
Nucleocytoplasmic partitioning of tobacco N receptor is ...plantpath.ibb.waw.pl/pdf/New Phytologist...
14
Nucleocytoplasmic partitioning of tobacco N receptor is modulated by SGT1 Rafal Hoser 1 , Marek _ Zurczak 1 , Malgorzata Lichocka 1 , Sabina Zuzga 1 , Michal Dadlez 1,2 , Marcus A. Samuel 3 , Brian E. Ellis 4 , Johannes Stuttmann 5 , Jane E. Parker 5 , Jacek Hennig 1 and Magdalena Krzymowska 1 1 Institute of Biochemistry and Biophysics PAS, Pawinskiego 5a, 02-106, Warsaw, Poland; 2 Biology Department, Institute of Genetics and Biotechnology, Warsaw University, Pawinskiego 5a, 02-106, Warsaw, Poland; 3 Department of Biological Sciences, University of Calgary, 2500 University Dr NW Calgary, Alberta, T2N 1N4, Canada; 4 Michael Smith Laboratories, University of British Columbia, 2185 East Mall, Vancouver, BC V6T 1Z4, Canada; 5 Department of Plant–Microbe Interactions, Max-Planck Institute for Plant Breeding Research, Carl-von-Linn e-Weg 10, 50829, Cologne, Germany Author for correspondence: Magdalena Krzymowska Tel: + 48 22 592 5721 Email: [email protected] Received: 27 March 2013 Accepted: 29 April 2013 New Phytologist (2013) doi: 10.1111/nph.12347 Key words: disease resistance, MAPK, NB-LRR, nucleocytoplasmic shuttling, protein phosphorylation, SGT1, tobacco (Nicotiana tabacum). Summary SGT1 ( Suppressor of G2 allele of SKP1) is required to maintain plant disease Resistance (R) proteins with Nucleotide- Binding (NB) and Leucine- Rich Repeat (LRR) domains in an inactive but signaling-competent state. SGT1 is an integral component of a multi-protein network that includes RACK1, Rac1, RAR1, Rboh, HSP90 and HSP70, and in rice the Mitogen- Activated Protein Kinase (MAPK), OsMAPK6. Tobacco (Nicotiana tabacum) N protein, which belongs to the Toll- Interleukin Receptor (TIR)-NB-LRR class of R proteins, confers resistance to Tobac- co Mosaic Virus (TMV). Following transient expression in planta, we analyzed the functional relationship between SGT1, SIPK – a tobacco MAPK6 ortholog – and N, using mass spectrometry, confocal micros- copy and pathogen assays. Here, we show that tobacco SGT1 undergoes specific phosphorylation in a canonical MAPK target-motif by SIPK. Mutation of this motif to mimic SIPK phosphorylation leads to an increased proportion of cells displaying SGT1 nuclear accumulation and impairs N-mediated resistance to TMV, as does phospho-null substitution at the same residue. Forced nuclear localization of SGT1 causes N to be confined to nuclei. Our data suggest that one mode of regulating nucleocytoplasmic partitioning of R proteins is by maintaining appropriate levels of SGT1 phosphorylation catalyzed by plant MAPK. Introduction Infection of resistant cultivars of tobacco plants with TMV trig- gers a multi-component defense that culminates in tissue collapse at pathogen entry sites. This programmed cell death-like process, known as the Hypersensitive Response (HR), is thought to restrict systemic spread of the virus (van Loon, 1983). Resistance to TMV is mediated by the tobacco N protein of the TIR-NB- LRR class of plant disease resistance proteins (Whitham et al., 1994). Recognition of a helicase (p50) domain of the viral 126- kDa replicase by N (Mestre & Baulcombe, 2006; Ueda et al., 2006; Burch-Smith et al., 2007) initiates downstream signaling pathways that include rapid activation of two tobacco MAP kin- ases, Salicylic acid- Induced Protein Kinase (SIPK) and Woun- ding- Induced Protein Kinase (WIPK), that share a common upstream kinase, NtMEK2 (Jin et al., 2003). A body of evidence suggests that SIPK and WIPK are involved in induction of defense-related genes and HR cell death (Zhang et al., 1998, 2000; Lee et al., 2001). N-mediated resistance requires SGT1 (Liu et al., 2002b), a conserved HSP90 co-chaperone that was originally identified in Saccharomyces cerevisiae as an essential component of cell cycle progression (Kitagawa et al., 1999). In yeast, SGT1 functions as an adaptor in assembly of a number of multi-protein complexes, including kinetochore CBF3 and ubiquitin ligase SCF (Catlett & Kaplan, 2006). In plants, SGT1, HSP90 ( Heat Shock Protein 90kD) and RAR1 ( Required for Ml a Resistance 1) form a dynamic molecular chaperone complex whose role is to maintain NB-LRR receptors in an inactive but signaling-competent form (Liu et al., 2003; Bot € er et al., 2007; Zhang et al., 2010; Kadota & Shirasu, 2012). Current evidence suggests that this chaperone complex both stabilizes NB-LRR proteins and mediates their subsequent degradation (Holt et al., 2005; Leister et al., 2005; Sangster & Queitsch, 2005; Azevedo et al., 2006; Kadota & Shirasu, 2012), thus maintaining a finely tuned balance between signal transmission and attenuation. In mammals, structurally similar NOD-LRR sensors of microbial infection also require SGT1 and HSP90 for proper function (Mayor et al., 2007; da Silva Correia et al., 2007). Few protein substrates have been unequivocally identified for plant MAPKs, although large-scale protein screens have demon- strated that many substrates are likely to exist in plants (Feilner Ó 2013 The Authors New Phytologist Ó 2013 New Phytologist Trust New Phytologist (2013) 1 www.newphytologist.com Research
-
Upload
hoangxuyen -
Category
Documents
-
view
216 -
download
0
Transcript of Nucleocytoplasmic partitioning of tobacco N receptor is ...plantpath.ibb.waw.pl/pdf/New Phytologist...

Nucleocytoplasmic partitioning of tobacco N receptor ismodulated by SGT1
Rafał Hoser1, Marek _Zurczak1, Małgorzata Lichocka1, Sabina Zuzga1, Michal Dadlez1,2, Marcus A. Samuel3,
Brian E. Ellis4, Johannes Stuttmann5, Jane E. Parker5, Jacek Hennig1 and Magdalena Krzymowska1
1Institute of Biochemistry and Biophysics PAS, Pawinskiego 5a, 02-106, Warsaw, Poland; 2Biology Department, Institute of Genetics and Biotechnology, Warsaw University, Pawinskiego 5a,
02-106, Warsaw, Poland; 3Department of Biological Sciences, University of Calgary, 2500 University Dr NW Calgary, Alberta, T2N 1N4, Canada; 4Michael Smith Laboratories, University of
British Columbia, 2185 East Mall, Vancouver, BC V6T 1Z4, Canada; 5Department of Plant–Microbe Interactions, Max-Planck Institute for Plant Breeding Research, Carl-von-Linn�e-Weg 10,
50829, Cologne, Germany
Author for correspondence:Magdalena Krzymowska
Tel: + 48 22 592 5721Email: [email protected]
Received: 27 March 2013
Accepted: 29 April 2013
New Phytologist (2013)doi: 10.1111/nph.12347
Key words: disease resistance, MAPK,NB-LRR, nucleocytoplasmic shuttling, proteinphosphorylation, SGT1, tobacco (Nicotianatabacum).
Summary
� SGT1 (Suppressor of G2 allele of SKP1) is required to maintain plant disease Resistance (R)
proteins with Nucleotide-Binding (NB) and Leucine-Rich Repeat (LRR) domains in an inactive
but signaling-competent state. SGT1 is an integral component of a multi-protein network that
includes RACK1, Rac1, RAR1, Rboh, HSP90 and HSP70, and in rice the Mitogen-Activated
Protein Kinase (MAPK), OsMAPK6. Tobacco (Nicotiana tabacum) N protein, which belongs
to the Toll-Interleukin Receptor (TIR)-NB-LRR class of R proteins, confers resistance to Tobac-
co Mosaic Virus (TMV).� Following transient expression in planta, we analyzed the functional relationship between
SGT1, SIPK – a tobacco MAPK6 ortholog – and N, using mass spectrometry, confocal micros-
copy and pathogen assays.� Here, we show that tobacco SGT1 undergoes specific phosphorylation in a canonical MAPK
target-motif by SIPK. Mutation of this motif to mimic SIPK phosphorylation leads to an
increased proportion of cells displaying SGT1 nuclear accumulation and impairs N-mediated
resistance to TMV, as does phospho-null substitution at the same residue. Forced nuclear
localization of SGT1 causes N to be confined to nuclei.� Our data suggest that one mode of regulating nucleocytoplasmic partitioning of R proteins
is by maintaining appropriate levels of SGT1 phosphorylation catalyzed by plant MAPK.
Introduction
Infection of resistant cultivars of tobacco plants with TMV trig-gers a multi-component defense that culminates in tissue collapseat pathogen entry sites. This programmed cell death-like process,known as the Hypersensitive Response (HR), is thought torestrict systemic spread of the virus (van Loon, 1983). Resistanceto TMV is mediated by the tobacco N protein of the TIR-NB-LRR class of plant disease resistance proteins (Whitham et al.,1994). Recognition of a helicase (p50) domain of the viral 126-kDa replicase by N (Mestre & Baulcombe, 2006; Ueda et al.,2006; Burch-Smith et al., 2007) initiates downstream signalingpathways that include rapid activation of two tobacco MAP kin-ases, Salicylic acid-Induced Protein Kinase (SIPK) and Woun-ding-Induced Protein Kinase (WIPK), that share a commonupstream kinase, NtMEK2 (Jin et al., 2003). A body of evidencesuggests that SIPK and WIPK are involved in induction ofdefense-related genes and HR cell death (Zhang et al., 1998,2000; Lee et al., 2001).
N-mediated resistance requires SGT1 (Liu et al., 2002b), aconserved HSP90 co-chaperone that was originally identified in
Saccharomyces cerevisiae as an essential component of cell cycleprogression (Kitagawa et al., 1999). In yeast, SGT1 functions asan adaptor in assembly of a number of multi-protein complexes,including kinetochore CBF3 and ubiquitin ligase SCF (Catlett &Kaplan, 2006). In plants, SGT1, HSP90 (Heat Shock Protein90kD) and RAR1 (Required for Mla Resistance 1) form adynamic molecular chaperone complex whose role is to maintainNB-LRR receptors in an inactive but signaling-competent form(Liu et al., 2003; Bot€er et al., 2007; Zhang et al., 2010; Kadota &Shirasu, 2012). Current evidence suggests that this chaperonecomplex both stabilizes NB-LRR proteins and mediates theirsubsequent degradation (Holt et al., 2005; Leister et al., 2005;Sangster & Queitsch, 2005; Azevedo et al., 2006; Kadota &Shirasu, 2012), thus maintaining a finely tuned balance betweensignal transmission and attenuation. In mammals, structurallysimilar NOD-LRR sensors of microbial infection also requireSGT1 and HSP90 for proper function (Mayor et al., 2007; daSilva Correia et al., 2007).
