
Non-thermal plasma inhibits human cervical cancer HeLa cells ... · Cervical cancer is the second...
11
1 SCIENTIFIC REPORTS | 6:19720 | DOI: 10.1038/srep19720 www.nature.com/scientificreports Non-thermal plasma inhibits human cervical cancer HeLa cells invasiveness by suppressing the MAPK pathway and decreasing matrix metalloproteinase-9 expression Wei Li 1 , K. N. Yu 3 , Lingzhi Bao 1 , Jie Shen 2 , Cheng Cheng 2 & Wei Han 1,4 Non-thermal plasma (NTP) has been proposed as a novel therapeutic method for anticancer treatment. However, the mechanism underlying its biological effects remains unclear. In this study, we investigated the inhibitory effect of NTP on the invasion of HeLa cells, and explored the possible mechanism. Our results showed that NTP exposure for 20 or 40 s significantly suppressed the migration and invasion of HeLa cells on the basis of matrigel invasion assay and wound healing assay, respectively. Moreover, NTP reduced the activity and protein expression of the matrix metalloproteinase (MMP)-9 enzyme. Western blot analysis indicated that NTP exposure effectively decreased phosphorylation level of both ERK1/2 and JNK, but not p38 MAPK. Furthermore, treatment with MAPK signal pathway inhibitors or NTP all exhibited significant depression of HeLa cells migration and MMP-9 expression. The result showed that NTP synergistically suppressed migration and MMP-9 expression in the presence of ERK1/2 inhibitor and JNK inhibitor, but not p38 MAPK inhibitor. Taken together, these findings suggested that NTP exposure inhibited the migration and invasion of HeLa cells via down-regulating MMP-9 expression in ERK1/2 and JNK signaling pathways dependent manner. These findings provide hints to the potential clinical research and therapy of NTP on cervical cancer metastasis. Non-thermal plasma (NTP), generated at room temperature by ionization of neutral gas molecules, results in a mixture of numerous short-lived but highly active chemical species 1 . ese active chemical species are essential for various biological processes in cells and human tissues. In recent years, NTP have been used in many biomed- ical applications such as wound healing, sterilization, blood coagulation and the ablation of cultured liver cancer cells 2–5 . In addition, newly developed NTP exert anti-tumor effects in various cancer cell types both in vitro and in vivo 6–9 . ese researches present exciting prospects for potential clinical applications of NPT on cancer therapy. Many studies have shown that NTP treatment induce apoptosis in various tumor models 6,10,11 . Further studies also demonstrate that NTP treatment inhibits colorectal and thyroid papillary cancer cells invasion and metasta- sis in vitro 12–14 . However, the underlying molecular signaling pathways for NTP-inhibited migration and invasion in cancer cells still need further investigation. Cervical cancer is the second leading cause of cancer-related mortality in women worldwide, accounting for approximately 270,000 deaths per year 15,16 . Approximately 50% of cervical cancer patients die from metastasis 1 Center of Medical Physics and Technology, Hefei Institutes of Physical Sciences, Chinese Academy of Sciences, 350 Shushanhu Road, Hefei, 230031, Anhui Province, P.R. China. 2 Institute of Plasma Physics, Hefei Institutes of Physical Sciences, Chinese Academy of Sciences, 350 Shushanhu Road, Hefei, 230031, Anhui Province, P.R. China. 3 Department of Physics and Materials Science, City University of Hong Kong, Tat Chee Avenue, Kowloon Tong, Hong Kong. 4 Collaborative Innovation Center of Radiation Medicine of Jiangsu Higher Education Institutions and School for Radiological and Interdisciplinary Sciences (RAD-X), Soochow University, Suzhou, Jiangsu, China. Correspondence and requests for materials should be addressed to W.H. (email: [email protected]) Received: 19 June 2015 Accepted: 27 October 2015 Published: 28 January 2016 OPEN
Transcript of Non-thermal plasma inhibits human cervical cancer HeLa cells ... · Cervical cancer is the second...

1Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
www.nature.com/scientificreports
Non-thermal plasma inhibits human cervical cancer HeLa cells invasiveness by suppressing the MAPK pathway and decreasing matrix metalloproteinase-9 expressionWei Li1, K. N. Yu3, Lingzhi Bao1, Jie Shen2, Cheng Cheng2 & Wei Han1,4
Non-thermal plasma (NTP) has been proposed as a novel therapeutic method for anticancer treatment. However, the mechanism underlying its biological effects remains unclear. In this study, we investigated the inhibitory effect of NTP on the invasion of HeLa cells, and explored the possible mechanism. Our results showed that NTP exposure for 20 or 40 s significantly suppressed the migration and invasion of HeLa cells on the basis of matrigel invasion assay and wound healing assay, respectively. Moreover, NTP reduced the activity and protein expression of the matrix metalloproteinase (MMP)-9 enzyme. Western blot analysis indicated that NTP exposure effectively decreased phosphorylation level of both ERK1/2 and JNK, but not p38 MAPK. Furthermore, treatment with MAPK signal pathway inhibitors or NTP all exhibited significant depression of HeLa cells migration and MMP-9 expression. The result showed that NTP synergistically suppressed migration and MMP-9 expression in the presence of ERK1/2 inhibitor and JNK inhibitor, but not p38 MAPK inhibitor. Taken together, these findings suggested that NTP exposure inhibited the migration and invasion of HeLa cells via down-regulating MMP-9 expression in ERK1/2 and JNK signaling pathways dependent manner. These findings provide hints to the potential clinical research and therapy of NTP on cervical cancer metastasis.
