
Northern Goshawk (Accipiter gentilis laingi)
23
Northern Goshawk (Accipiter gentilis laingi) Post-Fledging Area Size Estimation on Vancouver Island, BC Photo Credit: Todd Luoma by Erica L. McClaren and Christina L. Pendergast December 2002 Prepared for: Ministry of Water, Land and Air Protection, Canadian Forest Products, Weyerhaeuser, Timberwest, Western Forest Products, and B.C. Conservation Foundation
Transcript of Northern Goshawk (Accipiter gentilis laingi)

Northern Goshawk (Accipiter gentilis laingi)
Post-Fledging Area Size Estimation
on
Vancouver Island, BC
a
Decem PrepaWeye
Photo Credit: Todd Luom
by
Erica L. McClaren
and
Christina L. Pendergast
ber 2002
red for: Ministry of Water, Land and Air Protection, Canadian Forest Products, rhaeuser, Timberwest, Western Forest Products, and B.C. Conservation Foundation

Abstract Managing for avian species with long post-fledging parental care periods requires
knowledge of post-fledging area (PFA) size and habitat characteristics, and the length of post-fledging periods. This information was unavailable for the northern goshawk subspecies Accipiter gentilis laingi inhabiting Vancouver Island, British Columbia. Our objectives were: 1) to estimate PFA size and length of the post-fledging period at northern goshawk nests on Vancouver Island; and 2) to compare our results to those reported for other goshawk subspecies (A. g. atricapillus, A. g. gentilis). We used 95% adaptive kernels to estimate PFA size from 12 fledglings at 12 nests using visual and road telemetry between 29 June and 2 September, 2001-2002. A total of 236 radio telemetry locations were collected from 15 fledglings, although only 12 had sufficient locations to estimate PFA size. Ninety-three percent of fledgling locations were within 200 m of nests during the first three weeks post-fledging but only 42.4% of locations were within this distance during the remaining four weeks post-fledging. Northern goshawks dispersed from PFAs 45.9±1.3 days post-fledging. Mean PFA size was 58.6±16.2 ha (95% CI: 22.9, 94.3 ha). Our estimated mean PFA size for northern goshawk fledglings on Vancouver Island was smaller than size estimates reported for A. g. atricapillus in New Mexico and for A. g. gentilis in Sweden but was larger than estimates reported for A. g. atricapillus in interior British Columbia. We recommend leaving a minimum of 132 ha for PFAs, the upper confident limit size for PFAs plus a 100 m buffer. This area will include PFAs around multiple nest sites within goshawk nest areas and prevent edge influences such as predation on fledglings from edge-adapted predators and wind throw events from degrading PFA habitat.
Introduction
Northern goshawk (Accipiter gentilis), hereafter referred to as goshawk, breeding home ranges appear to be composed of a nest area (NA), a post-fledging area (PFA), and a foraging area (FA) (Reynolds et al. 1992). The NA often contains several alternative nest trees, roost trees, plucking posts, and serves as the center of courtship behaviours and fledgling movements (Reynolds et al. 1992, Kennedy et al. 1994). Goshawk nest areas may occur within one or multiple forest stands (Reynolds et al. 1992, Squires and Reynolds 1992). NAs vary in size and shape depending on topography and the availability of suitable habitat (Reynolds 1983, Ethier 1999).
Kennedy et al. (1994) described the PFA as the area surrounding and including the NA, with its size roughly corresponding with the female core-use area (Kennedy et al. 1994). In this paper, we define the PFA as the area used by fledglings after they fledge and before they become independent of adults and disperse from their natal territory. PFAs are considered to provide critical habitat for young goshawks which are extremely vulnerable to predation when they are learning to fly and hunt (Reynolds et al. 1992, Kennedy et al. 1994, Daw and DeStefano 2001). Because fledglings are being primarily fed by adults while in PFAs, their ability to access prey deliveries and feed without disturbance, may dramatically influence their dispersal success and first-year survivorship (Wood et al. 1998, Powell et al. 2002). Thus, PFAs are characterized by an abundance of the habitat attributes critical for goshawk prey, perching and feeding sites (snags, coarse woody debris), and by extensive canopy cover (> 50%) which provides
2

protection to fledglings from predators (Reynolds et al. 1992, Kennedy et al. 1994, Daw and DeStefano 2001).
FAs make up most of an individual’s breeding home range and are comprised of adult male and female hunting areas. FAs may or may not include the NA and PFA. Kennedy et al. (1994) suggest adult males do not hunt directly within the NA and PFA in order to maintain locally abundant food supplies for adult females and for fledglings while they are learning to hunt. FAs vary in size among locales and among individual goshawks according to an individual’s experience within its breeding home range, differences in hunting efficiency, food requirements (brood size) and the availability of food within home ranges (Kennedy et al. 1994). Context of this Study
The size and habitat characteristics of PFAs have been studied for several raptors including buzzards (Buteo buteo; Walls and Kenward 1998), bald eagles (Haliaeetus leucocephalus; Wood et al. 1998), golden eagles (Aquila chrysaetos; O’Toole et al. 1999) and spotted owls (Strix occidentalis lucida; Willey and van Riper 2000). To ensure suitable breeding habitat for avian species, it is essential to provide adequate nest sites and PFAs; these two aspects of an individual’s breeding habitat requirements are inextricably linked. This is especially important in species with long post-fledging parental care periods. Juveniles are vulnerable during the post-fledging period and PFA habitat characteristics likely influence the probability of young surviving to dispersal (Wood et al. 1998, Powell et al. 2002). Furthermore, the number of young that successfully disperse from a breeding area is often considered a measure of adult female reproductive output (Clutton-Brock 1988, Green and Cockburn 2001).
Goshawks are a species of concern within North America (Kennedy 1997, Crocker-Bedford 1998, DeStefano 1998) and Europe (Widen 1997). However, we know little about goshawk behaviour and habitat use during the post-fledging period and even less about goshawk natal dispersal (Kennedy and Ward In Press). In British Columbia, the Identified Wildlife Management Strategy, Volume I (BC Ministry of Forests and BC Ministry of Environment 1999) recommends a 240 ha PFA be placed around designated goshawk (A. g. laingi) nesting areas. The size of this PFA in BC was based on PFA size estimates for goshawks inhabiting the southwestern USA by Reynolds et al. (1992) and Kennedy et al. (1994). Similar PFA estimates were reported by Kenward et al. (1993a) for goshawks in Sweden. Goshawk research in the Kispiox/Lakes Districts of central BC suggests goshawk PFA size may be much smaller, averaging < 20 ha (Doyle and Mahon 2000, Mahon and Doyle 2001). Therefore, our objective was to estimate the size of goshawk PFAs on Vancouver Island, BC around nests in a variety of forest types, surrounded by different harvest regimes. We compare PFA size estimates for A. g. laingi within coastal forests to PFA size estimates for A. g. atricapillus (southwestern USA and interior BC) and A. g. gentilis (Sweden). Our knowledge of goshawk PFA size and habitat characteristics on Vancouver Island is crucial for us to adequately manage the breeding habitat of this goshawk subspecies which is designated by COSEWIC as ‘Threatened’ in Canada (Cooper and Chytyk 2000) and is provincially Red-listed (Ministry of Environment, Lands and Parks 2000).
3