Few protein substrates have been unequivocally identified forplant MAPKs, although large-scale protein screens have demon-strated that many substrates are likely to exist in plants (Feilner
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 1
www.newphytologist.com
Research

et al., 2005; Popescu et al., 2009) and that some of these areinvolved in controlling disease (Andreasson et al., 2005; Menkeet al., 2005; Bethke et al., 2009; Popescu et al., 2009). Also, celldeath elicited in Nicotiana benthamiana by transient overexpres-sion of AtMKK7 and AtMKK9, two defense-related MAPKkinases from Arabidopsis thaliana, depends on SGT1 (Popescuet al., 2009). Although previous studies have demonstrated anessential role for SIPK in N-gene mediated TMV resistance(Jin et al., 2003), it has remained unclear how SIPK and itsMAPK6 orthologs execute this function. Here we provideevidence that N. benthamiana SGT1 undergoes specific phos-phorylation by SIPK, thus establishing that a protein known toplay a central role in the plant defense is also the direct target of aMAPK cascade. We further show that SIPK phosphorylationof SGT1 is required for functional N-mediated resistance.We propose that phosphorylation of SGT1 fine-tunes thenucleocytoplasmic distribution of the N-receptor, a process thatis necessary for an effective plant resistance response to TMVinfection.
Materials and Methods
Plant material
Nicotiana tabacum L. cv Xanthi-nc plants of all genotypes, wild-type nn and transgenic NN N. benthamiana plants (BN3) weregrown in soil under controlled environmental conditions (21°C,16 h light, 8 h dark), as described previously (Talarczyk et al.,2002).
Recombinant protein preparation
The full-length cDNA of NbSGT1 (AY899199) was cloned intothe pET-28a vector. The full-length cDNA of SIPK was clonedinto the pGEX-6P-1 vector. Full-length cDNAs of NtMEK2DD
and NtMEK2KR were cloned into the pGEX-6P-2 vector.Escherichia coli (BL21) cells were induced with 0.25 mM IPTGat 18°C for 4 h. His-tagged recombinant protein was purifiedusing Ni-NTA resin (Qiagen). Glutathione S-transferase (GST)-tagged recombinant proteins were purified using Glutathione-Agarose resin (Sigma-Aldrich).
Protein analysis
Proteins were extracted in buffer containing 100 mM Tris HClpH 8.0, 1 mM EDTA, 150 mM NaCl, 7.2 mM b-mercaptoetha-nol, 0.5 mM 4-(2-aminoethyl) benzenesulfonyl fluoride hydro-chloride (AEBSF) and 0.03 lM PMSF (phenylmethylsulfonylfluoride). Protein content was measured by the Bradford methodusing a commercially available reagent (Bio-Rad, www.bio-rad.com). Extracts were fractionated by 12.5% SDS–PAGE and sub-jected to immunoblot analysis using specific primary antibodies:monoclonal mouse anti-c-myc (Sigma-Aldrich), polyclonal rab-bit anti-SGS or anti-TMV (Bioreba, Reinach BL1, Switzerland)and appropriate alkaline phosphatase-conjugated anti-mouse oranti-rabbit antibodies from Sigma-Aldrich. Immunoblots were
developed using the NBT/BCIP colorimetric detection kit fromRoche Applied Science.
In vitro phosphorylation assay
The purified recombinant proteins (NbSGT1, SIPK,NtMEKDD or NtMEK2KR; c. 2 lg each) were incubated with50 lM ATP supplemented with 1.5 lCi [c32P] ATP in the reac-tion buffer (20 mM Tris HCl pH 7.5, 40 mM MgCl2, 10 mMEGTA) at 30°C for 30 min. The reaction was terminated byadding 39 Laemmli sample buffer. In the nonradioactive reac-tion followed by mass spectrometry analysis, the concentrationof ATP was increased to 200 lM and the incubation timeextended to 2 h.
Strep-tag II affinity purification
The full-length cDNA of NbSGT1 was cloned into the pROK2vector with a (6xHis)-(Ala-Ala)-(Strep-tag II) coding sequence atthe 3′-end. The full-length cDNA of SIPK was cloned into thepROK2 vector. The pROK2 constructs and pTA7002 carryingcDNA of NtMEK2DD or NtMEK2KR were electroporated intoAgrobacterium tumefaciens (GV3101) cells, which were infiltratedinto 4-wk-old N. benthamiana leaves as described earlier (Romeiset al., 2001). Expression of the NtMEK2 variant cDNAs wasinduced by 30 lM dexamethasone 40–48 h later (Yang et al.,2001). Treated leaves were collected 5 h after dexamethasoneinfiltration. Ground leaf material (c. 3 g) was thawed in 10 mlEx-strep buffer (100 mM Tris HCl pH 8.0; 5 mM EGTA; 5 mMEDTA; 150 mM NaCl; 10 mM DTT; 0.5 mM AEBSF;5 lg ml�1 antipain; 5 lg ml�1 leupeptin; 50 mM NaF; 1% (v/v)Phosphatase Inhibitor Cocktail 1 (Sigma-Aldrich); 0.5% TritonX-100; and 100 lg ml�1 avidin) as described previously (Witteet al., 2004). The slurry was centrifuged for 10 min at 4°C(15 000 g), the supernatant filtered through Miracloth and 0.5 mlStrepTactin Sepharose (IBA GmbH, G€ottingen, Germany) wasadded. Binding was performed by incubation of this suspensionon a rotator for 1 h at 4°C. The slurry was transferred into aPoly-Prep column (Bio-Rad) and the flow-through was dis-carded. The resin was washed twice with 10 ml W-buffer(100 mM Tris HCl pH 8.0; 150 mM NaCl; 1 mM EDTA).Four times 250 ll E-buffer (100 mM Tris HCl pH 8.0; 150 mMNaCl; 1 mM EDTA; 2.5 mM desthiobiotin) was added and elu-ates were collected. The samples were concentrated on a Micro-con YM-10 (Millipore) for 30 min at 4°C (13 000 g) to a volumeof c. 20 ll and resolved by SDS-PAGE.
Mass spectrometry analysis
Gel bands containing the proteins of interest were subjected to astandard proteomic procedure as described in Supporting Infor-mation Methods S1. Briefly, reduced and alkylated proteins weresubjected to trypsin digestion. The resulting peptides were elutedfrom the gel and phosphopeptide enrichment was carried out onSwellGel Gallium-Chelated Discs (Phosphopeptide Isolation Kit,Thermo Fisher Scientific Rockford, IL, USA). LC-MS analyses
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

of the peptide mixtures were carried out on LTQ FTICR spec-trometer (Thermo). The MASCOT program was used for data-base searches, and MS/MS spectra of phosphorylated peptideswere also curated manually.
Heterologous transient complementation assay
For VIGS of NbSGT1, the TRV-based vector system wasadopted (Liu et al., 2002b). An NbSGT1 cDNA fragment wasPCR amplified using primers that add specific restriction sites(bold): TCTAGACGAGACATTTACAAAGATGCCG andGGATCCAGGGCTTCCTTCGACCTTCTTT and ligatedinto the pTRV2 vector. For the GFP construct, the first 130 bpof the GFP were PCR-amplified using primers that add specificrestriction sites (bold): GGATCCATGGTAGATCTGACTAGTAAAGG and CTCGAGGTATGTTGCATCACCTTCACCand cloned into the pTRV2 vector. To generate phospho-null(AP) and phospho-mimic (DP) variants of AtSGT1b by PCR,primers were designed that incorporated mutations T346A andT346D, respectively. Next, wild-type and both phospho-variantsof AtSGT1b cDNA were amplified by PCR with an upstreamprimer AGATCTGCTATGGAACAAAAGCTTATTTCTGAAGAAGACTTGCTCGAGATGGCCAAGGAATTAGCAGAGAAAGCT adding restriction sites and c-myc encoding sequence(bold) to the 5′ end, and a downstream primer GGTACCCGGGTCGACTCGAGTCAATACTCCCACTTCTTGAGCTCCAT adding restriction sites (bold) to the 3′ end of the product.All three inserts were cloned into the pROK2 vector. For silenc-ing of NbSGT1, Agrobacterium cultures (OD600 = 0.5) contain-ing pTRV1 or pTRV2 derivative plasmids were mixed at a 1 : 1ratio and infiltrated into the lower leaves of four-leaf-stageN. benthamiana plants using a 1-ml needleless syringe. For tran-sient expression of wild-type and phospho-variants of AtSGT1b,the systemic leaves of the plants silenced for NbSGT1 were infil-trated with Agrobacterium cultures containing c-myc-AtSGT1bconstructs. Twenty-four hours later, the same leaves were dustedwith carborundum and rubbed with TMV strain U1 suspension(1 lg ml�1). To detect the presence of TMV, total proteinsextracted from inoculated leaves 4 d after virus infection wereanalyzed by Western blots. Each experiment was repeated fourtimes, and each experiment included at least three independentbiological replicates.
Subcellular localization of AtSGT1b
The pCambia1302 vector was used to express AtSGT1b proteinvariants N-terminally tagged with GFP. cDNAs encodingwild-type, AP and DP variants of AtSGT1b (with T346A andT346D substitutions, respectively) were amplified by PCR withan upstream primer adding a NheI restriction site to the 5′ end,and a downstream primer adding a PmlI restriction site to the 3′end of the product. As controls, wild-type AtSGT1b cDNAwas PCR-amplified with the upstream primers adding a NheIrestriction site and ‘NLSSV40’ (PKKKRKV), ‘NLSSLCV’ (SYVKTVPNRTRTYIK), ‘NES’ (NELALKLAGLDINK) or ‘nes’ (NEL-ALKAAGADANK) encoding sequences to the 5′ ends of the
products, and the downstream primer as above. NcoI/NheI GFPand NheI/PmlI AtSGT1b inserts were cloned into the NcoI/PmlIpCambia1302 vector by a triple-ligation strategy.
In order to determine the subcellular localization of GFP-AtS-GT1b fusion proteins, leaves from 4-wk-old N. benthamianaplants were used for transient gene expression in epidermal cells.Plasmid DNA (2 lg) was adsorbed onto tungsten M17 particles(diameter 1.1 lm, 350 lg). Micro-bombardment was performedat a pressure of 1100 psi using a biolistic particle delivery system(model PDS-1000/He, Bio-Rad), and tissues were analyzed 24 hafter bombardment.
Hypersensitive response assay
In order to obtain N-NXS constructs, the Citrine sequence wasamplified with an upstream primer that added a SacI restrictionsite and a sequence encoding the last three amino acids (ING) ofthe N to the 5’ ends of the products, and downstream primersthat added ‘NLSSLCV’ ‘NLSSV40’ or ‘nlsSV40’ encoding sequenceswith a STOP-codon and a SacI restriction site to the 3′ ends ofthe products. The SacI inserts were then cloned into the SacIrestriction site of gN (Dinesh-Kumar et al., 2000).
In order to generate p50-U1-Cerulean-NLS and p50-U1-Cerulean-nls, the p50-U1-Cerulean construct (Burch-Smithet al., 2007) was PCR-amplified with primers including‘NLSSV40’ or ‘nlsSV40’ and the inserts were cloned into thepROK2 vector. The N and p50 derivative constructs were elec-troporated into A. tumefaciens (GV3101) cells, which were infil-trated into 4-wk-old N. benthamiana leaves, at OD600 = 1.7 and1.0, respectively. For co-infiltrations, Agrobacterium strains carry-ing N and p50 derivative constructs were mixed at a 1:1 ratio. Toenhance the expression levels, the infiltration suspensions weremixed at 10:1 ratio with an Agrobacterium suspension(OD600 = 2–3) expressing p19 (Witte et al., 2004). Photographsof leaves were taken 9 or 10 d after infiltration.