Non-thermal plasma (NTP), generated at room temperature by ionization of neutral gas molecules, results in a mixture of numerous short-lived but highly active chemical species1. These active chemical species are essential for various biological processes in cells and human tissues. In recent years, NTP have been used in many biomed-ical applications such as wound healing, sterilization, blood coagulation and the ablation of cultured liver cancer cells2–5. In addition, newly developed NTP exert anti-tumor effects in various cancer cell types both in vitro and in vivo6–9. These researches present exciting prospects for potential clinical applications of NPT on cancer therapy. Many studies have shown that NTP treatment induce apoptosis in various tumor models6,10,11. Further studies also demonstrate that NTP treatment inhibits colorectal and thyroid papillary cancer cells invasion and metasta-sis in vitro12–14. However, the underlying molecular signaling pathways for NTP-inhibited migration and invasion in cancer cells still need further investigation.
Cervical cancer is the second leading cause of cancer-related mortality in women worldwide, accounting for approximately 270,000 deaths per year15,16. Approximately 50% of cervical cancer patients die from metastasis
1Center of Medical Physics and Technology, Hefei Institutes of Physical Sciences, Chinese Academy of Sciences, 350 Shushanhu Road, Hefei, 230031, Anhui Province, P.R. China. 2Institute of Plasma Physics, Hefei Institutes of Physical Sciences, Chinese Academy of Sciences, 350 Shushanhu Road, Hefei, 230031, Anhui Province, P.R. China. 3Department of Physics and Materials Science, City University of Hong Kong, Tat Chee Avenue, Kowloon Tong, Hong Kong. 4Collaborative Innovation Center of Radiation Medicine of Jiangsu Higher Education Institutions and School for Radiological and Interdisciplinary Sciences (RAD-X), Soochow University, Suzhou, Jiangsu, China. Correspondence and requests for materials should be addressed to W.H. (email: [email protected])
Received: 19 June 2015
Accepted: 27 October 2015
Published: 28 January 2016
OPEN

www.nature.com/scientificreports/
2Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
after treatment with the current main therapeutic methods15. Cancer metastasis, spreading of cancer cells from the primary neoplasm to distant sites, forms the secondary tumor17. Metastasis occurs via a complex series of events, including invasion of cells from a primary tumor into the circulation system, immigration of these cells to distant organs, adhesion to endothelial cells, and infiltration into tissue17,18. In this process, degradation of the extracellular matrix (ECM) is mainly performed by matrix metalloproteinases (MMPs)19. In the MMP family, MMP-2 and MMP-9 are crucial for the invasion and metastasis of many types of cancer cells, and so several inhibitors of MMPs have been tested in clinical trials for prevention of tumor invasion and metastasis20–22. The expression and activity of MMP-9 and MMP-2 are regulated by various growth factors or mitogen-activated pro-tein kinase (MAPK)23,24. Many studies have demonstrated that MAPKs, including extracellular signal-regulated kinase (ERK1/2), c-Jun N-terminal kinase (JNK), and p38 MAPK, play important regulatory roles in cell invasion and metastasis24. As such, inhibition of MAPKs pathway is considered potential targets for preventing cancer metastasis.
In this study, we explored the inhibitory effects and the possible underlying molecular mechanisms of NTP on the migration and invasion of human cervical cancer HeLa cells. Our results demonstrated that NTP exposure inhibited the migration and invasion of human cervical cancer HeLa cells via inhibiting MAPK signaling path-way, which led to down-regulation of MMP-9 activity and expression. These findings provided a novel mechanis-tic insight into the potential of NTP on the suppression of cervical cancer invasion and metastasis.
ResultsNTP inhibited proliferation of HeLa cells. In this study, a non-thermal plasma (NTP) generating system was developed in our lab as previously described25. Helium gas was injected into the chamber through the gas inlet with a fixed flow rate of 80 L/h. In order to expel as much air as possible from the reactor chamber, helium was injected at 5 min before the experiment. The non-thermal plasma was generated by a voltage of 12 kV (peak to peak) at a frequency of 24 kHz.
Previous reports showed that NTP induced cell death in a exposure time dependent manner26. To determine the effect of NTP exposure time on the viability of Hela cells, the CCK-8 assay was used to measure cell viabil-ity. A gas-only treatment (helium) was used as a reference to exclude the gas effects of NTP. The results of the CCK-8 assay are shown in Fig. 1. The results showed that after 24 or 48 h incubation, NTP exposure from 10 to 40 s induced no distinct cytotoxic effects on HeLa cells (p > 0.05; Fig. 1). With the increase of exposure time (60 or 80 s), the cell viability decreased when compared with the control group (i.e., cells without NTP treatment) (p < 0.05; Fig. 1). However, for longer-time incubation (72 h), NPT exposures for 40 s significantly decreased the cell viability when compared with the control group (p < 0.05; Fig. 1). Therefore, NTP exposure durations of 10, 20 or 40 s were used in the subsequent experiments.
In addition, after 24 h incubation, NTP exposures from 10 to 40 s did not affect DNA damages (Fig. 2a). Flow cytometry analysis of apoptosis and mitochondrial transmembrane potential (Δ ψ m) showed that NTP expo-sures from 10 to 40 s had no significantly effects on cell apoptosis and mitochondrial transmembrane potential (p > 0.05; Fig. 2b,c). Cytoskeleton staining also showed that NTP exposures from 10 to 40 s did not change cel-lular morphology or cytoskeletal architecture (Fig. 2d). Western blot analysis showed that NTP treatment had no effects on the expressions of Bax, Bcl-2, Caspase-3, Bid, cytochrome c and γ -H2AX (Fig. 3). Taken together, after 24 h incubation, NTP exposure durations of 10, 20 or 40 s did not affect the viability of HeLa cells or cause physical damages to the cells.
