
Non- Mendelian inheritance Ms. Shivani Bhagwat Lecturer, School of Biotechnology DAVV.
53
Non- Mendelian inheritance Ms. Shivani Bhagwat Lecturer, School of Biotechnology DAVV
-
Upload
caren-mathews -
Category
Documents
-
view
222 -
download
1
Transcript of Non- Mendelian inheritance Ms. Shivani Bhagwat Lecturer, School of Biotechnology DAVV.

Non- Mendelian inheritance
Ms. Shivani BhagwatLecturer,School of BiotechnologyDAVV

• Mendelian inheritance patterns– Involve genes directly influencing traits– Obey Mendel’s laws
• Law of segregation• Law of independent assortment
– Include• Dominant / recessive relationships• Gene interactions• Phenotype-influencing roles of sex and environment
– Most genes of eukaryotes follow a Mendelian inheritance pattern

• Many genes do not follow a Mendelian inheritance pattern– e.g., Closely linked genes do not follow Mendel’s law
of independent assortment– non-Mendelian inheritance patterns are:
• Maternal effect• Epigenetic inheritance• Extranuclear inheritance

MATERNAL EFFECT• Maternal effect
– Inheritance pattern for certain nuclear genes– Genotype of mother directly determines phenotype
of offspring• Genotype of father and offspring are irrelevant
– Explained by the accumulation of gene products mother provides to developing eggs

5
MATERNAL EFFECTA. E. Boycott (1920s)• First to study an example of maternal effect• Involved morphological features of water snail
– Limnea peregra– Shell and internal organs can be either right- or left-
handed• Dextral or sinistral, respectively• Determined by cleavage pattern
of egg after fertilization– Dextral orientation is more
common and dominant

6
MATERNAL EFFECT
A. E. Boycott (1920s)• Began with two different true-breeding strains
– One dextral, one sinistral• Dextral ♀ x sinistral ♂ dextral offspring• Reciprocal cross sinistral offspring• Contradict a Mendelian pattern of inheritance

Reciprocal cross is a breeding experiment designed to test the role of parental sex on a given inheritance pattern.
All parent organisms must be true breeding to properly carry out such an experiment.
In one cross, a male expressing the trait of interest will be crossed with a female not expressing the trait. In the other, a female expressing the trait of interest will be crossed with a male not expressing the trait

8
MATERNAL EFFECTA. E. Boycott (1920s); Alfred Sturtevant (1923)• Sturtevant proposed that Boycott’s results could be explained
by a maternal effect gene– Conclusions drawn from F2
and F3 generations
– Dextral (D) is dominant to sinistral (d)
– Phenotype of offspring is determined by genotype of mother

9
EPIGENETIC INHERITANCE
• Epigenetic inheritance– Modification occurs to a nuclear gene or
chromosome– Occur during spermatogenesis, oogenesis, and early
stages of embryogenesis– Gene expression is altered
• May be fixed during an individual’s lifetime
– Expression is not permanently changed over multiple generations
• DNA sequence is not altered

10
EPIGENETIC INHERITANCE
• Two types of epigenetic inheritance: – Dosage compensation
• Offsets differences in the number of sex chromosomes• One sex chromosome is altered
– Genomic imprinting• Occurs during gamete formation• Involves a single gene or chromosome• Governs whether offspring express maternally- or
paternally-derived gene
11
DOSAGE COMPENSATION
• Males and females of many species have different numbers of certain sex chromosomes– e.g., X chromosomes– The level of expression
of many genes on sex chromosomes is similar in both sexes

12
DOSAGE COMPENSATION
• Apricot eye color in Drosophila– Conferred by an X-linked gene– Homozygous females resemble hemizygous males– Females heterozygous for the apricot allele and a
deletion have paler eye color– Two copies of the allele in a female produce a
phenotype similar to one copy in a male– The difference in gene dosage is being
compensated at the level of gene expression
13
DOSAGE COMPENSATION
• Dosage compensation does not occur for all eye color alleles in Drosophila– e.g., Eosin eye color
• Conferred by an X-linked gene• Homozygous eosin females have
darker eye color than hemizygous eosin males
– Dark eosin and light eosin
• Two copies of the allele in a female produce a phenotype different than one copy in a male

14
DOSAGE COMPENSATION
• Most X-linked genes show dosage compensation
• Some X-linked genes do not• Reasons for the difference are not understood

15
DOSAGE COMPENSATION• Mechanisms of dosage compensation
– Mammals• One X chromosome is inactivated in females
– “X inactivation”– Paternally derived in marsupial mammals– Paternal or random, depending on species of placental mammal
– Drosophila melanogaster• Twofold increase in expression of genes on the X chromosome
of males
– The nematode Caenorhabditis elegans• 50% reduction in expression of X-linked genes in XX individuals

16
DOSAGE COMPENSATION
Murray Barr and Ewart Bertram (1949)• Identified a highly condensed structure in interphase nuclei of
somatic cells of female cats– This structure was absent in male cats– “Barr body”– Later identified as a highly
condensed X chromosome– A normal human female has only one Barr body per
somatic cell, while a normal human male has none

17
DOSAGE COMPENSATIONMary Lyon (1961)• Aware of Barr and Bertram cytological evidence• Also aware of mammalian mutations producing a variegated
coat color pattern– e.g., Calico cats are heterozygous
for X-linked alleles determining coat color
• Possess randomly distributed patches of black and orange
• Lyon hypothesis: A single X chromosome was inactivated in the cells of females

Calico cats are almost always female because the X chromosome determines the color of the cat and female cats, much like all female mammals, have two X chromosomes, whereas male mammals, including common male cats, have one X and one Y chromosome.
Since the Y chromosome does not have any color genes, there is no chance he could have both orange and non-orange together.
One main exception to this is when, in rare cases, a male has XXY chromosomes (see Klinefelter's syndrome), in which case the male could have tortoiseshell or calico markings.
Male calico or tortoiseshell cats are sterile due to the abnormality of carrying two X chromosomes.
The orange mutant gene is found only on the X, or female, chromosome. The female cat, therefore, can have the orange mutant gene on one X chromosome and the genes for a black or white coat on the other, and those can affect or modify the orange mutant gene.
If that is the case, those several genes will be expressed in a blotchy coat of the tortoiseshell or calico kind.

