
New Basal Interatheriidae (Typotheria, Notoungulata ...
32
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3520, 32 pp., 16 figures, 4 tables July 31, 2006 New Basal Interatheriidae (Typotheria, Notoungulata, Mammalia) from the Paleogene of Central Chile RALPH B. HITZ, 1 JOHN J. FLYNN, 2 AND ANDRE ´ R. WYSS 3 ABSTRACT Two new basal interatheriids (‘‘notopithecines’’) are described from central Chile. Specimens of a new taxon, Johnbell hatcheri, derive from Abanico Formation deposits near Termas del Flaco, in the valley of the Rı ´o Tinguiririca, forming a distinctive element of the Tinguiririca Fauna. The Tinguiririca Fauna, which forms the basis of the recently defined Tinguirirican SALMA, documents the co-occurrence of species of archaic, dentally primitive herbivores and basal members of later diverging groups of more advanced hypsodont forms, and other transitional aspects of mammal evolution near the Eocene/Oligocene boundary. A specimen recovered from the Abanico Formation in the drainage of the Rı ´o Cachapoal (,100 km north of Termas del Flaco), SGOPV 3451 is referred to Johnbell hatcheri. This is the first time a specimen from elsewhere in the central Chilean Main Range has been assigned to a species represented in the stratotype sequence of the Tinguiririca Fauna (upper Rı ´o Tinguiririca Valley). Ignigena minisculus, the other new basal interatheriid recognized herein, is known only from older strata of the Abanico Formation, from the Tapado Fauna within the Rı ´o Tinguiririca Valley, estimated to be Casamayoran in age. Phylogenetic analysis shows these two new basal interatheriids to be outgroups to the Interatheriinae. Both new taxa are smaller than all other interatheriids known, except Punapithecus. Small body size may reflect geographic provincialism, as these diminutive forms are resticted to more northern latitudes compared to larger basal interatheriids, which derive from Patagonia. SGOPV 3604, from the Abanico Formation along the Rı ´o Azufre, several kilometers north of the Rı ´o Tinguiririca drainage, is referred to the early interatheriid Antepithecus brachystephanus, a taxon otherwise known only from Casamayoran (‘‘late’’ Barrancan subage) SALMA deposits in Patagonia. Copyright E American Museum of Natural History 2006 ISSN 0003-0082 1 Earth Sciences Department, Tacoma Community College, 6501 South 19th Street, Tacoma, WA 98466 ([email protected]). 2 Division of Paleontology, American Museum of Natural History ( [email protected]). 3 Department of Earth Science, University of California, Santa Barbara, CA 93106 ([email protected]).
Transcript of New Basal Interatheriidae (Typotheria, Notoungulata ...

PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY
CENTRAL PARK WEST A T 79TH STREET, NEW YOR K, NY 10024
Number 3520, 32 pp., 16 figures, 4 tables July 31, 2006
New Basal Interatheriidae (Typotheria,Notoungulata, Mammalia) from the Paleogene of
Central Chile
RALPH B. HITZ,1 JOHN J. FLYNN,2 AND ANDRE R. WYSS3
ABSTRACT
Two new basal interatheriids (‘‘notopithecines’’) are described from central Chile. Specimens ofa new taxon, Johnbell hatcheri, derive from Abanico Formation deposits near Termas del Flaco, inthe valley of the Rıo Tinguiririca, forming a distinctive element of the Tinguiririca Fauna. TheTinguiririca Fauna, which forms the basis of the recently defined Tinguirirican SALMA,documents the co-occurrence of species of archaic, dentally primitive herbivores and basalmembers of later diverging groups of more advanced hypsodont forms, and other transitionalaspects of mammal evolution near the Eocene/Oligocene boundary. A specimen recovered from theAbanico Formation in the drainage of the Rıo Cachapoal (,100 km north of Termas del Flaco),SGOPV 3451 is referred to Johnbell hatcheri. This is the first time a specimen from elsewhere in thecentral Chilean Main Range has been assigned to a species represented in the stratotype sequenceof the Tinguiririca Fauna (upper Rıo Tinguiririca Valley). Ignigena minisculus, the other new basalinteratheriid recognized herein, is known only from older strata of the Abanico Formation, fromthe Tapado Fauna within the Rıo Tinguiririca Valley, estimated to be Casamayoran in age.Phylogenetic analysis shows these two new basal interatheriids to be outgroups to theInteratheriinae. Both new taxa are smaller than all other interatheriids known, exceptPunapithecus. Small body size may reflect geographic provincialism, as these diminutive formsare resticted to more northern latitudes compared to larger basal interatheriids, which derive fromPatagonia.
