
Nikki van Sante 11673435 Supervisor: Timo Stein
30
1 Testing Unconscious Face Processing and its Specificity to Conspecifics Nikki van Sante 11673435 Supervisor: Timo Stein January 29, 2021
Transcript of Nikki van Sante 11673435 Supervisor: Timo Stein

1
Testing Unconscious Face Processing and its Specificity to Conspecifics
Nikki van Sante
11673435
Supervisor: Timo Stein
January 29, 2021

2
Abstract
Unconscious processing is studied extensively with detection paradigms, such as breaking
Continuous Flash Suppression (b-CFS), where a detection difference between conditions is
assumed to be caused by differential unconscious processing preceding detection. One effect
consistently found with this approach is that faces are detected faster in an upright orientation
compared to an inverted orientation. In a b-CFS study by Stein et al. (2012) with human
participants, this inversion effect was present for human faces as well as for chimpanzee
faces, with the inversion effect for human faces being stronger. However, as b-CFS has
recently been criticized for not being able to distinguish between unconscious and conscious
factors underlying a detection difference, it is not yet clear whether the face inversion effect
occurs unconsciously. Therefore, the current study aims to replicate these previous findings
by using a novel paradigm called detection-discrimination dissociation in combination with
backward masking. This method uses an additional discrimination task to exclude conscious
processes as a cause for a detection difference. Results revealed some evidence that human
participants processed the orientation of human faces unconsciously. However, other findings
indicated that this cannot be concluded with certainty. Further, no inversion effect was
present for chimpanzee faces. These findings shed light on previous b-CFS studies and
highlight the importance of using the novel detection-discrimination dissociation paradigm in
order to determine the scope and limits of unconscious processing.
Keywords: unconscious processing, faces, inversion, conspecifics

3
Testing Unconscious Face Processing and its Specificity to Conspecifics
Does the brain process information outside our awareness? And to what degree does
unconscious processing influence our behaviour? The phenomenon of blindsight clearly
demonstrates that not only conscious information influences actions. Blindsight can be
observed in patients with damage to primary visual cortex (Sanders et al., 1974; Weiskrantz,
1996). These patients have no conscious experience of seeing anything. However, when the
patients are forced to guess what they are looking at, they can accurately name objects
(Trevethan et al., 2007). Furthermore, these patients can accurately reach for objects or avoid
obstacles (de Gelder et al., 2008; Prentiss et al., 2018). This demonstrates that unconscious
processing plays an essential part in guiding behaviour in blindsight patients, which
challenged scientists to discover more about the role of unconscious processing in healthy
individuals.
However, in order to study unconscious processing in healthy participants, it is
required that stimuli can be made invisible. The rise of different experimental manipulations
made this possible. In masking paradigms, a visual stimulus is presented for a short duration
and afterwards another image, called the mask, is presented that renders the stimulus invisible
(Marcel, 1983). When the mask is not presented after the stimulus, participants will almost
always see the stimulus. Thus, by only minimally changing the task, a completely different
experience is induced, namely seeing a stimulus or being completely unaware of it. Another
experimental manipulation is interocular suppression, where a stimulus presented to one eye
is suppressed from awareness by another stimulus presented to the other eye (Izatt et al.,
2014; Lin & He, 2009). The two stimuli compete for access to awareness, such that only one
stimulus can be consciously perceived at the same time. Without changing the visual input,
the perception differs between seeing one stimulus or seeing another. These manipulations
made it possible to let all types of images and words vanish from sight and study unconscious

4
processing experimentally. Consequently, researchers began to investigate the scope and
limits of unconscious processing.
One of the main approaches to study unconscious processing is to measure the time it
takes for a stimulus to be detected. In these detection paradigms, a detection difference
between stimuli is assumed to be caused by differential unconscious processing preceding
detection. However, one problem with this approach is that low-level stimulus properties
could be responsible for a detection difference between stimuli (Pournaghdali & Schwartz,
2020). A solution to this problem is inversion, where a stimulus in an upright orientation is
compared with the same stimulus in an inverted orientation. In this way, low-level properties
of both stimuli are exactly the same. One effect that is consistently found with this approach
is that faces presented in an upright orientation are detected easier than inverted faces (Jiang
et al., 2007; Stein et al., 2012, 2016; Zhou et al., 2010). In other words, upright face stimuli
break faster into awareness compared to inverted faces. However, this inversion effect is not
found for houses (Albonico et al., 2018) or lamps (Stein et al., 2012). This led some
researchers believe that face processing is special compared to the processing of other stimuli
and that there is a specific neural circuitry for the processing of faces (Haxby et al., 2000).
This hypothesis implies that there is an inborn mechanism in the brain for processing faces.
However, inversion effects are also found for bodies (Reed et al., 2003) and objects of
expertise (Stein et al., 2016). For example, greater car expertise was related to larger
inversion effects (Stein et al., 2016). The expertise hypothesis states that face processing only
seems special because people have a lot of experience with face stimuli (McKone et al.,
2007).
Up to date, it is still a question whether people are born with a face processing module
or whether rapid face detection is accomplished because of our experience with these stimuli.
One interesting approach for resolving this debate is to study looking preferences in infants.

5
Newborn infants already have a preference for faces presented upright compared to inverted
(Johnson et al., 1991). This inborn face preference is probably mediated by subcortical
circuits, since cortical regions are not yet well developed in infancy (Johnson, 1990). Such an
inborn mechanism is thought to make fast detection of faces possible without conscious
awareness (Stein et al., 2011). Further, research found that there is a lot of overlap between
facial properties that attract infants’ gaze and face properties that influence the access to
awareness in adults (Stein et al., 2011). Thus, it seems that face detection is at least partly
mediated by inborn mechanisms. In order to study how experience also plays a role in face
detection it is interesting to look at the processing of faces of different species. One study
found that newborns cannot discriminate human faces from monkey faces, while they
preferred to look at monkey faces with an upright orientation compared to an inverted
orientation (Di Giorgio et al., 2012). This indicates that a face inversion effect is already
present at birth, but that is not yet specific to conspecifics. Based on these findings, it could
be hypothesized that there is an inborn face detection pathway operating unconsciously which
is not specific to conspecifics and is mediated by subcortical circuits. In contrast to infants,
adults show a conspecific advantage in attending to changes in human faces compared to
monkey faces and also visual search is faster for human faces (Neiworth et al., 2006;
Simpson et al., 2014). These findings could be a result of gaining experience with conspecific
faces. The influence of experience on face detection is hypothesized to be mediated by
cortical circuits and is thought to involve awareness (Jessen & Grossmann, 2015; Johnson,
2005). Therefore, it seems plausible that there is also an experience-dependent face detection
pathway mediated by cortical circuits which requires consciousness and is specific to
conspecifics. However, it is unclear how these two pathways influence face detection, as
inborn and experience-based factors to detection are often not dissociable. Further, conscious