19
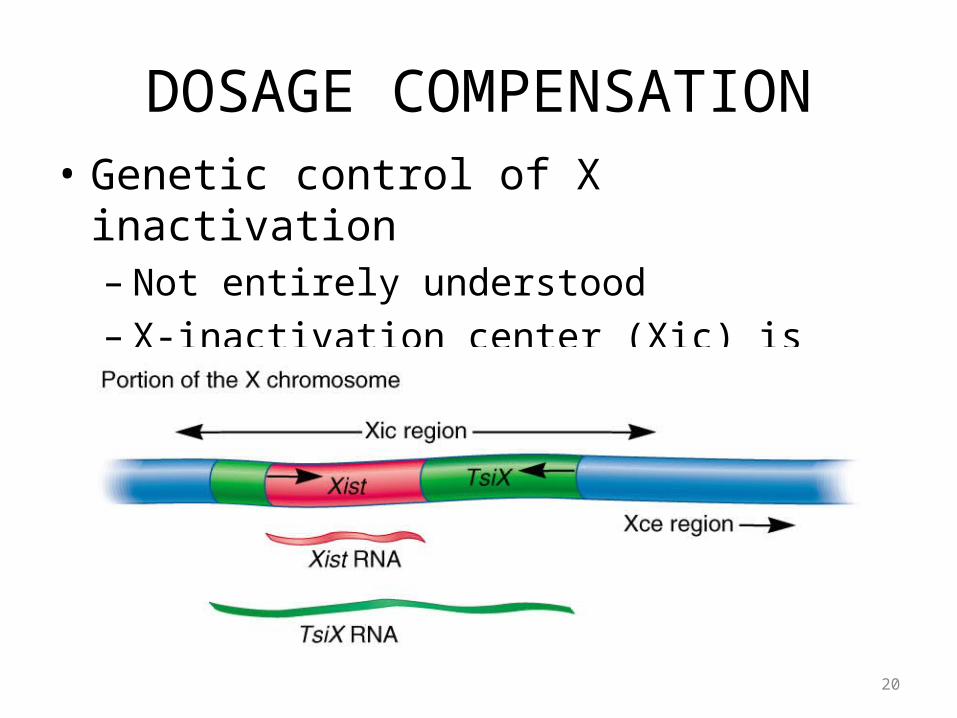
DOSAGE COMPENSATION
• Genetic control of X inactivation– Human cells (and those of other mammals)
possess the ability to count their X chromosomes– Only one is allowed to remain active
• XX females 1 Barr body• XY males 0 Barr bodies• XO females 0 Barr bodies (Turner syndrome)• XXX females 2 Barr bodies (Triple X syndrome)• XXY males 1 Barr body (Kleinfelter syndrome)
20
DOSAGE COMPENSATION• Genetic control of X inactivation
– Not entirely understood– X-inactivation center (Xic) is involved
• Short region of the X chromosome

21
Genomic Imprinting
• Genomic imprinting involves the physical marking of a segment of DNA– Mark is retained and recognized throughout the
life of the organism inheriting the marked DNA– Resulting phenotypes display non-Mendelian
inheritance patterns– Offspring expresses one allele, not both– “Monoallelic expression”

Extranuclear inheritance
Extranuclear inheritance (also known as cytoplasmic inheritance) is a form of non-Mendelian inheritance first discovered by Carl Correns in 1908. While working with Mirabilis jalapa. Correns observed that leaf color was dependent only on the genotype of the maternal parent. Based on these data, he determined that the trait was transmitted through a character present in the cytoplasm of the ovule.
Gene conversion
Gene conversion is a reparation process in DNA recombination, by which a piece of DNA sequence information is transferred from one DNA helix (which remains unchanged) to another DNA helix, whose sequence is altered. This may occur as a mismatch repair between the strands of DNA which are derived from different parents. Thus the mismatch repair can convert one allele into the other. This phenomenon can be detected through the offspring non-Mendelian ratios, and is frequently observed, e.g., in fungal crosses