Co-localization experiments
The pGWB 445 vector (Nakagawa et al., 2007a,b) was used toexpress AtSGT1b protein variants N-terminally tagged withCFP. cDNAs encoding wild-type, AP, DP and variants contain-ing ‘NLSSV40’ (PKKKRKV), ‘nlsSV40’ (PKTKRKV), ‘NES’ and‘nes’ coding sequences were amplified by PCR to produce blunt-end products for TOPO Cloning (pENTR/D-TOPO vector,Invitrogen). The resulting entry clones were LR recombined withthe Gateway pGWB 445 destination vector. cDNAs encodingTIR (amino acids from 1 to 179) or NB domains (amino acidsfrom 180 to 525) were amplified by PCR with primers contain-ing attB sequences. As a template for subcloning of the TIRdomain, a cDNA fragment consisting of exon 1 and exon 2 ofthe N gene from Nicotiana glutinosa was used. attB-flanked PCRproducts were BP-recombined with the pDONR201 donor vec-tor (Invitrogen). Subsequently generated entry clones were LR-recombined with the Gateway pGWB 441 destination vector toexpress TIR or NB domain polypeptides C-terminally taggedwith YFP. To obtain a cDNA encoding the N LRR domain
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 3

(amino acids from 589 to 1144), a fragment of exon 3 of the Ngene from N. glutinosa was PCR-amplified with an upstreamprimer adding an EcoRI site to the 5′ end of the product and adownstream primer adding the sequence of this domain fromexon 4 (up to BglII site) to the 3′ end of the product. Next, thesequence of exon 4 was PCR-amplified with the upper primerstarting from the beginning of this sequence and the lower primeradding the sequence of exon 5 and a XhoI site to the 3′ end ofthe product. EcoRI/BglII exon 3 and BglII/XhoI exon 4+5 insertswere cloned into EcoRI/XhoI pGEX-6P-1 vector by a triple-ligation strategy. The coding sequence of the LRR domain wasPCR-amplified to produce a blunt-end product for TOPO Clon-ing (pENTRTM/D-TOPO vector). The resulting entry clone wasLR-recombined with the Gateway pGWB 442 destination vectorto express the LRR domain N-terminally tagged with YFP. Themixture of CFP-AtSGT1b variants (1.5 lg) with either one ofthree YFP-N domain constructs (YFP-TIR, YFP-NB orYFP-LRR; 1.5 lg) and p19 (0.5 lg) plasmid DNAs was used formicrobombardment and tissue samples were analyzed 24–48 hafterwards.
For co-localization of AtSGT1b with N, a combination ofCFP-AtSGT1b variants (1.5 lg) with N-Citrine (1.5 lg) andp19 (0.5 lg) plasmid DNAs was used for microbombardmentand tissue samples were analyzed after 24–48 h.
Confocal laser scanning fluorescence microscopy
Transient intracellular fluorescence was observed by confocallaser scanning microscopy using a Nikon TE2000E EZ-C1inverted confocal microscope equipped with 609 oil immersionobjective lens (numerical aperture = 1.4). YFP, Citrine and GFPwere excited with the 488 nm line from an argon ion laser, andimages of CFP and Cerulean were obtained using 408 nm diodelaser excitation. The fluorescence signals were detected using the515/30 and 450/35 emission filters for the first and latter fluoro-phore groups, respectively. Scanning was performed in sequentialmode to prevent bleed-through. Images were collected from asingle optical section and processed using the EZ C1 program(Nikon Instruments B.V. Europe, Amstelveen, the Netherlands).Optimal imaging parameters were set up for each experimentand were equal for each image dataset. Quantification of fluores-cence intensities in the nuclear and cytoplasmic regions was per-formed using ImageJ software (Abramoff et al., 2004).
Results
SGT1 is phosphorylated by SIPK in vitro and in planta
Genetic analyses have linked SGT1 with diverse biological pro-cesses in plants, and the SGT1 protein was found to form part ofa multi-protein functional network that includes a rice MAPK,OsMAPK6 (Thao et al., 2007), whose tobacco ortholog is SIPK(NtSIPK). Amino acid sequence analysis of eukaryotic SGT1sequences (Fig. S1) revealed a canonical MAPK phosphorylationmotif (S/T-P) in the C-terminal SGS (SGT1-Specific) domain ofNbSGT1 and most known plant SGT1 proteins, as well as in the
Saccharomyces cerevisiae Sgt1p. In addition, NbSGT1 contains apredicted MAPK docking site of medium stringency (scansitealgorithm, http://scansite.mit.edu).
These observations prompted us to determine whether SGT1can be phosphorylated by SIPK. When recombinant SIPK wasco-incubated with either constitutively active NtMEK2DD or thecatalytically inactive form, NtMEK2KR (Jin et al., 2003), SIPKactivated by NtMEK2DD was able to phosphorylate the controlsubstrate (myelin basic protein, Fig. 1a, lane 4) and was also ableto phosphorylate recombinant NbSGT1 (Fig. 1a, lane 5). Twophosphopeptides derived from the C-terminal part of SIPK-
(a)
(b)
Fig. 1 Salicylic acid-Induced Protein Kinase (SIPK) phosphorylates SGT1(Suppressor of G2 allele of SKP1) in vitro. (a) Recombinant NbSGT1 with aC-terminal 6xHis epitope, as well as SIPK and its upstream kinase MEK2tagged with N-terminal Glutathione S-transferase (GST) epitopes, wereincubated with [c32P]ATP, resolved by SDS-PAGE and analyzed byautoradiography. Lanes 4 and 8 include myelin basic protein (MBP) as apositive control; for lanes 1, 3, 5, 7 and 9, SGT1 served as a substrate. Theband corresponding to phosphorylated NbSGT1 is designated by anasterisk. Constitutively active form of MEK2 (MEK2DD); catalyticallyinactive form of MEK2 (MEK2KR), heat-denatured SIPK (SIPKInactive). As aloading control, the Coomassie Brilliant Blue (CBB)-stained gel is shown inthe bottom panel. (b) Overall SGT1 structure showing phosphopeptides.In a parallel experiment, performed as above but with nonradioactive ATP,the band corresponding to SGT1 was excised from the Coomassie-stainedgel and subjected to LC-MS-MS/MS analysis. Two phosphorylatedpeptides identified by the MASCOT program are shown, and thephosphorylated S358 residue is underlined. The same analysis revealed dualphosphorylation of the TEY activation loop of SIPK (Fig. S2). Theexperiment was performed three times with similar results.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4

treated NbSGT1 were detected by mass spectrometry(LC-MS-MS/MS), and Ser358 within the KVEGSPPDGMELK-peptide that includes the predicted MAPK phosphorylation site(Fig. 1b) was unambiguously identified as phosphorylated(Fig. S2).
A transient expression assay was used to determine whetherSGT1 could also be phosphorylated in planta. Agrobacteriumstrains carrying either 35S::NbSGT1-Strep II – a dexamethasone(Dex)-inducible form of NtMEK2DD – or 35S::SIPK constructswere co-infiltrated into Nicotiana benthamiana leaves and after2 d incubation to ensure expression of the constitutive promoterconstructs, NtMEK2DD expression was induced by treatment ofthe infiltrated leaves with Dex (Fig. 2a). Protein was extractedfrom leaf samples collected 5 h after induction and SGT1 wasaffinity-purified via its Strep tag II. Only one phosphopeptide,that encompassing Ser358, was detected by mass spectrometryunder these conditions (Fig. 2b). As a control, we co-expressedkinase inactive NtMEK2KR with SGT1. In this case, SIPK wasnot included because its transient expression results in slightlyincreased SIPK activity and associated activation of defenseresponses (Zhang & Liu, 2001). Under these conditions, SGT1phosphorylation was not detected (Fig. 2c). We therefore
concluded that SGT1 can also be phosphorylated by SIPK invivo in N. benthamiana cells. However, we cannot fully excludethe possibility of SGT1 being phosphorylated by an endogenousN. benthamiana kinase.
(a)
(b)
(c)
Fig. 2 Salicylic acid-Induced Protein Kinase (SIPK) phosphorylates NbSGT1in planta. (a) NbSGT1 protein carrying a C-terminal Strep-tag II epitopewas transiently co-expressed with MEK2DD and SIPK in Nicotiana
benthamiana leaves. SGT1 (Suppressor of G2 allele of SKP1) was thenaffinity-purified, separated by SDS-PAGE and stained with CoomassieBlue. The excised bands corresponding to SGT1 were subjected to massspectrometric analysis. (b) Part of the fragmentation spectra of the twopeptides identified in the IMAC flow-through and bound fractions aspeptide KVEGSPPDGMELK either in the unphosphorylated form (upperpanel, flow through fraction – parent mass 1386.69 Da –MASCOT score92) or the phosphorylated form (lower panel, retentate fraction – parentmass 1466.66 Da –MASCOT score 58), in samples from leaves co-expressing SGT1, SIPK and MEK2DD. The region of the MS/MS spectrumfrom 770 to 1350m/z is shown because it contains the majority of thedaughter ion signals which allow for the unequivocal identification of thesite of SGT1 phosphorylation as S358. All major signals of the MS/MSspectra are identified by their corresponding fragment tags of the b, yseries but also of the y-H2O, b-H2O and, for the phosphorylated version(lower panel), the y-H3PO4 and b-H3PO4 series, expected in the case of aphosphorylated peptide. The dashed vertical lines indicate the massincrements of the y series ions of five consecutive amino acids denoted V,E, G, S, P, clearly pointing to the fragment mass increase of 80 Da at serine(pS) residue (note the shifted position of the y9, y10, y11 and y12 signals ascompared to the unchanged position of y7, y8 and shifted b8, b11 and b12signals). The positions of some of the signals overlap (b12 and y12-H2O,b11-H3PO4 and y10, y12-H3PO4 and y11) but the remaining unique signalsallow unambiguous localization of the modification. (c) In control samples,where SGT1 and MEK2KR were co-expressed without SIPK, nophosphorylated SGT1 peptides were detected. The fragmentationspectrum of nonphosphorylated KVEGSPPDGMELK peptide derived fromSGT1 transiently expressed in planta. Fragmentation (MS/MS) spectrumof the peptide of parent mass 1368.64 Da with all major peaks assigned toy, b or y-H2O, b-H2O (horizontal arrows) series of ions expected forpeptide of sequence KVEGSPPDGMELK. This analysis unequivocallyconfirms the peptide identity listed in MASCOT output file. Theinterpretation of the fragmentation spectrum in terms of the underlyingpeptide sequence is given in one-letter code for y-series and b-series ions.The experiment was performed three times with similar results.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 5
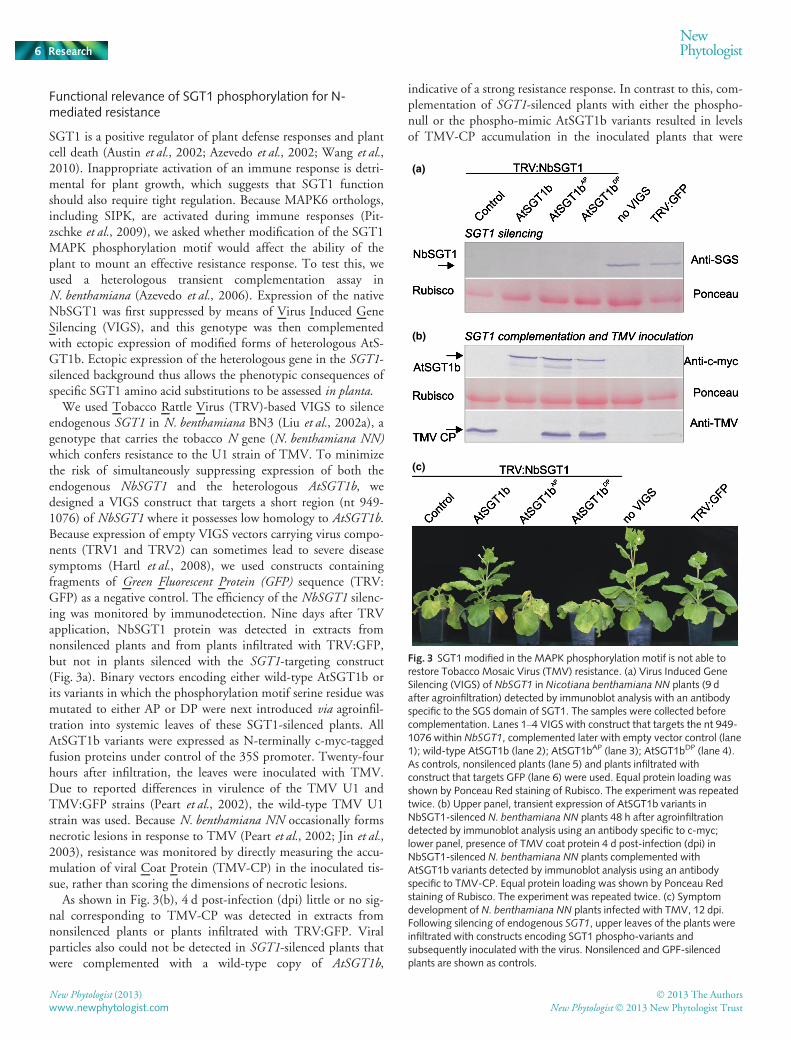
Functional relevance of SGT1 phosphorylation for N-mediated resistance
SGT1 is a positive regulator of plant defense responses and plantcell death (Austin et al., 2002; Azevedo et al., 2002; Wang et al.,2010). Inappropriate activation of an immune response is detri-mental for plant growth, which suggests that SGT1 functionshould also require tight regulation. Because MAPK6 orthologs,including SIPK, are activated during immune responses (Pit-zschke et al., 2009), we asked whether modification of the SGT1MAPK phosphorylation motif would affect the ability of theplant to mount an effective resistance response. To test this, weused a heterologous transient complementation assay inN. benthamiana (Azevedo et al., 2006). Expression of the nativeNbSGT1 was first suppressed by means of Virus Induced GeneSilencing (VIGS), and this genotype was then complementedwith ectopic expression of modified forms of heterologous AtS-GT1b. Ectopic expression of the heterologous gene in the SGT1-silenced background thus allows the phenotypic consequences ofspecific SGT1 amino acid substitutions to be assessed in planta.
