff as afunction of toehold length For technical details regardingthe simulations see Supplementary Section S4 andSupplementary Table S2 Multistrand captures the quali-tative dependence of keff on toehold length despite itsminimally parameterized rate model HoweverMultistrand predictions quantitatively diverge from ex-periment in two ways (Figure 5B) (i) the saturationvalue of keff for toeholds longer than five bases isroughly 20 times larger and (ii) increasing toehold lengthfrom 0 to 15 increases keff by a factor of 1038 in contrastto 1065 observed in experimentThe first issue could be addressed by uniformly
decreasing both kuni and kbi which would shift the pre-dicted curve down to match the data for toeholds longerthan five bases Indeed this re-scaling is also physicallyreasonable as there is considerable variation among hy-bridization and fraying rates in the literature for nucleicacids (32 3355ndash60)Scaling kuni and kbi uniformly would not resolve the
second challenge However like the IEL Multistrand isparticularly sensitive to the ratio kuni=kbi Decreasing kunislows down all unimolecular transitionsmdashwhich brings thebranch migration rate closer to experimentally inferredvalues but makes fraying too slow Two parameters kuniand kbi are simply not enough (36) to simultaneouslymatch the four distinct time scales involved rates of hy-bridization fraying branch migration and branch migra-tion initiation Therefore even unrealistically low choicesof kuni=kbi are unable to match observed acceleration instrand displacement rates due to toehold length (seeSupplementary Figure S7)The SSK analysis confirms that understanding what the
IELrsquos Gs and Gp represent requires examining featuresnot present in the NN model Multistrand models branchmigration as a frayndashandndashreplace process and interprets theIELrsquos sawtooth transition state as one in which the sub-strate-incumbent base pair at the junction is frayed Thischoice when coupled with a kuni calibrated to matchfraying rates (33) results in a branch migration rate thatis much faster than experimentally inferred step times(2829)Indeed the thermodynamics of the branch migration
junction eg states i and j in Figure 5A is not well
Nucleic Acids Research 2013 7
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013
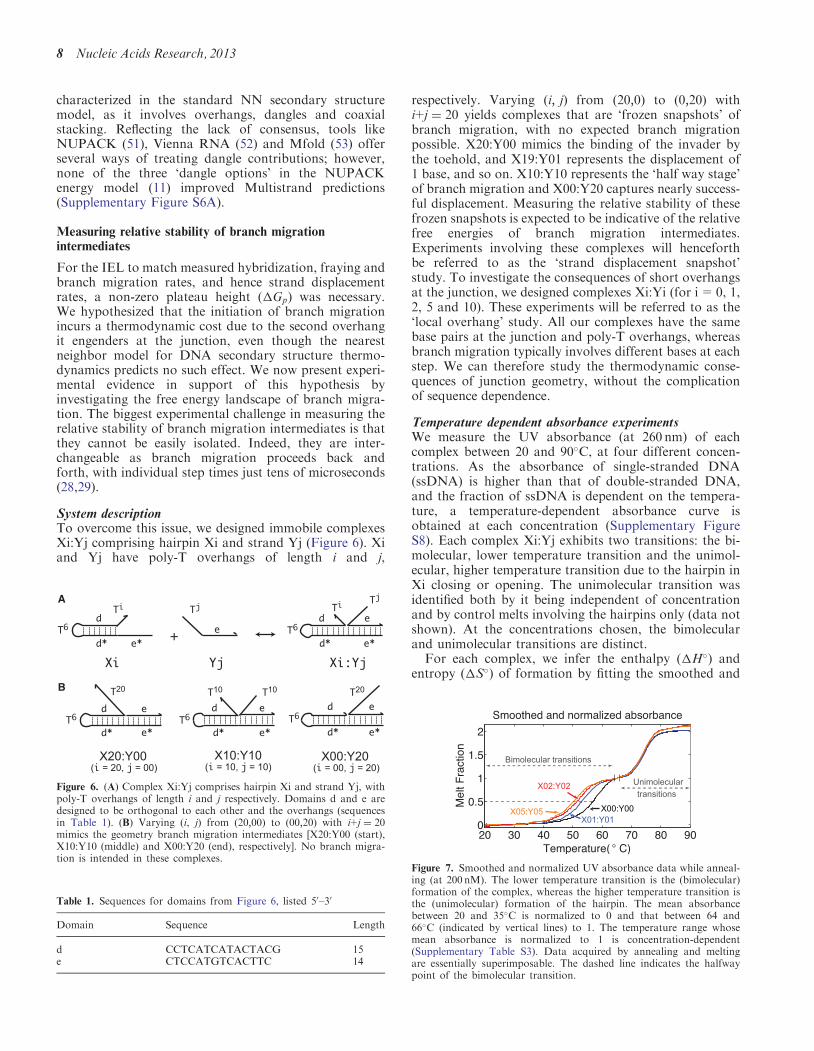
characterized in the standard NN secondary structuremodel as it involves overhangs dangles and coaxialstacking Reflecting the lack of consensus tools likeNUPACK (51) Vienna RNA (52) and Mfold (53) offerseveral ways of treating dangle contributions howevernone of the three lsquodangle optionsrsquo in the NUPACKenergy model (11) improved Multistrand predictions(Supplementary Figure S6A)
Measuring relative stability of branch migrationintermediates
For the IEL to match measured hybridization fraying andbranch migration rates and hence strand displacementrates a non-zero plateau height (Gp) was necessaryWe hypothesized that the initiation of branch migrationincurs a thermodynamic cost due to the second overhangit engenders at the junction even though the nearestneighbor model for DNA secondary structure thermo-dynamics predicts no such effect We now present experi-mental evidence in support of this hypothesis byinvestigating the free energy landscape of branch migra-tion The biggest experimental challenge in measuring therelative stability of branch migration intermediates is thatthey cannot be easily isolated Indeed they are inter-changeable as branch migration proceeds back andforth with individual step times just tens of microseconds(2829)
System descriptionTo overcome this issue we designed immobile complexesXiYj comprising hairpin Xi and strand Yj (Figure 6) Xiand Yj have poly-T overhangs of length i and j
respectively Varying ethi jTHORN from eth200THORN to eth020THORN withi+j frac14 20 yields complexes that are lsquofrozen snapshotsrsquo ofbranch migration with no expected branch migrationpossible X20Y00 mimics the binding of the invader bythe toehold and X19Y01 represents the displacement of1 base and so on X10Y10 represents the lsquohalf way stagersquoof branch migration and X00Y20 captures nearly success-ful displacement Measuring the relative stability of thesefrozen snapshots is expected to be indicative of the relativefree energies of branch migration intermediatesExperiments involving these complexes will henceforthbe referred to as the lsquostrand displacement snapshotrsquostudy To investigate the consequences of short overhangsat the junction we designed complexes XiYi (for i=0 12 5 and 10) These experiments will be referred to as thelsquolocal overhangrsquo study All our complexes have the samebase pairs at the junction and poly-T overhangs whereasbranch migration typically involves different bases at eachstep We can therefore study the thermodynamic conse-quences of junction geometry without the complicationof sequence dependence
Temperature dependent absorbance experimentsWe measure the UV absorbance (at 260 nm) of eachcomplex between 20 and 90C at four different concen-trations As the absorbance of single-stranded DNA(ssDNA) is higher than that of double-stranded DNAand the fraction of ssDNA is dependent on the tempera-ture a temperature-dependent absorbance curve isobtained at each concentration (Supplementary FigureS8) Each complex XiYj exhibits two transitions the bi-molecular lower temperature transition and the unimol-ecular higher temperature transition due to the hairpin inXi closing or opening The unimolecular transition wasidentified both by it being independent of concentrationand by control melts involving the hairpins only (data notshown) At the concentrations chosen the bimolecularand unimolecular transitions are distinct
For each complex we infer the enthalpy (H) andentropy (S) of formation by fitting the smoothed and
A
B
Figure 6 (A) Complex XiYj comprises hairpin Xi and strand Yj withpoly-T overhangs of length i and j respectively Domains d and e aredesigned to be orthogonal to each other and the overhangs (sequencesin Table 1) (B) Varying (i j) from eth2000THORN to eth0020THORN with i+j frac14 20mimics the geometry branch migration intermediates [X20Y00 (start)X10Y10 (middle) and X00Y20 (end) respectively] No branch migra-tion is intended in these complexes
Figure 7 Smoothed and normalized UV absorbance data while anneal-ing (at 200 nM) The lower temperature transition is the (bimolecular)formation of the complex whereas the higher temperature transition isthe (unimolecular) formation of the hairpin The mean absorbancebetween 20 and 35C is normalized to 0 and that between 64 and66C (indicated by vertical lines) to 1 The temperature range whosemean absorbance is normalized to 1 is concentration-dependent(Supplementary Table S3) Data acquired by annealing and meltingare essentially superimposable The dashed line indicates the halfwaypoint of the bimolecular transition
Table 1 Sequences for domains from Figure 6 listed 50ndash30
Domain Sequence Length
d CCTCATCATACTACG 15e CTCCATGTCACTTC 14
8 Nucleic Acids Research 2013
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013
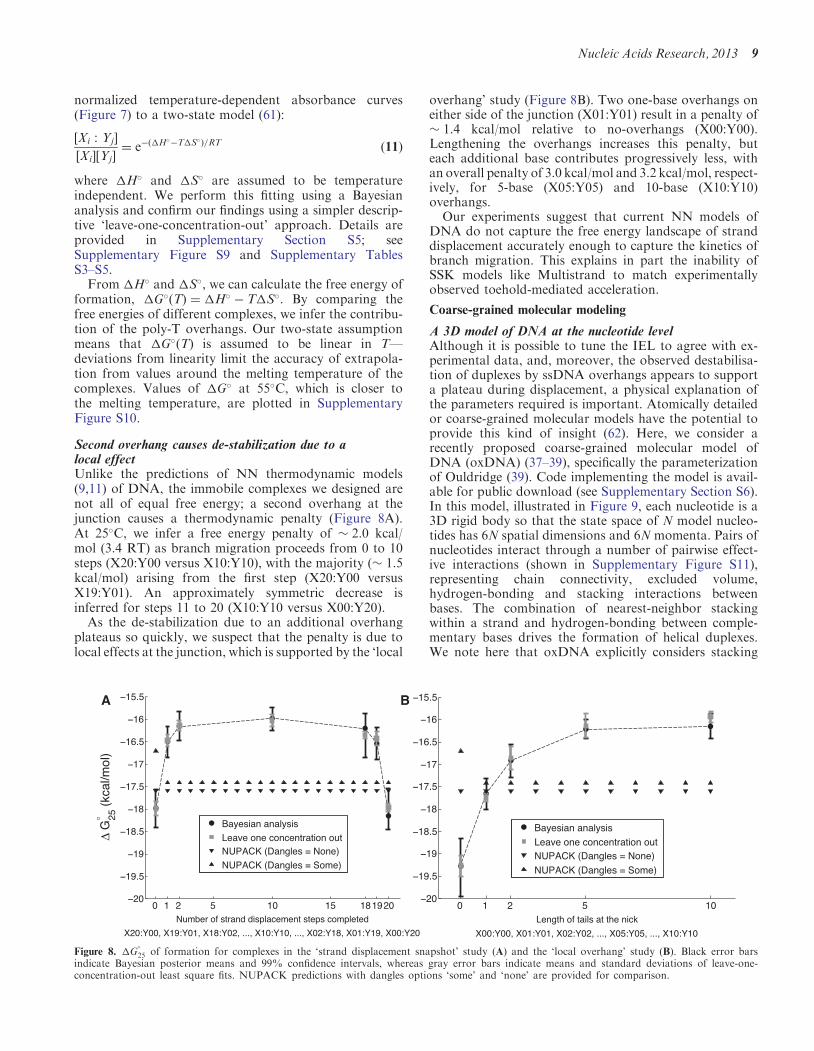
normalized temperature-dependent absorbance curves(Figure 7) to a two-state model (61)
frac12Xi Yj
frac12Xifrac12Yjfrac14 eethHTSTHORN=RT eth11THORN
where H and S are assumed to be temperatureindependent We perform this fitting using a Bayesiananalysis and confirm our findings using a simpler descrip-tive lsquoleave-one-concentration-outrsquo approach Details areprovided in Supplementary Section S5 seeSupplementary Figure S9 and Supplementary TablesS3ndashS5
