
Olfaction in Aging and Alzheimer's Disease : Event-related Potentials to a Cross-modal...
11
INTERNATIONAL SYMPOSIUM ON OLFACTION AND TASTE Olfaction in Aging and Alzheimer’s Disease Event-related Potentials to a Cross-modal Odor-Recognition Memory Task Discriminate ApoE ε4 + and ApoE ε4 − Individuals Claire Murphy a,b,c Ethan S. Solomon, a Lori Haase, a,c MiRan Wang, a and Charlie D. Morgan a a San Diego State University, b University of California, San Diego, and the c SDSU/UCSD Joint Doctoral Program in Clinical Psychology, San Diego, California, USA Alzheimer’s disease (AD) is a devastating neurodegenerative condition that affects more than 5 million Americans. Currently, a definitive and unequivocal diagnosis of AD can only be confirmed histopathogically via postmortem autopsy, demonstrating the need for objective measures of cognitive functioning for those at risk for AD. The single most important genetic risk factor of AD is the apolipoprotein E (ApoE) ε4 allele. The present study investigated olfactory and cognitive processing deficits in ApoE ε4 + indi- viduals using a cross-modal recognition memory task and an objective electrophysio- logical measure, the event-related potential (ERP). Ten ε4 + individuals (5 M, 5 F, mean [M] = 75.1 years) and 10 age- and gender-matched ε4 − individuals (5 M, 5 F, M = 71 years) sequentially encoded a set of 16 olfactory stimuli and were subsequently shown names of odors previously presented (targets) or not (foils). EEG activity was recorded from 19 electrodes as participants distinguished targets from foils using a two-button mouse. P3 latencies were significantly longer in ε4 + individuals, and intraclass correlations demonstrated differential activity between the two groups. These findings are consis- tent with a compensatory hypothesis, which posits that nondemented ε4 + individuals will expend greater effort in cognitive processing or engage in alternative strategies and therefore require greater activation of neural tissue or recruitment of different neu- ral populations. The findings also suggest that cross-modal ERP studies of recognition memory discriminate early neurocognitive changes in ApoE ε4 + and ApoE ε4 − indi- viduals and may contribute to identifying the phenotype of persons who will develop Alzheimer’s disease. Key words: aging; Alzheimer’s disease; ApoE ε4; olfaction; olfactory impairment; smell; recognition memory Introduction Alzheimer’s disease (AD) is a devastating neurodegenerative condition that affects more than 5 million Americans. Since 1980, the prevalence of AD in America has nearly dou- bled making it the most common form of de- Address for correspondence: Claire Murphy, Ph.D., SDSU/UCSD Joint Doctoral Program, 6363 Alvarado Ct., Suite 101, San Diego, CA 92120-4913. Voice: +619 594 4559; fax: +619 594 3773. [email protected] mentia, accounting for 50–60% of dementia- related cases. Current estimates suggest that by 2050, more than 13 million Americans will suf- fer from this debilitating disorder. 1 Diagnostic Criteria Both the National Institute of Neurologi- cal and Communicative Disorders and Stroke- Alzheimer’s Disease and Related Disorders As- sociation (NINDS–ADRDA) and the DSM-IV diagnostic criteria for AD include progressive International Symposium on Olfaction and Taste: Ann. N.Y. Acad. Sci. 1170: 647–657 (2009). doi: 10.1111/j.1749-6632.2009.04486.x c 2009 New York Academy of Sciences. 647
-
Upload
claire-murphy -
Category
Documents
-
view
213 -
download
0
Transcript of Olfaction in Aging and Alzheimer's Disease : Event-related Potentials to a Cross-modal...

INTERNATIONAL SYMPOSIUM ON OLFACTION AND TASTE
Olfaction in Aging and Alzheimer’s Disease
Event-related Potentials to a Cross-modalOdor-Recognition Memory Task Discriminate ApoE
ε4+ and ApoE ε4− Individuals
Claire Murphya,b,c Ethan S. Solomon,a Lori Haase,a,c
MiRan Wang,a and Charlie D. Morgana
aSan Diego State University, bUniversity of California, San Diego, and the cSDSU/UCSDJoint Doctoral Program in Clinical Psychology, San Diego, California, USA
Alzheimer’s disease (AD) is a devastating neurodegenerative condition that affects morethan 5 million Americans. Currently, a definitive and unequivocal diagnosis of AD canonly be confirmed histopathogically via postmortem autopsy, demonstrating the needfor objective measures of cognitive functioning for those at risk for AD. The singlemost important genetic risk factor of AD is the apolipoprotein E (ApoE) ε4 allele. Thepresent study investigated olfactory and cognitive processing deficits in ApoE ε4+ indi-viduals using a cross-modal recognition memory task and an objective electrophysio-logical measure, the event-related potential (ERP). Ten ε4+ individuals (5 M, 5 F, mean[M] = 75.1 years) and 10 age- and gender-matched ε4− individuals (5 M, 5 F, M = 71 years)sequentially encoded a set of 16 olfactory stimuli and were subsequently shown namesof odors previously presented (targets) or not (foils). EEG activity was recorded from 19electrodes as participants distinguished targets from foils using a two-button mouse.P3 latencies were significantly longer in ε4+ individuals, and intraclass correlationsdemonstrated differential activity between the two groups. These findings are consis-tent with a compensatory hypothesis, which posits that nondemented ε4+ individualswill expend greater effort in cognitive processing or engage in alternative strategiesand therefore require greater activation of neural tissue or recruitment of different neu-ral populations. The findings also suggest that cross-modal ERP studies of recognitionmemory discriminate early neurocognitive changes in ApoE ε4+ and ApoE ε4− indi-viduals and may contribute to identifying the phenotype of persons who will developAlzheimer’s disease.