We used Tobacco Rattle Virus (TRV)-based VIGS to silenceendogenous SGT1 in N. benthamiana BN3 (Liu et al., 2002a), agenotype that carries the tobacco N gene (N. benthamiana NN)which confers resistance to the U1 strain of TMV. To minimizethe risk of simultaneously suppressing expression of both theendogenous NbSGT1 and the heterologous AtSGT1b, wedesigned a VIGS construct that targets a short region (nt 949-1076) of NbSGT1 where it possesses low homology to AtSGT1b.Because expression of empty VIGS vectors carrying virus compo-nents (TRV1 and TRV2) can sometimes lead to severe diseasesymptoms (Hartl et al., 2008), we used constructs containingfragments of Green Fluorescent Protein (GFP) sequence (TRV:GFP) as a negative control. The efficiency of the NbSGT1 silenc-ing was monitored by immunodetection. Nine days after TRVapplication, NbSGT1 protein was detected in extracts fromnonsilenced plants and from plants infiltrated with TRV:GFP,but not in plants silenced with the SGT1-targeting construct(Fig. 3a). Binary vectors encoding either wild-type AtSGT1b orits variants in which the phosphorylation motif serine residue wasmutated to either AP or DP were next introduced via agroinfil-tration into systemic leaves of these SGT1-silenced plants. AllAtSGT1b variants were expressed as N-terminally c-myc-taggedfusion proteins under control of the 35S promoter. Twenty-fourhours after infiltration, the leaves were inoculated with TMV.Due to reported differences in virulence of the TMV U1 andTMV:GFP strains (Peart et al., 2002), the wild-type TMV U1strain was used. Because N. benthamiana NN occasionally formsnecrotic lesions in response to TMV (Peart et al., 2002; Jin et al.,2003), resistance was monitored by directly measuring the accu-mulation of viral Coat Protein (TMV-CP) in the inoculated tis-sue, rather than scoring the dimensions of necrotic lesions.
As shown in Fig. 3(b), 4 d post-infection (dpi) little or no sig-nal corresponding to TMV-CP was detected in extracts fromnonsilenced plants or plants infiltrated with TRV:GFP. Viralparticles also could not be detected in SGT1-silenced plants thatwere complemented with a wild-type copy of AtSGT1b,
indicative of a strong resistance response. In contrast to this, com-plementation of SGT1-silenced plants with either the phospho-null or the phospho-mimic AtSGT1b variants resulted in levelsof TMV-CP accumulation in the inoculated plants that were
(a)
(b)
(c)
Fig. 3 SGT1 modified in the MAPK phosphorylation motif is not able torestore Tobacco Mosaic Virus (TMV) resistance. (a) Virus Induced GeneSilencing (VIGS) of NbSGT1 in Nicotiana benthamiana NN plants (9 dafter agroinfiltration) detected by immunoblot analysis with an antibodyspecific to the SGS domain of SGT1. The samples were collected beforecomplementation. Lanes 1–4 VIGS with construct that targets the nt 949-1076 within NbSGT1, complemented later with empty vector control (lane1); wild-type AtSGT1b (lane 2); AtSGT1bAP (lane 3); AtSGT1bDP (lane 4).As controls, nonsilenced plants (lane 5) and plants infiltrated withconstruct that targets GFP (lane 6) were used. Equal protein loading wasshown by Ponceau Red staining of Rubisco. The experiment was repeatedtwice. (b) Upper panel, transient expression of AtSGT1b variants inNbSGT1-silenced N. benthamiana NN plants 48 h after agroinfiltrationdetected by immunoblot analysis using an antibody specific to c-myc;lower panel, presence of TMV coat protein 4 d post-infection (dpi) inNbSGT1-silenced N. benthamiana NN plants complemented withAtSGT1b variants detected by immunoblot analysis using an antibodyspecific to TMV-CP. Equal protein loading was shown by Ponceau Redstaining of Rubisco. The experiment was repeated twice. (c) Symptomdevelopment of N. benthamiana NN plants infected with TMV, 12 dpi.Following silencing of endogenous SGT1, upper leaves of the plants wereinfiltrated with constructs encoding SGT1 phospho-variants andsubsequently inoculated with the virus. Nonsilenced and GPF-silencedplants are shown as controls.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6
approximately equal to those seen in the empty vector controls(Fig. 3b). Moreover, plants complemented with mutated AtS-GT1b proteins or empty vector control, in contrast to plantsexpressing wild-type AtSGT1b, developed macroscopic diseasesymptoms manifested as chlorotic lesions and tissue desiccation(Fig. 3c). We concluded that neither the phospho-mimic nor thephospho-null variant of AtSGT1b is functional in N-mediatedresistance. This pattern of increased susceptibility is not a conse-quence of different levels of expression of the AtSGT1bphospho-variants (Fig. 3b). Notably, we found that the phospho-mimic and phospho-null alleles of AtSGT1b were still able tocomplement cell division defects of yeast sgt1-3 and sgt1-5mutant strains (Fig. S3), which argues against general loss offunction of these modified proteins. Because both the phospho-mimic and phospho-null AtSGT1b variants failed tocomplement NbSGT1 function, we reasoned that either thesimultaneous presence of both phospho-forms of SGT1 (nulland mimic) or dynamic changes in the phosphorylation state of
SGT1might be an important feature of N-mediated resistance toTMV.
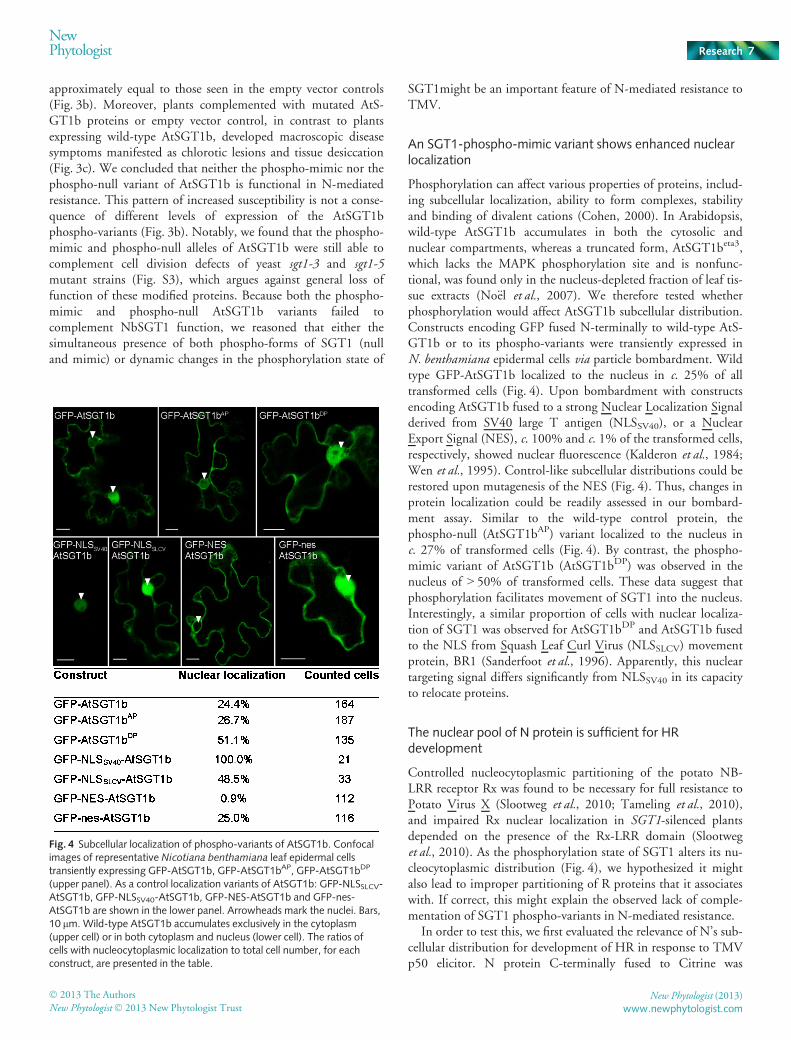
An SGT1-phospho-mimic variant shows enhanced nuclearlocalization
Phosphorylation can affect various properties of proteins, includ-ing subcellular localization, ability to form complexes, stabilityand binding of divalent cations (Cohen, 2000). In Arabidopsis,wild-type AtSGT1b accumulates in both the cytosolic andnuclear compartments, whereas a truncated form, AtSGT1beta3,which lacks the MAPK phosphorylation site and is nonfunc-tional, was found only in the nucleus-depleted fraction of leaf tis-sue extracts (No€el et al., 2007). We therefore tested whetherphosphorylation would affect AtSGT1b subcellular distribution.Constructs encoding GFP fused N-terminally to wild-type AtS-GT1b or to its phospho-variants were transiently expressed inN. benthamiana epidermal cells via particle bombardment. Wildtype GFP-AtSGT1b localized to the nucleus in c. 25% of alltransformed cells (Fig. 4). Upon bombardment with constructsencoding AtSGT1b fused to a strong Nuclear Localization Signalderived from SV40 large T antigen (NLSSV40), or a NuclearExport Signal (NES), c. 100% and c. 1% of the transformed cells,respectively, showed nuclear fluorescence (Kalderon et al., 1984;Wen et al., 1995). Control-like subcellular distributions could berestored upon mutagenesis of the NES (Fig. 4). Thus, changes inprotein localization could be readily assessed in our bombard-ment assay. Similar to the wild-type control protein, thephospho-null (AtSGT1bAP) variant localized to the nucleus inc. 27% of transformed cells (Fig. 4). By contrast, the phospho-mimic variant of AtSGT1b (AtSGT1bDP) was observed in thenucleus of > 50% of transformed cells. These data suggest thatphosphorylation facilitates movement of SGT1 into the nucleus.Interestingly, a similar proportion of cells with nuclear localiza-tion of SGT1 was observed for AtSGT1bDP and AtSGT1b fusedto the NLS from Squash Leaf Curl Virus (NLSSLCV) movementprotein, BR1 (Sanderfoot et al., 1996). Apparently, this nucleartargeting signal differs significantly from NLSSV40 in its capacityto relocate proteins.