From H and S we can calculate the free energy offormation GethTTHORN frac14 H TS By comparing thefree energies of different complexes we infer the contribu-tion of the poly-T overhangs Our two-state assumptionmeans that GethTTHORN is assumed to be linear in Tmdashdeviations from linearity limit the accuracy of extrapola-tion from values around the melting temperature of thecomplexes Values of G at 55C which is closer tothe melting temperature are plotted in SupplementaryFigure S10
Second overhang causes de-stabilization due to alocal effectUnlike the predictions of NN thermodynamic models(911) of DNA the immobile complexes we designed arenot all of equal free energy a second overhang at thejunction causes a thermodynamic penalty (Figure 8A)At 25C we infer a free energy penalty of 20 kcalmol (34 RT) as branch migration proceeds from 0 to 10steps (X20Y00 versus X10Y10) with the majority ( 15kcalmol) arising from the first step (X20Y00 versusX19Y01) An approximately symmetric decrease isinferred for steps 11 to 20 (X10Y10 versus X00Y20)
As the de-stabilization due to an additional overhangplateaus so quickly we suspect that the penalty is due tolocal effects at the junction which is supported by the lsquolocal
overhangrsquo study (Figure 8B) Two one-base overhangs oneither side of the junction (X01Y01) result in a penalty of 14 kcalmol relative to no-overhangs (X00Y00)Lengthening the overhangs increases this penalty buteach additional base contributes progressively less withan overall penalty of 30 kcalmol and 32 kcalmol respect-ively for 5-base (X05Y05) and 10-base (X10Y10)overhangsOur experiments suggest that current NN models of
DNA do not capture the free energy landscape of stranddisplacement accurately enough to capture the kinetics ofbranch migration This explains in part the inability ofSSK models like Multistrand to match experimentallyobserved toehold-mediated acceleration
Coarse-grained molecular modeling
A 3D model of DNA at the nucleotide levelAlthough it is possible to tune the IEL to agree with ex-perimental data and moreover the observed destabilisa-tion of duplexes by ssDNA overhangs appears to supporta plateau during displacement a physical explanation ofthe parameters required is important Atomically detailedor coarse-grained molecular models have the potential toprovide this kind of insight (62) Here we consider arecently proposed coarse-grained molecular model ofDNA (oxDNA) (37ndash39) specifically the parameterizationof Ouldridge (39) Code implementing the model is avail-able for public download (see Supplementary Section S6)In this model illustrated in Figure 9 each nucleotide is a3D rigid body so that the state space of N model nucleo-tides has 6N spatial dimensions and 6N momenta Pairs ofnucleotides interact through a number of pairwise effect-ive interactions (shown in Supplementary Figure S11)representing chain connectivity excluded volumehydrogen-bonding and stacking interactions betweenbases The combination of nearest-neighbor stackingwithin a strand and hydrogen-bonding between comple-mentary bases drives the formation of helical duplexesWe note here that oxDNA explicitly considers stacking
A B
Figure 8 G
25 of formation for complexes in the lsquostrand displacement snapshotrsquo study (A) and the lsquolocal overhangrsquo study (B) Black error barsindicate Bayesian posterior means and 99 confidence intervals whereas gray error bars indicate means and standard deviations of leave-one-concentration-out least square fits NUPACK predictions with dangles options lsquosomersquo and lsquononersquo are provided for comparison
Nucleic Acids Research 2013 9
interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

interactions between pairs of bases This is distinct fromthe base-pair stacks and coaxial stacks that appear intypical NN models which involve four bases (two fromeach strand) Multiple interactions in oxDNA then con-tribute to the effective strength of base-pair and coaxialstacking when the model is compared with NNdescriptionsoxDNA captures the thermodynamic and mechanical
changes associated with the formation of duplexes fromsingle strands under high salt conditions Specifically itquantitatively reproduces the oligomer length-dependenceof melting temperatures for the duplex transition thewidths of transitions the elastic moduli of duplexes andthe short persistence length of single strands oxDNAincludes the WatsonndashCrick rules of complementary basepairing but no further sequence dependence Duplex for-mation was therefore fitted to the behavior ofSantaLuciarsquos NN model (10) when averaged oversequence and we can only compare directly with theaverage-strength toehold data of Zhang and Winfree(30) oxDNA was fitted at 05M [Na+] where electrostaticinteractions are strongly screenedmdashthe repulsion of phos-phates is therefore incorporated into the backboneexcluded volume for simplicity The experiments ofZhang and Winfree (30) were performed in a buffer of125mM frac12Mg2+ which is known to result in similarduplex formation thermodynamics to high monovalentsalt buffers (10) It is plausible that kinetics of strand dis-placement might be significantly different in these twobuffersmdashthe migration of Holliday junctions forexample is known to vary between magnesium andsodium buffers (6364) Given that oxDNA was fittedonly to the duplex formation thermodynamics howeverthere is no reason to assume that its kinetics and the rep-resentation of transition intermediates are more suited toone buffer than the otherAlthough oxDNA contains many parameters none
were adjusted from those presented previously (39) forthe purposes of this particular study The parametervalues used have been seen to produce physically reason-able behavior for a range of systems Such systems includeDNA tweezers (39) a DNA walker (65) and the
interaction of kissing hairpins (66) and overstretching ofDNA under tension (67) Two of these systems thetweezers and the walker depend on strand displacementand so oxDNA has previously been shown to reproducethis phenomenon Furthermore a barrier to initiatingbranch migration was predicted when studying thetweezers (39) although it was not studied in great detail
Simulation techniquesWe provide a concise summary of our simulation tech-niques here for details see Supplementary Section S6We use two algorithms to simulate oxDNA the lsquoVirtualMoveMonte Carlorsquo (VMMC) algorithm ofWhitelam et al(6869) and the rigid-body Langevin Dynamics (LD)algorithm of Davidchack et al (70) The first approachrandomly attempts and accepts moves of clusters of nucleo-tides (illustrated in Supplementary Figure S12) in a mannerthat ensures the system samples from a Boltzmann distri-bution The second incorporates noise and damping termsinto Newtonrsquos equations in a self-consistent mannerthereby generating states drawn from the canonicalensemble The sampling of states is more efficient withVMMC and so it is used for all thermodynamicaverages Langevin algorithms however are explicitly dy-namical so they naturally give kinetic information that isharder to infer from VMMC Consequently all kineticresults quoted in this work use the LD algorithm