Infectious heredity
Infectious particles such as viruses may infect host cells and continue to reside in the cytoplasm of these cells. If the presence of these particles results in an altered phenotype, then this phenotype may be subsequently transmitted to progeny.Because this phenotype is dependent only on the presence of the invader in the host cell’s cytoplasm, inheritance will be determined only by the infected status of the maternal parent. This will result in a uniparental transmission of the trait, just as in extranuclear inheritance.
Genomic imprinting
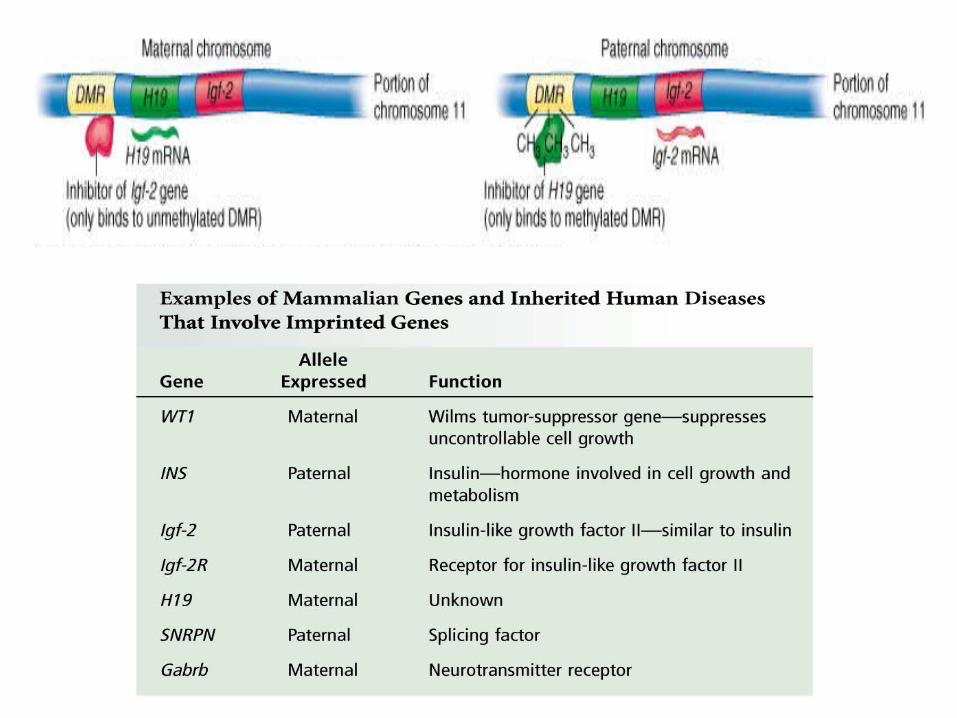
Genomic imprinting is a genetic phenomenon by which certain genes are expressed in a parent-of-origin-specific manner. Imprinted alleles are silenced such that the genes are either expressed only from the non-imprinted allele inherited from the mother (e.g. H19 or CDKN1C), or in other instances from the non-imprinted allele inherited from the father (e.g. IGF-2).
Forms of genomic imprinting have been demonstrated in insects, mammals and flowering plants.

Effects can includeA single geneA part of a chromosomeAn entire chromosomeAll the chromosomes from one parent
Genomic imprinting can also be correlated with the process of X inactivation
In some species, imprinting determines which X chromosome will be inactivatede.g., The paternal X chromosome is always inactivated in marsupialse.g., The paternal X chromosome is inactivated in extra embryonic tissue (e.g., the placenta) of placental mammals
X inactivation is random in the placental embryo itself.
Involves the physical marking of DNA. Also involves differentially methylated regions (DMRs) located near imprinted genes
Maternal or paternal copy is methylated, not both

Mosaicism
Individuals who possess cells with genetic differences from the other cells in their body are termed mosaics. These differences can result from mutations that occur in different tissues and at different periods of development.Mosaicism also results from a phenomenon known as X-inactivation. All female mammals have two X chromosomes. To prevent lethal gene dosage problems, one of these chromosomes is inactivated following fertilization. This process occurs randomly for all of the cells in the organism’s body. Because a given female’s two X chromosomes will almost certainly differ in their specific pattern of alleles, this will result in differing cell phenotypes depending on which chromosome is silenced. Calico cats, which are almost all female, demonstrate one of the most commonly observed manifestations of this process.
Trinucleotide repeat disorders
These diseases are all caused by the expansion of microsatellite tandem repeats consisting of a stretch of three nucleotides. In normal individuals, the number of repeated units is relatively low. With each successive generation, there is a chance that the number of repeats will expand. As this occurs, progeny can progress to permutation and ultimately affected status. Individuals with a number of repeats that falls in the permutation range have a good chance of having affected children.

Uniparental disomy
Uniparental disomy (UPD) occurs when a person receives two copies of a chromosome, or part of a chromosome, from one parent and no copies from the other parent.
1. UPD could involve isodisomy (meiosis II error)
2. heterodisomy (meiosis I error)
Uniparental inheritance of imprinted genes can also result in phenotypical anomalies. Though few imprinted genes have been identified, uniparental inheritance of an imprinted gene can result in the loss of gene function which can lead to delayed development, mental retardation, or other medical problems.The most well-known conditions include Prader-Willi syndrome and Angelman syndrome. Both of these disorders can be caused by UPD or other errors in imprinting involving genes on the long arm of chromosome 15.Other conditions, such as Beckwith-Wiedemann syndrome, are associated with abnormalities of imprinted genes on the short arm of chromosome 11.Chromosome 14 is also known to cause particular symptoms such as skeletal abnormalities, mental retardation and joint contractures among others.

Multifactorial (Complex) Inheritance
Expression of many traits may involve multiple genes.
Many such traits (eg, height) are distributed along a bell-shaped curve (normal distribution). Normally, each gene adds to or subtracts from the trait independently of other genes. In this distribution, few people are at the extremes and many are in the middle because people are unlikely to inherit multiple factors acting in the same direction. Environmental factors also add to or subtract from the final result.
Many relatively common congenital anomalies and familial disorders result from multifactorial inheritance. In an affected person, the disorder represents the sum of genetic and environmental influences. Risk of the occurrence of such a trait is much higher in 1st-degree relatives (siblings, parents, or children who share, on average, 50% of the affected person's genes) than in more distant relatives, who are likely to have inherited only a few high-liability genes.