SGOPV 3604, from the Abanico Formation along the Rıo Azufre, several kilometers north ofthe Rıo Tinguiririca drainage, is referred to the early interatheriid Antepithecus brachystephanus,a taxon otherwise known only from Casamayoran (‘‘late’’ Barrancan subage) SALMA deposits inPatagonia.
Copyright E American Museum of Natural History 2006 ISSN 0003-0082
1 Earth Sciences Department, Tacoma Community College, 6501 South 19th Street, Tacoma, WA 98466([email protected]).
2 Division of Paleontology, American Museum of Natural History ( [email protected]).3 Department of Earth Science, University of California, Santa Barbara, CA 93106 ([email protected]).

A phylogenetic definition for the name Interatheriidae is proposed on the strength of theresolution achieved in the phylogenetic analysis. This analysis also shows clearly that‘‘Notopithecinae’’ represents a paraphyletic assemblage. We suggest that taxa formerly termed‘‘notopithecines’’ are more appropriately referred to as ‘‘basal interatheriids’’.
INTRODUCTION
The Tinguiririca Fauna, the first indicationsof which were uncovered in 1988 (Novacek etal., 1989), was the first in a long series of newmammalian assemblages being discovered inthe Abanico Formation (and equivalents) inthe central Andean Main Range of Chile(Wyss et al., 1990, 1992, 1993, 1994; Charrieret al., 1990, 1996; Flynn et al., 1991, 1995,2003; Hitz et al., 2000; Croft et al., 2003;Reguero et al., 2003a; Flynn and Wyss, 2004).This formation and its lateral correlativesstrike parallel to the Main Range (i.e., north-south) and have yielded mammalian faunasacross a .400 km long band, between ap-proximately 33u459S (directly east of Santiagoin the Rıo Colorado and Cajon del Maiporegions) and ,37u 209S (Laguna del Laja)(fig. 1). These faunas represent many distincttemporal horizons, ranging in age from theCasamayoran (probably late Eocene, Kay etal., 1998) through the Colhuehuapian (earlyMiocene, Flynn and Swisher, 1995) or youn-ger SALMAs; radioisotopic data indicate thatdeposition of the Abanico Formation extend-ed until at least middle Miocene time(Charrier et al., 2002). Mammal faunas arepreserved in volcaniclastic intervals of theinfilling of long-lived extensional basin(s);these accumulations, approaching 5 km instratigraphic thickness, were uplifted and de-formed by compressional tectonics beginningin the middle-late Miocene (Charrier et al.,2002, 2005). Deposition of the unit may havebegun prior to the Casamayoran, but theprecise timing of initiation of sedimentation inthe basin is currently uncertain, given strati-graphic complexity arising from strong com-pressive deformation and often limited appli-cability of radioisotopic dating owing to low-temperature metamorphic alteration.
This paper extends description and phylo-genetic analysis of new taxa from the AbanicoFormation, both from the Tinguiririca valley(in the original Tinguiririca Fauna and theolder Tapado Fauna), and others more re-cently discovered elsewhere. Two of the speci-
mens described below fall in the lattercategory, coming from the Cachapoal Fauna(Rıo Cachapoal) discovered in 1996 and theAzufre Locality (Rıo Azufre) discovered in1997. Complementing our earlier work onTinguiririca interatheriids (Hitz et al., 2000)we here describe two new early divergingmembers of this group of small- to medium-sized notoungulates, and present a phylogenet-ic analysis of the Interatheriidae (Typotheria).Interatheriids, including both brachydont(presumably browsing) and hypsodont (pre-sumably grazing) forms, are generally wellrepresented in South American early-middleCenozoic land mammal faunas. Interatheriidsfirst appear in the Paleocene Riochican South
Fig. 1. Map of the central Chilean Andes,illustrating the broad swath of AbanicoFormation and equivalent deposits (shaded) andlocalities/faunas referred to in the text.