Processes like strand displacement are computationallydemanding to simulate and thus they require enhancedsampling techniques We use umbrella sampling (71) toimprove equilibration of thermodynamic averages inVMMC simulations This method involves applying anartificial bias to lower free-energy barriers therebyaccelerating the transitions between (meta)stable statesFor LD simulations of kinetics we use forward fluxsampling (7273) to obtain accurate estimates of therelative rates of strand displacement for different lengthsof toehold as shown in Supplementary Figure S13
oxDNArsquos energy mass and length scales imply a timescale Results in this section are quoted in terms of thistime scale for completeness As we are using a coarse-grained model with an approximate model of dynamicshowever the absolute times and rates reported should notbe overinterpreted We focus on relative rates whichshould be affected in a similar manner by the approxima-tions in oxDNA and the algorithms used to simulate it
Kinetic simulations of displacementWe consider a slightly truncated system (SupplementaryTable S6) based on that used by Zhang and Winfree (30)For computational simplicity we removed the majority ofthe tail of the incumbent strand which was used to interactwith a reporter Further to simplify the order parameterfor displacement we use an approach in which only theexpected (native) base pairs between the incumbent andthe substrate or the invading strand and the substrate aregiven a non-zero binding strength This simplification isreasonable because the sequences were designed toexhibit minimal secondary structure and cross-interactionswhen single-stranded Additional simulations of toeholdassociation were also performed to explore the
Figure 9 A 12-bp DNA duplex as represented by oxDNA Theenlarged section shows two rigid nucleotides highlighting thebackbone and base parts The planarity of bases is captured throughthe orientational dependence of interactions Image reproduced from[Ref 89]
10 Nucleic Acids Research 2013
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013
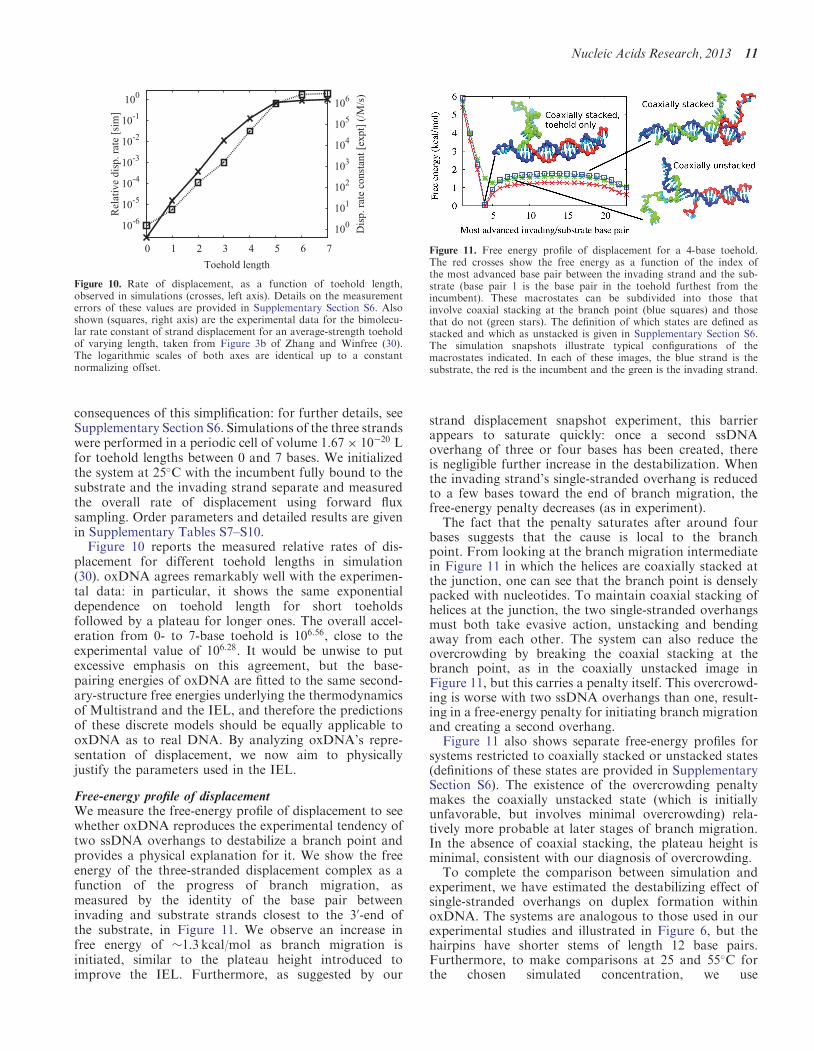
consequences of this simplification for further details seeSupplementary Section S6 Simulations of the three strandswere performed in a periodic cell of volume 167 1020 Lfor toehold lengths between 0 and 7 bases We initializedthe system at 25C with the incumbent fully bound to thesubstrate and the invading strand separate and measuredthe overall rate of displacement using forward fluxsampling Order parameters and detailed results are givenin Supplementary Tables S7ndashS10
Figure 10 reports the measured relative rates of dis-placement for different toehold lengths in simulation(30) oxDNA agrees remarkably well with the experimen-tal data in particular it shows the same exponentialdependence on toehold length for short toeholdsfollowed by a plateau for longer ones The overall accel-eration from 0- to 7-base toehold is 10656 close to theexperimental value of 10628 It would be unwise to putexcessive emphasis on this agreement but the base-pairing energies of oxDNA are fitted to the same second-ary-structure free energies underlying the thermodynamicsof Multistrand and the IEL and therefore the predictionsof these discrete models should be equally applicable tooxDNA as to real DNA By analyzing oxDNArsquos repre-sentation of displacement we now aim to physicallyjustify the parameters used in the IEL
Free-energy profile of displacementWe measure the free-energy profile of displacement to seewhether oxDNA reproduces the experimental tendency oftwo ssDNA overhangs to destabilize a branch point andprovides a physical explanation for it We show the freeenergy of the three-stranded displacement complex as afunction of the progress of branch migration asmeasured by the identity of the base pair betweeninvading and substrate strands closest to the 30-end ofthe substrate in Figure 11 We observe an increase infree energy of 13 kcalmol as branch migration isinitiated similar to the plateau height introduced toimprove the IEL Furthermore as suggested by our
strand displacement snapshot experiment this barrierappears to saturate quickly once a second ssDNAoverhang of three or four bases has been created thereis negligible further increase in the destabilization Whenthe invading strandrsquos single-stranded overhang is reducedto a few bases toward the end of branch migration thefree-energy penalty decreases (as in experiment)The fact that the penalty saturates after around four
bases suggests that the cause is local to the branchpoint From looking at the branch migration intermediatein Figure 11 in which the helices are coaxially stacked atthe junction one can see that the branch point is denselypacked with nucleotides To maintain coaxial stacking ofhelices at the junction the two single-stranded overhangsmust both take evasive action unstacking and bendingaway from each other The system can also reduce theovercrowding by breaking the coaxial stacking at thebranch point as in the coaxially unstacked image inFigure 11 but this carries a penalty itself This overcrowd-ing is worse with two ssDNA overhangs than one result-ing in a free-energy penalty for initiating branch migrationand creating a second overhangFigure 11 also shows separate free-energy profiles for
systems restricted to coaxially stacked or unstacked states(definitions of these states are provided in SupplementarySection S6) The existence of the overcrowding penaltymakes the coaxially unstacked state (which is initiallyunfavorable but involves minimal overcrowding) rela-tively more probable at later stages of branch migrationIn the absence of coaxial stacking the plateau height isminimal consistent with our diagnosis of overcrowdingTo complete the comparison between simulation and
experiment we have estimated the destabilizing effect ofsingle-stranded overhangs on duplex formation withinoxDNA The systems are analogous to those used in ourexperimental studies and illustrated in Figure 6 but thehairpins have shorter stems of length 12 base pairsFurthermore to make comparisons at 25 and 55C forthe chosen simulated concentration we use
Figure 11 Free energy profile of displacement for a 4-base toeholdThe red crosses show the free energy as a function of the index ofthe most advanced base pair between the invading strand and the sub-strate (base pair 1 is the base pair in the toehold furthest from theincumbent) These macrostates can be subdivided into those thatinvolve coaxial stacking at the branch point (blue squares) and thosethat do not (green stars) The definition of which states are defined asstacked and which as unstacked is given in Supplementary Section S6The simulation snapshots illustrate typical configurations of themacrostates indicated In each of these images the blue strand is thesubstrate the red is the incumbent and the green is the invading strand
Figure 10 Rate of displacement as a function of toehold lengthobserved in simulations (crosses left axis) Details on the measurementerrors of these values are provided in Supplementary Section S6 Alsoshown (squares right axis) are the experimental data for the bimolecu-lar rate constant of strand displacement for an average-strength toeholdof varying length taken from Figure 3b of Zhang and Winfree (30)The logarithmic scales of both axes are identical up to a constantnormalizing offset
Nucleic Acids Research 2013 11
complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

complementary lengths for interstrand binding of six andeight bases respectively (G can be inferred most accur-ately from experiments at 55C where the complexesmelt but we are most interested in the value at 25C)
