
Nrf2-activators Attenuate the Progression of Nonalcoholic
47
MOL #84269 Title page Nrf2-activators Attenuate the Progression of Nonalcoholic Steatohepatitis–Related Fibrosis in a Dietary Rat Model Rieko Shimozono, Yoshiji Asaoka, Yoshitaka Yoshizawa, Takumi Aoki, Hidetoshi Noda, Masateru Yamada, Mie Kaino, and Hidenori Mochizuki Toray Industries, Inc., Pharmaceutical Research Laboratories, Kanagawa, Japan. Molecular Pharmacology Fast Forward. Published on April 16, 2013 as doi:10.1124/mol.112.084269 Copyright 2013 by the American Society for Pharmacology and Experimental Therapeutics. This article has not been copyedited and formatted. The final version may differ from this version. Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269 at ASPET Journals on April 4, 2019 molpharm.aspetjournals.org Downloaded from
Transcript of Nrf2-activators Attenuate the Progression of Nonalcoholic

MOL #84269 1
Title page
Nrf2-activators Attenuate the Progression of Nonalcoholic Steatohepatitis–Related Fibrosis
in a Dietary Rat Model
Rieko Shimozono, Yoshiji Asaoka, Yoshitaka Yoshizawa, Takumi Aoki, Hidetoshi Noda,
Masateru Yamada, Mie Kaino, and Hidenori Mochizuki
Toray Industries, Inc., Pharmaceutical Research Laboratories, Kanagawa, Japan.
Molecular Pharmacology Fast Forward. Published on April 16, 2013 as doi:10.1124/mol.112.084269
Copyright 2013 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 2
Running Title Page
Running title
Nrf2-activators attenuate fibrosis progression in NASH model
Contact information
Rieko Shimozono. Pharmaceutical Research Laboratories, Toray Industries, Inc., 10-1,
Tebiro 6-chome, Kamakura, Kanagawa 248-8555, Japan. Telephone number: +81 467 32
9805; fax number: +81 467 32 9862; e-mail address: [email protected]
Number of items
number of text pages: 40
number of tables: 1
number of figures: 7
number of references: 40
number of words in Abstract: 246
number of words in Introduction: 449
number of words in Discussion: 1447
List of Abbreviations
ALT, alanine aminotransferase; ARE, antioxidant response element; AST, aspartate
aminotransferase; α-SMA, α smooth muscle actin; CDAA, choline-deficient L-amino
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 3
acid-defined; COL1A1, collagen α1(I); ECM, extracellular matrix synthesis; GST, glutathione
S-transferase; H&E, hematoxylin and eosin; HSC, hepatic stellate cell; Keap1, kelch-like
ECH-associated protein 1; NAFLD, nonalcoholic fatty liver disease; NASH, nonalcoholic
steatohepatitis; NQO1, NAD(P)H quinone oxidoreductase 1; Nrf2, nuclear
factor-erythroid-2-related factor 2; OPZ, oltipraz; qPCR, quantitative polymerase chain
reaction; ROS, reactive oxygen species; TGF-β1, transforming growth factor-β1; TIMP-1,
tissue inhibitor of metalloproteinase-1.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 4
Abstract
Oxidative stress is considered to be a key mechanism of hepatocellular injury and disease
progression in patients with nonalcoholic steatohepatitis (NASH). The transcription factor
Nrf2 plays a central role in stimulating expression of various antioxidant-associated genes in
the cellular defense against oxidative stress. As the cytosolic repressor Keap1 negatively
regulates Nrf2, activation of Nrf2 facilitated by its release from Keap1 may represent a
promising strategy in the treatment of NASH. To test this hypothesis, we utilized two
chemically distinct types of Nrf2-activator. One is the thiol-reactive agent oltipraz (OPZ), a
typical Nrf2-activator, and the other is a novel biaryl urea compound, termed NK-252.
NK-252 exhibits a greater Nrf2-activating potential than OPZ. Furthermore, in vitro binding
studies revealed that NK-252 interacts with the domain containing the Nrf2-binding site of
Keap1, whereas OPZ does not. This finding indicates that NK-252 is more potent than OPZ
due to its unique mechanism of action. For in vivo animal model studies, we employed rats
on a choline-deficient L-amino acid-defined (CDAA)-diet, which demonstrate pathological
findings similar to those seen in human NASH. The administration of OPZ or NK-252
significantly attenuated the progression of histological abnormalities in rats on a CDAA diet,
especially hepatic fibrosis. In conclusion, by using Nrf2-activators with independent
mechanisms of action, we show in a rat model of NASH that the activation of Nrf2 is
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 5
responsible for the anti-fibrotic effects of these drugs. This strategy of Nrf2 activation
presents new opportunities for treatment of NASH patients with hepatic fibrosis.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 6
Introduction
Nonalcoholic fatty liver disease (NAFLD) is a common, chronic disease of the liver. NAFLD
is mainly associated with the presence of obesity and multiple metabolic disorders.
Nonalcoholic steatohepatitis (NASH) is a progressive form of NAFLD and defined as the
presence of hepatic steatosis and inflammation with hepatocyte injury (ballooning) with or
without fibrosis (Chalasani et al., 2012). NASH can develop into cirrhosis, hepatic failure,
and hepatocellular carcinoma. With the growing epidemic of obesity, the prevalence and
impact of NAFLD continues to increase, making NASH potentially the most common cause
of advanced liver disease in coming decades (Vernon et al., 2011).
There is currently no broadly approved pharmacological therapy for NAFLD and the
management of patients with NAFLD consists of treating the associated metabolic disorders
(Chalasani et al., 2012). Several studies have investigated the effect of insulin sensitizing
agents in patients with NASH. For example, pioglitazone, one of the most well-established
thiazolidinediones, was demonstrated to improve liver histopathology in NASH. Furthermore,
antioxidant therapy with vitamin E has also been shown to have a beneficial effect on liver
histopathology. However, their efficacies are likely to be limited, and have not yet been
confirmed especially regarding fibrosis due to mixed results (Chalasani et al., 2012, Sanyal
et al., 2010, Comar and Sterling, 2006).
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 7
Oxidative stress is thought to be a major contributor to the pathogenesis and progression of
NASH (Koek et al., 2011). Oxidative stress has been defined as an imbalance between
oxidants and antioxidants in favor of the former, resulting in an overall increase in cellular
levels of reactive oxygen species (ROS) (Sies, 1997). In patients with histopathologically
progressive NASH, production of antioxidants is reduced and the total antioxidant capacity
is apparently insufficient to compensate for oxidative stress (Sreekumar et al., 2003).
Therefore, agents that promote cellular antioxidant defense mechanisms are likely to
improve NASH as well as or better than direct scavengers of ROS like vitamin E.