Methods Study Area
Goshawk nest areas were located on Vancouver Island, BC in forests dominated by western hemlock (Tsuga heterophylla) and Douglas-fir (Pseudotsuga menziesii), although western redcedar (Thuja plicata), amabalis fir (Abies amabalis), and red alder (Alnus rubra) were also abundant (Fig. 1). The predominant age of nest stands ranged from 45 to >250 years. Elevations of nest areas ranged from 150-850 m. Mean daily temperatures ranged from 4.1 C° in the winter (October to April) to 14.3 C° in the summer (May through September), and mean monthly precipitation ranged from 40 mm (July) - 234 mm (December), with a mean annual total of 1409 mm, of which most was rain (Environment Canada 1998).
Data Collection
The 15 goshawk nests used in this study were located either by using broadcast surveys of conspecific calls, or incidentally by forest company personnel and the public between 1994-2002. Broadcast survey methodology is described in detail in McClaren et al. (2002). Nest trees were visited in May to verify activity by the presence of incubating goshawks. When nests were verified as active, they were left undisturbed until a week prior to hatching (15 May – 1 June). During this time, we visited nests frequently to estimate hatching dates based on the adult female’s position within nests and behaviour. After hatching, we monitored the number and age of chicks. When nestlings were approximately 21 days, we climbed nest trees using the single rope technique (I. S. Jacobs, personal communication). Nestlings were lowered to the ground in a modified SilviCool III sac to ensure their protection and to regulate their temperature. Nestlings were weighed, measured, sexed and aged. The same individual (ELM) conducted all nestling procedures to maintain consistency in the data. Nestlings were sexed using tarsal width recommendations provided by Kenward et al. (1993a) where male tarsal widths were <6.5 mm and female tarsal widths were ≥6.5 mm. We aged nestlings from a photographic key developed by Boal (1994) and from our estimated hatch dates. Nestlings were banded with USFWS bands and with colour rivet bands (Acraft Sign and Nameplate Co. Ltd., Edmonton, Alberta). Radio-tagged nestlings received a unique band colour from their nest mates. We fitted the largest female nestling in each nest with a 9 g tarsal mount transmitter with a mortality switch (Advanced Technology Services, Isanti, Minnesota). There were two exceptions where we fitted males with transmitters: 1) all nestlings were male; and 2) the female fledgling’s transmitter battery died and so we captured it’s male sibling and affixed a new radio transmitter. The remaining nestlings were not radio-tagged. We chose the largest female to reduce variation in fledgling movements and dispersal dates caused by gender differences and because the extra weight from transmitters would have less impact on larger females, increasing their probability of surviving until dispersal. Radio transmitters were attached to nestling tarsi with a leather jesse that we designed so that fledglings could remove them after the 90 day transmitter battery expired (Ward and Kennedy 1996, Dewey and Kennedy 2001).
Prior to collecting fledgling location data, we laid out 600 x 600-m grids with location reference stations, centered around active nests (Fig. 2). After nestlings were
4
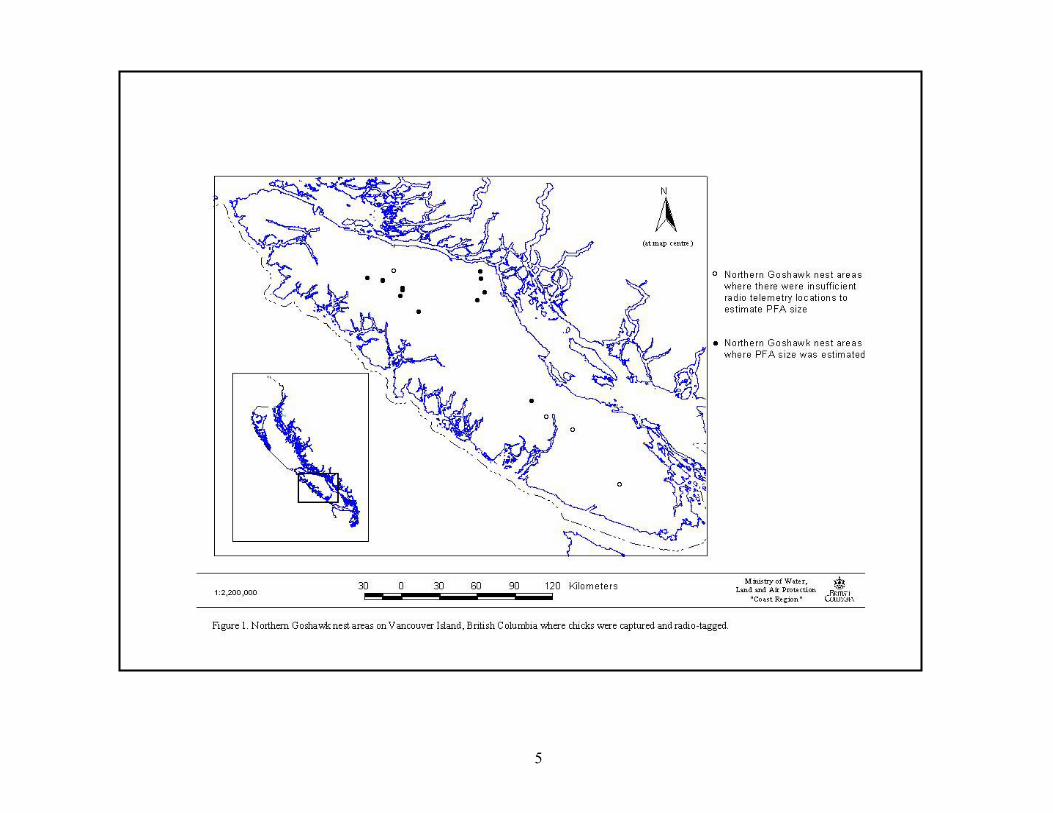
5

radio-tagged, nests were visited weekly to monitor chick development, survivorship, and transmitter operation. Once tagged nestlings fledged, nest areas were visited every 1-3 days to collect fledgling location data. We rotated sampling equally among nest areas and sampling periods (<800 - 1100, 1101 - 1400, 1400 - >1700) to ensure all times of day were equally represented. Teams of two listened for radio signals prior to entering nest stands. Using a 3-element Yagi antenna (Telonics Inc., Mesa, Arizona) and receiver (Models SRX-1000, SRX-400, Lotek Engineering Inc., Ontario), observers quietly approached fledglings on foot and obtained visual locations. We verified the observed fledgling was radio tagged with colour band identification. At one nest site where the radio-tagged fledgling was predated 5 days prior to dispersal, we continued locations on the remaining two siblings through audio locations. At another nest site where the radio transmitter battery failed prematurely, we continued locations on a sibling from the same nest that we captured and affixed a new transmitter. We measured distance and direction of the fledgling to the closest reference grid station using a meter-marked rope and compass. Fledgling behaviour, perch location and proximity to sibling(s) were recorded when possible. When fledglings became more mobile and visual locations were difficult to achieve without influencing their locations, we used triangulation from road stations to estimate fledgling locations. We estimated dispersal dates as mid-way between the last visit when fledglings were verified ≤1.6 km from nests and when no radio signal was heard within this distance on two consecutive telemetry sessions. Our dispersal definition parallels Kenward et al. (1993a) and Kennedy et al. (1994). We did not include locations collected from the Claud Elliot fledgling because this bird was ill and was recovered dead three weeks after fledging. We were unsure whether it exhibited a ‘normal’ activity pattern before it died.
N
Goshawk nest
Fledglinglocation
100 m
Reference station
100 m
Figure 2. Reference station grid layout around goshawk nests for calculating fledgling locations from visual observations on Vancouver Island 2001-2002.
6