6
and unconscious contributions to face detection are difficult to distinguish in detection
paradigms.
Currently, it is still unclear if the face inversion effect in detection is modulated by
conscious or unconscious factors. Previous detection studies about the face inversion effect
mainly use a method called breaking Continuous Flash Suppression (b-CFS). This is a strong
form of interocular suppression where participants are presented with a stimulus of interest to
one eye and the other eye is presented with a dynamic mask of high contrast which will
dominate vision for up to several seconds (Tsuchiya & Koch, 2005). At a given moment, the
stimulus will break through into awareness and becomes visible. The time it takes for a
stimulus to break through is thought to reflect unconscious processing prior to the conscious
experience (Del Río et al., 2018). According to this assumption, the finding that upright face
stimuli break through earlier than inverted face stimuli would indicate that there is enhanced
unconscious processing prior to the detection of upright faces. However, the assumption that
unconscious processing underlies detection differences during b-CFS has recently been
criticized (Moors et al., 2019; Stein & Sterzer, 2014). It has been argued that not only
unconscious factors but also conscious factors can play a role in the detection difference
between upright and inverted faces. For instance, later conscious factors related to
identification or recognition of the stimulus can cause the difference in detection times (Stein,
2019). Also, response biases can play a role in the detection of stimuli (Stein & Peelen,
2021). There are large individual differences in how inclined people are to say that they saw
something. Some people need to be entirely sure that they saw the stimulus in order to
indicate that they saw it, while others already state they saw the stimulus even if they are not
entirely sure. A third problem with inferring that unconscious processes underlie inversion
effects in detection is that it is automatically assumed that consciousness is discrete and that
participants can accurately state whether they saw a stimulus or not. However, it is highly

7
debated whether consciousness is really discrete and some studies argue that it can be better
explained as a continuous phenomenon (Fekete et al., 2018; Srinivasan, 2020; White, 2018).
Therefore, interpreting detection differences as a result of greater unconscious processing
may lead to an overestimation of unconscious processing.
These alternative explanations for inversion effects during b-CFS emphasize the
importance for developing a new, valid method that can dissociate conscious from
unconscious processing underlying detection. This would make it possible to investigate
whether the face inversion effect occurs unconsciously or whether it is only mediated by
conscious factors. One method that has recently been proposed as overcoming the problems
of b-CFS is the detection-discrimination dissociation paradigm (Stein & Peelen, 2021). This
paradigm consists of two tasks: detecting the location of the stimulus and discriminating the
stimulus on the critical stimulus dimension that underlies the detection difference. In case of
the inversion effect, this would mean that participants need to discriminate whether the face
is inverted or upright. If participants are unable to discriminate the orientation of a stimulus,
while there is still a detection difference between upright and inverted faces, this is probably
due to differential unconscious processing. This detection-discrimination dissociation method
can be used in combination with b-CFS, but it is important to find a condition where
participants perform at chance on the discrimination task and above chance on the detection
task. Because of large individual differences in breakthrough time, the b-CFS method is not
preferable (Gayet & Stein, 2017). Therefore, the current study will use the detection-
discrimination dissociation paradigm in combination with backward masking, as individual
differences are smaller and presentations times can be easily adjusted to obtain the optimal
condition.
A b-CFS study by Stein et al. (2012) found that human participants were faster in
detecting upright human faces compared to inverted human faces and that chimpanzee faces

8
were also detected faster in an upright orientation compared to an inverted orientation. The
inversion effect was larger for human faces, which indicates a conspecific advantage in face
detection. Because of the problems with the b-CFS method discussed earlier, the current
study will try to replicate the b-CFS findings of Stein et al. (2012) by adopting the detection-
discrimination paradigm in combination with backward masking. Human and chimpanzee
faces will be shown with different presentation times, which can either appear left or right on
the screen and have an upright or inverted orientation. It is predicted that human upright faces
are better localized than inverted faces when discrimination is at chance performance. The
same is predicted for chimpanzee faces. For presentation times where discrimination is at
chance, it is predicted that the size of the inversion effect of chimpanzee faces compared to
human faces does not differ, because it can be hypothesized that innate, unconscious
mechanisms are not specific to conspecifics. For longer presentation times, where
discrimination is above chance, it is predicted that the inversion effect for human faces is
larger compared to the inversion effect for chimpanzee faces, because conscious mechanisms
seem to be influenced by experience. If these predictions are accurate, it would indicate that
face perception can occur unconsciously and that it is not specific to conspecifics, while
conscious face perception is specific to our own species. This will provide some evidence
that there are two distinct face detection pathways that operate together in the detection of
faces: one innate, unconscious, subcortical pathway which is not specific to our own species
and one experience-dependent, conscious, cortical pathway that is specific to our own
species.
Methods
Participants
Participants were recruited via the Behavioural Science Lab subject pool of the University of
Amsterdam and were unaware of the purpose of the study. The sample consisted of 47

9
participants (35 female) and were between 18 and 45 years of age (M = 22.02, SD = 0.64).
Participants received a Psychology Research credit or 10 euros as a compensation for
participating. Inclusion criteria were normal or corrected-to-normal vision and an age of
above 16. Participants were excluded when their overall discrimination performance was
below a d’ of 0.5 at the longest presentation time. Based on this, no participants were
excluded. The study was approved by the Ethics Review Board of the Behavioural Science
Lab. Before the start of the experiment, participants were provided with an information letter
and signed informed consent. Some additional instructions about the set-up of the experiment
were provided by a researcher and participants were told to be as accurate as possible.
Practice trials with feedback were present in the experiment to make sure participants
understood the experiment.
Materials and stimuli
A 24-inch LCD monitor (1920 x 1080 pixels resolution) with a refresh rate of 120 Hz
was used to administer the experiment. All participants were seated in front of the screen at a
viewing distance of approximately 60 cm. The task was coded in MATLAB using
Psychtoolbox (Brainard, 1997). During the experiment six different frontal human faces and
six different frontal chimpanzee faces were used. The human face stimuli were collected via
the database of Ekman & Friesen (1976) and the chimpanzee faces were selected from the
internet (as in Stein et al., 2012). Inverted stimuli were created by turning the upright face
stimuli 180°. External facial features of the face stimuli were removed and the size was
adjusted to fit into a square image (100 × 100 pixels). A circular averaging filter was applied
to the outer edges to smooth the facial contours into the background. The contrast of all faces
was the same (SD 18.4) and the stimuli had the same luminance as the background (RGB
values [102 102 102]). Example stimuli are shown in Figure 1. The masks (284 x 284 pixels)
were constructed by creating randomly arranged circles in different shades of grey (diameter

