
Autotrophic Nitrogen Removal from Low Concentrated Effluents

Vol. 50, No. 6APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1985, p. 1519-15250099-2240/85/121519-07$02.00/0Copyright C) 1985, American Society for Microbiology
Nitrous Oxide Emission Associated with Autotrophic AmmoniumOxidation in Acid Coniferous Forest Soil
PERTTI J. MARTIKAINEN
Department of General Microbiology, University of Helsinki, SF-00280 Helsinki 28, Finland
Received 1 May 1985/Accepted 25 August 1985
Aerobic N20 production was studied in nitrifying humus from urea-fertilized pine forest soil. Acetylene andnitrapyrin inhibited both NH4' oxidation and N20 production, indicating that N20 production was closelyassociated with autotrophic NH4+ oxidation. N20 production was enhanced by low soil pH; it was negligibleabove pH 4.7. When soil pH decreased from 4.7 to 4.1, the relative amount of N20-N produced from NH4+-Noxidized increased exponentially to 20%. There was also some evidence that N20 formation was stimulated bysalts (potassium sulfate and sodium phosphates). The maximum rate of N20-N production was 0.17 ,ig ofN20-N per g of soil per h. When humus was treated with N02-, N20 evolved immediately, indicating chemicalformation, but no N20 was formed on the addition of N03-. The amount of N20-N evolved was 0.6 to 4.6%of N02--N added. A high concentration of N02- and low soil pH enhanced chemical production of N20. Therewas no accumulation of N02- during nitrification. The calculations indicated that chemical formation of N20was not the main source of N20 during NH4+ oxidation. After the addition of inhibitors of NH4+ oxidation thesoils contained N03 , but no N20 was produced. The results suggest that enhanced autotrophic NH4+ oxidationis a potential source of N20 in fertilized acid forest soil.
Besides being produced during biological denitrificationN20 is also formed during NH4' oxidation. Results ofstudies with pure cultures of NH4+ oxidizers have revealedthat 0.05 to 25% of the nitrogen from N02- (NO2--N)produced evolved as N20-N when NH4+-N is used as sub-strate (13, 16, 17, 41). N20 production from NH2OH, whichis an intermediate in NH4+-N oxidation (1), can be muchhigher. Nitrosomonas europaea can produce over 90% ofN20-N from NH2OH-N that is oxidized (41). In well-aeratedsoils N20-N production ranges from 0.01 to 2% of NH4+-Nadded or N03 -N produced (2, 3, 5, 8, 12). When NO02-Naccumulates in soil during NH4+-N oxidation, even higheramounts of N20-N may be formed (33) via chemodenitrifica-tion (9), in addition to direct N20 emission from cells ofNH4+-oxidizing bacteria. Although N20 emission duringnitrification may not be of general agronomic importance, itnevertheless is a source ofN20 postulated to be linked to thecatalytic destruction of stratospheric ozone (10, 40).Most studies of N20 emission from soil, using aerobic
atmosphere, have been made with agricultural soils, andonly a few results are available from acid forest soils.Goodroad and Keeney (12) have measured N20 fluxes indeciduous and coniferous forest sites and have found thatsignificantly more N20 is emitted from coniferous than fromdeciduous forest floors. They concluded that the lower pH ofthe coniferous forest soil might cause the difference.Robertson and Tiedje (30) have studied N20 production inforest soils using intact soil cores in a recirculating aerobicatmosphere. Their results suggest that in some acid forestsoils mechanisms other than denitrification might be impor-tant in N20 production. This report presents results fromlaboratory experiments in which N20 production was stud-ied in an aerobic atmosphere of fertilized acid coniferousforest humus.
MATERIALS AND METHODS
Soil samples were collected in 1983 from an experimentalMyrtillus pine forest site located at Tammela in southernFinland. The soil and the fertilization treatments have been
described previously (18). The urea fertilization applied inautumn 1978 was 200 kg ofN/ha. Some soil characteristics aregiven in Table 1. The samples were taken from the organichorizon of both unfertilized and urea-fertilized soils. Thesampling techniques and the soil homogenization methodhave been described previously (18). Storage and laboratorymanipulations are described below.
Analytical methods. Soil pH was measured in soil-watersuspensions (1/2 [vol/vol]). When exchangeable ammonium,nitrate, and nitrite were determined after the pH was mea-sured, 4 M KCl was added to the suspensions to obtain 2 MKCl for extraction. Ammonium and combined nitrate plusnitrite were determined by steam distillation (6). Nitrite wasmeasured by the method of Montgomery and Dymock (21).Total N was determined by the Kjehldahl method (7), andtotal C was determined with a Hewlett-Packard 185 C-H-Nanalyzer. N20 and 02 were determined with a Carlo Erba4200 gas chromatograph equipped with an electron capturedetector and a Porapak Q 80/100-mesh column (length, 2 m).Detector, injector, and oven temperatures were 200 to 300,130, and 50°C, respectively. The flow of carrier gas (eitherN2 or 90% argon-10% methane) was 30 ml/min. Analyses areexpressed on an oven dry weight basis. The results weretreated by the analysis of variance or correlation. Thedifferences between means of various treatments weretested, subsequent to analysis of variance, by the Tukeyhonestly significant difference method (34).