Key words: aging; Alzheimer’s disease; ApoE ε4; olfaction; olfactory impairment; smell;recognition memory
Introduction
Alzheimer’s disease (AD) is a devastatingneurodegenerative condition that affects morethan 5 million Americans. Since 1980, theprevalence of AD in America has nearly dou-bled making it the most common form of de-
Address for correspondence: Claire Murphy, Ph.D., SDSU/UCSDJoint Doctoral Program, 6363 Alvarado Ct., Suite 101, San Diego,CA 92120-4913. Voice: +619 594 4559; fax: +619 594 [email protected]
mentia, accounting for 50–60% of dementia-related cases. Current estimates suggest that by2050, more than 13 million Americans will suf-fer from this debilitating disorder.1
Diagnostic Criteria
Both the National Institute of Neurologi-cal and Communicative Disorders and Stroke-Alzheimer’s Disease and Related Disorders As-sociation (NINDS–ADRDA) and the DSM-IVdiagnostic criteria for AD include progressive
International Symposium on Olfaction and Taste: Ann. N.Y. Acad. Sci. 1170: 647–657 (2009).doi: 10.1111/j.1749-6632.2009.04486.x c© 2009 New York Academy of Sciences.
647

648 Annals of the New York Academy of Sciences
and global cognitive impairment in two ormore cognitive areas including memory. Phys-iological tests for diagnosing the disorder in aliving person are imperfect. Therefore, a defini-tive and unequivocal diagnosis of AD can onlybe confirmed histopathogically via postmortemautopsy. As pharmacological and other inter-ventions become available, distinguishing veryearly those individuals who will go on to de-velop AD will be very important for targetedtreatment; hence, efforts to identify these indi-viduals are keen. The current study was de-signed to investigate a new method for tar-geting these individuals using evoked brainpotentials.
Risk Factors
A number of risk factors for AD have beenidentified, with the most obvious being age,1,2 adecreased reserve capacity of the brain result-ing from reduced brain size, low educationaland occupational attainment, low mental abil-ity in early life, and reduced mental and phys-ical activity during later life,3,4 head trauma,5
and Down syndrome.6,7
The single most important genetic risk factoris the allelic variant apolipoprotein E (ApoE)ε4.8 The ApoE ε4 allele increases the risk ofdeveloping AD by three times in heterozygotesand by 15 times in homozygotes.9,10
ApoE ε4
ApoE is a key lipoprotein found in the brainas well as the cerebral spinal fluid (CSF).11,12
It has been mapped in the human genome tothe proximal long arm of chromosome 19.13,14
ApoE normally functions as a cholesterol trans-porter in the brain,12 but it also abnormallybinds to soluble and insoluble forms of B-amyloid15 and the βA4 peptide, which is theprimary component of neuritic plaques.16 Asshown by Poirier,17 the ApoE ε4 allele is lessefficient than other ApoE alleles in the reuseof membrane lipids and neuronal repair. ApoE
has also been found to contribute to the forma-tion of neurofibrillary tangles (NFTs).11
ApoE mRNA, which plays a compensatoryrole in neuronal sprouting and synaptoge-nesis in the central nervous system, is re-duced in the hippocampus in AD.15 Tau,the main component of NFTs, is elevatedin the CSF of AD patients who possess atleast one copy of the ε4 allele, comparedwith those without the ε4 allele.18 Soininenand Riekkinen19 showed a positive associationbetween the number of ε4 alleles and the in-creased deficit of acetylcholine-containing neu-rons in the brains of AD patients.
Prior research into global cognitive function-ing and ApoE allele genotype has been incon-clusive. A number of studies have shown thatthe ε4 allele predicted cognitive decline,20–26
and ε4+ persons perform poorly on tests ofprocessing speed, delayed free recall, and se-mantic long-term memory.27,56 However, per-formance is not impaired on all tests,28–30 andsome have reported no difference in cognitivedecline between the different allelic variants ofApoE.31,32
Olfaction and AD
AD produces neuropathological changes inperipheral and central areas of the brain thatprocess olfactory information. The first stagein the neurodegenerative progression of ADoccurs in the entorhinal and transentorhinalareas critical to olfactory information process-ing.33,34 The olfactory bulbs, anterior olfactorynucleus, and orbitofrontal cortex are all alsoaffected by AD.34,35 This pathology motivatesthe investigation of olfactory functioning in in-dividuals in the beginning stages of AD. Theolfactory deficits observed in AD include de-creases in odor threshold sensitivity,36–38 odoridentification,39–42 odor memory,43,44 odor flu-ency,45 and olfactory event-related potentials(ERPs).46
Deficits in olfactory functioning have alsobeen associated with those at risk for AD.Wilson46a discusses elegantly the olfactory

Murphy et al.: Olfaction in Aging and Alzheimer’s Disease 649
impairment in persons with mild cognitive im-pairment, an important risk factor for AD.He and his group have also shown a signif-icant relationship between olfactory impair-ment and the neuroanatomical bases of AD.Olfactory deficits are clearly associated withthe ApoE ε4 allele. Physiologically, a post-mortem study showed ApoE was localized inan increased number of olfactory receptor neu-rons, becoming more pervasive in the olfac-tory epithelium of individuals with AD com-pared to non-AD individuals.47 Behaviorally,Bacon and colleagues36 demonstrated that inthe first year of their AD diagnosis, ε4+ in-dividuals had inferior odor threshold sensitiv-ity. Murphy and colleagues42 showed that evenin nondemented elderly individuals, those withthe ε4 allele showed impaired odor identifica-tion, and Calhoun-Haney and Murphy48 foundsignificant decline in odor identification in ε4+
individuals who did not yet show significantdeclines in Dementia Rating Scale (DRS), sug-gesting that changes in olfactory function maypredict later overall decline.