10
23-46 pixels). A total of 100 masks were generated. Stimuli were presented in a grey box
(284 × 284 pixels, RGB values [102 102 102]) in the centre of the screen surrounded by
black.
Figure 1
Example Stimuli
Note: Stimuli were human faces or chimpanzee faces in an upright or inverted orientation.
Design
The experiment used a within-subjects design. The experiment consisted of two
blocks with 384 trials with human faces and 384 trials with chimpanzee faces. Before both
blocks, participants finished 8 practice trials (with the longest presentation time) which
included feedback on their accuracy. Participants alternately viewed the human trials or the
chimpanzee trials first. The order of the localization and discrimination task was
counterbalanced and trials were presented at random. Every combination of four presentation
times, two orientations (upright/inverted), two locations (left/right) and six different face
stimuli occurred four time in each block. In between the trials there were some breaks of 10 s
included to reduce fatigue. The duration of the experiment was approximately 60 minutes.
Procedure
Before the experiment started, participants’ age, gender and handedness was noted.
The experiment began with instructions on the screen. A trial began with a screen were the
box and a fixation cross was presented for 1.5 s. After this, a blank screen was presented for
0.5 s which marked the beginning of the stimulus presentation sequence. Then, a human or
chimpanzee face was shown on the left side or the right side of the box (centre-to-centre

11
distance 71 pixels) in an inverted or upright orientation. The face stimuli were presented with
four presentation times (8.3[8.3], 16.7, 25.0, and 50 ms). After the presentation of face
stimuli at the first presentation time, an additional blank screen was presented for 8.3 ms.
This value is presented in square brackets. Different presentation times were used to increase
the chance of finding a condition where discrimination is at or below chance performance.
The presentation times were determined based on pilot testing. Longer presentation times
were added to keep participants motivated to perform the task. After a face stimulus, three
backward masks were randomly selected and presented for 100 ms each.
Next, participants were asked to indicate whether the face was presented on the left or
the right side of the screen (localization task) and whether the face was in an upright or an
inverted orientation (discrimination task). Participants had to press the left or right arrow bar
on a keyboard to indicate the location of the face. To indicate if a face was presented upright
or inverted they respectively needed to press the arrow bar pointed upwards or the arrow bar
pointed downwards. The order of the localization task and the discrimination task was
counterbalanced between participants. The procedure was based on the study that introduced
the detection-discrimination dissociation paradigm (Stein & Peelen, 2021). An overview of
one trial can be found in Figure 2.
Figure 2
An Overview of a Trial

12
Note. The stimulus could appear on the left or on the right side of the grey box. After the
mask, participants had to perform the localization task first and then the discrimination task
or the other way around.
Analysis
Accuracy on the localization and discrimination task was transformed into SDT
measure d’. This was done separately for human and chimpanzee faces and for each
presentation time. In the localization task, pressing the left arrow bar was classified as a hit in
trials where the face was presented left, while it was classified as a false alarm when the face
was presented right. In the discrimination task, pressing the arrow bar pointing upwards was
classified as a hit in trials with an upright face orientation and was classified as a false alarm
when the face had an inverted orientation. When hit and false alarm rates were 0 or 1, they
were converted to 1/(2N) and 1−1/(2N), respectively, where N represents the number of trials
on which the rates were based (Macmillan & Creelman, 2005). To calculate d’, the z-
transformed false alarm rate was subtracted from the z-transformed hit rate. For the
localization task, d’ was divided by the square root of two, as the localization task is a 2-AFC
task while the discrimination task is a yes/no task (Macmillan & Creelman, 2005).
Discrimination d’ was analysed for different presentation times using a repeated-
measures ANOVA. To analyse localization d’ a repeated-measures ANOVA with the factors
presentation time and orientation was conducted. Further, with one-sample t-tests it was
investigated for different presentation times whether participants discriminated at chance
performance. After this, inversion effects were analysed for interesting presentation times by
conducting one-tailed paired-sample t-tests. If the face inversion effect is independent of
discrimination, we would expect that localization d’ follows the same pattern for trials with
correct vs. incorrect discrimination response. This was analysed by conducting a repeated-
measures ANOVA with the two factors orientation and presentation for correct

13
discrimination trials. After this, the same was done for incorrect discrimination trials. For
these two analysis, participants were included when they had at least five trials per condition
(upright-human, inverted-human, upright-chimpanzee and inverted-chimpanzee) at the three
shortest presentation times with correct and incorrect discrimination response. The longest
presentation time was not used for this analysis because of a ceiling effect. To compare the
inversion effects of human faces and chimpanzee faces, a repeated-measures ANOVA was
conducted with the factors species, orientation and presentation time. Normality was not
tested because it can be assumed that t-tests and ANOVA’s are robust for normality with a
sample size of above 30 (Kwak & Kim, 2017; Pituch & Stevens, 2016). For each ANOVA,
Mauchly’s test for sphericity was performed. Greenhouse-Geisser corrections were applied to
the degrees of freedom, when the assumption of sphericity was violated and corrected p-
values are reported. For all tests a significance level of .05 was used.
All analyses were performed in JASP (JASP Team, 2020) with standard frequentist
statistics and Bayesian statistics. For the Bayesian statistics, default prior scales were used.
BF10 represents the evidence for the alternative hypothesis, while BF01 can be interpreted as
the evidence in favour of the null hypothesis. BF0+ or BF+0 are reported in one-sided tests.
For multi-factorial ANOVAs, the inclusion BF is reported which can be interpreted as the
evidence for all models with a particular effect in comparison to all models without that
effect. The analysis of the current study was based on the study by Stein and Peelen (2021).
Results
Human faces
For human faces, discrimination performance was almost equal for the two shortest
presentation times and then increased, F(1.84, 83.58) = 247.65, p < .001, ηp2 = 0.85; BF10 =
1.67x1057. This is shown in Figure 3. Localization increased with presentation times, F(2.23,
100.47) = 284.61, p < .001, ηp2 = 0.86; BF10 = 1.39x10118. Human upright faces (M = 1.43,

14
SD = 1.13) were localized better than inverted human faces (M = 1.25, SD = 1.12), F(1, 45) =
32.13, p < .001, ηp2 = 0.42; BF10 = 152.41. Further, there was a significant interaction
between orientation and presentation time, F(3, 135) = 5.99, p < .001, ηp2 = 0.12; but BF01 =
2.79. As shown in Figure 3, the inversion effect was larger at the two intermediate
presentation times. There seems to be no inversion effect at the longest presentation time,
probably because performance was at ceiling.
Figure 3
Discrimination, Localization for Upright Faces and Localization for Inverted Faces for
Human and Chimpanzee Faces
Note. Mean localization d’ for upright faces (orange line) and localization d’ for inverted
faces (blue line) for different presentation times (a value in square brackets refers to the
duration of a blank box presented after the face stimulus) for human faces (left) and
chimpanzee faces (right). Error bars represent 95% CIs for the difference between upright
and inverted faces. For comparison, mean discrimination d’ (black line) with 95% CIs is also
shown in both panels.
Next, we looked for the longest presentation time where discrimination was at chance
performance. At the shortest presentation time, discrimination d’ was greater than zero, t(45)