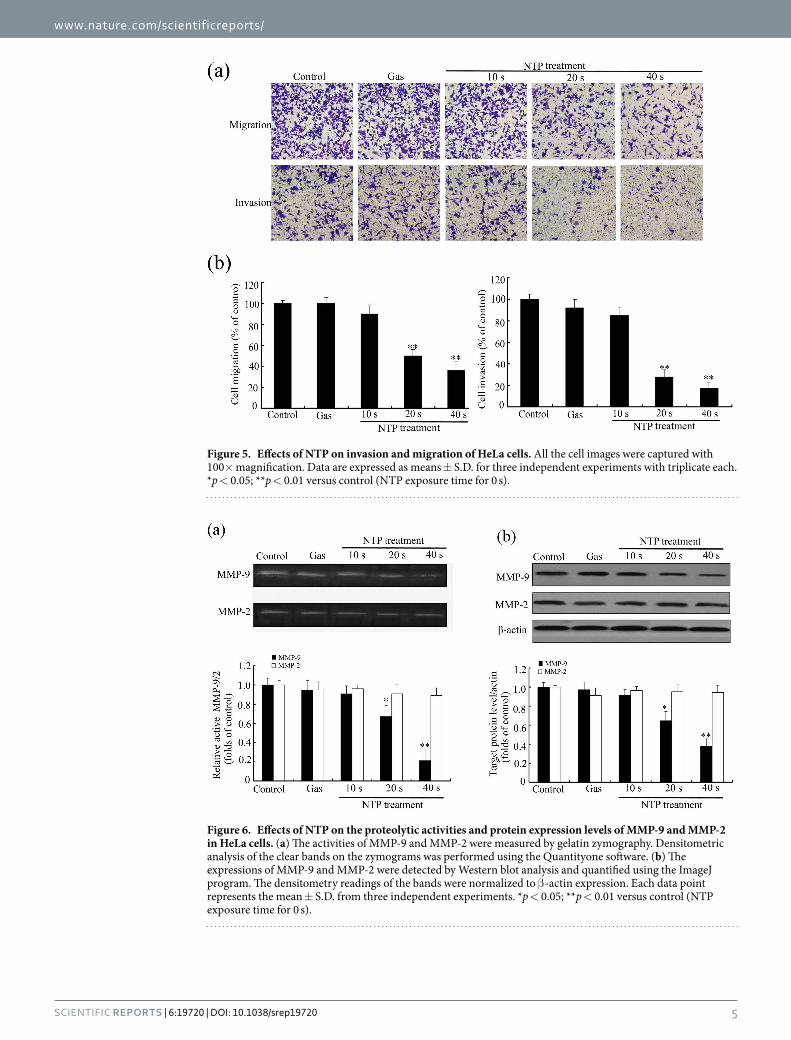
NTP inhibited migration and invasion of HeLa cells. The capabilities of migration and invasion of tumor cells are important characteristics of metastasis18. We first performed the wound-healing assay to measure the effect of NTP on HeLa cell motility. As shown in Fig. 4, NPT exposures significantly decreased the cell motility in a dose-dependent manner. NPT exposures for 20 or 40 s inhibited cell migration by 63% or 45%, respectively, when compared with the control group.
Transwell migration assay and Matrigel invasion assay were also used to examine the effects of NTP on migra-tory and invasive capacities of HeLa cells. As shown in Fig. 4, NPT exposures significantly inhibited the migration of HeLa cells in an exposure time-dependent manner. The percentage of transwell migrated cells decreased to
Figure 1. Effects of NTP on the growth of HeLa cells. HeLa cells were exposed to NTP for 10, 20 or 40 s. Data are expressed as means ± S.D. for three independent experiments with triplicate each. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).

www.nature.com/scientificreports/
3Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
Figure 2. Effects of NTP on DNA damage, apoptosis, mitochondrial transmembrane potential (Δψm) and cytoskeleton in HeLa cells. (a) Immunocytochemistry of γ -H2AX in cells and number of γ -H2AX foci per cell at 24 h after NTP treatment. DAPI was used to stain the cell nuclei. Scale bar = 20 μ m. (b) Annexin V-FITC/PI staining assay was used to determine the percentage of apoptotic cells in NTP-treated Hela cells. (c) The Δ ψ m was analyzed using a JC-1 Mitochondrial Potential Detection. (d) Immunofluorescence assays using FITC-conjugated phalloidin were performed to visualize the cytoskeleton (F-actin), and DAPI was used to stain the cell nuclei. Each data point represents the mean ± S.D. from three independent experiments. Scale bar = 20 μ m. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).
Figure 3. Effects of NTP on the expressions of Bax, Bcl-2, Caspase-3, Bid, cytochrome c and γ-H2AX in HeLa cells. The cells were exposed to NTP for 10, 20 or 40 s. After 24 h incubation, total protein was extracted and subjected to SDS-PAGE, followed by western blot analysis. The internal control was β -actin.

www.nature.com/scientificreports/
4Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
54% or 31% with NTP exposures of 20 or 40 s, respectively (see Fig. 5). Similarly, NPT exposures significantly decreased the number of invading cells through the matrigel when compared with control group. NTP exposures for 20 or 40 s inhibited the number of cells from invading to the lower chamber by 34% or 23%, respectively. Taken together, these results suggested that NTP could inhibit the migration and invasion of HeLa cells.
NTP decreased proteolytic activity and expression of MMP-9 in HeLa cells. Previous studies demonstrated that the expressions of MMP-2 and -9 were high in various malignant tumors and were closely related with tumor migration27,28. Therefore, in order to further test whether NTP treatment suppressed HeLa cell invasion via affecting the expression of matrix metalloproteinase, gelatin zymography assay was performed to measure MMP-9 and MMP-2 activities. As shown in Fig. 6a, NTP treatment significantly inhibited gelatinolytic activity of MMP-9 in an exposure-time-dependent manner, but the activity of MMP-2 did not change. The results of western blot (Fig. 6b) showed that NTP treatment also significantly decreased the protein expression level of MMP-9, but not MMP-2, in a time-dependent manner. These results indicated that NTP inhibited the proteolytic activity and protein expression of MMP-9.