The contribution of the single-stranded overhangs tothe free energy of association G is expected to beindependent of the length of the duplexes howeverallowing a direct comparison of this property to bemade The sequences used are given in SupplementaryTable S11 and further details are provided inSupplementary Section S6
The results of the hybridization simulations arereported in Table 2 The presence of two long ssDNAoverhangs is destabilizing by 13 and 075 kcalmol at25 and 55C respectively relative to the case with onesingle-stranded overhang We conclude that oxDNA hasa plateau height comparable with (but slightly smallerthan) that found in experiment with the caveat that wehave not measured sequence-dependent effects at thebranch point
Kinetics of branch migration and frayingHere we examine oxDNArsquos representation of the mech-anism of branch migration in detail Consistent with theliterature (282933) oxDNA predicts that branch migra-tion is slow compared to the rate at which base pairs in thetoehold fray and provides a mechanistic explanation forthis difference
Figure 12 Typical branch migration steps as represented by oxDNA for a toehold length of three bases In each case the nucleotide colored blackdisplaces the nucleotide colored gold (A) Displacement via invasion a base from the invading strand enters the incumbentsubstrate duplex andcompetes with a base from the incumbent for base pairing (i) Initial state with the gold-colored base on the incumbent bound to the substrate andthe black-colored base on the invading strand unbound (ii) Metastable intermediate with both the gold and black bases competing for base pairingwith the substrate (iii) Final state with the black base bound to the substrate and the gold base unbound (B) Displacement via sequential disruptionand formation of bonds a base pair in the incumbentsubstrate duplex breaks and the substrate base unstacks and moves across to the invadingsubstrate duplex (i) Coaxially unstacked initial state with the gold-colored base on the incumbent bound to the substrate and the black-colored baseon the invading strand unbound (ii) Metastable intermediate state in which the base pair involving the gold base has broken and the base from thesubstrate strand has unstacked (iii) Final state with the substrate base now bound to the black base in the invading strand Labeled distancesbetween hydrogen-bonding sites are used as coordinates to monitor branch migration steps in detail later in Figure 13
Table 2 Destabilizing Gmn due to dangling ssDNA sections as
found in simulations
Duplexlength
TC m n Gmn(kcalmol)
6 25 10 0 146 [12]0 10 138 [10]10 10 272 [32]
8 55 10 0 087 [060]0 10 094 [078]10 10 165 [179]
m is the length of the excess ssDNA attached to the hairpin and n thelength attached to the shorter strand Values for Gmn the free energyof formation of the duplex are extracted from simulations Thereported value Gmn is given by Gmn frac14 Gmn G00 Gmn isthen the destabilization of the duplex due to the ssDNA overhangsThe values in square brackets correspond to values inferred from ourexperimental studies for the closest equivalent lengths of danglingssDNA Note that G100 and G010 found here are compared toG020 and G200 from experiment ndash the local nature of the destabilisa-tion makes this reasonable
12 Nucleic Acids Research 2013
Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

Typical branch migration steps observed in oxDNA areshown in Figure 12 Two types of process occur mostcommonly
(1) Invading and incumbent duplexes remain coaxiallystacked at the branch point and a single-strandedbase from the invading strand enters the duplexregion competes for base-pairing to the substratewith a base from the incumbent and eventually dis-places it This process is illustrated in Figure 12AWe refer to this process as lsquobranch migration viainvasionrsquo
(2) Invading and incumbent duplexes coaxially unstackat the branch point and a base in the substrate istransferred from one duplex to the other In thisprocess one base pair must fully break before theother forms some time later An example of such astep is shown in Figure 12B We refer to this processas lsquobranch migration via sequential disruption andformation of bondsrsquo
Invasion is dominant at the start of branch migrationwhen the majority of systems are coaxially stackedBranch migration via sequential disruption and formationof bonds becomes more relevant at intermediate stageswhen coaxial stacking is less prevalent The two mechan-isms differ in whether the duplexes are coaxially stackedduring branch migration and whether the displacing baseinvades the duplex before disruption of the initial basepairing Invasion is geometrically infeasible from anunstacked state However the substrate strand couldremain stacked at the branch point whilst lsquoflipping outrsquoa base from the incumbent and only then replacing itwith a base from the invader This third type of processis rarer than the alternatives in oxDNA but cannot beruled out for real DNA Importantly all three processesrequire the disruption of favorable stacking interactionsbetween neighboring bases and considerable structural re-arrangement for each step of branch migration
We also note that the branch migration pathways haverelatively long-lived metastable intermediates (states rep-resented by Figure 12Aii and Bii These intermediatescan be clearly identified on free-energy landscapes for in-dividual steps of the branch migration process that areplotted in Figure 13A and B Typical trajectories ofbranch migration are shown in Figure 13C and DFigure 13C taken from the start of branch migrationclearly shows the system moving from one base-pairingconfiguration to another via a metastable intermediatein which both invading and incumbent bases are closeto the substrate base (branch migration via invasion)Figure 13D taken from a later stage of branch migrationshows a change in base pairing occurring via the diffusemetastable intermediate in which both invading andincumbent bases are distant from the substrate base(branch migration via sequential disruption and formationof bonds) We note that trajectories of both types arepossible at each stage but that invasion is particularlydominant for the first step of branch migration The factthat the intermediates are metastable despite their high
free energy indicates that they are not easily accessiblefrom the typical configurations of the systemBy contrast the fraying of a few base pairs in the
toehold and the subsequent detachment of the invadingstrand can occur relatively easily As illustrated inFigure 14AndashC it is not necessary to disrupt at least onestacking interaction for every base pair that is broken (asit is in branch migration) Further transition states infraying are much closer in configuration space to thetypical states of the system than they are for branch mi-gration making them easier to access This finding iscorroborated by Figure 14D which shows that the coord-inates of the system do not have to deviate far from theirtypical values before detachment occurs For comparisonwith Figure 13 Figure 15A shows a free-energy landscapeas a function of the separation of the base pairs in a twobase-pair toehold The minimum in the bottom left cor-responds to the fully bound state and the trajectoryshown in Figure 15B is the same as that in Figure 14illustrating both base pairs breaking in quick successionand the system leaving the bound state
A B
C D