15
= 2.99, p = .002 (one-tailed), Cohen’s d = 0.44; BF+0 = 15.53, while at the second shortest
presentation time discrimination d’ was not significantly different from zero, t(45) = 1.59, p
= .06 (one-tailed), Cohen’s d = 0.23; but BF0+ = 1.05. However, this finding could also be
interpreted as borderline significant (Tshikuka et al., 2016). Further, the Bayes Factor shows
that the null and the alternative hypothesis are almost equally likely. Therefore, it is
inconclusive whether discrimination d’ is at chance or above chance performance at the
second presentation time. At the two longest presentation times, discrimination d’ was above
chance performance, both t(45) ≥ 6.91, p < .001 (one-tailed), d ≥ 1.02; BF+0 > 1.84x106.
Next, it was tested whether an inversion effect was present at the three shortest presentation
times. For the second shortest presentation time, where discrimination was not significantly
different from 0, a significant inversion effect was found, which might indicates an
unconscious origin of this effect, t(45) = 3.07, p = .004, d = 0.45; BF10 = 9.26. This finding is
shown in Figure 4. To demonstrate that unconscious processing underlies the detection
difference between upright and inverted faces it is also important to perform an additional
analysis where the inversion effect is directly compared with the discrimination measure in
the same metric (Meyen et al., 2020; Schmidt & Vorberg, 2006). This analysis showed that
there was no significant difference between the inversion effect at the second presentation
time and discrimination at the second presentation time, t(45) = 1.22, p = .227, Cohen’s d =
0.18; BF01 = 3.11. Therefore, it is not clear whether the inversion effect at the second
presentation time is due to differential unconscious processing. At the shortest presentation
time, there was no significant inversion effect, t(45) = 1.24, p = .221, d = 0.18; BF01 = 3.05.
Further, a large inversion effect was present at the third presentation time, t(45) = 5.23, p
< .001, d = 0.77; BF10 = 4.33x103.
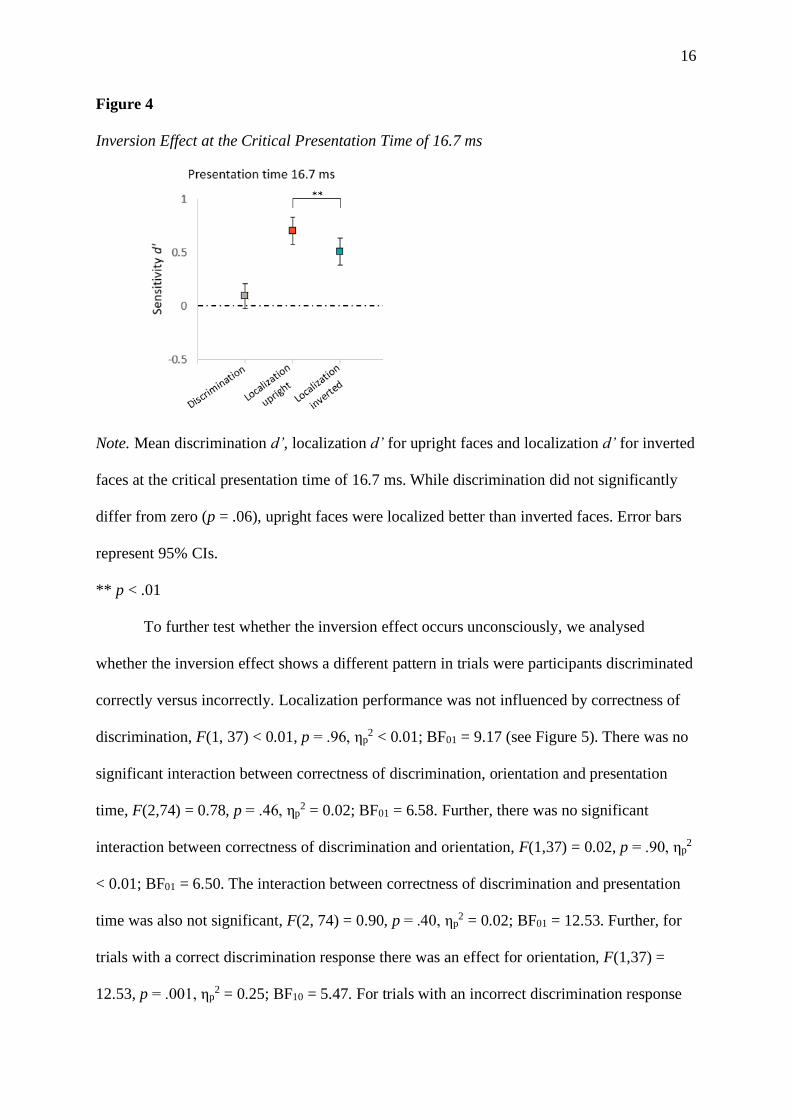
16
Figure 4
Inversion Effect at the Critical Presentation Time of 16.7 ms
Note. Mean discrimination d’, localization d’ for upright faces and localization d’ for inverted
faces at the critical presentation time of 16.7 ms. While discrimination did not significantly
differ from zero (p = .06), upright faces were localized better than inverted faces. Error bars
represent 95% CIs.
** p < .01
To further test whether the inversion effect occurs unconsciously, we analysed
whether the inversion effect shows a different pattern in trials were participants discriminated
correctly versus incorrectly. Localization performance was not influenced by correctness of
discrimination, F(1, 37) < 0.01, p = .96, ηp2 < 0.01; BF01 = 9.17 (see Figure 5). There was no
significant interaction between correctness of discrimination, orientation and presentation
time, F(2,74) = 0.78, p = .46, ηp2 = 0.02; BF01 = 6.58. Further, there was no significant
interaction between correctness of discrimination and orientation, F(1,37) = 0.02, p = .90, ηp2
< 0.01; BF01 = 6.50. The interaction between correctness of discrimination and presentation
time was also not significant, F(2, 74) = 0.90, p = .40, ηp2 = 0.02; BF01 = 12.53. Further, for
trials with a correct discrimination response there was an effect for orientation, F(1,37) =
12.53, p = .001, ηp2 = 0.25; BF10 = 5.47. For trials with an incorrect discrimination response
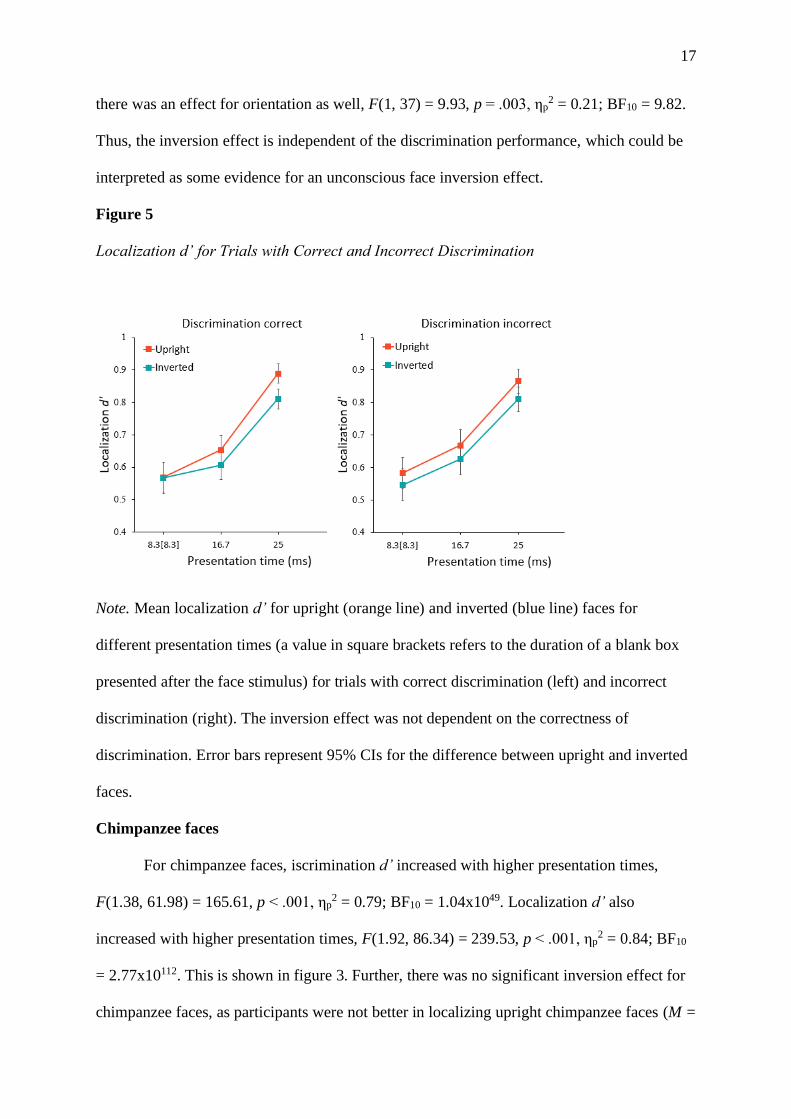
17
there was an effect for orientation as well, F(1, 37) = 9.93, p = .003, ηp2 = 0.21; BF10 = 9.82.
Thus, the inversion effect is independent of the discrimination performance, which could be
interpreted as some evidence for an unconscious face inversion effect.
Figure 5
Localization d’ for Trials with Correct and Incorrect Discrimination
Note. Mean localization d’ for upright (orange line) and inverted (blue line) faces for
different presentation times (a value in square brackets refers to the duration of a blank box
presented after the face stimulus) for trials with correct discrimination (left) and incorrect
discrimination (right). The inversion effect was not dependent on the correctness of
discrimination. Error bars represent 95% CIs for the difference between upright and inverted
faces.
Chimpanzee faces
For chimpanzee faces, iscrimination d’ increased with higher presentation times,
F(1.38, 61.98) = 165.61, p < .001, ηp2 = 0.79; BF10 = 1.04x1049. Localization d’ also
increased with higher presentation times, F(1.92, 86.34) = 239.53, p < .001, ηp2 = 0.84; BF10
= 2.77x10112. This is shown in figure 3. Further, there was no significant inversion effect for
chimpanzee faces, as participants were not better in localizing upright chimpanzee faces (M =