Location Determination We used the survey mapping extension of Road Engineering Software (Softree
Technical Systems Inc. 1998) to calculate UTM coordinates for visual fledgling locations based on the known GPSed nest tree location, grid reference station locations, and measured offsets from grid reference stations to fledglings. Fledgling locations from road telemetry and associated error ellipses from triangulation were estimated using Locate II version 1.5 (Nams 2000) based on the number of bearings, angles between bearings, and the distance from bearing locations to radio-tagged birds. Post-fledging Area Estimation
We used the HRE home range extension software (Rodgers and Carr 1998) for ArcView (version 3.2, Environmental Systems Research Institute, Inc., Redlands, California) to estimate PFA size. Although PFAs are not equivalent to home ranges, we assumed that the area used by fledglings prior to dispersal could be modeled using home range estimation techniques. Ninety-five percent adaptive kernel estimates were used to estimate PFA size. We felt adaptive kernels provided the most accurate reflection of fledgling use patterns around nests and were more conservative than minimum convex polygons because they used probability functions around bird locations to estimate boundaries rather than merely connecting the outermost data points. Triangulated locations with associated errors >1 km circle radius were not included in PFA estimates. As well we did not estimate PFA size for fledglings with <15 locations (n = 3).
Statistical Analyses
We considered fledgling location data collected from different nest sites within the same nest area in different years (i.e., Toad01, Toad02) to be independent. Mean and median hatching, fledging and dispersal dates were estimated from Julian days. We used the correlation procedure in SAS (SAS Institute, Inc. 1997) to examine the relationship between fledging date and dispersal date. We used a Fisher Exact Test to test whether fledglings moved farther from nests over time. Distances were calculated as the Euclidian distance from the nest tree to the fledgling location. We divided the distance we observed fledglings from nests into five categories (0-99 m, 100-199 m, 200-399 m, 400-799 m, >800 m) and three time intervals (1-3 weeks, 4&5 weeks, 6&7 weeks). We used Wilcoxon rank-sum tests (PROC NPAR1WAY; SAS Institute, Inc. 1997) to compare hatching, fledging, and dispersal dates, and PFA size between years. We used Wilcoxon rank-sum tests for pairwise these comparisons because small sample sizes and nonnormal data may invalidate the results provided by t-tests (Ott 1993). Results were considered statistically significant if P < 0.1. Means and standard errors are presented unless otherwise stated. Results
Forty-two goshawk nestlings from 17 nests were banded and measured. Mean age of banding, nest sex ratio, mass and tarsus widths are presented in Table 1. Nests contained 40% more females than males. Sex ratios were female-biased in 2001 but they were almost equal in 2002. Within broods of two, nestlings were often 1-2 days apart in age whereas within broods of three, there was frequently a runt that was 3-4 days younger
7
than its siblings. Although we used Boal (1994) to age nestlings, we constructed a local photographic aging key from our observations (Appendix I). Table 1. Mean goshawk nestling morphometrics collected from nests (n = 17 ) on Vancouver Island, 9-26 June 2001- 2002.
Sex
ratio (M:F)
Banding age
(days)
Male (n = 15)
Female (n = 27)
Mass Tarsus width Mass Tarsus Width 0.6:1.0 20.2 ± 0.8 530.4 ± 25.3 6.1 ± 0.1 574.6 ± 27.9 7.3 ± 0.1
Breeding Phenology
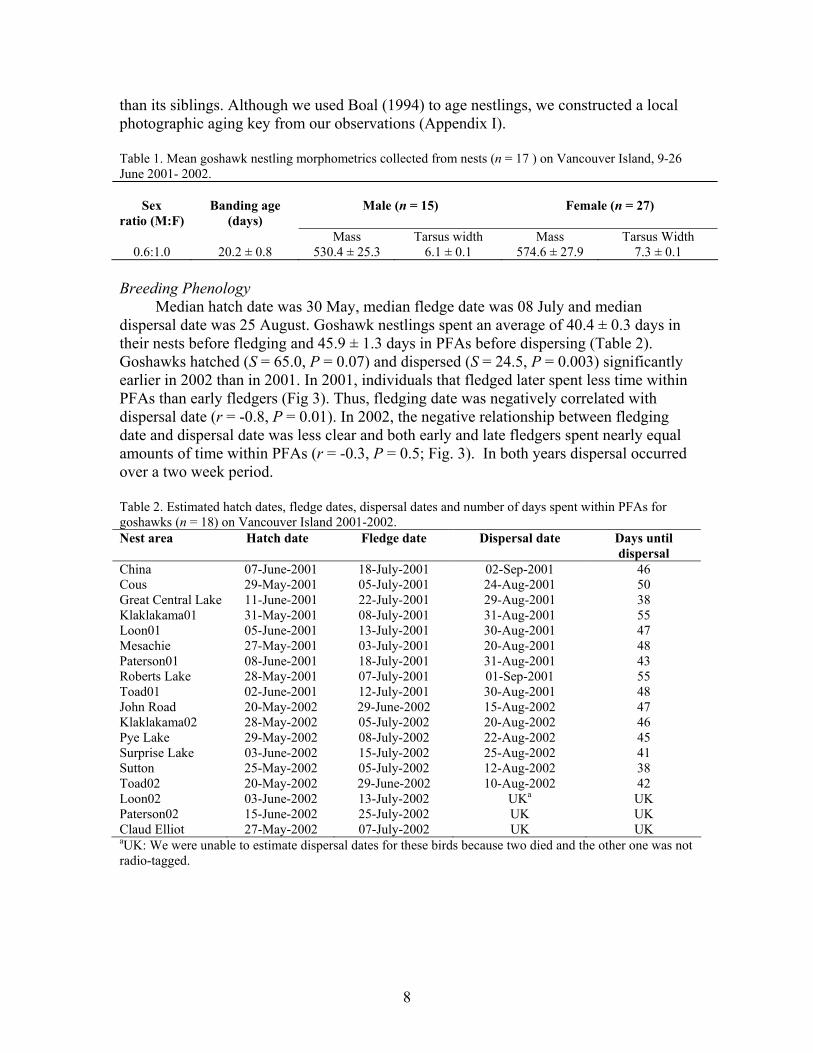
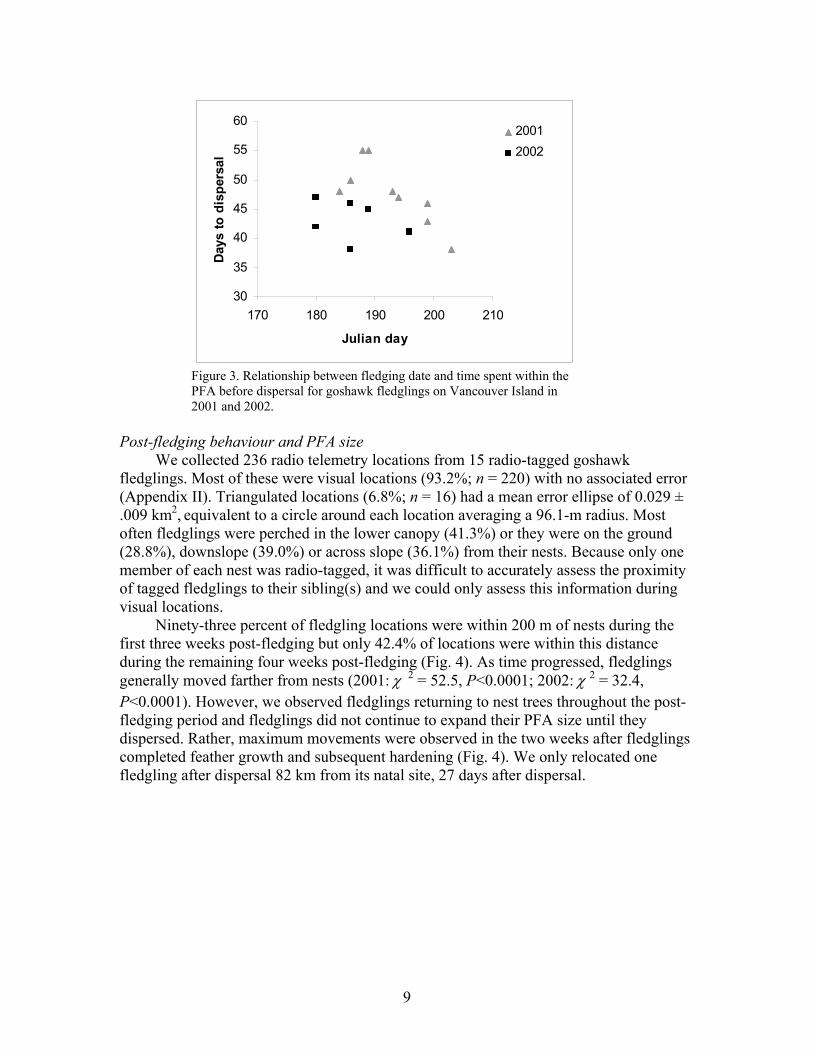
Median hatch date was 30 May, median fledge date was 08 July and median dispersal date was 25 August. Goshawk nestlings spent an average of 40.4 ± 0.3 days in their nests before fledging and 45.9 ± 1.3 days in PFAs before dispersing (Table 2). Goshawks hatched (S = 65.0, P = 0.07) and dispersed (S = 24.5, P = 0.003) significantly earlier in 2002 than in 2001. In 2001, individuals that fledged later spent less time within PFAs than early fledgers (Fig 3). Thus, fledging date was negatively correlated with dispersal date (r = -0.8, P = 0.01). In 2002, the negative relationship between fledging date and dispersal date was less clear and both early and late fledgers spent nearly equal amounts of time within PFAs (r = -0.3, P = 0.5; Fig. 3). In both years dispersal occurred over a two week period. Table 2. Estimated hatch dates, fledge dates, dispersal dates and number of days spent within PFAs for goshawks (n = 18) on Vancouver Island 2001-2002. Nest area Hatch date Fledge date Dispersal date Days until
dispersal China 07-June-2001 18-July-2001 02-Sep-2001 46 Cous 29-May-2001 05-July-2001 24-Aug-2001 50 Great Central Lake 11-June-2001 22-July-2001 29-Aug-2001 38 Klaklakama01 31-May-2001 08-July-2001 31-Aug-2001 55 Loon01 05-June-2001 13-July-2001 30-Aug-2001 47 Mesachie 27-May-2001 03-July-2001 20-Aug-2001 48 Paterson01 08-June-2001 18-July-2001 31-Aug-2001 43 Roberts Lake 28-May-2001 07-July-2001 01-Sep-2001 55 Toad01 02-June-2001 12-July-2001 30-Aug-2001 48 John Road 20-May-2002 29-June-2002 15-Aug-2002 47 Klaklakama02 28-May-2002 05-July-2002 20-Aug-2002 46 Pye Lake 29-May-2002 08-July-2002 22-Aug-2002 45 Surprise Lake 03-June-2002 15-July-2002 25-Aug-2002 41 Sutton 25-May-2002 05-July-2002 12-Aug-2002 38 Toad02 20-May-2002 29-June-2002 10-Aug-2002 42 Loon02 03-June-2002 13-July-2002 UKa UK Paterson02 15-June-2002 25-July-2002 UK UK Claud Elliot 27-May-2002 07-July-2002 UK UK aUK: We were unable to estimate dispersal dates for these birds because two died and the other one was not radio-tagged.
8
30
35
40
45
50
55
60
170 180 190 200 210
Julian day
Day
s to
dis
pers
al
20012002
Figure 3. Relationship between fledging date and time spent within the PFA before dispersal for goshawk fledglings on Vancouver Island in 2001 and 2002.
Post-fledging behaviour and PFA size
We collected 236 radio telemetry locations from 15 radio-tagged goshawk fledglings. Most of these were visual locations (93.2%; n = 220) with no associated error (Appendix II). Triangulated locations (6.8%; n = 16) had a mean error ellipse of 0.029 ± .009 km2, equivalent to a circle around each location averaging a 96.1-m radius. Most often fledglings were perched in the lower canopy (41.3%) or they were on the ground (28.8%), downslope (39.0%) or across slope (36.1%) from their nests. Because only one member of each nest was radio-tagged, it was difficult to accurately assess the proximity of tagged fledglings to their sibling(s) and we could only assess this information during visual locations.
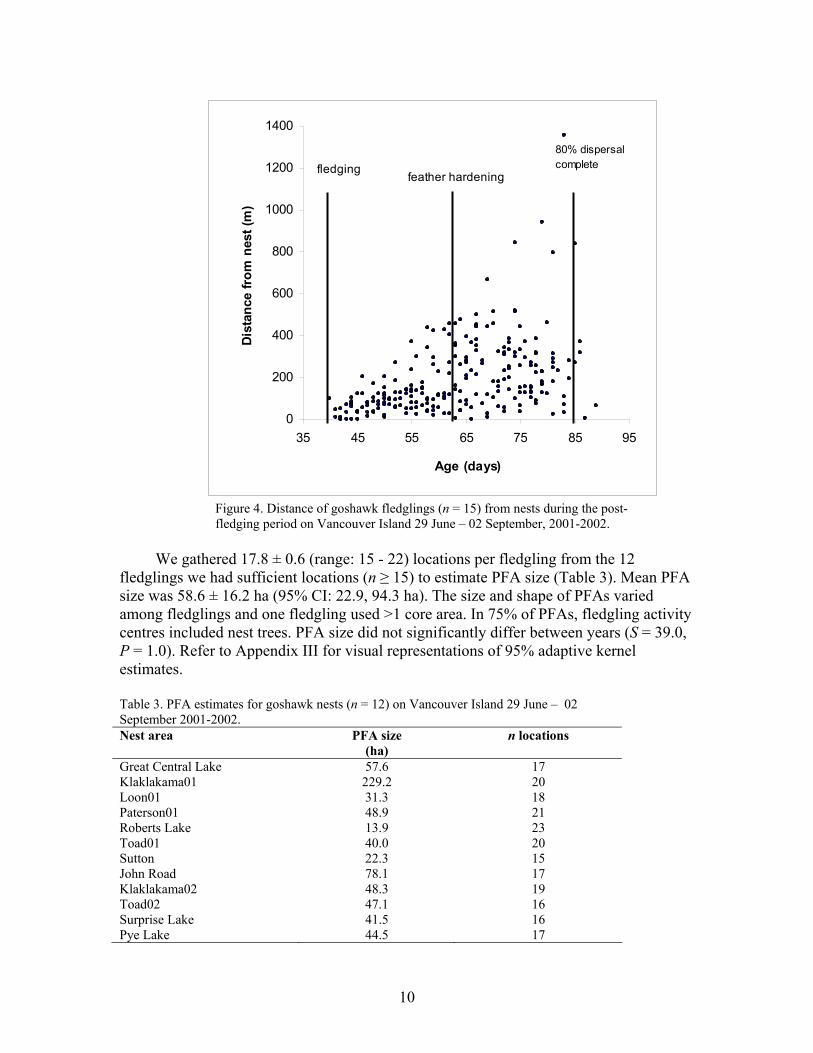
Ninety-three percent of fledgling locations were within 200 m of nests during the first three weeks post-fledging but only 42.4% of locations were within this distance during the remaining four weeks post-fledging (Fig. 4). As time progressed, fledglings generally moved farther from nests (2001: χ 2 = 52.5, P<0.0001; 2002: χ 2 = 32.4, P<0.0001). However, we observed fledglings returning to nest trees throughout the post-fledging period and fledglings did not continue to expand their PFA size until they dispersed. Rather, maximum movements were observed in the two weeks after fledglings completed feather growth and subsequent hardening (Fig. 4). We only relocated one fledgling after dispersal 82 km from its natal site, 27 days after dispersal.
9
0
200
400
600
800
1000
1200
1400
35 45 55 65 75 85 95
Age (days)
Dis
tanc
e fr
om n
est (
m)
fledgingfeather hardening
80% dispersalcomplete
Figure 4. Distance of goshawk fledglings (n = 15) from nests during the post-fledging period on Vancouver Island 29 June – 02 September, 2001-2002.
We gathered 17.8 ± 0.6 (range: 15 - 22) locations per fledgling from the 12
fledglings we had sufficient locations (n ≥ 15) to estimate PFA size (Table 3). Mean PFA size was 58.6 ± 16.2 ha (95% CI: 22.9, 94.3 ha). The size and shape of PFAs varied among fledglings and one fledgling used >1 core area. In 75% of PFAs, fledgling activity centres included nest trees. PFA size did not significantly differ between years (S = 39.0, P = 1.0). Refer to Appendix III for visual representations of 95% adaptive kernel estimates. Table 3. PFA estimates for goshawk nests (n = 12) on Vancouver Island 29 June – 02 September 2001-2002. Nest area PFA size
(ha) n locations
Great Central Lake 57.6 17 Klaklakama01 229.2 20 Loon01 31.3 18 Paterson01 48.9 21 Roberts Lake 13.9 23 Toad01 40.0 20 Sutton 22.3 15 John Road 78.1 17 Klaklakama02 48.3 19 Toad02 47.1 16 Surprise Lake 41.5 16 Pye Lake 44.5 17
10