Soil incubations for measuring nitrification and N20 pro-duction. Incubations were performed at 14°C in 100-mlbottles, stopped with rubber septa, containing 1.5 to 2 g ofsoil (oven dry weight). Both fresh and air-dried soils wereused. In all experiments soil moisture content during incu-bation was 60% of the water-holding capacity (WHC). Peri-odically, the bottles were aerated after determination of theamount of N20 produced. At intervals three replicate bottlesfor every treatment were analyzed for nitrate and nitrite(after measurement of soil pH; see above). The partialpressure of 02 was always > 0.18. Autotrophic ammoniumoxidation was repressed by adding nitrapyrin (31) in water
1519
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 1. Description of soils
Previous Soil parameterfertilizertreatmenta pH (in H20) Total C (%) Total N (%) C/N ratio
None 4.0-4.2 31.9 + 6.6" 0.83 + 0.13 38.3 + 4.7bUrea 4.4-4.7 35.0 + 5.1' 1.00 + 0.12C 35.1 ± 1.0cUABM 4.6-4.7 27.7 ± 5.8' 0.80 ± 0.11O 34.2 ± 4.6C
a For the amounts, see Martikainen (18).b Values are the mean ± standard deviation for nine determinations.C Values are the mean + standard deviation for three determinations.
solution (50 ,ug/g of soil; Dow N-serve [18]) or C2H2 (10%[vol/vol]) (15, 16). All nutrients were added in solution. SoilpH was adjusted by adding CaCO3.
In the first experiment samples were taken from the plotfertilized with urea, apatite, biotite, and micronutrients(UABM). It has already been shown that, on fertilization,nitrification activity is highest in the plot fertilized withUABM (18). Soil samples for this experiment were taken inmid-January 1983 (soil temperature at a depth of 5 cm was 0to 0.5°C). After homogenization soil was amended with urea(200 ,ug ofN per g of soil) and stored in polyethylene bags for3 weeks at 4°C to activate nitrification. Fresh soil (2 g ovendry weight) was placed in bottles, some of which weresupplemented with CaCO3 (18 mg/g of soil) to obtain a higherpH.
In the second experiment the same UABM soil as de-scribed above was used, but 40 ,ug of urea-N was added priorto a 3-week storage at 4°C. The effects of the addition ofK2SO4 on nitrification and N2O production were tested inthis experiment. It previously has been found that in UABM-fertilized soil nitrification is highly inhibited on the additionof 53.5 ,umol of K2SO4 per g of soil (19). Here, K2SO4 wasadded at 5.35 ,umol per g of soil.The experiments described above were done with UABM
soil. N20 production during nitrification was also measuredin urea-fertilized soil not treated with apatite, biotite, andmicronutrients. Samples for this experiment were taken inMay 1983. Lime, ammonium, phosphate, and micronutrientswere tested by the treatments shown in Table 2. Becausenutrients were added in water solutions, soil was air dried(after homogenization with a Moulinex homogenizer), sothat all solutions could be added to obtain a final soilmoisture content of 60% of the WHC. Soil pH was adjustedby adding CaCO3, after which the soil was added to 100-mlincubation flasks, and the nutrient solutions were thenadded.N20 production from added nitrite. Chemical production
of N20 from NO2- was tested by adding NO2- to freshhumus samples. Aerobic soil incubations were performed asdescribed above, except that each bottle contained 3 g of soil(oven dry weight). Soils were not sterilized before theaddition of NO2-. The possibility that evolution of N20 wasdue to biological denitrification was tested by determiningthe amount of N20 produced from added NO -. NO2-additions were 0, 50, and 200 ,ug of N per g of soil. ChemicalN2O evolution at low soil pH was studied by supplementingsamples with H2SO4. Additions of H+ were 0, 57, and 210,xmol of H+ per g of soil. The highest H+ level wasequivalent to an oxidation by autotrophic ammonium oxidiz-ers of 1,500 ,ug of NH4+-N to NO2--N (2 mol of H+ isproduced for every 1 mol of NH4' oxidized). An amount of1,500 ,ug of N is equivalent to an NH4+-N application of 200
Z O) :>
+txI
o 0
_- Z Z
z0
o
zr
6z
c0a
0t
0
C)00
C)C.0
C)
C)aeC)
0.tl O
a,a
L.a
00
0
a-
0
0
0e
C)CO
2
.000C)0t00
.0
.E00e
C)
a0
._
0
2
0_
2
C)
._
C)
S
ma
rsnWI)v c
00~ONNN 0 0
00 l0-,
0+1 +1±+1+1+100 en 00 00ON 00 WN 'n VI)
-OOW)ON
--VI 0+1 +1 +1 +1 +1 +1
IR'N "~ 00 Cl-
.- - . . .ON N_O ON 0
ONC 0 rN .0 ~0+1 tl +1 +1 +1 +1
++1 +1+1 +1+1
WI)CCl- 'I
+1 +1+1+1
rI eq en I al V-
d N oo _
£N_l_ -4
ON '. Vl_ 0
+l +1 +1 +1 +1 +1
ON%Cr-N 00 c
ClC'^N'ON ~N
C t Cl. -,-
CN cNC-l00C
+1 +1 t +1 +1 +1O N 0ONOtN00 ON 'f
+- +1 +1 +1 +1 +1'00 00 0VC
ON-4', Cl - -
+l +1 +1 +1 +1 +1
ON e'N '.0 Cl '.q ON
C* Cl 'C 0e
+1l +1 +1 +1 +1
-.00 Cl -n
NCN N N 00
- _. t CN _ 0
- +l- +l Cl-+
+1+1+ 1+1+1+- 'N N '.0 0N !'ON ^ 00 uN - N
Cl'N Clx. 00s C'
0l Cll-00 Cl+
_ ON l l /Nr'
N - r~ ~
++a CZZ
Oaaaaa
ZQQQQQm
.C
0
E0
4)._
C)a0
0.00
C)C)
o_
._w
z
z.