18
1.14, SD = 1.14) compared to inverted chimpanzee faces (M = 1.17, SD = 1.15), F(1, 45) =
1.71, p = .197, ηp2 = 0.04; BF01= 7.55. Moreover, there was no significant interaction
between orientation and presentation time, F(2.43, 109,34) = 0.476, p = .66, ηp2 = 0.01; BF01
= 25.71. As no inversion effect was found, no further analyses were performed.
Human vs. chimpanzee faces
Finally, it was tested whether the inversion effect was larger for human faces
compared to chimpanzee faces. Overall, localization d’ was higher for human faces compared
to chimpanzee faces, F(1,45) = 27.56, p < .001, ηp2 = 0.38; BF10 = 5.33x109. Further, there
was a significant interaction of species by orientation, F(1, 45) = 30.31, p < .001, ηp2 = 0.40;
BF10 = 7.61x1012. In other words, the inversion effect is larger for human faces compared to
chimpanzee faces. Furthermore, there was a significant three-way interaction of species by
orientation by presentation time, F(3, 135) = 3.74, p = .013, ηp2 = 0.08; BF10 = 5.73x1016. To
directly test our hypothesis, we compared the inversion effect at the potentially unconscious
presentation time of 16.7 ms between human and chimpanzee faces. Contrary to our
expectations, the inversion effect at the second presentation time was larger for human faces
compared to chimpanzee faces, t(45) = 3.05, p = .004, Cohen’s d = 0.45; BF10 = 8.91. The
inversion effects for human and chimpanzee faces for different presentation times are shown
in Figure 6.
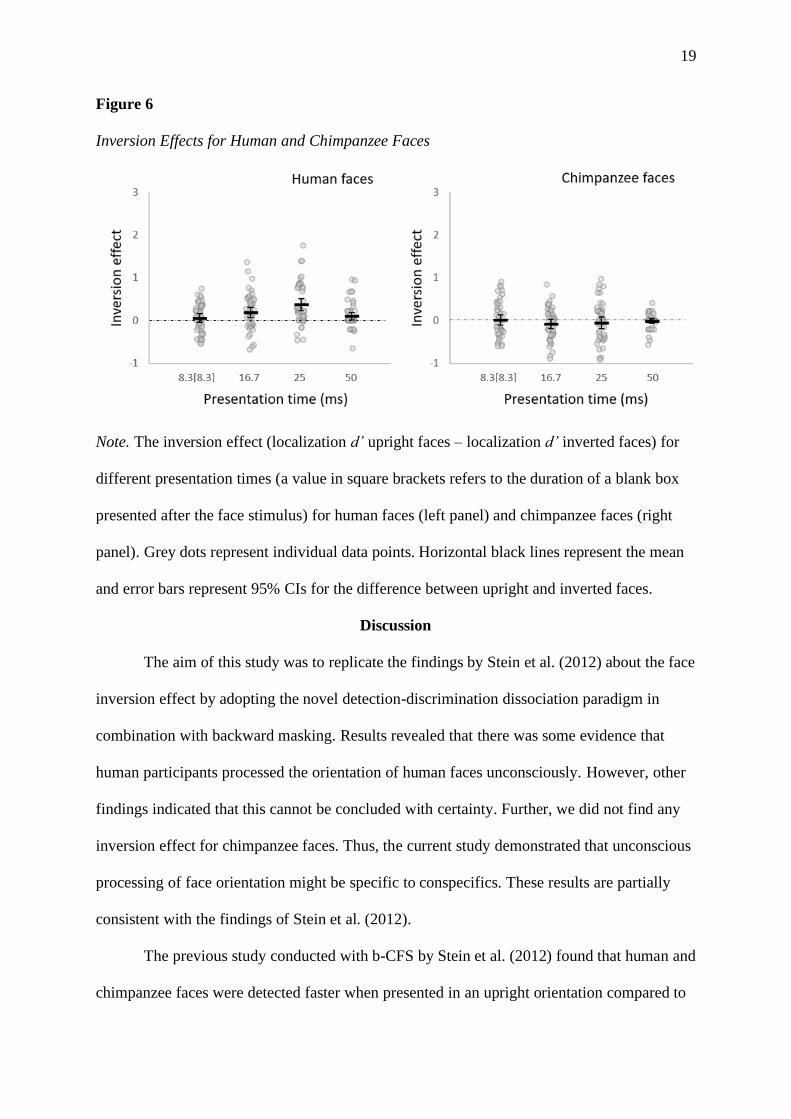
19
Figure 6
Inversion Effects for Human and Chimpanzee Faces
Note. The inversion effect (localization d’ upright faces – localization d’ inverted faces) for
different presentation times (a value in square brackets refers to the duration of a blank box
presented after the face stimulus) for human faces (left panel) and chimpanzee faces (right
panel). Grey dots represent individual data points. Horizontal black lines represent the mean
and error bars represent 95% CIs for the difference between upright and inverted faces.
Discussion
The aim of this study was to replicate the findings by Stein et al. (2012) about the face
inversion effect by adopting the novel detection-discrimination dissociation paradigm in
combination with backward masking. Results revealed that there was some evidence that
human participants processed the orientation of human faces unconsciously. However, other
findings indicated that this cannot be concluded with certainty. Further, we did not find any
inversion effect for chimpanzee faces. Thus, the current study demonstrated that unconscious
processing of face orientation might be specific to conspecifics. These results are partially
consistent with the findings of Stein et al. (2012).
The previous study conducted with b-CFS by Stein et al. (2012) found that human and
chimpanzee faces were detected faster when presented in an upright orientation compared to