Nuclear factor-erythroid-2-related factor 2 (Nrf2), a transcription factor that activates
antioxidant response elements (AREs), plays a central role in stimulating expression of
various antioxidant-associated genes in the cellular defense against oxidative stress (Itoh et
al., 1997). Under normal conditions, Kelch-like ECH-associated protein 1 (Keap1), a
cytosolic repressor of Nrf2, retains Nrf2 in the cytoplasm (Itoh et al., 1999, Itoh et al., 2004).
Thus, activation of Nrf2, facilitated by its release from Keap1, may represent a promising
strategy in the treatment of NASH.
To test this hypothesis, we utilized two chemically distinct types of Nrf2-activator. This study
characterized the respective mechanisms by which they activate Nrf2, and furthermore,
investigated their biochemical and histopathological effects on NASH using an established
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 8
model of diet-induced NASH: rats on a choline-deficient L-amino acid-defined (CDAA)-diet
(Nakae et al., 1992).
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 9
Materials and Methods
Chemicals. Oltipraz (OPZ, Fig. 1, left) was obtained from LKT Laboratories, Inc. (St. Paul,
MN). NK-252 (Fig. 1, right) was obtained from in-house synthesis. All compounds were
diluted in dimethyl sulfoxide (Sigma-Aldrich, St. Louis, MO) for in vitro assays, or suspended
in 0.5% (w/v) methyl cellulose (Nakarai tesque, Kyoto, Japan) for in vivo assays.
Plasmid Construntion. The 3-tandem repeat of ARE in the 5’-upstream region of NAD(P)H
quinone oxidoreductase 1 (NQO1) gene (Nioi et al., 2003) was inserted into pGL4.32
luciferase reporter vector (Promega, Madison, WI) via KpnI and HindIII sites to create ARE /
pGL4.32. A gene fragment encoding the partial length (residues 321-609) of human Keap1
(named Keap1-DC) was inserted into pGEX-6p-3 glutathione S-transferase (GST)-fusion
protein expression vector (GE Healthcare, Bucks, UK) via EcoRI and XhoI sites to create
Keap1-DC / pGEX-6p-3. All resultant plasmids were amplified in E. coli DH5α.
Stable Transformation, siRNA Transfection of Cells and Luciferase Reporter Gene
Assay. The Huh-7.5 cells, a subline derived from Huh-7 cells (Blight et al., 2002), were
obtained under license from Apath, LLC (St Louis, MO). Cells were transfected with ARE /
pGL4.32 by Lipofectamine™ LTX (Invitrogen, Carlsbad, CA). The stable clonal transfectant
was isolated by selection in hygromycin B (0.1 mg/mL). Cells derived from stable clones
were transfected with control or Nrf2 small interfering RNA (siRNA) (Stealth™ Select RNAi
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 10
for Nrf2, HSS107130, Invitrogen) by lipofectamine RNAiMAX (Invitrogen) (30 h), then
treated with OPZ, NK-252 (0.1−30 μmol/L, 16 h) or dimethyl sulfoxide alone (control). The
luciferase activity values were measured using the Steady-Glo® Luciferase Assay System
(Promega).
Hydrogen Peroxide (H2O2)-Induced Cytotoxicity Assays. The Huh-7 cells were treated
with OPZ, NK-252 (0.1−30 μmol/L, 24 h) or dimethyl sulfoxide alone (control). Cells were
then washed and treated with H2O2 (1 mmol/L, 24 h). Cell viability values were determined
using the Cell Titer 96® AQueous One Solution Reagent (Promega).
Protein Expression and Purification. pGEX-6p-3 or Keap1-DC / pGEX-6p-3 were
transformed into E. coli BL21 (DE3) cells. The induction of GST or GST-fused Keap1-DC
gene expression was done with isopropyl-1-thio-β-D-galactoside (0.2 mmol/L). Soluble
proteins were then extracted using the BugBuster® HT Protein Extraction Reagent
(Novagen, Madison, WI) and purified on GSTrap FF columns (1 mL, GE Healthcare).
Surface Plasmon Resonance (SPR) Interaction Analysis (BIAcore™). The SPR
measurements were performed on a BIAcore™ S51 instrument (Biacore AB, Uppsala,
Sweden). GST-fusioned Keap1-DC was bound onto the surface of flow cell 1 of a Series S
Sensor Chip CM5 (GE Healthcare) using the amine coupling kit (GE Healthcare). Flow cell 2
with GST was used as the reference flow cell. OPZ or NK-252 solutions (12.5−100 μmol/L)
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 11
were injected for 60 s at a flow rate of 10 μL/min. The resonance unit curves were
processed by subtracting the response in flow cell 1 from that in flow cell 2.
Animals and in vivo Experimental Design. The animal experiments were conducted
according to Guidelines for Animal Experiments, Research & Development Division, Toray
Industries, Inc. Six-week-old male Fischer 344 rats (Charles River Laboratories Japan Inc.,
Kanagawa, Japan) were randomly divided into four compound administration groups and
four control groups. Compound administration groups of rats fed with choline-deficient
L-amino acid-defined CDAA diet (Research Diets, Inc., New Brunswick, NJ) were orally
administered as follows: 1) with OPZ from one week after feeding at a dose of 60 mg/kg
once daily for nine weeks (CDAA+OPZ group; N=8), 2) with NK-252 from one week after
feeding at a dose of 20 mg/kg once daily for nine weeks (CDAA+NK-252_low group; N=8),
3) 2) at a dose of 60 mg/kg (CDAA+NK-252_high group; N=8), or 4) with NK-252 from six
weeks after feeding at a dose of 60 mg/kg once daily for four weeks (CDAA+NK-252_DA
[Delayed Administration] group; N=7). Two Control groups of rats fed with a CDAA diet for
six or ten weeks (pre-CDAA control or CDAA control group; N=9 each), and the other two
control groups of rats fed with CRF-1 diet (Charles River Laboratories Japan Inc.) for six or
ten weeks (pre-naive or naive; N=3 each). Laparotomy and blood sampling were performed
under isoflurane anesthesia. After blood sampling, rats were euthanized by exsanguination
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 12
under isoflurane anesthesia and the livers were immediately extirpated.
Histopathological Examination. In all experiments, 2 μm-thick slices of formalin-fixed and
paraffin-embedded liver were processed with hematoxylin and eosin (H&E) stain and Sirius
red stain using established methods. In addition, formalin-fixed paraffin sections were
applied to immunostaining for α smooth muscle actin (α-SMA). Sections were boiled for 20
min in Immunosaver (Wako, Osaka, Japan) before the overnight incubation of the primary
antibody (M0851, DAKO, Glostrup, Denmark). They were then exposed to Histofine®
Simple Stain RAT MAX PO (Nichirei Biosciences, Tokyo, Japan) for 30 min, visualized by
the peroxidase-diaminobenzidine (DAB) reaction.