z
z
z
z
z
z
z
z
z
z
z
C)aCZI-
1520 MARTIKAINEN
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from
N20 EMISSION IN FOREST SOIL 1521
kg/ha to the 0 horizon (depth, 5 cm; bulk density, 0.26g/cm3).
RESULTS
Nitrification and N20 production. In the first experimentthe lag period before nitrification was shorter in soils treatedwith CaCO3 than in nontreated soils (Fig. 1). Also themaximum rate of nitrification and the amount of (NO3- +NO2-)-N accumulated was higher in CaCO3-amended soils(P < 0.001; Fig. 1). NO2-N did not accumulate duringincubation (its concentration remained at <1 p.g/g of soil).After a 2-week incubation, when nitrification activity wasstill low (Fig. 1), soil pH was 5.2 and 6.8 without and withCaCO3, respectively. During nitrification soil pH decreasedbut was always higher in soils treated with CaCO3 than innontreated soils (P < 0.001; Fig. 1). Although accumulationof (NO3- + NO2)-N was higher in soils with lime, produc-tion of N20 was lower in these soils than in soils without lime(P < 0.001; Fig. 1). The addition of C2H2 inhibited nitrifica-tion and N20 production (Fig. 1). The maximum rate of N20production in soils without lime was 0.064 ,ug of N20-N perg of soil per h.
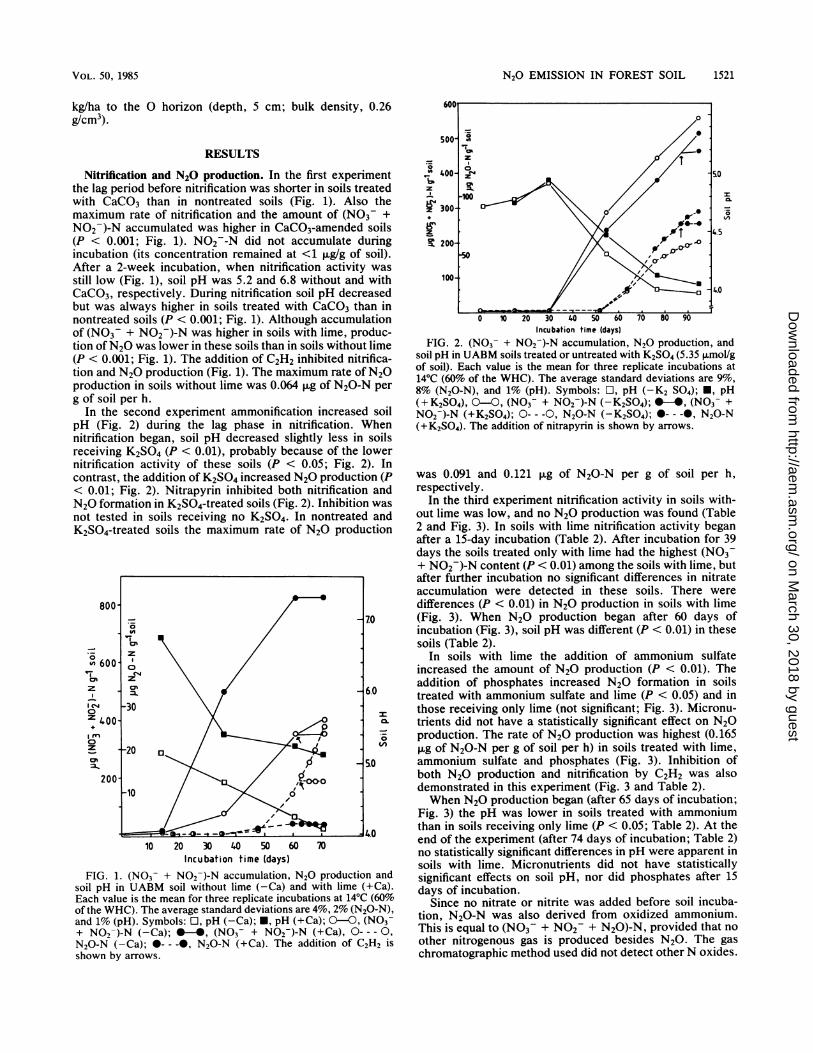
In the second experiment ammonification increased soilpH (Fig. 2) during the lag phase in nitrification. Whennitrification began, soil pH decreased slightly less in soilsreceiving K2SO4 (P < 0.01), probably because of the lowernitrification activity of these soils (P < 0.05; Fig. 2). Incontrast, the addition of K2SO4 increased N20 production (P< 0.01; Fig. 2). Nitrapyrin inhibited both nitrification andN20 formation in K2SO4-treated soils (Fig. 2). Inhibition wasnot tested in soils receiving no K2SO4. In nontreated andK2SO4-treated soils the maximum rate of N20 production
80070
z600 5
Z _0X 6.0C- 30
20 2 <4
2400 acod10
soil pH in UABM soil without lime (-Ca) and with lime (+Ca).Each value is the mean for three replicate incubations at 14°C (60%oof the WHC). The average standard deviations are 4%, 2% (N2A-N),and 1% (pH). Symbols: O, pH (-Ca); *, pH (+Ca); 0-0, (NOf-+ N02-)-N (-Ca); *-4, (NO3- + N02-)-N (+Ca), 0- -- 0,N20-N (-Ca); *---, N20-N (+Ca). The addition of C2H2 isshown by arrows.