2 AMERICAN MUSEUM NOVITATES NO. 3520

American Land Mammal ‘‘Age’’ (SALMA)(Marshall et al., 1983) and persist into theHuayquerian SALMA (Late Miocene), exhi-biting a radiation of hypsodont forms duringlate Oligocene time (Cifelli, 1985; Marshalland Cifelli, 1990). Interatheriids are tradition-ally divided into two groups (Simpson, 1945;McKenna and Bell, 1997), the Notopithecinae(early Paleogene, low-crowned forms) and theInteratheriinae (late Paleogene and Neogene,hypsodont and hypselodont forms). Phylo-genetic studies (Cifelli, 1993; Hitz, 1997; Hitzet al., 2000; Reguero et al., 2003b; this paper)have corroborated the monophyly of theInteratheriinae, whereas the taxa generallyreferred to the Notopithecinae form a para-phyletic assemblage. As detailed below, owingto the paraphyly of ‘‘Notopithecinae’’ wefavor referring to these forms collectively as‘‘basal interatheriids’’, reflecting their lack ofmembership in the Interatheriinae. Lastly, thenew Chilean basal interatheriids illustrate aninteresting biogeographic-morphologic pat-tern—an assemblage of unusually small-bod-ied forms having an extra-Patagonian distri-bution. The two new Chilean taxa, andPunapithecus, a recently described basal inter-atheriid from Catamarca and Salta Provinces,Argentina (Lopez and Bond, 1995; Alonso etal., 1988) are significantly smaller than pre-viously known interatheriids, and all threetaxa are known exclusively from localities farremoved from Patagonia.
Hitz et al. (2000) described two interather-iines, Santiagorothia chiliensis, from theTinguiririca Fauna and several Patagonianlocalities, and Proargyrohyrax, known onlyfrom Patagonia. Emended descriptions anddiagnoses for the Patagonian interatheriinesArgyrohyrax acuticostatus and Eopachyruchosplicifera were provided as well. Santiagorothiachiliensis and Proargyrohyrax are the earliest(Tinguirirican SALMA; Flynn et al., 2003)known interatheriines (sensu Hitz et al, 2000:3). Notably, specimens from various Argen-tine Patagonian localities (‘‘Astraponoteenplus superieure level’’, Chubut [Bond et al.,1996, 1997a]; Lomas Blancas, Chubut [Hitz etal., 2000]; Rocas Bayas, Rio Negro [Pascual etal., 1984; Bond et al., 1997b]; Laguna de laBombilla, Chubut [Pascual, 1965]) have beenreferred to Santiagorothia chiliensis (the holo-
type of which is from Chile; Hitz et al., 2000)thereby highlighting the biostratigraphic utili-ty of interatheriid taxa from the TinguiriricaFauna. The correlations offered in Hitz et al.(2000), and the occurrence of a conspecificoutside the Tinguiririca valley reported below,add to a growing body of evidence indicatingthat strata referable to the TinguiriricanSALMA (Flynn et al., 2003) occur beyondcentral Chile (see also Croft et al., 2003;Reguero et al., 2003a, 2003b).
Specimens examined in this study forsystematic comparison and character assess-ment in the phylogenetic analysis are housedin the American Museum of Natural History(AMNH); Field Museum of Natural History(FMNH); Florida Museum of NaturalHistory (FLMNH); Servicio Geologico deBolivia (GEOBOL); Los Angeles CountyMuseum (LACM); Museo Argentino deCiencias Naturales, Buenos Aires (MACN);Museo de La Plata (MLP); MuseoNacional de Historia Natural, Bolivia(MNHN); Museo Nacional de HistoriaNatural, Santiago (SGOPV); University ofCalifornia Museum of Paleontology (UCMP);and Yale Peabody Museum (YPM). Otherabbreviations include: R (Right), L (Left), i/I(lower/upper incisor), c/C (lower/upper ca-nine), p/P (lower/upper premolar), m/M (low-er/upper molar), dp/dP (deciduous lower/upper premolar), HI (hypsodonty index),SALMA (South American Land Mammal‘‘Age’’).
SYSTEMATIC PALEONTOLOGY
NOTOUNGULATA ROTH, 1903
TYPOTHERIA ZITTEL, 1893
INTERATHERIIDAE AMEGHINO, 1887
Johnbell hatcheri, new taxon
Figures 2–5
HOLOTYPE: SGOPV 3106: left maxilla,LI3-M3.
PARATYPE: SGOPV 2950: mandibles pre-serving symphysis, Ri2-3, Li1-m3.
REFERRED SPECIMENS: SGOPV 2910: Rp4-m2. SGOPV 2902: Rp1-m3. SGOPV 5001:a palate and mandibles, partially prepared,
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 3

LP2-M3 and Lp3-m3 exposed. SGOPV 3451:RP2-M2.
ETYMOLOGY: For John Bell Hatcher, a coalminer turned vertebrate paleontologist,known for his prodigious collecting andpioneering fieldwork in remote regions ofPatagonia, where he secured valuable Mio-cene fossil mammal collections includingmany interatheriid specimens.