20
an inverted orientation. Further, they found that this inversion effect was stronger for human
faces compared to chimpanzee faces. This is inconsistent with the current finding that there is
no inversion effect for chimpanzee faces. One explanation for this is that the previous study
used b-CFS, while the current study used backward masking in combination with the
detection-discrimination dissociation paradigm. Although both paradigms intent to measure
unconscious processing, invisibility is induced differently. Because of this, the underlying
neural mechanisms differ which could explain the different findings between the two
paradigms. Furthermore, b-CFS uses reaction times to measure a detection difference, which
could be influenced by decisional processes, while the current study uses a criterion-free
measure based on accuracy. It is possible that the inversion effect for chimpanzee faces in the
study of Stein et al. (2012) occurred because participants overall pressed a bit later for
inverted faces to indicate detection, just because those faces are less common. Thus, the
inversion effect for chimpanzee faces in the study of Stein et al. (2012) could have been
caused by conscious factors which are ruled out by the detection-discrimination dissociation
paradigm.
From an evolutionary perspective, it seems predictable that an inversion effect is
present for conspecific faces and not for faces of other species. For humans, detecting or
localizing other human faces efficiently is ecologically and socially relevant, for example for
communicating, reading other’s emotions and for connecting with each other. As we see
human faces throughout life almost always in an upright orientation, it is not surprising that
we are better in localizing upright faces compared to inverted ones. However, faces of other
species are not as relevant for humans which could explain why there is no inversion effect
for those stimuli. Support for this idea is provided by a study that showed that chimpanzees
also show an inversion effect for faces of conspecifics but not for human faces (Parr, 2011).

21
A related question is whether this conspecific inversion effect is innate or whether it is
due to experience and what brain circuits play a role. As discussed earlier, one idea is that
there is an innate subcortical face detection pathway that operates unconsciously and is not
specific to conspecifics while there is another experience-dependent cortical face detection
circuit which is specific to conspecifics where awareness is involved (Jessen & Grossmann,
2015; Mark H. Johnson, 2005; Stein et al., 2011). However, this hypothesis is contradictory
with the current results, as we found some evidence that face processing occurs
unconsciously, but that this was indeed specific to faces of our own species. Therefore,
another speculation could be that these unconsciously operating subcortical circuits that serve
face detection in infancy, are inhibited by cortical circuits during maturation (Johnson et al.,
2015; Pascalis & Kelly, 2009; Stein et al., 2011). It has become clear that a lot of cortical
regions also play a role in the processing of unconscious information (Dehaene, 2014). A
cortical brain region that is important in the processing of faces is the fusiform face area
(FFA). It is found that this brain area activates when faces are presented unconsciously
(Lehmann et al., 2004). Further, the FFA also activates for objects of expertise (Bilalić et al.,
2011; Gauthier et al., 2000; McGugin et al., 2012, 2014; Tarr & Gauthier, 2000). Thus, the
activation pattern of the FFA can be influenced by experience. If during adulthood the
subcortical circuit that served face detection during infancy is suppressed by the cortical brain
circuit involving the FFA, it is plausible that there is an unconscious inversion effect for
human faces and not for chimpanzee faces, as humans have far more experience with faces of
their own species. However, this is only a speculation and future studies should examine this
idea, as the current study did not look at any brain circuits.
A general methodological issue with this study is that the presentation times of the
face stimuli were not optimal. An important issue for this study was to find a presentation
time where subjects performed at chance on the discrimination task. However, results with

22
the human faces showed that at the shortest presentation time, subjects already performed
above chance at the discrimination task. The second shortest presentation time seemed to not
differ from 0 at discrimination significantly, but a p-value of 0.06 can also be interpreted as
borderline significant and Bayesian statistics showed that it was almost equally likely that
participants performed at chance versus above chance (Tshikuka et al., 2016). Further, a
direct comparison between the inversion effect and discrimination showed no evidence for an
unconscious inversion effect. However, analysing the inversion patterns for trials with correct
discrimination response separately from trials with incorrect discrimination response showed
some evidence for an unconscious origin of the inversion effect for human faces. Because of
these contradictory results, the conclusion that there is an unconscious inversion effect for
human faces should be treated with caution and future studies should try to find optimal
presentation times to determine whether the face inversion effect occurs unconsciously.
The current study shows that it is important to re-examine the b-CFS literature by
adopting the detection-discrimination dissociation paradigm, as the findings are inconsistent
with previous b-CFS results of Stein et al. (2012). As previously indicated, findings with b-
CFS cannot only be based on unconscious processing, but also on conscious influences.
Therefore, in order to clarify the scope and limits of unconscious processing, the detection-
discrimination dissociation paradigm should be used instead of b-CFS, as this paradigm
excludes conscious contributions to detection. Besides re-examining the b-CFS literature,
future research could also build upon this study by examining whether other facial properties
are processed unconsciously. For example, it would be interesting to study whether facial
expressions are processed unconsciously or whether familiarity of faces influence detection
unconsciously.
In conclusion, this study provided some evidence for an unconscious face inversion
effect specific to conspecifics. However, it cannot be concluded with certainty that this effect

23
is unconscious. This study points out that the extent of unconscious processing may be
somewhat limited and sheds light on previous b-CFS studies. Further, the current findings
highlight the importance of using the novel detection-discrimination dissociation paradigm in
research about unconscious processing. Adopting this new method in future research will
help unravelling the scope and limits of unconscious processing.