z0
z
z
Incubation time (days)FIG. 2. (NO3- + NO02)-N accumulation, N20 production, and
soil pH in UABM soils treated or untreated with K2SO4 (5.35 ,umol/gof soil). Each value is the mean for three replicate incubations at14°C (60% of the WHC). The average standard deviations are 9%,8% (N20-N), and 1% (pH). Symbols: O, pH (-K2 S04); *, pH(+K2SO4), 0-0, (NO3- + NO02)-N (-K2SO4); *-*, (NO3- +NO2-)-N (+K2SO4); 0--- 0, N20-N (-K2SO4); 0---0, N20-N(+K2SO4). The addition of nitrapyrin is shown by arrows.
was 0.091 and 0.121 ,ug of N20-N per g of soil per h,respectively.
In the third experiment nitrification activity in soils with-out lime was low, and no N20 production was found (Table2 and Fig. 3). In soils with lime nitrification activity beganafter a 15-day incubation (Table 2). After incubation for 39days the soils treated only with lime had the highest (NO3+ NO02)-N content (P < 0.01) among the soils with lime, butafter further incubation no significant differences in nitrateaccumulation were detected in these soils. There weredifferences (P < 0.01) in N20 production in soils with lime(Fig. 3). When N20 production began after 60 days ofincubation (Fig. 3), soil pH was different (P < 0.01) in thesesoils (Table 2).
In soils with lime the addition of ammonium sulfateincreased the amount of N20 production (P < 0.01). Theaddition of phosphates increased N20 formation in soilstreated with ammonium sulfate and lime (P < 0.05) and inthose receiving only lime (not significant; Fig. 3). Micronu-trients did not have a statistically significant effect on N20production. The rate of N20 production was highest (0.165,ug of N20-N per g of soil per h) in soils treated with lime,ammonium sulfate and phosphates (Fig. 3). Inhibition ofboth N20 production and nitrification by C2H2 was alsodemonstrated in this experiment (Fig. 3 and Table 2).When N20 production began (after 65 days of incubation;
Fig. 3) the pH was lower in soils treated with ammoniumthan in soils receiving only lime (P < 0.05; Table 2). At theend of the experiment (after 74 days of incubation; Table 2)no statistically significant differences in pH were apparent insoils with lime. Micronutrients did not have statisticallysignificant effects on soil pH, nor did phosphates after 15days of incubation.
Since no nitrate or nitrite was added before soil incuba-tion, N20-N was also derived from oxidized ammonium.This is equal to (NO3- + NO2 + N20)-N, provided that noother nitrogenous gas is produced besides N20. The gaschromatographic method used did not detect other N oxides.
VOL. 50, 1985
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
15
14
0,1
z0
zcm
\
(I
9
/31A'A
65 70Incubation time (days)
FIG. 3. Effects of calcium carbonate (Ca), ammonium s(N), sodium phosphates (P), and micronutrients (M) on N2Cduction in urea-fertilized soils. For application rates, see TaEach value is the mean for three replicate incubations at 14WCof the WHC). The average standard deviation is 22%. SynM- - M, no additions; O-E, Ca; -M, Ca + P; 0--- -0, Ca0-*, Ca + N + P; 0-0, Ca + N + P + M. The addition ofis shown by an arrow.
Also NO2- was not found. The amount of N2O-N prodwas 0.29 to 3.96% of the NH4+-N oxidized (Table 2).N20 production and soil pH. The results of the
experiments described above suggest that N2O productienhanced by low soil pH. The ratio of N20-N evolvlNH4+-N oxidized (see above) was plotted against soil p:different incubation periods of the three experiments.ratio ofN2O-N evolved to NH4+-N oxidized was deternbecause of variations in the nitrification rates. AveragmpHs for the measurement period were used. N20 produwas negligible above soil pH 4.7 (Fig. 4). The ratio of Nto NH4+-N oxidized increased exponentially with a deciin the soil pH from 4.7 to 4.1 (Fig. 4). Soils receiving Kwere excluded from this analysis. The addition of Kenhanced N2O production, even though K2SO4-treatedhad a higher pH (Fig. 2).Chemical production of N20 from added NO2-. OI
addition of NO2-, N2O production was manifest inunfertilized and UABM-fertilized soils (Fig. 5). Soibelow 4.7 had negligible effects on N20-N productionNO2--N concentration of 50 ,ug/g of soil but enhanceda concentration of 200 ,ug of NO2--N per g of soil (Fig. '
6). No N2O was produced from added NO3-. The amou
N2O-N produced from added NO2--N during the experiment(Fig. 5) ranged from 0.63 to 2.05% and 2.55 to 4.58% when 50and 200 ,ug of NO2--N per g of soil, respectively, wereadded. Maximum rates of N20-N production occurred im-mediately after the addition ofNO2 (Fig. 5), and these weremuch higher than the maximum rates in experiments inwhich N2O was produced during nitrification (see above).The rates of chemical N20 production decreased over theexperimental period (Fig. 5). After incubation the amount ofNO2-N remained below 0.05 ,ug/g of soil.
