
Nutrition ^ v ; and digestive physiology in monogastric ...
102
Nutrition ^ v ; 0 and digestive physiology in monogastric farm animals Reviews presented at the symposium on the occasion of the retirement of Dr Ir E.J. van Weerden, Wageningen, Netherlands, 26 May 1989 E.J. van Weerden & J. Huisman (Editors) Pudoc Wageningen 1989 \ T>Y\ !-"' i~*
Transcript of Nutrition ^ v ; and digestive physiology in monogastric ...

Nutrition ^ v ; 0
and digestive physiology in monogastric farm animals
Reviews presented at the symposium on the occasion of the retirement of
Dr Ir E.J. van Weerden, Wageningen, Netherlands, 26 May 1989
E.J. van Weerden & J. Huisman (Editors)
Pudoc Wageningen 1989
\ T> Y\ !-"' i~*

CIP data, Royal Netherlands Library, The Hague
Nutrition and digestive physiology in monogastric farm animals. Reviews presented at the symposium on the occasion of the retirement of Dr Ir E.J. van Weerden, Wageningen, Netherlands, 26 May 1989. / E.J. van Weerden &J. Huisman (Editors). - Wageningen: Pudoc. - x+IOI pp. ISBN 90-220-1011-2 UDC 636.2/.5.084:59l.l32 NUGI 835 Subject heading: animal nutrition
© Centre for Agricultural Publishing & Documentation (Pudoc), Wageningen, Netherlands, 1989
All rights reserved. Nothing from this publication may be reproduced, stored in a computerized system or published in any form or in any manner, including electronic, mechanical, reprographic or photographic, without prior written permission from the publisher, Pudoc, P.O. Box 4, 6700 AA Wageningen, Netherlands.
The individual contributions in this publication and any liabilities arising from them remain the responsibility of the authors.
Insofar as photocopies from this publication are permitted by the Copyright Act 1912, Article I6B and Royal Netherlands Decree of 20 June 1974 (Staatsblad 351) as amended in Royal Netherlands Decree of 23 August 1985 (Staatsblad 471 ) and by Copyright Act 1912, Article 17, the legally defined copyright fee for any copies should be transferred to the Stichting Reprorecht (P.O. Box 882, 1180 AW Amstelveen, Netherlands). For reproduction of parts of this publication in compilations such as anthologies or readers (Copyright Act 1912, Article 16), permission must be obtained from the publisher.
Printed in the Netherlands.
BIBLIOTHEEK1
LANDBOUWUNIVERSITEIT ' ''.GENINGEN

^ É É f •FF1? V-."äjaft».ATt
S i . - . -r i « . t J
Dr Ir E.J. van Weerden

PREFACE
Dr Ir E.J. van Weerden together with Dr Ir P. van der Wal, founded the institute now known as ILOB. Dr van Weerden started work there in 1960 and was responsible for nutritional and physiological research until 1984 when ILOB became part of the TNO organization. Dr van der Wal then left ILOB. Upon the departure of Dr van der Wal, Dr van Weerden became also responsible for the general management of ILOB.
TNO is an organization for applied scientific research and ILOB is an institute within TNO responsible for animal nutrition and physiology. It is partly subsidised by the government, mainly through the Ministry of Economic Affairs, but the major part of the work is concerned with applied contract research. This is often confidential, implying in many cases that the results can only be published with some delay; in some cases, no publications are allowed at all.
Research at ILOB is concerned with various topics, such as: efficacy and safety of raw materials, antibiotics, probiotics, new growth-promoting agents such as steroid hormones, /2-agonists and somatropin; digestive physiology and nutrition of monogastric farm animals, pharmacokinetics and biotransformation of additives and veterinary drugs. Dr. van Weerden had the ultimate responsibility for this research. Topics in which he was especially involved were amino acid requirement in poultry, pigs and veal calves, digestive physiology in pigs and veal calves, efficacy and safety of single-cell protein, new growth-promoting agents, antibiotics, probiotics, replacement of skimmed milk powder by other protein sources. /
Dr van Weerden is an excellent scientist, internationally recognized as a specialist in the fields mentioned who, in the period after 1984, also proved himself to be a good manager. Always sensitive to both sense and nonsense in research, he did much of the preliminary work on any new techniques or fields of research himself to see which points may be critical. He was invited to present his work at many scientific congresses and had considerable influence on many new developments in the fields of digestive physiology and nutrition of farm animals, advising various industries on the development of new products of application in the feed industry.
A symposium was organised in honour of Dr van Weerden to acknowledge all the effort he had put into the research carried out at ILOB for twenty-nine years and into management for the last five years. Two specialists from abroad were invited to present papers on digestive physiology in pigs and veal calves. Two of his co-workers contributed papers on developments in amino acids and antinutritional factors. A paper on the effects of somatropin in pigs, resulting from the research that has been done for several years in cooperation with the Agricultural University, was also present. The symposium was closed by Dr van Weerden with the presentation of his paper: Past and Future Developments of Protein Supply in Monogastric Farm Animals.
In this book, the papers presented are published in an extended form as reviews. These contributions are just a small expression of the appreciation felt for Dr van Weerden by his colleagues and co-workers from ILOB.
The organisers wish to thank Mr Haak, Mrs van den Berg-Volders and Mrs Haak-van den Brink for their excellent assistance in the organization.
J. Huisman, J.B. Schutte, S. Bakker and G.J.M, van Kempen (Organizing Committee)
Wageningen, Netherlands, May 1989.

CONTENTS
Research into the digestive physiology of pigs A.G. Low
Antinutritional factors (ANFs) in the nutrition of monogastric farm animals
J. Huisman 17
Research into the digestive physiology of the milk-fed calf R. Toullec, P. Guilloteau 37
Effect of porcine somatotropin on nitrogen gain and energy metabolism in fattening pigs
M.W.A. Verstegen, W. van der Hel 57
Practical application of (bio)synthetic amino acids in poultry and pig diets
J.B. Schutte /
Present and future developments in the protein/amino acid supply in monogastric farm animals
E.J. van Weerden °9

RESEARCH INTO THE DIGESTIVE PHYSIOLOGY OF PIGS
A.G. LOW
Pig Department, Agricultural and Food Research Council Institute for Grassland and Animal Production, Shinfield, Reading, Berkshire RG2 9AQ, U.K.
Summary
Historical development of digestive physiology is reviewed briefly. The importance of food intake, and in particular its neuro-endocrine relationship, is discussed. Gastric, biliary and pancreatic secretions and the effects of age and diet are described. The function of the small intestine both in terms of secretion and absorption are discussed with particular reference to practical measurement of protein quality and carbohydrate digestion and fermentation. The role of the large intestine in nutrient absorption is also discussed. Methods of measuring absorption using blood flow and sampling techniques are discussed. The review ends with some strategies for future research.
Keywords /
Pig, Digestion, Secretion, Absorption, Stomach, Small Intestine, Large Intestine, Protein, Carbohydrate.
1. Introduction
On the occasion of the retirement of E.J. Van Weerden after a distinguished career, it seems appropriate to review research in digestive physiology in the pig, because he has been instrumental in several important developments in the field. At the same time he has encouraged others to pursue innovative studies and he has put TNO in the forefront of research on this subject.
Historically, research into the digestive physiology of pigs was in progress at Rothamsted Research Station, Harpenden, in England in the middle of the nineteenth century, when observations were made on using an early form of cannula. However, major developments in this field in pigs were still nearly one hundred years away. In the last part of the nineteenth century and the early part of this century Pavlov and his colleagues in Russia laid the foundations of the subject of digestive physiology as we know it today. Their many experiments on the physiology of food consumption in dogs, and its reflex basis, and the subsequent series of digestive hydrolyses, involved the development of methods of surgery and physiological principles which were applied to pigs by Kvasnitskii and others from the 1930s onwards. Curiously enough these studies excited little interest in other countries until the early 1960s when Horszczaruk and colleagues in Poland began to examine hindgut function in pigs. Soon afterwards Cunningham, Friend & Nicholson (1963) began to make studies in Canada followed by the French group in Jouy-en-Josas led by Rérat (Auffray, Martinet & Rérat, 1967), and the English group at Shinfield formed by Braude during the 1960s. From then onwards development was rapid in all of these countries, and during the 1970s, further active new groups were formed in the USA, Australia, Cuba, Belgium, the Netherlands, Denmark, Sweden, the German Democratic Republic and West Germany. Since 1979 there have been

meetings on digestive physiology in the pig in Shinfield, England (1979), Jouy-en-Josas, France (1982), Copenhagen, Denmark (1985) and Jab^onna, Poland (1988). The next meeting will, appropriately, be held during 1991 in Wageningen. Each meeting has been followed by a publication containing important reviews and papers on new research topics.
The aim of this review is to discuss selected topics which have a particular interest for the author, and to make some suggestions for future developments in this field.
2. Food intake
The control of food intake in pigs, as in many other animals and in man, remains a topic of major interest and much uncertainty. The neuro-endocrine control of food intake is becoming progressively better understood, as reviews by Houpt (1982) for the pig, and Forbes (1988) for animals in general have shown. But much remains to be understood of what determines the pattern of food intake in relation to the physical, chemical and sensory properties of foods. While we know something of the way in which the central nervous system is integrated for the processes of digestion, through the action of regulatory peptides, glucostatic and gastrointestinal controls, it is nonetheless the case that most studies have been made under physiological conditions which are distant from those which occur when normal meals are eaten. These studies have revealed an extremely complex system, in which several of the regulatory peptides examined in detail appear to have several contrasting roles. Since food intake in practice appears to be the main limiting factor in pig production, better specific understanding of the neuro-endocrine regulation is much needed: in particular the roles of cholecystokinin (CCK), motilin, somatostatin and gastric inhibitory peptide merit study. One approach to this is specific immunological blocking of individual peptides to compare responses to their presence or absence, and this is the approach being currently developed at Shinfield. In addition, the central role of the vagus nerve in digestive function appears to be very complex and inadequately understood, as noted by Laplace (1989).
3. The mouth and salivary secretion
Although the pig is equipped with substantial teeth, under modern farming conditions very little food that requires much mastication is used, and thus food spends little time in the mouth. Perhaps for this reason, the amounts of saliva secreted are small, and there is little secretion except after meals (Arkhipovets, 1956; Corring, 1980). It is, however, difficult to measure salivary secretion from the individual ducts because of their small size and their location close to teeth which can easily damage catheters. The ratio of salivary amylase to pancreatic amylase is 1:250 000 according to Corring (1980) who collected saliva through an oesophageal cannula. It would be of interest to know how much saliva is secreted by wild pigs, who have to chew their food extensively, and by mature pigs, who visually appear to secrete substantial amounts of saliva, as do pigs under anaesthesia vhile undergoing gut surgery.
4. Gastric function
It has become clear in recent years that the stomach is an important reservoir of food, providing the intestines with a relatively constant supply of food, to some extent regardless of the times at which the

food was eaten, for 15-18h after a meal (Auffray, Martinet & Rérat, 1967; Zebrowska & Buraczewska, 1972; Braude, Fulford & Low, 1976). The stomach appears to be never completely empty and always displays motility and secretory activity.
Surgical methods of examining gastric function raise problems. Most of the work by Kvasnitskii (1951) involved either a simple stomach in the fundic region or a re-entrant cannula in the duodenum. A simple gastric cannula may be quite large and apparently does not markedly disturb motility (Cuber, Laplace & Villiers, 1980; Sissons & Rainbird, 1985), while re-entrant cannulae may be profoundly disturbing (Laplace, 1980). However it is difficult to make more than one observation per day with a simple gastric cannula if the entire contents are removed to provide a representative sample for analysis (Cuber et al., 1980; Low, Pittman & Elliott, 1985). A further major problem with re-entrant cannulation is that the cannula cannot be located proximal to the bile duct and it is not easy to place it proximal to the pancreatic duct. Hence the digesta collected from such cannulae inevitably contain post-gastric endogenous secretions in amounts which are considerable but which it is impossible to separate physically from gastric digesta.
The rate of gastric emptying is typically most rapid in the first hour after a meal, when elastic forces as well as physiological mechanisms, appear to be important influences; in general the larger the meal dry matter size, or the larger the volume of water given with the meal, the faster the initial rate of gastric emptying as shown in Table 1. (Low et al., 1985).
Table 1. Gastric Emptying (g/h) in 40kg Pigs Given 850g Diet and Varying Levels of Water
Diet and Water Hours after Meal Intake (g)
2337 958 304 47 70 2975 1220 565 238 -60 3612 1770 469 398 36
It is, however, not entirely clear what determines the emptying rate from a nutritional point of view: Braude et al. (1976) observed much more rapid emptying immediately after consumption of semi-purified diets than a diet based on barley, wheat and fishmeal, and this was confirmed by several other groups. The degree of osmolarity of digesta in the duodenum is thought to be directly proportional to an inhibitory effect on gastric emptying, but the putative mechanism has not been demonstrated in pigs. It is appealing to think that this was the mechanism involved in the work by Braude et al. (1976) because there was much more osmotically active material in the cereal-based diet. Deliberate modification of the composition of cereal-based diets e.g. by addition of oil, sucrose or cellulose did not lead to alterations in the rate of gastric emptying in the studies by Low et al. (1985) and these raised a question as to whether gastric emptying is readily manipulated by dietary changes. Subsequently we have examined the effect of a soluble non-starch polysaccharide (NSP), guar gum, on gastric emptying and again saw no effect of the NSP upon the rate of nutrient emptying from the stomach, though the rate of liquid emptying was delayed; this effect could have been due to changes in gastric secretion, or to the hydrophylic and viscous nature of guar gum

solutions (Rainbird & Low, 1986). Guar gum also depressed gastro-duodenal motility in corresponding studies (Rainbird & Sissons, unpublished).
The magnitude of gastric secretion of water containing electrolytes of both basic (in the oesophageal region) and acidic nature (in the fundic region) is remarkable. Pigs of 40 kg appear to secrete 10-20% of their total weight as gastric juice (based on measurement of duodenal digesta flow less separately measured outputs of bile and pancreatic juice) and of amounts of pepsin which could hydrolyse many times more peptide bonds than the diet contains (Zebrowska, Low & Zebrowska, 1983). Yet, in spite of the apparent overproduction of pepsin, there is clear evidence of adaptation of this enzyme to diet type in piglets (Cranwell, 1985) and growing pigs (Low, 1982). Earlier studies by Kvasnitskii (1951) provided an indirect indication of this. Secretion of gastric acid and water also appear to be diet-dependent: Kvasnitskii (1951) found that the higher the content of NSP in the diet, the greater the level of secretion, and this was confirmed by Zebrowska et al. (1983) and Cranwell, Low & Sambrook (unpublished). The latter authors prepared fully-innervated gastric pouches in 30 kg pigs. Such preparations do not permit complete collection of gastric secretion; the measurements are partly a function of the particular secretory tissue found in the chosen region of the stomach and are also unaffected by the mechanical stimulus that food particles are thought to exert on secretion. Even though the secretions in the gastric region are below pH2, the contents of the stomach lumen are greatly influenced by meal pH and buffering capacity. During the first 4 h after a meal when at least half of the meal leaves the stomach, the pH rarely falls below pH3.5, which is the upper optimum for the combined effect of the various pepsins found.
Insufficiency of gastric secretion, particularly of acid is conventionally thought to occur in piglets: this is a topic which deserves better understanding in relation to improved rearing systems.
Finally, it is often asked whether there is any significant gastric absorption of nutrients. Low & Rainbird (1986) were unable to detect 14C or 3H in peripheral blood following administration of amino acids or 3-o-methyl D-glucose, respectively, into the stomach of anaesthetized pigs with ligated oesophageal and pyloric sphincters. However Rérat et al. (unpublished) measured increased concentrations of amino acids and glucose in the efferent compared with the afferent gastric blood supply: however the concentrations of nutrients in their work were very much higher than used by Low & Rainbird (1986). It is possible that the transfer across the gastric mucosa seen by Rérat et al. (1989) was the result of passive diffusion, because Low & Rainbird (1986) could not demonstrate the presence of any active transport of amino acids or glucose in isolated gastric mucosa in vitro.
5. Duodenal Function
The digesta leaving the stomach have an increasingly low pH as time passes after a meal: they also tend to contain an increasing amount of soluble matter. However, the proportion of trichloroacetic acid-soluble nitrogen to total nitrogen rose from 12 to 50% over 12 h after a meal with casein as the sole protein source, whereas it was approximately 50% throughout this period for pigs receiving a barley-soya diet (Zebrowska et al., 1983). This digesta is immediately mixed with bile and then pancreatic juice and brought to a pH value between 6 and 7. Measurement of the contributions from these sources is not easy: while several authors have placed a catheter in one or

other duct, others have prepared a pouch into which the duct opened, sometimes in between a re-entrant cannula in the case of the pancreatic duct. Complex feedback mechanisms appear to regulate these secretions in part and return of the secretions is clearly essential for meaningful measurement (the effects of non-return have been demonstrated by Corring (1974) for pancreatic juice).
Both bile and pancreatic secretions have been clearly shown to respond to the nature of the diet but the relationships appear to be complex. For example, Aliev (1980) (cited by Juste, 1982) found that daily bile volume and lipid content rose as dietary lipid was increased to 5% (to 3.5 1 and 14.4 g, respectively) but then fell when the content rose further to 12%. These and other observations on biliary output were reviewed in detail by Juste (1982). Pancreatic output in pigs has been examined in more detail than biliary output and it has been found that the volume secreted increases as the meal size increases, or when the meal NSP content increases. Similarly, it is generally found that when the dietary fat or protein or carbohydrate content is increased there is a corresponding increase in the secretion of the appropriate enzymes. However, it is interesting to note that the amounts of trypsin and carboxypeptidase B did not decrease when pigs were given a protein-free diet after a diet with normal levels of protein, while the corresponding chymotrypsin and carboxypeptidase A activity disappeared almost completely after 2-3 days (Zebrowska, Low & Zebrowska, 1986). A remarkable aspect of pancreatic enzyme output is that the amounts secreted are often 10-20 times the amounts required under theoretically optimum conditions for the complete digestiorf of dietary protein, and yet there are numerous clear indications of adaptation to diet composition, as reviewed by Juste (1982), Simoes-Nunes (1982), or to trypsin inhibitors, as reviewed by Schneeman (1982). it remains an open question as to whether the amounts of enzyme secreted are not in fact large surpluses; under the conditions within the gut lumen very large variations in output are seen from day to day, implying relatively inexact control of synthesis or secretionof the enzymes, and perhaps a consequent need for surplus production. The neuro-endocrine regulation of pancreatic secretion in pigs has been the subject of much research in recent years by Corring and his group at Jouy-en-Josas, and the topic has been recently reviewed in detail, particularly at the level of putative molecular biological mechanisms (Corring, Juste & Lhoste, 1989). The pig pancreas offers a particularly exciting opportunity to understand the physiological and biochemical mechanisms of enzymic synthesis and secretion in eucaryotes since concurrent studies on secretion and on regualtory peptides are relatively straightforward. At a practical level, it is still not clear if there are ever instances of pancreatic insufficiency in normal animals, except soon after weaning when amounts of amylase and proteases may be limiting (though this is hard to prove).
Although the total volumes of endogenous inputs into the duodenum vary markedly they are substantial as shown in Table 2.
Table 2. Gastric, Biliary and Pancreatic Secretion (l/24h) in 40kg Pigs
Diet Gastric Bile Pancreatic Juice Juice
Barley-soya (high fibre) 8 1.8 2.5 Starch-Casein-cellulose (low fibre) 4 1.1 1.0

These secretions contain at least 6 g of nitrogen per 24 h, variable amounts of lipids and considerable quantities of electrolytes. A striking example of mineral secretion concerns the amounts of sodium passing the duodenum which were 400-638% higher than intake during 24 h periods; corresponding values for potassium were in the range 127-161% (Partridge, 1978). Th energy demand for synthesis of enzymes, epithelial cells, mucoprotein (formed throughout the gut as a protective agent) and movement of water and electrolytes has yet to be calculated but it is considerable. The integration of secretions to ensure that homeostasis is maintained in the gut lumen, even though different diets elicit very different endogenous responses, is as yet little understood in pigs. Attempts to measure the endogenous contribution to digesta in the duodenum have not always led to consistent answers, as noted by Low (1979a). The method of collection of digesta appears to have a major effect upon the result obtained: attempts to maintain the pressure which exists in the lumen of the duodenum during collection from re-entrant cannulas led to lower nitrogen flows than a free drainage system at room pressure (Low & Zebrowska, 1977). A later system for automatic measurement of digesta flow from re-entrant cannulas indicated that the attempts to maintain gut pressure may have inhibited digesta flow (Low, 1979b). These studies have led to some uncertainty about the extent to which there may be absorption in the proximal duodenum in pigs, or in the stomach, as discussed earlier. Zebrowska et al. (1982a,b) concluded that there is more gastric secretion than absorption in the proximal duodenum, but the methodology used makes it difficult to be certain that this was so.
6. Secretion and Absorption along the Small Intestine
Developments in the field of protein turnover in mammalian tissues and the whole body (Waterlow, Garlick & Millward, 1978; Simon, 1989) have provided a remarkable insight into the complex mechanisms which control the turnover of gut tissue. Nevertheless, the technical complications in such studies, which were discussed in the above reviews are formidable. Several different approaches have been taken in such work but it is clear that the turnover of gut tissue is of the order of 20% per day, while that of the pancreas has been estimated to be 75% per day, and muscle in the range of 2-3% per day. Even after allowance has been made for experimental inaccuracies, it is apparent that the digestive tract accounts for approximately 25% of the protein turnover in the whole body of pigs (Simon, Münchmeyer & Zebrowska, 1982). This process is clearly associated with a considerable demand for energy.
The wall of the small intestine is a major source of endogenous secretions; Horsczaruk, Buraczewska & Buraczewski (1974) estimated that 70 kg pigs secrete 6 1 of intestinal juice containing 8-12 g nitrogen. The nitrogen is a constituent of shed mucosal cells, plasma protreins and urea; the daily excretion of urea into the gut appears to be similar to the amounts excreted in urine in 24 h (Rérat & Buraczewska, 1986). It seems likely that the nature of the diet influences the amount and composition of these excretions; varying the composition of solutions used to perfuse isolated loops of jejunum in vivo led to marked changes in the output, of nitrogen (Buraczewska, 1579). Low & Rainbird (1984) found that NSP may also increase excretion of N into the jejunum.
Nutrient absorption along the small intestine of pigs has mainly been measured using re-entrant cannulas at various locations and calculating the difference between intake and output from cannulae. Thus Buraczewski, Buraczewska & Zebrowska (1975) found contrasting patterns

of disappearance of amino acids between different diets at seven sites along the small intestine. Information of this kind may be helpful in determining which parts of proteins are more or less readily digested by pigs. Measurements of the apparent absorption at several intestinal sites of amino acids (Low, 1979c), carbohydrates and lipid (Sambrook, 1979a,b) and minerals (Partridge, 1978) have served to demonstrate the complexities of inter-nutrient digestion. In most studies of this kind no attempt has been made to separate endogenous from dietary constituents of the digesta. The detailed physiological mechanisms involved in absorption have recently been reviewed by Friedrich (1989).
7. Absorption Measured at the End of the Small Intestine
Classical nutrition depends centrally upon measurements of the total and digestible content of nutrients in feedstuffs, and it will continue to do so in the foreseeable future. Until relatively recently, digestible amounts of nutrients have been measured by subtracting faecal loss from intake quantities, without correction for endogenous or bacterial contributions to faeces. Such measurements of apparent digestibility have been of major practical value especially for minerals, total carbohydrate and total lipid components of the diet. However, in the case of proteins, this is not so because of the role of the microflora of the large intestine. In what has become a classical study in pigs, Zebrowska (1973) maintained pigs on a protein-free diet given orally. During part of the experiment the pigs also received hydrolysed casein either through a duodenal cannula or through a cannula in the terminal ileum. The pigs were in negative nitrogen balance except when they received hydrolysed casein through the duodenal cannula as shown in Table 3.
Table 3. Nitrogen Balance (g/d) in Pigs Infused with Amino Acids into the Duodenum or Terminal Ileum while
Receiving a Protein-Free Diet Orally
! Input Urine
' Faeces
N balance
Duodenal Infusion
17.34 5.89 1.83
+9.62
Ileal Infusion
18.65 18.88 2.96
-3.19
NO Infusion
1.62 3.21 1.77
-3.36
This indicates that while the small intestine can absorb amino acids as such, the large intestine cannot: the metabolic fate of the amino acids is determined largely by bacteria which may incorporate them into their own protein or they may use them as energy or other metabolic fuels, with the release of ammonia and such compounds as diamines, which can be pharmacologically harmful to the animal. Furthermore, it is now evident that faecal nitrogenous compounds probably include less than 10% of undigested dietary components; most of the rest appear to be microbial in nature. Because of this microbial activity there have been many studies on the digestibility of protein, usually in terms of specific amino acids, at the end of the small intestine: indeed this has been the main focus of applied digestive physiology in the pig in recent years, and it has been reviewed several times (Just, 1980; Tanksley & Knabe, 1984; Sauer & Ozimek, 1986; Sibbald, 1987; Low &

Fisher, 1989). Much effort has been put into designing re-entrant cannulae which do not block up with fibrous feeds, by-passing the large intestine with ileo-rectal anastomoses, preserving the function of the ileo-caecal sphincter, and the use of simple cannulae coupled with a variety of non-absorbable and inert marker substances. However, it is important to re-assess critically the validity of such approaches in the light of current understanding of digestive function.
In most studies on ileal digestibility of amino acids measurements have been apparent and it is assumed that digesta simply contains amino acids of undigested dietary origin. However, it is clear that there are substantial quantities of endogenous amino acids in this region, and thus several attempts have been made to correct for them using protein-free diets. These raise problems as noted by Van weerden, Slump & Huisman (1980) in part because the amount of NSP which they contain markedly influences the amino acid content of digesta (Sauer et al., 1977). In addition, ileal digesta contain substantial amounts of Bacteria, which are able to incorporate urea secreted into the small intestine (Bergner et al,, 1986). It therefore seems necessary to remove all endogenous and bacterial components from digesta before the digestibility of the dietary amino acids can be measured, and this is no easy task.
From a practical viewpoint it can be seen that ileal digestibility values can help to improve the accuracy of diet formulation when unusual ingredients are used, or those which have been badly processed (Van Weerden et al., 1985) but not when feedstuff s used in studies leading to statements of requirements are used, since requirements automatically incorporate digestibility in empirical systems (Tanksley & Knabe, 1974). Fuller et al. (1981) compared dietary formulation on the basis of ileal digestible or gross dietary amino acids: free amino acids were added to the diet to meet ideal protein requirements and there were no clear advantages of basing this supplementation on ileal digestible rather than gross amino acid content. The response was measured as change in nitrogen retention; whether this was due to the unsuitability of ileal measurement or to shortcoming in the description of ideal protein. While measurements of the digestibility of amino acids in the ileum may be better than those made in faeces they may not be particularly accurate indicators of nutritive value. In the case of proteins containing poorly-available amino acids ileal digestibility values may seriously over-estimate their quality (E.S. Batterham; personal communication).
The marked microbial activity in the small intestine has considerable implications for carbohydrates as well. Millard & Chesson (1984) first demonstrated major disappearance of NSP monomers from swede anterior to the terminal ileum, as a result of microbial activity, or possibly non-enzymic hydrolysis (NSP are not digested by host carbohydrases, by definition). Graham, Hesselman & Aman (1986) found substantial NSP monomer disappearance from wheat bran, whole crop peas and beet pulp: uronic acids were particularly digestible. This largely microbial activity is associated with substantial production of volatile fatty acids. Longland, Close & Low (1989) also found that up to 70% of the uronic acids and 40% of the arabinose from diets containing sugar beet pulp were digested in pigs with ileo-rectal anastomoses as shown in Table 4. It seems difficult, therefore, to recommend that carbohydrates can be assessed in terms of digestibility at any specific sites within the gut. Faecal output of monomers gives the overall disappearance via either enzymic or microbial routes, but it is as yet not possible to make any accurate correction for any monomers which are of endogenous or bacterial origin, though clearly some of them must be. However, Longland & Low (1989) have concluded that less than 12% of
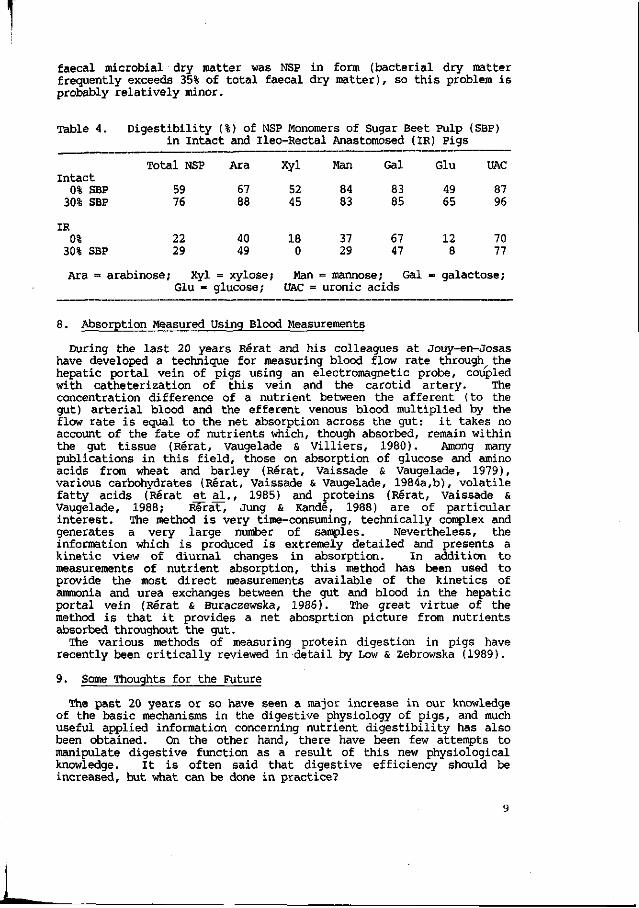
faecal microbial dry matter was NSP in form (bacterial dry matter frequently exceeds 35% of total faecal dry matter), so this problem is probably relatively minor.
Table 4. Digestibility (%) of NSP Monomers of Sugar Beet Pulp (SBP) in Intact and Ileo-Rectal Anastomosed (IR) Pigs
Intact 0% SBP
30% SBP
IR 0%
30% SBP
Total NSP
59 76
Ara = arabinose
22 29
; Xyl Glu =
8. Absorption Measured
Ara
67 88
40 49
= xylose; glucose;
Xyl
52 45
18 0
Man UAC =
Man
84 83
37 29
Gal
83 85
67 47
= mannose; Gal uronic acids
Using Blood Measurements
Glu UAC
49 87 65 96
12 70 8 77
= galactose;
During the last 20 years Rérat and his colleagues at Jouy-en-Josas have developed a technique for measuring blood flow rate through^ the hepatic portal vein of pigs using an electromagnetic probe, coupled with catheterization of this vein and the carotid artery. The concentration difference of a nutrient between the afferent (to the gut) arterial blood and the efferent venous blood multiplied by the flow rate is equal to the net absorption across the gut: it takes no account of the fate of nutrients which, though absorbed, remain within the gut tissue (Rérat, Vaugelade & Villiers, 1980). Among many publications in this field, those on absorption of glucose and amino acids from wheat and barley (Rérat, Vaissade & Vaugelade, 1979), various carbohydrates (Rérat, Vaissade & Vaugelade, 1984a,b), volatile fatty acids (Rérat et al., 1985) and proteins (Rérat, Vaissade & Vaugelade, 1988; Rerat, Jung & Kandé, 1988) are of particular interest. The method is very time-consuming, technically complex and generates a very large number of samples. Nevertheless, the information which is produced is extremely detailed and presents a kinetic view of diurnal changes in absorption. In addition to measurements of nutrient absorption, this method has been used to provide the most direct measurements available of the kinetics of ammonia and urea exchanges between the gut and blood in the hepatic portal vein (Rérat & Buraczewska, 1986). The great virtue of the method is that it provides a net abosprtion picture from nutrients absorbed throughout the gut.
The various methods of measuring protein digestion in pigs have recently been critically reviewed in detail by Low & Zebrowska (1989).
9. Some Thoughts for the Future
The past 20 years or so have seen a major increase in our knowledge of the basic mechanisms in the digestive physiology of pigs, and much useful applied information concerning nutrient digestibility has also been obtained. On the other hand, there have been few attempts to manipulate digestive function as a result of this new physiological knowledge. It is often said that digestive efficiency should be increased, but what can be done in practice?