Genetically determined predisposing factors, including a family history and specific biochemical pathways often identified by molecular markers (eg, high cholesterol), can sometimes identify people who are at risk and are likely to benefit from preventive measures.
Multigenic, multifactorial traits seldom produce clear patterns of inheritance; however, these traits tend to occur more often among certain ethnic and geographic groups or among one sex or the other.
Exceptions to Simple InheritancePolygenic Traits(stature, body shape, hair and skin color)Intermediate Expression(AA,Aa,aa)Codominance (A,B,AB blood gps)Multiple-allele Series (ABO blood grouping)Modifying and Regulator Genes: Modifying genes alter how certain genes are expressed in the phenotype. For instance, there is a dominant cataract gene which will produce varying degrees of vision impairment depending on the presence of a specific allele for a companion modifying gene. However, cataracts also can be promoted by diabetes and common environmental factors such as excessive ultraviolet radiation and alcoholism.

Regulator genes can either initiate or block the expression of other genes. They control the production of a variety of chemicals in plants and animals. For instance, the time of production of specific proteins that will be new structural parts of our bodies can be controlled by such regulator genes. they control the maturation and aging processes. Regulator genes that are involved in subdividing an embryo into what will become the major body parts of an individual are also referred to as control, homeotic , homeobox , or Hox genes. They are responsible for setting generalized cells on the path to become a head, torso, arms, legs, etc.
Incomplete Penetrance
Some genes are incompletely penetrant. That is to say, their effect does not normally occur unless certain environmental factors are present.
For example, you may inherit the genes that are responsible for type 2 diabetes but never get the disease unless you become greatly overweight, persistently stressed psychologically, or do not get enough sleep on a regular basis.
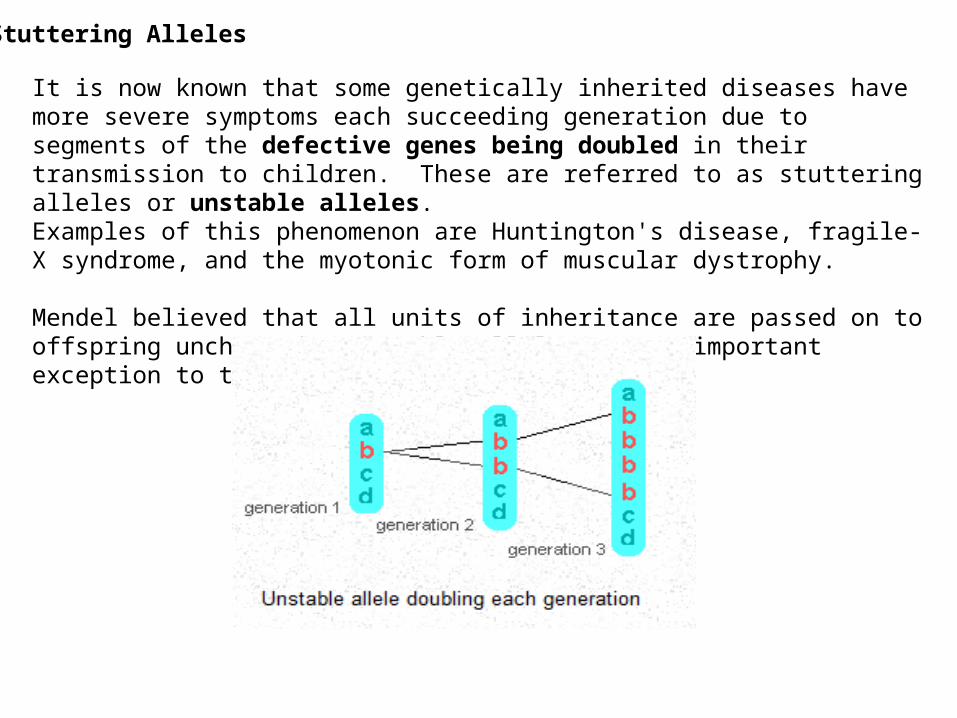
Stuttering Alleles
It is now known that some genetically inherited diseases have more severe symptoms each succeeding generation due to segments of the defective genes being doubled in their transmission to children. These are referred to as stuttering alleles or unstable alleles. Examples of this phenomenon are Huntington's disease, fragile-X syndrome, and the myotonic form of muscular dystrophy.
Mendel believed that all units of inheritance are passed on to offspring unchanged. Unstable alleles are an important exception to this rule.

Behavioural genetics
The field of study that examines the role of genetics in animal (including human) behaviour. Often associated with the "nature versus nurture" debate, behavioural genetics is highly interdisciplinary, involving contributions from biology, genetics, ethology, psychology, and statistics. Behavioural geneticists study the inheritance of behavioural traits. In humans, this information is often gathered through the use of the twin study or adoption study. In animal studies, breeding, transgenesis, and gene knockout techniques are common; psychiatric genetics.
Sir Francis Galton (1822-1911) was the first scientist to study heredity and human behaviour systematically.
What indications are there that behaviour has a biological basis? Behavior often is species specificBehaviors often breed trueBehaviors change in response to alterations in biological structures or processesIn humans, some behaviors run in familiesBehavior has an evolutionary history that persists across related species
Genes are evolutionary glue, binding all of life in a single history that dates back some 3.5 billion years. Conserved behaviors are part of that history.