DIAGNOSIS: Johnbell hatcheri is a smalltypothere possessing upper molars that arelonger than wide and have fossettes thatdisappear rapidly with wear, leaving a feature-less crown save for a persistent lingual sulcus.Among typotheres, this combination of fea-tures is found only in one subgroup ofinteratheriids (unnamed clade, node 2 infig. 6, this paper). The following distinctionsexclude Johnbell hatcheri from other groupsof small-bodied typotheres: archaeopithecidshave upper molars that are equidimensional orwider than long, with long-lived fossettes, and
archaeohyracids and hegetotheriids are muchlarger typotheres lacking a persistent lingualsulcus on upper molars. Assignment to theMesotheriidae is ruled out by their larger sizeand pronounced trilobed (only trilobed earlyin wear in trachytheriines, persistently trilobedin mesotheriines), hypsodont upper molars.
Johnbell hatcheri is an early divergingmember of the least inclusive clade (unnamed)including Interatheriinae (sensu Hitz et al.,2000, see fig. 6, node 4 this paper),Eopachyrucos, and Ignigena minisculus (newtaxon, below). This clade (clade 2, fig. 6) ischaracterized ancestrally, and distinguishedfrom all other interatheriids, by the followingderived features: smooth posterior ectolophon the upper premolars, and upper molarslonger than wide. Interatheriids excludedfrom this unnamed clade (Notopithecus,Transpithecus, Guiliemoscottia, Punapithecus,Antepithecus) lack these features, readilydistinguishing them from Johnbell hatcheri.
Fig. 3. Johnbell hatcheri, new taxon, SGOPV 3106 (holotype), left maxilla, LI3-M3, buccal view.Tinguiririca Fauna.
Fig. 2. Johnbell hatcheri, new taxon, SGOPV 3106 (holotype), left maxilla, LI3-M3, occlusal view.Tinguiririca Fauna.
4 AMERICAN MUSEUM NOVITATES NO. 3520

Conversely, Johnbell hatcheri is excludedfrom clade 3 of figure 6 (i.e., Interatheriinae—sensu Hitz et al., 2000—plus Eopachyrucos), inlacking many of the synapomorphic dentalfeatures diagnosing that clade (see phyloge-netic analysis, below): a low or flat ectolophon M1-3, and hypsodont or hypseledont cheekteeth (HI for Johnbell hatcheri SGOPV 3106 is0.86 as measured on the M2). Johnbell hatcheriis one of the smallest interatheriids known(rivaled only by Ignigena minisculus andPunapithecus). Its strikingly narrow premolars(in the transverse direction) and anteroposter-iorly narrow internal sulcus on the uppermolars nevertheless clearly differentiate itfrom these two other diminutive taxa. Inaddition, Johnbell hatcheri lacks the anteriorcingulae on the upper cheekteeth present inPunapithecus.
TYPE LOCALITY: The holotype derives fromLocality Set 3 (Flynn et al., 2003) in theAbanico (5 Coya Machalı) Formation, RıoTinguiririca valley, central Chile. Locality Set3 (5 ‘‘Locality C’’ of Charrier et al., 1996:figs. 6–8) occurs in purplish volcaniclasticsediments north of the Rıo Tinguiririca, lessthan 5 km north of the other two mainlocalities (Locality Sets 1 and 2, Flynn et al.,2003) yielding the Tinguiririca Fauna atTermas del Flaco (see Wyss et al., 1994;Charrier et al., 1996; Flynn et al., 2003).
KNOWN DISTRIBUTION: Tinguirirican SALMA(Flynn et al., 2003; 5 Tinguiririca faunalinterval or ‘‘Tinguirirican’’ of Flynn andSwisher, 1995). Known from the Tingui-ririca Fauna type locality (Locality Set 1;SGOPV 2902, 2910, and 5001) and LocalitySet 3 (the holotype and paratype, SGOPV
Fig. 4. Johnbell hatcheri, new taxon, SGOPV2950 (paratype), mandibular symphysis, Ri2-3, Li1-m3, occlusal view. Tinguiririca Fauna.
Fig. 5. Johnbell hatcheri, new taxon, SGOPV 3451 (referred specimen), partial maxillary with RP2-M2,occlusal view. Cachapoal Fauna.
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 5

3106 and 2950) of Flynn et al. (2003)(Localities ‘‘A’’ and ‘‘C’’ of Charrier et al.,1996, figs. 6–8), respectively lying 2 km southand 3 km northwest of the town of Termasdel Flaco and the Rıo Tinguiririca. Theoccurrence of Johnbell hatcheri at bothLocality Sets 1 and 3 is important in corre-lating between the discontinuously exposedfossil-bearing horizons of the AbanicoFormation from the north side of the Tin-guiririca valley to strata south of the river.Additionally, a single specimen (SGOPV3451) is known from a locality (CachapoalLocality and Fauna) within the AbanicoFormation in the Rıo Cachapoal drainage,some 100 km north of Termas del Flaco(Charrier et al., 1996 Flynn and Wyss, 2004).More detailed locality information is on fileat the American Museum of Natural History.Radioisotopic determinations from Tingui-ririca bracket the age of the fauna therebetween approximately 37.5–31.5 Ma, with
the fossiliferous horizons themselves directlydated at , 31.5 Ma (Wyss et al., 1994; Flynnet al., 2003) and the Tinguirirican SALMAlikely spanning at least 31–33 Ma (Flynn etal., 2003).