In the case of cereal-based diets it is usual for at least 20% of the dry matter ingested to be voided in faeces (including 20% of the nitrogen, with a further 25-30% appearing in urine). This faecal matter is largely microbial, endogenous or fibrous in form. Can these components be reduced? Antibiotics tend to increase nutrient digestibility, presumably because they lower bacterial fermentation of dietary compounds, allowing more digestion by host enzymes to absorbable monomers rather than bacterial mass. The use of antibiotics is already limited in some countries and it is possible that they will only be used for therapeutic purposes before long. If so, can microbial activity be reduced, or modified, by dietary change instead, or by modifying gut physiology e.g. by changing motility in such a way that passage rate is accelerated to an extent that host enzymic digestion is not impaired but microbial fermentation is suppressed by insufficient time for action? As yet we have no means of fine control of motility in pigs because we do not understand enough of the way this process is controlled in a neuro-endocrine context, and we do not know how the latter could be influenced by the nature of the diet. An alternative strategy might be to try and modify the microbial population so that its activities were mainly complete in the small intestine, followed by death and digestion of the cells and absorption of the nutritional components by the host. But the large intestine has evolved to provide a fermentation chamber for undigested dietary and endogenous residues and it may well be very difficult to change this. Attempts to modify the microbial population by diet appear to have limited success as found by Liu et al. (1985) who gave a wide range of NSP sources to pigs and found no significant evidence of adaptation. It may well be that the metabolic activity of bacteria is more sensitive to change that species distribution.
The major extent of endogenous secretion into the gut has been discussed in some detail earlier in this review. It is pertinent to examine whether protein turnover and excretion could be reduced, by dietary means, without detriment to the animal. As yet we cannot predict whether this is even a possibility.
The digestibility of dietary components such as starch and proteins appears, from various studies, to be essentially completed, unless these are closely associated in the diet with NSP and lignin: lignin in particular is known to determine the rate of fermentation of many feedstuffs (Van Soest, 1985) and so any reduction in the "barrier" effects of these compounds would be worthwhile, perhaps by adding exogenous NSP-digesting enzymes, as is already practised to some extent commercially, especially with young pigs. Such enzymic treatment might be effective before the diet is eaten in some cases, in order to provide the anterior part of the digestive tract with components which are likely to be digestible. This can only be done in a soundly-based way if the physico-chemical structure of feeds is well characterised; without such information, tailoring exogenous enzymes, or other physical treatments, is very haphazard.
In conclusion, I believe that fundamental research on digestive physiology, combined with better understanding of food structure and composition will continue to play an important role in raising the efficiency of pig production. At the same time it can be hoped that the environment will be improved through reducing effluent production. In addition there are good prospects for increasing the use that pigs can make, as omnivores, of foods that are inedible or at least unattractive for human consumption, to produce a highly palatable and nutritious food.
10

r References
Arkhipovets, A.I. 1956. Age characteristics of salivary secretion in young pigs. Journal of Physiology of USSR, 42, 882-886.
Auffray, P., J. Martinet & A. Rérat. 1967. Some aspects of gastrointestinal transit in the pig. Annales de Biologie Animale, Biochimie, Biophysique, 7, 261—269.
Bergner, H., 0. Simon, T. Zebrowska & R. Münchmeyer. 1986. Studies on the secretion of amino acids into the gastrointestinal tract of pigs. 3. Secretion of urea determined by continuous intravenous infusion of 1 N-urea. Archiv für Tierernährung, 36, 479-490.
Braude, R., R.J. Fulford & A.G. Low. 1976. Studies on digestion and absorption in the intestines of growing pigs. Measurements of the flow of digesta and pH. British Journal of Nutrition, 36, 497-510.
Buraczewska, L. 1979. Secretion of nitrogenous compounds in the small intestine of pigs. Acta Physiologica Polonica, 30, 319-326.
Buraczewska, L., S. Buraczewski & T. Zebrowska. 1975. Digestion and absorption in the small intestine of pigs. 2. Amino acid content in digesta and their absorption. Roczniki Nauk Rolniczych, 97B(1), 103-115.
Buraczewska, L., S. Buraczewski, B. Pastuczewska & T. Zebrowska (Editors). 1989. Digestive Physiology in the Pig. Polish Academy of Sciences, Warsaw, Poland. 407 pp.
Corring, T. 1974. Regulation of pancreatic secretion by negative feedback in the pig. Annales de Biologie Animale, Biochimie, Biophysique, 14, 487-498.
Corring, T. 1980. Endogenous secretions in the pig. In: Low, A.G. & I.G. Partridge (Editors), Current Concepts of Digestion and Absorption in Pigs. National Institute for Research in Dairying, Reading, U.K.
Corring, T., C. Juste & E.F. Lhoste. 1989. Nutritional regulation of pancreatic and biliary secretions. Nutrition Research Reviews, 2. (in press)
Cranwell, P.D. 1985. The development of acid and pepsin (E.C.3.4.23.1) secretory capacity in the pig: effects of age and weaning. British Journal of Nutrition, 54, 305-320.
Cuber, J.C., J.P. Laplace & P.A. Villiers. 1980. Fistulation of the stomach and residual gastric contents after intake of a semi-purified maize diet in the pig. Reproduction, Nutrition et Développement, 20, 1161-1172.
Cunningham, H.M., D.W. Friend & J.W.G. Nicholson. 1963. Observations on digestion in the pig using a re-entrant fistula. Canadian Journal of Animal Science, 43, 215-225.
Forbes, J.M. 1988. Metabolic aspects of the regulation of voluntary food intake and appetite. Nutrition Research Reviews, 1, 145-168.
Friedrich, M. 1989. Physiology of intestinal digestion and absorption. In: Bock, H.D., B.O. Eggum, A.G. Low, O. Simon & T. Zebrowska (Editors), Protein Metabolism in Farm Animals. Oxford University Press, Oxford, U.K. pp. 218-272.
Fuller, M.F., B.A. Baird, A. Cadenhead & R. Aitken. 1981. An assessment of amino acid digestibility at the terminal ileum as a measure of the nutritive value of proteins for pigs. Animal Production, 32, 396.
Graham, H., K. Hesselman & P. Aman. 1986. The influence of wheatbran and sugar-beet pulp on the digestibility of dietary components in a cereal based pig diet. Journal of Nutrition, 116, 242-251.
Horszczaruk, F., L. Buraczewska & S. Buraczewski. 1974. Amount and composition of intestinal juice collected from isolated intestinal loops of pigs. Roczniki Nauk Rolniczych, 95B(4), 69-77.
11

Houpt, T.R. 1982. The controls of food intake in the pig. In: Laplace, J.P., T. Corring & A. Rérat (Editors), Digestive Physiology in the Pig. Institut National de la Recherche Agronomique, Paris, France, pp. 17-28.
Just, A. 1989. Ileal digestibility of protein: applied aspects. In: Current Concepts of Digestion and Absorption in Pigs. National Institute for Research in Dairying, Reading, U.K. pp. 66-75.
Just, A., H. J0rgensen & J.A. Fernandez (Editors). 1985. Digestive Physiology in the Pig. National Institute of Animal Science, Copenhagen, Denmark. 401 pp.
Juste, C. 1982. Endogenous supplies from the digestive secretions in the pig. In: Laplace, J.P., T. Corring & A. Rérat (Editors), Digestive Physiology in the Pig. Institut National de la Recherche Agronomique, Paris, France, pp. 155-174.
Kvasnitskii, A.V. 1951. Problems of Digestive Physiology in Pigs. Sel'Khozgiz, Moscow, USSR. 200 pp.
Laplace, J.P. 1980. Stomach and small intestine motility in the pig: electromyography in nutritional studies. In: Low, A.G. & I.G. Partridge (Editors), Current Concepts of Digestion and Absorption in Pigs. National Institute for Research in Dairying, Reading, U.K. pp. 24-47.
Laplace, J.P., M.B. Felix, 0. Rampin & J.C. Marcilloux. 1989. Food intake - G.I. tract relationship in the pig. In: Buraczewska, L., S. Buraczewski, B. Pastuzewska & T. Zebrowska (Editors), Digestive Physiology in the Pig. Polish Academy of Sciences, Warsaw, Poland. pp. 18-35.
Laplace, J.P., T. Corring & A. Rérat (Editors). 1982. Digestive Physiology in the Pig. Institut National de la Recherche Agronomique, Paris, France. 317 pp.
Liu, Y.F., K. Fadden, E.A. Latymer, A.G. Low & M.J. Hill. 1985. The use of the cannulated pig to study the effect of dietary fibre -supplements on the bacterial flora of the porcine hindgut. in: Just, A., H. J0rgensen & J.A. Fernandez (Editors), Digestive Physiology in the Pig. National Institute of Animal Science, Copenhagen, Denmark, pp. 300-303.
Longland, A.C. & A.G. Low. 1989. Digestion of diets containing molassed or plain sugar beet pulp by pigs. Animal Feed Science and Technology, (in press)
Longland, A.C., W.H. Close & A.G. Low. 1989. Digestion of carbohydrates from sugar beet pulp in pigs with ileo-rectal anastomoses and energy balance measured by calorimetry. In: Buraczewska, L., S. Buraczewski, B. Pastuzewska & T. Zebrowska (Editors), Digestive Physiology in the Pig. Polish Academy of Sciences, Warsaw, Poland, pp. 108-119.
Low, A.G. 1979a. Studies on digestion and absorption in the intestines of growing pigs. 5. Measurements of the flow of nitrogen. British Journal of Nutrition, 41, 137-146.
Low, A.G. 1979b. A new automatic method for measuring dry matter and nitrogen flow through re-entrant cannulas in the duodenum of growing pigs. Proceedings of the Nutrition Society, 38, 129A.
Low, A.G. 1979c. Studies on digestion and absorption in the intestines of growing pigs. 6. Measurements of the flow of amino acids. British Journal of Nutrition, 41, 147-156.
Low, A.G. 1982. The activity of pepsin, chymotrypsin and trypsin during 24h periods in the small intestine of growing pigs. British Journal of Nutrition, 48, 147-159.
Low, A.G. & A.L. Rainbird. 1986. Lack of evidence for amino acid and glucose absorption in the stomach of pigs. Archiv für Tierernährung, 36, 327.
12

Low, A.G. & C. Fisher. 1989. Protein Evaluation in pigs and poultry. In: Wiseman, J. & D.J.A. Cole, Feedstuff Evaluation. Butterworths, London, U.K. (in press)
Low, A.G. & T. Zebrowska. 1977. Dry matter and nitrogen in the duodenal contents of growing pigs: a discrepancy explained. British Journal of Nutrition, 38, 145-147.
Low, A.G., R.J. Pittman & R.J. Elliott. 1985. Gastric emptying of barley-soya bean diets in the pig: effects of feeding level, supplementary maize oil, sucrose or cellulose, and water intake. British Journal of Nutrition, 54, 437-447.
LoWj A.G. & I.G. Partridge (Editors). 1979. Current Concepts of Digestion and Absorption in Pigs. National Institute for Research in Dairying, Reading, U.K. 222 pp.
Millard, P. & A. Chesson. 1984. Modifications to swede (Brassica napus L.) anterior to the terminal ileum of pigs: some implications for the analysis of dietary fibre. British Journal of Nutrition, 52, 583-594.
Partridge, I.G. 1978. Studies on digestion and absorption in the intestines of growing pigs. 3. Net movement of mineral nutrients in the digestive tract. British Journal of Nutrition, 39, 527-537.
Rainbird, A.L. & A.G. Low. 1986. Effect of guar gum on gastric emptying in growing pigs. British Journal of Nutrition, 55, 87-98.
Rérat, A. & L. Buraczewska. 1986. Postprandial quantitative kinetics of urea and ammonia nitrogen exchanges between the digestive tract and the portal blood in conscious pigs receiving a diet with or without urea. Archiv für Tierernährung, 36, 252-269.
Rérat, A., J. Jung & J. Kandé. 1988. Absorption kinetics of dietary hydrolysis products in conscious pigs given diets with different amounts of fish protein. 2. Individual amino acids. British Journal of Nutrition, 60, 105-120.
Rérat, A., M. Fiszlewicz, P. Herpin, P. Vaugelade & M. Durand. 1985. measurement of the appearance of volatile fatty acids in the portal vein during digestion in the pig. Comptes Rendus de la Académie des Sciences, Paris, 300, Serie 3, 467-470.
Rérat, A., P. Vaissade & P. Vaugelade. 1984. Absorption kinetics of some carbohydrates in conscious pigs. 2. Quantitative aspects. British Journal of Nutrition, 51, 517-529.
Rérat, A., P. Vaissade & P. Vaugelade. 1984. Absorption kinetics of some carbohydrates in conscious pigs. 1. Qualitative aspects. British Journal of Nutrition, 51, 505-515.
Rérat, A., P. Vaissade & P. Vaugelade. 1988. Absoprtion kinetics of dietary hydrolysis products in conscious pigs given diets with different amounts of fish protein. 1. Amino-nitrogen and glucose. British Journal of Nutrition, 60, 91-104.
Rérat, A., P. Vaugelade & P. Villiers. 1980. A new method for measuring the absorption of nutrients in the pig: critical examination. In: Low, A.G. & I.G. Partridge (Editors), Current Concepts of Digestion and Absorption in Pigs. National Institute for Research in Dairying, Reading, U.K. pp. 177-214.
Sambrook, I.E. 1979a. Studies on digestion and absorption in growing pigs. 7. Measurements of the flow of total carbohydrate, total reducing substances and glucose. British Journal of Nutrition, 42, 267-277.
Sambrook, I.E. 1979b. Studies on digestion and absorption in growing pigs. 8. Measurements of the flow of total lipid, acid detergent fibre and volatile fatty acids. British Journal of Nutrition, 42, 279-287.
13

Sauer, W.C. & L. Ozimek. 1986. Digestibility of amino acids in swine: results and their practical applications. Livestock Production Science, 15, 367-388.
Sauer, W.C., S.C. Stothers, G.D. Phillips & R.J. Parker. 1977. Apparent and true availability of amino acids in wheat and milling by-products for pigs. Canadian Journal of Animal Science, 57, 775-784.
Schneeman, B. 1982. Digestive enzyme activities from the pancreas in response to diet. In: Laplace, J.P., T. Corring & A. Rérat (Editors), Digestive Physiology in the Pig. Institut National de la Recherche Agronomique, Paris, France, pp. 125-131.
Sibbald, I.R. 1987. Estimation of bioavailable amino acids for feedingstuffs for poultry and pigs: a review with emphasis on balance experiments. Canadian Journal of Animal Science, 67, 221-300.
Simoes-Nunes, C. 1982. Some aspects of digestive enzyme development with age and diet composition adaptation. In: Laplace, J.P., T. Corring & A. Rérat (Editors), Digestive Physiology in the Pig. Institut National de la Recherche Agronomique, Paris, France. pp. 133-151.
Simon, 0. 1989. Metabolism of proteins and amino acids. In: Bock, H.D., B.0. Eggum, A.G. Low, 0. Simon & T. Zebrowska (Editors), Protein Metabolism in Farm Animals. Oxford University Press, Oxford, U.K. pp. 273-366.
Simon, 0., R. Münchmeyer & T. Zebrowska. 1982. Studies on the range of tissue protein synthesis in pigs: the effects of thyroid hormones. British Journal of Nutrition, 48, 571-582.
Tanksley, T.D. & D.A. Knabe. 1984. Ileal digestibilities of amino acids in pig feeds and their use in formulating diets. in: Haresign, W. & D.J.A. Cole (Editors), Recent Advances in Animal Nutrition - 1984. Butterworths, London, U.K.
Van Soest, P.J. 1985. Definition of fibre in animal feeds. In : Haresign, W. & D.J.A. Cole (Editors), Recent Advances in Animal Nutrition - 1985. Butterworths, London, pp. 55-70.
Van Weerden, E.J., J. Huisman, P. Van Leeuwen & P. Slump. 1985. The sensitivity of the ileal digestibility method as compared to the faecal digestibility to method. In: Just, A., H. Jorgensen & J.A. Fernandez (Editors), Digestive Physiology in the Pig. National Institute of Animal Science, Copenhagen, Denmark, pp. 392-395.
Van Weerden, E.J., P. Slump & J. Huisman. 1980. Amino acid digestion in different parts of the intestinal tract in pigs. In: Oslage, H.J. & K. Rohr (Editors), Protein Metabolism and Nutrition. Vol. 1, E.A.A.P. Publication No. 27, Braunschweig, West Germany. pp. 207-214.
Waterlow, J.C., P.J. Garlick & D.J. Millward. 1978. Protein Turnover in Mammalian Tissues and In the Whole Body. North-Holland Publishers, Amsterdam, Netherlands. 804 pp.
Zebrowska, T. 1973. Digestion and absorption of nitrogenous compounds in the large intestine of pigs. Roczniki Nauk Rolniczych, 95B(3), 85-90.
Zebrowska, T. & L. Buraczewska. 1972. Influence of dietary protein level on the rate of digestion in the small intestine of pigs. I. Amount and composition of digestion. Roczniki Nauk Rolniczych, 94B(1), 71-75.
Zebrowska, T., A.G. Low & H. Zebrowska. 1983. Studies on gastric digestion of protein and carbohydrate, gastric secretion and exocrine pancreatic secretion in the growing pig. British Journal of Nutrition, 49, 401-410.
14

Zebrowska, T., A.G. Low & H. Zebrowska. 1986. The flow and composition of duodenal digesta and pancreatic juice of pigs given a protein-free diet. Archiv für Tierernährung, 36, 331-332.
Zebrowska, T., O. Simon, R. Münchmeyer, E. Wolf, H. Bergner & H. Zebrowska. 1982a. Flow of endogenous amino acids along the gut of pigs. Archiv für Tierernährung, 32, 431-444.
Zebrowska, T., O. Simon, R. Münchmeyer, E. Wolf, H. Bergner & H. Zebrowska. 1982b. Investigation on the amino acid secretion and absorption in the stomach of the growing pig. Archiv für Tierernährung, 32, 703-710.
15
L

ANTINUTRITIONAL FACTORS (ANFs) IN THE NUTRITION OF MONOGASTRIC FARM ANIMALS
J. Huisman
TNO Institute for Animal Nutrition and Physiology (ILOB), P.O. Box 15, 6700 AA Wageningen
Summary
Many seeds contain ANFs. In soya, ANFs are trypsin inhibitors, lectins, Some examples of the effects of these animals are presented and discussed. Various points regarding the adequacy trypsin inhibitors and antigenic prot lectins and antigenic proteins have b Examples are given demonstrating that sensitivity to ANFs between animal sp studies into the effects of ANFs shou animals.
For future prospects the following po act in the animal, improvement of ana levels for ANFs.
beans and peas the most important tannins and antigenic proteins, factors in monogastric farm
of analytical methods for lectins, eins are discussed. Some assays for een shown to be inadequate.
there is a difference in ecies. The results show that Id be carried out in target
ints are discussed: the way ANFs lytical methods and threshold
1. Introduction
Many feedstuffs contain factors which produce different deleterious effects in animals (Liener, 1980; 1989; Marquardt, 1989; Pusztai, 1989). If these factors cause a negative effect on growth, feed conversion efficiency and/or health, they are referred to as "antinutritional factors" (ANFs). However, in this definition fibre may also be classified as an ANF. Therefore, one restriction is that ANFs have no feeding value. In plants and seeds these factors often act as biopesticides, protecting the seed against moulds, bacteria, and birds (Birk, 1987, 1989; Bond & Smith, 1989; Liener, 1980; 1989).
ANFs can be classified in various ways. In the following scheme they are classified on the basis of their effects on nutritive value of feedstuffs and biological response in the animal: - factors which have a depressive effect on protein digestion and on the
utilization of protein (trypsin and chymotrypsin inhibitors, lectins, polyphenolic compounds, saponins).
- factors which have a negative effect on digestion of carbohydrates (amylase inhibitors, polyphenolic coumpounds, flatulence factors).
- factors which have a negative effect on the utilization of minerals (glucosinolates, oxalic acid, phytic acid, gossypol).
- factors which inactivate vitamins or cause an increase in the animals' vitamin requirement (anti-vitamins).
In this overview the following points will be discussed: the occurrence of ANF in different seeds, some effects of ANFs in the animal, adequacy of ANF analysis, sensitivity of different animal species to ANFs and some future prospects.
2- Occurrence in seeds
Many seeds contain ANFs. The levels of different ANFs vary considerably between the different seeds. Table 1 summarizes the ANFs in the various seeds.
17

It shows that protease inhibitors and lectins are most important in the legume seeds soya, peas and beans, but some varieties of rye and triticale may also contain moderate levels of trypsin inhibitors. Tannins are mainly present in the coloured flowering varieties of Vicia faba beans and peas, and in sorghum and rapeseed. Glucosinolates are important in rapeseed, alkaloids in lupins, and gossypol in cottonseed. As demonstrated by Liener (1981), one particular seed often contains more than one ANF. For example soya contains: trypsin inhibitors, haemagglutinins (lectins), goitrogens, antivitamins, phytates, saponins, oestrogens, flatulence factors and allergens.
This overview will be restricted to the ANFs present in the legume seeds soya, beans and peas.
Table 1. Antinutritional factors in cereals and seeds.
Cereals/seeds Antinutritional factors
Protease inhibitor
Lectins Tannins/
Polyphenolic
compounds
Others
Cereal grains
Wheat Barley Rye Triticale Rice
Sorghum Corn
Legume see
Soya
Vicia faba
Phaseolus
ds
bean bean
-/ +
-/ + -/+/++ -/+/++ -/ + -/ + -/ +
++/+++
-/ + -/+/++
Peas
Lupins
Other seeds
+/++/+++
++ + ++/+++
+/++
_
-++/+++ +
+/++
_
---
-
+/++/+++ a)
Rapeseed
Sunflowerseed
Cottonseed
Peanut
-/ +
-/ +
+/++ +/++/+++
+ / + +d )
+ /++ e)
b)
+/++/+++ c)
Below detection limit + Low level ++ Medium level +++ High level
a) Alkaloids b) Glucosinolates c) Gossypol d) Phenolic compounds (3-3.5%) e) 16-182 tannins in the skin

3. Some effects of ANFs in monogastric farm animals
As mentioned, a seed of one particular species may contain various ANFs. Therefore, the negative effects in the animal is generally not attributed to one particular ANF. Moreover, the various ANFs have their own specific effects. In Table 2 a survey is given of the major effects of ANFs on digestion and the utilization of nutrients in monogastric animals.
Table 2. Major effects in monogastric farm animals of antinutritional factors present in legume seeds
Antinutritional factor
Major in vivo effect
References
Lectins -damage of gut wall
-immunological reaction
-metabolism toxicity
Donatucci, 1983; Donatucci et al., 1988; Greer, 1983; Kik, 1988; Liener, 1986; Pusztai, 1987.
Protease inhibitors -trypsin-chymotrypsin inhibitor
-reduction activity of (chymo)trypsin
-pancreas hypertrophy -decreased digestion
Burns, 1987; Liener, 1979; Liener and Kakade, 1980; Richardson, 1^80-81;Scarbieri & Whitaker, 1982; Birk, 1989.
-«-Amylase inhibitor -forming complex with Powers & Whitaker, amylase in salivary and pancreatic juice
-reduces starch avai-labilj ty
1977.
Tannins and polyphenol ic compounds
-forms complex with enzymes or feed protein
-reduces protein digestibility
Griffith, 1981; Philips et al., 1981; Aw & Swanson, 1985; Marquardt, 1989.
Flatulence factors -gastrointestinal Fleming, 1981; Fleming discomfort et al., 1988; Saini,
-increased maintenance 1989.
Phytic acid -forms complex with Reddy et al., 1982. minerals, and protein
-depresses absorption of minerals
Antigenic proteins -gut wall damage -immune response
Miller et al., 1984; Seegraber and Morril, 1982, 1986; Sissons and Smith, 1976; Kilshaw and Sissons, 1979.
Adapted with modifications from Van der Poel (1989).
19

Lectins are characterized by their unique ability to bind to specific sugars. Glycoproteins in the gut wall contain sugars to which lectins have affinity and, as a result, binding of lectins to the epithelial cells occur. Due to this binding, a series of effects occur, accumulating in growth depression. The main biological effects induced by the binding of lectins are: damage to the gut wall (Jaffe, 1980; Meyer et al., 1982; Pusztai, 1987; 1989; Torres-Pinedo, 1983), impaired transportation of nutrients across the intestinal wall (Donatucci et al., 1988; Jaffe, 1980; Liener, 1986), increased synthesis of mucosal protein (Liener, 1986; Pusztai, 1987), muscle atrophy (Pusztai, 1989), depressed blood insulin levels (Pusztai, 1989), inhibition of brush border hydrolases (Kim et al., 1976; Nakata & Kimura, 1980), and effects on the immune system (Pusztai, 1989). These effects may result in serious growth depression (Grant et al., 1983; Pusztai et al., 1981; King et al., 1983; Huisman et al., 1987, 1989; Huisman & van der Poel, 1989,a,b,c,).
Protease inhibitors are widely distributed in plant seeds. Legume seeds in general contain high levels of these inhibitors. There are various families of plant protein inhibitors (Birk, 1989). The main inhibitors in legume seeds are the trypsin and chymotrypsin inhibitor. These inhibitors are peptides which can form stable, inactive complexes with the proteolytic enzymes from the pancreas (Kakade & Liener, 1980). Due to this complex forming, the activity of the trypsin and chymotrypsin is decreased (Liener & Kakade, 1980; Rackis & Gumbmann, 1981; Rackis et al., 1985). Inactivation of the trypsin in the gut induces the endocrine cells in the mucosa to release more of the hormone cholecystokinin (CCK) which stimulates the pancreas to produce more digestive enzymes, such as trypsin, amylase and elastase (Birk, 1989; Liener & Kakade, 1980; Liener, 1989, Schuman et al., 1983). The net result is an endogenous loss of protein rich in S-containing amino acids which leads to depressed growth (Kakade & Liener, 1980; Liener, 1989). Because of the stimulated production of pancreatic enzymes, the pancreas becomes enlarged in small animal species due to hypertrophic and hyperplastic changes in morphology (Birk, 1989; Gallaher & Schneeman, 1986; Gumbmann et al., 1985; Liener & Kakade, 1980).
The growth depressing effect of protease inhibitors is mainly attributed to the loss of endogenous protein due to the negative feedback mechanism.
The oc-amylase inhibitor has been indicated as being responsible for the impaired digestion of starch in red kidney beans (Jaffe & Vega Letta, 1968). However, addition of this purified inhibitor did not affect the starch availability (Savaiano et al., 1977). Therefore, this factor seems to be of minor importance in these beans.
Tannins are polyphenolic compounds. The way these compounds act in the animal is not entirely clear. Tannins form complexes with proteins and carbohydrates in the feed but also with digestive enzymes. Due to this complex forming, the activity of digestive enzymes and the digestibility of nutrients is decreased (Griffiths & Mosely, 1980; Marquardt, 1989). Other antinutritional effects which have been attributed to tannins are damage to the gut wall, toxicity of tannins absorbed from the gut and interference with the absorption of some minerals (Liener, 1989; Mitjavila et al., 1977).
Flatulence factors are related to oligosaccharides which are broken down by intestinal bacteria in the large intestine. These oligosaccharides are not broken down in the small intestine due to the lack of appropriate enzymes, and so they flow into the large intestine where they are degraded by the action of bacterial a-galactosidae.
20
The cleavage products are converted methane, resulting in flatulence, d discomfort in the animals (Rackis,
Antigenic proteins cause gut wal in the gut linked with disorders in calves (Kilshaw & Sissons, 1979; Mi Morril, 1982, 1986; Sissons and Smi
Phytic acid forms complexes with resulting in a reduced availability and Fe. It also inhibits several of pancreatin and a-amylase.
into carbon dioxide, hydrogen and iarrhoea, nausea, cramp and 1975; Saini, 1989). 1 damage and immunological reactions
gut function in piglets and veal lier et al., 1984; Seegraber and th, 1976).
divalent and trivalent metal ions of minerals such as Ca, Mg, Zn, Cu digestive enzymes such as pepsin,
In the following some examples of the effects of ANFs in monogastric animals are presented. Table 3 summarizes the growth results of an experiment in which piglets and rats were fed a control diet or test diets containing ZOZ raw or toasted Phaseolus vulgaris beans. All diets were balanced for contents of protein and amino acids, net energy, and vitamins and minerals (for details see Huisman and Van der Poel, 1989b) . Phaseolus vulgaris beans were chosen as being representative of a seed containing high levels of toxic lectins.
Table 3. Growth and feed conversion efficiency in piglets and rats fed Ph. vulgaris bean for 3 weeks. ,
Diets
Piglets
Growth
g/day SD
Feed conversion efficiency
SD
control 137.7 19.3 100 1.74 0.12 100
20Z raw beans
in the diet -36.0
201 toasted beans
in the diet 111.8C
Rats
5.8 -26 negative
20.7 81 2.09c 0.11 120
control 6.68 0.24 100 2.99 0.21 100
2.0Z raw beans
in the diet 4.86 0.20 73 3.72 0.32 124
207. toasted beans
in the diet 6.32 0.46 95 3.10a 0.18 104
Data in the same column of each species with a different superscript differ significantly (P < 0.05).
21
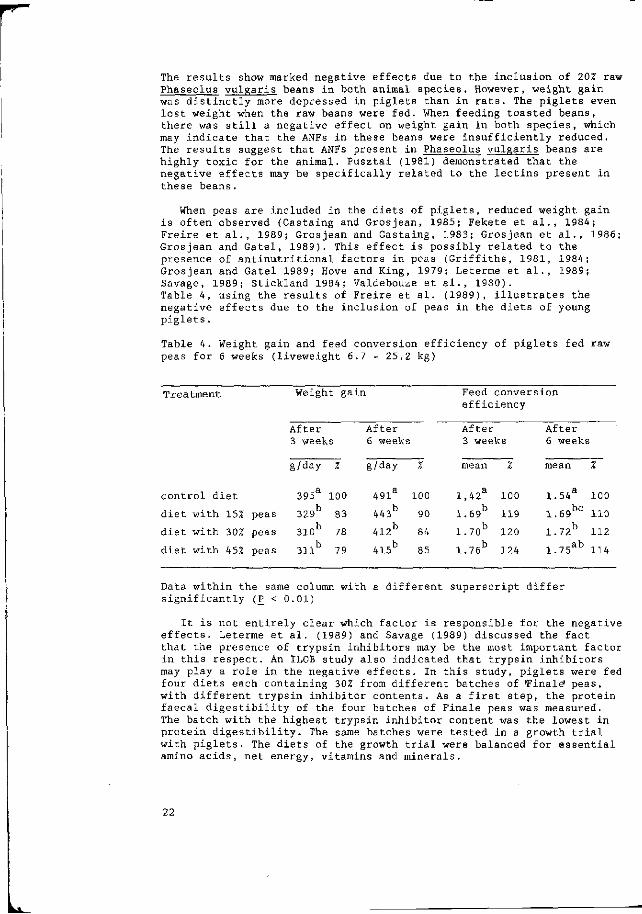
The results show marked negat Phaseolus vulgaris beans in b' was distinctly more depressed lost weight when the raw bean there was still a negative ef may indicate that the ANFs in The results suggest that ANFs highly toxic for the animal, negative effects may be speci these beans.
ive effects due to the inclusion of 20Z raw oth animal species. However, weight gain
in piglets than in rats. The piglets even s were fed. When feeding toasted beans, feet on weight gain in both species, which
these beans were insufficiently reduced. present in Phaseolus vulgaris beans are
Pusztai (1981) demonstrated that the fically related to the lectins present in
When peas are included in the diets of piglets, reduced weight gain is often observed (Castaing and Grosjean, 1985; Fekete et al., 1984; Freire et al., 1989; Grosjean and Castaing, 1983; Grosjean et al., 1986; Grosjean and Gatel, 1989). This effect is possibly related to the presence of antinutritional factors in peas (Griffiths, 1981, 1984; Grosjean and Gatel 1989; Hove and King, 1979; Leterme et al., 1989; Savage, 1989; Stickland 1984; Valdebouze et al., 1980). Table 4, using the results of Freire et al. (1989), illustrates the negative effects due to the inclusion of peas in the diets of young piglets .
Table 4. Weight gain and feed conversion efficiency of piglets fed raw peas for 6 weeks (liveweight 6.7 - 25.2 kg)
Treatment Weight gain
After 3 weeks
After 6 weeks
Feed conversion efficiency
After 3 weeks
After 6 weeks
g/day X g/day
control diet
diet with 15Z peas
diet with 30Z peas
diet with 452 peas
395 100
329b 83
~b 78
79
310 t
311
491
443t
412b
415t
00
90
84
85
1
1
1
1
42"
69b
70b
76b
100
119
120
124
1
1
1
1
54"
69 b C
72b
75 a b
100
110
112
114
Data within the same column with a different superscript differ significantly (P < 0.01)
It is not entirely clear which factor is responsible for the negative effects. Leterme et al. (1989) and Savage (1989) discussed the fact that the presence of trypsin inhibitors may be the most important factor in this respect. An ILOB study also indicated that trypsin inhibitors may play a role in the negative effects. In this study, piglets were fed four diets each containing 30Z from different batches of 'Finale1 peas, with different trypsin inhibitor contents. As a first step, the protein faecal digestibility of the four batches of Finale peas was measured. The batch with the highest trypsin inhibitor content was the lowest in protein digestibility. The same batches were tested in a growth trial with piglets. The diets of the growth trial were balanced for essential amino acids, net energy, vitamins and minerals.
22
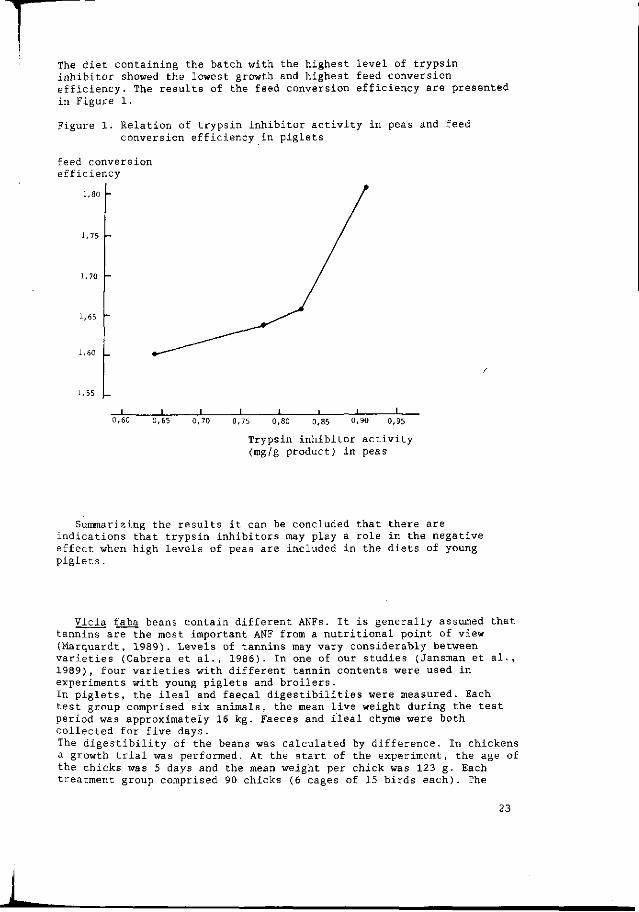
The diet containing the batch with the highest level of trypsin inhibitor showed the lowest growth and highest feed conversion efficiency. The results of the feed conversion efficiency are presented in Figure 1.
Figure 1. Relation of trypsin inhibitor activity in peas and feed conversion efficiency in piglets
feed conversion efficiency
1,75
1,70
1,65
1,60
1,55 [_
0,70 0,75 0,80 0,90 0,95
Trypsin inhibitor activity (mg/g product) in peas
Summarizing the results it can be concluded that there are indications that trypsin inhibitors may play a role in the negative effect when high levels of peas are included in the diets of young piglets .
Vicia faba beans contain different ANFs. It is generally assumed that tannins are the most important ANF from a nutritional point of view (Marquardt, 1989). Levels of tannins may vary considerably between varieties (Cabrera et al., 1986), In one of our studies (Jansman et al., 1989), four varieties with different tannin contents were used in experiments with young piglets and broilers. In piglets, the ileal and faecal digestibilities were measured. Each test group comprised six animals, the mean live weight during the test period was approximately 16 kg. Faeces and ileal chyme were both collected for five days. The digestibility of the beans was calculated by difference. In chickens a growth trial was performed. At the start of the experiment, the age of the chicks was 5 days and the mean weight per chick was 123 g. Each treatment group comprised 90 chicks (6 cages of 15 birds each). The
23
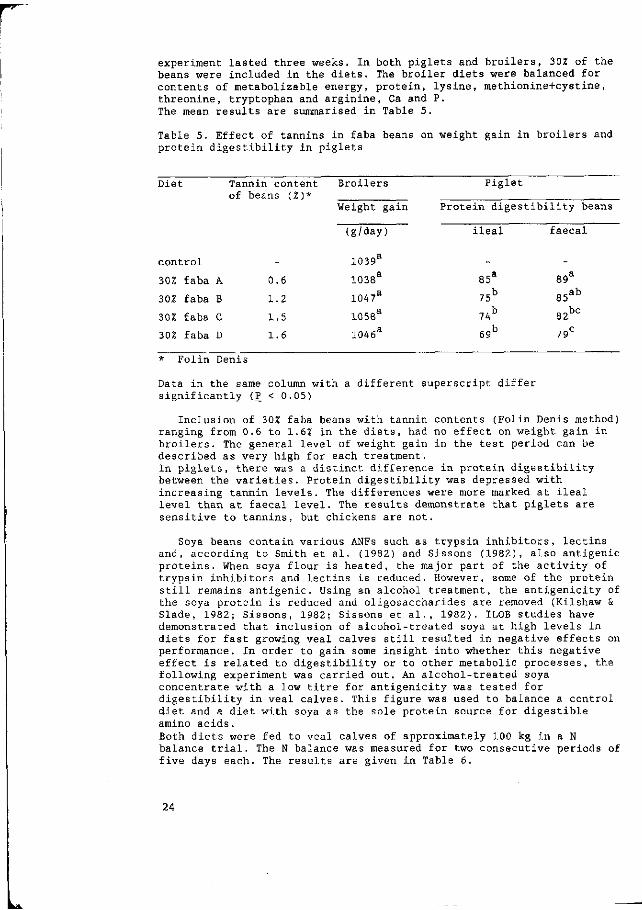
experiment lasted three weeks. In both piglets and broilers, 302 of the beans were included in the diets. The broiler diets were balanced for contents of metabolizable energy, protein, lysine, methionine+cystine, threonine, tryptophan and arginine, Ca and P. The mean results are summarised in Table 5.
Table 5. Effect of tannins in faba beans on weight gain in broilers and protein digestibility in piglets
Diet Tannin content Broilers Piglet
control
302 faba A
301 faba B
302 faba C
302 faba D
Tannin content of beans (2)*
-
0.6
1.2
1.5
1.6
Broilers
Weight gain
(g/day)
1039a
1038a
1047a
1058a
1046a
Protein digestibility beans
ileal faecal
85a
75b
74b
69b
89a
8 5 a b
82 b C
79C
* Folin Denis
Data in the same column with a different superscript differ significantly (P < 0.05)
Inclusion of 302 faba beans with tannin contents (Folin Denis method) ranging from 0.6 to 1.62 in the diets, had no effect on weight gain in broilers. The general level of weight gain in the test period can be described as very high for each treatment. In piglets, there was a distinct difference in protein digestibility between the varieties. Protein digestibility was depressed with increasing tannin levels. The differences were more marked at ileal level than at faecal level. The results demonstrate that piglets are sensitive to tannins, but chickens are not.
Soya beans contain various ANFs such as trypsin inhibitors, lectins and, according to Smith et al. (1982) and Sissons (1982), also antigenic proteins. When soya flour is heated, the major part of the activity of trypsin inhibitors and lectins is reduced. However, some of the protein still remains antigenic. Using an alcohol treatment, the antigenicity of the soya protein is reduced and oligosaccharides are removed (Kilshaw & Slade, 1982; Sissons, 1982; Sissons et al., 1982). ILOB studies have demonstrated that inclusion of alcohol-treated soya at high levels in diets for fast growing veal calves still resulted in negative effects on performance. In order to gain some insight into whether this negative effect is related to digestibility or to other metabolic processes, the following experiment was carried out. An alcohol-treated soya concentrate with a low titre for antigenicity was tested for digestibility in veal calves. This figure was used to balance a control diet and a diet with soya as the sole protein source for digestible amino acids. Both diets were fed to veal calves of approximately 100 kg in a N balance trial. The N balance was measured for two consecutive periods of five days each. The results are given in Table 6.
24