Twin study
Twin studies help disentangle the relative importance of environmental and genetic influences on individual traits and behaviors.
Twins are a valuable source for observation due to their genotypes and family environments tending to be similar. More specifically, monozygotic (MZ) or "identical" twins, share nearly 100% of their genetic polymorphisms, which means that most variation in pairs' traits (measured height, susceptibility to boredom, intelligence, depression, etc.) is due to their unique experiences. Dizygotic (DZ) or "fraternal" twins share only about 50% of their polymorphisms. DZ twins are helpful to study because they tend to share many aspects of their environment (e.g., parenting style, education, wealth, culture, community) by virtue of being born in the same time and place.
The basic logic of the twin study can be understood with very little mathematics beyond an understanding of correlation and the concept of variance.

Quantitative genetics
Quantitative genetics is the study of continuously measured traits (such as height or weight) and their mechanisms. It can be an extension of simple Mendelian inheritance in that the combined effects of one or more genes and the environments in which they are expressed give rise to continuous distributions of phenotypic values.Basic principlesThe phenotypic value (P) of an individual is the combined effect of the genotypic value (G) and the environmental deviation (E):P = G + E The genotypic value is the combined effect of all the genetic effects, including nuclear genes, mitochondrial genes and interactions between the genes.
Resemblance between relatives
Central in estimating the variances for the various components is the principle of relatedness. A child has a father and a mother. Consequently, the child and father share 50% of their alleles, as do the child and the mother. However, the mother and father normally do not share alleles as a result of shared ancestors.

The principle of relationship (R) is central to understanding the resemblances within families and can be useful when calculating inbreeding.
Relationship has two definitions that can be applied:1. The probable portion of genes that are same for two individuals due to common ancestry exceeding that of the base population.
2. Additive/numerator relationship: the relationship coefficient (Rxy¬) = twice the probability of two genes at loci in different individuals being identical by descent.
Rxy values can range from 0 to 2.
Relationship can be calculated in several ways;
a.from the known relationships of the individualb. from bracket pedigreesc.from pedigree path diagrams.
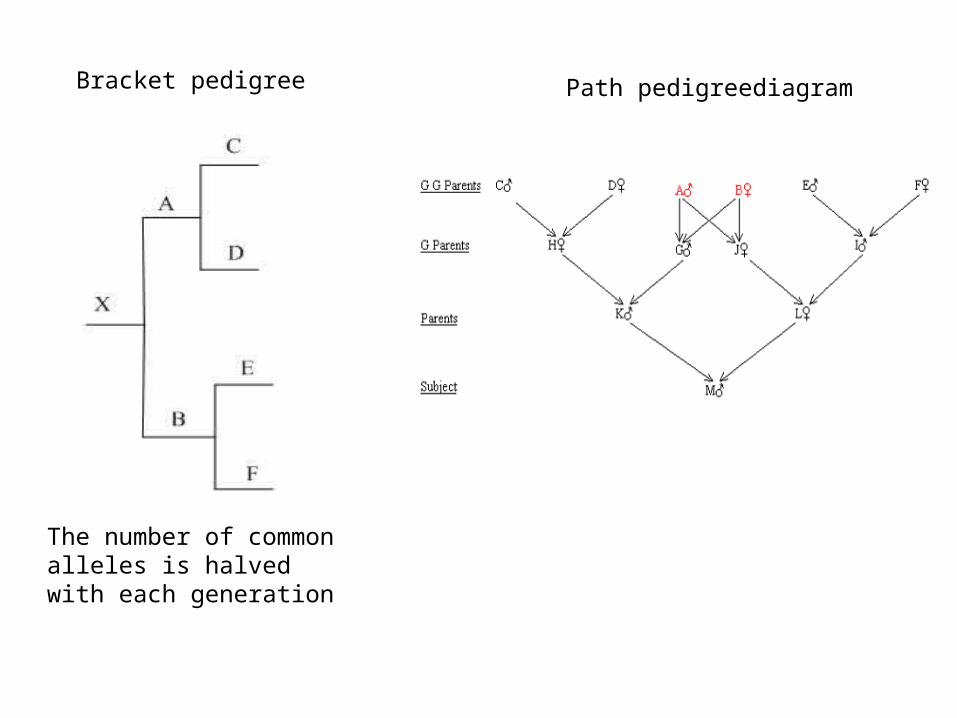
The number of common alleles is halved with each generation
Bracket pedigree Path pedigreediagram