Histopathological evaluation was performed using H&E and Sirius red staining. The fibrosis
score was defined with a modified Brunt’s method (Brunt, 2001) as follows: score-1) slight
fibrosis is observed sporadically in the centrilobular or periportal area, score-2) slight fibrosis
is observed frequently in the centrilobular and periportal area, score-3) bridging fibrosis is
formed between the centrilobular and periportal area, score-4) pseudolobule is formed. The
Nano-Zoomer Digital Pathology (Hamamatsu Photonics KK, Shizuoka, Japan) was used to
acquire digital high-resolution images through the 50× objectives. Image analysis was
performed using Definiens XD software (Definiens, Munich, Germany). Three images per
specimen were examined blindly and randomly and Sirius red-positive areas were quantified
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 13
as rate of fibrosis area.
Measurement of Plasma Transaminases. Blood samples from the abdominal aorta were
collected in tubes containing lithium heparin and plasma separator (Becton Dickinson and
Co., Franklin, NJ), centrifuged at 1500 x g, for 15 min to obtain the supernatant plasma.
Plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were
measured using a DRI-CHEM system (Fujifilm Corp., Tokyo, Japan).
Real-Time qPCR. Total RNA was extracted from each liver using the RNeasy® Mini Kit
(Qiagen, Valencia, CA) and translated into complementary DNA (cDNA) with High-Capacity
cDNA Reverse Transcription Kits (Applied Biosystems, Foster City, CA). Each cDNA was
subjected to real-time quantitative polymerase chain reaction (qPCR) using the ABI-Prism
7500 Sequence Detection System (Applied Biosystems) and amplified by SYBR® Premix
Ex Taq™ (TaKaRa Bio, Tokyo, Japan) reaction mixture utilizing gene specific primers
(Supplemental Table 1).The relative amount of each mRNA was determined using the
standard curve method and normalizing with GAPDH mRNA expression levels.
Measurement of Carbonyl Content of Proteins. Liver samples were rinsed with
phosphate-buffered saline to remove blood and homogenized. Following the measurement
of protein concentration by Bradford assay using the protein dye reagent concentrate
(Bio-Rad Laboratories, Richmond, CA), protein carbonyl derivatives in each homogenate
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 14
were analyzed using the OxiSelect™ Protein Carbonylation ELISA kit (Cell Biolabs, Inc.,
San Diego, CA) based on equal protein loading (10 μg/mL).
Statistical Analysis. In vivo quantitative data were analyzed by either Dunnett’s test or
Steel’s test with unequal variance, histological scores were analyzed by Steel’s test, and P <
0.05 was considered statistically significant.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 15
Results
NK-252 and OPZ Activate Nrf2 and Protect against H2O2-induced Cytotoxicity. As an
indicator of Nrf2 activation, we examined activation of the NQO1-ARE, which is
predominantly regulated by Nrf2 (Kwak et al., 2001, Reisman et al., 2009a, Reisman et al.,
2009b), using a luciferase reporter gene assay. The luciferase activity in Huh-7.5 cells
treated with OPZ or NK-252 showed activation of the NQO1-ARE in a dose-dependent
manner (Fig. 2, solid line). NK-252 displayed this effect with higher potency than OPZ based
on the fact that the EC2 value (concentration for a two-fold induction above background),
calculated with linear extrapolation from the values above and below the induction threshold,
was 20.8 μmol/L for OPZ and 1.36 μmol/L for NK-252. We also confirmed that the luciferase
activities induced by OPZ or NK-252 were almost completely inhibited by Nrf2 siRNA (Fig. 2,
dotted line). These results indicate that OPZ and NK-252 have potential as Nrf2-activators in
hepatic cells. Prototypical Nrf2-activators that include OPZ have been reported to protect
microglial cells from H2O2-induced cytotoxicity (Konwinski et al., 2004). In this study, we
examined and compared the protective effects of OPZ and NK-252 against H2O2-induced
cytotoxicity using Huh-7 cells to evaluate their antioxidant properties. The cells treated with
OPZ or NK-252 showed increased resistance to H2O2-induced cytotoxicity compared to
control cells (Fig. 3). Furthermore, the protective effect of NK-252 was also considerably
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 16
stronger than that of OPZ.
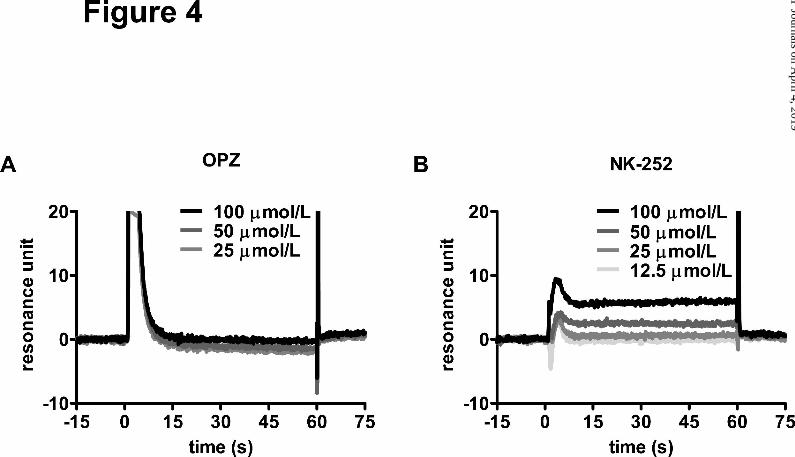
NK-252 but Not OPZ Interacts Directly with the Keap1-DC Domain that Contains the
Nrf2-binding Site. Sulfhydryl reactive compounds, such as dithiolethiones (including OPZ)
have been assumed to modify the sulfhydryl groups of Keap1 cysteines directly, and alter
the conformation of Keap1 (Kwak et al., 2003, Kobayashi et al., 2009). On the other hand,
NK-252, with no thiol reactive group, was not expected to interact with cysteine residues of
Keap1. Thus, we examined the binding affinity of NK-252 for the Keap1-DC domain
harboring the Kelch/double-glycine repeat motif, which has been reported to associate with
Nrf2 (Padmanabhan et al., 2006, Tong et al., 2006), and the C-terminal region. Using
BIAcore™ technology, the binding of NK-252 to the recombinant protein, consisting of the
Kelch domain of Keap1, was detected as an increase in resonance units in a
dose-dependent manner (Fig. 4B). In contrast, binding of OPZ to Keap1-DC was not
detected (Fig. 4A).