Figure 13 (A) The free-energy landscape of the first branch migrationstep (for a three-base toehold) as a function of the separation ofhydrogen-bonding sites involved obtained from umbrella samplingsimulations The distances HB1 and HB2 between hydrogen-bondingsites of nucleotides are illustrated in Figure 12Ai In essence they aredistances of the competing bases in the invading (HB2) and incumbent(HB1) strands from their complement in the substrate In these figuresthe black rectangles and squares highlight the same regions in eachgraph roughly corresponding to the initial final and intermediatestates involved in branch migration Free energy landscapes aremeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shownin light green (B) An equivalent landscape to (A) but obtainedat a later stage (step 11) of migration when branch migration viasequential disruption and formation of bonds is more common(C) An example trajectory showing the first step of branch migrationoccurring via invasion (D) An example trajectory from the 11th stepshowing branch migration via sequential disruption and formation ofbonds
Nucleic Acids Research 2013 13
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013
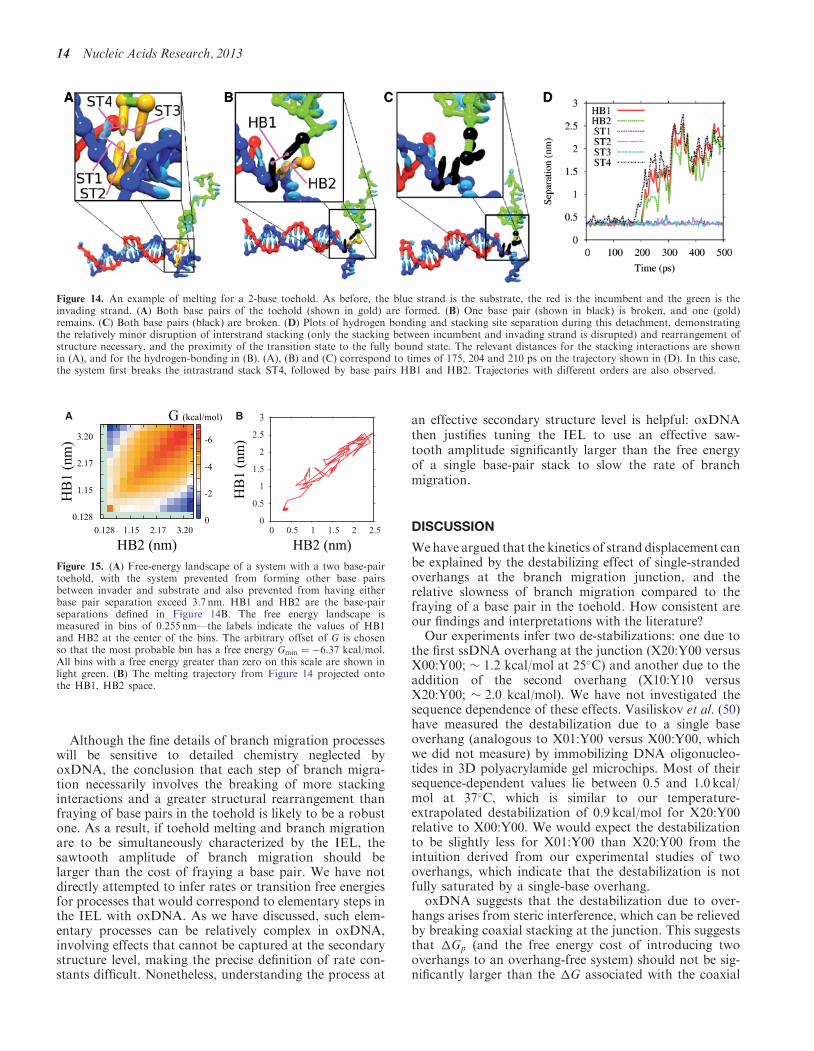
Although the fine details of branch migration processeswill be sensitive to detailed chemistry neglected byoxDNA the conclusion that each step of branch migra-tion necessarily involves the breaking of more stackinginteractions and a greater structural rearrangement thanfraying of base pairs in the toehold is likely to be a robustone As a result if toehold melting and branch migrationare to be simultaneously characterized by the IEL thesawtooth amplitude of branch migration should belarger than the cost of fraying a base pair We have notdirectly attempted to infer rates or transition free energiesfor processes that would correspond to elementary steps inthe IEL with oxDNA As we have discussed such elem-entary processes can be relatively complex in oxDNAinvolving effects that cannot be captured at the secondarystructure level making the precise definition of rate con-stants difficult Nonetheless understanding the process at
an effective secondary structure level is helpful oxDNAthen justifies tuning the IEL to use an effective saw-tooth amplitude significantly larger than the free energyof a single base-pair stack to slow the rate of branchmigration
DISCUSSION
Wehave argued that the kinetics of strand displacement canbe explained by the destabilizing effect of single-strandedoverhangs at the branch migration junction and therelative slowness of branch migration compared to thefraying of a base pair in the toehold How consistent areour findings and interpretations with the literature
Our experiments infer two de-stabilizations one due tothe first ssDNA overhang at the junction (X20Y00 versusX00Y00 12 kcalmol at 25C) and another due to theaddition of the second overhang (X10Y10 versusX20Y00 20 kcalmol) We have not investigated thesequence dependence of these effects Vasiliskov et al (50)have measured the destabilization due to a single baseoverhang (analogous to X01Y00 versus X00Y00 whichwe did not measure) by immobilizing DNA oligonucleo-tides in 3D polyacrylamide gel microchips Most of theirsequence-dependent values lie between 05 and 10 kcalmol at 37C which is similar to our temperature-extrapolated destabilization of 09 kcalmol for X20Y00relative to X00Y00 We would expect the destabilizationto be slightly less for X01Y00 than X20Y00 from theintuition derived from our experimental studies of twooverhangs which indicate that the destabilization is notfully saturated by a single-base overhang
oxDNA suggests that the destabilization due to over-hangs arises from steric interference which can be relievedby breaking coaxial stacking at the junction This suggeststhat Gp (and the free energy cost of introducing twooverhangs to an overhang-free system) should not be sig-nificantly larger than the G associated with the coaxial
Figure 14 An example of melting for a 2-base toehold As before the blue strand is the substrate the red is the incumbent and the green is theinvading strand (A) Both base pairs of the toehold (shown in gold) are formed (B) One base pair (shown in black) is broken and one (gold)remains (C) Both base pairs (black) are broken (D) Plots of hydrogen bonding and stacking site separation during this detachment demonstratingthe relatively minor disruption of interstrand stacking (only the stacking between incumbent and invading strand is disrupted) and rearrangement ofstructure necessary and the proximity of the transition state to the fully bound state The relevant distances for the stacking interactions are shownin (A) and for the hydrogen-bonding in (B) (A) (B) and (C) correspond to times of 175 204 and 210 ps on the trajectory shown in (D) In this casethe system first breaks the intrastrand stack ST4 followed by base pairs HB1 and HB2 Trajectories with different orders are also observed
A B
Figure 15 (A) Free-energy landscape of a system with a two base-pairtoehold with the system prevented from forming other base pairsbetween invader and substrate and also prevented from having eitherbase pair separation exceed 37 nm HB1 and HB2 are the base-pairseparations defined in Figure 14B The free energy landscape ismeasured in bins of 0255 nmmdashthe labels indicate the values of HB1and HB2 at the center of the bins The arbitrary offset of G is chosenso that the most probable bin has a free energy Gmin frac14 637 kcalmolAll bins with a free energy greater than zero on this scale are shown inlight green (B) The melting trajectory from Figure 14 projected ontothe HB1 HB2 space
14 Nucleic Acids Research 2013
stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

stacking transition in a nicked duplex Several attemptsto measure this quantity (485074) have inferred valuesof jGj 02 23 kcalmol at 37C and frac12Na+ frac14 1 to15mM Increasing frac12Na+ to 100mM was observed toincrease stability of coaxial stacks by 04 kcalmol Wetherefore conclude that our temperature-extrapolateddestabilization of 28 kcalmol between X10Y10 andX00Y00 at 37C (and in high salt conditions) is notinconsistent with these data
What do our findings imply for RNA Walter et al (75)have measured de-stabilization due to a single baseoverhang at a junction of duplexes as well as two singlebase overhangs on either side of a junction for RNA Theyreport a 28 kcalmol de-stabilization for their analog ofX01Y01 versus X00Y00 at 37C which is much higherthan our extrapolation of 14 kcalmol for DNA As thegeometry (A-form helix) and thermodynamics of RNA aresignificantly different from that of DNA we would notexpect analogous measurements for RNA to be quantita-tively similar For DNA we find that X20Y01 is signifi-cantly less stable than X01Y01 and if this holds true forRNA we would expect Gp for RNA to be significantlyhigher than the 20 kcalmol at 25C we infer for DNAHowever efn2 a NN thermodynamics model for RNAthat has been expanded to include experimentallymeasured terms for coaxial stacking and 1ndash and 2-nt over-hangs at junctions (7677) does not predict a free energypenalty similar to that which we infer for DNA stranddisplacement (see Supplementary Section S7 andSupplementary Figure S14) possibly due to incorporatingthese terms only in lsquointerior loopsrsquo