NTP inhibited phosphorylation of ERK1/2 and JNK in HeLa cells. MAPKs, including ERK1/2, p38 and JNK, regulate cell invasion as well as migration in various cell types23,29,30. To elucidate the mechanisms responsible for the inhibitory effect of NTP on metastasis, phosphorylation of MAPKs in HeLa cells was meas-ured by western blot. The results (Fig. 7) showed that NTP exposures significantly decreased the phosphorylation of ERK1/2 and JNK in an exposure-time-dependent manner but not the total level of these two proteins, whereas no distinct change was observed in the phosphorylation level of p38. These results suggested that NTP inhibited the metastatic activity of HeLa cells via inactivating JNK and ERK1/2 signaling pathways.
NTP inhibits HeLa cells invasion by suppressing phosphorylation of JNK and ERK1/2. We pro-ceed to further confirm whether the NTP-induced changes of MAPK signaling pathway were directly related to NTP-dependent HeLa cell invasion. The cells were pretreated with JNK inhibitors (SP600125), ERK1/2 inhibi-tors (SCH772984) or p38 MAPK inhibitors (SB203580) for 2 h and then exposed to NTP for 10, 20 or 40 s. As shown in Fig. 8, treatment with NTP alone or MAPK inhibitors significantly suppressed HeLa cell migration, as
Figure 4. Effects of NTP on migration of HeLa cells. All cell images were captured with 100× magnification. The relative migration of each sample was calculated from the difference in the area of unclosed wound region measured as compared to those of the untreated control (NTP exposure time for 0 s). Data are expressed as means ± S.D. for three independent experiments with triplicate each. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).
www.nature.com/scientificreports/
5Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
Figure 5. Effects of NTP on invasion and migration of HeLa cells. All the cell images were captured with 100× magnification. Data are expressed as means ± S.D. for three independent experiments with triplicate each. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).
Figure 6. Effects of NTP on the proteolytic activities and protein expression levels of MMP-9 and MMP-2 in HeLa cells. (a) The activities of MMP-9 and MMP-2 were measured by gelatin zymography. Densitometric analysis of the clear bands on the zymograms was performed using the Quantityone software. (b) The expressions of MMP-9 and MMP-2 were detected by Western blot analysis and quantified using the ImageJ program. The densitometry readings of the bands were normalized to β -actin expression. Each data point represents the mean ± S.D. from three independent experiments. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).

www.nature.com/scientificreports/
6Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
compared with that of the control. Furthermore, NTP synergistically suppressed migration after pre-treating with ERK1/2 and JNK inhibitors, but not so with the p38 MAPK inhibitor. The result illustrated that ERK1/2and JNK signaling pathways played a key role in HeLa cell migration.
Expression of MMP-9 was partly dependent on ERK1/2 and JNK activities in HeLa cells. To determine the role of MAPK signaling pathway in MMP-9 expression of HeLa cells, we evaluated MMP-9 expres-sion in HeLa cells using MAPK inhibitor supplementation. Western blot results (Fig. 9) showed that inhibi-tors of p38MAPK, ERK1/2 and JNK all reduced MMP-9 expression. In addition, NTP could synergistically down-regulate MMP-9 expression with the ERK1/2 inhibitor and the JNK inhibitor, but not with the p38MAPK inhibitor. The results suggested that inhibition of MMP-9 expression by NTP could be mediated through inacti-vation of ERK1/2 and JNK signaling pathways in HeLa cells.
DiscussionNTP has been extensively studied for its biomedical applications and has recently emerged as a potential novel tool in cancer treatment. Previous studies focused mainly on the apoptotic cancer cell death mediated by NTP as a potential cancer therapy strategy5. However, the mechanisms involved in the NTP-induced inhibition of cell migration and invasion have not been elucidated. In the present study, we investigated the inhibitory effects of NTP on the migration and invasion of human cervical cancer HeLa cells, and the underlying mechanisms involved in this process. Our results provided evidence that NTP exerted an anti-tumor effect via inhibiting can-cer cell migration and invasion.
Invasion and migration are important features of malignant tumors18. Tumor metastasis remains the main cause of treatment failure and death in a variety of tumors. To identify the anti-metastatic effects of NTP, we
Figure 7. Effects of NTP on the MAPK pathway in HeLa cells. The cells were exposed to NTP for 10, 20 or 40 s. After 24 h incubation, total protein was extracted and subjected to SDS-PAGE, followed by Western blot analysis. β -actin was used as an internal control. The expressions of ERK1/2 and p-ERK1/2 (a), JNK and p-JNK (b), p38 MAPK and p-p38 MAPK (c) proteins were determined using Western blot analysis and quantified using the ImageJ program. The upper panel shows a representative Western blot result. The lower panel illustrated the fold difference of integrated absorbance of protein after normalization with β -actin. Each data point represents the mean ± S.D. from three independent experiments. *p < 0.05; **p < 0.01 versus control (NTP exposure time for 0 s).

www.nature.com/scientificreports/
7Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
compared the migratory and invasive activity of HeLa cells using the scratch wound healing, transwell migration assay, and matrigel invasion assay. Our results showed that NTP significantly inhibited the migration and invasion of HeLa cells. These results were consistent with previous findings that NTP treatment decreased cell migration and invasion in several cell types.
MMPs, a family of zinc-binding proteases, can degrade the extracellular matrix (ECM) and basement mem-brane for cancer cells to enable migration and invasion27,28. Among MMPs, expressions of MMP-2 and MMP-9 are high in various malignant tumors and are closely related to tumor migration28. Therefore, early suppression of expression and/or proteolytic activity of MMP-9/2 can be the target for preventing cancer metastasis. To further explore the specific effects of NTP on MMP-2 and MMP-9, Gelatin zymography assay was used to detect the activities of MMP-2 and MMP-9 in HeLa cells. The results showed that NTP inhibited activities of MMP-9 in a time-dependent manner. The results of western bolt further revealed that the reduction of MMP-9 activity was a result of down-regulation at the protein level. These results suggested that NTP could inhibit or delay cancer invasion and migration in HeLa cells via reducing MMP-9 expression and activity.