REMARKS: In a preliminary description ofthese specimens (Hitz, 1997), SGOPV 2950,a mandible, was designated as the principalspecimen representing this new taxon, andSGOPV 3106, a nearly complete maxilla, wasdesignated as the equivalent of the paratype,although it was not formally named in thatdissertation. Upon further consideration, weformally designate SGOPV 3106 the holotypeand SGPOV 2950 the paratype, as maxillaetend to be more abundant in the notoungulatefossil record and preserve a greater number ofdiagnostic characters. Supporting the latterclaim is the fact that in the phylogeneticanalysis of Interatheriidae presented below,we recognize eighteen distinct characters forthe upper dentition but only ten for the lower.
Fig. 6. Consensus of 623 equally parsimonious trees of 70 steps (branch and bound search, all charactersunordered, equally weighted; for individual trees: CI 5 0.74, RI 50.86). Labeled nodes are referenced fromthe text. Character states supporting nodes are listed adjacent to the node (if no number is given, it is thesingle derived state for the character, coded as ‘‘1’’). Character states with an asterisk indicate an equivocalcondition in which either the most basal member of the clade in question is missing data for that character orthe outgroup to that clade is missing the data for that character. Boxed character states represent conditionsidentified as diagnostic of Interatheriinae sensu Hitz et al., (2000) but are shown in this study to havea broader distribution.
6 AMERICAN MUSEUM NOVITATES NO. 3520

In addition to the dental features notedabove, interatheriines (sensu Hitz et al., 2000)are diagnosed by two derived cranial features,an anteriorly projecting sliver of the frontal onthe superoanterior orbit rim, and a posteriorportion of the auditory bulla that laps ontothe paraoccipital process. Given the scarcity ofknown cranial material, these features cannotbe scored for most early interatheriids, in-cluding Eopachyrucos and Johnbell hatcheri.Nevertheless, the known distribution of thesefeatures argues that they diagnose a clade atleast as inclusive as Interatheriinae, but lessinclusive than Interatheriidae.
DESCRIPTION
Johnbell hatcheri is known from nearlycomplete maxillary and mandibular dentitions(figs. 2–5). Only SGOPV 5001 shows anarticulated upper and lower dentition, butthe teeth of this specimen are tightly clenchedand embedded within an extremely hard andbrittle volcaniclastic matrix, making thecrowns of the lower dentition inaccessible to
preparation (without risking destruction of theteeth). The remaining suite of upper and lowerdentitions is allocated to the same taxon basedon similarities of the labial faces of thecheekteeth (visible in 5001), common mor-phological features of the occlusal surfacesamong those specimens with crowns prepared,similarity in size, and the presence of featuresidentifying all specimens as basal interather-iids. Mensural information is presented intable 1.
UPPER DENTITION
The description of the upper dentition isbased principally on the holotype SGOPV3106, the specimen with the best preserved andmost completely exposed upper dentition.
Only I3 is preserved among the upperincisors. It is incisiform; a shallow, verticallyoriented groove divides the tooth into anteriorand posterior portions labially, a division thatis less apparent lingually. The upper canine isincisiform and similar in size to I3. Theanteroexternal swelling (anterior portion of
TABLE 1
Dental Measurements (in mm) for Upper and Lower Jaws of Johnbell hatcheri
Jaw
SGOPV 2950 SGOPV 3106 SGOPV 2910 SGOPV 5001 SGOPV 5001 SGOPV 3451
lower upper lower lower upper upper
AP I1 0.56 — — — — —
TR I1 0.43 — — — — —
AP I2 0.63 — — — — —
TR I2 0.48 — — — — —
AP I3 0.88 1.59 — — — —
TR I3 0.62 0.86 — — — —
AP C 1.16 1.79 — — — —
TR C 0.92 0.86 — — — —
AP P1 1.95 2.01 — — 3.08 —
TR P1 1.14 1.44 — — — —
AP P2 2.35 2.91 — — — 2.30
TR P2 1.37 1.87 — — — 1.40
AP P3 2.60 2.95 — — — 2.80
TR P3 1.71 2.45 — — — 2.10
AP P4 2.81 3.12 2.95 — — 2.90
TR P4 1.97 2.97 1.59 — — 2.60
AP M1 3.22 3.90 3.41 3.53 3.40 3.50
TR M1 2.05 3.01 1.95 — — 3.30
AP M2 3.47 4.08 — 3.81 3.83 3.70
TR M2 2.29 2.72 — — — —
AP M3 3.98 — — 5.01 — —
TR M3 2.09 — — — — —
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 7

tooth) is better developed on the canine thanon I3.