Is the IELrsquos inferred value of Gs consistent withmeasured branch migration rates Using the experimen-tally measured plateau height Gp frac14 20 kcalmol and thecorresponding inferred values of Gs frac14 53 kcalmol forthe IEL and Gs frac14 36 kcalmol for the AEL thesemodels predict branch migration step times of 103 and53 ms respectively These values are now more in linewith previous experimental measurements Raddinget al (28) report an average branch migration step timeof 12 ms at 37C in 10mM Na+ and Green and Tibbetts(29) estimate an upper limit of 20 ms at 65C in 03M Na+Our estimates are therefore somewhat larger although itwould be expected that the low temperature high salt con-ditions of Zhang and Winfree (30) which stabilize heliceswould reduce branch migration rates Indeed such aneffect is reported by Radding et al (28) Finally we notethat Zhang and Winfree (30) infer a branch migration steptime of 25ms from their phenomenological model12ndash24 times larger still than our estimates We attributethis difference to the absence in their model of a freeenergy penalty for initiating branch migration ie whatwe here call Gp
Are the IELrsquos values for kuni and Gbp compatible withmeasured fraying rates Wetmur and Davidson (33) haveinferred fraying rates from temperature-jump experimentsThey predict 004ms for fraying of a base pair at 25Cwhich is faster than our values (02 ms for the IEL 2 ms forAEL) and consistent with fraying being at least an orderof magnitude faster than branch migration steps We notehowever that these reported values were not directly
measured in experiments and required modeling toextract Furthermore related measurements of thekinetics of RNA base pair fluctuations found significantlyslower rates with fraying times on the order of microsec-onds at 25C (55ndash57) We therefore conclude that our par-ameterization is not inconsistent with well-establishedkinetic properties of nucleic acidsWe argue that the slowness of branch migration
initiation relative to fraying is a key aspect in understand-ing strand displacement In contrast Reynaldo et al(35) explained the low rate constant of zero-toeholddisplacement by positing that a certain number of basepairs n=3 or 4 are necessary to form a stable duplexand thus shorter duplexes can be ignored or assumed todetach instantly Our models differ by assigning favorableand increasingly strong thermodynamic energies fortoeholds attaching by 1 2 or more base pairs there isno penalty for short duplexes Instead we can reinterprettheir n as the value for which the rate of fraying n basepairs (kunie
njGbpj=RT) equals that of initiating branchmigration (kunie
Gs+p=RT) which for the IEL givesn frac14 43 A similar calculation for the AEL gives n frac14 38
CONCLUSIONS
We explain the dependence of strand displacement kineticson toehold length using two factors (i) the physicalprocess by which a single step of branch migrationoccurs is significantly slower than the fraying of a singlebase pair and (ii) initiating branch migration incurs athermodynamic penalty not captured by state-of-the-artnearest neighbor models of DNA due to the additionaloverhang it engenders at the junction The slowness ofbranch migration relative to fraying is captured in theIEL by a sawtooth amplitude Gs frac14 53 kcalmol whichis significantly larger than a single base-pair stack(jGbpj frac14 17 kcalmol) oxDNA provides physical justifi-cation for this by suggesting that branch migration stepsare slower than fraying as each stage necessarily involvesgreater structural rearrangement and disruption of favor-able stacking interactions Initiating branch migration isslower than the average branch migration step because ofthe free energy penalty (Gp) incurred from ourexperiments we infer Gp 20 kcalmol oxDNA repro-duces this penalty and suggests that it arises from stericinterference of the single strands and duplexes at thebranch migration junction Specifically the presence ofan extra single-stranded overhang after the first step ofbranch migration causes additional disruption of(coaxial) stacking and reduction of conformationalfreedom as the strands are forced to bend away fromeach otherOur work shows that toehold-mediated strand displace-
ment involves four distinct time scalesmdashrates of hybrid-ization fraying branch migration and branch migrationinitiationmdashcoupled with the energetics of base pairing inthe toehold The IEL model captures these rates via theparameters kbi kuni Gs Gp and Gbp Presuming thatthese rates and energies are the essential determinants ofstrand displacement kinetics the IEL analysis provides aframework for making kinetic predictions about different
Nucleic Acids Research 2013 15
strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

strand displacement-based systems For example the IELpredicts that for short toeholds keff scales as 1=b whereb is the branch migration domain length (seeSupplementary Section S2 and Supplementary FigureS15) However the dependence of kbi kuni Gs Gp
and Gbp on reaction conditions (eg salt concentrationsand temperature) are only partially understood impedingquantitative predictions for differing reaction conditionsNonetheless our current understanding does suggest thatdetermining just those five parameters should be sufficientto predict toehold-mediated strand displacement undernew conditionsWe expect that the IEL framework is also suitable for
describing other nucleic acids Toehold-mediated stranddisplacement has been demonstrated with RNA (7879)but the dependence of kinetics on toehold length has notbeen characterized Although several experimental studieshave reported hybridization and fraying rates for RNAthat are within the range observed for DNA (555780ndash82) branch migration and branch migration initiationrates are not well understood There is however someevidence that with 1-nt toeholds strand displacementrates for RNA are considerably slower than those forDNA (83) whereas the average base pairing energy isstronger (76) these observations are enough to predictthat a plot of log10 keffethhTHORN for RNA would have a lowery-intercept but higher initial slope than that for DNABroadly then we would expect the qualitative featuresof RNA strand displacement kinetics to be similar tothat of DNA quantitative understanding will requirefurther experimentsSimilarly the effect of structural variations of toehold-
mediated strand displacement such as associative andremote toeholds (8485) or initial mismatches in thebranch migration domain (86) could be understood interms of their effects on the fundamental rates andenergies These particular variants are likely to slowdown branch migration initiation and hence increaseA150 which could in turn permit greater design flexibilitywhile engineering nucleic acid devices and systemsAlthough the IEL provides a simple framework for
understanding strand displacement it does not predictor explain the underlying biophysical mechanisms norcan it easily be adapted to handle more complex structuralvariants or explain sequence-dependent behavior in largerstrand displacement cascades and dynamic DNA nano-technology (24) where the kinetics of both on- and off-pathway reactions (such as lsquoleakrsquo reactions analogous tozero-toehold strand displacement) are of great currentinterest oxDNA is well suited to studying the underlyingbiophysical mechanisms and behaviors of structuralvariants but it is computationally expensive for largersystemsEfficient quantitative simulation of sequence-dependent
kinetics for both desired and spurious strand displace-ment pathways would facilitate design and optimizationof large systems in silico SSK models based on NNthermodynamics such as Multistrand would be well-suited for such purposes but as found here currentimplementations require adjustment to match criticalthermodynamic and kinetic features Thermodynamically