The nuclear pool of N protein is sufficient for HRdevelopment
Controlled nucleocytoplasmic partitioning of the potato NB-LRR receptor Rx was found to be necessary for full resistance toPotato Virus X (Slootweg et al., 2010; Tameling et al., 2010),and impaired Rx nuclear localization in SGT1-silenced plantsdepended on the presence of the Rx-LRR domain (Slootweget al., 2010). As the phosphorylation state of SGT1 alters its nu-cleocytoplasmic distribution (Fig. 4), we hypothesized it mightalso lead to improper partitioning of R proteins that it associateswith. If correct, this might explain the observed lack of comple-mentation of SGT1 phospho-variants in N-mediated resistance.
In order to test this, we first evaluated the relevance of N’s sub-cellular distribution for development of HR in response to TMVp50 elicitor. N protein C-terminally fused to Citrine was
Fig. 4 Subcellular localization of phospho-variants of AtSGT1b. Confocalimages of representative Nicotiana benthamiana leaf epidermal cellstransiently expressing GFP-AtSGT1b, GFP-AtSGT1bAP, GFP-AtSGT1bDP
(upper panel). As a control localization variants of AtSGT1b: GFP-NLSSLCV-AtSGT1b, GFP-NLSSV40-AtSGT1b, GFP-NES-AtSGT1b and GFP-nes-AtSGT1b are shown in the lower panel. Arrowheads mark the nuclei. Bars,10 lm. Wild-type AtSGT1b accumulates exclusively in the cytoplasm(upper cell) or in both cytoplasm and nucleus (lower cell). The ratios ofcells with nucleocytoplasmic localization to total cell number, for eachconstruct, are presented in the table.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 7
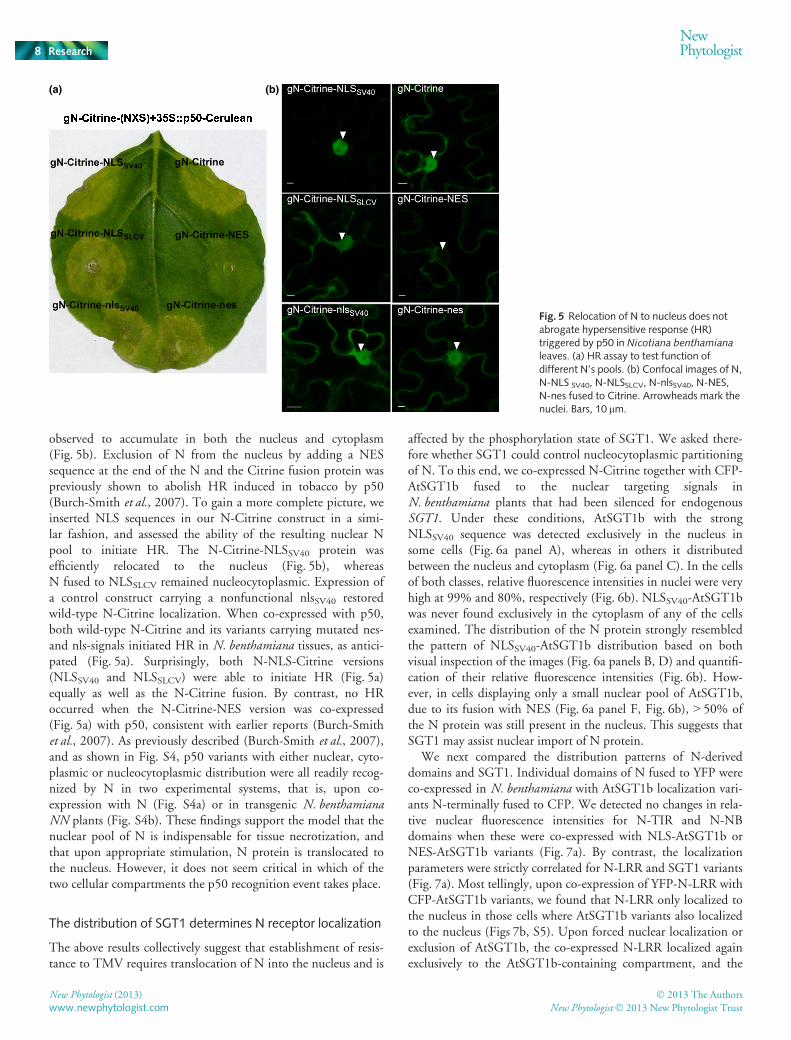
observed to accumulate in both the nucleus and cytoplasm(Fig. 5b). Exclusion of N from the nucleus by adding a NESsequence at the end of the N and the Citrine fusion protein waspreviously shown to abolish HR induced in tobacco by p50(Burch-Smith et al., 2007). To gain a more complete picture, weinserted NLS sequences in our N-Citrine construct in a simi-lar fashion, and assessed the ability of the resulting nuclear Npool to initiate HR. The N-Citrine-NLSSV40 protein wasefficiently relocated to the nucleus (Fig. 5b), whereasN fused to NLSSLCV remained nucleocytoplasmic. Expression ofa control construct carrying a nonfunctional nlsSV40 restoredwild-type N-Citrine localization. When co-expressed with p50,both wild-type N-Citrine and its variants carrying mutated nes-and nls-signals initiated HR in N. benthamiana tissues, as antici-pated (Fig. 5a). Surprisingly, both N-NLS-Citrine versions(NLSSV40 and NLSSLCV) were able to initiate HR (Fig. 5a)equally as well as the N-Citrine fusion. By contrast, no HRoccurred when the N-Citrine-NES version was co-expressed(Fig. 5a) with p50, consistent with earlier reports (Burch-Smithet al., 2007). As previously described (Burch-Smith et al., 2007),and as shown in Fig. S4, p50 variants with either nuclear, cyto-plasmic or nucleocytoplasmic distribution were all readily recog-nized by N in two experimental systems, that is, upon co-expression with N (Fig. S4a) or in transgenic N. benthamianaNN plants (Fig. S4b). These findings support the model that thenuclear pool of N is indispensable for tissue necrotization, andthat upon appropriate stimulation, N protein is translocated tothe nucleus. However, it does not seem critical in which of thetwo cellular compartments the p50 recognition event takes place.
The distribution of SGT1 determines N receptor localization
The above results collectively suggest that establishment of resis-tance to TMV requires translocation of N into the nucleus and is
affected by the phosphorylation state of SGT1. We asked there-fore whether SGT1 could control nucleocytoplasmic partitioningof N. To this end, we co-expressed N-Citrine together with CFP-AtSGT1b fused to the nuclear targeting signals inN. benthamiana plants that had been silenced for endogenousSGT1. Under these conditions, AtSGT1b with the strongNLSSV40 sequence was detected exclusively in the nucleus insome cells (Fig. 6a panel A), whereas in others it distributedbetween the nucleus and cytoplasm (Fig. 6a panel C). In the cellsof both classes, relative fluorescence intensities in nuclei were veryhigh at 99% and 80%, respectively (Fig. 6b). NLSSV40-AtSGT1bwas never found exclusively in the cytoplasm of any of the cellsexamined. The distribution of the N protein strongly resembledthe pattern of NLSSV40-AtSGT1b distribution based on bothvisual inspection of the images (Fig. 6a panels B, D) and quantifi-cation of their relative fluorescence intensities (Fig. 6b). How-ever, in cells displaying only a small nuclear pool of AtSGT1b,due to its fusion with NES (Fig. 6a panel F, Fig. 6b), > 50% ofthe N protein was still present in the nucleus. This suggests thatSGT1 may assist nuclear import of N protein.
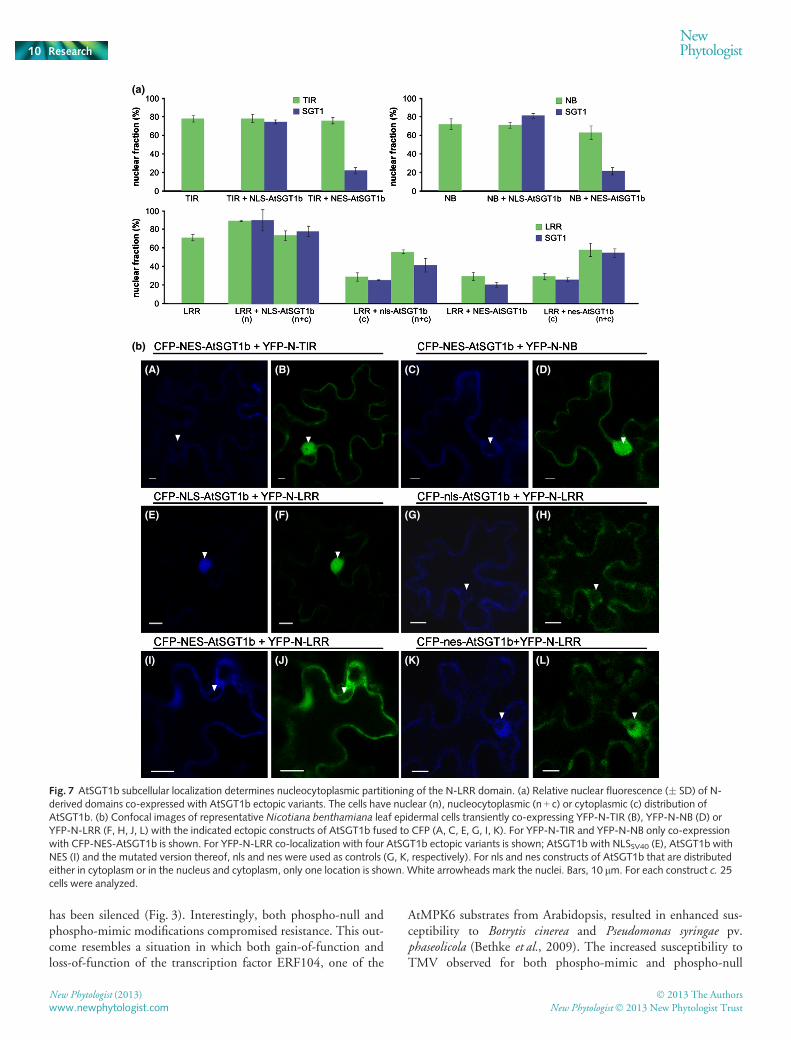
We next compared the distribution patterns of N-deriveddomains and SGT1. Individual domains of N fused to YFP wereco-expressed in N. benthamiana with AtSGT1b localization vari-ants N-terminally fused to CFP. We detected no changes in rela-tive nuclear fluorescence intensities for N-TIR and N-NBdomains when these were co-expressed with NLS-AtSGT1b orNES-AtSGT1b variants (Fig. 7a). By contrast, the localizationparameters were strictly correlated for N-LRR and SGT1 variants(Fig. 7a). Most tellingly, upon co-expression of YFP-N-LRR withCFP-AtSGT1b variants, we found that N-LRR only localized tothe nucleus in those cells where AtSGT1b variants also localizedto the nucleus (Figs 7b, S5). Upon forced nuclear localization orexclusion of AtSGT1b, the co-expressed N-LRR localized againexclusively to the AtSGT1b-containing compartment, and the
(a) (b)
Fig. 5 Relocation of N to nucleus does notabrogate hypersensitive response (HR)triggered by p50 in Nicotiana benthamianaleaves. (a) HR assay to test function ofdifferent N’s pools. (b) Confocal images of N,N-NLS SV40, N-NLSSLCV, N-nlsSV40, N-NES,N-nes fused to Citrine. Arrowheads mark thenuclei. Bars, 10 lm.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8
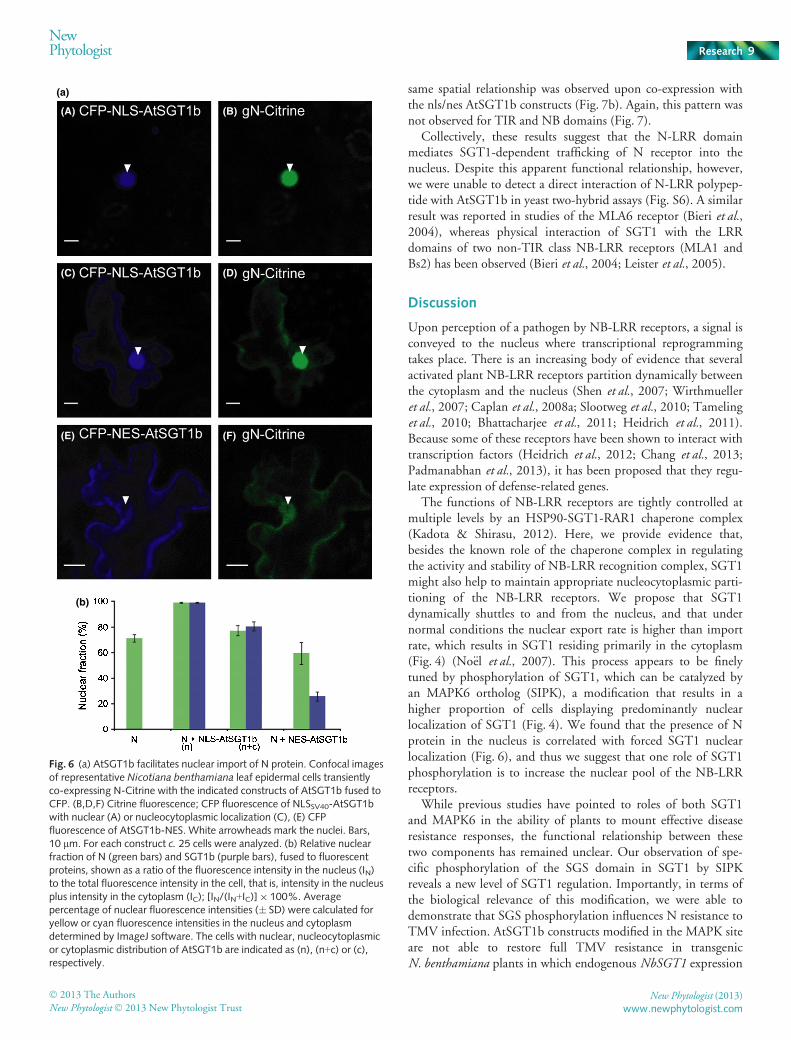
same spatial relationship was observed upon co-expression withthe nls/nes AtSGT1b constructs (Fig. 7b). Again, this pattern wasnot observed for TIR and NB domains (Fig. 7).