The first upper premolar bears a distinctanterolabial swelling on the ‘‘ectoloph’’ anda posterolingual shelf heel, the latter of whichprobably represents a diminutive protocone.Otherwise the tooth is relatively narrow.
The succeeding premolars are progressivelylarger but otherwise share the same distinctivemorphology seen in the premolars of otherinteratheriids. A distinct parastyle and prom-inent paracone occur on P2-4, producing a foldor groove between the two (although it doesnot penetrate the crown deeply). A smallprotocone occupies a position near the meta-cone. The lingual premolar margins aresmooth and straight, being oriented nearlyanteroposteriorly. There is no hypocone,thereby making the three posterior upperpremolars rather narrow. Anterior cingulaeare lacking (but a small emargination may bepresent on P3-4). A high posterior cingulummay have occurred on P3-4, as one is presenton P2, but it has since merged with theremainder of the crown through wear. Astrong undulation on the ectoloph corre-sponds to the parastyle and paracone, butthe remainder of the ectoloph is nearly straightposteriorly.
The first two upper molars are very similarin morphology, the most significant differencebeing the greater length of M2. These teeth areexpanded anteroexternally, lending this por-tion of the teeth a wedge shape. A smallparastyle is present, as is a shallow parastyle/paracone inflection. The ectoloph shows mildundulations; the parastyle, paracone, andmetacone have approximately an equalamount of relief on M1, but on M2 themetacone shows decidedly less relief comparedto the parastyle and paracone. The protolophextends posterolingually from the parastyle; itdoes not bear a cingulum (or if it did, thisfeature coalesced with the remainder of thecrown early in wear). The protocone andhypocone are separated by a deep sulcus,which extends nearly to the roots. Theposterior border is oriented transversely andis marked by a high cingulum, which appearsto merge with the remainder of the crownearly in wear. Anterior and posterior fossettesare seen on M1, but wear has nearly erased
them. A posterior fossette is visible on M2, butalso is nearly obliterated by wear. A smallirregular feature is visible on the anteroexter-nal quadrant of the crown, which may be theremnant of an anterior fossette, but thisstructure is faint and we refrain from identi-fying it conclusively as a fossette. Given thefaintness of the fossettes in SGOPV 3106 (theholotype, which exhibits an early wear stage),we conclude that even modest wear in thistaxon would likely have rendered the molarocclusal surfaces featureless. The lingualsulcus on both molars is deep and probablypersisted until wear neared the roots.
The third upper molar is a smaller versionof M1-2. The main morphological contrastbetween it and the two preceding teeth is in theposterior margin. The hypocone is less promi-nent on M3, making the posterior marginangle anterointernally from the posteriormargin of the ectoloph, rather than trans-versely as in M1-2. Fossettes are not visible onM3, and were probably absent entirely, giventhe early wear stage of this tooth on SGOPV3106. The lingual sulcus appears to be deepand probably persisted into late wear stages.
Teeth of the upper dentition are rooted andmay be described as brachydont, but thecrowns of the cheekteeth are heightenedsomewhat. The hypsodonty index (HI; great-est ectoloph height divided by greatest ecto-loph length) for M2 on Johnbell hatcheri(SGOPV 3106) is 0.85. For comparison, HIvalues of the M2 of other small basalinteratheriids are: Notopithecus adapinus(specimen not identified), 0.81 (Simpson,1967); Antepithecus adapinus (specimen notidentified), 0.72 (Simpson, 1967); SGOPV3168 (new taxon, Ignigena minisculus, below),0.79; Punapithecus minor (MLP-V-10-1), 0.61;Antepithecus brachystephanus (SGOPV 3604,below), 0.45. All values were calculated basedon teeth with trace to slight wear.
LOWER DENTITION
This description is based on the paratype(SGOPV 2950), SGOPV 2910, and SGOPV5001, all of which possess well-preserved lowerteeth.
The first two lower incisors are both small,each having a faint, vertically oriented groove
8 AMERICAN MUSEUM NOVITATES NO. 3520

lingually. The i3, which is slightly larger thani1-2, bears a similar groove. The lower canineis very poorly preserved and provides littleinformation.