accurate coaxial stacking terms and penalties for overhang-ing single-stranded sections of DNA at junctions should beincorporated into the model Incorporating the relativerates of fraying and branch migration within theMultistrand model faces several challenges Because everysecondary structure state has a well-defined free energybranch migration intermediates at the lsquotop of thesawtoothrsquo (frayed junctions with two tails) cannot simplybe pushed to higher energies as was done for the IEL usingGs Moreover Multistrand currently incorporates onlyone of the two possible branch migration mechanisms sug-gested by oxDNA (sequential disruption and formation ofbonds) Three independent approaches could conceivablybe pursued to better capture branch migration kineticsFirst an additional unimolecular rate constant parametercould be used for transitions local to the junction Secondnew transitions such as lsquoshiftrsquo moves in Kinfold (4387)could be incorporated to model alternative branch migra-tion pathways Third Multistrandrsquos state space could beaugmented possibly by including features such as non-base-pair stacking interactions
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [88ndash91]
ACKNOWLEDGEMENTS
The authors thank Zhen-Gang Wang Andrew JTurberfield David Yu Zhang Niles A Pierce Brian RWolfe David Soloveichik and Georg Seelig for helpfuldiscussions The authors thank the anonymous refereesfor feedback which significantly improved this work
FUNDING
National Science Foundation [CCF-0832824] theEngineering and Physical Sciences Research Council[EPI0013521] the Gordon and Betty MooreFoundation through the Caltech ProgrammableMolecular Technology Initiative the ScatcherdEuropean Trust and University College OxfordFunding for open access charge lsquoCaltech ProgrammableMolecular Technology Initiativersquo through the Gordonand Betty Moore Foundation
Conflict of interest statement None declared
REFERENCES
1 KallenbachNR MaRI and SeemanNC (1983) An immobilenucleic acid junction constructed from oligonucleotides Nature305 829ndash831
2 WinfreeE LiuFR WenzlerLA and SeemanNC (1998)Design and self-assembly of two-dimensional DNA crystalsNature 394 539
3 RothemundPWK (2006) Folding DNA to create nanoscaleshapes and patterns Nature 440 297ndash302
4 HeY YeT SuM ZhangC RibbeA JiangW and MaoC(2008) Hierarchical self-assembly of DNA into symmetricsupramolecular polyhedra Nature 452 198ndash201
16 Nucleic Acids Research 2013
5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

5 SeemanNC (2007) An overview of structural DNAnanotechnology Mol Biotech 37 246ndash257
6 PinheiroAV HanD ShihWM and YanH (2011) Challengesand opportunities for structural DNA nanotechnology NatNanotechnol 6 763ndash772
7 WeiB DaiM and YinP (2012) Complex shapes self-assembledfrom single-stranded DNA tiles Nature 485 623ndash627
8 SantaLuciaJ Jr AllawiHT and SeneviratnePA (1996)Improved nearest-neighbor parameters for predicting DNA duplexstability Biochemistry 35 3555ndash3562
9 SantaLuciaJ Jr (1998) A unified view of polymer dumbbell andoligonucleotide DNA nearest-neighbor thermodynamicsProc Natl Acad Sci USA 17 1460ndash1465
10 SantaLuciaJ Jr and HicksD (2004) The thermodynamics ofDNA structural motifs Annu Rev Biophys Biomol Struct 33415ndash440
11 DirksRM BoisJS SchaefferJM WinfreeE and PierceNA(2007) Thermodynamic analysis of interacting nucleic acidstrands SIAM Rev 49 65ndash88
12 HagermanPJ (1988) Flexibility of DNA Annu Rev BiophysBiophys Chem 17 265ndash286
13 StojanovicMN SemovaS KolpashchikovD MacdonaldJMorganC and StefanovicD (2005) Deoxyribozyme-based ligaselogic gates and their initial circuits J Am Chem Soc 1276914ndash6915
14 PenchovskyR and BreakerRR (2005) Computational designand experimental validation of oligonucleotide-sensing allostericribozymes Nat Biotechnol 23 1424ndash1433
15 SeeligG SoloveichikD ZhangDY and WinfreeE (2006)Enzyme-free nucleic acid logic circuits Science 314 1585ndash1588
16 QianL and WinfreeE (2011) Scaling up digital circuitcomputation with DNA strand displacement cascades Science332 1196ndash1201
17 ZhangDY TurberfieldAJ YurkeB and WinfreeE (2007)Engineering entropy-driven reactions and networks catlayzed byDNA Science 318 1121ndash1125
18 YinP ChoiHM CalvertCR and PierceNA (2008)Programming biomolecular self-assembly pathways Nature 451318ndash323
19 YurkeB TurberfieldAJ MillsAP SimmelFC andNeumannJ (2000) A DNA-fueled molecular machine made ofDNA Nature 406 605ndash608
20 DirksRM and PierceNA (2004) Triggered amplification byhybridization chain reaction Proc Natl Acad Sci USA 10115275ndash15278
21 PeiR TaylorSK StefanovicD RudchenkoS MitchellTEand StojanovicMN (2006) Behavior of polycatalytic assembliesin a substrate-displaying matrix J Am Chem Soc 12812693ndash12699
22 GreenSJ BathJ and TurberfieldAJ (2008) Coordinatedchemomechanical cycles a mechanism for autonomous molecularmotion Phys Rev Lett 101 238101
23 BathJ and TurberfieldAJ (2007) DNA nanomachinesNat Nanotechnol 2 275ndash284
24 ZhangDY and SeeligG (2011) Dynamic DNA nanotechnologyusing strand-displacement reactionsrsquo Nat Chem 3 103ndash113
25 KrishnanY and SimmelFC (2011) Nucleic acid basedmolecular devices Angew Chem Int Ed 50 3124ndash3156
26 SoloveichikD SeeligG and WinfreeE (2010) DNA as auniversal substrate for chemical kinetics Proc Natl Acad SciUSA 107 5393ndash5398
27 CardelliL (2013) Two domain DNA strand displacementMath Struct Comput Sci 23 247ndash271
28 RaddingCM BeattieKL HollomanWK and WiegandRC(1977) Uptake of homologous single-stranded fragments bysuperhelical DNA J Mol Biol 116 825ndash839
29 GreenC and TibbettsC (1981) Reassociation rate limiteddisplacement of DNA strands by branch migration Nucleic AcidsRes 9 1905ndash1918
30 ZhangD and WinfreeE (2009) Control of DNA stranddisplacement kinetics using toehold exchange J Am Chem Soc131 17303ndash17314
31 YurkeB and MillsA (2003) Using DNA to powernanostructures Genet Prog Evol Mach 4 111ndash122
32 MorrisonLE and StolsLM (1993) Sensitive fluorescence-basedthermodynamic and kinetic measurements of DNA hybridizationin solution Biochemistry 30 3095ndash3104
33 WetmurJG and DavidsonN (1968) Kinetics of renaturation ofDNA J Mol Biol 31 349ndash370
34 FellerW (1968) An Introduction to Probability Theory and itsApplications Vol 1 Wiley and Sons New York
35 ReynaldoLP VologodskiiAV NeriBP and LyamichevVI(2000) The kinetics of oligonucleotide replacements J Mol Biol297 511ndash520
36 SchaefferJM (2013) Stochastic simulation of the kinetics ofmultiple interacting nucleic acid strands PhD Thesis CaliforniaInstitute of Technology
37 OuldridgeTE LouisAA and DoyeJPK (2010) DNAnanotweezers studied with a coarse-grained model of DNA PhysRev Lett 104 178101
38 OuldridgeTE LouisAA and DoyeJPK (2011) Structuralmechanical and thermodynamic properties of a coarse-grainedDNA model J Chem Phys 134 085101
39 OuldridgeTE (2011) Coarse-grained modelling ofDNA and DNA nanotechnology PhD Thesis University ofOxford
40 MetropolisN RosenbluthAW RosenbluthMN TellerAHand TellerE (1953) Equation of state calculation by fastcomputing machines J Chem Phys 21 1087ndash1092
41 RudolphG (1999) The fundamental matrix of the generalrandom walk with absorbing boundaries Technical reportCollaborative Research Center lsquoComputational Intelligencersquo CI-75University of Dortmund
42 KawasakiK (1966) Diffusion constants near the critical point fortime-dependent Ising models Phys Rev 145 224ndash230