r Table 6. N balance in veal calves of approximately 100 kg live weight
Protein source N balance
g/day SD
Skimmed milk powder
Soya concentrate
42.8
34.4l
a 1.4
2.5
100
80
a - b : significantly different (P < 0.001)
The results clearly show that in spite of the alcohol treatment of soya and balancing the diets on digestible amino acids, the N balance in the soya-fed calves was distinctly lower compared with the control calves fed a diet with skimmed milk powder as the sole protein source. In a study with calves fed the same batch of soya, gut wall damage was observed in spite of a low titre for antigenicity. It is not clear which factor is responsible for the reduced N balance when soya is fed to calves. However, it was found that the analytical method commonly used to determine of antigenic compounds was inadequate. Using an ELISA method, antigenic compounds were still analysed in this soya (see chapter 4: Discussion points on analytical procedures). Moreover, this soya also contained low levels of trypsin inhibitors and lectins. It may be that veal calves are also sensitive to low levels of these factors.
4. Discussion points on analytical procedures
The adequacy of analytical methods for ANFs is essential for research into the effects of ANFs in animals and for studies into reducing the negative effects of ANFs. When the analytical procedures are critically evaluated, one comes to the conclusion that the methods used are often inadequate. •Some of the most important analytical methods for ANF will be discussed.
4.1 Lectins
Haemagglutination of red blood cells is most commonly used to measure lectin activity. This method is based on the the sugar binding properties of lectins to glycoproteins present on the surface of the red blood cells. Due to this binding, the cells agglutinate and the amount of agglutinated cells is used to measure lectin activity. Marquardt et al. (1975) reported that red blood cells from different animal species haemagglutinate differently with lectins from the same sample. It is not clear which type of red blood cells gives the most reliable results. In our studies, we also found differences in the haemagglutination activity of different red blood cells with lectins present in the same sample (Table 7).
As previously discussed by Huisman et al. (1987), important discussion points concerning this method are: - do the glycoproteins in the red blood cells contain the same sugars as
those in the gut wall? In other words: to what extent does the haemagglutination method predict the binding of lectin to the surface of the gut wall?
- not all lectins are pathogenic to the same extent, e.g. lectins from peas are less pathogenic than those in the Phaseolus vulgaris bean. Moreover, one particular seed can contain different types of (iso)lectins. An important question is whether these lectins are all (and to the same extent) pathogenic for the animal.
25

Table 7. Comparison of haemagglutinin activity of three red blood cell origins.
Haemagglutinin activity (units*/mg sample)
Rabbit Pig Horse
Ph. vulgaris cv. Processor - raw 80 - 20 min. steam heated 5
Ph. vulgaris cv. Procol 20
raw soya bean meal 20
20 1
5
1
640 100
n o t determined
0 . 1
* One haemagglutinin activity unit is defined as the smallest amount of sample required for agglutination under test conditions. Haemagglutinin activity is expressed in haemagglutinin units per milligram sample (Valdebouze et al., 1980).
- lectins with one binding site which can originate due to processing, do not haemagglutinate red blood cells. However, they can bind to the gut wall and may therefore cause damage.
These discussion points indicate that the haemagglutination method cannot, in principle be adequate. A first step to improve the analysis of lectins was the development of an ELISA (Enzyme-Linked Immuno-Assay) for lectins. In this method, lectin antibodies are raised and used to determine lectins in the samples. The advantage of the ELISA method compared to the haemagglutination assay is the higher sensitivity and the fact that all lectins are determined. Using the ELISA method, lectins having no specificity to erythrocytes but to leucocytes, and the so-called mono-lectins can also be analysed. One disadvantage of ELISA is that no differentiation can be made between the different types of lectins, including toxic and non-toxic lectins. A very promising assay for determining of different types of lectins is FLIA (Functional Lectin Immuno-Assay), developed by Hamer et al. (1989). This method is based on the ability of lectins to bind to microtitre plates which have been coated with either a carbohydrate matrix or a gut wall brush border membrane preparation. Principally, this method can be adapted to measure, the binding of various lectins from different legumes to the gut wall of the animal.
4.2 Antigenic proteins
Soya proteins have been shown to cause immuno-responses accompanied by disorders in gut function in young piglets and veal calves (Kilshaw and Sissons, 1979; Miller et al., 1984; Seegraber, 1982, 1986; Sissons & Smith, 1976, Toullec & Guilloteau, this volume). It is not entirely clear whether these responses should be attributed to the low levels of lectins present in treated soya or to the fact that soya proteins have been proven to be antigenic. Sissons (1982) reported that antigenicity of soya was eliminated after an alcohol treatment. The antigenicity of the soya tested in that study was measured using a passive haemagglutination method. In this method tanned red blood cells are coated with soya. The coated cells are mixed with blood samples containing soya antibodies. If antigenic compounds are present in the soya, antibodies bind to the coated soya and, as a result, the red blood cells agglutinate (Kilshaw & Sissons, 1979). In one of our studies, we
26

r used this method to determine the'antigenicity in two batches of soya. One batch was split up into two parts. One part was mildly treated with alcohol in order to get a "high-antigen soya" concentrate and the other part treated according to the adequate procedure in order to get a "low-antigen soya" concentrate. The alcohol treatments were carried out by a manufacturer of soya concentrates for veal calves. Both batches were analysed by the manufacturer and by our own laboratory according the passive haemagglutination procedure. The mean analysed values of both laboratories for antigenicity of soya were: a titre of <1 for the "low-antigen soya concentrate" and a titre of 6/7 for the "high-antigen soya concentrate". Both batches were tested for ileal and faecal digestibility, immune response and gut wall damage. There were no differences in ileal and faecal digestibility in either batch, nor was an immune response or gut wall damage observed. These results did not correspond with the analytical data for antigenicity because no immune response or gut wall damage should be expected for the "low-antigen batch". As a follow up, an ELISA method for determining antigenic proteins was developed. Calves were fed soya and soya protein antibodies were raised. The positive serum of these calves was used to determine antigenic active proteins in the soya concentrates. The titres for both batches using the ELISA method were: 7 for the "low-antigen soya" and 8 for the "high antigen soya". These analytical results corresponded well with the results from the animal study. Based on these results, it must be concluded that the passive haemagglutination method is inadequate for determining of antigenic proteins. ,,
4.3 Trypsin inhibitors
One of the problems with this analysis is that there are many different methods applied and, moreover, the units in which the trypsin inhibitor activity is expressed are often different. Therefore, it is difficult to compare the results of one laboratory with those from another. Further point is that mainly bovine trypsin is used to determine trypsin inhibitor activity. Boisen (1989) demonstrated that the use of trypsins from different origins in the assay gives different values. Therefore, it may be that the use of bovine trypsin gives misleading results when related to the biological response in non-bovine species. Thus, trypsins related to the target animal should be recommended for use in the trypsin inhibitor assay.
Another discussion point is that some other factors can inhibit trypsin activity, e.g. tannins. Using the common trypsin inhibitor assay (mostly based on Kakade et al., 1974), the true trypsin inhibitor activity may be overestimated. At the ANF Workshop of 1988 there was a strong feeling that an international committee should be set up to develop standardized protocols for determining trypsin inhibitor activity.
4.4 Tannins
Different methods are described to determine tannin contents in feeds and feed ingredients. Most are based on the colorimetric principle of determination. However, the results of these assays depend on the colouring agents, reaction conditions and the standard agents used. Furthermore, tannins consist of a heterogenic group of chemical substances with assumed distinct nutritional effects. Therefore, the colorimetric methods for tannin determination do not seem to be appropriate. To be able to determine "toxic" tannins, assays based on the HPLC principle seem promising. With this type of assay, tannins can be separated according to their chemical structure.
27
i_

Further research should clarify the relationship between the presence of different types of tannins in diets for pigs and poultry and their nutritional effects. The results of tannin assay should quantitatively predict the nutritional harm of the tannins involved.
5. Animal species differences
The mode of action of ANFs in the animal is mainly studied in rats and chickens. Also, the effects of technological treatments for the inactivation of ANFs are often tested in rats using parameters such as PER, NPU, N balance and weight gain. Based on these results, conclusions are drawn regarding the optimal treatments for inactivating of the ANFs. An important question is whether the results obtained with rats and chickens can be extrapolated to other animal species such as pigs. It is crucial to know to which levels the ANFs have to be reduced in order to minimize their harmful effects. In relation to this, it is also essential to know whether the animals are equal in their sensitivity to ANFs. There is also hardly any information on threshold levels in target animals. So far only a few data have been published on the effects of ANFs in the different animal species. Studies by Combs et al. (1967) and Yen et al. (1977) suggest that rats and piglets respond differently when fed raw soyabeans. Visitpanich et al. (1985) found that the rat and piglet respond differently to the feeding of chickpeas. Liener & Kakade (1980) demonstrated that the response of the pancreas to trypsin inhibitors varies between animal species (Table 8).
Table 8. Relationship between size of pancreas of various species of animals and response of pancreas to raw soybeans of trypsin inhibitor.
Species Size of pancreas Pancreatic (Z of body weight) hypertrophy
Reference
Mouse Rat Chick
Guinea Dog Pig
Human Calf
pig
0.6 0.5 0.4
0 0.21 0.10
0.09 0.06
- 0.8 - 0.6 - 0.6
29 -0.24 - 0.12
- 0.12 -0.08
< - ) v
Schingoethe et al. (1970 ) Kakade et al. (1973) Lepkovsky et al. (1959)
Patten et al. (1973) Patten et al. (1971) Yen et al. (1971)
Kakade et al. (1976)
Observed in young guinea pigs but not in adults
Taken from Long (1961)
Predicted response
Table adapted from Liener and Kakade (1980)
As demonstrated in this table, pancreas hypertrophy due to trypsin inhibitors occurs in smaller animals but not in larger ones such as pigs. However, we should not concluded from these results that the pancreas in larger animals is not stimulated to produce more pancreatic enzymes due to the negative feedback mechanism induced by trypsin inhibitors. In a study by Schuman et al. (1983), increased secretion of
28

pancreas enzymes was found in pigs fed raw soyabean meal. Summarizing this, one must conclude that there is a significant lack
of knowledge regarding the sensitivity of target animals to ANFs. A series of experiments was, therefore, carried out by ILOB-TNO together with the Animal Nutrition Department of the Agricultural University, to study the effects of ANFs in piglets, rats and chickens. In these studies, the Phaseolus vulgaris bean was chosen as representative of a seed containing high levels of•toxic lectins, and the pea (Pisum sativum) as an example of a seed containing moderate levels of trypsin inhibitors and lectins. Moreover, according to Bertrand et al. (1988) the pea lectins are assumed to be non- or low toxic. With Ph. vulgaris beans two experiments were carried out in which 20% raw beans were incorporated into the diets. In the first experiment, the diets were balanced on total protein contents and in the second the diets were balanced on contents of digestible protein (details see: Huisman & Van der Poel, 1989b,c). An experiment was also carried out with diets containing 301 peas. These diets were balanced on digestible protein content (for details see Huisman & Van der Poel, 1989d). The results of these studies are summarized in Table 9.
Table 9. Effects of ANFs in different animal species
Inclusion rate in the diet
Criteria In vivo effects
20£ Ph. vulgaris Weight gain Pancreas weight* Spleen weight* Protein digestibility
30Z Pisum sativum Weight gain Pancreas weight* Spleen weight*
piglets
-10 • I -
a t s
/o + 0 -
0 + 0
c h i c k e n s
/ 0 + 0
N.D.
0 + 0
0 = no effect + = increased weight -/--/ / = decreased weight N.D. = not determined * = 2 of live weight
The results show marked differences between piglets on the one hand and rats and chickens on the other. With Ph. vulgaris beans, piglets did not gain weight, they even lost it. Weight loss was also evident when the diets were balanced on digestible protein, even when extra casein was added to the diets. In rats, there was some weight loss with the diets balanced on total protein, but when balanced on digestible protein there was no weight loss. In chickens there was no negative effect on weight gain due to feeding the beans.
The pancreas weight was increased in rats and chickens but not in piglets. The increased weight of the pancreas in rats and chickens may be related to the presence of trypsin inhibitors in the beans. In both experiments, spleen weight was markedly reduced in the piglets but not in rats or chickens. In the second experiment, the same was observed for the thymus weight. These results may indicate that the immune system in piglets is more sensitive to ANFs than in rats or chickens. Protein digestibility of the diets was distinctly more depressed in piglets compared to rats. With respect to the experiment with peas, a reduction in weight gain was
29
L

observed in piglets. In rats and chickens weight gain was not affected. The pancreas weight was increased in the rats and chickens but not in the piglets.
The results presented here clearly demonstrate that there is a difference in sensitivity between animal species to ANFs present in Ph. vulgaris beans and Pisum sativum. For Vicia faba beans a difference in sensitivity to ANFs was also found between piglets and chickens (see chapter 3, Table 5 ) . Piglets are distinctly more sensitive to ANFs in beans and peas than rats and chickens. Thus, the results obtained with rats and chickens cannot be extrapolated to piglets. Therefore, studies into the effects of ANFs should be carried out in the target animal species.
6. Future prospects
The use of legume seeds in diets for monogastric farm animals is seriously hampered by the fact that these seeds contain ANFs which have negative effects on animal performance. The nutritional value of these seeds can be improved if we are able to reduce these negative effects. However, there is insufficient knowledge about the way these factors act in the target animal. For the future it would be worthwhile to consider the following points:
Studies into the way ANFs act in the animal An important part of present knowledge is based on results obtained in smaller animals such as rats and chickens. Since it has been demonstrated here that these results cannot be extrapolated to other animal species, it is important to carry out more research on the target animal. A difficult problem is that one legume seed contains more than one ANF. Therefore, it is impossible to draw conclusions about a particular ANF when the whole seed is fed. In order to understand the effects of each of the various ANFs, it is necessary to isolate and purify them and to study them separately in target animals. This may not always be realistic because large quantities of purified ANFs would be required which, in many cases, are very expensive. Therefore, animal models should be developed in which only small amounts of ANFs are required to study their effects. Promising models, developed by ILOB in cooperation with the Agricultural University in Wageningen and the University of Utrecht, are pancreas-canulated pigs which survive small intestinal biopsy techniques for studying effects of trypsin inhibitors and lectins. In these models only small amounts of ANF are needed.
Improvement of analytical methods Adequate analyses are essential for research into the effects of ANFs in animals, for evaluating technological treatments in order to reduce the ANF activity, and for plant breeders to develop new varieties low in ANFs. As discussed, there is major concern about the analysis for lectins, antigenic proteins and tannins. For lectins, the newly developed FLIA method looks very promising. Regarding trypsin inhibitors there is a need to standardize assays. Research into the development of adequate analytical methods should be given high priority because other research depends on the validity of these assays.
Threshold levels There is hardly any information concerning threshold levels of ANF in feedstuffs and diets. To improve the quality of feedstuffs, diet formulation, technology and plant breeding it is important to know which levels of ANFs can be tolerated without causing negative effects m the animal. It is questionable whether the ANF content should be reduced to zero. There are indications that in some animals low levels of ANFs can be tolerated without causing negative effects
30

on performance. However, this will depend on the target animal. As already demonstrated, piglets are much more sensitive to ANFs than chickens. The age of the animals will also play a role, e.g. young piglets are more sensitive to ANFs than older pigs. Veal calves may be classified as being very sensitive to ANFs.
Knowledge of threshold levels is also important for technologists. They need to know to which levels ANFs should be reduced. It could be that different products with varying ANF levels can be manufactured for different animal species. For plant breeders, it is important to know which levels of ANFs are acceptable from a nutritional point of view. Whether these levels can be reached by plant breeding is uncertain because seeds with low levels of ANF are generally less disease resistant.
8. References
Bertrand, G., Sève, B., Gallant, D.J. and Tomé, R. (1988). Absence d'effets antinutritionel des lectines de pois, sous forme native ou purifee chez porcelet. Sciences des Aliments, 8, 187-212.
Birk, Y. (1987). Proteinase inhibitors. In: Hydrolytic enzymes, pp 257-305. [A.Neuberger and K. Brocklehurst, editors]. Elsevier, Amsterdam.
Birk, Y. (1989). In: Recent advances of research in antinutritional factors in legume seeds, pp 83-94. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors], Pudoc, Wageningen, The Netherlands.
Boisen, S. (1989). Comparative studies on trypsin inhibitors in legumes and cereals. In: Recent advances of research in antinutritional' factors in legume seeds, pp 118-120. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Bond, D.A. and Smith, D.B. (1989). Possibilities for the reduction of antinutritional factors in grain legumes by breeding. In: Recent advances of research in antinutritional factors in legume seeds, pp 285-296. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Cabrera, A., Lopez-Medina, J. and Martin, A. The genetics of tannin content in faba bean (Vicia faba L.). In: Recent advances of research in antinutritional factors in legume seeds., pp 297-300. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Castaing, J. and Grosjean, F. (1985). Effet de forts percentages de pois de printemps, dans de régimes pour porcs charcutiers, a base de maïs ou d'orge et en complément de tourteau de colza. Journées recherce porcine de France, 17, 407-418.
Combs, G.E., Connes, R.G., Berry, T.H. and Wallace, H.D. (1967). Effect of raw and heated soyabean meal on gain, nutrient digestibility, plasma amino acids and other blood constituents of growing swine. Journal of Animal Sciences 26, 1067-1071.
Donatucci, D.A., Liener, I.E. and Gross, C.J. (1987). Binding of navy bean (Phaseolus vulgaris) lectin to the intestinal cells of the rat and its effect on the absorption of glucose. Journal of Nutrition, 117, 2154-2160.
Fekete, J., Castaing, J., Lavorel, 0., Leuillet, M. and Quemere, P. (1984). Utilisation des pois protéagineux par le porcelet sevré.
Journées recherche porcine en France, 16, 393-400. Freire, J.B., Hulin, J.C., Peiniau, J. and Aumaitre, A. (1989). Effet de
la cuisson-extrusion du pois de printemps sur la digestibilite des aliments de sevrage précoce du porcelet et consequences sur les performances jusqu'à l'abattage. Journées recherche porcine en France, 21, 75-82.
Gallaher, D. and Schneeman, B.0. (1986). Nutritional and metabolic response to plant inhibitors of digestive enzymes. In: Nutritional and
31

Toxicological significance of enzyme inhibitors in foods, pp 167-185. [M.Friedman, editor]. Plenum Press, New York.
Grant, G., More, L.J., McKenzie N.H., Stewart, J.C. and Pusztai, A. (1983). A survey of the nutritional and haemagglutination properties
of legume seeds generally available in the UK. British Journal of Nutrition, 50, 207-214.
Griffiths, D.W. (1981). The polyphenolic content and enzyme inhibitory activity of testas from bean (Vicia faba) and pea (Pisum Sativum) varieties. Journal of the Science of Food and Agriculture. 32, 797-804.
Griffiths and Mosely, 1980. The effect of diets containing field beans of high or low polyphenolic content on the activity of digestive enzymes in the intestines of rats. Journal of the Science of Food and Agriculture, 31, 255 - 259.
Griffiths, D.W. (1984). The trypsin and chymotrypsin inhibitor activities of various pea (Pisum spp) and field bean (Vicia faba) cultivars. Journal of the Science of Food and Agriculture, 35, 481-486.
Grosjean, F. and Castaing, J. (1983). Recherche d'amélioration de la valeur alimentaire du pois d'hiver pour le porc charcutier. Journées recherche porcine en France, 15, 335-346.
Grosjean, F., Castaing, J. and Gatel, F. (1986). Utilisation comparée de différentes variétés de pois et d'une association pois de printemps-féverole par le porc charcutier. Journées recherche porcine en France, 18, 47-56.
Grosjean F. and Gatel, F. (1989). Feeding value of Pisum Sativum for pigs: - influence of technology, - influence of genotype (trypsin inhibitor activity). In: Recent advances of research in antinutritional factors in legume seeds, pp 239-242. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Gumbmann, M.R., Spangler, W.L., Dugan, G.M., Rackis, J.J. and Liener, I.E. (1985). The USDA trypsin inhibitor study. IV. The chronic effects of soy flour and soy protein isolate on the pancreas in rats after two years. Qualitas Plantanum. Plant Foods for Human Nutrition. 35, 275-315.
Hamer, R.J., Koninkx, J.F.J.G., Van Oort, M.G., Mouwen, J.M.V.M. and Huisman, J. (1989). New developments in lectin analysis. In: Recent advances of research in antinutritional factors in legume seeds, pp 30-33. [J.Huisman, A.F.B, van der Poel and Liener, I.E., editors). Pudoc, Wageningen, The Netherlands.
Hove, E.I. and King, S. (1979). Trypsin inhibitor content of lupin seeds and other grain legumes. New Zealand Journal of Agricultural Research, 22, 41-41.
Huisman, J., van der Poel, A.F.B., ten Haaf, L.A.M., Smits, C.H.M, and de Jong, J. (1987). Species differences: different negative effects of antinutritional factors (ANF) in the Phaseolus vulgaris bean in young pigs and rats. Proc. 5th. int. symp. on protein metabolism and nutrition, Rostock, DDR.
Huisman, J., van der Poel, A.F.B., Verstegen, M.W.A. and van Weerden, E.J. (1989). Antinutritional factors (ANF) in pig nutrition. Accepted for publication in: World Review of Animal Production.
Huisman J. and van der Poel, A.F.B. (1989a). Comparison of effects of antinutritional factors in legume seeds. In: Recent advances of research in antinutritional factors in legume seeds, pp 317-327. [J.Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Huisman, J. and van der Poel, A.F.B. (1989b). Comparison of zootechnical characteristics in piglets and rats fed diets containing Phaseolus vulgaris. In publication.
32

r Huisman, J and van der Poel, A.F.B. (1989c). Effect of variable protein
contents in diets containing Phaseolus vulgaris on performance, organ weights and blood parameters in piglets, rats and chickens. In publication.
Huisman, J. and van der Poel, A.F.B. (1989d). Performance and organ weights of piglets, rats and'chickens fed diets containing Pisum Sativum. In preparation.
Jaffe, W.G. and Vega Letta, C.V. (1968). Heat stable growth inhibiting fractions in beans (Phaseolus vulgaris). Journal of Nutrition, 94. 203.
Jaffe, W. (1980). Hemagglutinins, (lectins). In: Toxic constituents of Plant Foodstuffs, pp 73-102. (I.E.Liener, editor). Academic Press, New York.
Jansman, A.J.M., Huisman, J. and Van der Poel, A.F.B. (1989). Faba beans with different tannin contents: ileal and faecal digestibility in piglets and growth in chickens. In: Recent advances of research in antinutritional factors in legume seeds, pp 176-180. [J. Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Kakade, M.L., Rackis, J.J., Mc Ghee, J.E. and Puski, G. (1974). Determination of trypsin inhibitor activity of soy products: A collaborative analysis of an improved procedure. Cereal chemistry, 51 376-382.
Kilshaw, P.J. and Sissons, J.W. (1979). Gastrointestinal allergy t/> soyabean protein in preruminant calves. Antibody production and digestive disturbances in calves fed heated soyabean flour. Research in Veterinary Science, 27, 361-365.
Kilshaw, P.J. and Slade, H. (1982). Villus atrophy and crypt elongation in the small intestine of preruminant calves fed with heated soyabean flour or wheat glutin. Research in Veterinary Science, 33, 305-308.
Kim, Y.S., Borphy, E.J. and Nicholson, J.A. (1976). Rat intestinal brush border membrane peptidases. II. Enzymatic properties, immunochemistry and interactions with lectins of two different forms of the enzyme. Journal of Biological Chemistry, 251, 3206-3212.
King , T.P., Begbie, R. and Cadenhead, A. (1983). Nutritional toxicity of raw kidney beans in pigs. Immunocytochemical and cytopathological studies on the gut and the pancreas. Journal of the Science of Food and Agriculture, 34, 1404-1412.
Liener, I.E. Book: Toxic constituents of plant foodstuffs. Academic press, New York, US.
Liener, I.E., (1986). Nutritional significance of lectins in the diet. In: The lectins: Properties, functions and applications in Biology and Medicine. [I.E. Liener, N. Sharon and I.J. Goldstein, editors). Academic press. New York, US.
Leterme, P., Beckers, Y. and Thewis, A. Inter- and intravarietal and variability of the trypsin inhibitors content of peas and his influence on apparent digestibility of crude proteins by growing pigs. In: Recent advances of research of antinutritional factors in legume seeds, pp 121-124. [J. Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, the Netherlands.
Liener, I.E. (1989). Antinutritional factors in legume seeds: State of the art. In: Recent advances of research in antinutritional factors in legume seeds, pp 6-13. [ J. Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Marquardt, R.R. (1989). Dietary effects of tannins, vicine and convicine. In: Recent advances of research in antinutritional research in legume seeds, [ J. Huisman, A.F.B, van der Poel and I.E. Liener, editors). Pudoc, Wageningen, The Netherlands.
33

Meyer, O.M., Froseth, J.A. and Coon, C.N. (1982). Protein utilization and toxic effects of raw beans (Phaseolus vulgaris) for young pigs.
Journal of Animal Science, 55, 1087-1098. Miller, B.G., Newby, T.J., Stokes, C R . , Hampson, D.J., Brown, P.J. and
Bourne, F.J. (1984). The importance of dietary antigen in the case of postweaning diarrhoea in pigs. American Journal Veterinary Research, 45, 1730-1733.
Mitjavila, S., Lacombe, C , Carrera, G. and Derache, R. (1977). Tannic acid and oxidized tannic acid on the functional state of rat intestinal epithelium. Journal of Nutrition, 107, 2113 - 2121.
Nakata, S. and Kimura, T. (1985). Effect of ingested toxic bean lectins on the gastrointestinal tract in the rat. Journal of Nutrition, 115, 1621-1629.
Poel, A.F.B, van der, (1989). Effect of processing on antinutritional factors (ANF) and protein nutritional value for pigs of dry beans (Phaseolus vulgaris). A review. Animal Feed Science and Technology, submitted.
Pusztai, A., Clarke, E.M.W., Grant, G. and King, T.P. (1981). The toxicity of Phaseolus vulgaris lectins. Nitrogen and immunochemical studies. Journal of the Science of Food and Agriculture, 32, 1037-1046.
Pusztai, A. (1987). Plant lectins-biological functions. Acta Biochemica et Biophysica, Hungary, 99, 355-375.
Pusztai, A. (1989). Biological effects of dietary lectins. In: Recent advances of research in antinutritional factors in legume seeds, pp 17-29.[ J. Huisman, A.F.B, van der Poel and I.E. Liener, editors). Pudoc, Wageningen, The Netherlands.
Rackis, J.J. (1975). Oligosaccharides o£ food legumes: Alpha-galactosidases activity and flatus problems. In: Physiological effects of food carbohydrates, pp 207-222. [J. Allen and J. Heiige, editors). American Chem. S o c , Washington, DC, US
Rackis, J.J. and Gumbmann, M.R. (1981). Protease inhibitors: Physiological properties and nutritional significance. In: Antinutrients and Natural Toxicants in Foods, pp 203-237. [R.L. Ory, editor]. Food and Nutrition Press, Westport, Connecticut, US.
Rackis, J.J., Gumbmann, M.R. and Liener, I.E. (1985). The USDA trypsin inhibitor study. I. Background, objectives and procedural details. Qualitas Plantanum Plant Foods for Human Nutrition, 35, 213-242.
Saini, H.S. (1989). Legume seed oligosaccharides. In: Recent advances of research in antinutritional factors in legume seeds, pp 329-341. [J. Huisman, A.F.B, van der Poel and I.E. Liener, editors). Pudoc, Wageningen, the Netherlands.
Savage, G.P. (1989). Antinutritive factors in peas. In: Recent advances of research of antinutritional factors in legume seeds, pp 342-350. [J. Huisman, A.F.B, van der Poel and I.E. Liener, editors]. Pudoc, Wageningen, The Netherlands.
Savaiano, D.A., Powers, J.R., Castello, M.J. Withaker, J.R. and Clifford, A.J. (1977). The effect of an a-amylase inhibitor on the growth rate of weanling rats. Nutrition Reports International, 15, 443-449.
Schumann, B., Souffrant, W.B., Matkowitz, R. and Gebhardt, G. (1983). Untersuchungen zur endogenen N-Sekretion im Pankreassekret beim Schwein. Wissenschaftlich Zeitschrift Karl-Marx-Universitat Mathematik Naturwissenschaft, 32, 6, 570-575.
Seegraber, F.J. and Morril, J.L. (1982). Effect of soy protein on calves'intestinal absorptive ability and morphology determined by scanning electron microscopy. Journal of Dairy Science, 65, 1962-1970.
34

r Seegraber, F.J. and Morril, J.L. (1986). Effect of protein source in
calf milkreplacres on morphology and absorptive ability of the small intestine. Journal of Dairy Science, 69, 460-469.
Sissons, J.W. and Smith, R.H. (1976). The effect of different diets including those containing soya-bean products, on the passage of digesta movement and water and nitrogen absorption on the small intestine of the pre-ruminant calf. British Journal of Nutrition, 36, 421-438.
Sissons, J.W. (1982). Effects of soya-bean products on digestive processes in the gastrointestinal tract of preruminant calves. Proceedings Nutrition Society, 41, 53-61.
Sissons, J.W., Smith, R.H., Hewitt, D. and Nyrup, A. (1982). Prediction of the suitability of soy-bean products for feeding to preruminant calves by an invitro immunochemical method. British Journal of Nutrition, 47, 311-318.
Stickland, R.G. (1984). Condensed tannins of pea seeds. Plant Science Letters, 34, 403-410.
Torres-Pinedo, R. (1983). Lectins and the intestine. Journal of the Pediatric Gastroenterology and Nutrition, 2, 588-594.
Valdebouze, P., Bergeron, P., Gaborit, T. and Delort-Laval, J. (1980). Content and distribution of trypsin inhibitors and hemagglutinins in some legume seeds. Canadian Journal of Plant Science, 60, 695-701.
Visitpanich, T., Batterham, E.S. and Norton, B.W. (1985). Nutritional value of chickpea (Cicer arietinum) and pigeon pea (Cajanus cajan) meals for growing pigs and rats. I. Energy content and protein quality. Australian Journal of Agricultural Research, 36, 327-325.
Yen, J.T., Jensen, A.H. and Simon, J. (1977). Effect of dietary raw soybean and soybean trypsin inhibitor on trypsin and chymotrypsin activities in the pancreas and in the small intestinal juice of growing swine. Journal of Nutrition, 107, 156-165.
35