OPZ and NK-252 Exert Histopathological Anti-fibrotic and Anti-inflammatory Property
in the Liver of NASH Model Rats. On histopathological examination (Fig. 5A, B), the
median score of fibrosis in rats fed with a CDAA diet for 10 weeks (CDAA control) was 4,
indicating that most rats exhibited pseudolobule formation. However, rats on a CDAA diet
given OPZ or NK-252 displayed decreased fibrosis scores compared with CDAA control rats,
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 17
with median scores of 3, corresponding to bridging fibrosis (Fig. 5D). In addition, CDAA
control rats displayed approximately 20-fold augmentation of the liver fibrosis area
compared with rats fed with a normal control diet (naive) (14.7%, 0.72%, respectively). This
augmentation was also drastically reduced by administration of OPZ or NK-252 (Fig. 5E,
5.80% for OPZ, 6.20% [low] and 4.97% [high] for NK-252). The effects of NK-252 on both
fibrosis score and fibrosis area were dose-dependent. Furthermore, immunohistochemistry
for α-SMA (Fig. 5C), an indicator of activation of hepatic stellate cells (HSCs), demonstrated
that there was a marked increase in HSCs activation in livers of CDAA control rats,
especially at the advancing edge of the fibrosis. In contrast, livers of rats on a CDAA diet
given OPZ or NK-252 showed markedly reduced numbers of α-SMA positive cells.
Regarding inflammation, cellular infiltration of liver tissue was also assessed on H&E
stained liver sections. Infiltration with inflammatory cells, which was rarely observed in the
liver of naive rats, appeared occasionally (slight, +) or frequently (moderate, ++) in the
peribiliary region of CDAA control rats. On the other hand, the incidence and severity of
inflammatory lesions were decreased in rats given OPZ or NK-252, demonstrating the
anti-inflammatory properties of OPZ and NK-252 (Supplemental Table 2).
OPZ and NK-252 Have a Hepatoprotective Effect as Manifested by Levels of Plasma
Aminotransferases in NASH Model Rats. The biochemical liver function tests showed
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 18
mild elevation of plasma aminotransferase (ALT and AST) levels, indicators of
hepatocellular damage, in CDAA control rats (Table 1). In contrast, a significant reduction in
plasma ALT levels were observed in rats on a CDAA diet with high-dose administration of
NK-252, and plasma AST levels were decreased significantly in each of the compound
administration groups when compared with CDAA control rats (Table 1), which indicates the
hepatoprotective effects of OPZ and NK-252.
OPZ and NK-252 Upregulate NQO1 Gene Expression and Exert Antioxidant Property
in the Liver of NASH Model Rats. To demonstrate Nrf2 activation by OPZ and NK-252 in
vivo, using real-time qPCR we examined the level of gene expression of NQO1, one of the
prototypic Nrf2-regulated antioxidant enzymes, in the liver of rats. There was no significant
difference between CDAA control rats and naive rats. However, administration of OPZ or
NK-252 significantly up-regulated NQO1 gene expression in the liver of rats on a CDAA diet
(Fig. 6A). Carbonyl content of proteins as a marker of protein oxidation was also measured
to validate oxidative stress in the liver of rats on a CDAA diet and antioxidant properties of
OPZ and NK-252 in vivo. CDAA control rats showed markedly higher carbonyl content in
liver than Naive rats, which indicates elevation of oxidative damage. In contrast, the
carbonyl content in liver of rats on a CDAA diet given OPZ or NK-252 were lower compared
to these of CDAA control rats (Fig. 6B). These results suggest that OPZ and NK-252 exerted
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 19
both Nrf2-activating and antioxidant properties in those livers affected by NASH.
OPZ and NK-252 Downregulate Fibrogenic Gene Expression in the Liver of NASH
Model Rats. To investigate the molecular mechanism of Nrf2-activator’s anti-fibrotic
properties, we examined levels of gene expression of fibrogenic mediators including
transforming growth factor-β1 (TGF-β1), collagen α1(I) (COL1A1) and tissue inhibitor of
metalloproteinase-1 (TIMP-1). The expression of all of these genes was elevated in the liver
of rats on a CDAA diet compared to naive rats (4.73, 52.5, 13.4-fold, respectively). On the
other hand, rats on a CDAA diet given OPZ or NK-252 showed substantially lower
expression of all three genes, although not all changes were statistically significant (Fig.
6C-E). The effect of NK-252 on the expression of these genes and the associated
histopathological properties (Fig. 5D, E) were both dose-dependent, suggesting that these
genes are involved in the anti-fibrotic mechanism of NK-252.
NK-252 Prevents Further Progression of Established Fibrosis in the Liver of NASH
Model Rats. Thus far, we used a dosing regimen in which administration of the test
compounds was started after one week of pre-feeding with a CDAA diet. In a preliminary
examination, we had confirmed that feeding of a CDAA diet for one week induced nothing
other than histological steatosis and increases in plasma aminotransferases (unpublished
observation). To examine the therapeutic, rather than preventive, anti-fibrotic effect of an
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 20
Nrf2-activator, we used the delayed-administration regimen with NK-252 as a representative
agent, a regimen that consisted of four-week administration of the drug after six-weeks of
pre-feeding with a rats on a six-week CDAA diet before administration (pre-CDAA control)
already showed a fibrosis score of at least 2, with slight fibrosis frequently present in the
centrilobular and periportal area (Fig. 7A). They also displayed approximately four-fold
augmentation of the liver fibrosis area compared with pre-naive rats (3.39%, 0.86%,
respectively, Fig. 7B). Four-week administration of NK-252 resulted in substantial
retardation of the progression of fibrosis compared with CDAA control rats, as reflected in
both the fibrosis score (median score-3) and fibrosis area (5.20%) (Fig. 7A, B). In addition,
infiltration of inflammatory cells in the peribiliary region was almost completely inhibited by
the administration of NK-252 (Supplemental Table 2).
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 21
Discussion
Published studies using knockout or knockdown animals have demonstrated a relationship
between NASH and the Keap1-Nrf2 system. That is, loss of Nrf2 leads to rapid onset and
exacerbation of steatohepatitis in animal models of NASH (Chowdhry et al., 2010, Sugimoto
et al., 2010). In contrast, loss of Keap1, with enhanced expression of Nrf2, attenuates
hepatic steatosis (Zhang et al., 2010). However, potential contribution of Nrf2 to progression
of fibrosis remains unknown. In addition, some chemical Nrf2-activators have been reported
to prevent obesity and insulin resistance (Shin et al., 2009, Yu et al., 2011) —the disorders
that underlie NASH—while there is still no clear-cut evidence whether Nrf2-activators are
clinically effective in NASH.