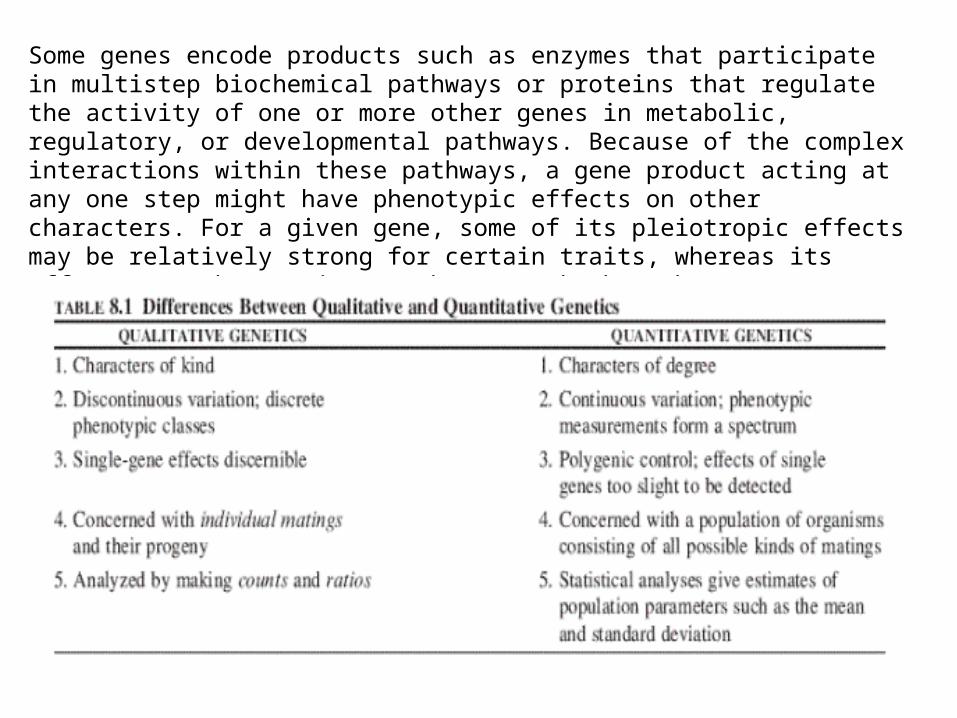
Qualitative vs. Quantitative Traits
The classical Mendelian traits encountered have been qualitative in nature; i.e., traits that are easily classified into distinct phenotypic categories. These discrete phenotypes are under the genetic control of only one or a very few genes with little or no environmental modification to obscure the gene effects. In contrast, the variability exhibited by many traits fails to fit into discrete phenotypic classes (discontinuous variability), but instead forms a spectrum of phenotypes that blend imperceptively (continuous variability). Economically important traits such as body weight gains in livestock, mature plant heights, egg or milk production, and yield of grain per acre are quantitative or complex traits with continuous variability.
The basic differences between qualitative and quantitative traits involve the number of genes contributing to the phenotypic variability and the degree to which the phenotype can be modified by environmental factors.

Quantitative traits are governed by many genes (perhaps 10–100 or more), each contributing a small amount to the phenotype are their individual effects cannot be detected by Mendelian methods.
For this reason, quantitative traits are also referred to as polygenic traits. Stretches of DNA with closely linked genes responsible for phenotypes associated with quantitative traits are called quantitative trait loci or QTLs.
Each gene usually has effects on more than one trait. The idea that each character is controlled by a single gene (the one-gene-one-trait hypothesis) has often been falsely attributed to Mendel. But even he recognized that a single factor (or gene) might have manifold effects on more than one trait.
For example, he observed that purple flowers are correlated with brown seeds and a dark spot on the axils of leaves; similarly, white flowers are correlated with light-colored seeds and no axillary spots on the leaves. In Drosophila, many loci (e.g., genes named dumpy, cut, vestigial, apterous) contribute to a complex character such as wing development. Each of these genes also has pleiotropic effects on other traits. For example, the gene for vestigial wings also effects the halteres (balancers), bristles, egg production in females, and longevity.
Some genes encode products such as enzymes that participate in multistep biochemical pathways or proteins that regulate the activity of one or more other genes in metabolic, regulatory, or developmental pathways. Because of the complex interactions within these pathways, a gene product acting at any one step might have phenotypic effects on other characters. For a given gene, some of its pleiotropic effects may be relatively strong for certain traits, whereas its effects on other traits may be so weak that they are difficult or impossible to identify by Mendelian techniques. It is the totality of these pleiotropic effects of numerous loci that constitutes the genetic base of a quantitative character.
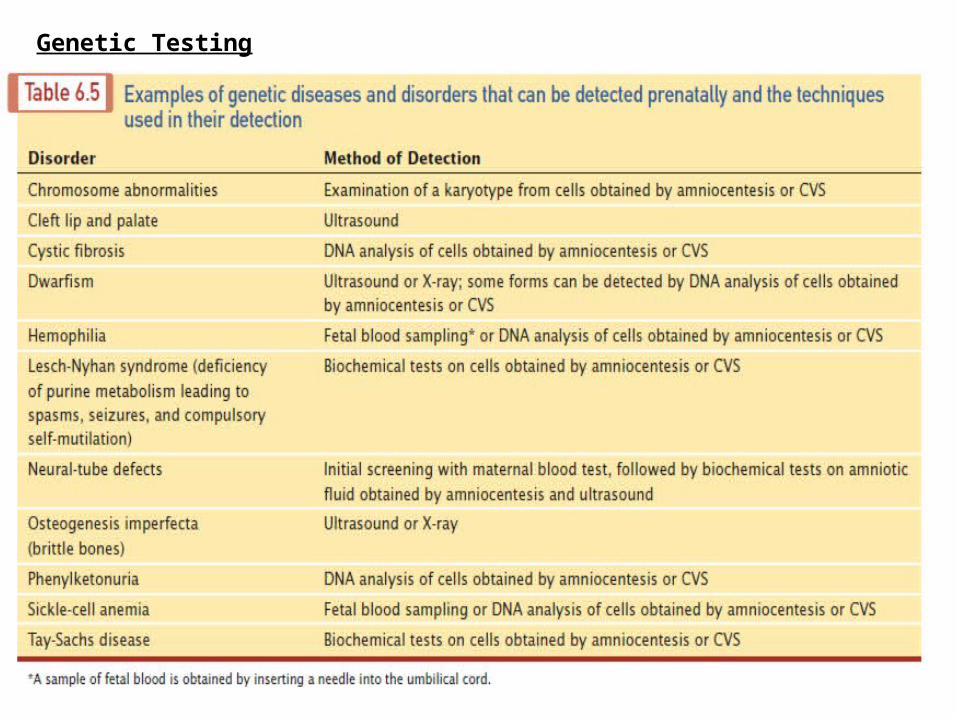
Genetic Testing

Non-disjunction
Nondisjunction ("not coming apart") is the failure of chromosome pairs to separate properly during meiosis stage 1 or stage 2. This could arise from a failure of homologous chromosomes to separate in meiosis I, or the failure of sister chromatids to separate during meiosis II or mitosis. The result of this error is a cell with an imbalance of chromosomes. Such a cell is said to be aneuploid. Loss of a single chromosome (2n-1), in which the daughter cell(s) with the defect will have one chromosome missing from one of its pairs, is referred to as a monosomy. Gaining a single chromosome, in which the daughter cell(s) with the defect will have one chromosome in addition to its pairs is referred to as a trisomy (2n+1).