Survivorship
Brood reduction occurred at a minimum of seven nests prior to banding. At least three nestlings died post-banding but pre-fledging. In 2001, 100% of radio-tagged fledglings survived through the post-fledging period until dispersal and we did not observe any mortality of untagged fledglings (Table 4). However, in 2002, 37.5% of radio-tagged fledglings and at least one untagged fledgling died prior to dispersal. One tagged and one untagged fledgling were predated. Another fledgling was collected while it was dying from an unknown cause and a fourth fledgling’s transmitter signal was on mortality in a tree, approximately 100 m from its nest shortly after it fledged (Table 4). Seventy-five percent of the fledgling mortalities occurred one month post-fledging. We experienced one premature battery failure with John Road_a and so we captured its sibling and placed a new transmitter on it. We continued locations on the John Road_b fledgling and it survived to disperse but we relocated it via air after dispersal and the transmitter mortality switch was on. Thus, we can not conclude whether this bird dropped its transmitter or died. Additionally, the Sutton fledgling removed its transmitter just prior to dispersing.
Table 4. Survivorship information from radio-tagged goshawk fledglings 29 June – 11 September, 2001-2002. Year tagged Fledgling ID Date last observed Fate 2001 China 27-Aug-01 dispersed 2001 Cous 22-Aug-01 dispersed 2001 Great Central Lake 27-Aug-01 dispersed 2001 Mesachie 16-Aug-01 dispersed 2001 Klaklakama01 24-Aug-01 dispersed 2001 Loon01 24-Aug-01 dispersed 2001 Paterson01 25-Aug-01 dispersed 2001 Roberts Lake 25-Aug-01 dispersed 2001 Toad01 24-Aug-01 dispersed 2002 Claud Elliot 29-July-02 dead (unknown cause) 2002 Toad02 05-Aug-02 dead (predated) 2002 John Road_a 03-Aug-02 battery failed 2002 John Road_b 11-Sep-02 mortality switch on 2002 Loon02 14-July-02 dead (unknown cause) 2002 Sutton 10-Aug-02 dispersed 2002 Klaklakama02 19-Aug-02 dispersed 2002 Pye Lake 21-Aug-02 dispersed 2002 Surprise Lake 23-Aug-02 dispersed Discussion Post-fledging and dispersal behaviour
Goshawk fledglings on Vancouver Island exhibited similar behaviour during the post-fledging period to goshawks in Sweden (Kenward et al. 1993a), New Mexico (Kennedy et al. 1994), and Nevada (Shipman 1997). Most fledglings experienced a behavioural transition period approximately 24 days post-fledging. In the first 21 days post-fledging, fledglings were within 200 m of nests, either on the ground or perched in the lower to mid-canopy. Beyond 28 days post-fledging, fledglings were farther from nests, they were adept fliers and they were perched in the upper canopy, often on treetops. This change in fledgling behaviour corresponds with the completion of primary
11