Memory
Neuroimaging and electrophysiologicalstudies have revealed several different brainregions involved in encoding and retrieval ofepisodic memory. These studies demonstratedthat neural activity is observed in differentbrain regions depending on a number ofconditions, including depth of encoding,49
sensory modality,50 and task set.51
Recognition Memory and ApoE
A number of studies have shown significantdeficits in odor-recognition memory and odoridentification in ε4+ individuals.52–54 They alsocommit more false-positive errors in response toolfactory stimuli than ε4− individuals.52 A false-positive error is defined as erroneously choos-ing a new stimulus (foil) as a member of theoriginal stimulus set (target). Analysis of false-positive errors and poor performance on odor-
recognition memory tasks may be useful waysto distinguish between normal older individualsand those at risk for developing AD.
ERP Components
Early components of the ERP waveform,N1/P2, represent exogenous sensory mech-anisms that have been associated with psy-chophysical measures of olfactory functioning,such as odor threshold sensitivity.57,58 Latercomponents of the ERP waveform, especiallyP3, represent endogenous mechanisms, such asstimulus classification speed as well as cogni-tive attending and evaluation of stimuli.59,60
P3 latency is correlated with performance onneuropsychological tests that measure mem-ory and cognitive processing speed.61 P3 am-plitude is proportional to the amount of neu-ral resources allocated to a given task.62,63 P3latency increases and P3 amplitude decreasesas individuals age.57,58,64,65 Wetter and Mur-phy66 also demonstrated that ApoE ε4+ indi-viduals performing an oddball task show ab-normally increased P3 latency and decreasedP3 amplitude. These findings suggest the po-tential of electroencephalographic (EEG) mea-sures for assessing cognitive functioning andmore specifically recognition memory in ε4+
individuals.However, to date, an ERP recognition mem-
ory study using a cross-modal odor-recognitiontask (presenting olfactory stimuli during encod-ing and visually presenting names of those sameodors during recognition) has never been em-ployed to investigate differences in cortical ac-tivation in ApoE ε4+ and ε4− individuals. Thiswas our aim.
Methods
Participants were 20 nondemented olderadults, 10 of whom were ε4+ (5 M, 5 F,mean [M] = 75.1 years, SD = 8.3) and 10of whom were ε4−, age and gender matched(M = 71 years, SD = 6.1). All gave informedconsent.
650 Annals of the New York Academy of Sciences
Psychophysical and NeuropsychologicalMeasures
Psychophysical measures included the SanDiego Odor Identification Test (SDOIT),70
measures of verbal fluency including letter andcategory fluency, and tests of odor fluency.45
The odor threshold test37 screened for poten-tial anosmia or severe hyposmia.
Neuropsychological measures included theCalifornia Odor Learning Test (COLT),71 theDRS,72 and the Mini-Mental State Exam(MMSE).73 These measures assess participants’global cognitive functioning, cognitive impair-ment, and odor learning, recall, and recogni-tion memory abilities.
Electrophysiological Data Collection
EEG activity was recorded with an elec-trode cap from 19 electrode sites, Fp1/2, Fz,F3/4, F7/8, Cz, C3/4, T7/8, Pz, P3/4, P7/8,and O1/2, for 2000 ms, including a 100-msprestimulus base line, amplified via a GrassTechnologies (Warwick, RI) RPS 197 amplifiersystem, filtered (0.1–30 Hz bandpass, 6 dB oc-tave/slope), and digitized at 256 Hz. Data wereaveraged offline using inhouse software to rejectartifactual responses of ± 150 μV and a re-gression analysis–based electro-ocular (EOG)correction procedure to remove any remain-ing artifact.74 Neuroscan software was used tocalculate intraclass correlations.
Procedure
The experimental paradigm used in thisstudy is identical to the paradigm used in a par-allel functional MRI (fMRI) study investigatingcross-modal recognition memory for odors,69
and is based on a design of Stark and Squire.50
Stimuli were presented in three blocks, eachwith 72 trials. During encoding, participantswere presented with 16 familiar odors from theCOLT.71 Each was presented for 5 s, with a15-s interstimulus interval.
During EEG recording, participants werepresented every 4 s with words displayed ona computer screen that corresponded to namesof odors previously presented (targets) or not(foils), followed by a fixation cross. Participantsdistinguished targets from foils using a two-button mouse.
Hypotheses
The purpose of the proposed study was to ex-amine whether ApoE ε4+ and ε4− individualsexhibit differences in the neural allocation of re-sources during a cross-modal recognition mem-ory task. We analyzed P3 latency and intra-class correlations of activity at each electrodesite. We hypothesized differences in ERP dataas a function of allele status, longer P3 laten-cies, and differences in activity in ApoE ε4+
individuals, dependent on response type.