Once in a Blue Moon
One of the best-known facts of genetics is that a cross between a horse and a donkey produces a mule.
Actually, it’s a cross between a female horse and a male donkey that produces the mule; the reciprocal cross, between a male horse and a female donkey, produces a hinny, which has smaller ears and a bushy tail, like a horse . Both mules and hinnies are sterile because horses and donkeys are different species with different numbers of chromosomes: a horse has 64 chromosomes, whereas a donkey has only 62. There are also considerable differences in the sizes and shapes of the chromosomes that horses and donkeys have in common. A mule inherits 32 chromosomes from its horse mother and 31 chromosomes from its donkey father, giving the mule a chromosome number of 63.
The maternal and paternal chromosomes of a mule are not homologous, and so they do not pair and separate properly in meiosis; consequently, a mule’s gametesare abnormal and the animal is sterile.
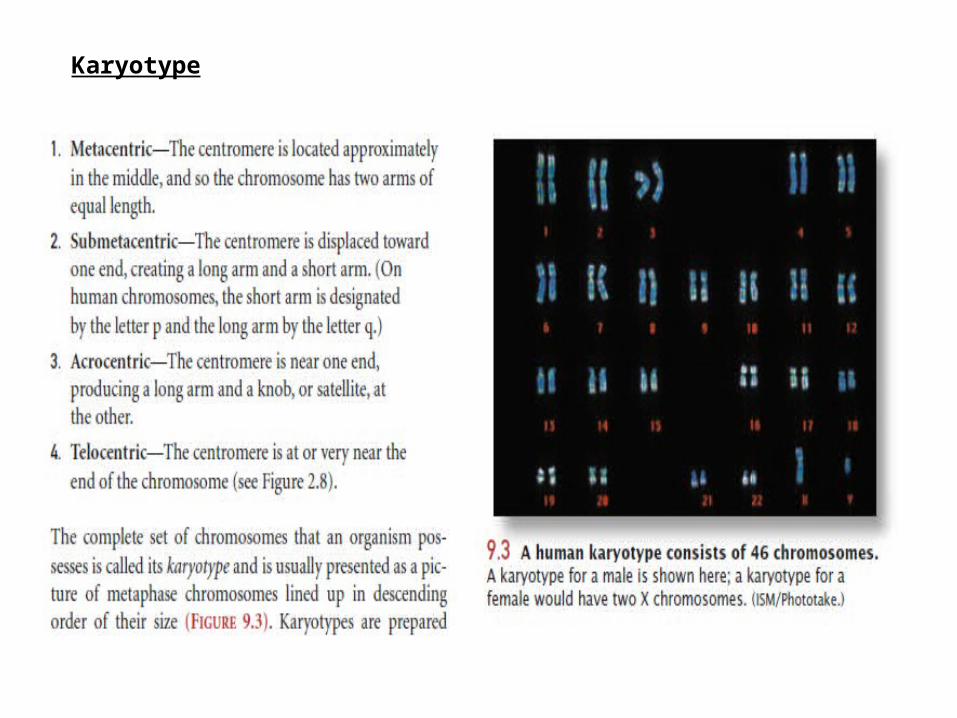
Karyotype

Preparation and staining techniques help to distinguish among chromosomes of similar size and shape.
1. chromosomes may be treated with enzymes that partly digest them; staining with a special dye called Giemsa reveals G bands, which distinguish areas of DNA that are rich in adenine–thymine base pairs.
2. Q bands are revealed by staining chromosomes with quinacrine mustard and viewing the chromosomes under UV light.
3. C bands which are regions of DNA occupied by centromeric heterochromatin.
4. R band, which are rich in guanine–cytosine base pairs.

Chromosome abnormality
A chromosome anomaly, abnormality or aberration reflects an atypical number of chromosomes or a structural abnormality in one or more chromosomes. A Karyotype refers to a full set of chromosomes from an individual which can be compared to a "normal" Karyotype for the species via genetic testing. Chromosome anomalies usually occur when there is an error in cell division following meiosis or mitosis. There are many types of chromosome anomalies.
They can be organized into two basic groups:a. numerical b. structural anomalies.

Numerical Disorders
This is called Aneuploidy (an abnormal number of chromosomes), and occurs when an individual is missing either a chromosome from a pair (monosomy) or has more than two chromosomes of a pair (Trisomy, Tetrasomy, etc.).In humans an example of a condition caused by a numerical anomaly is Down Syndrome, also known as Trisomy 21 (an individual with Down Syndrome has three copies of chromosome 21, rather than two). Turner Syndrome is an example of a monosomy where the individual is born with only one sex chromosome, an XO.