24
References
Albonico, A., Furubacke, A., Barton, J. J. S., & Oruc, I. (2018). Perceptual efficiency and the
inversion effect for faces, words and houses. Vision Research, 153, 91–97.
https://doi.org/10.1016/j.visres.2018.10.008
Axelrod, V., Bar, M., & Rees, G. (2015). Exploring the unconscious using faces. Trends in
Cognitive Sciences, 19(1), 35–45. https://doi.org/10.1016/j.tics.2014.11.003
Bilalić, M., Langner, R., Ulrich, R., & Grodd, W. (2011). Many Faces of Expertise: Fusiform
Face Area in Chess Experts and Novices. Journal of Neuroscience, 31(28), 10206–
10214. https://doi.org/10.1523/JNEUROSCI.5727-10.2011
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10(4), 433–436.
https://doi.org/10.1163/156856897X00357
de Gelder, B., Tamietto, M., van Boxtel, G., Goebel, R., Sahraie, A., van den Stock, J.,
Stienen, B. M. C., Weiskrantz, L., & Pegna, A. (2008). Intact navigation skills after
bilateral loss of striate cortex. Current Biology, 18(24), R1128–R1129.
https://doi.org/10.1016/j.cub.2008.11.002
Dehaene, S. (2014). Consciousness and the Brain: Deciphering How the Brain Codes Our
Thoughts. Pinguin Group.
Del Río, M., Greenlee, M. W., & Volberg, G. (2018). Neural dynamics of breaking
continuous flash suppression. NeuroImage, 176, 277–289.
https://doi.org/10.1016/j.neuroimage.2018.04.041
Ekman, P., & Friesen, W. V. (1976). Pictures of Facial Affect. Consulting Psychologists
Press.
Fahrenfort, J. J., Scholte, H. S., & Lamme, V. A. F. (2007). Masking Disrupts Reentrant
Processing in Human Visual Cortex. Journal of Cognitive Neuroscience, 19(9), 1488–
1497. https://doi.org/10.1162/jocn.2007.19.9.1488

25
Faivre, N., Berthet, V., & Kouider, S. (2012). Nonconscious Influences from Emotional
Faces: A Comparison of Visual Crowding, Masking, and Continuous Flash
Suppression. Frontiers in Psychology, 3. https://doi.org/10.3389/fpsyg.2012.00129
Fekete, T., Van de Cruys, S., Ekroll, V., & van Leeuwen, C. (2018). In the interest of saving
time: A critique of discrete perception. Neuroscience of Consciousness, 2018(1).
https://doi.org/10.1093/nc/niy003
Gauthier, I., Skudlarski, P., Gore, J. C., & Anderson, A. W. (2000). Expertise for cars and
birds recruits brain areas involved in face recognition. Nature Neuroscience, 3(2),
191–197. https://doi.org/10.1038/72140
Gayet, S., & Stein, T. (2017). Between-Subject Variability in the Breaking Continuous Flash
Suppression Paradigm: Potential Causes, Consequences, and Solutions. Frontiers in
Psychology, 8. https://doi.org/10.3389/fpsyg.2017.00437
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The distributed human neural system
for face perception. Trends in Cognitive Sciences, 4(6), 223–233.
https://doi.org/10.1016/S1364-6613(00)01482-0
Izatt, G., Dubois, J., Faivre, N., & Koch, C. (2014). A direct comparison of unconscious face
processing under masking and interocular suppression. Frontiers in Psychology, 5.
https://doi.org/10.3389/fpsyg.2014.00659
JASP Team (2020). JASP (Version 0.12.2) [Computer software].
Jessen, S., & Grossmann, T. (2015). Neural signatures of conscious and unconscious
emotional face processing in human infants. Cortex, 64, 260–270.
https://doi.org/10.1016/j.cortex.2014.11.007
Jiang, Y., Costello, P., & He, S. (2007). Processing of Invisible Stimuli: Advantage of
Upright Faces and Recognizable Words in Overcoming Interocular Suppression.

26
Psychological Science, 18(4), 349–355. https://doi.org/10.1111/j.1467-
9280.2007.01902.x
Johnson, M. H., Dziurawiec, S., Ellis, H., & Morton, J. (1991). Newborns’ preferential
tracking of face-like stimuli and its subsequent decline. Cognition, 40, 1–19.
https://doi.org/10.1016/0010-0277(91)90045-6
Johnson, M. H. (1990). Cortical maturation and the development of visual attention in early
infancy. Journal of Cognitive Neuroscience, 2(2), 81–95.
https://doi.org/10.1162/jocn.1990.2.2.81
Johnson, M. H. (2005). Subcortical face processing. Nature Reviews Neuroscience, 6(10),
766–774. https://doi.org/10.1038/nrn1766
Johnson, M. H., Senju, A., & Tomalski, P. (2015). The two-process theory of face
processing: Modifications based on two decades of data from infants and adults.
Neuroscience & Biobehavioral Reviews, 50, 169–179.
https://doi.org/10.1016/j.neubiorev.2014.10.009
Kwak, S. G., & Kim, J. H. (2017). Central limit theorem: The cornerstone of modern
statistics. Korean Journal of Anesthesiology, 70(2), 144–156.
https://doi.org/10.4097/kjae.2017.70.2.144
Lamme, V. A. F., & Roelfsema, P. R. (2000). The distinct modes of vision offered by
feedforward and recurrent processing. Trends in Neurosciences, 23(11), 571–579.
https://doi.org/10.1016/S0166-2236(00)01657-X
Lehmann, C., Mueller, T., Federspiel, A., Hubl, D., Schroth, G., Huber, O., Strik, W., &
Dierks, T. (2004). Dissociation between overt and unconscious face processing in
fusiform face area. NeuroImage, 21(1), 75–83.
https://doi.org/10.1016/j.neuroimage.2003.08.038

27
Lin, Z., & He, S. (2009). Seeing the invisible: The scope and limits of unconscious
processing in binocular rivalry. Progress in Neurobiology, 87(4), 195–211.
https://doi.org/10.1016/j.pneurobio.2008.09.002
Macmillan, N. A., & Creelman, C. D. (2005). Detection theory: A user’s guide, 2nd ed (pp.
xix, 492). Lawrence Erlbaum Associates Publishers.
Marcel, A. J. (1983). Conscious and unconscious perception: Experiments on visual masking
and word recognition. Cognitive Psychology, 15(2), 197–237.
https://doi.org/10.1016/0010-0285(83)90009-9
McGugin, R. W., Gatenby, J. C., Gore, J. C., & Gauthier, I. (2012). High-resolution imaging
of expertise reveals reliable object selectivity in the fusiform face area related to
perceptual performance. Proceedings of the National Academy of Sciences of the
United States of America, 109(42), 17063–17068.
https://doi.org/10.1073/pnas.1116333109
McGugin, R. W., Newton, A. T., Gore, J. C., & Gauthier, I. (2014). Robust expertise effects
in right FFA. Neuropsychologia, 63, 135–144.
https://doi.org/10.1016/j.neuropsychologia.2014.08.029
McKone, E., Kanwisher, N., & Duchaine, B. C. (2007). Can generic expertise explain special
processing for faces? Trends in Cognitive Sciences, 11(1), 8–15.
https://doi.org/10.1016/j.tics.2006.11.002
Meyen, S., Zerweck, I. A., Amado, C., von Luxburg, U., & Franz, V. H. (2020). The
unconscious priming fallacy: When can scientists truly claim an indirect task
advantage? ArXiv. http://arxiv.org/abs/2004.14987
Moors, P., Gayet, S., Hedger, N., Stein, T., Sterzer, P., Ee, R. van, Wagemans, J., &
Hesselmann, G. (2019). Three Criteria for Evaluating High-Level Processing in