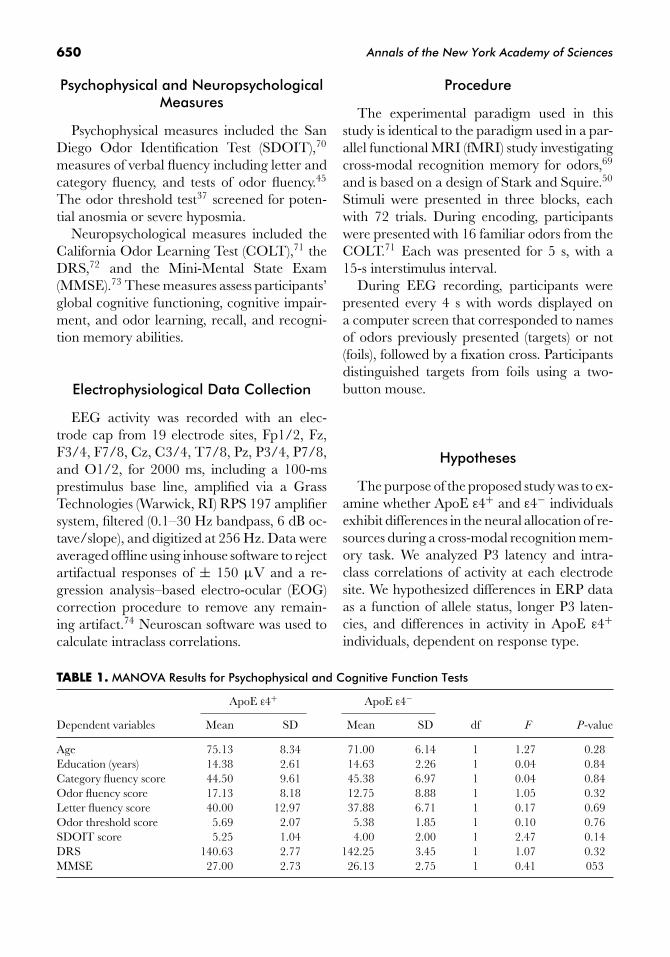
TABLE 1. MANOVA Results for Psychophysical and Cognitive Function Tests
ApoE ε4+ ApoE ε4−
Dependent variables Mean SD Mean SD df F P-value
Age 75.13 8.34 71.00 6.14 1 1.27 0.28Education (years) 14.38 2.61 14.63 2.26 1 0.04 0.84Category fluency score 44.50 9.61 45.38 6.97 1 0.04 0.84Odor fluency score 17.13 8.18 12.75 8.88 1 1.05 0.32Letter fluency score 40.00 12.97 37.88 6.71 1 0.17 0.69Odor threshold score 5.69 2.07 5.38 1.85 1 0.10 0.76SDOIT score 5.25 1.04 4.00 2.00 1 2.47 0.14DRS 140.63 2.77 142.25 3.45 1 1.07 0.32MMSE 27.00 2.73 26.13 2.75 1 0.41 053

Murphy et al.: Olfaction in Aging and Alzheimer’s Disease 651
Figure 1. P3 latency for ApoE ε4+ and ε4− participants.
Results
Psychophysical and CognitiveFunctioning Tests
Multivariate analyses of variance(MANOVA) on age, years of education,category fluency, odor fluency, letter fluency,mean odor threshold, SDOIT, DRS, andMMSE scores demonstrated no significant
differences between groups (Table 1). In par-ticular, analyses demonstrated no significantdifference in odor threshold scores betweenApoE ε4+ and ε4− participants. This findingsuggests that ε4+ and ε4− participants had sim-ilar sensory functioning and, therefore, similarpotential to encode the olfactory stimuli. Thisresult was not surprising since participantswere screened to exclude participants withanosmia or severe hyposmia.
TABLE 2. ANOVA Results Comparing P3 Latency for ApoE ε4+ and ApoE ε4− Participants
P3 Latency
Effect df F P-value ηp2
Allele status (AS) 1 9.59 0.01 0.35Site (S) 2 17.63 0.00 0.50Response (R) 3 1.18 0.32 0.06(R)(AS) 3 0.53 0.62 0.03(S)(R) 6 0.88 0.46 0.05(S)(R)(AS) 6 0.57 0.64 0.03

652 Annals of the New York Academy of Sciences
TABLE 3. Intraclass Correlations at the 19 Electrode Sites, Comparing Brain Response in ε4+ andε4− Individuals during the Four Memory Response Types: Correct Rejections, False Positives, Hits, andMisses, Respectively
Electrode Correct rejections: False positives: Hits: Misses:site correlation (r) correlation (r) correlation (r) correlation (r)
Fz 0.95 0.93 0.61 0.18∗
Cz 0.94 0.25∗ 0.89 0.26∗
Pz 0.89 0.62 0.92 0.34∗
F3 0.28∗ 0.54 0.17∗ 0.50F4 0.29∗ 0.42∗ 0.30∗ 0.64F7 0.00∗ 0.17∗ 0.01∗ 0.08∗
F8 0.07∗ 0.13∗ 0.01∗ 0.32∗
C3 0.58 0.50 0.65 0.41∗
C4 0.35∗ 0.82 0.63 0.87T7 0.29∗ 0.30∗ 0.18∗ 0.56T8 0.13∗ 0.29∗ 0.18∗ 0.19∗
P3 0.73 0.66 0.75 0.80P4 0.68 0.96 0.91 0.96P7 0.60 0.71 0.72 0.98P8 0.78 0.67 0.60 0.76Fp1 0.10∗ 0.73 0.12∗ 0.03∗
Fp2 0.01∗ 0.06∗ 0.47 0.11∗
O1 0.74 0.51 0.95 0.68O2 0.80 0.71 0.95 0.75
Correlations that indicate significantly different responses are indicated by ∗.