Previous reports from various laboratories demonstrated that MMP-9 expression was critically mediated by the MAPK pathway in various cell lines23,31. The activation of ERK1/2 and/or p38 MAPK played important roles in controlling MMP-9 gene expression through stimulating transcription factors (AP-1 and NF-κ B)24,30. The JNK signaling was essential for a variety of biological processes such as morphogenesis, cell proliferation, migration, invasion and cell death32,33. Several studies indicated that suppression of MAPKs had the potential to prevent invasion and metastasis for various tumors23,24. Our results showed that NTP exposures inhibited phosphoryla-tion of ERK1/2 and JNK in a time-dependent manner. These results suggested that NTP inhibited the metastatic activity of HeLa cells via inactivating the MAPK pathway. However, how NTP inhibited these intracellular signal pathways to initiate the response remained unclear to date. Previous studies also demonstrated that reactive oxygen species (ROS) generated by NTP had selective damages on cancer cells through induction of apoptosis
Figure 8. Effects of NTP and MAPK inhibitors on migration of HeLa cells. The cells were pretreated with or without MAPK inhibitors (ERK1/2 inhibitor SCH772984, p38 MAPK inhibitor SB203580 and JNK inhibitor SP600125) for 2 h. The cells were then exposed to NTP for 20 s. After 24 h incubation, the migration of HeLa cells with different treatments was tested by cell migration assay. The data represent the mean ± S.D of three independent experiments. *p < 0.05, **p < 0.01.

www.nature.com/scientificreports/
8Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
or loss of cell adhesion7,34,35. Extracellular ROS could activate cell surface receptors to regulate cell migration and invasion36,37. Various chemical or physical stimulators could regulate MMP-9 expression via increasing the ROS level29,36. Based on the above discussion, it was postulated that ROS generated by NTP could be the most effective major factors to initiate the suppressive effect of NTP on cell migration and invasion. Nevertheless, further studies to investigate the signaling initiated by NTP would be needed for a better understanding on the mechanism of NTP inhibition of migration and invasion.
In conclusion, our results demonstrated that NTP exposure effectively inhibited the migration and inva-sion of human cervical cancer HeLa cells via inhibiting the phosphorylation of JNK and ERK1/2, which led to down-regulation of MMP-9 activity and expression. It is anticipated that these results will provide a mechanistic insight into the potential effects of NTP on the suppression of cervical cancer invasion and metastasis.
MethodsCell culture and reagents. Human cervical cancer HeLa cells (ATCC, CCL-2) were obtained from American Type Culture Collection and cultured in DMEM supplied with 10% FCS, 100 U/mL of penicillin and 100 μ g/mL of streptomycin. All cultures were maintained in a 37 °C and 5% CO2 humidified atmosphere. Matrigel was purchased from BD Biosciences (Bedford, MA). The primary antibodies for ERK1/2, p-ERK1/2, JNK, p-JNK, p38, p-p38, MMP-2, MMP-9, Bax, Bcl-2, Caspase-3, Bid, cytochrome c and β -actin were purchased from Cell Signaling Technology Inc. (Beverly, MA, USA). Horseradish peroxidase (HRP)-conjugated secondary antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). FITC-conjugated phalloidin was pur-chased from Sigma (Sigma -Aldrich, St. Louis, MO). γ -H2AX antibody was purchased from Abcam (Abcam, Cambridge, MA, USA). BCA Protein Assay Kit and enhanced chemiluminescence (ECL) reagents were pur-chased from Pierce (Rockford, IL, USA). Cell counting kits-8 (CCK-8), Annexin V-FITC/propidium iodine (PI) apoptosis detection kit, JC-1 Mitochondrial Potential Detection Kit, RIPA lysis buffer and phenylmethyl sulfon-ylfluoride (PMSF) were purchased from Beyotime Inst. Biotech (Beijing, China). DMEM medium was purchased from Invitrogen Life Technologies (Carlsbad, CA, USA). Fetal calf serum (FCS) was purchased from Thermo Scientific (Rockford, IL, USA). Polyvinylidene difluouride (PVDF) membrane were purchased from Millipore Corp (Bedford, MA, USA). MAPK inhibitors (ERK1/2 inhibitor SCH772984, p38 MAPK inhibitor SB203580 and JNK inhibitor SP600125) were purchased from Selleck Chemicals (Houston, TX, USA). All other chemicals and reagents were of the highest quality and obtained from standard commercial sources.
DBD plasma device. The atmospheric pressure dielectric barrier discharge (DBD) plasma was schematically illustrated previously25. The four DBD plasma reactors were sealed in a hollow plexiglass cylinder as a reactor chamber with two air orifices. One was used to inject the working gas, while the other was used to expel air from the hollow plexiglass cylinder. The high voltage electrode was a 32 mm diameter copper cylinder, which was cov-ered by 1 mm thick quartz glass as an insulating dielectric barrier. The ground electrode was a 37 mm diameter copper cylinder. The discharge gap between the bottom of the quartz and the treated sample surface was fixed at 5 mm. The alternating current power supply was a commercial transformer capable of continuous and tunable output voltages and frequencies. The applied voltage and discharge current of the DBD plasma were monitored using a Tektronix MSO 5104 digital oscilloscope equipped with a high voltage probe (Tektronix P6015A) and
Figure 9. Effects of NTP and MAPK inhibitors on MMP-9 expression of HeLa cells. The cells were pretreated with or without MAPK inhibitors (ERK1/2 inhibitor SCH772984, p38 MAPK inhibitor SB203580 and JNK inhibitor SP600125) for 2 h. The cells were then exposed to NTP for 20 s. After 24 h incubation, the protein levels of MMP-9 were determined by Western blot. The data represent the means ± S.D. of three independent experiments. The lower panel illustrated the fold difference of integrated absorbance of protein after normalization with β -actin. *p < 0.05; **p < 0.01.