43 FlammC FontanaW HofackerIL and SchusterP (2000)RNA folding at elementary step resolution RNA 6 325ndash338
44 von AhsenN WittwerCT and SchutzE (2001) Oligonucleotidemelting temperatures under PCR conditions nearest-neighborcorrections for Mg2+ deoxynucleotide triphosphate and dimethylsulfoxide concentrations with comparison to alternative empiricalformulas Clin Chem 47 1956ndash1961
45 OwczarzyR MoreiraBG YouY BehlkeMA andWalderJA (2008) Predicting stability of DNA duplexes insolutions containing magnesium and monovalent cationsBiochemistry 47 5336ndash5353
46 BommaritoS PeyretN and SantaLuciaJ Jr (2000)Thermodynamic parameters for DNA sequences with danglingends Nuclei Acids Res 28 1929ndash1934
47 PeyretN (2000) Prediction of nucleic acid hybridizationparameters and algorithms PhD Thesis Wayne State University
48 ProtozanovaE YakovchukP and Frank-KamenetskiiMD(2004) Stacked-unstacked equilibrium at the nick site of DNAJ Mol Biol 342 775ndash785
49 PyshnyiDV and IvanovaEM (2002) Thermodynamicparameters of coaxial stacking on stacking hybridizationof oligodeoxyribonucleotides Russ Chem B 51 1145ndash1155
50 VasiliskovVA ProkopenkoDV and MirzabekovAD (2001)Parallel multiplex thermodynamic analysis of coaxial basestacking in DNA duplexes by oligodeoxyribonucleotidemicrochips Nucleic Acids Res 29 2303ndash2313
51 ZadehJN SteenbergCD BoisJS WolfeBR PierceMBKhanAR DirksRM and PierceNA (2011) NUPACKanalysis and design of nucleic acid systems J Comput Chem32 170ndash173
52 HofackerIL (2003) Vienna RNA secondary structure serverNuclei Acids Res 31 3429ndash3431
53 ZukerM (2003) Mfold web server for nucleic acid folding andhybridization prediction Nuclei Acids Res 31 3406ndash3415
54 GillespieDT (1977) Exact stochastic simulation of coupledchemical reactions J Phys Chem 81 2340ndash2361
55 CraigME CrothersDM and DotyP (1971) Relaxationkinetics of dimer formation by self-complementaryoligonucleotides J Mol Biol 62 383ndash401
56 PorschkeD UhlenbeckOC and MartinFH (1973)Thermodynamics and kinetics of the helix-coil transitionof oligomers containing GC base pairs Biopolymers 121313ndash1335
Nucleic Acids Research 2013 17
57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013

57 PorschkeD (1974) A direct measurement of the unzipperingrate of a nucleic acid double helix Biophys Chem 2 97ndash101
58 KuhnH DemidovVV CoullJM FiandacaMJ GildeaBDand Frank-KamenetskiiMD (2002) Hybridization of DNA andPNA molecular beacons to single-stranded and double-strandedDNA targets J Am Chem Soc 124 1097ndash1103
59 TsourkasA BehlkeMA RoseSD and BaoG (2003)Hybridization kinetics and thermodynamics of molecular beaconsNuclei Acids Res 31 1319ndash1330
60 GaoY WolfLK and GeorgiadisRM (2006) Secondarystructure effects on DNA hybridization kinetics a solution versussurface comparison Nuclei Acids Res 34 3370ndash3377
61 AlbergoDD MarkyLA BreslauerKJ and TurnerDH(1981) Thermodynamics of (dG-dC)3 double-helix formation inwater and deuterium oxide Biochemistry 20 1409ndash1413
62 ElberR and KirmizialtinS (2013) Molecular machinesCurr Opin Struct Biol 23 206ndash211
63 PanyutinIG and HsiehP (1994) The kinetics of spontaneousDNA branch migration Proc Natl Acad Sci USA 912021ndash2025
64 JooC McKinneySA LilleyDM and HaT (2004) Exploringrare conformational species and ionic effects in DNA HollidayJunctions using single-molecule spectroscopy J Mol Biol 342739ndash751
65 OuldridgeTE HoareRL LouisAA DoyeJPK BathJ andTurberfieldAJ (2013) Optimizing DNA nanotechnology throughcoarse-grained modelling a two-footed DNA walker ACS Nano26 2479ndash2490
66 RomanoF HudsonA DoyeJPK OuldridgeTE andLouisAA (2012) The effect of topology on the structure andfree energy landscape of DNA kissing complexes J Chem Phys136 215102
67 RomanoF ChakrabortyD DoyeJPK OuldridgeTE andLouisAA (2013) Coarse-grained simulations of DNAoverstretching J Chem Phys 138 085101
68 WhitelamS and GeisslerPL (2007) Avoiding unphysical kinetictraps in Monte Carlo simulations of strongly attractive particlesJ Chem Phys 127 154101
69 WhitelamS FengEH HaganMF and GeisslerPL (2009)The role of collective motion in examples of coarsening andself-assembly Soft Matter 5 1251ndash1262
70 DavidchackRL HandelR and TretyakovMV (2009) Langevinthermostat for rigid body dynamics J Chem Phys 130 234101
71 TorrieGM and ValleauJP (1977) Nonphysical samplingdistributions in Monte Carlo free-energy estimation umbrellasampling J Comp Phys 23 187ndash199
72 AllenRJ WarrenPB and ten WoldePR (2005) Sampling rareswitching events in biochemical networks Phys Rev Lett 94018104
73 AllenRJ ValerianiC and ten WoldePR (2009) Forward fluxsampling for rare event simulations J Phys Condens Matter 21463102
74 YakovchukP ProtozanovaE and Frank-KamenetskiiMD(2006) Base-stacking and base-pairing contributions into thermalstability of the DNA double helix Nucleic Acids Res 34 564ndash574
75 WalterAE TurnerDH KimJ LyttleMH MullerPMathewsDH and ZukerM (1994) Coaxial stacking of helixesenhances binding of oligoribonucleotides and improves predictionsof RNA folding Proc Natl Acad Sci USA 91 9218ndash9222
76 MathewsDH SabinaJ ZukerM and TurnerDH (1999)Expanded sequence dependence of thermodynamic parametersimproves prediction of RNA secondary structure J Mol Biol288 911ndash940
77 MathewsDH DisneyMD ChildsJL SchroederSJZukerM and TurnerDH (2004) Incorporating chemicalmodification constraints into a dynamic programming algorithmfor prediction of RNA secondary structure Proc Natl Acad SciUSA 101 7287ndash7292
78 ChoiHMT ChangJY TrinhLA PadillaJE FraserSEand PierceNA (2010) Programmable in situ amplification formultiplexed imaging of mRNA expression Nat Biotechnol 281208ndash1212
79 CalluraJM DwyerDJ IssacsFJ CantorCR and CollinsJJ(2010) Tracking tuning and terminating microbial physiologyusing synthetic riboregulators Proc Natl Acad Sci USA 10715898ndash15903
80 WetmurJG (1976) Hybridization and renaturation kinetics ofnucleic acids Annu Rev Biophys Bioeng 5 337ndash361
81 PorschkeD (1977) Elementary steps of base recognition andhelix-coil transitions in nucleic acids Mol Biol BiochemBiophys 24 191ndash218
82 LimaWF (1992) Implication of RNA structure on antisenseoligonucleotide hybridization kinetics Biochemistry 31 12055
83 HomannM NedbalW and SczakielG (1996) Dissociation oflong-chain duplex RNA can occur via strand displacementin vitro biological implications Nucleic Acids Res 244395ndash4400
84 ChenX (2011) Expanding the rule set of DNA circuitry withassociative toehold activation J Am Chem Soc 134 263ndash271
85 GenotAJ ZhangDY BathJ and TurberfieldAJ (2011)Remote toehold a mechanism for flexible control of DNAhybridization kinetics J Am Chem Soc 133 2177ndash2182
86 ZhangDY and WinfreeE (2010) Robustness and modularityproperties of a non-covalent DNA catalytic reaction NucleicAcids Res 38 4182ndash4197
87 PorschkeD (1974) Model calculations on the kinetics ofoligonucleotide double helix coil transitions Evidence for a fastchain sliding reaction Biophys Chem 2 83ndash96
88 PuglisiJD and TinocoIT Jr (1989) Absorbance melting curvesof RNA Methods Enzymol 180 304ndash325
89 GelmanA CarlinJB SternHS and RubinDB (2004)Bayesian Data Analysis CRC Press Boca Raton Florida
90 OuldridgeTE SulcP RomanoF DoyeJPK and LouisAA(2013) DNA hybridization kinetics zippering internaldisplacement and sequence dependence Nucleic Acids Res (doi101093nargkt687 epub ahead of print August 8 2013)
91 FerrenbergAM and SwendsenRH (1988) New Monte Carlotechnique for studying phase transitions Phys Rev Lett 612635ndash2638
18 Nucleic Acids Research 2013


