P3 Latency
Olfactory ERP studies have focused almostexclusive on the Fz, Cz, and Pz electrode sitesand thus, to place the data in the contextof the literature, we first conducted ANOVAon midline electrode sites (Table 2). ANOVAof P3 latency showed that latency was signi-ficantly delayed in ApoE ε4+ individuals(F [1,18] = 9.6, P < 0.05, eta2 = 0.35)(Fig. 1).
Intraclass Correlations
To investigate differences in activity betweengroups, intraclass correlations of the positivearea measured from 525–675 ms post stimu-lus from ApoE ε4+ and ε4− participants wereexamined at each electrode site and for eachresponse type. The 525–675 ms poststimulusresponse epoch was used to capture the epochduring which the P3s for the participants weregenerated (see Fig. 1). The intraclass correla-
tion expresses the degree of overlap and re-lated variability between two waveforms. Thestatistic is generated by comparing two ERPwaveforms, taking into account both wave-shape and absolute voltage values. Correlationvalues below 0.44 (P < 0.05) indicated stati-cally significant differences in amplitude (seeTable 3). As illustrated in Table 3, therewere significant differences in activity betweengroups. Intraclass correlations for each of theresponse types are given in Figure 2, withblue indicating the largest differences betweengroups and red indicating greatest similaritybetween groups. The topographical maps inFigure 3 illustrate the group differences inP3 amplitude, with warmer colors indicatinggreater amplitude.
Discussion
This study was designed to investigatewhether EEG correlates of cross-modal

Murphy et al.: Olfaction in Aging and Alzheimer’s Disease 653
Figure 2. Intraclass correlations for electrode sites for the four response types for ε4+ and ε4− individuals.Blue indicates the greatest differences between groups, and red the highest correlations between groups.
Figure 3. P3 amplitude for the four response types for ε4+ and ε4− individuals. Warmer colors indicategreater amplitude.
odor-recognition memory discriminate be-tween ε4+ and ε4− individuals. The results sup-port significant differences between ε4+ andε4− individuals in P3 latency and in intraclasscorrelations of activity.
Psychophysical and CognitiveFunctioning Tests
No significant differences were found inpsychophysical and cognitive functioning test

654 Annals of the New York Academy of Sciences
scores between ε4+ and ε4− participants, sug-gesting that participants had similar olfactoryfunctioning supporting similar sensory encod-ing of the olfactory stimuli. Participants withanosmia or serious hyposmia were screened outusing the odor threshold test, thus this result isnot surprising.
P3 Latency and Intraclass Correlations
P3 Latency
The original hypothesis that ε4+ participantswould demonstrate longer P3 latencies thanε4− participants was supported. This is the firstreport of longer latencies in a cross-modal odor-recognition memory task. Latency differenceshave been observed in ERP studies using odd-ball tasks66,75; however, this is the first observa-tion that a cross-modal odor-recognition mem-ory task discriminates between ε4+ and ε4−
individuals. Longer P3 latencies in ε4+ indi-viduals are consistent with the theory that ε4+
individuals demonstrate slower cognitive pro-cessing than normal older adults and are at riskfor subsequently developing AD.
Amplitude and Intraclass Correlations
Figure 3 illustrates the differences in P3 am-plitude between ε4+ and ε4− participants at thedifferent electrode sites.
Intraclass correlations were employed in or-der to compare waveshape and voltage valuesat each individual electrode site between ε4+
and ε4− participants. These correlations weregenerated for each response type using a 525–675 ms poststimulus response epoch to capturethe epoch during which the P3s for the partic-ipants were generated (see Fig. 1). For correctrejection responses, ε4+ and ε4− participantsshowed significant differences in intraclass cor-relations in electrode sites over frontal (F3, F4,F7, F8, Fp1, Fp2) and temporal (T7, T8) areas.This effect was also observed when evaluatinghit responses (F3, F4, F7, F8, T7, T8, Fp1).The ε4+ and ε4− individuals showed signifi-cantly different intraclass correlations at a num-
ber of electrode sites for false-positive responses(Cz, F4, F7, F8, T7, T8, Fp2) and misses (Fz,Cz, Pz, F7, F8, C3, T8, Fp1, Fp2), suggestingthat differential activity underlies these perfor-mance errors (see Table 3 and Fig. 2). Thesefindings are consistent with the compensatoryhypothesis that ε4+ individuals expend greatereffort in cognitive processing in a cross-modalodor-recognition memory task than ε4− indi-viduals. This increased effort observed in ε4+
individuals requires activation of a greater arealextent of neural tissue, or an increased firingrate within a given area, or a larger num-ber of neurons recruited to perform the task.This pattern of correlations is also compatiblewith the hypothesis that ε4+ individuals mayengage in different strategies in order to per-form the task. Preliminary data from our labo-ratory using fMRI are also consistent with thesehypotheses (Murphy, Haase, and Wang,manuscript in preparation).