and rectrice feather growth and subsequent feather hardening (Kenward et al. 1993a). The farthest distance fledglings were observed from nests peaked within 1-2 weeks after they completed feather growth. Kenward et al. (1993a) also described this pattern for goshawks in Sweden and Minguez et al. (2001) for Bonelli’s eagles (Hieraaetus fasciatus). This suggests that fledglings don’t continue to expand their PFA size indefinitely until dispersal. Rather, they typically expand their PFA size to its maximum just after completion of feather growth and spend the remaining post-fledging period within these boundaries until dispersal. Thus, fledgling locations should be collected equally throughout the post-fledging period to accurately estimate PFA size and not concentrated in the time just before dispersal.
The breeding season began earlier on Vancouver Island in 2002 than in 2001 with goshawk chicks hatching approximately 1 week earlier and dispersing 10 days earlier than in 2001. Most fledglings dispersed abruptly between 80-96 days of age (10 Aug – 2 Sep) in both years. This appears to be a common pattern for goshawks (Kenward et al. 1993a, Dewey and Kennedy 2001) and for many other raptors (spotted owls: Willey and van Ripper 2000; Bonelli’s eagles: Minguez et al. 2001), although Walls and Kenward (1998) reported a biomodal dispersal pattern for buzzards. In 2001, fledging date was negatively correlated with dispersal date with late fledgers spending less time within PFAs than early fledgers. Kenward et al. (1993b) reported a similar relationship between hatch date and dispersal age among female goshawks in Sweden. However, in 2002, early fledgers and late fledgers spent a similar amount of time within PFAs on Vancouver Island. The difference among years may have resulted from a later onset of breeding in 2001. A shorter post-fledging period could result in reduced survival for late fledgers because they have fewer days to develop flight and hunting skills and they spend less time accumulating mass while being fed by parents. In several bird species, an early onset of breeding indicates higher food availability within the nest area which often results in higher fledgling mass and survival (Dewey and Kennedy 2001, Naef-Daenzer et al. 2001, Aparicio and Bonal 2002).
Because we did not manipulate any proximate factors that may have influenced the length of the post-fledging period we can only speculate on what caused fledglings to disperse. Food supplementation experiments performed by Kenward et al. (1993a) and Dewey and Kennedy (2001) demonstrated increased food availability significantly increased the length of goshawk post-fledging periods. Kenward et al. (1993a) believed goshawk dispersal was caused by behavioural maturation but was accelerated by food shortages. Thus, in 2001, reduced provisioning rates by adults may have forced young to leave the PFA. A later onset of breeding in 2001 may have resulted in reduced prey availability within breeding home ranges and a time crunch for parents to physically recover from breeding and complete their molt before winter (Kenward et al. 1993a). In contrast, when food availability does not decrease within breeding home ranges, post-fledging periods are fixed in length, with late fledgers spending similar amounts of time within PFAs as early fledgers (Florida: bald eagle, Wood et al. 1998; Spain: Bonelli’s eagle, Minguez et al. 2001). This scenario closely resembles what we observed in 2002. We found no evidence that parental aggression caused fledglings to disperse on Vancouver Island which supports experimental results provided by Kenward et al. (1993a).
12