www.nature.com/scientificreports/
9Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
current probe (Tektronix P6021). Helium gas was injected into the chamber through the gas inlet with a fixed flow rate of 80 L/h. In order to expel as much air as possible from the reactor chamber, helium was injected at 5 min before the experiment. The non-thermal plasma was generated by a voltage of 12 kV (peak to peak) at a frequency of 24 kHz. The discharge power density was measured to be about 0.9 W/cm2.
Non-thermal plasma treatment. For plasma exposures, 3.5 × 105 cells were seeded into 30 mm diameter Petri dishes. The medium was replaced with 2 ml fresh complete culture medium just before exposures. The dis-tance between the nozzle and the surface of culture medium was fixed at 10 mm. The cells were then exposed to NTP for 10, 20 or 40 s. The control cells (NTP exposure for 0 s) were subjected to identical procedures except the plasma treatment. A gas-only treatment (helium) was used to confirm the presence or absence of effects from the gas alone. At the chosen time points, the cells were harvested for further experiments.
Cell viability assay. The cell viability was measured by using the CCK-8 assay (Beyotime Biotech, China) according to the manufacture’s recommendations. Briefly, HeLa cells (3 × 105 per well) were seeded into 30 mm dishes with 1.8 mL complete culture medium and exposed to NTP for 10, 20 or 40 s. After 24, 48 or 72 h incuba-tion, 200 μ L of CCK-8 solution was added into the cell culture medium and incubated for a further 4 h at 37 °C. After plate vibrating for 20 s, the supernatants (100 μ L) were transported into a 96-well plate. Absorbance at 450 nm was read with a microplate reader (ELx800, BioTek Instruments Inc., USA).
Immunocytochemistry. HeLa cells (3 × 105 per well) were seeded into 30 mm dishes with 1.8 mL complete culture medium and exposed to NTP for 10, 20 or 40 s. After 24 h incubation, the cells were fixed with 4% for-maldehyde for 30 min. and then permeabilized with 0.1% Triton X-100. After blocking in bovine serum albumin (BSA) in 5% PBS for 45 min, the dishes were incubated with a polyclonal rabbit anti-γ -H2AX antibody (1:50) for 2 h. The dishes were then washed again in PBS and incubated with FITC-labeled goat anti-rabbit antibody (1:200) for 60 min. After rinsing in PBS, DAPI was added to dishes for 10 min to counter stain nuclei. The dishes were washed with PBS and then visualized using a fluorescence microscope (Leica DMI 40008, Germany).
In addition, cytoskeleton staining was performed using FITC-conjugated phalloidin according to the manu-facturer’s instructions.
Mitochondrial transmembrane potential (Δψm) assessment. The mitochondrial transmembrane potential Δ ψ m was analyzed using a JC-1 Mitochondrial Potential Detection Kit (Biotium Inc., Hayward, CA, US). Briefly, HeLa cells (3 × 105 per well) were seeded into 30 mm dishes with 1.8 mL complete culture medium and exposed to NTP for 10, 20 or 40 s. After 24 h incubation, cells were harvested and stained by JC-1 in PBS for 30 min at room temperature in the dark, followed by flow cytometric analysis.
Flow cytometry analysis of apoptosis. Annexin V-Fluorescein isothiocyanate/PI apoptosis detection kit (KeyGen Biotech Co., Ltd., Nanjing, China) was used for apoptosis detection. Briefly, HeLa cells (3 × 105 per well) were seeded into 30 mm dishes with 1.8 mL complete culture medium and exposed to NTP for 10, 20 or 40 s. After 24 h incubation, cells were collected and washed twice with PBS, and then suspended with 500 μ L of 1× Annexin V binding buffer containing 5 μ L of Annexin V-FITC and 5 μ L of PI. After incubation in the dark for 10 min at room temperature, the cells were analyzed using flow cytometry (Beckman Coulter, Fullerton, CA).
Wound-healing assay. HeLa cells were seeded into 30-mm dishes and grown to 90% confluence. A linear wound was created in the confluent monolayer by using a 200 μ L micropipette tip, and followed by washing with PBS to remove the cell debris. The cells with 2 ml culture medium were then exposed to NTP for 10, 20 or 40 s. After 24 h incubation, cell migration was observed under a microscope (Olympus IX71, Tokyo, Japan) at a mag-nification of 100× . The cells migrated across the black lines were counted in five randomly chosen fields of view in each sample. The inhibition effect was expressed after normalization (taking the amount for the control group as 100%).
Cell migration and invasion assays. The transwell insert for 24-well plate (8 μ m-pore size, Corning, New York, USA) was used to measure the migratory and invasive ability of cells. For transwell migration assays, HeLa cells (2.5 × 104) were seeded into the transwell insert (upper chamber) with serum-free medium after NTP expo-sure for 10, 20 or 40 s, and the culture medium with 10% FCS was added in the lower chamber (the space between the well bottom and the insert) for chemo-attractant. The invasion assays were performed similarly except that the upper chambers of 24-well cell culture inserts were coated with 200 mg/ml of Matrigel (BD Biosciences, Bedford, MA). After 24 h incubation, the insert was taken out and the cells were fixed with 4% paraformaldehyde for 30 min and stained with 1% crystal violet. The non-invading cells were carefully removed from the upper sur-face of the insert membrane with cotton wool. The number of cells migrated to the lower surface of the membrane was determined by counting 10 randomly selected fields of view under the microscope. Data were expressed as the percentage of invasion of treated cells as compared to the control cells.