Collectively, these results suggest that the N-LRR domainmediates SGT1-dependent trafficking of N receptor into thenucleus. Despite this apparent functional relationship, however,we were unable to detect a direct interaction of N-LRR polypep-tide with AtSGT1b in yeast two-hybrid assays (Fig. S6). A similarresult was reported in studies of the MLA6 receptor (Bieri et al.,2004), whereas physical interaction of SGT1 with the LRRdomains of two non-TIR class NB-LRR receptors (MLA1 andBs2) has been observed (Bieri et al., 2004; Leister et al., 2005).
Discussion
Upon perception of a pathogen by NB-LRR receptors, a signal isconveyed to the nucleus where transcriptional reprogrammingtakes place. There is an increasing body of evidence that severalactivated plant NB-LRR receptors partition dynamically betweenthe cytoplasm and the nucleus (Shen et al., 2007; Wirthmuelleret al., 2007; Caplan et al., 2008a; Slootweg et al., 2010; Tamelinget al., 2010; Bhattacharjee et al., 2011; Heidrich et al., 2011).Because some of these receptors have been shown to interact withtranscription factors (Heidrich et al., 2012; Chang et al., 2013;Padmanabhan et al., 2013), it has been proposed that they regu-late expression of defense-related genes.
The functions of NB-LRR receptors are tightly controlled atmultiple levels by an HSP90-SGT1-RAR1 chaperone complex(Kadota & Shirasu, 2012). Here, we provide evidence that,besides the known role of the chaperone complex in regulatingthe activity and stability of NB-LRR recognition complex, SGT1might also help to maintain appropriate nucleocytoplasmic parti-tioning of the NB-LRR receptors. We propose that SGT1dynamically shuttles to and from the nucleus, and that undernormal conditions the nuclear export rate is higher than importrate, which results in SGT1 residing primarily in the cytoplasm(Fig. 4) (No€el et al., 2007). This process appears to be finelytuned by phosphorylation of SGT1, which can be catalyzed byan MAPK6 ortholog (SIPK), a modification that results in ahigher proportion of cells displaying predominantly nuclearlocalization of SGT1 (Fig. 4). We found that the presence of Nprotein in the nucleus is correlated with forced SGT1 nuclearlocalization (Fig. 6), and thus we suggest that one role of SGT1phosphorylation is to increase the nuclear pool of the NB-LRRreceptors.
While previous studies have pointed to roles of both SGT1and MAPK6 in the ability of plants to mount effective diseaseresistance responses, the functional relationship between thesetwo components has remained unclear. Our observation of spe-cific phosphorylation of the SGS domain in SGT1 by SIPKreveals a new level of SGT1 regulation. Importantly, in terms ofthe biological relevance of this modification, we were able todemonstrate that SGS phosphorylation influences N resistance toTMV infection. AtSGT1b constructs modified in the MAPK siteare not able to restore full TMV resistance in transgenicN. benthamiana plants in which endogenous NbSGT1 expression
(a)
(A) (B)
(C) (D)
(E) (F)
(b)
Fig. 6 (a) AtSGT1b facilitates nuclear import of N protein. Confocal imagesof representative Nicotiana benthamiana leaf epidermal cells transientlyco-expressing N-Citrine with the indicated constructs of AtSGT1b fused toCFP. (B,D,F) Citrine fluorescence; CFP fluorescence of NLSSV40-AtSGT1bwith nuclear (A) or nucleocytoplasmic localization (C), (E) CFPfluorescence of AtSGT1b-NES. White arrowheads mark the nuclei. Bars,10 lm. For each construct c. 25 cells were analyzed. (b) Relative nuclearfraction of N (green bars) and SGT1b (purple bars), fused to fluorescentproteins, shown as a ratio of the fluorescence intensity in the nucleus (IN)to the total fluorescence intensity in the cell, that is, intensity in the nucleusplus intensity in the cytoplasm (IC); [IN/(IN+IC)]9 100%. Averagepercentage of nuclear fluorescence intensities (� SD) were calculated foryellow or cyan fluorescence intensities in the nucleus and cytoplasmdetermined by ImageJ software. The cells with nuclear, nucleocytoplasmicor cytoplasmic distribution of AtSGT1b are indicated as (n), (n+c) or (c),respectively.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 9
has been silenced (Fig. 3). Interestingly, both phospho-null andphospho-mimic modifications compromised resistance. This out-come resembles a situation in which both gain-of-function andloss-of-function of the transcription factor ERF104, one of the
AtMPK6 substrates from Arabidopsis, resulted in enhanced sus-ceptibility to Botrytis cinerea and Pseudomonas syringae pv.phaseolicola (Bethke et al., 2009). The increased susceptibility toTMV observed for both phospho-mimic and phospho-null
(a)
(b)
(A) (B) (C) (D)
(E) (F) (G) (H)
(I) (J) (K) (L)
Fig. 7 AtSGT1b subcellular localization determines nucleocytoplasmic partitioning of the N-LRR domain. (a) Relative nuclear fluorescence (� SD) of N-derived domains co-expressed with AtSGT1b ectopic variants. The cells have nuclear (n), nucleocytoplasmic (n + c) or cytoplasmic (c) distribution ofAtSGT1b. (b) Confocal images of representative Nicotiana benthamiana leaf epidermal cells transiently co-expressing YFP-N-TIR (B), YFP-N-NB (D) orYFP-N-LRR (F, H, J, L) with the indicated ectopic constructs of AtSGT1b fused to CFP (A, C, E, G, I, K). For YFP-N-TIR and YFP-N-NB only co-expressionwith CFP-NES-AtSGT1b is shown. For YFP-N-LRR co-localization with four AtSGT1b ectopic variants is shown; AtSGT1b with NLSSV40 (E), AtSGT1b withNES (I) and the mutated version thereof, nls and nes were used as controls (G, K, respectively). For nls and nes constructs of AtSGT1b that are distributedeither in cytoplasm or in the nucleus and cytoplasm, only one location is shown. White arrowheads mark the nuclei. Bars, 10 lm. For each construct c. 25cells were analyzed.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10

modifications does not necessarily imply that both SGT1phospho-variants play the same role in defense response or thatboth are nonfunctional. On the contrary, the fact that these twovariants differ in their impact on N protein nucleocytoplasmicpartitioning might indicate that two sub-pools of SGT1, phos-phorylated and nonphosphorylated, are required in the cell inorder to establish resistance. Because either of the mutated SGT1proteins is able to restore viability in sgt1 mutant yeast strains(Fig. S3), the overall protein architecture does not appear to beunduly disturbed by the introduced amino acid substitutions. Itwas previously shown that a frame-shift mutation that eliminatedthe target phosphorylation motif (TP) in the distal part ofAtSGT1b had a dominant negative effect on resistance to PVX ina heterologous complementation assay in N. benthamiana (Bot€eret al., 2007). Also, the resistance phenotype of the sgt1beta3 Ara-bidopsis mutant, which produces a truncated protein lacking theC-terminal 36 aa containing the SIPK phosphorylation site, isindistinguishable from that of the sgt1b-3 null mutant (Grayet al., 2003; No€el et al., 2007).
These observations, taken together with our results, stronglysuggest that reversible phosphorylation of SGT1 plays an impor-tant role in plant NB-LRR resistance to pathogen attack. Thefinding that the phospho-mimic substitution of SGT1 results ina marked increase in its partitioning into the nucleus (Fig. 4)suggests that phosphorylation at this residue dynamically regu-lates the cytoplasmic and nuclear SGT1 pools, and that theplant’s ability to mount an effective resistance response is condi-tioned by an appropriate balance between the phosphorylatedand nonphosphorylated forms.