These findings are complemented byanatomical studies that have demonstrated thatε4+ individuals experience greater temporalatrophy than ε4− individuals.77,78 Finally, in-traclass correlations lend further support toneuropsychological studies that suggest false-positive responses were able to discriminate be-tween ε4+ and ε4− individuals.52,79
Conclusion
ApoE ε4+ individuals demonstrated signif-icantly longer P3 latency than ε4− individu-als, and differential activity for all responsetypes. Differential activity in ApoE ε4+ andε4− individuals, demonstrated by the intra-class correlation coefficients, is consistent witha compensatory hypothesis, which posits thatε4+ individuals expend greater effort in cog-nitive processing and therefore require greateractivation of neural tissue during retrieval at-tempts. Finally, in accordance with previousERP and fMRI studies involving odor familiar-ity, participants exhibited greater positive areain electrode sites over the right hemisphere in

Murphy et al.: Olfaction in Aging and Alzheimer’s Disease 655
comparison with the left hemisphere. Thesefindings suggest that cross-modal ERP stud-ies of recognition memory in ApoE ε4+ andε4− individuals are a useful measure for in-dexing functional brain integrity, for under-standing the neurocognitive changes associatedwith the ApoE ε4 allele, and for discriminat-ing between brain response in ε4+ and ε4−
individuals.
Acknowledgments
We thank Drs. Paul Gilbert, John Polich,and Wendy Smith and Mr. Brian Lopez forexpertise and assistance. Supported by the Na-tional Institute of Health Grants DC02064 andAG04085 to C. M.
Conflicts of Interest
The authors declare no conflicts of interest.
References
1. Hebert, L.E. et al. 2003. Alzheimer disease in theUS population: prevalence estimates using the 2000census. Arch. Neurol. 60: 1119–1122.
2. Farrer, L.A. et al. 1995. Apolipoprotein E genotypein patients with Alzheimer’s disease: implications forthe risk of dementia among relatives. Ann. Neurol. 38:797–808.
3. Mayeux, R. 2003. Epidemiology of neurodegenera-tion. Annu. Rev. Neurosci. 26: 81–104.
4. Mortimer, J.A., D.A. Snowdon & W.R. Markesbery.2003. Head circumference, education and risk of de-mentia: findings from the Nun Study. J. Clin. Exp.
Neuropsychol. 25: 671–679.5. Jellinger, K.A. 2004. Head injury and dementia. Curr.
Opin. Neurol. 17: 719–723.6. Motte, J. & R.S. Williams. 1989. Age-related changes
in the density and morphology of plaques and neu-rofibrillary tangles in Down syndrome brain. Acta
Neuropathol. 77: 535–546.7. Wisniewski, K.E., H.M. Wisniewski & G.Y. Wen.
1985. Occurrence of neuropathological changes anddementia of Alzheimer’s disease in Down’s syndrome.Ann. Neurol. 17: 278–282.
8. Raber, J., Y. Huang & J.W. Ashford. 2004. ApoEgenotype accounts for the vast majority of AD
risk and AD pathology. Neurobiol. Aging 25: 641–650.
9. Farrer, L.A. et al. 1997. Effects of age, sex, and eth-nicity on the association between apolipoprotein Egenotype and Alzheimer disease. A meta-analysis.APOE and Alzheimer Disease Meta Analysis Con-sortium. JAMA 278: 1349–1356.
10. Corder, E.H. et al. 1995. The apolipoprotein E E4 al-lele and sex-specific risk of Alzheimer’s disease. JAMA
273: 373–374.11. Namba, Y. et al. 1991. Apolipoprotein E immunore-
activity in cerebral amyloid deposits and neurofibril-lary tangles in Alzheimer’s disease and kuru plaqueamyloid in Creutzfeldt-Jakob disease. Brain Res. 541:163–166.
12. Saunders, A.M. et al. 1993. Apolipoprotein E epsilon4 allele distributions in late-onset alzheimer’s diseaseand in other amyloid-forming diseases. Lancet 342:710–711.
13. Das, H.K. et al. 1985. Isolation, characterization,and mapping to chromosome 19 of the humanapolipoprotein E gene. J. Biol. Chem. 260: 6240–6247.
14. Olaisen, B., P. Teisberg & T. Gedde-Dahl, Jr. 1982.The locus for apolipoprotein E (apoE) is linked to thecomplement component C3 (C3) locus on chromo-some 19 in man. Hum. Genet. 62: 233–236.
15. Poirier, J. et al. 1993. Apolipoprotein E polymor-phism and Alzheimer’s disease. Lancet 342: 697–699.
16. Strittmatter, W.J. et al. 1993. Apolipoprotein E:high-avidity binding to beta-amyloid and increasedfrequency of type 4 allele in late-onset familialAlzheimer disease. Proc. Natl. Acad. Sci. USA 90: 1977–1981.
17. Poirier, J. 1994. Apolipoprotein E in animal mod-els of CNS injury and in Alzheimer’s disease. Trends
Neurosci. 17: 525–530.18. Tapiola, T. et al. 1998. CSF tau is related to
apolipoprotein E genotype in early Alzheimer’s dis-ease. Neurology 50: 169–174.
19. Soininen, H.S. & P.J. Riekkinen, Sr. 1996. Apolipo-protein E, memory and Alzheimer’s disease. Trends
Neurosci. 19: 224–228.20. Feskens, E.J. et al. 1994. Apolipoprotein e4 allele and
cognitive decline in elderly men. BMJ 309: 1202–1206.
21. Haan, M.N. et al. 1999. The role of APOE epsilon4 inmodulating effects of other risk factors for cognitivedecline in elderly persons. JAMA 282: 40–46.