RESEARCH INTO THE DIGESTIVE PHYSIOLOGY OF THE MILK-FED CALF
R. Toullec & P. Guilloteau
Laboratoire du Jeune Ruminant Institut National de la Recherche Agronomique 65 rue de Saint-Brieuc, 35042 Rennes Cedex, France
Summary
In the preruminant calf, the activities of many abomasal (chymosin and lysozyme) and intestinal (lactase and aminopeptidases A and N) enzymes are maximum at 2 d of age and subsequently decline. In contrast, the activities of most pancreatic enzymes and of intestinal maltase increase with age, whilst that of pepsin does not appear to change. The use of severely heated skim milk or of replacement protein may have depressive effects on protease secretion. These adaptations as well as gut motility are probably regulated by many gastro-entero-pancreatic hormones whose plasma concentrations change with age and protein source or after feeding. The specific activities of pancreatic enzymes increases with age more rapidly than the relative levels of the corresponding mRNAs. The closure of the oesophageal groove may be impaired by poor management conditions. The use of non-clotting diets results in faster abomasal emptying rate and absorption of fat and protein. Replacement proteins are less completely digested than milk protein in the small intestine and can also induce increased endogenous protein losses. The development of hypersensitivity reactions in the gastro-intestinal tract appears to be one limiting factor in the use of many replacement proteins.
Key-words Preruminant calf, digestive secretions, gut hormones, pancreatic mRNAs, oesophageal groove, abomasal emptying, ileal digestion, dietary allergy.
Introduction
The young calf is normally fed colostrum and then whole milk or milk substitutes. Usually only liquid diets are used for veal calf production until slaughter. Solid food is given early to animals to be weaned, but digestion is essentially preruminant until at least 4 - 5 weeks of age. Thus, in the absence of rumen function the preruminant calf depends on its digestive secretions for the assimilation of dietary fat, carbohydrate and protein. Whole milk and milk substitutes based on skim milk and replacement fat are efficiently digested by healthy preruminant calves.However, with the shortage of skim milk powder in the European Community, alternative sources of protein and carbohydrate are being used more widely. Therefore, it is important to understand how the preruminant calf adapts to these changes. The main aspects of fat, carbohydrate and protein digestion have been reviewed between 1979-1983 (Thivend et al., 1979 ; Roy, 1980 ; Sissons, 1981 ; Toullec et al., 1983). This report will deal with recent advances in digestive secretions, regulation of digestive function, passage of digesta, ileal digestion and protein intolerance.
37
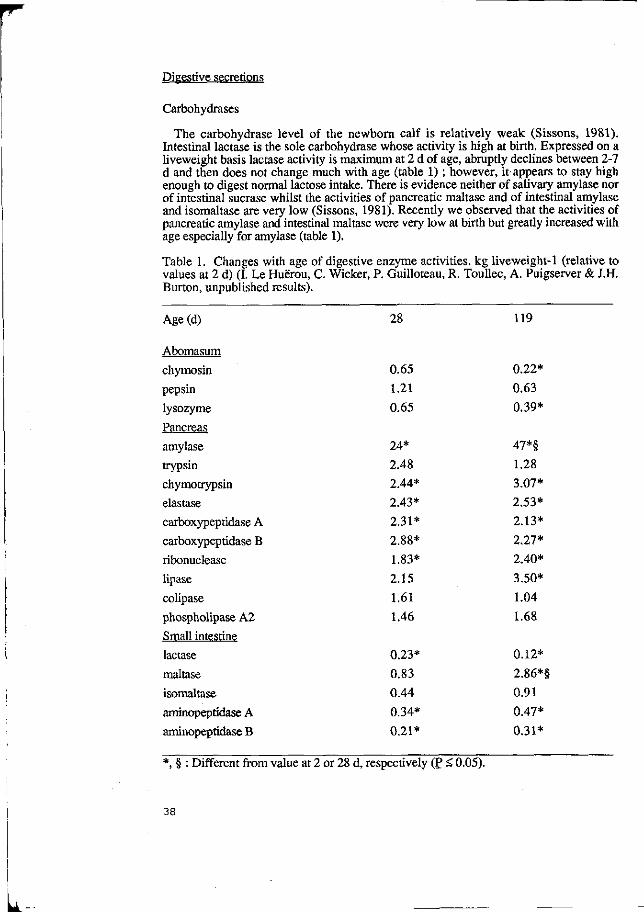
Digestive secretions
Carbohydrases
The carbohydrase level of the newborn calf is relatively weak (Sissons, 1981). Intestinal lactase is the sole carbohydrase whose activity is high at birth. Expressed on a liveweight basis lactase activity is maximum at 2 d of age, abruptly declines between 2-7 d and then does not change much with age (table 1) ; however, it appears to stay high enough to digest normal lactose intake. There is evidence neither of salivary amylase nor of intestinal sucrase whilst the activities of pancreatic maltase and of intestinal amylase and isomaltase are very low (Sissons, 1981). Recently we observed that the activities of pancreatic amylase and intestinal maltase were very low at birth but gready increased with age especially for amylase (table 1).
Table 1. Changes with age of digestive enzyme activities, kg liveweight-1 (relative to values at 2 d) (I. Le Huërou, C. Wicker, P. Guilloteau, R. Toullec, A. Puigserver & J.H. Burton, unpublished results).
Age(d) 28 119
Abomasum
chymosin
pepsin
lysozyme
Pancreas
amylase
trypsin
chymotrypsin
elastase
carboxypeptidase A
carboxypeptidase B
ribonuclease
lipase
colipase
phospholipase A2
Small intestine
lactase
maltase
isomaltase
aminopeptidase A
aminopeptidase B
*, § : Different from value at 2 or 28 d, respectively (P < 0.05).
38
0.65 1.21
0.65
24*
2.48 2.44*
2.43*
2.31*
2.88*
1.83*
2.15
1.61 1.46
0.23*
0.83
0.44
0.34*
0.21*
0.22* 0.63
0.39*
47*§
1.28
3.07*
2.53*
2.13*
2.27*
2.40*
3.50*
1.04
1.68
0.12*
2.86*§
0.91
0.47*
0.31*

Fat hydrolases and bile
Salivary lipase largely contributes to the hydrolysis of triglycerides. It is particularly active on triglycerides containing short-chain fatty acids but splits ester bonds of long chain fatty acids too (Edwards-Webb, 1983). The activity of pancreatic lipase per unit liveweight is low at birth but increases thereafter, whilst the activities of colipase and phospholipase A2 do not appear to vary with age (table 1). Little is known about bile secretion ; however, total bile salts secretion during the first 7 h after a meal was found to increase by 36% when sorbitol was introduced in the diet (Thivend et al., 1984).
Hydrochloric acid and enzymes involved in the digestion of nitrogenous products
Hydrochloric acid secretion in the abomasum, estimated from CI" - Na+ difference in duodenal digesta, was found to rise by 50% between 1-4 weeks of age (Ternouth et al. 1976). In contrast, between 3-32 weeks little change was observed in the quantity of H+
secreted per kg liveweight by abomasal pouches (Guilloteau et al., 1980b). Coagulating activity is mainly due to chymosin. The amount of chymosin per kg liveweight is maximum at 2 d of age and subsequently decreases (table 1). In contrast, pepsin activity does not appear to vary with age. Lysozyme is a muramidase which splits the bonds between N-acetylmuramic acid and N-acetylglucosamine in bacterial peptidoglycan. This enzyme is more abundant in ruminants than in non-ruminants and is mainly concentrated in gastric mucosa (Dobson et al., 1984). In the preruminant calf, lysozyme activity was found to follow a pattern similar to that of chymosin (table 1).
In contrast, in the pancreas most proteolytic enzyme activities as well as ribonuclease activity are minimum at 2 d of age and subsequently increase during the first 2 months (table 1), exhibiting a pattern which is roughly the reverse of that observed for chymosin and lysozyme. Little is known about peptidase activities in the small intestine. The activities of aminopeptidases A and N are maximum at 2 d of age, abruptly decline between 2-7 d and do not appear to change much thereafter.
Many digestive secretions involved in protein digestion may be influenced by management practices and dietary factors (teat versus bucket-feeding, excessive heat-treatment of skim milk, use of non-milk protein) (Toullec et al., 1983). Inconsistent results have been reported on the effects of non-milk protein on gastric secretions. These variations are probably due to different techniques for estimating secretions and treatments used for processing the non-milk protein. In contrast, feeding replacement protein usually results in a reduction of trypsin and chymotrypsin secretions. For example, the activities per kg liveweight present in the pancreatic tissue were found to be depressed by 40% when skim milk was replaced by a soyabean protein concentrate and whey derivatives (Guilloteau et al., 1986a).
Microbial digestion
In calves given whole milk or diets based on skim milk and replacement fat, fat and lactose digestion is apparently finished at the end of ileum and only 2-4% of total nitrogen apparently absorbed in the whole digestive tract disappears in the hindgut (Van Hellemond & Van Weerden, 1973 ; Van Weerden et al., 1977 ; Besle et al., 1980 ; Guilloteau et al., 1986c).Endogenous carbohydrates are still present in the ileal digesta in amounts equivalent to about 7-9% of lactose intake (Besle et al., 1980) and are largely fermented in the hindgut since the apparent digestibility of nitrogen-free-extract is close to 0.99.
39
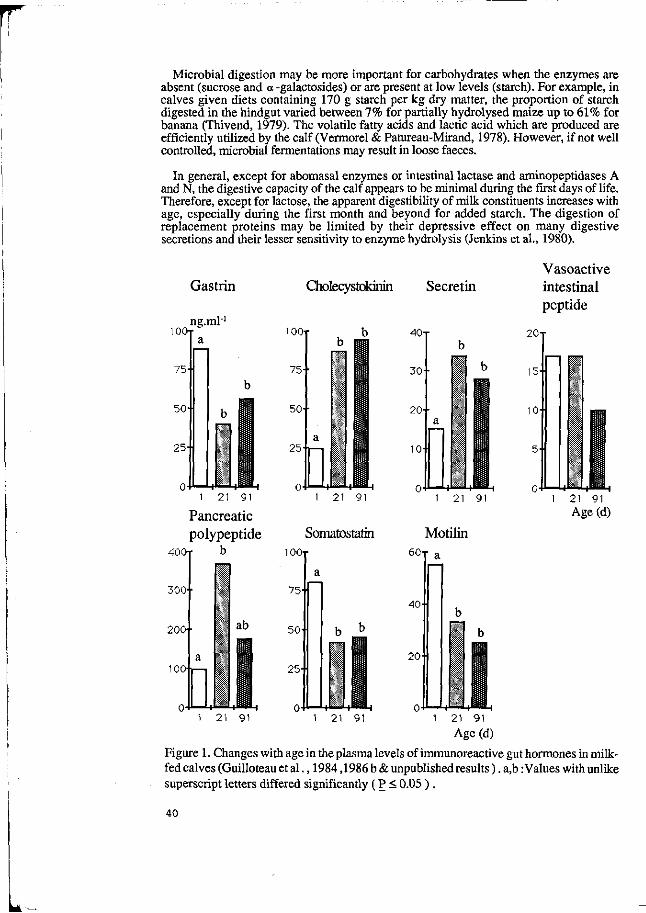
Microbial digestion may be more important for carbohydrates when the enzymes are absent (sucrose and a -galactosides) or are present at low levels (starch). For example, in calves given diets containing 170 g starch per kg dry matter, the proportion of starch digested in the hindgut varied between 7% for partially hydrolysed maize up to 61% for banana (Thivend, 1979). The volatile fatty acids and lactic acid which are produced are efficiently utilized by the calf (Vermorel & Patureau-Mirand, 1978). However, if not well controlled, microbial fermentations may result in loose faeces.
In general, except for abomasal enzymes or intestinal lactase and aminopeptidases A and N, the digestive capacity of the calf appears to be minimal during the first days of life. Therefore, except for lactose, the apparent digestibility of milk constituents increases with age, especially during the first month and beyond for added starch. The digestion of replacement proteins may be limited by their depressive effect on many digestive secretions and their lesser sensitivity to enzyme hydrolysis (Jenkins et al., 1980).
1 21 91
Pancreatic polypeptide
4 0 0 T b
300'
2 0 0 "
100-
0
Cholecystokinin Secretin
100'
75
50
25
04*—H-1 21 91
Somatostatin 1 0 0 T
a
75 '
504
25-
1 21 0'
4 0 T
30-•
20
10
0 1 ' ' i ' i |WW| 1 21 91
Motilin 60
40-
20 '
1 21 0+
Vasoactive intestinal peptide
2 0 T
15-
10-
l ' ' I 21 91 Age (d)
1 21 91
Age (d)
Figure 1. Changes with age in the plasma levels of immunoreactive gut hormones in milk-fed calves (Guilloteau et al., 1984,1986 b & unpublished results ) . a,b :Values with unlike superscript letters differed significantly ( P < 0.05 ) .
40

Regulation of digestive function
Hormonal control
The development of the digestive tract, its secretions and its motor activity are regulated by complex neuro-hormonal mechanisms. For example, acid secretion in the abomasum is stimulated by gastrin and cholecystokinin (CCK) and inhibited by secretin, whilst pepsin secretion appears to be stimulated by secretin (Mc Leay & Bell, 1981). Pancreatic enzyme secretions are favoured by CCK and may be inhibited by pancreatic polypeptide (PP) (Davicco, 1978). Gastrin, CCK and secretin decrease the myoelectric activity of the abomasum which results in a slowing down of abomasal emptying (Mc Leay & Bell, 1981). Gastrin has a trophic effect on the fundic mucosa and the pancreas (Wiener et al., 1987) and CCK on the pancreas (Marx et al., 1987). Somatostatin has numerous inhibitory effects on digestive secretions, the release of the other gastro-entero-pancreatic hormones and gut motility (Newman et al., 1987).
In the preruminant calf large changes with age were observed for the basal plasma concentrations of most assayed gut hormones during the first 3 weeks of life (Guilloteau et al., 1984 & 1986a ; P. Guilloteau, R. Toullec, J.A. Chayvialle & C. Bernard, unpublished results). For example, at 21 d, values were 2.3-, 3.5- and 3.6-fold higher than at birth for secretin, CCK and PP, respectively and 2.3-, 1.6- and 1.9-fold lower for gastrin, motilin and somatostatin (fig. 1). The decrease was particularly rapid for somatostatin since it occured between the first and the second day. No significant trend was apparent thereafter but, in another experiment, the plasma level of somatostatin^ was found to increase by 6.3-fold between 7-70 d of age (P.Guilloteau et al., unpublished results). The early changes of CCK, somatostatin and secretin levels could favour the development of pancreatic tissue and its secretions. The decrease in gastrin level may have resulted in the reduction of chymosin secretion. The decrease of motilin level could be related to the development of gut motility due to enteral feeding. However, little is known about the changes with age of the receptors corresponding to gut peptides. Also, the effect of a gut peptide on the synthesis of the different enzymes in a tissue either depends on the enzyme and/or is altered by other factors : for example, if CCK favours enzyme synthesis in the pancreas, amylase production increases much more with age than the production of the other enzymes.
One hour after feeding a diet based on skim milk powder, the plasma levels of gastrin, CCK and gastric inhibitory peptide (GIP) were increased by 2.6-, 1.8- and 2.3-fold, respectively, compared with the levels before (fig. 2). In contrast, the plasma level of secretin was decreased by 2.1-fold, whilst no systematic trend could be evidenced for motilin, PP and somatostatin. The increase in gastrin and CCK levels after the meal should favour abomasal and pancreatic enzyme secretions. Secretin appears to be particularly involved in the regulation of the pH of small intestine digesta by enhancing pancreatic bicarbonate secretion and reducing abomasal acid secretion. Its release is increased when the duodenal pH is lower than 4.5 (Doyle et al., 1987). This value, which is 2-3 before the meal in the preruminant calf, reaches 5-6 immediately after and 2-3 h are required before it becomes lower than 4.5.
The total replacement of skim milk protein by soya and whey proteins led to a decrease in the basal plasma concentrations of secretin and GIP (fig. 2). After the meal GIP concentration was still reduced whilst CCK concentration was increased. Similar trends were observed for secretin and CCK with fish protein (Guilloteau et al., 1984). These changes could be partially related to the faster abomasal emptying rate of protein and fat. CCK release is favoured by products of fat and protein digestion (Marx et al., 1987) which are more abundant within the intestinal lumen during the first after feeding hours with non-coagulating diets than with milk.As far as the soya diet was concerned, the
41
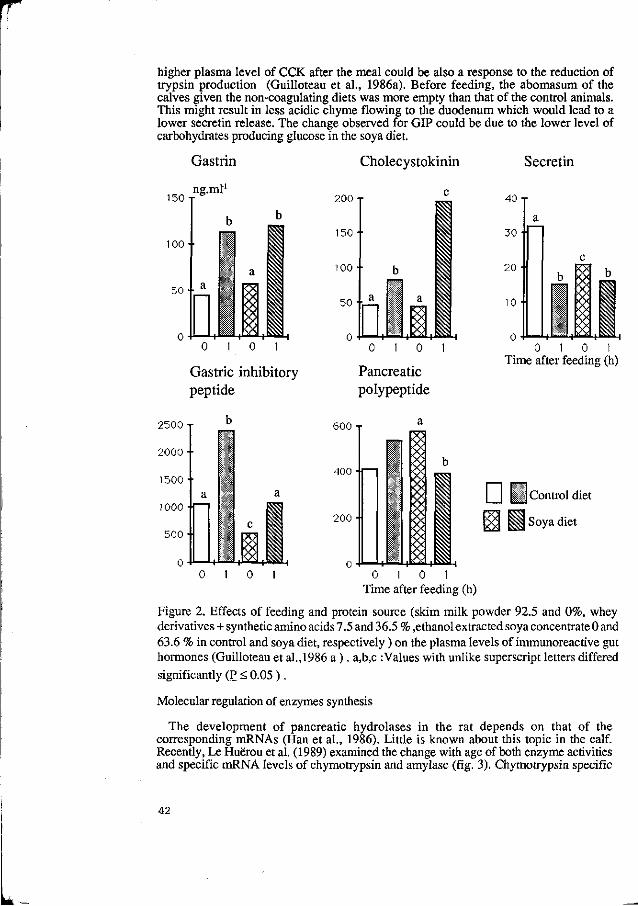
higher plasma level of CCK after the meal could be also a response to the reduction of trypsin production (Guilloteau et al., 1986a). Before feeding, the abomasum of the calves given the non-coagulating diets was more empty than that of the control animals. This might result in less acidic chyme flowing to the duodenum which would lead to a lower secretin release. The change observed for GIP could be due to the lower level of carbohydrates producing glucose in the soya diet.
150
Gastrin
ng.ml1
100
50
I
0 1 0
Gastric inhibitory peptide
Cholecystokinin
200 T
150
100 ••
50
J 0 1 0 1
Pancreatic polypeptide
Secretin
40 T
30
20 • b
1
A o 0 1 0 1
Time after feeding (h)
2500
2000
1500
1000
500
0
• •
a
b
a
i l
600
0 1 0 1
400 -1
200
JA m
I I [ j Control diet
Da §s | Soya diet
0 1 0 1 Time after feeding (h)
Figure 2. Effects of feeding and protein source (skim milk powder 92.5 and 0%, whey derivatives + synthetic amino acids 7.5 and 36.5 % ,ethanol extracted soya concentrate 0 and 63.6 % in control and soya diet, respectively ) on the plasma levels of immunoreactive gut hormones (Guilloteau et al.,1986 a ) . a,b,c :Values with unlike superscript letters differed significantly (P < 0.05 ) .
Molecular regulation of enzymes synthesis
The development of pancreatic hydrolases in the rat depends on that of the corresponding mRNAs (Han et al., 1986). Little is known about this topic in the calf. Recently, Le Huërou et al. (1989) examined the change with age of both enzyme activities and specific mRNA levels of chymotrypsin and amylase (fig. 3). Chymotrypsin specific
42

activity was found to be 20 and 50% higher at 28 and 119 d of age than at birth. In contrast, the relative level of mRNA was 44% lower at 28 d than at birth but had almost regained its birth value at 119 d. Chymotrypsin synthesis appeared to be determined more by some translational and /or post-translational regulation than by the transcriptional control mechanism. Amylase activity, which was very low at birth, was 22 and 43 fold higher, respectively, at 28 and 119 d. During the same time, the level of mRNA increased, but less than the enzyme activity (by 2.7 and 10.2 fold, respectively). Thus, amylase synthesis was probably regulated at the transcription step as well as by translation and/or post-translation events.
Chymotrypsin Amylase 2,0
1 ,0"
0 , 5 -
0,0
50
25 *
*
• i;::.;..;;J ,
*§
- H
*§
/" —1
28 119 28 119 Age (d)
Figure 3. Changes with age of the specific activities ( |_J) and mRNAs levels ( | j | ) of chymotrypsin and amylase ( relative to values at the birth ) in the pancreas of milk-fed calves ( Le Huërou et al, 1989 & unpublished results).*,§ : Different from value at birth and at 28 d, respectively (P< 0.05).
Therefore, new methods can be used to get a better understanding of digestive function. After the present descriptive phase,they should lead to other ways of interpreting and improving digestive adaptation.
Passage of digesta
Closure of the oesophageal groove
Milk and milk-substitutes normally by-pass the rumen thanks to the closure of the oesophageal groove. However some veal calves present problems characterized by inappétence, abdominal distension, a long hirsute hair-coat and abundant clay-like faeces (Tadeu dos Santos et al., 1986 ; Breukink et al., 1988). These symptoms appears 2-4 weeks after the beginning of the fattening period. Using polyethylene glycol as a marker in rumen fistulated calves, Tadeu dos Santos et al. (1986) found that 57% of the milk ingested fell into the rumen in "abnormal" calves, instead of 3% in "normal" animals. Breukink et al. (1988) observed that almost all the milk entered the reticulorumen and was slowly transferred to the abomasum in "abnormal" calves. Other characteristics of the "abnormal" calves were decreases in the pH of the rumen contents (5.1 instead of 6.7,2 h
43

after the meal) and in apparent digestibility (0.73 instead of 0.92 for fat) (Tadeu dos Santos et al., 1986), as well as a shortening of the intestinal villi (Breukink et al., 1988).
In an attempt to simulate these phenomenons, Nunes do Prado et al. (1987 & 1988a) introduced 25% of the milk into the rumen via a canula. The pH of the rumen contents decreased whilst their volatile fatty acids and lactic acid concentrations increased. Lactose disappeared from the rumen within 6 h whilst milk protein still constituted about 40% of total protein in rumen contents 16 h after milk introduction. However, there was impairment neither in oesophageal groove function nor in feed intake even after 3 weeks, perhaps because the canula could be an outlet. According to the observations of Breukink et al. (1988), "abnormal" calves exhibit a drinking behaviour different from that of "normal" calves : they are "gulpers" instead of "sippers". The determinant factor involved in the non-closure of the oesophageal groove is not known but the frequency of the symptoms appears to depend on the management. For example, transportation of the young calves on long distances is unfavourable in this respect. "Abnormal" calves could be treated by emptying the rumen via a stomach tube and by training the calves to suck small amounts of whole milk or good quality milk-substitute after initiating intensive sucking on the herdsman's fingers (Breukink et al., 1988). However, feed intake and liveweight gain of successfully treated calves remained lower than those of normal calves.
Abomasal emptying
In calves given whole milk duodenal digesta contains no intact casein except during the first 10 min after feeding, indicating that considerable hydrolysis of this protein takes place in the abomasum (Yvon et al., 1984). The phosphopeptides fractions of casein chains, except for the caseinomacropeptide resulting from the hydrolysis of K casein, are retained in the abomasum for a longer time than the non-phosphorylated fractions (Yvon et al., 1986). In contrast, whey protein enters rapidly the duodenum ; a-lactalbumin is largely hydrolysed whilst ß-Iactoglobulin is only slightly split before leaving the abomasum (Yvon et al., 1984).
The use of non-casein protein results in a faster abomasal emptying rate for fat and protein due to absence of a clot. This rapid emptying rate could partially contribute to the lower digestibility of most of the diets containing protein substitutes : although not always significant, such a trend has been usually observed for nitrogen and/or fat at the ileal or faecal level by impairing the clotting ability of milk protein-based diets (Toullec et al., 1974 ; Van Weerden et al., 1977 ; Jenkins & Emmons, 1982 ; Strudsholm, 1988 ; Petit et al., 1989). Similarly with diets infused into the duodenum, a rapid flow rate was found to depress nitrogen and fat digestibility (Guilloteau et al.,1981).
Fat and amino acid absorption is strongly influenced by abomasal emptying rate (Beynen & Van Gils, 1983 ; Petit et al., 1987 ; Nunes do Prado et al., 1989c). Therefore, with non-clotting diets, fat and free amino acids accumulate in the blood during the first postprandial hours, although the liver is able to use larger amounts of amino acids (Houlier et al., 1989).
Results of protein digestion in the small intestine
Digestibility of nitrogen and amino acids
Usually, the apparent digestibilities of total nitrogen and amino acid nitrogen are lower at the end of the ileum than values obtained for the entire digestive tract (table 2). However, with a diet containing a pregelatinized pea flour, similar values were recorded at the two levels, probably because large amounts of starch escaped digestion in the small
44
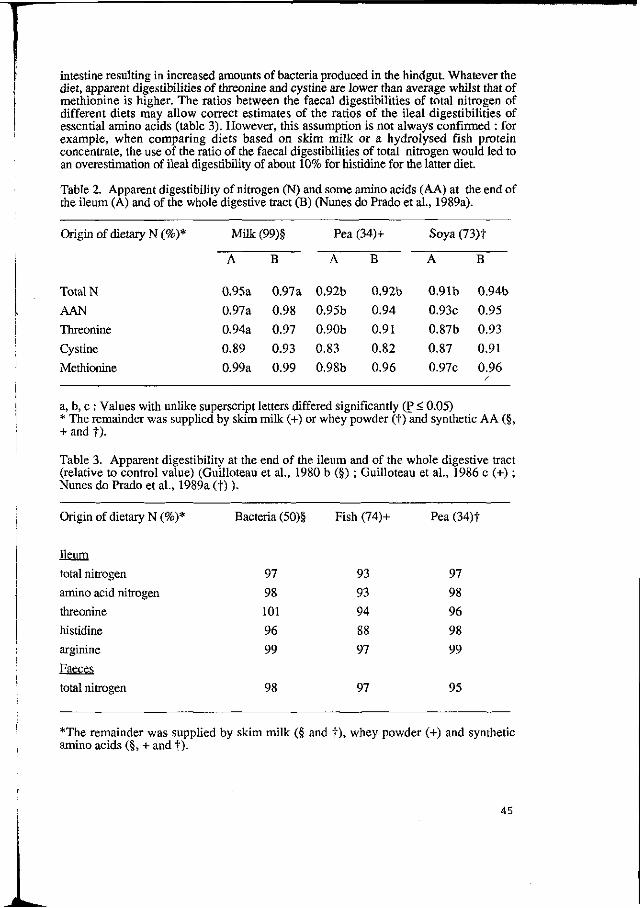
A
0.95a
0.97a
0.94a
0.89
0.99a
B
0.97a
0.98
0.97
0.93
0.99
A
0.92b
0.95b
0.90b
0.83
0.98b
B
0.92b
0.94
0.91
0.82
0.96
A
0.91b
0.93c
0.87b
0.87
0.97c
B
0.94b
0.95
0.93
0.91
0.96
intestine resulting in increased amounts of bacteria produced in the hindgut. Whatever the diet, apparent digestibilities of threonine and cystine are lower than average whilst that of methionine is higher. The ratios between the faecal digestibilities of total nitrogen of different diets may allow correct estimates of the ratios of the ileal digestibilities of essential amino acids (table 3). However, this assumption is not always confirmed : for example, when comparing diets based on skim milk or a hydrolysed fish protein concentrate, the use of the ratio of the faecal digestibilities of total nitrogen would led to an overestimation of ileal digestibility of about 10% for histidine for the latter diet.
Table 2. Apparent digestibility of nitrogen (N) and some amino acids (AA) at the end of the ileum (A) and of the whole digestive tract (B) (Nunes do Prado et al., 1989a).
Origin of dietary N(%)* Milk (99)§ Pea (34)+ Soya (73)t
Total N
AAN
Threonine
Cystine
Methionine
a, b, c : Values with unlike superscript letters differed significantly (P < 0.05) * The remainder was supplied by skim milk (+) or whey powder (f ) and synthetic A A (§, + and f).
Table 3. Apparent digestibility at the end of the ileum and of the whole digestive tract (relative to control value) (Guilloteau et al., 1980 b (§) ; Guilloteau et al., 1986 c (+) ; Nunes do Prado et al., 1989a (t) )•
Origin of dietary N (%)*
Ileum
total nitrogen
amino acid nitrogen
threonine
histidine
arginine
Faeces
total nitrogen
*The remainder was supplied by skim milk (§ and f ), whey powder (+) and synthetic amino acids (§, + and f ).
45
>ria(50)§
97 98 101 96
99
98
Fish (74)+
93 93 94 88 97
97
Pea (3
97 98 96 98 99
95

m m
®
E Arg
m^m Factor 1
Asp Ala Gly
S2,
(S3 ©
©
Leu
He Met
Tyr
Val
His
Glu Thr Ser
u o u
® [10
Pro
© Figure 4. Comparison of the amino acid composition (g/100g assayed amino acids) of protein of ileal digesta (C~J) collected from preruminant calves receiving diets containing different protein sources with that of dietary (| |),endogenous(| | ) and gut bacteria ( ( _ ) ) proteins. Each point is the projection of one protein on the principal plane (1-2) of the factorial correspondance analysis. The seventeen assayed amino acids (Ala, Arg, etc.) are also projected. Dietary protein source and ileal digesta B, C, F, M and P : methanol-grown bacteria, casein, hydrolysed fish, milk and pregelatinized pea flour. S diet: heated soya meal; S digesta: S1 and S2 alcohol-extracted soya concentrate, S3 soya isolate, S4 and S5 allergenic heated soya meal before and after sensitization of the calves (Guilloteau et al, 1980 b & 1986c; Nunes do Prado et al., 1989a, C. Duvaux, J.W. Sissons, R. Toullec & P. Guilloteau, unpublished results) .( B )'• mean composition of pig and sheep faecal bacteria (Mason etal., 1976; Mason, 1979).CM:calfmeconium(Grongnetetal., 1981). AFraxenic lamb faeces (Combe, 1976). Ileal digesta were representative of 24 h or 96 h collection periods, except C, S2, S4 and S5 which corresponded to the first 3h after arrival of feed residues as indicated by a phenol red marker in the effluent.
46

Nature of protein escaping digestion in the small intestine
Assessment of endogenous protein in ileal digesta can be approached by comparing their amino acid composition to those of calf meconium (Grongnet et al., 1981) and faeces of the germ-free lamb (Combe, 1976). Protein of bacterial origin can be evaluated by comparisons with compositions of faecal bacteria isolated from pig (Mason et al, 1976) or sheep faeces (Mason, 1979). The amino acid profiles of the ileal digesta corresponding to three control diets based on skim milk and nine experimental diets containing different replacement proteins were submitted to a factorial correspondence analysis together with dietary, endogenous and bacterial proteins (fig. 4).
With the control diets the ileal digesta were always near the endogenous products, confirming the high true digestibility of milk protein. The protein of Ml, M2 and M3 digesta could be assumed to be representative of the undigested mixture of endogenous and bacterial proteins at the end of ileum in the preruminant calf. By iterative calculation the mean theoretical proportions of endogenous and bacterial proteins in that mixture were found to be 0.68 and 0.32, respectively. All of the other digesta were farther from the area containing the endogenous products and closer to the dietary proteins than Ml, M2 and M3 digesta (fig. 4). However B, C, F, SI and S2 digesta were not near the line joining Ml, M2 or M3 digesta to the corresponding diet. Therefore, the protein from methanol-grown bacteria, casein, fish and alcohol-extracted soya concentrate which escaped digestion were particular fractions whose amino acid composition was largely different from that of the whole dietary protein. In contrast, S4 and S5 digesta were very close to dietary protein, suggesting that large proportions of little degraded soya protein were present at the end of ileum in calves given heated soya meal. Iterative calculations indicated that the proportions of dietary protein could be about 0.58, 0.13, 0.35, 0.18, 0.88 and 0.72 in C, P, S2, S3, S4 and S5 digesta, respectively, the rest being the mixture of endogenous and bacterial proteins obtained with milk protein. Calculations of the distance of %^ (Guilloteau et al., 1980a) between the amino acid composition of the theoretical mixtures of dietary, endogenous and bacterial proteins and the actual composition of digesta showed a good fit for P, S3, S4 and S5 but not for C and S2. These results confirmed that the dietary protein escaping digestion was particular fractions for C and S2 but not for S4 and S5. The proportions of dietary protein with P and S3 were too low to draw conclusions. Replacement proteins are less completely digested than milk protein in the small intestine but can also induce increased amounts of endogenous losses.
Allergenic effects of dietary protein
General aspects
It is well established that unrefined soya products are unsuitable as major sources of protein in calf milk replacers (Sissons, 1982). Even heated soya flour, in which antinutritionnal factors considered to be harmful for monogastric animals (antitryptic factors and lectins) have been inactivated may cause gastrointestinal hypersensitive reactions involving disturbances in gut motility, digesta movement and nutrient absorption. Tissue inflammatory responses were evidenced in histological studies of the intestinal mucosa ; biopsy samples revealed villous atrophy, oedema and lymphocytic inlfiltration (Roy et al., 1977 ; Barratt et al., 1979 ; Seegraber & Morill, 1982 ; Pedersen, 1986). Also, a dramatic increase of gut permeability to ß-lactoglobulin was evidenced in calves sensitized to soya and then challenged with heated soya flour mixed with whole milk (Kilshaw & Slade, 1980). These disorders are probably invoked by immunologically active globular proteins, especially glycinin and ß-conglycinin (Kilshaw &
47