With the exception of one fledgling, we were unable to locate all fledglings after they dispersed, suggesting fledglings initially traveled far. Kenward et al. (1993b) documented goshawk fledglings dispersing from one nest area to another and being fed by other adults. We did not document this in our study, although in previous investigations we trapped three goshawk fledglings within a nest area that fledged only two (E. L. McClaren, personal observation 1997). Initial dispersal distances appear to be moderated by local food availability whereby fledglings within food-rich areas move shorter distances immediately after dispersal than fledglings from food-poor areas (Kenward et al. 1993b, Kennedy and Ward In Press).
Post-fledging Area Size and Habitat
Most fledglings included nest trees within their activity centres and some had more than one activity centre. Fledglings returned to their nest tree activity centres throughout the post-fledging period, i.e., this area wasn’t only used in the early post-fledging period. This pattern was also reported by Kennedy and Ward (1996: goshawks), Wood et al. (1998: bald eagles), and Belthoff and Ritchison (1989: eastern screech-owls [Otus asio]). These behaviours suggest that nest areas are important throughout the post-fledging period for several raptors. Current goshawk management guidelines in BC recommend reduced disturbance levels around goshawk nests until young fledge (BC Ministry of Forests and BC Ministry of Environment, Lands and Parks 1999). However, the vulnerability of young during the early post-fledging period and their continued use of the nest area throughout the post-fledging period, suggest disturbance recommendations should be adhered to until young disperse. Disturbance near nest areas during the post-fledging period may interfere with adult prey deliveries to young and may consequently affect juvenile survival. Wood et al. (1998) suggest goshawk fledglings rely almost entirely on their parents for food during the post-fledging period, although Kenward et al. (1993a) and Kennedy et al. (1994) indicate fledglings provide themselves with some food during this time. We observed fledglings in pursuit of prey and we trapped one fledgling with a live lure. In other investigations on Vancouver Island, we have trapped fledglings in the latter part of the post-fledging period using falling-end Swedish goshawk traps with rock doves (Columba livia) as lures (Kenward et al. 1983). Although goshawks may practice hunting during the late post-fledging period, they probably experience low success rates and receive most food from adults.
Our estimated mean PFA size of 58.6 ha is smaller than PFA sizes reported by Kenward et al. (1993a) and Kennedy et al. (1994), but larger than those reported by Doyle and Mahon (2000) and Mahon and Doyle (2001) within BC’s interior forests. PFA size estimates were variable but they were consistent among 2001 and 2002 with the exception of Klaklakama01 being larger than most other PFA estimates in our study. This estimate was probably larger because 45% of locations were obtained from road telemetry with mean associated error polygons of 104.4-m radius, thus artificially expanding the PFA size. Mahon and Doyle (2001) may have obtained smaller PFA estimates than other studies because they primarily used fledgling vocalizations to aurally locate fledglings, rather than using radio telemetry. Fledglings were silent on 52% of our telemetry sessions and many that were vocal as we approached became silent as we drew near. As well, fledglings that are closer to nests are more likely to be detected aurally and so fledglings located far from nests may go undetected. Often radio telemetry is essential
13

to locate fledglings when they move greater distances from nest trees. Our PFA estimate may be smaller than Kennedy et al. (1994) because they used the adult female core use area to approximate PFA size rather than calculating PFA size directly from fledgling locations. Similarly, PFA estimates provided by Kenward et al. (1993a) were probably inflated because their level of resolution for telemetry locations was only 100 m. It has been well documented in the literature that home range estimates vary depending on the techniques used to collect locational data and on the home range estimation program used to calculate home range size (Lawson and Rodgers 1997, Seaman et al. 1999, Kenward et al. 2001). Therefore, it is difficult to compare PFA size among studies that use different data collection and size estimation techniques.
PFA size likely varies with the habitat composition around nest sites, prey availability within and surrounding nest areas, nest stand size and the degree of habitat contrast at nest stand edges. Kennedy and Ward (In Press) documented food-supplemented goshawk fledglings leaving PFAs on short-term exploratory forays more often than fledglings that weren’t fed. As well, PFAs may be smaller in nest stands with hard edge boundaries such as forest-clearcut edges relative to forest-secondary road or forest-meadow edges. We observed fledglings congregating at the edges of nest stands before dispersing but they rarely crossed roads or clearcuts into other forest stands prior to dispersal. Research examining the habitat use patterns of goshawk fledglings within PFAs is lacking. Daw and DeStephano (2001) characterized goshawk PFAs in Oregon as having dense canopies but also wet and dry openings for foraging habitat. Although valuable, this study used hypothetical 170 ha circular PFAs around active nest sites and did not use radio telemetry data to examine habitat use/availability patterns. Within four of the 12 nest areas in our study, PFAs included relatively dense, younger second-growth forests (30-45 years) near nest trees. Ethier (1999) noted that forests immediately surrounding nest trees (0.04 ha) within the same study area on Vancouver Island had large dbh (79 ± 4.4 cm) and forests within a 300-m radius of the nest tree had significantly smaller dbh (43 ± 2.7 cm). Fledglings spent much of their time feeding on the ground or perched in the lower canopy of trees. Thus, dense forests may provide extra cover for fledglings and protection from predators. As well, we rarely located fledglings upslope of nest trees. We suspect this was an artefact of our steep study area where flying upslope would be more energetically demanding and challenging for fledglings, however similar observations were reported in BC’s interior forests with lower gradients (F. Doyle, Wildlife Dynamics Consulting, personal communication).
Survivorship
Our sample size was too small to conduct a rigorous survivorship analysis. However, we observed a dramatic difference in fledgling survival between 2001 and 2002. Annual variability in survival has been well documented for goshawks (Ward and Kennedy 1996, Dewey and Kennedy 2001) and several other avian species (Newton 1998, Suryan and Irons 2001). Goshawk food provisioning experiments have documented direct and indirect influences of prey availability on goshawk survivorship from hatching to fledging and from fledging to dispersal (Kenward et al. 1993a, Ward and Kennedy 1996, Dewey and Kennedy 2001). The amount of time adults can allocate to hunting and nest defense is moderated by food availability. If food is scarce, adults will spend more time hunting and less time protecting their young; predation pressures may also be
14