DISCUSSION
The results of this study show that N2O may form innitrifying forest humus under well-aerated conditions. N2Oproduction was closely associated with autotrophic NH4'oxidation and was enhanced by low soil pH. These resultssupport the observations of Robertson and Tiedje (30), whofound that in some forest soils N20 might not be produced bydenitrifiers. They have found that in some intact forest soilcores N20 is produced only in the absence of C2H2. It isinteresting that these soils had lower pHs than those thatproduced N20 only in the presence of C2H2 (4.6 versus 5.2).Goodroad and Keeney (12) have found that a coniferousforest site produces more N2O in situ than does a deciduoussite. Soil pH was lower in the coniferous site (4.5 versus 5.0).The effects of nutrients on N20 production must be
considered in terms of nutrient effects on nitrification andsoil pH. A low soil pH favored N20 production (Fig. 4). InK2SO4-treated soils N20 production and soil pH werehigher, but (NO3- + NO2)-N accumulation was lower thanthat in nontreated soils (Fig. 2). These results indicate that
75 K2SO4 has a stimulative salt effect on N20 production.(NH4)2SO4 also increased N20-N production (Fig. 3 andTable 2), possibly by decreasing soil pH and not by stimu-
ulfate lating nitrification (Table 2; see above). It has already beenpler2- found that the addition of K2SO4 and (NH4)2SO4 to acid soils
-(60% inhibits (NO3- + NO2-)-N accumulation (19). In this study,nbols: sodium phosphates had no statistically significant effect on+ N, soil pH and nitrification during N2O production (see above).C2H2 The slight increase in N20 production because of phosphates
might thus be a salt or nutrient effect.N2O production by NH4+ oxidizers may vary with cultural
luced
threeion ised toH forThe
ninede soilction20-Nrease,2SO4
soils
n theboth
11 pHat a
i it at5 andint of
zI
0
z
mEN
01-
z0
z
25~
z 20_a
40 4.
zS\
,10 _r,z \zO e
O\
40 .S 5
Soil pH
4.0 4.5 5.0Soil pH
55 60
FIG. 4. Percentage production of N20-N from NH4+-N oxidizedat pH 6.1 to 4.1. The values were calculated by the results shown inFig. 1 (-), Fig. 2 (0), Fig. 3 (l), and Table 2 (O). The insert showsthe percentage of production of N20-N from NH4+-N oxidized atpH 4.6 to 4.1. In y = -2.387 x + 11.184, y = log% N20-N fromNH4+-N oxidized, x = soil pH, r = -0.985 (P < 0.001, df = 7).
1522 MARTIKAINEN
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

N20 EMISSION IN FOREST SOIL 1523
0
6
zIU
3
2 0~~~~~~~~~~~~~~1~~~~~~~~~~~~~~~~~~~00
10 20 30 40 50 10 20 30 40 50 10 20 30 40 50Incubation time (hours)
FIG. 5. Cumulative production of N20-N from NO,--N in unfertilized (0, 0) and UABM-fertilized (I, *) soils after different additionsof H+ as H2SO4. Open symbols, 50 ,ug of N02--N per g of soil; closed symbols, 200 ,ug of NO2--N per g of soil. Added NO3--N (50 ,ug ofN per g of soil) produced no N20. Each treatment had two replicates (means are shown). Incubations were at 14WC (60% of the WHC).
conditions, because it is dependent on the buffer systemsused (16, 41) and on cell age. Old cells produce more N20than young ones (41). This observation may be of someimportance in this study, in which N20 production occurredat the end of the experiments when most cells were aged.However, at or above soil pH 4.7, N20 production wasnegligible even after a long incubation. Thus, low soil pH isessential to N20 production.
Autotrophic NH4' oxidizers have been reported to pro-duce N20 in both oxidative and reductive processes (16, 29).It has been suggested that N20 evolves from a labileintermediate between NH2OH and NO2- (possibly N202H2)during NH4+ oxidation (1, 29). A low oxygen level supports
2.5k
.a_
z
'NI
08
2.0
1.5h
1.01
0.51-
2.0
-
3.0 4.0Soil pH
FIG. 6. Maximum rates of chemical N2O-N formation fromadded NO2--N at different soil pH calculated by the results shownin Fig. 5. Symbols as described in the legend to Fig. 5.