Sissons, 1979 a & b) and perhaps a-conglycinin (J.W. Sissons, personal communication), since the denaturation of these proteins prevents the troubles.
Mechanisms
Detection of IgG, and more rarely of IgE, antibodies specific to soya proteins in the blood of calves given feeds containing soya products has led to speculation that the deleterious effects could have been due to a gastrointestinal allergic reaction (Smith & Sissons, 1975 ; Barratt et al., 1979).
Dietary allergy could result from three different types of hypersensitivity reactions (type I, III or IV) (Gell & Coombs, 1968). The type I reaction or immediate hypersensitivity involves an antigen reacting with mast cells passively sensitized by IgE, inducing the release of histamine or other vasoactive amines which causes tissue damage. The type III reaction or semi-delayed hypersensitivity is due to the formation of immune complexes between the antigen and systemic IgG, complement activation and platelets aggregation, leading to the release of vasoactive amines and proteolytic enzymes by leucocytes. The type IV or delayed hypersensitivity is mediated by T - lymphocytes specific of the antigen which release lymphokines. Porter et al. (1981) reported that 50% of the calves exhibited the type III reaction and only 2 - 3% the type I reaction when given immunologically active products. In contrast, Heppell et al. (1987) observed from cutaneous tests that heated soya flour induced reaginic responses corresponding to type I in all the calves ; however the type IV reaction did not appear to occur.
Disorders similar to those reported in the calf have been evidenced at weaning in piglets with diets containing heated soya flour (Miller et al., 1984). However, piglets rapidly become tolerant to soya whilst preruminant calves do not (Heppell et al., 1987). Failure of localised immuno-defence mechanisms in the small intestine of the calf against the uptake of soya protein has been ascribed to inadequate synthesis of secretory IgA and IgM (Barratt et al., 1979).
The effect of age on the development of allergy symptoms is not well documented. As far as antibody production against soya protein is concerned, Barratt et al. (1979) found higher titres when the soya diet was introduced at 1 week instead of 4 weeks of age. In contrast, Srihara (1984) did not show evidence of any differences when the soya diet started at 6, 18 or 30 d of age.
Effects of soya products on gut motility
Studies of intestinal motility have been made in calves given a series of test feeds containing casein, heated soya flour (HSF) or antigen free soya protein (Sissons et al., 1987). Giving HSF induced a progressive reduction of immediate postprandial abomasal emptying rate and an accelerated passage of digesta in the small intestine, together with a reduced antral motility and an increased number and velocity of migrating myoelectric complexes (MMC) along the intestine. These disorders occured with neither casein nor antigen free soya protein. They appeared to develop together with the anti-soya IgG antibodies. The rapid effect of a test feed containing HSF on abomasal motility (within 30 min) in sensitized calves is indicative of type I anaphylactic hypersensitivity. The disorders induced by sucrose addition in the control diet were distinct from those observed with HSF (table 4) : the number of MMC was reduced, mainly because the duration of the phase of irregular spike activity was increased. Therefore, the disturbances observed with HSF were probably not due to an osmotic effect of its oligosides. The hypothesis of an allergenic reaction was strenghtened by the preventive effect of an antiallergic drug (Nedocromil sodium, Fisons, pic) given for 3 days before a test feed (Duvaux et al., 1988). In calves which had been given an intraperitoneal injection of
48
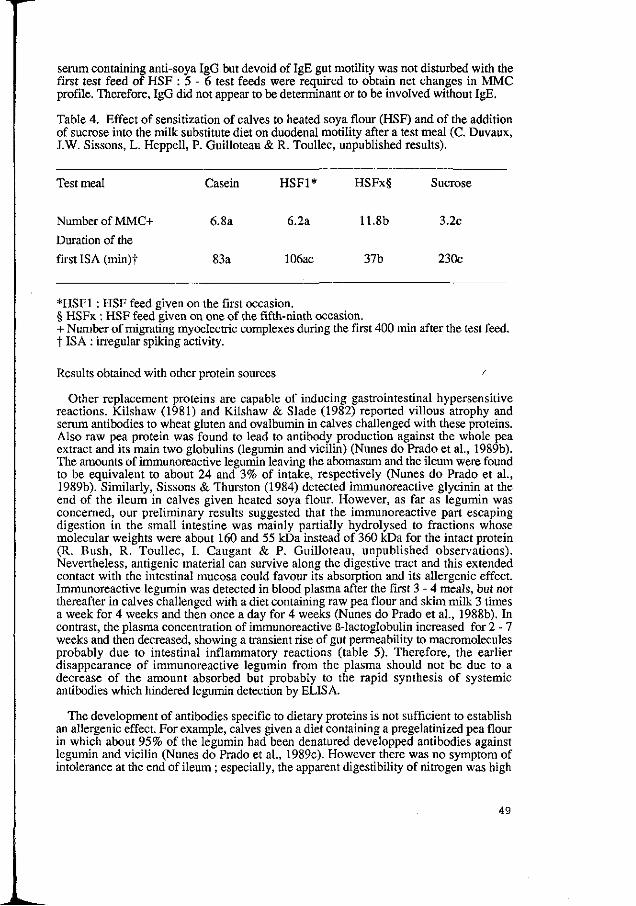
serum containing anti-soya IgG but devoid of IgE gut motility was not disturbed with the first test feed of HSF : 5 - 6 test feeds were required to obtain net changes in MMC profile. Therefore, IgG did not appear to be determinant or to be involved without IgE.
Table 4. Effect of sensitization of calves to heated soya flour (HSF) and of the addition of sucrose into the milk substitute diet on duodenal motility after a test meal (C. Duvaux, J.W. Sissons, L. Heppell, P. Guilloteau & R. Toullec, unpublished results).
Test meal
Number of MMC+
Duration of the
first ISA (min)t
Casein
6.8a
83a
HSF1*
6.2a
106ac
HSFx§
11.8b
37b
Sucrose
3.2c
230c
*HSF1 : HSF feed given on the first occasion. § HSFx : HSF feed given on one of the fifth-ninth occasion. + Number of migrating myoelectric complexes during the first 400 min after the test feed. t ISA : irregular spiking activity.
Results obtained with other protein sources /
Other replacement proteins are capable of inducing gastrointestinal hypersensitive reactions. Kilshaw (1981) and Kilshaw & Slade (1982) reported villous atrophy and serum antibodies to wheat gluten and ovalbumin in calves challenged with these proteins. Also raw pea protein was found to lead to antibody production against the whole pea extract and its main two globulins (legumin and vicilin) (Nunes do Prado et al., 1989b). The amounts of immunoreactive legumin leaving the abomasum and the ileum were found to be equivalent to about 24 and 3% of intake, respectively (Nunes do Prado et al., 1989b). Similarly, Sissons & Thurston (1984) detected immunoreactive glycinin at the end of the ileum in calves given heated soya flour. However, as far as legumin was concerned, our preliminary results suggested that the immunoreactive part escaping digestion in the small intestine was mainly partially hydrolysed to fractions whose molecular weights were about 160 and 55 kDa instead of 360 kDa for the intact protein (R. Bush, R. Toullec, I. Caugant & P. Guilloteau, unpublished observations). Nevertheless, antigenic material can survive along the digestive tract and this extended contact with the intestinal mucosa could favour its absorption and its allergenic effect. Immunoreactive legumin was detected in blood plasma after the first 3 - 4 meals, but not thereafter in calves challenged with a diet containing raw pea flour and skim milk 3 times a week for 4 weeks and then once a day for 4 weeks (Nunes do Prado et al., 1988b). In contrast, the plasma concentration of immunoreactive ß-lactoglobulin increased for 2 - 7 weeks and then decreased, showing a transient rise of gut permeability to macromolecules probably due to intestinal inflammatory reactions (table 5). Therefore, the earlier disappearance of immunoreactive legumin from the plasma should not be due to a decrease of the amount absorbed but probably to the rapid synthesis of systemic antibodies which hindered legumin detection by ELISA.
The development of antibodies specific to dietary proteins is not sufficient to establish an allergenic effect. For example, calves given a diet containing a pregelatinized pea flour in which about 95% of the legumin had been denatured developped antibodies against legumin and vicilin (Nunes do Prado et al., 1989c). However there was no symptom of intolerance at the end of ileum ; especially, the apparent digestibility of nitrogen was high
49

and the amino acid composition of ileal digesta was not much altered (fig. 4), suggesting that the digestion of dietary protein was not impaired. Besides the methods quoted above the preparation of antibodies specific of bovine IgE would provide a tool very helpful to assess allergenic effects (Nielsen & Wilkie, 1977 ; Gershwin & Dygert, 1983 ; Barrat et al., 1985). Also the changes in the ability of the gut to absorb xylose (Seegraber & Morrill, 1986) or mannitol and lactulose (André et al., 1987) could be useful.
Table 5. Effect of the partial replacement of skim milk powder by a raw pea flour on the plasma concentration of immunoreactive ß-lactoglobulin (Nunes do Prado et al., 1989c).
ß-lactoglobulin (ng. ml-1) Diet
initial minimum § maximum+ final
Pea* 212ad 119b 502a 213cd Control 337a - - 97b
* The pea diet was given 3 times per week during the first 4 weeks and then each morning for 4 weeks. § During the first or the second week. + Between the second and the seventh week.
Conclusion
The influence of age and diet on abomasal, pancreatic and intestinal secretions is well documented. However little attention has been given to the effect of diet on certain pancreatic proteases, including carboxypeptidases, and intestinal aminopeptidases. Also few data appear to be available on bile secretion. Studies are developping on the hormonal control of the digestive function and on the molecular control of enzyme synthesis. The closure of the oesophageal groove can be disturbed by poor management conditions. The effect of protein source on abomasal emptying and its consequence on digestion are well understood. Measurements of ileal digestibility can provide useful informations on the protein fractions resistant to digestion and on the availability of amino acids. The development of hypersensitivity reactions in the gastrointestinal tract with certain replacement proteins has been evidenced. Further work is required to improve the methods of diagnosis and to adapt the treatments reducing the allergenicity to the different protein sources.
References
André, C , F. André, L. Colin & S. Cavagna, 1987. Measurement of intestinal permeability to mannitol and lactulose as means of diagnosing food allergy and evaluating therapeutic effectiveness of disodium cromoglycate. Annals of Allergy 59 : 127-130.
Barratt, M.E.J., S.J. Senior, K. May, M. Hall & P. Porter, 1985. Hypersensitivity to dietary components in young farm animals : isolation and partial purification of bovine immunoglobulin E. Research in Veterinary Science 37 :93-96.
50

Barratt, M.E.J., P.J. Strachan & P. Porter, 1979. Immunologically mediated nutritionnal disturbances associated with soya protein antigens. Proceedings of the Nutrition Society 38: 143-150.
Besle, J.M., B. Lassalas & P. Thivend, 1981. Digestion des glucides cytoplasmiques de la féverole par le veau préruminant. Reproduction Nutrition Développement 21 : 629-649.
Beynen A.C. & L.G.M. Van Gils, 1983. Postprandial changes in the levels of lipids, glucose, urea and nonprotein nitrogen in the serum of veal calves fed milk replacers containing either skim milk powder or soyabean protein concentrate. Zeitschrift für Tierphysiologie, Tierernährung und Futtermittelkunde 49,49-56.
Breukink, HJ., T. Wensing, A. Van Weeren-Keverling Buisman, E.G. Van Bruinessen-Kapsenberg & N. A.P.C. De Visser, 1988. Consequences of failure of the oesophageal groove reflex in veal calves fed milk replacer. The Veterinary Quaterly 10:126-135.
Combe, E., 1976. Influence de la microflore intestinale sur la composition en acides aminés des fèces des agneaux. Compte-rendus de la Société de Biologie 170,787-793.
Davicco, M.J., 1978. Régulation du pancréas exocrine du jeune veau. Thesis n° 452. University of Clermont-Ferrand II. 102 p.
Dobson, D.E., E.M. Prager & A.C. Wilson, 1984. Stomach lysozyme of ruminants. I. Distribution and catalytic properties. Journal of Biological Chemistry 259 : 11607-11616.
Doyle, H.R., F. Lluis & P.L. Rayford, 1987. Secretin. In : Thompson, J.C., G.H. Greeley, P.L. Rayford & C.M.Townsend : Gastrointestinal endocrinology. McGraw-Hill, New-York. 223-233.
Duvaux, C, J.W. Sissons, L. Heppell, R. Toullec & P. Guilloteau, 1988. Mécanismes impliqués dans la réaction allergique intestinale aux protéines de soja chez le veau préruminant. Reproduction Nutrition Développement 28 (supplément 1) : 159-160.
Edwards-Webb, J.D., 1983. Digestive lipolysis in the preruminant calf. The abomasal hydrolysis of butter oil, coconut oil, palm oil and tallow. Journal of the Science of Food and Agriculture 34 : 930-936.
Geil, P.G.H. & R.R.A. Coombs, 1968. Classification of allergic reactions responsible for clinical hypersensitivity and disease. In : Clinical aspects of immunology. Blackwell Scientific Publications, Oxford. 575.
Gershwin, L.J. & B.S. Dygert, 1983. Development of a semiautomated microassay for bovine immunoglobulin E : definition and standardization.American Journal of Veterinary Research 44 : 891-895.
Guilloteau, P., J.A. Chayvialle, R. Toullec, J.F. Grongnet & C. Dardillat, 1984. Early life pattern of plasma secretin level in calves. Canadian Journal of Animal Science 64 (supplement) : 100-101.
Guilloteau, P., J.A. Chayvialle, R. Toullec, J.F. Grongnet & C. Dardillat, 1986b. Evolution du taux plasmatique de CCK avec l'âge et le régime alimentaire chez le jeune veau. Reproduction Nutrition Développement 26 : 377-378.
51

Guilloteau, P., T. Corring, J.A. Chayvialle, C. Bernard, J.W. Sissons & R. Toullec, 1986a. Effect of soya protein on digestive enzymes, gut hormone and anti-soya antibody plasma levels in the preruminant calf. Reproduction Nutrition Développement 26:717-728.
Guilloteau, P., P. Patureau-Mirand, R. Toullec & J. Prugnaud, 1980a. Digestion of milk protein and of methanol-grown bacteria protein in the preruminant calf. II. Amino acid composition of ileal digesta and faeces and blood levels of free amino-acids. Reproduction Nutrition Développement 20 : 615-629.
Guilloteau, P., R. Toullec, P. Garnot, P. Martin & G. Brule, 1980b. Influence de l'âge sur les sécrétions de suc gastrique et d'électrolytes chez le veau préruminant. Reproduction Nutrition Développement 20:1279-1284.
Guilloteau, P., R. Toullec, J.F. Grongnet, P. Patureau-Mirand, J. Prugnaud & D. Sauvant, 1986c. Digestion of milk, fish and soya-bean protein in the preruminant calf : flow of digesta, apparent digestibility at the end of the ileum and amino acid of ileal digesta. British Journal of Nutrition 55 : 571-592.
Guilloteau, P., R. Toullec, P. Patureau-Mirand & J. Prugnaud, 1981. Importance of the abomasum in digestion in the preruminant calf. Reproduction Nutrition Développement 21 : 885-899.
Grongnet, J.F., P. Patureau-Mirand, R. Toullec & J. Prugnaud, 1981. Utilisation des protéines du lait et du lactosérum par le jeune veau préruminant. Influence de l'âge et de la dénaturation des protéines du lactosérum. Annales de Zootechnie 30 :443-464.
Han, J.H., L. Rail & W.J. Rutter, 1986. Selective expression of rat pancreatic genes during embryonic development. Proceedings of the National Academy of Sciences USA 83: 110-114.
Heppell, L.M.J., J.W. Sissons, I.J.F. Stobo, S.M. Thurston & C. Duvaux, 1987. Immunological intolerance in calves fed with antigenic soyabean protein. In : Chandra, R.K. : Food allergy. Nutrition Research Education Foundation, St John's, Newfounland. 109-115.
Houlier, M.L., P. Patureau-Mirand, D. Durand, D. Bauchart, G. Bayle & J. Lefaivre, 1989. Influence de la vitesse d'absorption des acides aminés sur leur bilan hépatique chez le veau préruminant. Reproduction Nutrition Développement 29 : in press.
Jenkins, K.J. & D.B. Emmons, 1982. Evidence for beneficial effect of chymosin-casein clots in abomasum on calf performance.Nutrition Reports International 26 : 635-643.
Jenkins, K.J., S. Mahadevan & D.B. Emmons, 1980. Susceptibility of proteins used in calf milk replacers to hydrolysis by various proteolytic enzymes. Canadian Journal of Animal Science 60 : 907-914.
Kilshaw, K.J., 1981. Gastrointestinal hypersensitivity in the preruminant calf. In : Bourne, F.J. : The mucosal immune system. Current topics in veterinary medicine and animal science. Vol. 12. Martinus Nijhoff Publishers, The Hague. 203-223.
Kilshaw, P.J. & J.W. Sissons, 1979a. Gastrointestinal allergy to soyabean protein in preruminant calves. Antibody production and digestive disturbances in calves fed heated soyabean flour. Research in Veterinary Science 27 : 361-365.
52

Kilshaw, K.J. & J.W. Sissons, 1979b. Gastrointestinal allergy to soyabean protein in preruminant calves. Allergenic constituents of soyabean products. Research in Veterinary Science 27 : 366-371.
Kilshaw, K.J. & H. Slade, 1982. Villus atrophy and crypt elongation in the small intestine of preruminant calves fed with soyabean flour or wheat gluten. Research in Veterinary Science 33 : 305-308.
Le Huërou, I., C. Wicker, P. Guilloteau, R. Toullec & A. Puigserver, 1989. Modulation de 1'ARN messager de la chymotrypsine pancréatique au cours du développement postnatal et du sevrage chez le veau. Reproduction Nutrition Développement 29 : in press.
Mc Leay, L.M. & F.R. Bell, 1981. Effect of cholecystokinin, secretin, glucagon and insulin on gastric emptying and acid secretion in the calf. American Journal of Veterinary Research 41:1590-1594.
Marx, M., G. Gomez, J. Lonovics & J.C. Thompson, 1987. Cholecystokinin. In : Thompson, J.C, G.H. Greeley, P.L. Rayford & CM. Townsend : Gastrointestinal endocrinology. McGraw-Hill, New-York. 213-222.
Mason, V.C., 1979. The quantitative importance of bacterial residues in the non-dietary faecal nitrogen of sheep. I. Methodology studies. Zeitschrif für Tierphysiologie, Tierernährung und Futtermittelkunde 41 : 131-139.
Mason, V.C, A. Just & S. Bech-Andersen, 1976. Bacterial activity in the hindgut of pigs. II. Its influence on the apparent digestibility of nitrogen and amino-acids. Zeitschrif für Tierphysiologie, Tierernährung und Futtermittelkunde 36 : 310-324.
Miller, B.G., A.D. Phillips, T.J. Newby, C. Stokes & F.J. Bourne, 1984. Immune hypersensitivity and post-weaning diarrhoea in the pig. Proceedings of the Nutrition Society 43 : 116 A.
Newman, J.B., F. Lluis & CM. Townsend, 1987. Somatostatin. In : Thompson, J.C, G.H. Greeley, P.L. Rayford & CM. Townsend : Gastrointestinal endocrinology. McGraw-Hill, New-York. 286-299.
Nielsen, K.H. & B.N. Wilkie, 1977. Bovine reaginic antibody. II. Preparation of an anti-reaginic immunoglobulin. International Archives Allergy applied Immunology 53 : 174-179.
Nunes do Prado, I., R. Toullec, P. Guilloteau & J.P. Barlet, 1988a. Origine des protéines du contenu de rumen du veau préruminant. Reproduction Nutrition Développement 28: 123-124.
Nunes do Prado, L, R. Toullec, P. Guilloteau & J. Guéguen, 1989a. Digestion des protéines de pois et de soja chez le veau préruminant. II. Digestion apparente à la fin de l'iléon et du tube digestif. Reproduction Nutrition Développement 29 : in press.
Nunes do Prado, L, R. Toullec, P. Guilloteau & J. Guéguen, 1989b. Digestion des protéines de pois dans la caillette et l'intestin grêle du veau préruminant : résultats préliminaires. Reproduction Nutrition Développement 29 : in press.
Nunes do Prado, L, R. Toullec, J.P. Lallès, J. Guéguen, L. Hingand & P. Guilloteau, 1989c. Digestion des protéines de pois et de soja chez le veau préruminant. I. Taux circulants de nutriments, formation d'anticorps et perméabilité intestinale aux macromolécules. Reproduction Nutrition Développement 29 : in press.
53

Nunes do Prado, I., R. Toullec, J.P. Lallès, L. Hingand & J. Guéguen, 1988b. Anticorps contre les protéines alimentaires et perméabilité intestinale aux macromolécules chez le veau préruminant recevant de la farine de pois. Reproduction Nutrition Développement 28 (supplement 1) : 157-158.
Nunes do Prado, I., R. Toullec, Y. Manis & P. Guilloteau, 1987. Effet de l'introduction d'une partie du lait dans le rumen du veau préruminant sur le fonctionnement de la gouttière oesophagienne et la composition du jus de rumen. Reproduction Nutrition Développement 27 :253-254.
Pedersen, H.E., 1986. Studies of soyabean protein intolerance in the preruminant calf. Ph. D. Thesis, University of Reading. 323 p.
Petit, H.V., M. Ivan & G.J. Brisson, 1988. Digestibility and blood parameters in preruminant calves fed a clotting or a nonclotting milk replacer. Journal of Animal Science 66:986-991.
Petit, H.V., M. Ivan & G.J. Brisson, 1989. Digestibility measured by fecal and ileal collection in preruminant calves fed a clotting or a nonclotting milk replacer. Journal of Dairy Science 72 : 123-128.
Porter, P., M.E.J. Barratt & W.D. Allen, 1981. Intestinal response to dietary and bacterial antigens affecting health and performance in the calf. Advances in Experimental Biology and Medicine 137 : 649-661.
Roy, J.H.B., 1980. The calf. 4th edition. Butterworths, London, 442 p.
Roy, J.H.B., I.J.F. Stobo, S.M. Shotton, P. Ganderton & CM. Gillies, 1977. The nutritive value of non-milk proteins for the preruminant calf. The effect of replacement of milk protein by soya-bean flour or fish-protein concentrate. British Journal of Nutrition 38: 167-187.
Seegraber, F.J. & J.L. Morrill, 1986. Effect of protein source in calf milk replacers on morphology and absorptive ability of small intestine. Journal of Dairy Science 69 :460-469.
Sissons, J.W., 1981. Digestive enzymes of cattle. Journal of the Science of Food and Agriculture 32: 105-114.
Sissons, J.W., 1982. Effects of soya-bean products on digestive processes in the gastrointestinal tract of the preruminant calf. Proceedings of the Nutrition Society 41 : 53-61.
Sissons, J.W., H.E. Pedersen, C. Duvaux, S.M. Thurston, S. Starkey & J.A.H. Wass, 1987. Abnormalities in gastrointestinal motility in calves fed antigenic soyabean protein. In : Chandra, R.K. : Food allergy. Nutrition Research Education Foundation, St John's, Newfoundland. 95-108.
Sissons, J.W. & S.M. Thurston, 1984. Survival of dietary antigens in the digestive tract of calves intolerant to soyabean products. Research in Veterinary Science 37 : 242-246.
Smith, R.H. & J.W. Sissons, 1975. The effect of different feeds, including those containing soya-bean products, on the passage of digesta from the abomasum of the preruminant calf. British Journal of Nutrition 33 : 329-349.
54

Srihara, P., 1984. Processing to reduce the antigenicity of soyabean products for preruminant calf diets. Ph D. Thesis, University of Guelph. 162 p.
Strudsholm, F., 1988. The effect of curd formation in the abomasum on the digestion of milk replacers in preruminant calves. Acta Agricultura Scandinavica 38 : 321-327.
Ternouth, J.H., J.H.B. Roy & S.M. Shptton, 1976. Concurrent studies of the flow of digesta in the duodenum and of exocrine secretion of calves. 4. The effect of age. British Journal of Nutrition 36 : 523-535.
Tadeu dos Santos, G., R. Toullec, C. Roger, H. de la Grange & P. Guilloteau, 1986. Caractéristiques digestives des veaux de boucherie s'adaptant mal en ateliers d'engraissement. Reproduction Nutrition Développement 26 : 1217.
Thivend, P., 1979. La digestion des glucides chez le veau sevré. Annales de la Nutrition et de l'Alimentation 33 : 233-245.
Thivend, P., M. Debarre, J. Lefaivre & R. Toullec, 1984. Influence of sorbitol on biliary secretion in the preruminant calf. Canadian Journal of Animal Science 64 (supplement): 102-103.
Thivend, P., R. Toullec & P. Guilloteau, 1979. Digestive adaptation in the preruminant. In : Ruckebusch, Y. & P. Thivend : Digestive physiology and metabolism in ruminants. MTP Press, Lancaster. 561-585.
Toullec, R., J.F. Frantzen & CM. Mathieu, 1974. Influence de la coagulation des protéines sur l'utilisation digestive d'un lait de remplacement chez le veau préruminant. Annales de Zootechnie 33 : 359-364.
Toullec, R., P. Guilloteau, P. Patureau-Mirand & J.W. Sissons, 1983. Digestion and absorption of protein in the preruminant. In : Arnal, M., R. Pion & D. Bonin : Protein metabolism and nutrition. INRA Publications, Paris. 245-261.
Van Hellemond, K.K. & E.J. Van Weerden, 1973. Milk protein substitutes in rations for veal calves. Proceedings of the Nutrition Society 32 : 231-235.
Van Weerden, E.J., J. Huisman & K.K. Van Hellemond, 1977. Verteringsfysiologisch onderzoek. Enkele nitkomsten ten danzien van het verterings proces in het maagdarmkanaal van het meskalf. Landbouwkunding Tijdschrift 89 : 217-224.
Vermorel, M. & P. Patureau-Mirand, 1978. Utilisation des produits terminaux de la digestion des glucides et des lipides par le veau préruminant. In : INRA : Le veau de boucherie. INRA Publications, Versailles. 63-73.
Wiener, I., T. Khalil, J.C. Thompson & P.L. Rayford, 1987. Gastrin. In : Thompson, J.C., G.H. Greeley, P.L. Rayford & CM. Townsend : Gastrointestinal endocrinology. McGraw-Hill, New-York. 194-212.
Yvon, M., J.P. Pélissier, P. Guilloteau & R. Toullec, 1986. Digestion des protéines du lait dans la caillette du veau préruminant. Reproduction Nutrition Développement 26 : 705-715.
Yvon, M., I. Van Hille, J.P. Pélissier, P. Guilloteau & R. Toullec, 1984. In vivo milk digestion in the calf abomasum. II. Milk and whey proteolysis. Reproduction Nutrition Développement 24:835-843.
55

EFFECT OF PORCINE SOMATOTROPIN ON NITROGEN GAIN AND ENERGY METABOLISM IN FATTENING PIGS
M.W.A. Verstegen and W. van der Hel
Ag r i cu l tu ra l Univers i ty Wageningen, Department of An imal Nu t r i t i on and Department of An imal Husbandry
Summary
From many studies i t has become clear that at s imi lar in take of p rote in growing pigs deposit more p ro te in w i th porcine somatotropin (pST). This increase can amount to above 30% ext ra p rote in gain. This increase seems to be more or less independent of energy in take. However i f pST is associated w i th reduced feed in take as occurs o f ten then the increased protein gain w i l l be less. The resul t ing e f fect of pST is increased rate of gain, decreased feed conversion ra t io and increased lean % in the carcass. Associated w i th pST is also a c lear ly increased metabol ic ra te. I t is not c lear to which aspect of energy metabol ism this can be a t t r i bu ted . When we assume maintenance is the only reason the increase is about 20%. I f pST however is not associated w i th maintenance but w i th e f f ic iency the deposit ion of p rote in and/or f a t w i l l require more metabol izable energy. When a l l ex t ra heat is associated w i th the cost of p rote in the e f f ic iency is lowered f rom 0.52 to 0.48. When a l l heat is associated w i th fa t the e f f ic iency is lowered f rom 0.77 to 0.68. I t was also discussed how ac t iv i ty and a l tered thermal demand may be associated w i th ex t ra heat product ion.
Keywords: porcine somatot ropin, prote in ga in, metabol ic ra te , maintenance, par t ia l e f f ic iency.
In t roduct ion
Exogenous admin is t ra t ion of porcine growth hormone increases growth rate and also changes carcass composit ion markedly (Boyd et a l . , 1987; Ether ton et a l . , 1987; McLaren et a l . , 1987). Especial ly in pigs between 50 and 105 kg these studies have been made. Most of these studies showed that e f fect of pST
- increased rate of gain provided that feed in take increased - decreased feed in take - increased the % of lean tissue in the carcass - reduced the ra t io of feed to gain
However not a l l studies showed this e f fect (see Kanis et a l . , 1990). More recent ly Campbel l et a l . (1988) showed that also in animals f rom 25-55 kg exogenous admin is t ra t ion improved performance. Most ly the ear l ier studies have been made on castrated male pigs. In la ter studies w i th animals of d i f fe ren t sexes a s imi lar increase in performance was found. Campbel l and K ing (1989; personal communicat ion) showed that w i th appl icat ion of somatotropine to pigs fed various feeding levels the feed/gain rat ios between sexes became s imi lar . I t was also shown that in general feed in take is d iminished in a ra t io proport ional to the dose
57

of application. It has also been derived that maintenance feed probably increased (Campbell et al., 1988). In this paper we will discuss some aspects of energy metabolism and N gain.
In this context we will focus on the effects of porcine somatotropin on protein gain in relation to
- pST applied - sex - energy intake - protein intake.
Moreover we will focus on aspects of energy metabolism in somatotropin t reated pigs.
Protein gain
One of the major aspects of pST application is the change in carcass composition in t reated animals. Campbell et al. (1988) noted that growth hormone reduced body and carcass fat at an increased protein and water concentration of pigs at 55 kg. In Table 1 some of their data are given.
Table 1. Effect of porcine growth hormone (pGH) administration on the composition of the carcass and the empty body at 55 kg (from Campbell et al., 1988).
Feeding level
Ad lib.
80%
60%
Dose pGH g kg-ld-1
0 100
0 100
0 100
Rate of gain g/d-1
905 1052
670 842
543 681
Carcass water
%
53.8 59.3
56.0 63.3
58.9 64.3
protein %
14.5 16.1
15.3 17.0
16.2 17.3
fat %
28.0 20.0
23.9 16.0
20.5 14.6
Empty body water protein
% %
56.6 14.6 61.7 16.2
57.7 15.4 64.2 16.9
60.2 16.3 65.6 17.3
fat %
25.8 18.8
23.1 15.3
19.6 14.1
Data on Table 1 show that since rate of gain was increased also rate of body protein deposition in these pigs was increased. As a consequence lean % can be much higher in pST t reated animals (Kanis et al., 1989). It has been shown that the capacity to sustain N gain is highest in boars compared to sows. This in turn is higher then in castrated males. From a recent experiment Campbell et al. (1989) reported that from 60 to 100 kg boars gained 32% more protein then sows and 42% more then castrated males. They found that after administration with pST during this period protein deposition was raised to the same absolute level regardless of sex .
58
Table 2. Protein and fat accretion of animals of 31 days from 60 kg onwards (Campbell et al., 1989).
Boars
Gilts
Castrated males
Treatment
control 100 mg/kg d
control 100 mg/kg d
control 100 mg/kg d
Fat gain g/d
317 203
411 185
462 223
Protein gain g/d
196 238
148 235
139 225
In their experiment animals received ad lib. feed. From this it is not clear how protein gain would have been at different intake levels and in different genotypes. These results suggest that the differences between sexes diminish with pST. It can be derived that probably also different genotypes may react differently on pST. In an experiment of the Wageningen group (see Verstegen et al., 1989) pigs of three genotypes Duroc, Pietrain and a crossbred between Dutch Landrace and Dutch Yorkshire (DYxDL) were t reated with pST from 55 kg onwards. During the period of application animals were placed on metabolism cages during 10 weeks and N balances were measured during the last 6 weeks. Animals received a feeding level of about 2.6 t imes maintenance. The diet is given in Table 3.
Table 3. Composition of d ie t a )
Ingredient
Yellow corn Barley Peas Extracted soybean meal (48.5% crude protein) Corn gluten feed Tapioca Citrus pulp Sugarcane molasses Animal meal Animal fat Mono calcium phosphate Salt Limestone Mineral + Vitamin mixture^) Mixturec)
% by weight
3.7 11.8 10
21 5
35 0.7 5 4 1.2 0.9 0.2 0.2 0.5 0.8
a)Calculated: Dig. lysine, 0.92%; Dig. methionine + cystine, 0.55%; net energy for pigs, 9.0 MJ/kg; analysed dm 88.6%; crude protein, 19.4%;
Ca, 0.90%; P, 0.69%.
Results on ra te of gain and feed to gain ratio are presented in Table 4.
59