greater. However, early nest initiation dates and high nest productivity in 2002 suggest prey were abundant (Minguez et al. 2001, Naef-Daenzer 2001). As well, similar lengths of post-fledging periods among early and late fledgers did not indicate reduced provisioning rates later in the breeding season. Therefore, lower survival may have resulted from higher predation rates caused by abundant predator populations, also benefiting from high food availability. Because we didn’t measure prey or predator abundance, we can only speculate on the relationship between prey availability, predator abundance, and fledgling survivorship during our study.
Conclusions and Management Implications
Our study suggests that the timing and length of goshawk post-fledging periods on Vancouver Island vary among years but generally, fledglings spend 6-7 weeks (July – September) within PFAs. Most fledglings dispersed from PFAs by the end of August. PFAs varied in shape and size, however, most included nest trees within activity centres throughout the post-fledging period and some contained more than one activity centre. Fledglings were near nests in the early post-fledging period and moved farthest from nests 65-80 days post-fledging, corresponding with the completion of their feather growth. Relatively high predation rates in our second year, support earlier conclusions that fledglings are vulnerable to predators during this time when they are poor fliers, relatively vocal and spend much time on or near the ground. Thus, PFA habitat is likely extremely important to the successful dispersal of fledglings, perhaps a better estimate of reproductive output than the number of young fledged.
The mean PFA size for goshawks within our study area on Vancouver Island was 58.6 ± 16.2 ha (95% CI: 22.9, 94.3 ha). We believe our results reflect PFAs within a variety of habitat types including continuous old-growth (>250 years), continuous second-growth (45-70 years), fragmented old-growth and fragmented second-growth forests, and from a diversified topography and aspect. Because the subspecies of goshawk on Vancouver Island are Red-listed provincially and Threatened federally, we recommend a conservative PFA size of 132 ha, which combines the upper 95% CI for the estimate with a 100 m buffer, to reduce edge effects. Within temperate rainforests, the zone of influence from edge effects has been measured to approximately 100 m (Chen et al. 1993, Brand and George 2001). Importantly, this area should also be large enough to enable goshawks to use multiple nest sites within nest areas, which will have different PFAs. The boundaries of PFAs should be designated to include habitat with high canopy closure and without roads or clearcuts, which may reduce habitat suitability for fledglings. Our results should not be interpreted as the only habitat necessary for goshawks to successfully reproduce. Prey availability in habitat near PFAs is also essential in order for adults to rear young (Reynolds et al. 1992, Kennedy and Ward In Press). Future Research Directions
We recommend that habitat use and availability studies occur to examine if fledgling locations are associated with certain habitat characteristics around nest sites. This information may assist with designating suitable PFAs around nests when it is not feasible to collect radio telemetry data.
15

Acknowledgements We thank Don Doyle for his dedication to this project, invaluable
recommendations, and skilled abilities to secure funding. We appreciate Forest Renewal BC and the Forest Investment Account funding provided by Canadian Forest Products, Weyerhaeuser, TimberWest and Western Forest Products, as well as the cooperation of these companies, enabling us to work on their tree farm licenses and private lands. This project would not have been possible without the labour intensive data collection provided by: Jeanine Bond, Nick Davey, Rian Dickson, Ian Jacobs, Marty Krkosek, Paul Levesque, Josh Malt, James Strain, and Aubrey Zeeman. We extend our gratitude to Ian Jacobs for fearlessly climbing goshawk nest trees and to Aubrey Zeeman for tirelessly leading a goshawk crew. We thank Dena Cator for keeping her eyes and ears cued for goshawks while canoeing on Surprise Lake. Pat Kennedy, Joni Ward and Sarah Dewey provided technical advice for this project. Literature Cited Aparicio, J. M. and R. Bonal. 2002. Effects of food supplementation and habitat
selection on timing of lesser kestrel breeding. Ecology 83:873-877. Belthoff, J. R., and G. Ritchison. 1989. Natal dispersal of eastern screech-owls. Condor
91:254-265. Boal. C. W. 1994. A photographic and behavioural guide to aging nestling northern
goshawks. Studies in Avian Biology 16:32-40. Brand, A. L., and T. L. George. 2001. Response of passerine birds to forest edge in
coast redwood forest fragments. Auk 118:678-686. British Columbia Ministry of Forests and British Columbia Ministry of Environment,
Lands and Parks. 1999. Managing Identified Wildlife: Procedures and Measures. Volume I. Victoria, BC.
Chen, J., Franklin, J. F., And T. A. Spies. 1993. Contrasting microclimates among
clearcut, edge and interior old growth Douglas-fir forest. Agricultural and Forest Meteorology 63:219-237.
Clutton-Brock, T. H., ed. 1988. Reproductive success. In: studies of individual variation
in contrasting breeding seasons. University of Chicago Press, Chicago.
Cooper, J. M. and P. Chytyk. 2000. Draft status report on the Queen Charlotte goshawk. Committee on the Status of Endangered Wildlife in Canada, Ottawa, ON.
Crocker-Bedford, D. C. 1998. The value of demographic and habitat studies in determining the status of northern goshawks (Accipiter gentilis atricapillus) with special reference to Crocker-Bedford (1990) and Kennedy (1997). Journal of Raptor Research 32:329-335.
16