The N protein function during the tobacco TMV defenseresponse can be separated temporally and spatially into two dis-tinct phases: pathogen recognition, which is thought to occur inthe cytoplasm but requires the chloropastic protein NRIP1(N Receptor-Interacting Protein 1); and subsequent defense-ori-ented physiological reprogramming, which is orcheastratedmainly in the nucleus (Burch-Smith et al., 2007; Caplan et al.,2008b). This model is consistent with the observed distributionof the receptor and its ligand, because the N protein is found inthe host cytoplasm and nucleus, whereas the TMV 126-kDareplicase protein (and its p50 fragment that is sensed by N) isassociated with the viral replication complex at the ER(Padmanabhan et al., 2006). Unexpectedly, however, our resultsshow that restriction of N to the nucleus does not compromiseHR development (Fig. 5). Re-direction of p50 into the nucleusalso did not change its elicitation activity, and a typical HR wasconsistently observed when p50-NLSSV40 was co-expressed withN in N. benthamiana (Fig. S4a). Similarly, p50-NLSSV40 infiltra-tion into transgenic N. benthamiana BN3 plants (expressing N)triggered HR (Fig. S4b). These data suggest that the nuclear poolof N is itself competent for association with ligand (Figs 5, S4),and that events occurring subsequent to p50 recognition, such asconformational changes and oligomerization of the receptor(Mestre & Baulcombe, 2006), can also proceed outside the cyto-plasmic milieu. A recent report showed that recognition of theCoat Protein of Potato Virus X (CP-PVX) mediated by thepotato Coiled-Coil (CC)-NB-LRR type Rx protein occurs
mainly in the cytoplasm (Tameling et al., 2010). At this point,we cannot exclude the possibility that establishment of full resis-tance to TMV also requires a small cytoplasmic pool of Nbecause nuclear-targeted proteins have to travel through the cyto-plasm. Indeed, bifurcating cytoplasmic cell death and nuclearpathogen resistance pathways were recently reported for the Ara-bidopsis RPS4 and barley MLA10 NB-LRR receptors (Heidrichet al., 2011; Bai et al., 2012).
The finding that nuclear localization of N is critical for itsfunction in TMV resistance (Burch-Smith et al., 2007) and Fig. 5raises the question of how trafficking of the receptor complexesinto the nucleus is controlled. It was previously hypothesized thattransport of the human NB-LRR type NOD1 receptor into thenucleus could be facilitated by SGT1 (da Silva Correia et al.,2007). Consistent with this idea, and with the evolutionarily con-served nature of SGT1 (Fig. S1), nuclear localization of thepotato Rx receptor protein was dramatically reduced in plantssilenced for SGT1 expression (Slootweg et al., 2010). Nucleo-cytoplasmic partitioning of an Rx variant lacking the LRRdomain was not affected by SGT1 silencing, as predicted by amodel in which SGT1 interacts with the LRR domain of NB-LRR proteins (Bieri et al., 2004; Leister et al., 2005). Reduced
(a) (b)
Fig. 8 A model of maintenance of N’s nucleocytoplasmic partitioning.(a) Pre-recognition complex, made up of N, SGT1, RAR1, HSP90 andpossibly other host factors including p50 docking proteins, assemblies incytoplasm. SGT1 interacts directly or indirectly with Leucine-Rich Repeat(LRR) domains of R receptors (Bieri et al., 2004; Leister et al., 2005). Forsimplicity, only monomeric SGT1 is shown participating in this complex. Inintact cells, the complex shuttles between the cytoplasm and the nucleus.(b) Upon TMV infection, p50 binds first indirectly to N’s TIR domain, thendirectly to the NB and LRR domains, leading to conformational changesand oligomerization of the N protein (Mestre & Baulcombe, 2006; Burch-Smith et al., 2007). Activated SIPK phosphorylates the highly conservedSGS domain located in the C-terminal part of SGT1, thereby shifting thebalance towards the nuclear distribution of SGT1, and consequently, ofthe N receptor complex. Within the nucleus, N interacts withtranscriptional factors (TF), leading to modulation of expression of defenserelated genes.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 11

nuclear localization of N protein lacking the LRR domain hasalso been observed (Burch-Smith et al., 2007).
Our data support the notion that SGT1 plays an importantrole in maintaining an appropriate subcellular distribution of theN receptor. When the N-derived LRR domain was transientlyexpressed either with wild-type AtSGT1b or its ectopic forms,both proteins co-localized (Fig. 7). Interestingly, when AtSGT1bwas co-expressed with the full-length N protein, AtSGT1b wasable to relocate N in only one direction, towards the nucleus.This distinction might indicate that control of the normal direc-tion of signal flow upon TMV recognition involves factors inaddition to SGT1. Alternatively, there might be intramolecularconstraints within the N receptor that normally hinder nuclearimport, but which are relieved upon ligand recognition. Furtherexperiments are necessary to establish which scenario operateswithin the N signaling cascade.
The co-localization pattern of N-LRR and AtSGT1b was simi-lar for all the inspected cells, independently of whether theyexpressed AtSGT1b phospho-variants or AtSGT1b with forcedcytoplasmic or nuclear localization, which suggests that presenceor absence of phosphorylation probably does not change the asso-ciation of AtSGT1b with N (Fig. 7 and data not shown). How-ever, because we were able to show that phospho-mimicsubstitution, in particular, results in a more pronounced redistri-bution of SGT1 between the nucleus and the cytoplasm, we pro-pose a model in which phosphorylation of SGT1 is required forestablishing a pattern of distribution of SGT1, and consequentlyN, between the nucleus and cytoplasm, and that this distributionpattern enables the host cell to mount an effective TMV resis-tance response (Fig. 8). SGT1 seems to contribute to two steps ofN shuttling. First, by maintaining an appropriate level of cyto-plasmic receptors (Holt et al., 2005; Li et al., 2010), SGT1 mightaffect nuclear flow rate of N, as has been shown for other cargoesand their chaperones (Timney et al., 2006). Second, it mightcontrol the translocation process itself (Fig. 6). However, theobservation that the N protein still shows nucleocytoplasmic par-titioning in SGT1-silenced plants (Fig. 6) suggests that, in con-trast to Rx (Slootweg et al., 2010), the critical role for SGT1resides not in control of N-shuttling per se, but rather in its abilityto shift the equilibrium of the receptor distribution towards thenucleus upon effector recognition.
In summary, our results provide new insights into the mecha-nism(s) by which SGT1 modulates plant responses to pathogenattack, and reveal exciting new avenues for exploring the underly-ing regulatory network.
Acknowledgements
We thank: S. Zhang for MEK2 constructs; N-H. Chua for thepTA7002 vector; B. Baker for BN3 seeds; S. P. Dinesh-Kumarfor the TRV VIGS system and gN-Citrine and p50-Ceruleanconstructs; M. Taube for N cDNA subclone; K. Kitagawa foryeast strains; I. Fijałkowska for plasmids used as a positive controlin yeast two-hybrid assays; A. Anielska-Mazur for assistancein confocal analysis; and members of the G. Muszynska andG. Dobrowolska labs for help and stimulating discussions. This
work was supported by grants from Ministry of Science andHigher Education Republic of Poland to M.K. (No N302 01531/1618 and NN301 163235) and J.H. (N N301 318039), andfrom NSERC Canada to B.E. The fellowship for M.Z. wasfunded by the Foundation for Polish Science, project MPD/2009-3/2. J.S. and J.E.P. were supported by a Deutsche Fors-chungsgemeinschaft ‘SFB 635′ grant.
References
Abramoff MD, Magalhaes PJ, Ram SJ. 2004. Image processing with ImageJ.
Biophotonics International 11: 36–41.Andreasson E, Jenkins T, Brodersen P, Thorgrimsen S, Petersen NH, Zhu S,
Qiu JL, Micheelsen P, Rocher A, Petersen M et al. 2005. The MAP kinase
substrate MKS1 is a regulator of plant defense responses. EMBO Journal 24:2579–2589.
Austin MJ, Muskett P, Kahn K, Feys BJ, Jones JD, Parker JE. 2002.
Regulatory role of SGT1 in early R gene-mediated plant defenses. Science295: 2077–2080.
Azevedo C, Betsuyaku S, Peart J, Takahashi A, No€el L, Sadanandom A, Casais
C, Parker J, Shirasu K. 2006. Role of SGT1 in resistance protein accumulation
in plant immunity. EMBO Journal 25: 2007–2016.Azevedo C, Sadanandom A, Kitagawa K, Freialdenhoven A, Shirasu K,
Schulze-Lefert P. 2002. The RAR1 interactor SGT1, an essential component
of R gene-triggered disease resistance. Science 295: 2073–2076.Bai S, Liu J, Chang C, Zhang L, Maekawa T, Wang Q, Xiao W, Liu Y, Chai J,
Takken FL et al. 2012. Structure-function analysis of barley NLR immune
receptor MLA10 reveals its cell compartment specific activity in cell death and
disease resistance. PLoS Pathogens 8: e1002752.Bethke G, Unthan T, Uhrig JF, Poschl Y, Gust AA, Scheel D, Lee J. 2009.
Flg22 regulates the release of an ethylene response factor substrate from MAP
kinase 6 in Arabidopsis thaliana via ethylene signaling. Proceedings of theNational Academy of Sciences, USA 106: 8067–8072.
Bhattacharjee S, Halane MK, Kim SH, Gassmann W. 2011. Pathogen effectors
target Arabidopsis EDS1 and alter its interactions with immune regulators.
Science 334: 1405–1408.Bieri S, Mauch S, Shen QH, Peart J, Devoto A, Casais C, Ceron F, Schulze S,
Steinbiss HH, Shirasu K et al. 2004. RAR1 positively controls steady statelevels of barley MLA resistance proteins and enables sufficient MLA6
accumulation for effective resistance. Plant Cell 16: 3480–3495.Bot€er M, Amigues B, Peart J, Breuer C, Kadota Y, Casais C, Moore G,
Kleanthous C, Ochsenbein F, Shirasu K et al. 2007. Structural and functional
analysis of SGT1 reveals that its interaction with HSP90 is required for the
accumulation of Rx, an R protein involved in plant immunity. Plant Cell 19:3791–3804.
Burch-Smith TM, Schiff M, Caplan JL, Tsao J, Czymmek K, Dinesh-Kumar
SP. 2007. A novel role for the TIR domain in association with
pathogen-derived elicitors. PLoS Biology 5: e68.Caplan J, Padmanabhan M, Dinesh-Kumar SP. 2008a. Plant NB-LRR immune
receptors: from recognition to transcriptional reprogramming. Cell Host &Microbe 3: 126–135.
Caplan JL, Mamillapalli P, Burch-Smith TM, Czymmek K, Dinesh-Kumar SP.
2008b. Chloroplastic protein NRIP1 mediates innate immune receptor
recognition of a viral effector. Cell 132: 449–462.Catlett MG, Kaplan KB. 2006. Sgt1p is a unique co-chaperone that acts as a
client adaptor to link Hsp90 to Skp1p. Journal of Biological Chemistry 281:33 739–33 748.
Chang C, Yu D, Jiao J, Jing S, Schulze-Lefert P, Shen QH. 2013. Barley MLA
immune receptors directly interfere with antagonistically acting transcription
factors to initiate disease resistance signaling. Plant Cell 25: 1158–1173.Cohen P. 2000.The regulation of protein function by multisite phosphorylation –a 25 year update. Trends in Biochemical Sciences 25: 596–601.
Dinesh-Kumar SP, ThamWH, Baker BJ. 2000. Structure-function analysis of
the tobacco mosaic virus resistance gene N. Proceedings of the National Academyof Sciences, USA 97: 14 789–14 794.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12

Feilner T, Hultschig C, Lee J, Meyer S, Immink RG, Koenig A, Possling A,
Seitz H, Beveridge A, Scheel D et al. 2005.High throughput identification of
potential Arabidopsis mitogen-activated protein kinases substrates.Molecular &Cellular Proteomics 4: 1558–1568.
Gray WM, Muskett PR, Chuang H, Parker JE. 2003. Arabidopsis SGT1b is
required for SCFTIR1-mediated auxin response. Plant Cell 15: 1310–1319.Hartl M, Merker H, Schmidt DD, Baldwin IT. 2008.Optimized virus-induced
gene silencing in Solanum nigrum reveals the defensive function of leucine
aminopeptidase against herbivores and the shortcomings of empty vector
controls. New Phytologist 179: 356–365.Heidrich K, Blanvillain-Baufume S, Parker JE. 2012.Molecular and spatial
constraints on NB-LRR receptor signaling. Current Opinion in Plant Biology15: 385–391.