Among prototypical Nrf2-activators, OPZ, previously studied in a phase II trial of patients
with liver fibrosis or cirrhosis (Kim et al., 2011), was assumed to be a representative tool to
investigate the efficacy of chemical Nrf2-activators on NASH. However, OPZ may have
various target molecules other than Keap1 (Kang et al., 2002, Brooks et al., 2009)
presumably due to its mechanism of action, i.e., reacting with the sulfhydryl groups of
cysteines. Thus, OPZ probably has various effects independent of Nrf2 activation and OPZ
alone appears to be insufficient to confirm our hypothesis. To create a more specific
Nrf2-activator, we designed and synthesized a novel biaryl urea compound NK-252, with no
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 22
sulfhydryl reactive group. In cell-based assays, NK-252 was found to have both Nrf2
activation effects and an antioxidant capability greater than those of OPZ. More surprising is
that an in vitro BIAcore™ binding study revealed that NK-252 interacts directly with the
Keap1-DC domain containing the Nrf2-binding site, whereas OPZ does not. This result
indicates that NK-252 activates Nrf2 through a unique mechanism of action, probably in a
more specific manner than prototypical Nrf2-activators like OPZ. Although further verification
is needed, including structural analysis of binding of NK-252 to Keap1 (e.g. X-ray
crystallography), it is possible that NK-252 represents the first chemical compound that
competitively inhibits the binding of Nrf2 to Keap1. Regarding drug discovery, it appears that
the OPZ-based design of Nrf2-activators may not be achievable because some
modifications of its chemical structure are thought to result in interaction with cysteine
residues in proteins other than Keap1. However, the NK-252-based design seems to
represent a more feasible approach to the production of specific Nrf2-activators.
The CDAA-diet model has been developed as a nutritional animal model of NASH
representing the progression of NASH that occurs when associated with worsening
oxidative damage (Nakae et al., 1992). In this study, rats on a CDAA diet exhibited an
increased level of protein oxidation in the liver, as occurs in patients with NASH (Valenti et
al., 2008); this model was therefore assumed to be the most appropriate for evaluating the
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 23
efficiency of Nrf2-activators. OPZ and NK-252 commonly attenuated development of fibrosis
in rats on a CDAA diet in both qualitative (histopathological scoring) and quantitative
(assessment of fibrosis area) terms, likely due to suppression of HSCs activation, which play
a pivotal role in fiber synthesis and degradation. Dose-dependency of NK-252 was observed
for Nrf2-activating, antioxidant and anti-fibrotic properties, which suggests some correlation
between Nrf2 activation, suppression of oxidative stress and attenuation of liver fibrosis.
Protein carbonyl measurement did not detect sufficient antioxidant property of OPZ to
explain its anti-fibrotic property, which indicates at least a partial contribution by the
nonspecific effect of OPZ discussed above. However, the possibility remains that other
markers of oxidative stress might reveal its antioxidant property more precisely.
Moreover, the delayed-administration NK-252 regimen demonstrated that it can prevent
further progression of established fibrosis. This finding supports the possibility that
Nrf2-activators delay the development of liver fibrosis in patients with NASH. In this study,
we did not demonstrate recovery from fibrosis with NK-252 administration, compared with
baseline. However, further investigations using a different regimen or other animal models
are necessary to confirm whether or not Nrf2-activators can reverse liver fibrosis
The cell-based assay revealed the preventive effects of OPZ and NK-252 against
H2O2-induced cytotoxicity in the Huh-7 cells, which originate from hepatocytes. Therefore,
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 24
the in vivo hepatoprotective effects of these compounds in the liver of rats on a CDAA diet
were likely to be due to their direct actions on hepatocytes. It is generally accepted that
there is sequential crosstalk between hepatocytes, inflammatory cells and HSCs. Hepatic
injury causes accelerated recruitment of inflammatory cells (e.g., Kupffer cells) and their
pro-fibrogenic cytokine secretion stimulates extracellular matrix synthesis (ECM) synthesis
by activating HSCs. In this study, OPZ and NK-252 were found to block the infiltration of
inflammatory cells, probably due to attenuation of hepatic injury. Furthermore, there is a
possibility that the attenuation of fibrosis seen with these compounds was also attributable
to their hepatoprotective properties.
HSCs may also be a direct target of Nrf2-activators. It is widely assumed that persistent
auto-/paracrine stimulation of activated HSCs by TGF-β1, a potent pro-fibrogenic cytokine,
is the key mechanism in liver fibrogenesis. TGF-β1 has been reported to activate SMAD
family of transcription factors, which lead to induction of fibrogenic genes expression
including COL1A1 and TIMP-1 (Verrecchia et al., 2001) and autoinduction of TGF-β1
(Roberts et al., 2006). TIMP-1, an inhibitor of ECM-degrading proteases, not only prevents
the degradation of ECM, but also inhibits the apoptosis of activated HSCs (Murphy et al.,
2002), resulting in the synergistic inductuion of ECM synthesis. These previous findings,
taken together with our analysis of gene expression, suggest that Nrf2-activators modulate
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 25
TGF-β1-mediated fiber synthesis and degradation in the HSCs of rats on a CDAA-diet.
Moreover, published studies have reported cross-talk between TGF-β1 and ROS signaling
in HSCs. TGF-β1 induces the accumulation of H2O2, and this ROS act as intracellular signal
mediator of the pro-fbrogenic action of TGF-β1 (De et al., 1999, García-Trevijano et al.,
1999), which suggests that ROS stimulates induction of fibrogenic genes expression
including TGF-β1 itself. Therefore, once ROS has stimulated TGF-β1 signal transduction, a
vicious circle between ROS and TGF-β1 presumably promotes HSCs activation and ECM
synthesis. Nrf2-activators with antioxidant properties may prevent progression of liver
fibrosis through the interruption of this vicious circle. On examination of the molecular
mechanism underlying anti-fibrotic properties of Nrf2-activators on HSCs activation, a
further contrasting point is that another prototypical Nrf2-activator, sulforaphane, has
recently been reported to suppress the TGF-β signaling pathway by inhibiting
phosphorylation and transcriptional activation of SMAD3, leading to a reduction in
TGF-β-induced expression of fibrogenic genes (Oh et al., 2012). Although the contributions
made by their anti-fibrotic effects remain unclear, OPZ and NK-252 may also have the same
effect.
There are many animal models of NAFLD, but most are limited as they either simply
demonstrate obesity/insulin resistance-metabolic syndrome-related phenotypes (e.g., fatty
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 26
liver) without replicating the progressive stages of liver pathology exhibited by patients with
NASH (e.g., steatohepatitis and fibrosis), or they show the typical pathology of NASH but the
etiology is not the metabolic syndrome. We therefore need to establish different models that
can be used for appropriate evaluation of target compounds. The CDAA-diet model is
assumed to fall into the latter category, as our preliminary examination revealed that this
model did not show any body weight gain and exhibited markedly reduced levels of
triglycerides in the plasma throughout the 10-weeks study period (unpublished observation),
likely due to the blockage of very-low-density lipoprotein (VLDL) secretion induced by
CDAA-diet (Kawaguchi et al., 2004, Fon and Rozman, 2011).These results indicate that the
metabolic profile of CDAA-diet model is generally not the same or partially converse of
human NASH. In this study, we did not detect any histological changes of fat accumulation
induced by administration of NK-252 or OPZ. However, there is possibility that such
anti-steatotic effects would be detectable using another animal model.