Daw, S. K. and S. DeStefano. 2001. Forest characteristics of northern goshawk nest
stands and post-fledging areas in Oregon. Journal of Wildlife Management 65:59-65.
DeStefano, S. 1998. Determining the status of Northern Goshawks in the west: is our
conceptual model correct? Journal of Raptor Research 32:342-348. Dewey, S. R. and P. L. Kennedy. 2001. Effects of supplemental food on parental-care
strategies and juvenile survival of northern goshawks. Auk 118:352-365.
Doyle, F. I. and T. Mahon. 2000. Inventory of the northern goshawk (Accipiter gentilis) in the Kispiox Forest District. Ministry of Environment, Lands and Parks, Smithers B.C.
Environment Canada [online]. 1998. Canadian climate normals 1961–1990: Campbell River airport, British Columbia. <http://www.cmc.ec.gc.ca/climate/normals/British ColumbiaC002.htm> (29 March 2000).
Ethier, T. J. 1999. Breeding ecology and habitat of northern goshawks (Accipiter
gentilis laingi) on Vancouver Island: a hierarchical approach. Thesis. University of Victoria, Victoria, BC.
Green, D. J., and A. Cockburn. 2001. Post-fledging care, philopatry and recruitment in
brown thornbills. Journal of Animal Ecology 70:505-514. Kennedy, P. L. 1997. The northern goshawk (Accipiter gentilis atricapillus): is there
evidence of a population decline? Journal of Raptor Research 31:95-106. Kennedy, P. L. and J. M. Ward. In Press. Effects of experimental food supplementation
on movements of juvenile northern goshawk (Accipiter gentilis atricapillus). Oecologia.
Kennedy, P. L., J. M. Ward, G. A. Rinker, and J. A. Gessaman. 1994. Post-fledging
areas in northern goshawk home ranges. 1994. Studies in Avian Biology 16:75-82. Kenward, R. E., Clarke, R. T., Hodder, K. H., and S. Walls. 2001. Density and linkage
estimators of home range: nearest-neighbor clustering defines multinuclear cores. Ecology 82:1905-1920.
Kenward, R. E., M. Karlbom, and V. Marcstrom. 1983. The price of success in goshawk
trapping. Journal of Raptor Research 17:84-91.
17

Kenward, R. E., V. Marcstrom, and M. Karlbom. 1993a. Post-nestling behaviour in goshawks, Accipiter gentilis: I. The causes of dispersal. Animal Behaviour 46:365-370.
Kenward, R. E., V. Marcstrom, and M. Karlbom. 1993b. Post-nestling behaviour in
goshawks, Accipiter gentilis: II. Sex differences in sociality and nest-switching. Animal Behaviour 46:371-378.
Lawson, E. J. G., and A. R. Rodgers. 1997. Differences in home-range size computed in
commonly used software programs. Wildlife Society Bulletin 25:721-729.
Mahon, T. and F. I. Doyle. 2001. Inventory of the northern goshawk in the Lakes Forest District. Annual Report 2000/2001. Ministry Environment, Lands and Parks, Smithers, BC.
McClaren, E. L., P. L. Kennedy, and S. R. Dewey. 2002. Do some northern goshawk nest areas consistently fledge more young than others? Condor 104: 343-352.
Minguez, E., E. Angulo, and V. Siebering. 2001. Factors influencing length of the post-
fledging period and timing of dispersal in Bonelli’s eagle (Hieraaetus fasciatus) in southwestern Spain. 2001. Journal of Raptor Research 35:228-234.
Ministry of Environment, Lands and Parks 2000. 2000 Red and Blue lists for amphibians,
reptiles, birds and mammals. Victoria, B.C. Naef-Daenzer, B., F. Widmer and M. Nuber. 2001. Differential post-fledging survival of
great and coal tits in relation to their condition and fledging date. Journal of Animal Ecology 70:730-738.
Nams, V. O. 2000. Locate II user’s guide. Pacer, Truro, Nova Scotia, Canada. Newton, I. 1998. Population limitation in birds. Academic Press Ltd., London. O’Toole, L. T., Kennedy, P. L., Knight, R. L., and L. C. McEwan. 1999. Postfledging
behaviour in golden eagles. Wilson Bulletin 111:472-477. Ott, R. L. 1993. An introduction to statistical methods and data analysis, 4th ed. Duxbury
Press, Belmont, CA. Powell, L. A., Calvert, D. J., Barry, I. M., and L. Washburn. 2002. Post-fledging
survival and dispersal of peregrine falcons during a restoration project. Reynolds, R. T. 1983. Management of western coniferous forests habitat for nesting
accipiter hawks. U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Stat. Fort Collins, CO. Gen. Tech. Rep. RM-102.
18

Reynolds, R. T., R. T. Graham, M. H. Reiser, R. L. Bassett, P. L. Kennedy, D. A. Boyce,
G. Goodwin, R. Smith, and E. L. Fisher. 1992. Management recommendations for the Northern Goshawk in the southwestern United States. USDA Forest Service General Technical Report RM-GTR-217.
Rodgers, A. R. and A. P. Carr. 1998. HRE: the home range extension for ArcView
user’s manual. Centre for Northern Forest Ecosystem Research, Ontario Ministry of Natural Resources, Ontario, Canada.
SAS Institute Inc. 1997. SAS/STAT Software: changes and enhancements through
release of 6.12. SAS Institute Inc., Carey, NC. Seaman, D. E., Millspaugh, J. J., Kernohan, B. J., Brundige, G. C., Raedeke, K. J., and R.
A. Gitzen. 1999. Effects of sample size on kernel home range estimates. Journal of Wildlife Management 63:739-747.
Shipman, M [online]. 1997. The northern goshawk (Accipiter gentilis) from egg to the
air. <http://www.blueplanetphoto.com/feature/gost.htm> (January 2002). Squires, J. R. and R. T. Reynolds. 1997. Northern goshawk (Accipiter gentilis). In The
Birds of North America, No. 298 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologists’ Union, Washington, D.C.
Suryan, R. M. and D. B. Irons. 2001. Colony and population dynamics of black-legged
kittiwakes in a heterogeneous environment. Auk 118:636-649. Walls, S. S. and R. E. Kenward. 1998. Movements of radio-tagged buzzards Buteo
buteo in early life. Ibis 140:561-568. Ward, J. M. and P. L. Kennedy. 1996. Effects of supplemental food on size and survival
of juvenile northern goshawks. Auk 113:200-208. Widen, P. 1997. How, and why, is the goshawk (Accipiter gentilis) affected by modern
forest management in Fennoscandia? Journal of Raptor Research 31:107-113. Willey, D. W. and C. van Riper III. 2000. First-year movements by juvenile Mexican
spotted owls in the Canyonlands of Utah. Journal of Raptor Research 34:1-7. Wood, P. B., M. W. Callopy, and C. M. Sekerak. 1998. Postfledging nest dependence
period for bald eagles in Florida. Journal of Wildlife Management 62:333-339.
19

APPENDIX I
PHOTOGRAPHIC AGING KEY
TO NORTHERN GOSHAWKS
ON
VANCOUVER ISLAND
20

15 days (Photo Credit: Erica McClaren) 10 days (Photo Credit: Jeanine Bond)
21
15 days (Photo Credit: Ian Jacobs)
15 day (Photo Credit: Ian Jacobs) 17 days (Photo Credit: Ian Jacobs)
25 days (Photo Credit: Ian Jacobs) 22 days (Photo Credit: Christina Pendergast)

APPENDIX II
NORTHERN GOSHAWK
FLEDGLING LOCATIONS
AND
ASSOCIATED ERROR POLYGONS
22

APPENDIX III
95% ADAPTIVE KERNEL ESTIMATES
FOR
NORTHERN GOSHAWK FLEDGLINGS
35


