Table 4. Ra te of gain (kg/d) and feed conversion of animals of three genotypes at constant feeding level during 6 weeks (5-10 weeks after initial administration).
Rate of gain Feed/gain
Pietrain
Duroc
DYxDL
C T C T C T
.87b
.99a
.72d
.80C
.92b l.Ofa
2.94b 2.79c 3.40a 3.19a 2.90b 2.70d
* Different superscripts in a column means significantly different (P^O.01).
As an average animals with pST increased their rate of gain with about 100 g per day. Since feed intakes were similar ratio of feed to gain was decreased with about 0.17. Results on performance as presented in Table 4 do not give information on al teration of body composition. In the same experiment therefore N gain was also measured. In Table 5 the results in N gain has been recalculated to protein. Since energy balances were also made fat gain could be derived from the data on intake of metabolizable energy, heat production and energy associated with protein gain. This can be done as follows:
Metabolizable energy (ME) = Heat production (H) + Retained energy (RE)
Moreover retained energy (RE) = energy retained in protein + energy retained in fat. Since energy retained in protein can be obtained from N balance as Energy retained in protein = N balance x 6.25 x 23.7 the resulting fat gain can be calculated. Results have been given in Table 5. The results show that there may be differences between genotypes.
Table 5. Effect of pST on ra te of gain in protein and fat (g/d).
Genotype Treatment
Control pST
Control pST
Control pST
Protein (g/d)
142 191
119 166
168 210
Fat (g/d)
263 249
275 215
280 218
Pietrain
Duroc
DYxDL
On an average protein gain was increased with about 40 g/d and fat gain was decreased with about 40 g/d in these experiments. Detailed data on this have been presented by Van der Hel et al. (1988). Under normal circumstances increases in protein accretion of the magnitudes shown in Table 5 would be expected to require concomitant increases in the levels of dietary protein and amino acids. By using the factorial approach Boyd et al. (1988) predicted an almost two-fold increase in the dietary lysine requirement of pigs administered pST between 55 and 100 kg liveweight. However, this requires accurate information on the
60
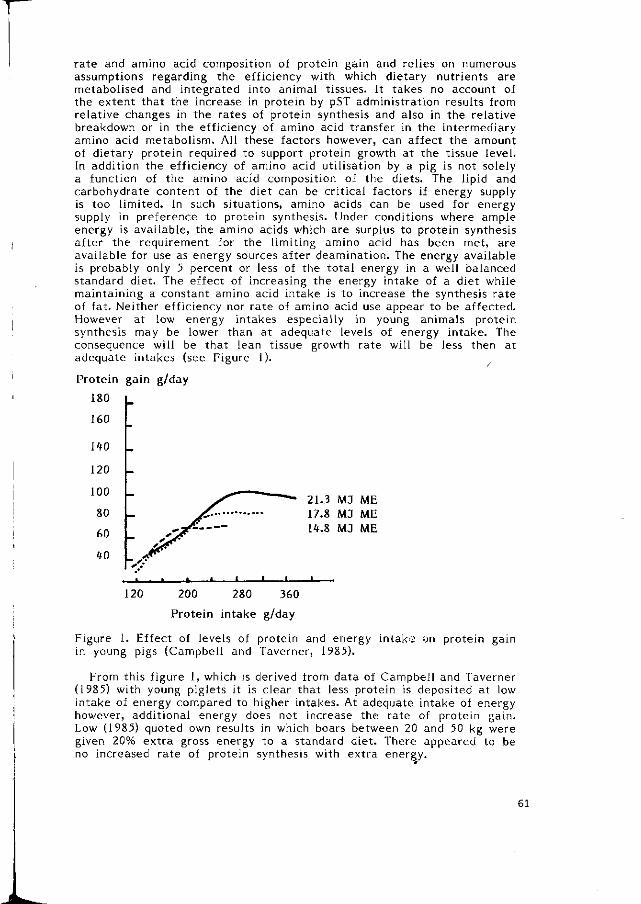
rate and amino acid composition of protein gain and relies on numerous assumptions regarding the efficiency with which dietary nutrients are metabolised and integrated into animal tissues. It takes no account of the extent that the increase in protein by pST administration results from relative changes in the rates of protein synthesis and also in the relative breakdown or in the efficiency of amino acid transfer in the intermediary amino acid metabolism. All these factors however, can affect the amount of dietary protein required to support protein growth at the tissue level. In addition the efficiency of amino acid utilisation by a pig is not solely a function of the amino acid composition of the diets. The lipid and carbohydrate content of the diet can be critical factors if energy supply is too limited. In such situations, amino acids can be used for energy supply in preference to protein synthesis. Under conditions where ample energy is available, the amino acids which are surplus to protein synthesis after the requirement for the limiting amino acid has been met, are available for use as energy sources after deamination. The energy available is probably only 5 percent or less of the total energy in a well balanced standard diet. The effect of increasing the energy intake of a diet while maintaining a constant amino acid intake is to increase the synthesis ra te of fat. Neither efficiency nor rate of amino acid use appear to be affected. However at low energy intakes especially in young animals protein synthesis may be lower than at adequate levels of energy intake. The consequence will be that lean tissue growth ra te will be less then at adequate intakes (see Figure 1).
Protein gain g/day
180
160
140
120
100
80
60
40 _ A
21.3 MJ ME 17.8 MJ ME 14.8 MJ ME
-L. 120 200 280 360
Protein intake g/day
Figure 1. Effect of levels of protein and energy intako in young pigs (Campbell and Taverner, 1985).
<jn protein gain
From this figure 1, which is derived from data of Campbell and Taverner (1985) with young piglets it is clear that less protein is deposited at low intake of energy compared to higher intakes. At adequate intake of energy however, additional energy does not increase the rate of protein gain. Low (1985) quoted own results in which boars between 20 and 50 kg were given 20% extra gross energy to a standard diet. There appeared to be no increased rate of protein synthesis with extra energy.
61
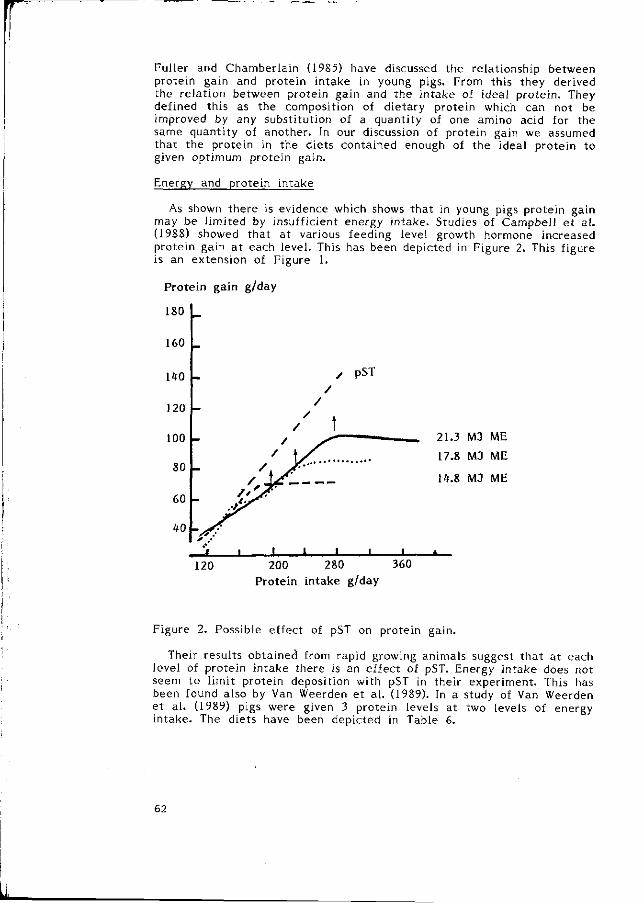
Fuller and Chamberlain (1985) have discussed the relationship between protein gain and protein intake in young pigs. From this they derived the relation between protein gain and the intake of ideal protein. They defined this as the composition of dietary protein which can not be improved by any substitution of a quantity of one amino acid for the same quantity of another. In our discussion of protein gain we assumed that the protein in the diets contained enough of the ideal protein to given optimum protein gain.
Energy and protein intake
As shown there is evidence which shows that in young pigs protein gain may be l imited by insufficient energy intake. Studies of Campbell e t al. (1988) showed that at various feeding level growth hormone increased protein gain a t each level. This has been depicted in Figure 2. This figure is an extension of Figure 1.
Protein gain g/day
180 |_
160
140
120
100
80 - / ,
60
40
pST
/.'
/ /
/ t - 21.3 M3 ME
17.8 M3 ME
14.8 M3 ME
I -J- J _ J _
120 200 280 360 Protein intake g/day
Figure 2. Possible effect of pST on protein gain.
Their results obtained from rapid growing animals suggest that at each level of protein intake there is an effect of pST. Energy intake does not seem to limit protein deposition with pST in their experiment. This has been found also by Van Weerden et al. (1989). In a study of Van Weerden et al. (1989) pigs were given 3 protein levels at two levels of energy intake. The diets have been depicted in Table 6.
62
Table 6. Dietary composition in triais of Van Weerden et al. (1989).
Groups Treatment Composition of feed Net energy Crude protein Lysine
MJ/kg (%) (%)
I II
III IV
V VI
VII VIII
IX X
Control pST t reated
Control pST t reated
Control pST t reated
Control pST t reated
Control pST t reated
9.1 9.1
9.1 9.1
9.1 9.1
10.8 10.8
10.8 10.8
16.0 16.0
18>.0 18.0
20.0 20.0
18.0 18.0
20.0 20.0
0.80 0.80
0.90 0.90
0.98 0.98
0.92 0.92
1.07 1.07
The results of their study showed that N gain intake increases N gain and at each level of protein intake this increase is of similar magnitude regardless of energy intake (Table 7). /
Table 7. N balance (g/day) in experiments with various lysine and energy levels (Van Weerden et al., 1989).
Diets Control pST pST as % Energy of control
normal energy, 23.8 29.4 124 Low 16% protein
normal energy 25.4 33.0 130 Medium 18% protein
normal energy 26.9 35.9 134 High 20% protein
high energy 25.4 33.8 133 Medium IS% protein
high energy 26.3 35.6 135 High 20% protein
Thus at each level of increased protein intake we can expect an increased protein gain with pST. This means that N gain from N in feed results in increased efficiency with use of pST. Van Weerden and Verstegen (1989) showed that pST may reduce N output (N feed - N retained in body) in growing finishing pigs (60-110 kg) in the order of 20% due to increased efficiency.
In conclusion it can be derived that the application of pST to growing and finishing pigs
- improves N gain relative of N in feed, this improvement occurs at each level of N intake.
- improves N in gain relative independent on energy in feed
63

- improves N gain to such extent that differences between sexes diminish.
As a result Jean % in the carcass may be increased by 0 to 4% (Kanis et al., 1989).
Energy metabolism
In the tr ials reported by Campbell et al. (1988) they calculated by regression of energy retained on intake of digestible energy that maintenance requirement in energy was increased with pST. This suggests that metabolic rate was increased after application of pST. In the study of Van der Hel et al. (1988) energy metabolism was measured and in Table 8 the heat production and retained energy are given.
Table 8. Heat production (H) and retained energy (RE) in animals of 55 to 100 kg and t reated with pST (T) or sham injected (C) (Verstegen et al., 1989).
Pietrain
Duroc
Crossbred
C T
C T
C T
Heat production kJ/kgO.75
632 653
611 669
623 682
(H) Retained energy (RE) kJ/kgO.75
448 427
469 418
469 427
It appears that in all three genotypes heat production is increased with pST. A summary of the results on energy balances is given in Table 9.
Table 9. Energy metabolism (kJ/kg) and pST in growing pigs of 55-100 kg. C = sham injected, pST = t reated (Van der Hel e t al., 1988).
C pST
Intake (ME) Energy gain RE Heat production
1092 470 622
1084 416 668
At similar intake heat production is as an average increased with 44 kJ. Consequently retained energy of the same amount of feed intake will be decreased with 44 kJ. It is not clear why increased energy expenditure is associated with the application of somatotropine to animals.
According to Machlin's work (1972) with pigs, pST directs nutrients to the muscle during growth. In understanding the metabolic effect of somatotropine on metabolism it is important to note that any intervention that alters the ra te of major route of nutrient (or energy) storage will also alter the rate of storage pathways (Reeds, 1987). Therefore it is important to distinguish between the mechanisms which are responsible for a reduction in e.g. fat deposition that might arise from a specific stimulus of protein deposition and that appear to stem primarily from an increase in energy expenditure. It is not clear how the increase in serum glucose and insulin level after somatotropin applications in pigs (Etherton et al., 1986a and Boyd, 1987) is associated with the increased energy expenditure.
64

Some of the possibilities and options to explain this increase have been given by Verstegen et al. (1989). We first assumed that animals are kept at thermoneutral conditions. Heat (or thermal losses) is produced as a result of the many metabolic processes occurring within the animal, the extent to which it occurs is not only characteristic of the animal per se but is dependent upon nutritional, productive, environmental and other related factors. Thus no simple system can be used to describe the contribution made by the various factors to metabolic heat production.
In practice, and in order to facili tate the application of energy evaluation systems, it has been customary to partition the thermal losses into those associated with maintenance, on the one hand, and those resulting from the deposition of tissue or products formed within the body, on the other. The former, the maintenance heat loss, represents an animal in a s tate of energy equilibrium. This means that it is neither losing nor gaining energy, so that the intake of dietary energy exactly balances the animal's heat output. The heat arising from the accretion of tissues or products within the body represents the amount of work done in their deposition and varies with the nutritional s ta te of the animal, so that the higher the level of feed intake, the higher the rate of tissue accretion and the greater the heat output associated with these processes, i.e. the heat increment of feeding (which is synonymous with "specific dynamic action" and "dietary induced thermogenesis"). Both the maintenance heat loss and heat produced as a result of tissue deposition are influenced by a number of factors. As the ra te of the animal's heat loss increases at any given level of feed intake, there will be a reduction in the ra te at which energy is retained and hence a change in the energetic efficiency of growth. For practical purposes it is important to know to what extent heat production varies in relation to those factors which influence it, since this determined the extent and efficiency of energy utilisation.
It is usual to express the energy retained by an animal as a function of its bodyweight and the quality and quantity of the ration provided. The food can then be described in its capacity to sustain maintenance (MEm) and to promote energy gain (RE). This is i llustrated in Figure 3, where metabolizable energy (ME) intake, i.e. the gross energy of the feed minus the energy lost in faeces, urine and methane, is related to energy retained (RE).
We assume that partial efficiency (tgC£- in Figure 3) was not a l tered with increase in ME intake. The inefficiency I-tgoi is for heat increment. Above the basal or fasting level of metabolism, each increment in ME is associated with an increment in heat production (H). However, the increment in ME exceeds the increment in H so that the animal has the capacity to retain energy (RE), although RE only becomes positive a t intakes above the maintenance energy requirement, that is MEm = zero RE. The efficiency with which energy is retained is equal to RE/(ME-MEm). As we are working with growing animals feed intake is normally above maintenance. We assumed linearity between energy retention and intake of metabolizable energy. Energy retention can be described as:
RE = kg*ME - b in which: RE = retained energy
ME = metabolizable energy kg = dER/(ME-MEm) b/k g = maintenance requirement (MEm)
This procedure was applied by Campbell et al. (1988). They regressed ER on energy intake. They used digestible energy instead of metabolizable energy. This is justified because there is a nearly fixed ratio of digestible
65

Intake of metabolizable energy
Figure 3. The relation between energy retention (kJ /kg 0 - 7 5 per day) and metabolizable energy (kJ /kg 0 - 7 5 per day) in the pig at thermoneutral condition, tg is partial efficiency above maintenance. MEm is metabolizable energy needed for maintenance.
to metabolizable energy above maintenance. Since there is a very high correlation between metabolizable energy and digestible energy (DE) the formula above also holds for DE. The value (1-kg) represents the increase in energy expenditure with feed intake above maintenance (MEp). The efficiency is representative for total energy gain from ME and does not account for differences in its composition, reflected in the rates of protein and fat deposition. Separate estimates of the energetic efficiency of protein deposition (kp) and fat deposition (k(S) can be calculated with the respective heat increments being MEp r o t . (1-kp) and MEfat ( 1 - kp . Thus the total thermal loss associated with the metabolism of dietary energy is the sum of the energy costs of maintenance (MEm) and the heat increment associated with the deposition of tissue, calculated as MEp
(1-kg) or partitioned into that associated with protein deposition, ME p r o t . (1-kp) and fat deposition, MEfat (1-kjp. There are thus three options for increased metabolis rate - a l tered maintenance (MEm) - a l tered partial efficiency of protein deposition (kp) - a l tered partial efficiency of fat deposition (k^). In all three cases the result should be a reduced retained energy as shown in Figure 4. Maintenance requirement increased metabolic rate may result in an increased maintenance requirement. Maintenance requirement (MEm) was calculated as MEm = ME i n t a ke - En in fat r e t a i n ed / ^ - En in prot. r e t a ined /kp We used efficiency given by ARC (1981) to calculate MEm. The principle of this calculation is also depicted in Figure 5. It has been assumed that the efficiencies of conversion of ME above maintenance into energy retained in fat and protein are not a l tered. Resulting maintenance requirement is calculated as follows:
Controls: MEm = 393 k3 ME/kg"-^ pST treated: MEm = 491 k3 ME/kgu- /:>
66
RE
I • C
• pST
ME
Figure 4. Effect of pST on retained energy RE at the same intake of metabolizable energy.
Figure 5. Effect of pST on maintenance requirement (ME).
Campbell e t al. (1988) derived a higher maintenance requirement for pST t reated animals from their calculations. Various possibilities can be the reason for increase a maintenance requirement. One reason may be activity. It can be expected that maintenance requirement will be increased by increased activity. However in various discussions it has been suggested that pST t reated animals are more lethargic then control animals (Curtis, 1989). Since in studies on N gain (Huisman et al., 1988) animals were kept in metabolic crates it was decided to measure metabolic ra te in growing animals which were kept in groups. Van der Hel et al. (1989) studied effect of metabolic rate in two trials with each 2k castrated male pigs of 70 to 90 kg for a period of 3 weeks each. In each trial 12 pigs
67
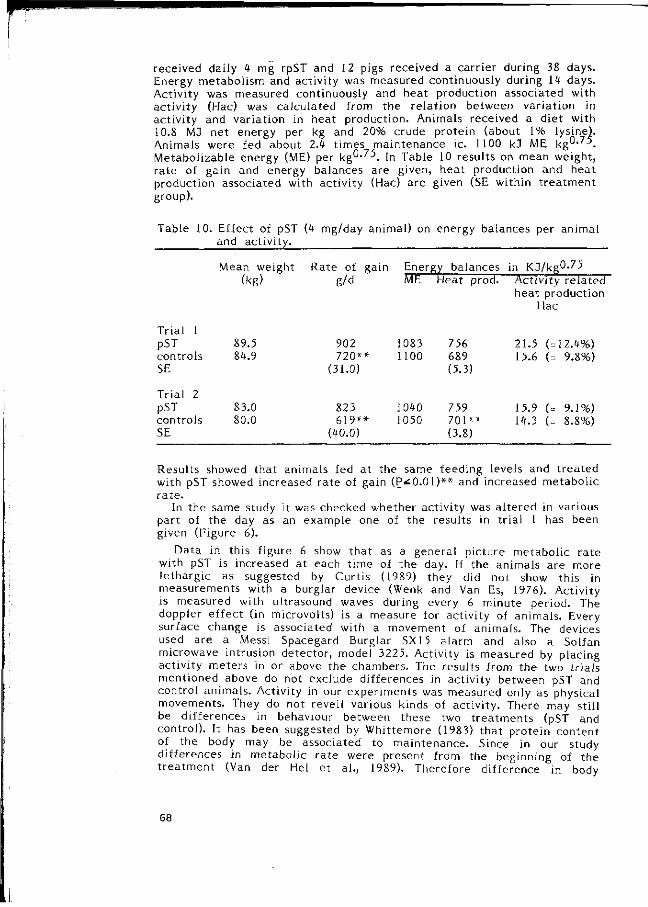
received daily 4 mg rpST and 12 pigs received a carrier during 38 days. Energy metabolism and activity was measured continuously during 14 days. Activity was measured continuously and heat production associated with activity (Hac) was calculated from the relation between variation in activity and variation in heat production. Animals received a diet with 10.8 Ml net energy per kg and 20% crude protein (about 1% lysine). Animals were fed about 2.4 times maintenance ic. 1100 kJ ME kg 0 , 7 * . Metabolizable energy (ME) per k g 0 , 7 5 . In Table 10 results on mean weight, rate of gain and energy balances are given, heat production and heat production associated with activity (Hac) are given (SE within t reatment group).
Table 10. Effect of pST (4 mg/day animal) on energy balances per animal and activity.
Mean weight Rate of gain Energy balances in K3/kg0-7-^ (kg) g/d ME Heat prod. Activity related
heat production Hac
Trial 1 pST 89.5 902 1083 756 21.5 (=12.4%) controls 84.9 720** 1100 689 15.6 (= 9.8%) SE (31.0) (5.3)
Trial 2 pST 83.0 823 1040 759 15.9 (= 9.1%) controls 80.0 619** 1050 701** 14.3 (= 8.8%) SE (40.0) (3.8)
Results showed that animals fed at the same feeding levels and t reated with pST showed increased rate of gain (P^O.01)** and increased metabolic ra te .
In the same study it was checked whether activity was altered in various part of the day as an example one of the results in trial 1 has been given (Figure 6).
Data in this figure 6 show that as a general picture metabolic rate with pST is increased at each time of the day. If the animals are more lethargic as suggested by Curtis (1989) they did not show this in measurements with a burglar device (Wenk and Van Es, 1976). Activity is measured with ultrasound waves during every 6 minute period. The doppler effect (in microvolts) is a measure for activity of animals. Every surface change is associated with a movement of animals. The devices used are a Messl Spacegard Burglar SX15 alarm and also a Solfan microwave intrusion detector, model 3225. Activity is measured by placing activity meters in or above the chambers. The results from the two trials mentioned above do not exclude differences in activity between pST and control animals. Activity in our experiments was measured only as physical movements. They do not reveil various kinds of activity. There may still be differences in behaviour between these two t reatments (pST and control). It has been suggested by Whittemore (1983) that protein content of the body may be associated to maintenance. Since in our study differences in metabolic rate were present from the beginning of the t rea tment (Van der Hel et al., 1989). Therefore difference in body
68
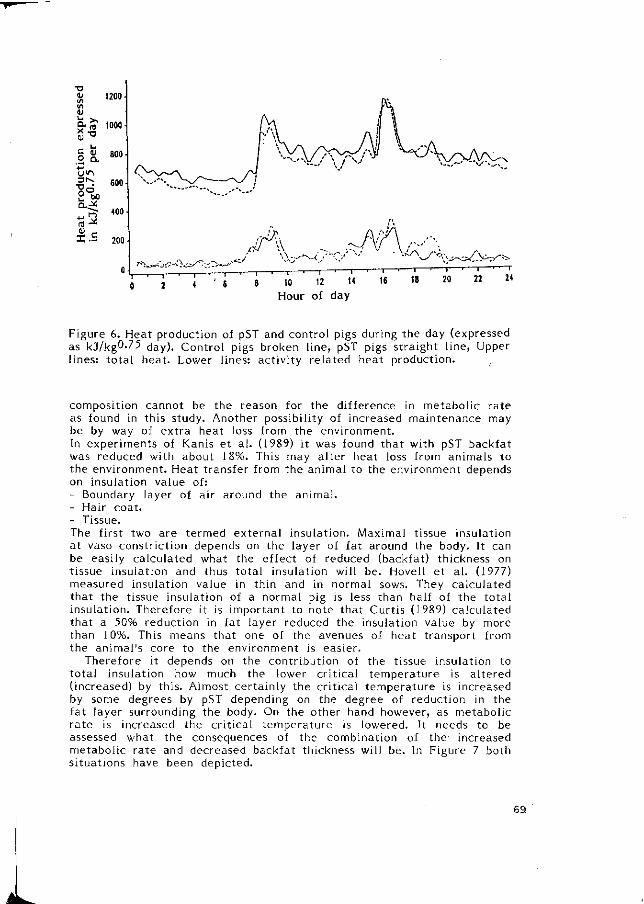
"O l/>
<U
*- :>* 2"° .2 D-<JU"N
O . * * J "
1200
tooo
800
600
400
200
\ \ . „ , - . * - < , V/\_/^\ ,_ — i —
to 12 14 Hour of day
—\— 18 20 22 24
Figure 6. Heat production of pST and control pigs during the day (expressed as k J /kg 0 - 7 5 day). Control pigs broken line, pST pigs straight line, Upper lines: total heat. Lower lines: activity related heat production. /
composition cannot be the reason for the difference in metabolic ra te as found in this study. Another possibility of increased maintenance may be by way of extra heat loss from the environment. In experiments of Kanis et al. (1989) it was found that with pST backfat was reduced with about 18%. This may alter heat loss from animals to the environment. Heat transfer from the animal to the environment depends on insulation value of: - Boundary layer of air around the animal. - Hair coat. - Tissue. The first two are termed external insulation. Maximal tissue insulation at vaso constriction depends on the layer of fat around the body. It can be easily calculated what the effect of reduced (backfat) thickness on tissue insulation and thus total insulation will be. Hovell et al. (1977) measured insulation value in thin and in normal sows. They calculated that the tissue insulation of a normal pig is less than half of the total insulation. Therefore it is important to note that Curtis (1989) calculated that a 50% reduction in fat layer reduced the insulation value by more than 10%. This means that one of the avenues of heat transport from the animal's core to the environment is easier.
Therefore it depends on the contribution of the tissue insulation to total insulation how much the lower critical temperature is altered (increased) by this. Almost certainly the critical temperature is increased by some degrees by pST depending on the degree of reduction in the fat layer surrounding the body. On the other hand however, as metabolic ra te is increased the critical temperature is lowered. It needs to be assessed what the consequences of the combination of the increased metabolic rate and decreased backfat thickness will be. In Figure 7 both situations have been depicted.
69

Heat production
pST
Tl T2
Temperature
Figure 7. Effect of pST on metabolic ra te in pigs (horizontal lines) and possible effects on thermal demand. C and pST represents increase of heat production with low temperature. The values Tl and T2 represents lower critical temperature.
The tpcC in Figure 7 is for increased thermal demand below thermoneu-trali ty and as insulation in pST animals is less than in controls, thermal demand will be probably increased. Traditionally it has been derived that metabolic rate may not be the same in various selected lines. Differences in maintenance requirement have been reported for pigs with different genetic capacities for growth (Campbell and Taverner, 1985) and in pigs selected for different backfat thickness (Sundst^l et al., 1979). Also in mice selected for low body weight Van der Wal et al. (1976) reported a higher maintenance compared to control animals.
Alteration of efficiency of protein synthesis
In Figure 8 it is shown how with similar maintenance differences in efficiency of retained energy can occur. Furthermore this efficiency can be calculated from energy in protein and/or from energy in fat gain. Retained energy
— ME
MEm is similar
Figure 8. Possible effect of pST on efficiency of synthesis of retained energy, assuming constant maintenance.
70

We assume that net protein synthesis is increased with the administration of rpST. However this does not mean that the partial energetic efficiency of protein accretion is also increased. A decreased protein turnover which is associated with higher protein gain would be expected to yield a lower heat production. It may be that an increase in protein accretion as a result of rpST is additionally associated with a more rapid protein turnover. This would lead to a changed metabolic rate depending on whether synthesis or turnover is changed most. If we assumed the same maintenance (420 k3 ME/kg u -^ ) in all animals and the same partial efficiency for fat deposition (kf = 0.74) in all animals, then the resulting kp were as follows:
* No change in maintenance 420 kJ ME/kg^-'-\ * No change in energy required per kJ fat deposited (kf = 0.74). * Change in energy required per kJ protein retained:
Controls kp = 0.59 Treatment kp = 0.48
The higher metabolic rate with pST may also result from higher rate of protein accretion. Our results showed that the partial efficiency of protein accretion in control animals (kp) was similar to the ARC (1981) es t imate. However kp in pST animals was markedly reduced, thus this is similar to the argument of Campbell et al. (1988).
Alterations in efficiency of fat synthesis
There may still be a third reason for the increase in heat production. Increased lypolysis (Etherton et al., 1986a) may increase fat "turnover" and as a result the partial efficiency of fat deposition from metabolizable energy is reduced. This implies that kf in rpST animals should be less than .74 compared to control animals. When we calculated kf by assuming that maintenance is equal in all groups (420 kJ ME/kg^-'-5) and partial protein efficiency was also equal (kp = 0.54) then the resulting kf was as follows. * No change in maintenance 420 kJ ME/kg^ - ' - ' . * No change in energy required per kJ in protein (kp = 0.54). * Change in energy required per kJ in fat:
Controls kf = 0.77 Treatment kf = 0.68
Turnover however is found at all levels of metabolism. According to Reeds (1987) at virtually every "key" reaction both forward and reverse reaction (or both) can be increased. The result will be that metabolic control of the cycles will determine what are the changes in heat production. In addition the net result of the forward and reverse cycles. Campbell et al. (1988) argued that the inhibition of lipogenesis may cause the decrease in fat accretion. They however also suggest the reduced amount of energy available for lipogenesis may contribute in a passive way to reduction in fat accretion. Their results of increased fat accretion with extra energy intake and pST showed tha t the reduced availability of energy above that for protein synthesis and/or maintenance may be responsible for the lower deposition. It may be that energy requirement for in maximum protein gain with pST may be increased. Thus metabolic changes in pST animals need to be assessed with regard to requirements of protein and energy. It can be concluded that pST increased protein gain, reduces fat gain and
71

increases metabolic rate. Until now it is not clear, whether the cause is a l tered maintenance and/or lower efficiency of energy gain. It need to be studied if the altered metabolic rate associated with pST is of a similar magnitude to the a l teration brought about by selection towards a similar combination of protein and fat gain as with pST. In that respect it is also of interest to note that the effects of pST on protein and fat gain are sustained after withdrawal of pST t reatment . Campbell et al. (1989) found that after administration during 30-60 kg and withdrawal ra te of gain after 60 kg was sustained to be superior compared to control animals. In addition feed intake was similar during the withdrawal period.
It is not clear how pST effects metabolic ra te . It would be of interest to know if metabolic rate still remains increased at those conditions. From the discussion on application of pST in growing and fattening pigs it can be concluded that pigs with pST - at same intake more protein gain ( ? ' 2 5 % extra) - at same intake less fat gain - at same intake more rate of gain - at same intake more lean % - higher heat production - maintenance and/or efficiencies al tered - thermal demand probably altered.
References
ARC, 1980. The nutrient requirement of pigs. Agricultural Research Council Commonwealth Agricultural Bureaux, Slough, England, 341 pp.
Boyd, R.D., 1987. Somatotropin and productive efficiency in swine. Animal Health & Nutrition, Febr. 1987, p. 23.
Boyd, R.D., D. Way-Cahen, D.E. Bauman, D.H. Beerman, A.F. de Neergard, L. South, 1987. In: Proceedings of the Maryland Nutrition Conference for Feed manufacturers pp. 58-66.
Boyd, D.R., D. Way-Cahen and B. Kirk, 1988. Implications of somatotropin on nutrient requirements of growing swine. Proceedings of International Seminar on the Science of Porcine Somatotropin, p. 122-143.
Campbell, R.G., N.C. Steele, T.J. Caperna, J.P. McMurtry, M.B. Solomon and A.D. Mitchell, 1988. Interrelationship between energy intake and endogenous porcine growth hormone administration (pGH) on the performance body composition and protein and energy metabolism of growing pigs weighing 25 to 55 kg live weight. J.Anim.Sci. 66: 1643-1655.
Campbell, R.G., N.C. Steele, T.J. Caperna, J.P. McMurtry, M.B. Solomon and A.D. Mitchell, 1989. Interrelationship between sex and exogenous growth hormone administration on performance, body composition and protein and fat accretion of growing pigs. J.Anim.Sci. 67: 177-186.
Campbell, R.G. and M.R. Taverner, 1985. The effects of strain and sex on protein and energy metabolism in growing pigs. In: P.W. Moe and H.F. Tyrell and P.J. Reynolds, Ed. EAAP-publ.no. 31. Proc.En.Met. Virginia, p. 78-81. Roman and Littlefield, USA.
Curtis, S.E., 1989. Potential side-effects of exogenous somatotropin in pigs. In: P. van der Wal, G.J. Nieuwhof and R.D. Politiek Eds. Biotechnology for control of growth and product quality in swine. Implications and acceptability, p. 155-158. Pudoc Wageningen.
72

Etherthon, T.D., C.M. Evock, C S . Chung, P.E. Walton, M.N. Sillence, K.A. Magri, R.E. Ivy, 1986a. Stimulation of pig growth performance by long-term t reatment with pituitary porcine growth hormone (pGH) and a recombinant pGH. J.Anim.Sci. 63 (suppl. 1): 219 (abstract).
Etherton, T.D., J.P. Wiggins, C S . Chung, CM. Evock, J.F. Rebhun, P.E. Walton, 1986b. Stimulation of'pig growth performance by porcine growth hormone and growth hormone releasing-factor. J.Anim.Sci. 63: 1389.
Etherton, T.D., 3.P. Wiggins, CM. Evock, C S . Chung, J.F. Rebhun, P.E. Walton, N .C Steele, 1987. Stimulation of pig growth performance by porcine growth hormone: determination of the dose-response relationship. J.Anim.Sci. 64: 433.
Fuller, M.F. and A.G. Chamberlain, 1985. Protein requirements of pigs, p. 85-96. In: D.J.A. Cole and W. Haresign, Eds. Recent Developments in pig nutrition. Butterworths, London.
Hel, W. van der, M.W.A. Verstegen, J. Huisman, E. Kanis, E.J. van Weerden and P. van der Wal, 1988. Effect of pST t reatment on energy balance t rai ts and metabolic rate in pigs. J.Anim.Sci. 66, suppl. 1: 255 abstract 93.
Hel, W. van der, M.W.A. Verstegen, E. Kanis, E.J. van Weerden and P. van der Wal, 1989. Effect of pST on metabolic rate and daily pattern of activity. J.Anim.Sci. In press.
Hovell, F.D. de, J.G. Gordon and R.M. MacPherson, 1977. Thin sows. 2 Observations on the energy and nitrogen exchanges of thin and normal sows in environmental temperatures of 20 and 5 °C J.Agric.Sci. 89: 523-533.
Huisman, J., E.J. van Weerden, W. van der Hel, M.W.A. Verstegen, E. Kanis and P. van der Wal, 1988. Effect of rpST t reatment on ra te of gain in protein and fat in two breeds of pigs and crossbreds. J.Anim.Sci. 66, suppl. 1: p. 255, abstract 93.
Kanis, E., G.J. Nieuwhof, K.H. de Greef, W. van der Hel, M.W.A. Verstegen, J. Huisman and P. van der Wal, 1989. Effect of recombinant porcine somatotropin on growth and carcass quality in growing pis. Interaction with genotype and slaughter weight. J.Anim.Sci. In press.
Low, A.G., 1985. Amino acid use by growing pigs. p. 108. In: D.J.A. Cole and W. Haresign, Eds. Recent developments in pig nutrition. Butterworths, London.
Machlin, L.J., 1972. Effect of porcine growth hormone on growth and carcass composition of the pig. J.Anim.Sci. 35: 794.
McLaren, D.G., G.L. Grebner, P.J. Bechtel, F.K. McKeith, J.E. Novakofski and R.A. Easter, 1987. Effects of graded levels of natural porcine somatotropin (pST) on growth performance of 57 to 103 kg pigs. J.Anim.Sci. 65 (suppl. 1): 245 (abstract).
Reeds, P.J., 1987. Metabolic control and future opportunities for growth regulation. Anim.Prod. 46: 149-160.
Sundst{$l, F., N. Standal and O. Vangen, 1979. Energy metabolism in lines of pigs selected for thickness of backfat and rate of gain. Acta Agricul-tura Scandinavica 29: 337-345.
Verstegen, M.W.A., W. van der Hel, A.M. Henken, J. Huisman, E. Kanis, P. van der Wal and E.J. van Weerden, 1989. Effect of exogenous porcine somatotropine administration on partitioning nitrogen and energy metabolism in three genotypes of pigs. J.Anim.Sci. In press.
Wal, H. van der, M.W.A. Verstegen and W. van der Hel, 1976. Protein and fat deposition in selected lines of mice in relation to feed intake. EAAP publ.no. 19. Proc.En.Metab. Vichy: 125-129.
73

n
Weerden, E.3. van, M.W.A. Verstegen, J.M. Fentener van Vlissingen, W. van der Hel, E. Kanis and P. van der Wal, 1989. Effect of pST on N gain in pigs at various protein and energy intake levels. J.Anim.Sci. In press.
Weerden, E.3. van and M.W.A. Verstegen, 1989. Effect of porcine somatotropin (pST) on environmental N pollution. In: Biotechnology for control and growth and product quality in swine. Implications and acceptability. P. van der Wal, G.J. Nieuwhof and R.D. Politiek, Eds. p. 237-246.
Wenk, C. and A.J.H. van Es, 1976. Eine methode zur Bestimmung des Energieaufwandes für die Körperliche Aktivität von wachsenden Kühen. Monatshefte 54: 232-236.
Whittemore, CT. , 1983. Development of recommended energy and protein allowances for growing pigs. Agricultural Systems 11: 159-186.
74
IL.