The p1 is transversely narrow; a deep,vertically oriented groove opens lingually,giving the crown a V-shaped appearance. Theposterior arm of the V is slightly larger thanthe anterior arm and bears a small posteriorheel set off from the remainder of the crownby a shallow vertical groove labially.
The p2 closely resembles p1, except that theposterior heel is larger and more distinct. Thisheel is accentuated by a shallow groove on thelabial face of the posterior crest, as well asa vertical groove lingually.
The p3 consists of a distinct trigonid andtalonid. The trigonid is dominated by a gentlyconvex protolophid (opening lingually) anda small metalophid. This convexity producesa shallow internal vertical trough between themetaconid and the anterior portion of theprotolophid. A small talonid attaches directlyto the metaconid and is demarcated from thetrigonid by a strong labial vertical groove anda mild lingual one.
The p4 is larger than the p3 and bearsa strongly curved protolophid. The anteriormargin of the protolophid is oriented trans-versely and, given its size, may be termeda paralophid. The metalophid is also orientedtransversely; a deep vertical groove lies be-tween it and the paralophid. A small talonidarises from the metaconid and is separatedfrom the trigonid by a deep labial grooveand a shallower, more posterior lingualgroove.
The trigonids of m1-3 are similar, consistingof a diagonal paralophid (the lingual marginof which is posteriorly positioned relative tothe labial margin), a longitudinal protolophid,and a diagonal metalophid (parallel to theparalophid) displaying a prominent metaco-nid. The vertical groove between the para-lophid and the metalophid is very shallow,reaching midway down the crown toward theroots. The molar talonids are demarcatedfrom the trigonids by deep labial and lingualvertical grooves, the former being slightlymore posteriorly situated than the latter.These grooves extend nearly to the base ofthe crown. No hypoconulid is seen on m1 or
m2 and the talonids are essentially ovate incrown view, with the long axis orientedtransversely. The m1 talonid attaches to thetrigonid just labial to the metaconid. The m2-3talonids attach more labially, nearer the centerof the metalophid. The m3 talonid is largerthan those of the preceding two molars anddisplays a prominent hypoconulid, whichproduces a slight groove on the tooth’s lingualsurface.
MANDIBLE
SGOPV 5001, the paratype SGOPV 2950,and SGOPV 2910 preserve partial mandibles.SGOPV 2910 displays a mental foramen nearthe ventral margin of the horizontal ramus,positioned directly below p4. The other twospecimens do not clearly preserve mentalforamina, although a possible trace of one ispresent below p4 on SGOPV 5001. Themandible is 7.47 mm and 7.90 mm deep belowm2 on SGOPV 5001 and SGOPV 2910respectively.
DISCUSSION
SGOPV 3451, consisting of a portion ofa right maxilla containing P2-M2 (fig. 5), isremarkably similar to specimens of Johnbellhatcheri from the Rıo Tinguiririca valley (typearea of the Tinguiririca Fauna) in size andmorphology, and the anatomical descriptionprovided above applies to this specimen asclosely as it does to specimens from theTinguiririca region. SGOPV 3451 derives froman as yet little studied locality (yielding theCachapoal Fauna; Charrier et al., 1996; Flynnand Wyss, 2004) in the Rıo Cachapoaldrainage, about 5 km NW of the Rıo LasLenas fossiliferous sites (Flynn et al., 1995)and ,100 km N of the fossiliferous stratacropping out near Termas del Flaco that bearthe Tinguiririca and Tapado faunas.
The Cachapoal specimen, SGOPV 3451,shares a key diagnostic feature with Johnbellhatcheri (transversely narrow upper premo-lars), and the specimen otherwise displays notaxonomically significant features or sizedifferences differentiating it from all otherspecimens here referred to Johnbell hatcheri,substantiating inclusion of this specimen in the
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 9

hypodigm. This is the first Andean specimenfrom outside the stratotype sequence of theTinguiririca Fauna (upper Rıo Tinguiriricavalley) referable to a species from that fauna,demonstrating, among other things, the utilityof Johnbell hatcheri for long distance biochro-nologic correlation.
SGOPV 3451 is in an earlier ontogeneticwear state than is the holotype of Johnbellhatcheri (SGOPV 3106). The posterior cin-gulum on the M2 of SGOPV 3451, forexample, has not yet completely merged withthe crown, as it has on SGOPV 3106. The M1and M2 of SGOPV 3451 both appear todisplay a faint anterior fossette, and the M2also has a posterior fossette. The lack ofa posterior fossette on M1 in SGOPV 3451 isin contrast to the condition in SGOPV 3106,which still retains a posterior fossette on thistooth despite greater wear. This variationseems taxonomically inconsequential, likelyreflecting only the extremely ephemeralnature of the molar fossettes in this taxon.