N20 formation (13, 16, 17), indicating participation of areductive enzyme system. Recently, Poth and Focht (27)have shown that Nitrosomonas europaea produces N20mainly by NO2 reduction (nitrifier denitrification) and havepointed out that simultaneous NH4+ oxidation is needed totransfer electrons from NH4' to NO2- (NO3- was notreduced). This coupling of NO2- reduction to oxidativeprocesses can explain why C2H2, which inhibits the first stepin NH4' oxidation, has been reported to inhibit N20 pro-duction from NH4' in the presence of NO2- (16). C2H2 doesnot inhibit NH2OH oxidation (15, 16), which explains why itdoes not inhibit N20 production from NH2OH (16).The observations from this study that no N20 is produced
on the addition of inhibitors, even when the soil containsNO3- for possible denitrification by heterotrophic bacteria,support nitrifier denitrification. However, the favorable ef-fects of low soil pH and salts cannot be unambiguouslyexplained by these results. Enhanced N20 production at alow pH was not detected in studies with pure cultures ofNitrosomonas europaea, but sterile soil was found to greatlystimulate this process (16). It is not known what effect lowpH or salts have on N20 production by Nitrosospira spp.,which probably is responsible for NH4' oxidation in fertil-ized forest soil (20). It is possible that Nitrosospira enzymesthat catalyze the oxidation of the intermediate betweenNH2OH and NO2 are sensitive to low pH and some salts athigh concentrations. If the intermediate (N202H2) accumu-lates in cells, N20 may be formed during hydrolysis ofN202H2 (29). The labile intermediate theory is not supportedby the results of Poth and Focht (27). They have pointed outthat the use of NO2- as a terminal electron acceptor con-serves 02 for the first step ofNH4+ oxidation. This oxidationneeds molecular oxygen. Nitrite, which is a toxic endproduct, can also be removed by its reduction to N20. In thisstudy the oxygen hypothesis did not fit well into the ob-served N20 production pattern. For example, in the firstexperiment (Fig. 1), oxygen depletion probably was greaterin soils with lime because of their higher level of nitrification
VOL. 50, 1985
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1524 MARTIKAINEN
(and respiration of heterotrophic microbes?), and N20 for-mation thus must be higher in soils with lime than in soilswithout lime. It is more likely that under the extremeconditions caused by low soil pH (or salts) Nitrosospira spp.are sensitive to N02 and will eliminate N02 by reducing itto N20.
Besides direct evolution from cells of NH4' oxidizers,other mechanisms also exist for the production of N20 inaerated soils. Biological denitrification in anaerobicmicrosites, even during incubation of nonwaterlogged soil inan aerobic atmosphere, may occur. This possibility couldnot be totally excluded in this study because nonsterile soilwas used. However, when nitrification was inhibited bynitrapyrin or C2H2, N20 production ceased, although the soilstill contained NO3. It was to be expected that NO3 woulddiffuse from soil solution into anaerobic microsites. Thepossibility that both C2H2 and nitrapyrin inhibit denitrifica-tion is unlikely. It has been shown that the addition of 50 pugof nitrapyrin per g of soil does not inhibit denitrification (14).One possible explanation is that denitrifiers in the micrositesuse N02 but not NO3 as electron acceptor (37, 39). A Kmvalue as low as 0.20 mg of 02 per liter at 27°C has been foundfor Nitrosospira sp. in forest soil (20). It is possible thatN02 is produced by NH4+ oxidizers in a microenvironmentand that part of it is reduced to N20 by denitrifiers in thesame niche. Denitrification is known to occur at 02 concen-trations of 0.1 to 0.2 mg/liter in sewage, ocean, and purecultures (11). Some denitrifiers can use both 02 and NO2 aselectron acceptors simultaneously (39). Pang and Cho (25)recently have reported that biological denitrification ofadded NO2- can occur in the L-H layer of coniferous forestsoil, even under aerobic conditions.
In soil, N20 is also produced from NO2- by chemical
denitrification (9). High organic matter content and low soilpH favor this process (22, 23); forest soil may thus be a goodenvironment for chemical denitrification. In this study theamount of N20-N was 0.6 to 4.6% of the amount of N02 -Nadded. This agrees with observations made with other typesof soil (4, 23, 28, 32). In this study a high concentration ofN02 at low soil pH enhanced chemical N20 formation. Therates of chemical N20 formation immediately after theaddition of NO2- were much higher than those of N20formation during nitrification, but when NO2- concentra-tions had decreased (final concentrations, <0.05 jig ofN per
g of soil) the rates were of the same magnitude.In the second experiment (Fig. 2), chemical N20 forma-
tion could not explain the rate of N20 production. In soilsthat received K2SO4, N20-N and NO3-N productions av-