22. Hyman, B.T. et al. 1996. Apolipoprotein E and cog-nitive change in an elderly population. Ann. Neurol.
40: 55–66.23. Jonker, C. et al. 1998. Association between apolipo-
protein E epsilon4 and the rate of cognitive decline

656 Annals of the New York Academy of Sciences
in community-dwelling elderly individuals with andwithout dementia. Arch. Neurol. 55: 1065–1069.
24. Yaffe, K. et al. 1997. Apolipoprotein E phenotypeand cognitive decline in a prospective study of elderlycommunity women. Arch. Neurol. 54: 1110–1114.
25. Riley, K.P. et al. 2000. Cognitive function andapolipoprotein E in very old adults: findings fromthe Nun Study. J. Gerontol. B Psychol. Sci. Soc. Sci. 55:S69–S75.
26. Carmelli, D. et al. 2000. The joint effect ofapolipoprotein E epsilon4 and MRI findings onlower-extremity function and decline in cognitivefunction. J. Gerontol. A Biol. Sci. Med. Sci. 55: M103–M109.
27. Staehelin, H.B. et al. 1999. Apolipoprotein E geno-types and cognitive functions in healthy elderly per-sons. Acta Neurol. Scand. 100: 53–60.
28. Henderson, A.S. et al. 1995. Apolipoprotein E alleleepsilon 4, dementia, and cognitive decline in a pop-ulation sample. Lancet 346: 1387–1390.
29. O’Hara, R. et al. 1998. The APOE epsilon4 alleleis associated with decline on delayed recall perfor-mance in community-dwelling older adults. J. Am.
Geriatr. Soc. 46: 1493–1498.30. Small, B.J., H. Basun & L. Backman. 1998. Three-
year changes in cognitive performance as a functionof apolipoprotein E genotype: evidence from veryold adults without dementia. Psychol. Aging 13: 80–87.
31. Brayne, C. et al. 1996. Apolipoprotein E genotype inthe prediction of cognitive decline and dementia in aprospectively studied elderly population. Dementia 7:169–174.
32. Helkala, E.L. et al. 1996. Memory functions in humansubjects with different apolipoprotein E phenotypesduring a 3-year population-based follow-up study.Neurosci. Lett. 204: 177–180.
33. Braak, H. & E. Braak. 1997. Staging of Alzheimer-related cortical destruction. Int. Psychogeriatr. 9(Suppl1): 257–261; discussion 269–272.
34. Price, J.L. et al. 1991. The distribution of tangles,plaques and related immunohistochemical markersin healthy aging and Alzheimer’s disease. Neurobiol.
Aging 12: 295–312.35. Esiri, M.M. & G.K. Wilcock. 1984. The olfactory
bulbs in Alzheimer’s disease. J. Neurol. Neurosurg. Psy-
chiatry 47: 56–60.36. Bacon, A.W. et al. 1998. Very early changes in olfac-
tory functioning due to Alzheimer’s disease and therole of apolipoprotein E in olfaction. Ann. N. Y. Acad.
Sci. 855: 723–731.37. Murphy, C. et al. 1990. Olfactory thresholds are as-
sociated with degree of dementia in Alzheimer’s dis-ease. Neurobiol. Aging 11: 465–469.
38. Nordin, S. & C. Murphy. 1996. Impaired sen-
sory and cognitive olfactory function in questionableAlzheimer’s disease. Neuropsych. 10: 112–119.
39. Serby M. 1986. Olfaction and Alzheimer’s disease.Prog. Neuropsychopharmacol. Biol. Psychiatry 10: 579–586.
40. Doty R.L. et al. 1991. Odor identification deficitof the parkinsonism-dementia complex of Guam:equivalence to that of Alzheimer’s and idiopathicParkinson’s disease. Neurology 5(Suppl 2): 77–80; dis-cussion 80–81.
41. Morgan, C.D., S. Nordin & C. Murphy. 1995. Odoridentification as an early marker for Alzheimer’s dis-ease: impact of lexical functioning and detection sen-sitivity. J. Clin. Exp. Neuropsychol. 17: 793–803.
42. Murphy, C., A. Bacon, M.W. Bondi & D.P. Salmon.1998. Apolipoprotein E status is associated with odoridentification deficits in nondemented older persons.Ann. N. Y. Acad. Sci. 855: 744–750.
43. Murphy, C., S. Nordin & S. Jinich. 1999. Veryearly decline in recognition memory for odors inAlzheimer’s disease. Aging Neuropsychol. Cogn. 6: 229–240.
44. Schiffman S.S., B.G. Graham, E.A. Sattely-Miller,et al. 2002. Taste, smell and neuropsychologi-cal performance of individuals at familial risk forAlzheimer’s disease. Neurobiol. Aging 23: 397–404.
45. Bacon, A.W., J.S. Paulsen & C. Murphy. 1999. A testof odor fluency in patients with Alzheimer’s diseaseand Huntington’s chorea. J. Clin. Exp. Neuropsychol.
21: 341–351.46. Morgan, C.D. & C. Murphy. 2002. Olfactory event-
related potentials in Alzheimer’s disease. J. Int. Neu-
ropsychol. Soc. 8: 753–763.46a. Wilson, R.S., S.E. Arnold, J.A. Schneider, et al.
2009. Olfactory impairment in presymptomaticAlzheimer’s disease. Ann. N. Y. Acad. Sci. InternationalSymposium on Olfaction and Taste. 1170: 730–735.
47. Yamagishi, M. et al. 1998. Increased density ofolfactory receptor neurons immunoreactive forapolipoprotein E in patients with Alzheimer’s dis-ease. Ann. Otol. Rhinol. Laryngol. 107: 421–426.