Gelatin zymography. Gelatin zymography was performed with the MMP Zymography assay kit (Applygen Technologies Inc.) to measure the activities of MMP-9 and MMP-2 according to the manufacturer’s instructions. Briefly, HeLa cells were washed with PBS, and then serum-free culture medium was added before the NTP expo-sure for 10, 20 or 40 s. After 24 h incubation, the supernatants were electrophoresed on 8% SDS-PAGE gels con-taining 0.1% gelatin. The gels were washed twice with washing buffer (2.5% Triton X-100) for 1 h to remove SDS, and were then incubated in developing buffer (50 mM Tris-Cl, pH 7.6, 200 mM NaCl, 10 mM CaCl2) at 37 °C for 12 h for digestion of gelatin. The gelatinolytic activity of MMP-9 and MMP-2 was visualized by staining the gels with Coomassie blue for 30 min and destaining with 45% methanol/10% acetic acid until clear bands suggestive of

www.nature.com/scientificreports/
1 0Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
gelatin digestion appeared. The subsequent zymographic band intensities were quantified using the QuantityOne software (Bio-Rad, Hercules, CA).
Western blot analysis. HeLa cells (3 × 105 per well) were seeded into 30-mm dishes with 2 mL complete culture medium and then exposed to NTP for 0, 10, 20 and 40 s. After 24 h incubation, the total protein was extracted from NTP-treated cells as described previously38. The protein concentration was determined with the BCA Protein Assay Kit (Pierce, Rockford, IL, USA). Equivalent amounts of protein samples were uploaded and separated by 12% SDS-PAGE, and then electro-transferred to polyvinylidene difluouride (PVDF) membrane (Millipore Corp, Atlanta, GA, US). The membranes were blocked in 5% non-fat dry milk at room temperature for 1 h, and then incubated with primary antibodies overnight at 4 °C, followed by HRP-conjugated secondary antibodies at room temperature for 1 h. The signals were developed using the ECL (enhanced chemilumines-cence) reagents (Pierce, Rockford, IL, USA) and exposed to an X-ray film (Kodak, Tokyo, Japan). Each band was quantified by densitometric scanning using the free software ImageJ. The densitometry readings of the bands were normalized to β -actin expression.
Statistical analysis. All data were expressed as mean ± standard deviation (S.D). The difference between the treated samples and the untreated controls was analyzed by Student’s t-test and those differences corresponding to p < 0.05 were considered significant.
References1. Moisan, M. et al. Plasma sterilization. Methods mechanisms. Pure Appl Chem 74, 349–358 (2002).2. Isbary, G. et al. Successful and safe use of 2min cold atmospheric argon plasma in chronic wounds: results of a randomized
controlled trial. Brit J Dermatol 167, 404–410 (2012).3. Shashurin, A., Keidar, M., Bronnikov, S., Jurjus, R. A. & Stepp, M. A. Living tissue under treatment of cold plasma atmospheric jet.
Appl Phys Lett 93 (2008).4. Fridman, G. et al. Blood coagulation and living tissue sterilization by floating-electrode dielectric barrier discharge in air. Plasma
Chenm Plasma P 26, 425–442 (2006).5. Neha, K. et al. Non-thermal plasma with 2-deoxy-D-glucose synergistically induces cell death by targeting glycolysis in blood cancer
cells. Sci Rep 5, 8726 (2015).6. Kang, S. U. et al. Nonthermal plasma induces head and neck cancer cell death: the potential involvement of mitogen-activated
protein kinase-dependent mitochondrial reactive oxygen species. Cell Death and Dis 5, e1056 (2014).7. Neha, K. et al. Responses of Solid Tumor Cells in DMEM to Reactive Oxygen Species Generated by Non-Thermal Plasma and
Chemically Induced ROS Systems. Sci Rep 5, 8587 (2015).8. Vandamme, M. et al. Antitumor Effect of Plasma Treatment on U87 Glioma Xenografts: Preliminary Results. Plasma Process Polyms
7, 264–273 (2010).9. Donadelli, M. et al. Gemcitabine/cannabinoid combination triggers autophagy in pancreatic cancer cells through a ROS-mediated
mechanism. Cell Death and Dis 2, e152 (2011).10. Keidar, M. et al. Cold atmospheric plasma in cancer therapy a). Phys.Plasmas 20, 057101 (2013).11. Keidar, M. et al. Cold plasma selectivity and the possibility of a paradigm shift in cancer therapy. Br. J. Cancer 105, 1295–1301
(2011).12. Kim, C.-H. et al. Induction of cell growth arrest by atmospheric non-thermal plasma in colorectal cancer cells. J Biotechnol 150,
530–538 (2010).13. Kim, J. Y., Wei, Y., Li, J. & Kim, S. O. 15-mum-sized single-cellular-level and cell-manipulatable microplasma jet in cancer therapies.