eraged 2.2 and 7.0 ,ug of N per g of soil per day, respectively.The amount of N20-N produced from NO2-N did notexceed 5% (see above). As estimated by the amounts of N20and NO3 formed, the average rate of N02 -N productionshould be at least (100/5 x 2.2) + 7.0 = 51 ,ug of N per g ofsoil per day, to provide N02 for chemical denitrificationand nitrite oxidation during the whole 40-day period. Over2,000 ,ug of NH4+-N thus must be oxidized during thisperiod. In some K2SO4-treated soils ammonium oxidationand N20 production were inhibited by nitrapyrin during the95 days of incubation (data not shown in Fig. 2). Theammonium accumulation rates determined for these soilsshowed that the highest possible amount of NH4+-N oxida-tion was about 600 pg of NH4+-N per g of soil. Furthermore,after 95 days of incubation the cumulative amount of (NH4++ NO3- + NO2- + N20)-N was higher in soils withoutnitrapyrin than in soils with nitrapyrin (673 and 609 jig of Nper g of soil, respectively). With a high chemical N20
production, the amount of N should be lower in the formerthan in the latter soils because N20 is a minor component ofthe nitrogenous gases that evolve during chemicaldenitrification (4, 23, 28, 35). It must also be mentioned thatthe reduction of NO - to N02- could not provide enoughN02- to make the observed N20 formation a chemicalprocess. So, in these soils, the chemical N20 formationseems to be of relatively limited importance in the total N20flux.The long-term laboratory incubations in this study indicate
a potential for aerobic N20 production in nitrifying acid soil.A comparison of this potential with the denitrification poten-tial in the UABM soil with anaerobic incubations and anoptimum NO3- level (38) showed that the maximum aerobicN20-N production (0.17 Lg/g of soil per h) was about 50% ofthe denitrification potential. The production ofN20 by forestsoil Nitrosospira spp. must be examined at various condi-tions for better understanding of the mechanisms of N20production. Additional work must also be done to evaluateincreases in in situ evolution of N20 in terms of fertilization,which stimulates nitrification in acid forest soil. Evidencefrom other types of soil with higher pHs indicated thatammonium or urea fertilization increases N20 emission insitu and that this N20 is produced by ammonium oxidizers(5, 9). Urea fertilization is likely to increase N20 evolutionfrom coniferous forest soil because urea can stimulate nitri-fication for many years (18). It recently has been discussedhow lime applied to forest soil to neutralize acid depositionaffects tree growth (26) and soil microbes (24, 36). It isknown that the addition of lime can stimulate nitrification inacid forest soil (24). The results of this study indicate that theaddition of lime may be environmentally harmful because ofan increase in N20 evolution, especially when nitrification isstimulated by lime but the soil pH remains below 5.
LITERATURE CITED1. Anderson, J. H. 1964. The metabolism of hydroxylamine to
nitrite by Nitrosomonas. Biochem. J. 91:8-17.2. Aulakh, M. S., D. A. Rennie, and E. A. Paul. 1984. Acetylene
and N-serve effects upon N20 emission from NH4' and NO3treated soils under aerobic and anaerobic conditions. Soil Biol.Biochem. 16:351-356.
3. Blackmer, A. M., J. M. Bremner, and E. L. Schmidt. 1980.Production of nitrous oxide by ammonia-oxidizing chemoauto-trophic microorganisms in soil. Appl. Environ. Microbiol.40:1060-1066.
4. Bollag, J.-M., S. Drzymala, and L. T. Kardos. 1973. Biologicalversus chemical nitrite decomposition in soil. Soil Sci.116:44-50.
5. Breitenbeck, G. A., A. M. Blackmer, and J. M. Bremner. 1980.Effects of different nitrogen fertilizers on emission of nitrousoxide from soil. Geophys. Res. Lett. 7:85-88.
6. Bremner, J. M. 1965. Inorganic forms of nitrogen, p. 1179-1237.In C. A. Black (ed.), Methods of soil analysis, part II. AmericanSociety of Agronomy, Madison, Wis.
7. Bremner, J. M. 1965. Total nitrogen, p. 1149-1178. In C. A.Black (ed.), Methods of soil analysis, part II. American Societyof Agronomy, Madison, Wis.
8. Bremner, J. M., and A. M. Blackmer. 1979. Effects of acetyleneand soil water content on emission of nitrous oxide from soils.Nature (London) 280:380-381.
9. Chalk, P. M., and C. J. Smith. 1983. Chemodenitrification, p.65-89. In J. R. Freney and J. R. Simson (ed.), Gaseous loss ofnitrogen. Martinus Nijhoff/Dr W. Junk Publishers, The Hague.
10. Crutzen, P. J., and D. W. Ehalt. 1977. Effects of nitrogenfertilizers and combustion on the stratospheric ozone layer.Ambio 6:112-117.
11. Focht, D. D., and W. Verstraete. 1977. Biochemical ecology ofnitrification and denitrification. Adv. Microb. Ecol. 1:135-214.
APPL. ENVIRON. MICROBIOL.
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from

N20 EMISSION IN FOREST SOIL 1525
12. Goodroad, L. L., and D. R. Keeney. 1984. Nitrous oxideemission from marsh and prairie ecosystems. J. Environ. Qual.13:448-452.
13. Goreau, T. J., W. A. Kaplan, S. C. Wofsy, M. B. McElroy,F. W. Valois, and S. W. Watson. 1980. Production of NO2- andN20 by nitrifying bacteria at reduced concentration of oxygen.Appl. Environ. Microbiol. 40:526-532.
14. Henninger, N. M., and J.-M. Bollag. 1976. Effect of chemicalsused as nitrification inhibitors on the denitrification process.Can. J. Microbiol. 22:668-672.
15. Hynes, R. K., and R. Knowles. 1982. Effect of acetylene onautotrophic and heterotrophic nitrification. Can. J. Microbiol.28:334-340.
16. Hynes, R. K., and R. Knowles. 1984. Production of nitrous oxideby Nitrosomonas europaea: effects of acetylene, pH, andoxygen. Can. J. Microbiol. 30:1397-1404.