Heidrich K, Wirthmueller L, Tasset C, Pouzet C, Deslandes L, Parker JE. 2011.
Arabidopsis EDS1 connects pathogen effector recognition to cell
compartment–specific immune responses. Science 334: 1401–1404.Holt BF, Belkhadir Y, Dangl JL. 2005. Antagonistic control of disease resistance
protein stability in the plant immune system. Science 309: 929–932.Jin H, Liu Y, Yang K-Y, Kim CY, Baker B, Zhang S. 2003. Function of a
mitogen-activated protein kinase pathway in N gene-mediated resistance in
tobacco. Plant Journal 33: 719–731.Kadota Y, Shirasu K. 2012. The HSP90 complex of plants. Biochimica etBiophysica Acta 1823: 689–697.
Kalderon D, Roberts BL, Richardson WD, Smith AE. 1984. A short amino acid
sequence able to specify nuclear location. Cell 39: 499–509.Kitagawa K, Skowyra D, Elledge SJ, Harper JW, Hieter P. 1999. SGT1 encodes
an essential component of the yeast kinetochore assembly pathway and a novel
subunit of the SCF ubiquitin ligase complex.Molecular Cell 4: 21–33.Lee J, Klessig DF, Nurnberger T. 2001. A harpin binding site in tobacco plasma
membranes mediates activation of the pathogenesis-related gene HIN1
independent of extracellular calcium but dependent on mitogen-activated
protein kinase activity. Plant Cell 13: 1079–1093.Leister RT, Dahlbeck D, Day B, Li Y, Chesnokova O, Staskawicz BJ. 2005.
Molecular genetic evidence for the role of SGT1 in the intramolecular
complementation of Bs2 protein activity in Nicotiana benthamiana. Plant Cell17: 1268–1278.
Li Y, Li S, Bi D, Cheng YT, Li X, Zhang Y. 2010. SRFR1 negatively regulates
plant NB-LRR resistance protein accumulation to prevent autoimmunity. PLoSPathogens 6: e1001111.
Liu Y, Jin H, Yang K-Y, Kim CY, Baker B, Zhang S. 2003. Interaction between
two mitogen-activated protein kinases during tobacco defense signaling. PlantJournal 34: 149–160.
Liu Y, Schiff M, Marathe R, Dinesh-Kumar SP. 2002a. Tobacco Rar1, EDS1and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco
mosaic virus. Plant Journal 30: 415–429.Liu Y, Schiff M, Serino G, Deng XW, Dinesh-Kumar SP. 2002b. Role of SCF
ubiquitin-ligase and the COP9 signalosome in the N gene-mediated resistance
response to Tobacco Mosaic Virus. Plant Cell 14: 1483–1496.van Loon LC. 1983.Mechanism of resistance in virus-infected plants. In: Bailey
J, Deverall B, eds. The dynamics of host defense. Sydney, Australia: Academic
Press, 123–190.Mayor A, Martinon F, De Smedt T, Petrilli V, Tschopp J. 2007. A crucial
function of SGT1 and HSP90 in inflammasome activity links mammalian and
plant innate immune responses. Nature Immunology 8: 497–503.Menke FLH, Kang H-G, Chen Z, Park JM, Kumar D, Klessig DF. 2005.
Tobacco transcription factor WRKY1 is phosphorylated by the MAP kinase
SIPK and mediates HR-like cell death in tobacco.Molecular Plant-MicrobeInteractions 18: 1027–1034.
Mestre P, Baulcombe DC. 2006. Elicitor-mediated oligomerization of the
tobacco N disease resistance protein. Plant Cell 18: 491–501.Nakagawa T, Kurose T, Hino T, Tanaka K, Kawamukai M, Niwa Y, Toyooka
K, Matsuoka K, Jinbo T, Kimura T. 2007a.Development of series of
Gateway binary vectors, pGWBs, for realizing efficient construction of fusion
genes for plant transformation. Journal of Bioscience and Bioengineering 104:34–41.
Nakagawa T, Suzuki T, Murata S, Nakamura S, Hino T, Maeo K, Tabata R,
Kawai T, Tanaka K, Niwa Y et al. 2007b. Improved Gateway binary vectors:
high-performance vectors for creation of fusion constructs in transgenic analysis
of plants. Bioscience, Biotechnology, and Biochemistry 71: 2095–2100.No€el LD, Cagna G, Stuttmann J, Wirthmuller L, Betsuyaku S, Witte CP, Bhat
R, Pochon N, Colby T, Parker JE. 2007. Interaction between SGT1 and
cytosolic/nuclear HSC70 chaperones regulates Arabidopsis immune responses.
Plant Cell 19: 4061–4076.Padmanabhan MS, Ma S, Burch-Smith TM, Czymmek K, Huijser P,
Dinesh-Kumar SP. 2013. Novel positive regulatory role for the SPL6
transcription factor in the N TIR-NB-LRR receptor-mediated plant innate
immunity. PLoS Pathogens 9: e1003235.Padmanabhan MS, Shiferaw H, Culver JN. 2006. The Tobacco Mosaic Virus
replicase protein disrupts the localization and function of interacting Aux/IAA
proteins.Molecular Plant-Microbe Interactions 19: 864–873.Peart JR, Cook G, Feys BJ, Parker JE, Baulcombe DC. 2002. An EDS1orthologue is required for N mediated resistance against tobacco mosaic virus.
Plant Journal 29: 569–579.Pitzschke A, Schikora A, Hirt H. 2009.MAPK cascade signalling networks in
plant defence. Current Opinion in Plant Biology 12: 421–426.Popescu SC, Popescu GV, Bachan S, Zhang Z, Gerstein M, Snyder M,
Dinesh-Kumar SP. 2009.MAPK target networks in Arabidopsis thalianarevealed using functional protein microarrays. Genes & Development 23: 80–92.
Romeis T, Ludwig AA, Martin R, Jones JD. 2001. Calcium-dependent protein
kinases play an essential role in a plant defence response. EMBO Journal 20:5556–5567.
Sanderfoot AA, Ingham DJ, Lazarowitz SG. 1996. A viral movement protein as a
nuclear shuttle. The geminivirus BR1 movement protein contains domains
essential for interaction with BL1 and nuclear localization. Plant Physiology110: 23–33.
Sangster T, Queitsch C. 2005. The HSP90 chaperone complex, an emerging
force in plant development and phenotypic plasticity. Current Opinion in PlantBiology 8: 86–92.
Shen QH, Saijo Y, Mauch S, Biskup C, Bieri S, Keller B, Seki H, Ulker B,
Somssich IE, Schulze-Lefert P. 2007. Nuclear activity of MLA immune
receptors links isolate-specific and basal disease-resistance responses. Science315: 1098–1103.
da Silva Correia J, Miranda Y, Leonard N, Ulevitch R. 2007. SGT1 is essential
for Nod1 activation. Proceedings of the National Academy of Sciences, USA 104:
6764–6769.Slootweg E, Roosien J, Spiridon LN, Petrescu AJ, Tameling W, Joosten M,
Pomp R, van Schaik C, Dees R, Borst JW et al. 2010. Nucleocytoplasmic
distribution is required for activation of resistance by the potato NB-LRR
receptor Rx1 and is balanced by its functional domains. Plant Cell 22: 4195–4215.
Talarczyk A, Krzymowska M, Borucki W, Hennig J. 2002. Effect of yeast CTA1
gene expression on response of tobacco plants to tobacco mosaic virus
infection. Plant Physiology 129: 1032–1044.Tameling WI, Nooijen C, Ludwig N, Bot€er M, Slootweg E, Goverse A, Shirasu
K, Joosten MH. 2010. RanGAP2 mediates nucleocytoplasmic partitioning of
the NB-LRR immune receptor Rx in the Solanaceae, thereby dictating Rxfunction. Plant Cell 22: 4176–4194.
Thao NP, Chen L, Nakashima A, Hara S, Umemura K, Takahashi A, Shirasu K,
Kawasaki T, Shimamoto K. 2007. RAR1 and HSP90 form a complex with
Rac/Rop GTPase and function in innate-immune responses in rice. Plant Cell19: 4035–4045.
Timney BL, Tetenbaum-Novatt J, Agate DS, Williams R, Zhang W, Chait BT,
Rout MP. 2006. Simple kinetic relationships and nonspecific competition
govern nuclear import rates in vivo. Journal of Cell Biology 175: 579–593.Ueda H, Yamaguchi Y, Sano H. 2006. Direct interaction between the tobacco
mosaic virus helicase domain and the ATP-bound resistance protein, N factor
during the hypersensitive response in tobacco plants. Plant Molecular Biology61: 31–45.
Wang K, Uppalapati SR, Zhu X, Dinesh-Kumar SP, Mysore KS. 2010. SGT1
positively regulates the process of plant cell death during both compatible and
incompatible plant–pathogen interactions.Molecular Plant Pathology 11: 597–611.
WenW, Meinkoth JL, Tsien RY, Taylor SS. 1995. Identification of a signal for
rapid export of proteins from the nucleus. Cell 82: 463–473.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 13

Whitham S, Dinesh-Kumar SP, Choi D, Hehl R, Corr C, Baker B. 1994. The
product of the tobacco mosaic virus resistance gene N: similarity to toll and the
interleukin-1 receptor. Cell 78: 1101–1115.Wirthmueller L, Zhang Y, Jones JD, Parker JE. 2007. Nuclear accumulation of
the Arabidopsis immune receptor RPS4 is necessary for triggering
EDS1-dependent defense. Current Biology 17: 2023–2029.Witte CP, No€el LD, Gielbert J, Parker JE, Romeis T. 2004. Rapid one-step
protein purification from plant material using the eight-amino acid StrepII
epitope. Plant Molecular Biology 55: 135–147.Yang KY, Liu Y, Zhang S. 2001. Activation of a mitogen-activated protein kinase
pathway is involved in disease resistance in tobacco. Proceedings of the NationalAcademy of Sciences, USA 98: 741–746.
Zhang M, Kadota Y, Prodromou C, Shirasu K, Pearl LH. 2010. Structural basis
for assembly of Hsp90-Sgt1-CHORD protein complexes: implications for
chaperoning of NLR innate immunity receptors.Molecular Cell 39: 269–281.Zhang S, Du H, Klessig DF. 1998. Activation of the tobacco SIP kinase by both
a cell wall-derived carbohydrate elicitor and purified proteinaceous elicitins
from Phytophthora spp. Plant Cell 10: 435–450.Zhang S, Liu Y. 2001. Activation of salicylic acid-induced protein kinase, a
mitogen-activated protein kinase, induces multiple defense responses in
tobacco. Plant Cell 13: 1877–1889.Zhang S, Liu Y, Klessig DF. 2000.Multiple levels of tobacco WIPK activation
during the induction of cell death by fungal elicitins. Plant Journal 23: 339–347.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Sequence comparison of the SGS domains of variousSGT1 proteins.
Fig. S2Mascot analysis of the in vitro phosphorylated NbSGT1.
Fig. S3Complementation of yeast sgt1 mutants with phospho-variants of SGT1.
Fig. S4 Ability of ectopically expressed p50 to trigger HRresponse.
Fig. S5 Subcellular localization of AtSGT1b phospho-variantsdetermines nucleocytoplasmic partitioning of the N-LRRdomain.
Fig. S6 Yeast two-hybrid analysis of the interaction between N-LRR and AtSGT1b.
Methods S1Mass spectrometry details, yeast complementationand yeast two-hybrid assay.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist (2013) � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist14


