Structural abnormalities
When the chromosome's structure is altered, this can take several forms:
Deletions: A portion of the chromosome is missing or deleted. Duplications: A portion of the chromosome is duplicated, resulting in extra genetic material. Translocations: When a portion of one chromosome is transferred to another chromosome. In a reciprocal translocation, segments from two different chromosomes have been exchanged. Inversions: A portion of the chromosome has broken off, turned upside down and reattached, therefore the genetic material is inverted.Insertions: A portion of one chromosome has been removed from its normal place and inserted into another chromosome.
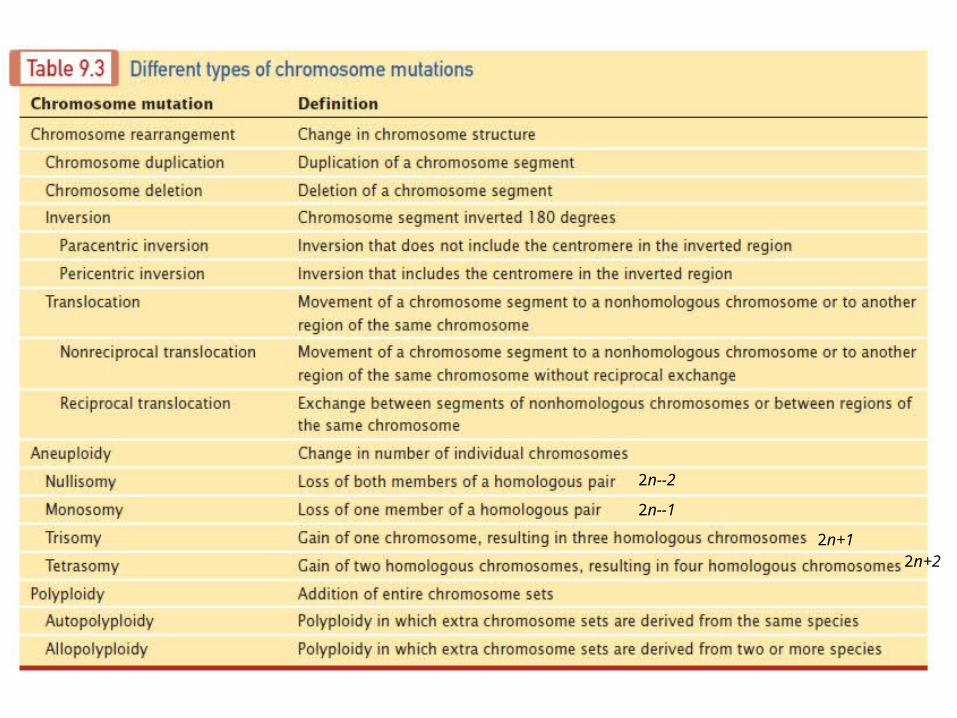
2n--2
2n--1
2n+12n+2

Y chromosome
“Functional wasteland,” “Nonrecombining desert” and “Gene-poor chromosome” are some of the different definitions given to the Y chromosome in the last decade.
The Y chromosome is one of the two sex-determining chromosomes in most mammals, including humans. In mammals, it contains the gene SRY((Sex-determining region Y), which triggers testis development if present. The human Y chromosome is composed of about 60 million base pairs. DNA in the Y chromosome is passed from father to son, and Y-DNA analysis may thus be used in genealogy research. With a 30% difference between humans and chimpanzees, the Y chromosome is one of the fastest evolving parts of the human genome
Before Y chromosomeMany ectothermic vertebrates have no sex chromosomes. If they have different sexes, sex is determined environmentally rather than genetically. For some of them, especially reptiles, sex depends on the incubation temperature; others are hermaphroditic (meaning they contain both male and female gametes in the same individual).

Origin
The X and Y chromosomes are thought to have evolved from a pair of identical chromosomes, termed autosomes, when an ancestral mammal developed an allelic variation, a so-called 'sex locus' – simply possessing this allele caused the organism to be male.The chromosome with this allele became the Y chromosome, while the other member of the pair became the X chromosome. Over time, genes which were beneficial for males and harmful to (or had no effect on) females either developed on the Y chromosome, or were acquired through the process of translocation.
Recombination inhibitionRecombination between the X and Y chromosomes proved harmful—it resulted in males without necessary genes formerly found on the Y chromosome, and females with unnecessary or even harmful genes previously only found on the Y chromosome. As a result, genes beneficial to males accumulated near the sex-determining genes, and recombination in this region was suppressed in order to preserve this male specific region.Over time, the Y chromosome changed in such a way as to inhibit the areas around the sex determining genes from recombining at all with the X chromosome. As a result of this process 95% of the human Y chromosome is unable to recombine.

Shrinking theory
The human Y chromosome has lost 1,393 of its 1,438 original genes over the course of its existence. With a rate of genetic loss of 4.6 genes per million years, the Y chromosome may potentially lose complete function within the next 10 million years.
High mutation rateThe human Y chromosome is particularly exposed to high mutation rates due to the environment in which it is housed. The Y chromosome is passed exclusively through sperm, which undergo multiple cell divisions during gametogenesis. Each cellular division provides further opportunity to accumulate base pair mutations. Additionally, sperms are stored in the highly oxidative environment of the testis, which encourages further mutation. These two conditions combined, put the Y chromosome at a risk of mutation 4.8 times greater than the rest of the genome.

Human Y chromosome
In humans, the Y chromosome spans about 58 million base pairs (the building blocks of DNA) and represents approximately 2% of the total DNA in a male cell. The human Y chromosome contains 86 genes, which code for only 23 distinct proteins. Traits that are inherited via the Y chromosome are called holandric traits.
The bulk of the Y chromosome which does not recombine is called the "NRY" or non-recombining region of the Y chromosome. It is the SNPs in this region which are used for tracing direct paternal ancestral lines.


