PRACTICAL APPLICATION OF (BIO)SYNTHETIC AMINO ACIDS IN POULTRY AND PIG DIETS
J.B. Schutte
TNO Institute of Animal Nutrition and Physiology (ILOB) P.O. Box 15, 6700 AA Wageningen
Summary
In this paper some aspects with regard to the practical application of (bio) synthetic amino acids in poultry and pig diets are discussed. - The dietary level of protein in layer and pig diets can be reduced
with approximately 2.5 units when the diets are adequately supplied by the first limiting amino acids.
- The replacement of 42 protein from soya by synthetic amino acids did not effect chick performance. In laying hens, however, the substitution of 3.52 soya protein by synthetic amino acids lead to a significantly lower performance. The difference in reaction to dietary supplementation with synthetic amino acids between chicks and layers may be explained by the differences in time during which feed was available to the birds; chickens 24 h /day, layers 16 h /day.
- D- and DL-methionine are utilized as well as L-methionine in chicks, independent of SAA level or nature of the diet.
- The D-isomer of tryptophan is only partially utilized by chicks. In pigs indications were found that the utilization of the D-isomer of tryptophan depends partly on the dietary level of L-tryptophan. At a dietary L-tryptophan level of approximately 0.15%, equal utilization was found for DL and L tryptophan.
- In pigs the maximum replacement value of cystine for methionine was found to be approximately 502.
1. Introduction
At present the utilization of protein is an important subject of discussion in the Netherlands because of the serious environmental problems related with the intensive production of pigs and poultry in certain parts of the country. One of the most serious contaminants of the environment originating from animal production is nitrogen; for example the contamination of the ground water with nitrate and the contamination of the air with ammonia. The principal reason why animal production is largely responsible for this contamination, is the fact that animals are inefficient in converting of feed protein into animal protein. In pigs and poultry on an average only 35 to 451 of the protein cq. nitrogen (N) consumed is used for the production of meat and/or eggs. One of the most important factors affecting the utilization of dietary protein is the balance of the amino acids in the protein. The closer the amino acid composition of the diet matches the requirement for maintenance and production of meat and eggs, the less protein the animal needs. However, a typical problem in feed formulation is the achievement of an optimal balance of amino acids. In pig and poultry diets the most critical amino acids in this connection are methionine, lysine, threonine, tryptophan, isoleucine and, in broiler diets also arginine. Therefore, supplementation of a diet with these amino acids provides a means for increasing the efficiency of the utilization of dietary protein.
75
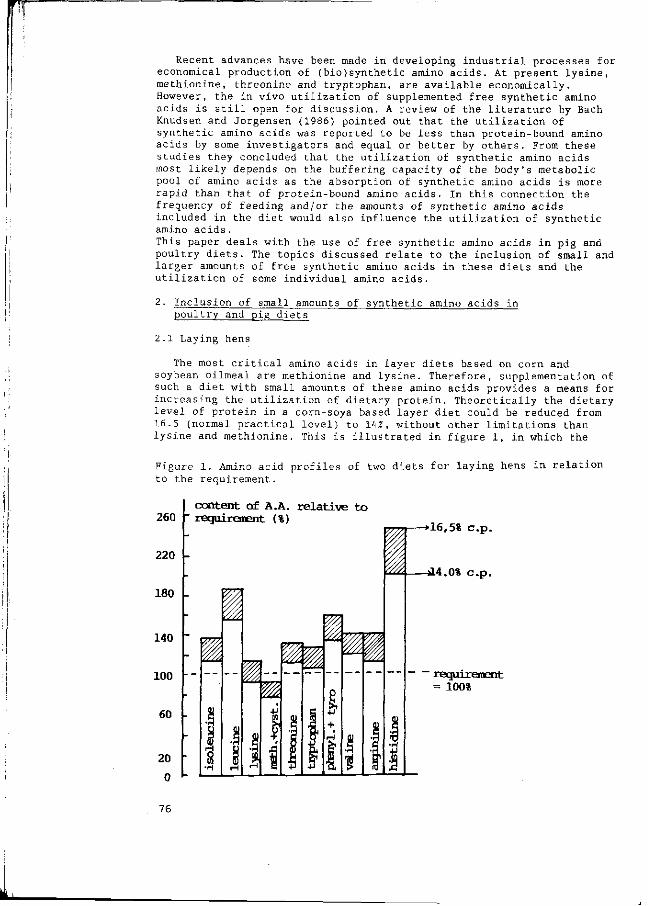
Recent advances have been made in developing industrial processes for economical production of (bio)synthetic amino acids. At present lysine, methionine, threonine and tryptophan, are available economically. However, the in vivo utilization of supplemented free synthetic amino acids is still open for discussion. A review of the literature by Bach Knudsen and Jorgensen (1986) pointed out that the utilization of synthetic amino acids was reported to be less than protein-bound amino acids by some investigators and equal or better by others. From these studies they concluded that the utilization of synthetic amino acids most likely depends on the buffering capacity of the body's metabolic pool of amino acids as the absorption of synthetic amino acids is more rapid than that of protein-bound amino acids. In this connection the frequency of feeding and/or the amounts of synthetic amino acids included in the diet would also influence the utilization of synthetic amino acids.
This paper deals with the use of free synthetic amino acids in pig and poultry diets. The topics discussed relate to the inclusion of small and larger amounts of free synthetic amino acids in these diets and the utilization of some individual amino acids.
2. Inclusion of small amounts of synthetic amino acids in poultry and pig diets
2.1 Laying hens
The most critical amino acids in layer diets based on corn and soybean oilmeal are methionine and lysine. Therefore, supplementation of such a diet with small amounts of these amino acids provides a means for increasing the utilization of dietary protein. Theoretically the dietary level of protein in a corn-soya based layer diet could be reduced from 16.5 (normal practical level) to 14Z, without other limitations than lysine and methionine. This is illustrated in figure 1, in which the
Figure 1. Amino acid profiles of two diets for laying hens in relation to the requirement.
260
220
180
140
100
content of A.A. relative to requirement (%)
>16,5% c.p.
J.4.0% c.p.
requirement = 100%
76

amino acid profiles of a 16.5 and 14.01 crude protein (c.p.) corn-soya diet in comparison with the requirement are presented. The requirement figures for each essential amino acid is set at 1001. The amino acid contents of both protein diets are expressed as a percentage of the requirement. If the content of an amino acid in the diet is below 1002, this means that the diet is deficient in that amino acid, if the content is above 1002, the diet has a surplus of that amino acid. The requirement figures for amino acids are derived from those presently used in the Netherlands. For some of the most important amino acids these requirement figures (in 2 of the diet) are as follows: SAA (methionine + cystine), 0.682; methionine, 0.341; lysine, 0.762; threonine, 0.502; tryptophan, 0.152 and isoleucine 0.552. These levels hold only when using well digestible diets containing approximately 11.7 MJ.ME/kg. The theoretical calculation that by reducing the dietary level of protein from 16.5 to 142, only methionine and lysine will become limiting, was tested in a series of four experiments. In these Trials the performance of layers on a 16.52 c.p. diet supplemented with methionine up to a level of 0.652 SAA was compared with those on a 142 c.p. diet supplemented with methionine (0.652 SAA) and lysine up to a level of approximately 0.752. Both diets were based on corn and soya. The birds were housed individually in battery cages. The hens were placed in the cages 2-4 weeks before the Trial started at approximately 26 weeks of age. Groups of 15 adjacent cages containing birds of similar initial body weight were used as replicates. Six replicates of each treatment were used in a randomised block design. Light was provided for a period of 16 hours/day. /
The birds were fed the experimental diets ad libitum as dry mash for three months. However, two of the Trials were extended over a full laying period of 52 weeks. The results achieved in the Trials are summarized in Tables 1 and 2.
Table 1. Laying hen performance (egg mass per hen in g per day and feed conversion) on a diet with 16.5 and 14% protein. Age period 26-38 weeks.
Experiment Egg mass Feed/egg mass
16.57. c.p1 142 c.p.2 16.52 c.p.1 142 c.p.2
111.03 111.04 111.05 111.07
51.4 51.1 55.7 54.7
50.4 49.4 56.1 53.0
2.37 2.17 2.10 2.07
2.37 2.25 2.13 2.08
Mean 53.2 52.4 2.18 2.21
1 Supplemented with methionine. 2 Supplemented with methionine and lysine.
In none of the trials was there a significant difference between both diets in egg mass production and feed conversion efficiency. On a composite basis, performance of the hens fed the 16.57. c.p. diet was somewhat better than that of the hens fed the 142 c.p. diet. However, the small difference in favour of the 16.52 c.p. diet was not significant. So, from the results of these trials it can be concluded, that it is
77

Table 2. Laying hen performance (egg mass per hen in g per day and feed conversion) on a diet with 16.5 and 14% protein. Age period 21-78 weeks.
Experiment Egg mass
1 2 16.52 c.p. 142 c.p.
Feed/egg mass
1 2 16.5% c.p. 142 c.p.
111 . 03 111 .04
5 1 . 1 52 . 0
5 0 . 3 5 1 . 8
2 . 47 2 . 25
2 . 4 8 2 .29
Mean 51.6 51.0 2.36 2.38
1 Supplemented with methionine 2 Supplemented with methionine and lysine.
Table 3. Effect of protein level on N excretion.
Layer 1 Layer 2
Crude protein content (2) 16.5 14.0
Intake of N (g/day), N retention (g/day)j N excretion (g/day)~
3.04 1.12 1.92
2.60 1.12 1.48
1 Feed intake 115 g/day 2 1.10 g in eggs and 0.02 g in growth 3 via faeces + urine.
possible to attain nearly the same performance with a 142 c.p. corn soya based diet than with a standard 16.52 c.p. diet, provided the low-protein diet is in addition to methionine also supplemented with lysine. The latter will not only improve the utilization of the dietary protein, but as a result also influence N excretion. The consequences of the reduction in dietary protein from 16.5 to 14Z in relation to the excretion of N are illustrated in Table 3.
The results presented in this table show that reducing the dietary protein level from 16.5 to 142, results in a reduction of N excretion of 0.44 g/day. This reduction corresponds with approximately 252.
2.2 Pigs
In practical Dutch pig diets, the most critical amino acids are lysine, methionine, threonine and tryptophan. Following these amino acids isoleucine, valine and histidine will most likely become limiting by reducing the dietary protein level. Theoretically the dietary level of protein in a barley, corn, tapioca and soya bean oilmeal based pig
78

Table 4. Utilization of free synthetic amino acids (FSAA) in pigs.
Daily weight gain
17.52 c.p. 15.02 c.p. FSAA
707 696 711 725
+
Feed/gain
17.52 c.p.
1.905 1.919
15.01 c.p. FSAA
1.918 1.904
+
Mean 709 710 1.912 1.911
diet could be reduced with 2.5 units without other limitations than the above mentioned amino acids. This assumption was tested in two Trials with pigs in the live weight period of 20-40 kg. In addition to a positive control diet containing 17.52 crude protein (normal practical level) a negative diet was formulated containing 15.02 crude protein. The low protein diet was supplemented with lysine, methionine, threonine and tryptophan up to a level present in the 17.52 protein diet. To be sure that no other amino acids would become limiting in the low protein diet, also small amounts of isoleucine, valine and histidine were included in this diet. In total 0.942 of synthetic amino acids were added to the low protein diet; 0.432 lysine, 0.142 methionine, 0.122 threonine, 0.042 tryptophan, 0.102 isoleucine, 0.052 valine and 0.062 histidine. Each experimental diet was fed to four replicate pens of eight pigs each, two pens with group-housed barrows and two with group-housed gilts. The experimental diets were fed ad libitum as pellets for a period of four weeks (age period of 9-13 weeks). The trial was repeated once. The results of this study (Table 4) show that with both diets almost equal performance was achieved. So, the inclusion of small amounts of synthetic amino acids in the diet did not affect pig performance when the diets were fed ad libitum as pellets.
In conclusion it can be stated that the utilization of dietary protein in pigs can be improved considerably by including the first limiting amino acids in the diet. As a result this will contribute substantially to the reduction of the N excretion. The practical potentials, however, depend on the price relations between feed components and synthetic amino acids in connection with the amounts they have to be included in the diet. The amino acids isoleucine, valine and histidine are not available commercially. So, an addition of these amino acids to the diet will increase the price of feed considerably. Therefore a Trial was performed to study whether or not the diet has to be supplied with isoleucine, valine and histidine when reducing protein level from 17.5 to 152 in a barley, corn, tapioca and soya bean oilmeal based young pig diet. This study was carried out in cooperation with the Institute for Livestock Feeding and Nutrition Research (IVVO). The results of this study pointed out that valine and histidine will not become limiting by reducing the dietary level of protein with 2.5 units. The results with regard to isoleucine are not yet available.
79

3. Inclusion of larger amounts of synthetic amino acids in poultry diets
3.1 Broiler chicks
To study the effect of larger inclusions of synthetic amino acids on chick performance, two diets were formulated and calculated to contain 20 and 162 crude protein (c.p.), respectively. The main components of the diets were corn and soybean oilmeal. Both diets were supplemented with L-lysine and DL-methionine in order to obtain a dietary level of 1.252 lysine and 0.861 SAA. The 162 c.p. diet was fed without and with a supplementation of all other essential amino acids and non-essential amino acids (as glutamic acid and glycine) up to the levels present in the 202 c.p. diet. Each experimental diet was fed to four replicate floor pens of 40 birds each. The diets were fed ad libitum as pellets for a period of three weeks (7-28 days of age). The trial was repeated once. Reducing the dietary level of protein from 20 to 16% resulted in both Trials in a significant decrease of chick performance (Table 5 ) . However, this negative effect could completely be overcome by an addition of free synthetic amino acids to the 162 c.p. diet. This suggest that free synthetic amino acids are utilized as well as the protein-bound amino acids in chicks. Table 5. Utilization of free synthetic amino acids (FSAA) in chicks.
Treatment Weight gain (g) Feed/gain
202 c.p. 162 c.p.
diet diet 1)
162 c.p. diet+FSAA' 2)
Exp. A
810 666 813
Exp. B
840 618 836
Exp. A Exp.
1 .61 1.92 1.59
1 .54 1.80 1.55
1) SAA and lysine at the 202 c.p. level. 2) All essential and non-essential amino acids at the 202 c.p. level.
3.2 Laying hens
In the study with laying hens two diets, based on corn and soybean oilmeal, were formulated and calculated to contain 14 and 10.52 c.p., respectively. Both diets were supplemented with L-lysine and DL-methionine in order to obtain dietary levels of 0.732 lysine and 0.65% SAA. The 10.52 c.p. diet was fed without and with an addition of all other essential amino acids and non-essential amino acids up to a level present in the 142 c.p. diet. Each experimental diet was fed to 60 individually caged birds. The diets were fed ad libitum as dry mash for a period of 12 weeks (26-38 weeks of age). Light was provided for a period of 16 hours/day. Reducing the dietary protein level from 14.0 to 10.52 resulted in a significant decrease of layer performance. The addition of essential and non-essential amino acids to the 10.52 c.p. diet did improve performance significantly. However, egg mass production and feed conversion efficiency were still significantly less favourable than on the 14.02 c.p. diet. These results suggest that

Table 6. Utilization of free synthetic amino acids (FSAA) in laying hens.
Treatment Daily Daily Feed/Egg egg' mass (g) feed intake (g) mass
14.02 c.p. diet 53.8 111 2.06 10.52 c.p. diet } 41.9 102 2.44 10.52 c.p. diet + FSAA ' 50.8 109 2.15
1) SAA and lysine at the 142 c.p. level. 2) All essential and non-essential amino acids at the 142 c.p. level.
synthetic amino acids are less well utilized than protein-bound amino acids in laying hens. The difference in reaction to dietary supplementation with synthetic amino acids between chicks and laying hens may be explained by the difference in time during which feed was available to the birds, i.e. chicks 24 hours/days and layers 16 hours/day. That the frequency of feeding may influence the utilization of synthetic amino acids is supported by the literature. This was first demonstrated by Batterham (1974), who found that in/pigs free synthetic lysine was utilized much better by feeding the diet six times per day than by feeding the lysine supplemented diet once a day. Buraczewska et al (1980) studied once and four times daily feeding of a diet supplemented with synthetic lysine on the absorption rate of N and lysine in pigs. When feeding once a day, the rate of passage of nitrogen through the intestine varied from 0.4 to 3.2 g/h, while the lysine content of the digesta was 3.2 to 4.5 g/16 g N. When feeding four times a day, the rate of passage of nitrogen was less variable, ranging from 0.7 to 1.8 g/h and the lysine content in protein (N x 6.25) of the digesta was in the range of 3.4 to 4.1. In the case of laying hens a possible time lag effect might be acting in the change from light to dark. This hypothesis was tested in a study with broiler chicks. Two diets were formulated, a positive control diet containing 202 c.p. and a diet containing 162 c.p. supplemented with all essential and non-essential amino acids up to a level present in the 202 c.p. diet. At a lighting scheme of 24 h/day, on both diets the same performance was achieved. However, with a lighting scheme of 16 h light and 8 h darkness/day, feed conversion efficiency of the birds fed the 162 c.p. diet supplemented with synthetic amino acids was significantly less favourable (32) than that of the birds fed the 202 c.p. diet. No differences in weight gain were observed between both groups. So, in conclusion it can be stated that in broiler chicks a lighting scheme of 16 h light and 8 h darkness/day may influence the utilization of synthetic amino acids negatively.
4. Utilization of the D-isomer of methionine in broiler chicks
Considering the literature data there still seems to be a lack of agreement on the utilization of the D-isomer of methionine in chicks. Some investigators reported equal utilization of the D- and L-isomer (Leveille et al; 1960; Featherston et al; 1962; Bauriedel, 1963). On the other hand, Brüggemann et al (1962), Smith (1966) and Baker and Boebel (1980) reported the D-isomer to be inferior to the L-isomer. In
81

m
Table 7. Biological activity of DL-methionine by using a crystalline amino acid basal diet (trial 1).
Treat- Methionine Added SAA Average Average Feed/ Ana-ment source methionine level weight daily feed gain ly-?
equivalents (Z of ~-j- _•_,._,__ _J_ (Z of diet) } diet)
1 2 3 4 5 6 7 8
-L-methionine DL-methionine -L-methionine DL-methionine -
L-methionine
-0 0 -0 0
0
10 10
10 10
10
0 0 0 0 0 0 0 0
40 50 50 45 55 55 50 60
gain
(g)
375* 564b
546b
500?. 622b
634b
564* 619b
632b
intake (g/bird)
40.4 51.2 49.8 49.0 54.5 55.1 51.2 53.6 53.8
2 1 1 2 1 1 1 1 1
2 6 K
92b
<
82 b
90^
79b 9 DL-methionine 0.10 0.60
Composite data
L-methionine 602a 53.1 1.85a 4
DL-methionine 604a 52.9 1.84a
1) The crystalline amino acid basal diet contained 0.40, 0.45 or 0.501 SAA.
2) Within analysis, means followed by different superscripts are significantly different (P < 0.05).
addition, some authors reported that the utilization of the D-isomer depends on the diet composition. Baker & Boebel (1980) reported that the D-isomer is as well utilized as the L-form using an intact protein diet, but not when tested in a crystalline amino acid diet. Katz and Baker (1975) found that the utilization of the D-isomer depends on the total dietary level of SAA (methionine + cystine). At SAA levels near the requirement they found equal efficacy of D and L methionine, but at SAA levels below the requirement, L-methionine was found to be superior to D-methionine. In order to obtain more information about the influence of the diet composition and the total level of SAA on the utilization of the D-isomer, three Trials with broiler chicks were performed. In Trial 1 the biological activity of L and DL methionine was compared using a crystalline amino acid diet. The two methionine forms were tested in the basal diet at equimolar levels of 0.102 methionine at total SAA levels of 0.40, 0.45 and 0.502, respectively. Trial 2 and 3 were designed to compare the biological activity of L-, D and DL methionine using a semi-purified and a practical type basal diet, respectively. In Trial 2 each of the methionine forms were tested at equimolar additions of 0.05, 0.10, 0.15 and 0.20Z methionine, and in Trial 3 at equimolar additions of 0.05 and 0.102.
Each experimental diet was fed to six replicates (= cages) of 13 birds each for a period of 21 days (6 to 27 days of age). The diets were fed ad libitum as dry mash.
The results obtained in the three trials are summarised in Tables 7 8 and 9.
In none of the trials were significant differences in performance observed between the methionine sources. So, in conclusion it can be stated that the isomers of methionine are equally utilized in broiler chicks, independent of SAA level or nature of the basal diet.
82

Table 8. Biological activity of L, purified basal diet (Trial 2).
D and DL methionine by using a semi-
Treatment
1 2 3
4 5 6
7 8 9
0 1 2
Methionine Source
D-methionine L-methionine
DL-methionine
D-methionine L-methionine
DL-methionine
D-methionine L-methionine
DL-methionine
D-methionine L-methionine
DL-methionine
Added Methionine equivalents
(I of diet ; )
0.05 0.05 0.05
0.10 0.10 0.10
0.15 0.15 0.15
0.20 0.20 0.20
Average Weight gain
(g)
398 412 428
497 498 494
554 566 564
594 614 600
Average daily feed intake (g/bird)
43.0 44.9 46.6
49.4 49.1 49.8
52.2 52.7 51.9
52.8 54.0 53.8
Feed/ gain
2.27 2.29 2.29
2.09 2.07 2.12
1.98 1.96 1.93
1.87 1.85 1.88
f aT 2) lysis
1
2
3
4 /
Composite data
D-methionine L-methionine
DL-methionine
511 522 521
49.4 50.2 50.5
2.05 2.04 2.06
5
1) The semi-purified basal diet contained 0.40Z SAA.
2) Within analysis, differences in weight gain and feed/gain ratio are not significant (]? < 0.05).
5. Utilization of the D-isomer of tryptophan in chicks and pigs
Data on the biological activity of the D-isomer of tryptophan in chickens and pigs are limited and variable. Ohara and Ariyoshi (1979) found that the biological effectiveness of D-tryptophan relative to the L-isomer was only 15% in chicks. In a subsequent study (Ohara et al., 1980) they estimated the relative biological utilization of DL and D tryptophan to be 47 and 211, respectively, of that of L-tryptophan. Much higher values are reported by Liebert & Gebhardt (1979). They estimated the biological activity of DL and D tryptophan relative to L-tryptophan in chicks to be 86 and 72Z, respectively. Shelton et al. (1951) and Thompson et al. (1952) fed DL-tryptophan to pigs and demonstrated partial use of the D-isomer. Arentsen & Zimmerman (1985) reported that the nutritive value of DL-tryptophan is approximately about 701 of that of L-tryptophan for weaning pigs. Kirchgessner & Roth (1985), however, reported an equal biological activity of DL and L tryptophan for pigs over the weight range of 20 to 55 kg. At ILOB one trial with chickens and two with pigs were carried out to obtain more information about the biological activity of DL-tryptophan. The results of these trials are reported briefly below.
83

Table 9. Biological activity of D and DL methionine by using a practical type basal diet xTrial 3 ) .
Treatment
Methionine source
Added methionine equivalents (1 of diet) ;
Average Average Feed/ weight daily gain gain feed intake (g) (g/bird)
Analysis
2)
1 2 3
4 5 6
D-methionine L-methionine
DL-methionine
D-methionine L-methionine
DL-methionine
D-methionine L-methionine
DL-methionine
0.05 0.05 0.05
0.10 0.10 0.10
Composite
657 669 663
679 683 682
data 668 676 672
54.4 55.1 55.2
54.6 55.0 54.6
54.5 55.0 54.9
1.74 1.73 1.75
1.69 1.69 1.68
1.72 1.71 1.72
1
2
3
1) The practical type basal diet contained 0.68% SAA.
2) Within analysis, differences in weight gain and feed/gain ratio are not significant (P > 0.05).
5.1 Chickens
In the study with broiler chicks, a semi-purified basal diet was used and calculated to contain 0.08Z tryptophan. The treatments consisted of additions of 0.0, 0.02, 0.04, 0.06 and 0.082 L or DL tryptophan. Each treatment was comprised of 112 birds, subdivided in eight replicates (= cages) of 14 birds each. The experimental diets were fed ad libitum as dry mash for a period of 18 days (7 to 25 days of age). The results of this trial (Table 10) indicate that both tryptophan sources did improve chick performance up to the highest supplemented level of tryptophan. However, at each supplemented level weight gain and feed conversion efficiency of the birds fed the L-tryptophan diets was significantly better than that of the birds fed the DL-tryptophan diets. From the results it was calculated that the biological activity of DL-tryptophan is approximately 65Z of that of L-tryptophan.
5.2 Pies
In the two pig trials, the biological activity of DL-tryptophan was compared with L-tryptophan by feeding diets severely to moderately deficient in tryptophan (trial 1) and moderately deficient to adequately supplemented with tryptophan (trial 2). In Trial 1, L- and DL-tryptophan were added in equimolar additions of 0.03 and 0.062 tryptophan to a practical type basal diet containing 0.092 tryptophan. Each experimental diet was fed to four replicate pens of 12 pigs each; two pens with group-housed barrows and two pens with group-housed gilts. The diets were fed twice a day according to a
84
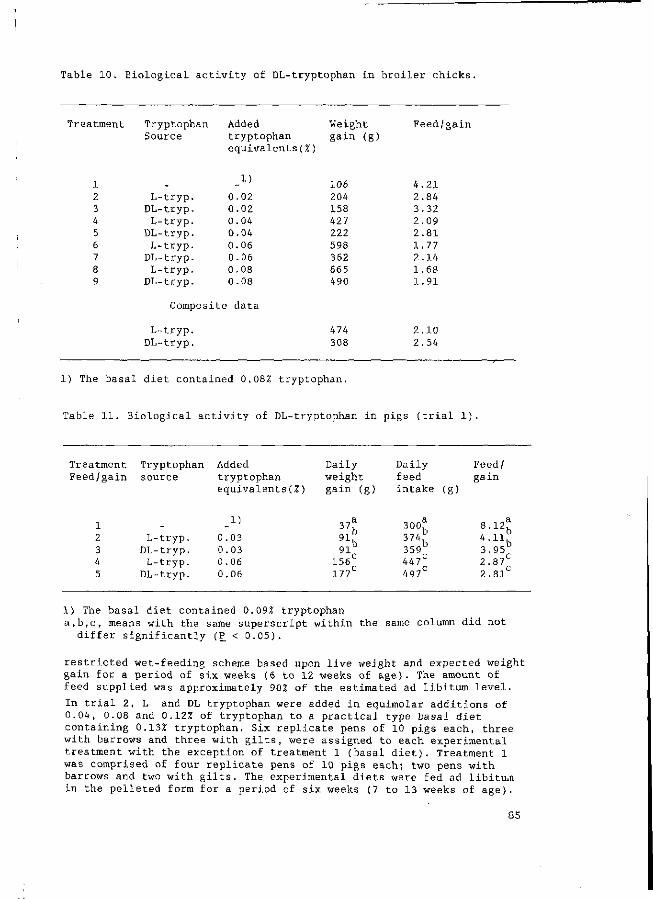
Table 10. Biological activity of DL-tryptophan in broiler chicks.
Treatment Tryptophan Added Weight Source tryptophan gain (g
equivalents(Z)
Feed/gain
1) 1 2 3 4 5 6 7 8 9
-L-tryp
DL-tryp L-tryp
DL-tryp L-tryp
DL-tryp L-tryp
DL-tryp
0.02 0.02 0.04 0.04 0.06 0.06 0.08 0.08
106 204 158 427 222 598 362 665 490
4.21 2.84 3.32 2.09 2.81 1.77 2.14 1.68 1.91
Composite data
L-tryp. DL-tryp.
474 308
2.10 2.54
1) The basal diet contained 0.08Z tryptophan.
Table 11. Biological activity of DL-tryptophan in pigs (trial 1)
Treatment Feed/gain
Tryptophan source
Added tryptophan equivalents (Z)
Daily weight gain (g)
Daily feed intake (g)
Feed/ gain
1)
L-tryp. DL-tryp.
L-tryp. DL-tryp.
0 0 0 0
03 03 06 06
3 7 t 9 1 t 91
156C
c 177
300, 374 359 447C
497C
8.12,
3.95 2.87C
1) The basal diet contained 0.09X tryptophan a,b,c, means with the same superscript within the same column did not
differ significantly (P < 0.05).
restricted wet-feeding scheme based upon live weight and expected weight gain for a period of six weeks (6 to 12 weeks of age). The amount of feed supplied was approximately 90Z of the estimated ad libitum level.
In trial 2, L and DL tryptophan were added in equimolar additions of 0.04, 0.08 and 0.1ZZ of tryptophan to a practical type basal diet containing 0.132 tryptophan. Six replicate pens of 10 pigs each, three with barrows and three with gilts, were assigned to each experimental treatment with the exception of treatment 1 (basal diet). Treatment 1 was comprised of four replicate pens of 10 pigs each; two pens with barrows and two with gilts. The experimental diets were fed ad libitum in the pelleted form for a period of six weeks (7 to 13 weeks of age).
85

Table 12. Biological activity of DL-tryptophan in pigs (Trial 2 ) .
Treatment Tryptophan Added Daily Daily Feed/gain source tryptophan weight feed
equivalents(2) gain (g) intake (g)
1) 1 2 3 4 5 6 7
-L- t ryp .
DL-tryp. L - t ryp .
DL-tryp. L - t ryp .
DL-tryp.
0.04 0.04 0.08 0.08 0.12 0.12
150 390f; 343^ 588
605 619
633e
de de
391 680e
618 , 971C
1008 1029fc
1053e
de
2.61" 1.74 1.80 1.64C
1.67C
1.66C
1.66C
1) The basal diet contained 0.132 tryptophan.
a, b, c, d, e, means with the same superscript within the same column
did not differ significantly (P < 0.05).
The results of the two Trials are summarized in Tables 11 and 12. The results show that in Trial 1, almost equal performance was achieved with both tryptophan levels at a dietary level of 0.031, whereas at a level of 0.06%, DL-tryptophan produced slightly better performance than did L-tryptophan. In Trial 2, the supplemental level of 0.042 L-tryptophan produced weight gain and feed intake results which were significantly superior to those of DL-tryptophan. However, at supplemental levels of 0.08 and 0.0122, diets with DL-tryptophan produced somewhat better performance than did L-tryptophan.
In summary, in both trials the same trend of increasing utilization of DL-tryptophan by increasing the supplemental level was observed. These results seem to indicate that the utilization of the D-isomer of tryptophan partly depends on the dietary level of L-tryptophan. Based on the results of trial 2, young pigs may already have problems with the utilization of the D-isomer when the diet contains less than approximately 0.152 tryptophan. For practical feed formulation, this point is of minor relevance since practical diets for young pigs contain generally more than 0.152 natural tryptophan.
6. Utilization of cystine in pigs
The estimated extent to which cystine can be used to meet the requirement for total SAA ranges from 40 to 702. Mitchell et al. (1968) reported that cystine can replace at least 702 of the need for total SAA. This value is considerably higher than earlier estimates of 402 by Becker et al. (1955) and 502 by Shelton et al. (1951). Baker et al. (1959) reported that the replacement value of cystine depends on the feeding system used. They found that when using an ad libitum system, 562 of SAA could be cystine. However, when using restricted feeding conditions, a replacement value of 662 was found. Recently, Roth & Kirchgessner (1987) reported that considerable losses in performance occur if the proportion of cystine exceeds 552 of the dietary level of SAA. At ILOB a study was carried out to obtain more information about the replacement value of cystine for methionine in pigs. For this study two practical type basal diets were formulated containing 0.252 methionine + 0.252 cystine and 0.202 methionine + 0.302 cystine, respectively. Both
86
Table 13. Utilization of cystine in pigs.
0 0
0 0
0 0
20 25
25 30
30 35
0.30 0.25
0.30 0.25
0.30 0.25
0.50 0.50
0.55 0.55
0.60 0.60
60 50
55 45
50 42
Treatment Dietary level (in Z) of: Cystine Daily Feed/gain as Z of weight
Meth. Cyst. SAA SAA gain (g)
1 0.20 0.30 0.50 60 610* 2.382!* 2 0.25 0.25 0.50 50 693 2.238
3 0.25 0.30 0.55 55 686 2.264b, 4 0.30 0.25 0.55 45 736C 2.136
5 0.30 0.30 0.60 50 729° 2.164^° 6 0.35 0.25 0.60 42 738° 2.092
a, b, c, d, means with the same superscript within the same column did not differ significantly (P < 0.05).
diets were supplemented with 0.05 and 0.10Z DL-methionine resulting in dietary SAA levels of 0.55 and 0.602 respectively. Each experimental diet was fed to six replicate pens of 9 pigs each; three pens with group-housed barrows and three with group-housed gilts. The experimental diets were fed ad libitum as pellets for a period of six weeks (8.5 to 14.5 weeks of age). The results of the study are summarized in table 13. From these results it can be deduced that an increase of the dietary SAA level with cystine did not have any effect on pig performance (Treatment 2 vs 3, Treatment 4 vs 5 ) . Further, it may be concluded from the results that in pigs not more than 501 of the total dietary SAA should be furnished by cystine. The 502 replacement value of cystine in pigs agrees well with those reported by Schutte et al. (1984) in layer diets.
7. References
Arentson, B.E. and Zimmerman, D.R. 1985. Nutritive value of D-tryptophan for the growing pig. Journal of Animal Science 60:474-479.
Bach Knudsen, K.E. and Jorgensen, H. 1986. Use of synthetic amino acids in pig and poultry diets. In: Recent Advances in animal nutrition. Hazesign, W. and Cole, D.J.A. (Eds.) Butterworths, London, p.215-225.
Baker, D.H., Clausing, W., Harmon, B.G., Jensen, B.G. and Becker, A.H. 1969. Replacement value of cystine for methionine for the young pigs. Journal of Nutrition 29:581-584.
Baker, D.H., and K.P. Boebel, 1980. Utilization of the D- and L-isomers of methionine and methionine hydroxy analogue as determined by chick bioassay. Journal of Nutrition 110:959-964.
Batterham, E.S., 1974. The effect of frequency of feeding on the utilization of free lysine by growing pigs. British Journal of Nutrition 31:237-242.
Bauriedel, W.R., 1963. The effect of feeding D-methionine on the D-amino oxidase activity of chick tissues. Poultry Science 42:214-217.
Becker, D.E., Jensen, A.H., Terril, S.W. and Norton, H.W. 1955. The methionine-cystine need of the young pigs. Journal of Animal Science 14:1086-1094.
87

Brüggemann, J.K., K. Drepper, and H. Zucker, 1962. Quantitative determination of the utilization of D-, L- and DL-methionine and DL-2-hydroxy-4-methylthiobutric acid-Ca by the chick. Naturwissenschaften 49:344.
Buraczewska, L. Lachowicz, J. and Buraczewski, S. 1980. The rate of absorption of synthetic lysine and dietary protein in the upper half of the small intestine of pigs. Arch. Tierernährung 30:751-758.
Featherston, W.R., H.R. Bird and A.E. Harper, 1962. Ability of the chick to utilize D- and excess L-indispensable amino acid nitrogen in the synthesis of dispensable amino acids. Journal of Nutrition 78:95-100.
Katz, R.S., and D.H. Baker, 1975. Efficacy of D-, L- and DL-methionine for growth of chicks fed crystalline amino acid diets. Poultry Sei. 54:1667-1674.
Kirchgessner, M., and Roth, F.X. 1985. Biologische Wirksamkeit von DL-tryptophan bei Mastschweinen. Zeitschrift fur Tierphysiologie, Tierernährung und Futtermittelkunde 54:135-141.
Leveille, G.A., R. Shapiro and H. Fischer, 1960. Amino acid requirements for maintenance in the adult rooster. IV. The requirements for methionine, cystine, phenylalanine, tyrosine and tryptophan; the adequacy of the determined requirements. Journal of Nutrition 72:8-15.
Liebert, F. and Gebhardt G. 1979. N-Bilanzuntersuchungen zur Verwertung von DL-Threonin und DL-Tryptophan am Broilerküken. Archiv Tierernährung 29:581-588.
Mitchell, J.R. , Becker, D.E., Harmon, B.G., Norton, H.W. and Jensen, A.H. 1968. Some amino acid needs of the young pig fed a semi-synthetic diet. Journal of Animal Science 27(2): 1322-1326.
Ohara, J. and Ariyoshi, S. 1979. Nutritive value of L-, DL- and D-tryptophan in the chick. Journal of Nutrition Science: Vitaminol. 25:185-193.
Ohara, I., Otsuka, S., Yugari, Y. and Aryoshi. S. 1980. Comparison of the nutritive values of L-, DL- and D-tryptophan in the rat and chick. Journal of Nutrition 110:634-640.
Roth, F.X. and Kirchgessner, M. 1987. Biological efficiency of dietary methionine or cystine supplementation with growing pigs. Journal of Animal physiology and Animal Nutrition 58(5): 267-280.
Schutte, J.B., Weerden van E.J. and Bertram, H.L. 1984. Protein and sulphur amino acid nutrition of the hen during the early stage of laying. Archiv Geflügelkunde 48 (5):165-170.
Shelton, D.C., Beeson, W.M. and Mertz, E.T. 1951. Quantitative DL-tryptophan requirement of the weanling pig. Journal of Animal Science 10:73-79.
Smith. R.E., 1966. The utilization of L-methionine, Dl-methionine andmethionine hydroxy analogue by the growing chick. Poultry Science 45:571-577.
Thompson. C M . , Reber, E., Whitehair, C.K. andMacVicar, R. 1952. Utilization of D-tryptophan by swine. Journal of Animal Science 11: 712-720.