48. Calhoun-Haney, R. & C. Murphy. 2005. Apolipo-protein epsilon4 is associated with more rapid de-cline in odor identification than in odor threshold orDementia Rating Scale scores. Brain Cogn. 58: 178–182.
49. Craik, F.I.M. & R.S. Lockhart. 1972. Levels of pro-cessing: a framework for memory research. J. Verb.
Learn. Verb. Behav. 11: 671–684.50. Stark, C.E. & L.R. Squire. 2000. Functional mag-
netic resonance imaging (fMRI) activity in the hip-pocampal region during recognition memory. J. Neu-
rosci. 20: 7776–7781.51. Morcom, A.M. & M.D. Rugg. 2002. Getting ready
to remember: the neural correlates of task set duringrecognition memory. Neuroreport 13: 149–152.

Murphy et al.: Olfaction in Aging and Alzheimer’s Disease 657
52. Gilbert, P.E. & C. Murphy. 2004. The effect of theApoE epsilon4 allele on recognition memory for ol-factory and visual stimuli in patients with patho-logically confirmed Alzheimer’s disease, probableAlzheimer’s disease, and healthy elderly controls.J. Clin. Exp. Neuropsychol. 26: 779–794.
53. Nordin, S. & C. Murphy. 1998. Odor memory innormal aging and Alzheimer’s disease. Ann. N. Y. Acad.
Sci. 855: 686–693.54. Sundermann, E.E., P.E. Gilbert & C. Murphy. 2007.
Apolipoprotein E epsilon4 genotype and gender: ef-fects on memory. Am. J. Geriatr. Psychiatry 15: 869–878.
55. Bondi, M.W. et al. 1995. Episodic memory changesare associated with the APOE-epsilon 4 allele in non-demented older adults. Neurology 45: 2203–2206.
56. Morgan, C.D. et al. 1997. Olfactory event-related po-tentials: older males demonstrate the greatest deficits.Electroencephalogr. Clin. Neurophysiol. 104: 351–358.
57. Murphy, C. et al. 1994. Olfactory-evoked potentials:assessment of young and elderly, and comparison topsychophysical threshold. Chem. Senses 19: 47–56.
58. Donchin, E. & M.G. Coles. 1988. Is the P300 com-ponent a manifestation of context updating? Behav.
Brain Sci. 11: 357–427.59. Polich, J. 1998. P300 clinical utility and control of
variability. J. Clin. Neurophysiol. 15: 14–33.60. Geisler, M.W. et al. 1999. Neuropsychological per-
formance and cognitive olfactory event-related brainpotentials in young and elderly adults. J. Clin. Exp.
Neuropsychol. 21: 108–126.61. Kramer, A.F. & D.L. Strayer. 1988. Assessing the de-
velopment of automatic processing: an application ofdual-task and event-related brain potential method-ologies. Biol. Psychol. 26: 231–267.
62. Wickens, C. et al. 1983. Performance of concur-rent tasks: a psychophysiological analysis of the reci-procity of information-processing resources. Science
221: 1080–1082.63. Morgan, C.D. et al. 1999. Olfactory P3 in young and
older adults. Psychophysiology 36: 281–287.
64. Murphy, C. et al. 2000. Olfactory event-related po-tentials and aging: normative data. Int. J. Psychophysiol.
36: 133–145.65. Wetter, S. & C. Murphy. 2001. Apolipoprotein E
epsilon4 positive individuals demonstrate delayed ol-factory event-related potentials. Neurobiol. Aging 22:439–447.
66. Cerf-Ducastel, B. & C. Murphy. 2006. Neural sub-strates of cross-modal olfactory recognition memory:an fMRI study. NeuroImage 31: 386–396.
67. Murphy, C. et al. 2002. Prevalence of olfactory im-pairment in older adults. JAMA 288: 2307–2312.
68. Murphy, C., S. Nordin & L. Acosta. 1997.Odor learning, recall, and recognition memory inyoung and elderly adults. Neuropsychology 11: 126–137.
69. Mattis, S. 1988. Dementia Rating Scale: Professional man-
ual. Psychological Assessment Resources. Odessa,FL.
70. Folstein, M.F., S.E. Folstein & P.R. McHugh. 1975.“Mini-mental state”. A practical method for grad-ing the cognitive state of patients for the clinician.J. Psychiatr. Res. 12: 189–198.
71. Semlitsch, H.V. et al. 1986. A solution for reliableand valid reduction of ocular artifacts, applied to theP300 ERP. Psychophysiology 23: 695–703.
72. Green, J. & A.I. Levey. 1999. Event-related potentialchanges in groups at increased risk for Alzheimerdisease. Arch. Neurol. 56: 1398–1403.
73. Geroldi, C. et al. 1999. APOE-epsilon4 is associatedwith less frontal and more medial temporal lobe at-rophy in AD. Neurology 53: 1825–1832.
74. Rosso, S.M. et al. 2002. Apolipoprotein E4 in the tem-poral variant of frontotemporal dementia. J. Neurol.
Neurosurg. Psychiatry 72: 820.75. Gilbert, P.E. & C. Murphy. 2004. Differences be-
tween recognition memory and remote memory forolfactory and visual stimuli in nondemented elderlyindividuals genetically at risk for Alzheimer’s disease.Exp. Gerontol. 39: 433–441.


