28
Continuous Flash Suppression. Trends in Cognitive Sciences, 23(4), 267–269.
https://doi.org/10.1016/j.tics.2019.01.008
Neiworth, J. J., Hassett, J. M., & Sylvester, C. J. (2006). Face processing in humans and new
world monkeys: The influence of experiential and ecological factors. Animal
Cognition, 10(2), 125. https://doi.org/10.1007/s10071-006-0045-4
Parr, L. A. (2011). The inversion effect reveals species differences in face processing. Acta
Psychologica, 138(1), 204–210. https://doi.org/10.1016/j.actpsy.2011.06.005
Pascalis, O., & Kelly, D. J. (2009). The Origins of Face Processing in Humans: Phylogeny
and Ontogeny. Perspectives on Psychological Science, 4(2), 200–209.
https://doi.org/10.1111/j.1745-6924.2009.01119.x
Pituch, K. A., & Stevens, J. (2016). Applied multivariate statistics for the social sciences:
Analyses with SAS and IBM’s SPSS (6th edition). Routledge/Taylor & Francis Group.
Pournaghdali, A., & Schwartz, B. L. (2020). Continuous flash suppression: Known and
unknowns. Psychonomic Bulletin & Review, 27(6), 1071–1103.
https://doi.org/10.3758/s13423-020-01771-2
Prentiss, E. K., Schneider, C. L., Williams, Z. R., Sahin, B., & Mahon, B. Z. (2018).
Spontaneous in-flight accommodation of hand orientation to unseen grasp targets: A
case of action blindsight. Cognitive Neuropsychology, 35(7), 343–351.
https://doi.org/10.1080/02643294.2018.1432584
Reed, C. L., Stone, V. E., Bozova, S., & Tanaka, J. (2003). The Body-Inversion Effect.
Psychological Science, 14(4), 302–308. https://doi.org/10.1111/1467-9280.14431
Sanders, M. D., Warrington, E.K., Marshall, J., & Wieskrantz, L. (1974). ‘Blindsight’: Vision
in a field defect. The Lancet, 303(7860), 707–708. https://doi.org/10.1016/S0140-
6736(74)92907-9

29
Schmidt, T., & Vorberg, D. (2006). Criteria for unconscious cognition: Three types of
dissociation. Perception & Psychophysics, 68(3), 489–504.
https://doi.org/10.3758/BF03193692
Simpson, E. A., Husband, H. L., Yee, K., Fullerton, A., & Jakobsen, K. V. (2014). Visual
search efficiency is greater for human faces compared to animal faces. Experimental
Psychology, 61(6), 439–456. https://doi.org/10.1027/1618-3169/a000263
Srinivasan, N. (2020). Consciousness Without Content: A Look at Evidence and Prospects.
Frontiers in Psychology, 11. https://doi.org/10.3389/fpsyg.2020.01992
Stein, T. (2019). The Breaking Continuous Flash Suppression Paradigm. In G. Hesselmann
(Ed.), Transitions between Consciousness and Unconsciousness (pp. 1–38).
Routledge. https://doi.org/10.4324/9780429469688-1
Stein, T., & Peelen, M. (2021). Dissociating conscious and unconscious influences on visual
detection effects. Nature Human Behaviour. https://doi.org/10.1038/s41562-020-
01004-5
Stein, T., Peelen, M. V., & Sterzer, P. (2011). Adults’ Awareness of Faces Follows
Newborns’ Looking Preferences. PLOS ONE, 6(12), e29361.
https://doi.org/10.1371/journal.pone.0029361
Stein, T., Reeder, R. R., & Peelen, M. V. (2016). Privileged access to awareness for faces and
objects of expertise. Journal of Experimental Psychology: Human Perception and
Performance, 42(6), 788–798. https://doi.org/10.1037/xhp0000188
Stein, T., & Sterzer, P. (2014). Unconscious processing under interocular suppression:
Getting the right measure. Frontiers in Psychology, 5.
https://doi.org/10.3389/fpsyg.2014.00387

30
Stein, T., Sterzer, P., & Peelen, M. V. (2012). Privileged detection of conspecifics: Evidence
from inversion effects during continuous flash suppression. Cognition, 125(1), 64–79.
https://doi.org/10.1016/j.cognition.2012.06.005
Tarr, M. J., & Gauthier, I. (2000). FFA: A flexible fusiform area for subordinate-level visual
processing automatized by expertise. Nature Neuroscience, 3(8), 764–769.
https://doi.org/10.1038/77666
Trevethan, C. T., Sahraie, A., & Weiskrantz, L. (2007). Form discrimination in a case of
blindsight. Neuropsychologia, 45(9), 2092–2103.
https://doi.org/10.1016/j.neuropsychologia.2007.01.022
Tshikuka, J.-G., Magafu, M. G. M. D., Molefi, M., Masupe, T., Matchaba-Hove, R. B.,
Mbongwe, B., & Tapera, R. (2016). Addressing the Challenge of P-Value and Sample
Size when the Significance is Borderline: The Test of Random Duplication of
Participants as a New Approach. International Journal of Statistics in Medical
Research, 5(3), 214-218. https://doi.org/10.6000/1929-6029.2016.05.03.7
Tsuchiya, N., & Koch, C. (2005). Continuous flash suppression reduces negative afterimages.
Nature Neuroscience, 8(8), 1096–1101. https://doi.org/10.1038/nn1500
Weiskrantz, L. (1996). Blindsight revisited. Current Opinion in Neurobiology, 6(2), 215–220.
https://doi.org/10.1016/S0959-4388(96)80075-4
White, P. A. (2018). Is conscious perception a series of discrete temporal frames?
Consciousness and Cognition, 60, 98–126.
https://doi.org/10.1016/j.concog.2018.02.012
Zhou, G., Zhang, L., Liu, J., Yang, J., & Qu, Z. (2010). Specificity of face processing without
awareness. Consciousness and Cognition: An International Journal, 19(1), 408–412.
https://doi.org/10.1016/j.concog.2009.12.009


