Collectively, these studies revealed that OPZ and NK-252 attenuate the progression of
fibrosis and indicated that their mechanism of action involves both a direct action on HSCs,
and hepatoprotective and anti-inflammatory properties. In other words, Nrf2-activators
probably counteract NASH-related fibrosis in an additive manner. Using Nrf2-activators with
independent mechanisms of action, we demonstrated that the activation of Nrf2 is
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 27
responsible for their anti-fibrotic effects. It has been reported in a model based on a high-fat
diet that rosiglitazone, a peroxisome proliferator-activated receptor (PPAR)-γ ligand,
protects against liver fibrosis in concert with induction of hepatic NQO1 gene expression
(Gupte et al., 2010). Meanwhile, our results provide the first evidence that Nrf2 activation
alone is sufficient to attenuate the progression of NASH-related fibrosis. The strategy of Nrf2
activation is expected to present new opportunities for treatment of NASH with hepatic
fibrosis. Taking into consideration the efficacy of Nrf2-activators against the metabolic
syndrome (Shin et al., 2009, Yu et al., 2011), Nrf2-activators may act as potential
therapeutic agents for patients at any stage of NASH as it progresses.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 28
Authorship Contributions
Participated in research design: Asaoka, Yoshizawa, Aoki, Noda, Yamada, Kaino, Mochizuki,
and Shimozono.
Conducted experiments: Asaoka, Yoshizawa, Aoki, Noda, and Shimozono.
Contributed new reagents or analytic tools: Aoki, and Noda.
Performed data analysis: Asaoka, and Shimozono.
Wrote or contributed to the writing of the manuscript: Shimozono.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 29
References
Blight KJ, McKeating JA, Rice CM (2002) Highly permissive cell lines for subgenomic and
genomic hepatitis C virus RNA replication. J Virol 76: 13001-13014.
Brooks SC 3rd, Brooks JS, Lee WH, Lee MG, Kim SG (2009) Therapeutic potential of
dithiolethiones for hepatic diseases. Pharmacol Ther 124: 31-43.
Brunt EM (2001) Nonalcoholic steatohepatitis: definition and pathology. Semin Liver Dis 21:
3-16.
Chalasani N, Younossi Z, Lavine JE, Diehl AM, Brunt EM, Cusi K, Charlton M, Sanyal AJ
(2012) The diagnosis and management of non-alcoholic fatty liver disease: practice
Guideline by the American Association for the Study of Liver Diseases, American College
of Gastroenterology, and the American Gastroenterological Association. Hepatology 55:
2005-2023.
Chowdhry S, Nazmy MH, Meakin PJ, Dinkova-Kostova AT, Walsh SV, Tsujita T, Dillon JF,
Ashford ML, Hayes JD (2010) Loss of Nrf2 markedly exacerbates nonalcoholic
steatohepatitis. Free Radic Biol Med 48: 357-371.
Comar KM, Sterling RK (2006) Review article: Drug therapy for non-alcoholic fatty liver
disease. Aliment Pharmacol Ther 23: 207-215.
De Bleser PJ, Xu G, Rombouts K, Rogiers V, Geerts A (1999) Glutathione levels discriminate
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 30
between oxidative stress and transforming growth factor-beta signaling in activated rat
hepatic stellate cells. J Biol Chem 274: 33881-33887.
Fon Tacer K, Rozman D (2011) Nonalcoholic Fatty liver disease: focus on lipoprotein and
lipid deregulation. J Lipids 2011: 783976.
García-Trevijano ER, Iraburu MJ, Fontana L, Domínguez-Rosales JA, Auster A,
Covarrubias-Pinedo A, Rojkind M (1999) Transforming growth factor beta1 induces the
expression of alpha1(I) procollagen mRNA by a hydrogen
peroxide-C/EBPbeta-dependent mechanism in rat hepatic stellate cells. Hepatology 29:
960-970.
Gupte AA, Liu JZ, Ren Y, Minze LJ, Wiles JR, Collins AR, Lyon CJ, Pratico D, Finegold MJ,
Wong ST, Webb P, Baxter JD, Moore DD, Hsueh WA (2010) Rosiglitazone attenuates
age- and diet-associated nonalcoholic steatohepatitis in male low-density lipoprotein
receptor knockout mice. Hepatology 52: 2001-2011.
Itoh K, Chiba T, Takahashi S, Ishii T, Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K,
Hatayama I, Yamamoto M, Nabeshima Y (1997) An Nrf2/small Maf heterodimer mediates
the induction of phase II detoxifying enzyme genes through antioxidant response
elements. Biochem Biophys Res Commun 236: 313-322.
Itoh K, Tong KI, Yamamoto M (2004) Molecular mechanism activating Nrf2-Keap1 pathway
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 31
in regulation of adaptive response to electrophiles. Free Radic Biol Med 36: 1208-1213.
Itoh K, Wakabayashi N, Katoh Y, Ishii T, Igarashi K, Engel JD, Yamamoto M (1999) Keap1
represses nuclear activation of antioxidant responsive elements by Nrf2 through binding
to the amino-terminal Neh2 domain. Genes Dev 13: 76-86.
Kang KW, Kim YG, Cho MK, Bae SK, Kim CW, Lee MG, Kim SG (2002) Oltipraz regenerates
cirrhotic liver through CCAAT/enhancer binding protein-mediated stellate cell inactivation.
FASEB J 16: 1988-1990.
Kawaguchi K, Sakaida I, Tsuchiya M, Omori K, Takami T, Okita K (2004) Pioglitazone
prevents hepatic steatosis, fibrosis, and enzyme-altered lesions in rat liver cirrhosis
induced by a choline-deficient L-amino acid-defined diet. Biochem Biophys Res Commun
315: 187-195.
Kim SG, Kim YM, Choi JY, Han JY, Jang JW, Cho SH, Um SH, Chon CY, Lee DH, Jang JJ,
Yu E, Lee YS (2011) Oltipraz therapy in patients with liver fibrosis or cirrhosis: a
randomized, double-blind, placebo-controlled phase II trial. J Pharm Pharmacol 63:
627-635.
Kobayashi M, Li L, Iwamoto N, Nakajima-Takagi Y, Kaneko H, Nakayama Y, Eguchi M,
Wada Y, Kumagai Y, Yamamoto M (2009) The antioxidant defense system Keap1-Nrf2
comprises a multiple sensing mechanism for responding to a wide range of chemical
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 32
compounds. Mol Cell Biol 29: 493-502.