PRESENT AND FUTURE DEVELOPMENTS IN THE PROTEIN/AMINO ACID SUPPLY OF MONOGASTRIC FARM ANIMALS
E.J. van Weerden
TNO Institute of Animal Nutrition and Physiology (ILOB) P.O. Box 15, 6700 AA Wageningen, NL
1. Introduction
In this paper, after a short overview of the present state of affairs of research into protein and amino acids in monogastric farm animals, possible developments in this area in the near future are discussed. The emphasis is placed on perspectives and problems of research aimed at solving the technical imperfections in present-day practical feed formulation.
2. Present state of affairs
In the last decades, our knowledge of protein/amino acid nutrition of monogastric farm animals has increased dramatically. These developments will be discussed on the basis of the two main criteria relevant to practical nutrition: 1) the contents of available amino acids in feed components 2) the requirements of the animals for these amino acids. '
The data presently available to nutritionists, are summarized below.
Contents of available amino acids in feed components
- Reliable tables with contents of total amino acids in common feed components
- Minimal data on contents of available/digestible amino acids in feed components.
Requirement figures
- Reasonably reliable requirement figures for lysine and sulphur amino acids (SAA) for the most important categories of poultry and pigs.
- Insufficiently reliable requirement figures for the other amino acids.
- No requirement figures based on determined availability/digestibility of the amino acids.
From the tables it is clear where the gaps are in the present knowledge. First, there is a lack of data on the contents of digestible amino acids in feedstuff s used in diet formulation for pigs and poultry. The question which of the digestibility parameters, faecal or ileal, is the best estimate for the protein feeding value, will be discussed later. Second, our present knowledge on amino acid requirements for the different categories of poultry and pigs is incomplete. This concerns especially the almost complete lack of data from requirement studies in which the digestibility of the amino acids in the test diets is determined rather than calculated.
The figures for requirements of digestible amino acids presently used in the Netherlands are all based on data calculated from these incomplete tables.
Therefore, the conclusion with regard to the present state of affairs, must be that although great progress in amino acid nutrition research has been made, there are still many questions, which are also relevant to practical nutrition, left unanswered. In the following, special attention will be paid to these questions and to the possibilities and problems related to further research in this field.
89

3. Future developments
3.1 Available amino acids in feedstuffs
This topic will be discussed on the basis of the following three items : - which criteria should be used: faecal or ileal digestibility? (3.1.1) - what are the advantages of using the method chosen? (3.1.2) - what are the problems, especially with regard to the methods of
determination (3.1.3)..
3.1.1 Faecal or ileal digestibility
Many years ago, researchers had already come to the generally accepted conclusion that the criterion "digestible amino acid" is a better measure for "available amino acid" than the criterion "total amino acid" as determined in the standard chemical analysis. However, even since it was demonstrated (Zebrowska, 1973; Just et al. 1981) that the protein digestion process taking place in the large intestine of pigs does not yield amino acids that can be used by the animal, the question has been raised as to which type of parameter for digestibility, faecal of ileal, is to be preferred. It was realized that especially for feed components and diets in which a considerable part of the protein is broken down by the microflora in the large intestine, faecal digestibility is not a reliable estimate for the availability of amino acids. After approximately ten years of research in different places of the world, general agreement was reached, stating that in pigs ileal digestibility of amino acids is a better estimate for availability than faecal digestibility.
In the Netherlands, a provisional table with ileal digestibility coefficients of lysine, methionine + cystine, threonine and tryptophan in several of the most important feed components used in pig diets, will most probably be published at the end of this year. This table will be based on results of research carried out by ILOB/CIVO during the last eight years, together with data from the literature.
The question as to whether or not ileal digestibility of amino acids in poultry is also a better measure of availability than faecal digestibility, cannot yet be answered.
Whereas it was generally accepted that the caeca and large intestine in fowl play only a minor role in the protein digestion process, recent results of Green & Kiener (1989) and of Schutte et al. (1987) created doubts about this concept. Green & Kiener (1989) compared digestibilities of protein and amino acids in intact and caecectomized cocks and found with soya and sunflower seed meal only small differences between both digestibilities. However, in meat meals, digestibilities with intact birds were higher. Schutte et al. (1987) compared digestibilities of protein and amino acids of Phaseolus vulgaris beans heat-treated in two different ways (105°C for 40 minutes, 105°C for 110 minutes) in intact cocks and birds canulated at the terminal ileum. The results are summarized in Table 1. With the 40-min autoclaved beans, the differences between faecal and ileal digestibilities are relatively small, with the beans heated for 110 minutes, the digestibilities are lower, especially the ileal digestibilities of threonine and tryptophan. This phenomenon of a distinct depressive effect of overheating on ileal amino acid digestibility, but not on faecal digestibility, was also observed in pigs by Van Weerden et al. (1987). Whether these findings indicate that ileal digestibility is in general also the preferable estimate for available amino acids in poultry remains to be seen.
90

Table 1. Faecal and ileal digestibilities of Phaseolus beans in cocks.
Beans autoclaved Beans autoclaved at 105 °C, 40 min at 105 °C, 110 min
Crude protein Lysine Methionine + cystine Threonine Tryptophan
faecal
68-74 53 68 72
ileal
71 78 58 65 71
faecal
69 65 50 62 66
ill
62 63 51 45 54
Table 2. Correlations (r) between N deposition (g/day) and intake of faecal or ileal digestible protein on amino acids (g/day).
Faecal digestible Ileal digestible
Crude protein 0.75 0.81 Lysine 0.87 0.88 Methionine 0.76 0.78 Threonine 0.89 0.91
Table 3. Correlations (r) between weight gain or feed conversion and faecal or ileal digestible protein or organic matter.
Protein
faecal dig.
0.34 -0.65
ileal dig.
0.76 -0.87
Organic Mat
faecal dig.
0.89 -0.88
ter
ileal dig.
0.80 -0.77
Weight gain Feed Conversion
3.1.2 Advantages of ileal digestibility
Before introducing the concept of ileal digestible amino acids in practical diet formulation, it is desirable to acquire information on the advantages of this new system. For this purpose Just et al. (1985) carried out a series of N balance experiments with 24 diets of different composition (crude fibre contents of 3.9 to 12.5% in the dry matter, different levels of tapioca and potato starch, etc.). The results are summarized in Table 2 and show that protein deposition is slightly higher when correlated with the intake of ileal digestible protein/ amino acids, but the differences with intake of faecal digestible amino acids are small and in general not significant. Dierick et al. (1987) compared diets with an increasing share of the large intestine in the total protein digestion and found (Table 3) that weight gain and feed conversion are correlated considerably higher with ileal than with faecal digestible protein. On the other hand, performance is higher when correlated with faecal rather than with ileal digestible organic matter. According to Dierick et al. (1987) this last finding again stresses the importance of hind gut fermentation for the energy supply of pigs,
91

especially when fed high-fibre diets.
The conclusion drawn from both studies could be that it is indeed advantageous to formulate pig diets on the basis of ileal digestible rather than faecal digestible amino acids. The extent of the difference between both methods will depend on the type of diet; as the contribution of the large intestine in the total protein digestion process increases, the differences between both methods will become more marked.
3.1.3 Problems
The method of determining ileal digestible amino acids is more complicated than determining of faecal digestible amino acids and leads, therefore, to a number of problems. Three of these problems will be discussed briefly below.
- Determining of ileal digestibility in pigs, and even more so in fowl, is complex, labourious and hence expensive. The experimental animals must be canulated at the end of the small intestine, the collection of intestinal chyme is time-consuming and indicators are usually necessary to estimate intestinal flow. In the course of the years, several techniques have been developed to measure ileal digestibility in pigs: T-canulae with spot-sampling of ileal chyme using indicators, re-entrant canules with quantitative chyme collection, semi-quantitative chyme collection via a large caecum canule, the newly developed ilea-rectal shunt method. In each method, also different modifications are applied. Until now it has not been possible to evaluate all these techniques or to judge the reliability of the results obtained with them. This is because a direct comparison has not yet been carried out.
- A second problem, strongly related to the former is that in each technique, modifications have been introduced in order to save time and labour. However, it is not always clear whether or not such modifications are justified, especially when the periods of chyme collection are reduced to such an extent that it is questionable whether the figures obtained are reliable. An extensive evaluation and strict standardization of techniques is a prerequisite to enable a clear comparison of the data obtained from the different research institutes. As an example of the differences obtained from the different research institutes, Table 4 gives figures of ileal digestibilities of protein and four important amino acids in maize, measured at institutes in the USA and Canada (four batches) and at ILOB (six batches) .
Table 4. Ileal digestibilities of maize.
Source Number of batches Protein Lysine Methionine Threonine Tryptophan
X
79
67
sd
2
4
X
76
54
sd
5
8
X
89
82
sd
2
3
X
73
59
sd
4
6
X
72
45
sd
3a
12
USA/Canada 4
ILOB 6
a. For tryptophan the number of batches was two
92

The data from ILOB are in all criteria lower than the American data. These differences, sometimes more than 20 units, could be explained by the difference in origin of the maize (ILOB-data are from maize of French origin), but the techniques used may have also played a role.
- A third point which is especially relevant for the practical application of ileal digestibility figures, is the necessity for simple, but reliable screening methods, preferably in vitro techniques, to become available for predicting ileal amino acid digestibilites in feed components and diets. In this respect it is likely that in the near future a choice can be made between the combined vitro-vivo technique of the modified nylon bag method and one of the vitro techniques under development at CIVO-Zeist and IVVO-Lelystad. The first results indicate that a reliable ranking between batches within one feed component will be possible.
The conclusion of this paragraph can be summarized as follows. Poultry - At this stage, no choice based on experimental data between faecal and ileal digestibility of amino acids possible. - More data needed on both criteria of digestibility. Pigs - Ileal digestibility of amino acids criterion of choice. - More data on ileal digestibility needed. - Standardization of techniques necessary. - Development of simple screening methods for ileal digestible amino acids needed.
In the future, research should also solve the problems related to factors responsible for obstructing the optimal process of digestion, especially that taking place in the small intestine. Particularly relevant in this respect are the "antinutritional factors" (ANFs) (Huisman, 1989, this volume) and some of the non-starch polysaccharides (NSPs) that interfere with the normal digestion process in the small intestine. In this last respect it is likely that the application of certain enzymes will be advantageous.
3.2 Amino acid requirements
In this paragraph attention is focussed on two aspects of this broad field of amino acid research, 1) the methods used in determining amino acid requirements, 2) the effect of the hormonal status of the animal on the amino acid requirements, especially in connection with the effects on environmental N pollution.
3.2.1 Methods of determination
It is evident that the requirement for amino acids should be expressed on the same basis as that used for the contents of available amino acids. This means that in the concept of ileal digestible amino acids, the requirements should also be given as ileal digestible components. Until now amino acid needs in poultry as well as in pigs are almost exclusively determined as "total amino acids". Very few results have appeared in the literature with requirement figures based on experimentally determined digestibility data. The recommended levels of faecal digestible amino acids in poultry and pig feeds in the CVE-tables presently used in the Netherlands, are based on total amino acid, converted to faecal digestible amino acid using the coefficients mentioned in the tables. It is evident that this situation is very unsatisfactory and it is highly desirable that in the near future reliable data on amino acid requirements based on experimentally measured digestibility coefficients of the amino acids should be available.
93

Below, the criteria used in determining amino acid requirements are listed. - Protein deposition: N balance
N in carcass - Weight gain, feed conversion, slaughter quality - Amino acids in blood - Urea in blood , ,
14 - Amino acid oxidation via CO .
Whereas, theoretically, the methods directly determining protein deposition are to be preferred, in young animals weight gain and other performance parameters can also give reliable estimates of amino acid requirement. This is because of the high correlation with protein deposition. The indirect measures of amino acid requirements, blood content of amino acids and blood content of urea, are not widely applied in monogastric animals as the results are, in general, irregular. Methods determining amino acid oxidation by measuring the production of
CO. have been introduced. The merits of this technique are not yet proven, but figures for requirements of young growing pigs, obtained with these methods, are definitely too low, as discussed by Henry et al. (1987) .
It is generally known that literature data on the amino acid requirements of the different categories of farm animals are rather different. One of the reasons, apart from the variation in methods used, is the different way in which the results of the experiments are analysed. The relevance of this point has been discussed by Fisher et al. (1973) on the basis of the results from a layer experiment described by Bray (1969), in which the effect of lysine intake on egg production was measured. In Figure 1, the relation between lysine intake and egg output as calculated by Fisher et al. (1973) is shown by using four different statistical methods: the linear "broken line" method (D) and three slightly different, non-linear methods (A, B, C ) . The non-linear methods give a better picture of the real situation than the linear "broken line" model (Figure 1). In ILOB amino acid studies, with poultry as well as with pigs this is also generally observed. The reason for the non-linearity of the response to graded levels of amino acid intakes is illustrated by Fisher et al. (1973) in Figure 2. In Figure 2, part (a) describes the reaction of an individual laying hen to increasing intakes of an amino acid. It is assumed that the hen responds in a linear way until a plateau is reached. In part (b) of the figure the variability in the production level of a small group of laying hens is indicated, resulting in the characteristic S-shape mean response curve of the group. This S-curve was generally found in our broiler studies when a broad range of amino acid intake levels was covered.
Nowadays, there is general agreement among researchers that an analysis of the results of experiments on amino acid requirement on the basis of the "broken line" model usually gives too low figures for the requirement (Figure 1). However, when non-linear models are applied, often too high estimates of the requirement are obtained. Despite many discussions among statisticians, a satisfactory, practical solution to this problem is not yet available. At ILOB, the procedure followed is that the results of these types of studies are statistically analysed according to a "best-fitting" model - generally a non-linear curve, frequently a quadratic - and the response curve is presented together with the "confidence limits" of the suggested requirement figure. For practical application it is important to give the complete response curve because this gives an indication of the negative effects when feeding below (or above) the optimum. The detrimental effects of a supply below the optimum can be different for the individual amino acids
94
Figure 1. Typical set of input—output data (o) from an experiment in which pullets were allowed free access to diets with a series of contents of an essential amino acid. After Bray (1969). Curve B (— — ) was fitted by the procedure described in this paper. The other plots are an exponential curve A (---), a quadratic curve C (— - — ) and two straight lines fitted by Bray D (- — ) . The curves were fitted by least-squares procedures.
50 Egg production per bird (g/day)
40
30 300 400 500 600 700
Lysine intake, mg/bird day
Figure 2. Model proposed for the response of laying hens to amino acid intake, a. Response of a single hen, b, with individual and mean response in a small group. Mean response marked by broken line.
Egg production per bird (g/day)
Amino acid intake, mg/bird day
95

Figure 3. Lysine requirement of young veal calves.
N balance(%) X
X
X 100 -
1.20 2.20 1.40 1.60 1.80 2.00 analysed lysine content (g/100 feed)
Figure 4. Requirement of methionine and cystine for young veal calves.
N balance(%) ^
100L _ ^ *
95-
90
85
80-
0.70 0.90 1.10 analysed m+c content ( g/100 g feed)
as shown in Figures 3 and 4 for lysine and methionine + cystine in young (four - seven weeks) veal calves (van Weerden & Huisman, 1978). When the lysine content of the milk replacer based on skimmed milk was 10 i below the optimum, N deposition was 51 depressed, but for methionine + cystine the N deposition was nearly 10Z lower when the dietary methionine + cystine content was 202 below the optimum.
3.2.2 Effect of hormonal status on amino acid requirements
In the paper by Schutte (1989, this volume) it is clearly shown that the possibilities of reducing environmental N pollution caused by poultry and pigs, are highly dependent on an exact knowledge of the
96
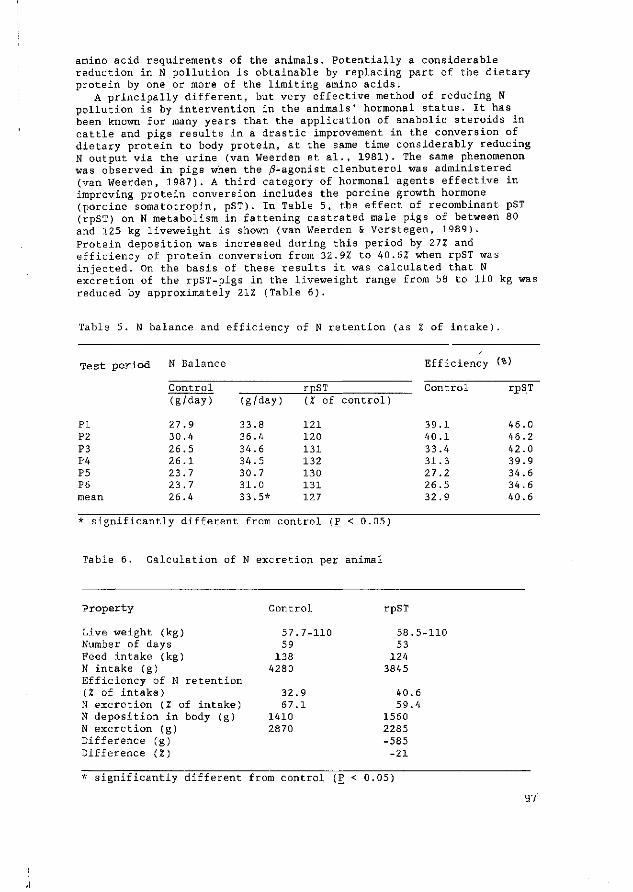
amino acid requirements of the animals. Potentially a considerable reduction in N pollution is obtainable by replacing part of the dietary protein by one or more of the limiting amino acids.
A principally different, but very effective method of reducing N pollution is by intervention in the animals' hormonal status. It has been known for many years that the application of anabolic steroids in cattle and pigs results in a drastic improvement in the conversion of dietary protein to body protein, at the same time considerably reducing N output via the urine (van Weerden et al., 1981). The same phenomenon was observed in pigs when the /3-agonist clenbuterol was administered (van Weerden, 1987). A third category of hormonal agents effective in improving protein conversion includes the porcine growth hormone (porcine somatotropin, pST). In Table 5, the effect of recombinant pST (rpST) on N metabolism in fattening castrated male pigs of between 80 and 125 kg liveweight is shown (van Weerden & Verstegen, 1989). Protein deposition was increased during this period by 27% and efficiency of protein conversion from 32.91 to 40.6% when rpST was injected. On the basis of these results it was calculated that N excretion of the rpST-pigs in the liveweight range from 58 to 110 kg was reduced by approximately 21Z (Table 6).
Table 5. N balance and efficiency of N retention (as X of intake).
Test period N Balance Efficiency (%'
PI P2 P3 P4 P5 P6 mean
Control (g/day)
27.9 30.4 26.5 26.1 23.7 23.7 26.4
(g/day)
33.8 36.4 34.6 34.5 30.7 31.0 33.5*
rpS' (Z .
121 120 131 132 130 131 127
(Z of control) Control
39.1 40.1 33.4 31.3 27.2 26.5 32.9
rpST
46.0 46.2 42.0 39.9 34.6 34.6 40.6
* significantly different from control (P < 0.05)
Table 6. Calculation of N excretion per animal
Property
Live weight (kg) Number of days Feed intake (kg) N intake (g) Efficiency of N retention (X of intake) N excretion (Z of intake) N deposition in body (g) N excretion (g) Difference (g) Difference (2)
Control
57.7-59
138 4280
32.9 67.1
1410 2870
• 110
rpST
58.5-110 53
124 3845
40.6 59.4
1560 2285 -585
-21
* significantly different from control (P < 0.05)
97

P excretion in these animals was reduced by 162 (van Weerden & Verstegen, 1989).
Whereas it is proven that compounds of the three mentioned categories of hormones or hormone-like agents can considerably increase protein deposition and decrease N excretion in pigs, it is still not clear whether or not the increased amounts of protein gained by the animal require higher levels of amino acids in the diet. As discussed by Campbell et al. (1989), the factorial approach applied by Boyd et al. (1988) predicting an almost doubling of the lysine requirement of pST-treated pigs, is of doubtful value. This approach is based on a number of assumptions which, especially in the case of agents affecting intermediary metabolism, are of questionable significance.
Experimental data on the protein/amino acid requirements of pST-treated pigs published so far, are incomplete. They indicate that protein/amino acid requirements may be somewhat, but not drastically, increased after pST treatment (Steele et al. 1989; Campbell et al. 1989). The results of two ILOB studies, one with veal calves and one with pigs, tend to confirm this preliminary conclusion.
In the study with milk-replacer-fed male veal calves of between i25 and 140 kg liveweight, placebos as well as calves implanted with oestradiol-17/3 + trenbolone acetate (20 mg + 140 mg) were given diets with increasing contents of methionine + cystine. Each treatment group included six individually fed animals. The results for N balance, measured over three consecutive 5-day test periods, are given in Table 7.
Whereas the number of methionine + cystine levels is only limited and the highest meth. + cyst, contents are apparently too low to enable reliable conclusions on the meth. + cyst, requirement of the steroid-treated calves, the slopes of both response curves are not very different. This suggests that the methionine + cystine need of the steroidimplanted calves is not considerably higher than that of the placebo animals, despite the fact that N deposition in the treated calves is, on average, 281 higher.
In the experiment with pigs, the effects on N balance in castrated animals of between 70 and 95 kg liveweight were measured of injections of recombinant porcine somatotropine (rpST). Ten groups of six barrows each were fed five test diets: three diets with "normal" energy content (2175 kcal net energy/kg) and different protein contents where lysine was the limiting factor, and two diets with a high energy level (2570 kcal/kg) with the two higher protein/lysine contents. Each diet was fed on a restricted scheme to a placebo and to a rpST-treated group. The average results of two 5-day test periods are given in Table 8. In the placebo groups fed the "normal" energy diets, protein deposition as well as weight gain and feed conversion were improved when the protein/lysine level in the diet was increased. This means that for these fast growing pigs the dietary lysine level of 0.80 - 0.90Z recommended in Dutch practice is already too low. It also stresses the need to regularly retest amino acid requirements in order to cope with genetic and other improvements influencing the animals capacity for protein gain/growth. Increasing the energy content of the diet did not affect protein gain. However, weight gain and feed conversion were more favourable. Recombinant pST increased protein deposition by 31Z, weight gain by 18% and feed conversion was 15Z lower. When dietary protein/lysine was increased, the effect of rpST on protein gain was increased, but the differences were not significant; the effects on weight gain and feed conversion were irregular.
The results of both pig and veal calf studies indicate the tendency for the protein/amino acid requirement after manipulation of the
98
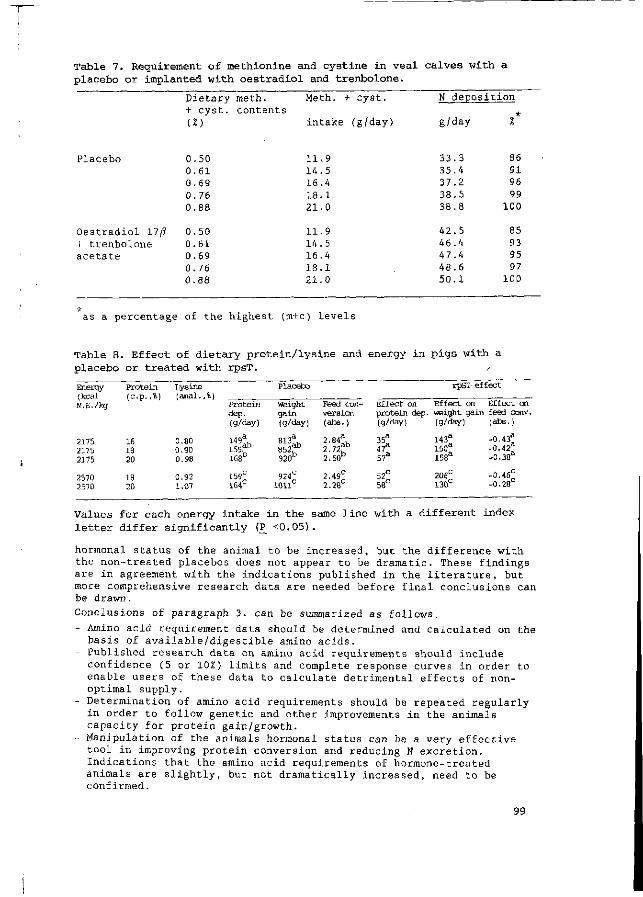
Table 7. Requirement of methionine and cystine in veal calves with a placebo or implanted with oestradiol and trenbolone.
P l acebo
O e s t r a d i o l 17/3 + t r e n b o l o n e a c e t a t e
D i e t a r y + c y s t . (Z)
0 .50 0 . 61 0 . 69 0 .76 0 .88
0 .50 0 . 61 0 .69 0 .76 0 .88
me th . c o n t e n t s
Meth.
i n t a k e
11 .9 14 . 5 16 .4 1 8 . 1 21 .0
11 .9 14 .5 16 .4 1 8 . 1 21 . 0
+ c y s t .
( g / d ay )
N d e p o s i t i o n
g / day
3 3 . 3 35 .4 37 . 2 3 8 . 5 38 .8
4 2 . 5 46 .4 47 . 4 48 . 6 5 0 . 1
I
86 91 96 99
100
85 93 95 97
100
as a percentage of the highest (m+c) levels
Table 8. Effect of dietary protein/lysine and energy in pigs with a placebo or treated with rpsT. ,
Energy
N .E . /kg
2175 2175 2175
2570 2570
P r o t e i n ( c . p . ,%)
16 18 20
18 20
Lys ine ( ana l . ,%)
0 .80 0 .90 0 .98
0 .92 1.07
P r o t e i n dep . (g /day)
149 aK
159?* 168 b
159C
164C
Placebo
Weight g a i n (g /day)
8 1 3 1 8 5 2 ^ 920°
924= 1011
Feed conv e r s i o n ( a b s . )
2 . 84 a . 2 . 7 2 ^ 2 . 5 0 b
2.28
E f fec t on p r o t e i n dep . (g /day)
K 4 7 a
57 a
5 2 c 58°
rpST- e f f e c t
E f f e c t on we igh t g a i n (g /day)
143 a
150 a
158 a
206° 130 c
Ef fec t on feed conv, ( a b s . )
- 0 . 4 3 a
- 0 . 4 2 a
- 0 . 3 8 a
- 0 . 4 6 C
- 0 . 2 8
Values for each energy intake in the same line with a different index letter differ significantly (]? <0.05).
hormonal status of the animal to be increased, but the difference with the non-treated placebos does not appear to be dramatic. These findings are in agreement with the indications published in the literature, but more comprehensive research data are needed before final conclusions can be drawn. Conclusions of paragraph 3. can be summarized as follows. - Amino acid requirement data should be determined and calculated on the
basis of available/digestible amino acids. - Published research data on amino acid requirements should include
confidence (5 or 10Z) limits and complete response curves in order to enable users of these data to calculate detrimental effects of non-optimal supply.
- Determination of amino acid requirements should be repeated regularly in order to follow genetic and other improvements in the animals capacity for protein gain/growth.
- Manipulation of the animals hormonal status can be a very effective tool in improving protein conversion and reducing N excretion. Indications that the amino acid requirements of hormone-treated animals are slightly, but not dramatically increased, need to be confirmed.
99

4. References
Boyd, D.R., Wray-Cahen, D. and Kirk, B., (1988). In: Campbell R.G., Johnson, R.J. and King, R.H. (1989). Implications of biotechnological techniques for manipulating animal growth and development on tissue and dietary nutrient requirements of pigs. Proc. Symp. "Biotechnology for control of growth and product quality in swine, implications and acceptability", p.137-144. (P. van der Wal, G.J. Nieuwhof, R.D. Politiek, editors). Pudoc, Wageningen, the Netherlands.
Bray, D.J., (1969). Studies with Corn-Soya Laying Diets. 8. Requirements for limiting amino acids. The basal diet and the requirements for isoleucine, lysine and tryptophan. Poultry Science 48, 674-684.
Campbell, R.G., Johnson, R.J. and King, R.H., 1989. Proc. Symp. "Biotechnology for control of growth and product quality in swine, implications and acceptability", Pudoc, Wageningen, p. 137.
Centraal Veevoederbureau in Nederland, 1986. Veevoedertabel. Centraal Veevoederbureau in Nederland, 1984. Voorlopige tabel
verteerbare aminozuren in veevoedergrondstoffen voor varkens. Dierick, N.A., Vervaeke, J., Decuypere, J., van der Heyde, H. and
Henderickx, H., (1987). Correlation of ileal and faecal digested protein and organic matter to production performance in growing pigs. In: European Association for Animal Production Publication No. 35, Session 3, 50-51.
Fisher, C , Morris, T.R. and Jennings, R.C., 1973. A model for the description and prediction of the response of laying hens to amino acid intake. British Poultry Science 14, 469-484.
Green, S. and Kiener, T., 1989. Digestibilities of nitrogen and amino acids in soya-bean, sunflower, meat and rapeseed meals measured with pigs and poultry. Animal Production 48, 157-179.
Henry, Y., Arnal, M., Obled, C. and A. Rérat, 1987. Protein and Amino Acid Requirements of Pigs. In: European Association for Animal Production Publication No. 35, Session 3, 9-18.
Just, A., Jorgensen, H. and Fernandez, J.A., 1981. The digestive capacity of the caecum-colon and the value of the nitrogen absorbed from the hind gut for protein synthesis in pigs. British Journal of Nutrition 46, 209-219.
Just, A., Jorgensen, H. and Fernandez, J.A., 1985. Correlations of protein deposited in growing female pigs to ileal and feacal digestible crude protein and amino acids. Livestock Production Science 12, 145-159.
Schutte, J.B., van Leeuwen, P. and van Weerden, E.J., 1987. •i: The ileal and faecal digestibility of protein and amino acids of heat
treated beans in poultry. In: European Association for Animal jjlji Production Publication No. 35, Session 3, 103.
i Steele, N.C., Campbell, R.G., Caperna, T.J., McMurtry, J.P. and | !• Solomon, M.B., 1989. PST efficacy in North America: management i'i; variables and advantages. In: Proc.Symp. "Biotechnology for j control of growth and product quality in swine, implications and
acceptability", pp 51-63. (P. van der Wal, E.J. Nieuwhof, R.D. Politiek, editors), Pudoc, Wageningen, the Netherlands.
Van Weerden, E.J., Berende, P.L.M, and Huisman, J., 1981. Application of endogenous and exogenous anabolic agents in veal calves. In: Proceedings E.C. Workshop "Anabolic agents in beef and veal production", pp. 1-26.
100

Van Weerden, E.J. and Huisman, J., 1978. De aminozuurbehoefte van het vleeskalf. In: Kalverdag Wageningen, 25 april 1978, pp. 4-21.
Van Weerden, E.J., 1987. Effects of Clenbuterol on N deposition and carcass composition in castrated male pigs. In: Beta-agonists and their effects on animal growth and carcass quality, pp. 152-162. Elsevier Applied Science, London and New York.
Van Weerden, E.J. and Verstegen, M.W.A., 1989. Effect of PST on environmental N pollution. In: Proc.Symp. "Biotechnology for control of growth and product quality in swine, implications and acceptability", 237-243 , Pudoc, Wageningen 1989, 237-243.
Zebrowska, T., 1973. Digestion and absorption of nitrogenous compounds in the large intestine of pigs. Rocz. Nauk. Roln., B.95, 80.
101


