17. Lipschultz, F., 0. C. Zafiriou, S. C. Wofsy, M. B. McElroy,F. W. Valois, and S. W. Watson. 1981. Production of NO andN20 by soil nitrifying bacteria. Nature (London) 294:641-643.
18. Martikainen, P. J. 1984. Nitrification in two coniferous forestsoils after different fertilization treatments. Soil Biol. Biochem.16:577-582.
19. Martikainen, P. J. 1985. Nitrification in forest soil of differentpH as affected by urea, ammonium sulphate and potassiumsulphate. Soil Biol. Biochem. 17:363-367.
20. Martikainen, P. J., and E.-L. Nurmiaho-Lassila. 1985.Nitrosospira, an important ammonium-oxidizing bacterium infertilized coniferous forest soil. Can. J. Microbiol. 31:190-197.
21. Montgomery, H. A. C., and J. F. Dymock. 1961. The determi-nation of nitrite in water. Analyst 86:414-416.
22. Nelson, D. W., and J. M. Bremner. 1969. Factors affectingchemical transformations of nitrite in soils. Soil Biol. Biochem.1:229-239.
23. Nelson, D. W., and J. M. Bremner. 1970. Gaseous products ofnitrite decomposition in soils. Soil Biol. Biochem. 2:203-215.
24. NihlgArd, B., and B. Popovic. 1984. Effects of different limingagencies in forest-a literature review. Statens NatursvardsverkPM 1851, p. 1-85. Statens NaturvArdsverk, Information-senheten, Solna, Sweden.
25. Pang, P. C.-K., and C. M. Cho. 1984. Oxygen consumption anddenitrification activity of a conifer forest soil profile. Soil Sci.Soc. Am. J. 48:393-399.
26. Popovic, B., and F. Andersson. 1984. Liming and forest produc-tion-literature review and revision of Swedish liming experi-ments. Statens Naturv&rdsverk PM 1792, p. 1-107. StatensNaturvardsverk, Informationsenheten, Solna, Sweden.
27. Poth, M., and D. D. Focht. 1985. "5N kinetic analysis of N20
production by Nitrosomonas europaea: an examination ofnitrifier denitrification. Appl. Environ. Microbiol. 49:1134-1141.
28. Reuss, J. O., and R. L. Smith. 1965. Chemical reactions ofnitrites in acid soils. Soil Sci. Soc. Am. Proc. 29:267-270.
29. Ritchie, G. A. F., and D. J. D. Nicholas. 1972. Identification ofthe sources of nitrous oxide produced by oxidative and reduc-tive processes in Nitrosomonas europaea. Biochem. J.126:1181-1191.
30. Robertson, G. P., and J. M. Tiedje. 1984. Denitrification andnitrous oxide production in successional and old-growth Mich-igan forests. Soil Sci. Soc. Am. J. 48:383-389.
31. Shattuck, G. E., and M. Alexander. 1963. A differential inhibitorof nitrifying microorganisms. Soil Sci. Soc. Am. Proc. 27:600-601.
32. Smith, C. J., and P. M. Chalk. 1980. Fixation and loss ofnitrogen during transformations of nitrite in soils. Soil Sci. Soc.Am. J. 44:288-291.
33. Smith, C. J., and P. M. Chalk. 1980. Gaseous nitrogen evolutionduring nitrification of ammonia fertilizer and nitrite transforma-tions in soils. Soil Sci. Soc. Am. J. 44:277-282.
34. Sokal, R. R., and F. J. Rohlf. 1981. Biometry, 2nd ed. W. H.Freeman and Co., San Francisco.
35. Stevenson, F. J., R. M. Harrison, R. Wetselaar, and R. A.Leeper. 1970. Nitrosation of soil organic matter. III. Nature ofgases produced by reaction of nitrite with lignins, humic sub-stances, and phenolic constituents under neutral and slightlyacidic conditions. Soil Sci. Soc. Am. Proc. 34:430-435.
36. Soderstrom, B. 1984. Some microbial data on acidification andliming of forest soils-a literature review. StatensNaturvfardsverk PM 1860, p. 1-15. Statens Naturvairdsverk,Informationsenheten, Solna, Sweden.
37. Tiedje, J. M. 1982. Denitrification, p. 1011-1026. In A. L. Page(ed.), Methods of soil analysis, part II. American Society ofAgronomy, Madison, Wis.
38. Uomala, P., P. Martikainen, L. Skold, and K. Kari. 1982. Effectof fertilization on forest soil denitrification potential, p. 329-336.In The Second National Symposium on Biological NitrogenFixation, Helsinki, 8-10 June 1982. The Finnish National Fundfor Research and Development.
39. Vangnai, S., and D. A. Klein. 1974. A study of nitrite-dependentdissimilatory micro-organisms isolated from Oregon soils. SoilBiol. Biochem. 6:335-339.
40. Weiss, R. F. 1981. The temporal and spatial distribution oftropospheric nitrous oxide. J. Geophys. Res. 86:7185-7195.
41. Yoshida, T., and M. Alexander. 1970. Nitrous oxide formationby Nitrosomonas europaea and heterotrophic microbes. SoilSci. Soc. Am. Proc. 34:880-882.
VOL. 50, 1985
on March 30, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