Koek GH, Liedorp PR, Bast A (2011) The role of oxidative stress in non-alcoholic
steatohepatitis. Clin Chim Acta 412: 1297-1305.
Konwinski RR, Haddad R, Chun JA, Klenow S, Larson SC, Haab BB, Furge LL (2004)
Oltipraz, 3H-1,2-dithiole-3-thione, and sulforaphane induce overlapping and protective
antioxidant responses in murine microglial cells. Toxicol Lett 153: 343-355.
Kwak MK, Itoh K, Yamamoto M, Sutter TR, Kensler TW (2001) Role of transcription factor
Nrf2 in the induction of hepatic phase 2 and antioxidative enzymes in vivo by the cancer
chemoprotective agent, 3H-1, 2-dimethiole-3-thione. Mol Med 7: 135-145.
Kwak MK, Wakabayashi N, Itoh K, Motohashi H, Yamamoto M, Kensler TW (2003)
Modulation of gene expression by cancer chemopreventive dithiolethiones through the
Keap1-Nrf2 pathway. Identification of novel gene clusters for cell survival. J Biol Chem
278: 8135-8145.
Murphy FR, Issa R, Zhou X, Ratnarajah S, Nagase H, Arthur MJ, Benyon C, Iredale JP
(2002) Inhibition of apoptosis of activated hepatic stellate cells by tissue inhibitor of
metalloproteinase-1 is mediated via effects on matrix metalloproteinase inhibition:
implications for reversibility of liver fibrosis. J Biol Chem 277: 11069-11076.
Nakae D, Yoshiji H, Mizumoto Y, Horiguchi K, Shiraiwa K, Tamura K, Denda A, Konishi Y
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 33
(1992) High incidence of hepatocellular carcinomas induced by a choline deficient
L-amino acid defined diet in rats. Cancer Res 52: 5042-5045.
Nioi P, McMahon M, Itoh K, Yamamoto M, Hayes JD (2003) Identification of a novel
Nrf2-regulated antioxidant response element (ARE) in the mouse NAD(P)H:quinone
oxidoreductase 1 gene: reassessment of the ARE consensus sequence. Biochem J 374:
337-348.
Oh CJ, Kim JY, Min AK, Park KG, Harris RA, Kim HJ, Lee IK (2012) Sulforaphane attenuates
hepatic fibrosis via NF-E2-related factor 2-mediated inhibition of transforming growth
factor-β/Smad signaling. Free Radic Biol Med 52: 671-682.
Padmanabhan B, Tong KI, Ohta T, Nakamura Y, Scharlock M, Ohtsuji M, Kang MI,
Kobayashi A, Yokoyama S, Yamamoto M (2006) Structural basis for defects of Keap1
activity provoked by its point mutations in lung cancer. Mol Cell 21: 689-700.
Reisman SA, Aleksunes LM, Klaassen CD (2009a) Oleanolic acid activates Nrf2 and
protects from acetaminophen hepatotoxicity via Nrf2-dependent and Nrf2-independent
processes. Biochem Pharmacol 77: 1273-1282.
Reisman SA, Buckley DB, Tanaka Y, Klaassen CD (2009b) CDDO-Im protects from
acetaminophen hepatotoxicity through induction of Nrf2-dependent genes. Toxicol Appl
Pharmacol 236: 109-114.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 34
Roberts AB, Tian F, Byfield SD, Stuelten C, Ooshima A, Saika S, Flanders KC (2006)
Smad3 is key to TGF-beta-mediated epithelial-to-mesenchymal transition, fibrosis, tumor
suppression and metastasis. Cytokine Growth Factor Rev 17: 19-27.
Sanyal AJ, Chalasani N, Kowdley KV, McCullough A, Diehl AM, Bass NM,
Neuschwander-Tetri BA, Lavine JE, Tonascia J, Unalp A, Van Natta M, Clark J, Brunt EM,
Kleiner DE, Hoofnagle JH, Robuck PR (2010) Pioglitazone, vitamin E, or placebo for
nonalcoholic steatohepatitis. N Engl J Med 362: 1675-1685.
Shin S, Wakabayashi J, Yates MS, Wakabayashi N, Dolan PM, Aja S, Liby KT, Sporn MB,
Yamamoto M, Kensler TW (2009) Role of Nrf2 in prevention of high-fat diet-induced
obesity by synthetic triterpenoid CDDO-imidazolide. Eur J Pharmacol 620: 138-144.
Sies H (1997) Oxidative stress: oxidants and antioxidants. Exp Physiol 82: 291-295.
Sreekumar R, Rosado B, Rasmussen D, Charlton M (2003) Hepatic gene expression in
histologically progressive nonalcoholic steatohepatitis. Hepatology 38: 244-251.
Sugimoto H, Okada K, Shoda J, Warabi E, Ishige K, Ueda T, Taguchi K, Yanagawa T,
Nakahara A, Hyodo I, Ishii T, Yamamoto M (2010) Deletion of nuclear factor-E2-related
factor-2 leads to rapid onset and progression of nutritional steatohepatitis in mice. Am J
Physiol Gastrointest Liver Physiol 298: G283-294.
Tong KI, Katoh Y, Kusunoki H, Itoh K, Tanaka T, Yamamoto M (2006) Keap1 recruits Neh2
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 35
through binding to ETGE and DLG motifs: characterization of the two-site molecular
recognition model. Mol Cell Biol 26: 2887-2900.
Valenti L, Rametta R, Dongiovanni P, Maggioni M, Fracanzani AL, Zappa M, Lattuada E,
Roviaro G, Fargion S (2008) Increased expression and activity of the transcription factor
FOXO1 in nonalcoholic steatohepatitis. Diabetes 57: 1355-1362.
Vernon G, Baranova A, Younossi ZM (2011) Systematic review: the epidemiology and
natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in
adults. Aliment Pharmacol Ther 34: 274-285.
Verrecchia F, Chu ML, Mauviel A (2001) Identification of novel TGF-beta /Smad gene targets
in dermal fibroblasts using a combined cDNA microarray/promoter transactivation
approach. J Biol Chem 276: 17058-17062
Yu Z, Shao W, Chiang Y, Foltz W, Zhang Z, Ling W, Fantus IG, Jin T (2011) Oltipraz
upregulates the nuclear factor (erythroid-derived 2)-like 2 [corrected] (NRF2) antioxidant
system and prevents insulin resistance and obesity induced by a high-fat diet in C57BL/6J
mice. Diabetologia 54: 922-934.
Zhang YK, Yeager RL, Tanaka Y, Klaassen CD (2010) Enhanced expression of Nrf2 in mice
attenuates the fatty liver produced by a methionine-and choline-deficient diet. Toxicol Appl
Pharmacol 245: 326-334.