Biosen bioelectron 26, 555–559 (2010).14. Cao, J. et al. Non-Thermal Atmospheric Pressure Plasma Inhibits Thyroid Papillary Cancer Cell Invasion via Cytoskeletal
Modulation, Altered MMP-2/-9/uPA Activity. PLoS ONE 9, e92198 (2014).15. Wright, T. C. & Kuhn, L. Alternative approaches to cervical cancer screening for developing countries. Best Pract Res 26, 197–208
(2012).16. Ferlay, J. et al. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 127, 2893–2917 (2010).17. Friedl, P. & Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat Rev Cancer 3, 362–374 (2003).18. Entschladen, F., Drell, T. L., Lang, K., Joseph, J. & Zaenker, K. S. Tumour-cell migration, invasion, and metastasis: navigation by
neurotransmitters. Lancet Oncol 5, 254–258 (2004).19. Johnson, J. L. et al. Regulation of Matrix Metalloproteinase Genes by E2F Transcription Factors: Rb-Raf-1 Interaction as a Novel
Target for Metastatic Disease (vol 72, pg 516, 2012). Cancer Res 72, 1317–1317 (2012).20. Weng, C.-J. & Yen, G.-C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids,
monophenol, polyphenol, and their derivatives. Cancer Treatment Reviews 38, 76–87 (2012).21. Sun, W. H. et al. Expression of cyclooxygenase-2 and matrix metalloproteinase-9 in gastric carcinoma and its correlation with
angiogenesis. Jpn J Clin Oncol 35, 707–713 (2005).22. Noh, S. et al. MMP-2as a Putative Biomarker for Carcinomatosis in Gastric Cancer. Hepato-Gastroenterol 58, 2015–2019 (2011).23. Chun, J. & Kim, Y. S. Platycodin D inhibits migration, invasion, and growth of MDA-MB-231 human breast cancer cells via
suppression of EGFR-mediated Akt and MAPK pathways. Chem-biol interact 205, 212–221 (2013).24. Yang, Y.-T., Weng, C.-J., Ho, C.-T. & Yen, G.-C. Resveratrol analog-3,5,4′ -trimethoxy-trans-stilbene inhibits invasion of human lung
adenocarcinoma cells by suppressing the MAPK pathway and decreasing matrix metalloproteinase-2 expression. Mol Nutr Food Res 53, 407–416 (2009).
25. Zhang, H. et al. Effects and Mechanism of Atmospheric-Pressure Dielectric Barrier Discharge Cold Plasma on Lactate Dehydrogenase (LDH) Enzyme. Sci. Rep 5, 10031 (2015).
26. Li, W. et al. Swainsonine Induces Caprine Luteal Cells Apoptosis via Mitochondrial-Mediated Caspase-Dependent Pathway. J Biochem Mol Toxicol 28, 456–464 (2014).
27. Kim, W., Woo, K.-C., Kim, G.-C. & Kim, K.-T. Nonthermal-plasma-mediated animal cell death. Journal of Physics D: Applied Physics 44, 13001 (2011).
28. Nelson, A. R., Fingleton, B., Rothenberg, M. L. & Matrisian, L. M. Matrix metalloproteinases: Biologic activity and clinical implications. J Clin Oncol 18, 1135–1149 (2000).
29. Jinga, D. C. et al. MMP-9 and MMP-2 gelatinases and TIMP-1 and TIMP-2 inhibitors in breast cancer: correlations with prognostic factors. J Cell Mol Med 10, 499–510 (2006).
30. Chiu, W.-T. et al. Contribution of reactive oxygen species to migration/invasion of human glioblastoma cells U87 via ERK-dependent COX-2/PGE2 activation. Neurobiology of Dis 37, 118–129 (2010).

www.nature.com/scientificreports/
1 1Scientific RepoRts | 6:19720 | DOI: 10.1038/srep19720
31. Langlois, B. et al. LRP-1 Promotes Cancer Cell Invasion by Supporting ERK and Inhibiting JNK Signaling Pathways. PLoS ONE 5 (2010).
32. Chun, J. et al. Platycodin D inhibits migration, invasion, and growth of MDA-MB-231 human breast cancer cells via suppression of EGFR-mediated Akt and MAPK pathways. Chem-biol interact 205, 212–221 (2013).
33. Igaki, T., Pagliarini, R. A. & Xu, T. Loss of cell polarity drives tumor growth and invasion through JNK activation in Drosophila. Curr Biol 16, 1139–1146 (2006).
34. Veal, E. A., Day, A. M. & Morgan, B. A. Hydrogen peroxide sensing and signaling. Mol Cell 26, 1–14 (2007).35. Kim, D., Gweon, B., Kim, D. B., Choe, W. & Shin, J. H. A Feasibility Study for the Cancer Therapy Using Cold Plasma. Ifmbe Proc 23,
355–357 (2009).36. Stoffels, E. et al. Plasma needle for in vivo medical treatment: recent developments and perspectives. Plasma Sources Sci T 15,
S169–S180 (2006).37. Tochhawng, L., Deng, S., Pervaiz, S. & Yap, C. T. Redox regulation of cancer cell migration and invasion. Mitochondrion 13, 246–253
(2013).38. Lu, T. Y. & Gabrilovich, D. I. Molecular Pathways: Tumor-Infiltrating Myeloid Cells and Reactive Oxygen Species in Regulation of
Tumor Microenvironment. Clin Cancer Res 18, 4877–4882 (2012).
AcknowledgementsThis work was funded by the Natural Science Fund of Anhui Province (No. 1408085MH180), Chinese Academy of Sciences (CASHIPS) Dean Fund (No. YZJJ201331) and Natural Science Foundation of China (No. 81227902), and the Innovative Program of Development Foundation of Hefei Center for Physical Science and Technology, China (No. 2014FXCX008).
Author ContributionsW.L. and W.H. designed/supervised the study. W.L. and W.H. wrote the manuscript, performed the study and interpreted the results. J.S. and C.C. contributed the materials. K.N.Y. and L.B. reviewed the manuscript. All authors discussed the results and commented on the manuscript.
Additional InformationCompeting financial interests: The authors declare no competing financial interests.How to cite this article: Li, W. et al. Non-thermal plasma inhibits human cervical cancer HeLa cells invasiveness by suppressing the MAPK pathway anddecreasing matrix metalloproteinase-9 expression. Sci. Rep. 6, 19720; doi: 10.1038/srep19720 (2016).
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license,
unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
![Thymoquinoneloadedinnanostructured ... · SH-SY5Y human neuroblastoma cells [10], SW 626 human colon cancer cells [11], ES-2 human ovarian cancer cells [12], HeLa human cervical carcinoma](https://static.fdocuments.net/doc/165x107/604dba5a6c0cbc4067664fea/thymoquinoneloadedinnanostructured-sh-sy5y-human-neuroblastoma-cells-10-sw.jpg)

