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 36
Figure Legends
Fig. 1. Chemical structures of OPZ and NK-252.
Fig. 2. OPZ and NK-252 activate NQO1-ARE, which are inhibited by Nrf2 knock-down.
Huh-7.5 cells with stable expression of an ARE-luciferase reporter gene were transfected
with negative control (NC) siRNA or Nrf2 siRNA. After transfection (30 h), cells were treated
for 16 h with 0.3-30 μmol/L OPZ or NK-252. The average luciferase activity value for control
cells with NC siRNA transfection was arbitrarily set to 100%, and all other results were
expressed relative to this. The results shown represent the means ± SD from duplicate
assays for OPZ (A) and NK-252 (B).
Fig. 3. OPZ and NK-252 have protective effects against H2O2-induced cytotoxicity. Huh-7
cells were treated for 24 h with 0.3-30 μmol/L OPZ or NK-252, and then cells were washed
and treated for 24 h with 1 mmol/L H2O2. The average cell viability value for control cells
without H2O2 was arbitrarily set to 100%, and all other results were expressed relative to this.
The results shown represent the means ± SD from triplicate assays for OPZ (A) and NK-252
(B).
Fig. 4. NK-252 but not OPZ interacts directly with the Keap1-DC domain which contains the
Nrf2-binding site. Interaction between Keap1-DC domain and OPZ or NK-252 was
determined by the BIAcore™ assay. 12.5−100 μmol/L OPZ or NK-252 were applied to a
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 37
Keap1-DC-bounded sensor chip. The resonance units reflect the number of attached
molecules. Each line indicates the resonance unit curve of 12.5, 25, 50, 100 μmol/L
compounds with Keap1-DC, respectively from singlet assays for OPZ (A) and NK-252 (B).
Fig. 5. OPZ and NK-252 exert histopathological anti-fibrotic efficacies in the liver of NASH
model rats. (A-E) Histopathological evaluation was performed with the liver of rats fed a diet
of CRF-1 for 10 weeks (naive), rats fed a diet of CDAA for 10 weeks (CDAA control), or rats
on a CDAA diet with nine-week administration of OPZ or NK-252 (CDAA+OPZ,
CDAA+NK-252_low or CDAA+NK-252_high). (A-C) Serial or near-serial sections stained
with H&E (A) Sirius red (B) and α-SMA (C) (scale bar, 1 mm). (D) Histopathological fibrosis
scores were evaluated with the modified Brunt’s method consisting of score-1 to -4. (E)
Quantitative evaluation of Sirius red-positive fibrosis area. The results shown represent
individual scores or means ± SEM (N=3-8). † P < 0.05, and †† P < 0.01 between each
compound administration group versus CDAA control (Steel’s test).
Fig. 6. OPZ and NK-252 upregulate NQO1 gene expression, exert antioxidant property and
downregulate fibrogenic gene expression in the liver of rats on a CDAA diet. (A, C-E) Total
RNA was isolated from the liver of rats described in the legend of Fig. 5, and then real-time
qPCR was performed. The results shown represent means ± SEM (N=3-8) of the gene
expression of NQO1 (A), TGF-β1 (C), COL1A1 (D) and TIMP-1 (E) relative to those of naive
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 38
rats. (B) Protein carbonyl derivatives in each liver homogenate of rats described in the
legend of Fig. 5 were detected by an ELISA assay. The results shown represent means ±
SEM (N=3-8) of nanomole of carbonyl derivatives per milligram of total proteins assayed.* P
< 0.05, ** P < 0.01, and *** P < 0.001 between each compound administration group versus
CDAA control (Dunnett’s test). † P < 0.05, and †† P < 0.01 between each compound
administration group versus CDAA control (Steel’s test).
Fig. 7. NK-252 prevents further progression of established fibrosis in the liver of NASH
model rats. (A, B) Histopathological evaluation was performed with the liver of rats on a
six-week CRF-1 diet rats (pre-naive), rats on a six-week CDAA diet (pre-CDAA control), rats
on a 10-week CRF-1 diet (naive), rats on a 10-week CDAA diet (CDAA control), or rats on a
CDAA diet with four-week administration of NK-252 (CDAA+NK-252_DA). The procedure
was described in the legend of Figure 5. The results shown represent fibrosis score (A) and
means ± SEM (N=3-8) of rate of liver fibrosis area (B). †P < 0.05 between each compound
administration group versus CDAA control (Steel’s test).
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

MOL #84269 39
Tables
Table 1
Weighing data and biochemical data
Group N
Age
(week)
Body weight
(g)
Liver wet weight
(g)
Plasma ALT
(IU/L)
Plasma AST
(IU/L)
naive 3 16 259.3 ± 3.5 7.57 ± 0.29 28.0 ± 2.2 104.0 ± 11.1
CDAA control 9 16 212.4 ± 3.7 10.32 ± 0.20 170.7 ± 9.6 344.4 ± 17.3
CDAA+OPZ 8 16 227.0 ± 3.4 * 13.88 ± 0.3 *** 152.6 ± 8.1 256.8 ± 14.4 ***
CDAA+NK-252_low 8 16 216.5 ± 3.2 12.25 ± 0.18 *** 164.4 ± 11.8 285.3 ± 11.8 *
CDAA+NK-252_high 8 16 226.4 ± 4.2 * 14.09 ± 0.37 *** 120.8 ± 3.2 ** 238.5 ± 9.6 ***
pre-naive 3 12 245.0 ± 0.9 8.63 ± 0.23 22.0 ± 0.8 94.0 ± 18.5
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
Molecular Pharm
acology Fast Forward. Published on A
pril 16, 2013 as DO
I: 10.1124/mol.112.084269
at ASPET Journals on April 4, 2019 molpharm.aspetjournals.org Downloaded from

MOL #84269 40
pre-CDAA control 9 12 198.9 ± 2.9 12.94 ± 0.24 245.0 ± 16.1 346.3 ± 9.2
CDAA+NK-252_DA 7 16 223.7 ± 3.6 13.27 ± 0.36 *** 150.4 ± 10.6 219.0 ± 12.5 ***
* P < 0.05, ** P < 0.01, and *** P < 0.001 between each compound administration group versus CDAA control (Dunnett's test)
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
Molecular Pharm
acology Fast Forward. Published on A
pril 16, 2013 as DO
I: 10.1124/mol.112.084269
at ASPET Journals on April 4, 2019 molpharm.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from
This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from

This article has not been copyedited and formatted. The final version may differ from this version.Molecular Pharmacology Fast Forward. Published on April 16, 2013 as DOI: 10.1124/mol.112.084269
at ASPE
T Journals on A
pril 4, 2019m
olpharm.aspetjournals.org
Dow
nloaded from


