Ignigena minisculus, new taxon
Figures 7–10
HOLOTYPE: SGOPV 3168: left maxilla, in-cluding partial zygoma and C-M3.
REFERRED: SPECIMENS SGOPV 3167: leftmandible with p1-p3, m1.
ETYMOLOGY: Ignigena, meaning ‘‘born offire’’, in reference to the volcanoclastic depos-its from which the specimens derive; andminisculus, meaning ‘‘small’’, in reference tothe diminutive size of the specimens.
DIAGNOSIS: The same criteria listed in theprevious section marking Johnbell hatcheri asa basal interatheriid, and distinguishing itfrom all known interatheriines, apply to thistaxon as well. Fundamental differences intooth characters differentiate Ignigena minis-culus from other groups of small bodiedtypotheres: archaeopithecids have equidimen-sional upper molars with long-lived fossettes,and molars wider than long; archaeohyracidsand hegetotheriids are much larger typotheres,and lack a persistent lingual sulcus on uppermolars; even small mesotheriids are clearlylarger and have pronounced trilobed (exceptfor late wear stages in trachytheriines, wherethese teeth become bilobed) hypsodont uppermolars.
Ignigena minisculus belongs to the leastinclusive clade (unnamed) containing Intera-theriinae (sensu Hitz et al., 2000, see fig. 6,node 4 this paper) plus Eopachyrucos andJohnbell hatcheri, characterized ancestrally,
Fig. 7. Ignigena minisculus, new taxon, SGOPV 3168 (holotype), left maxillary fragment, partial zygoma,C-M3, occlusal view. Tapado Fauna.
10 AMERICAN MUSEUM NOVITATES NO. 3520
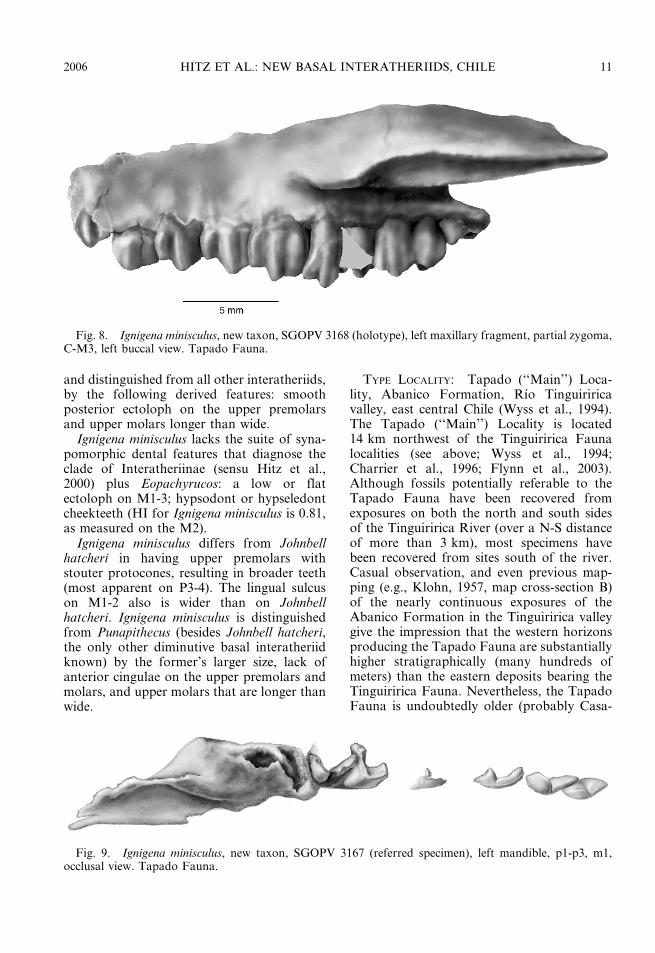
and distinguished from all other interatheriids,by the following derived features: smoothposterior ectoloph on the upper premolarsand upper molars longer than wide.
Ignigena minisculus lacks the suite of syna-pomorphic dental features that diagnose theclade of Interatheriinae (sensu Hitz et al.,2000) plus Eopachyrucos: a low or flatectoloph on M1-3; hypsodont or hypseledontcheekteeth (HI for Ignigena minisculus is 0.81,as measured on the M2).
Ignigena minisculus differs from Johnbellhatcheri in having upper premolars withstouter protocones, resulting in broader teeth(most apparent on P3-4). The lingual sulcuson M1-2 also is wider than on Johnbellhatcheri. Ignigena minisculus is distinguishedfrom Punapithecus (besides Johnbell hatcheri,the only other diminutive basal interatheriidknown) by the former’s larger size, lack ofanterior cingulae on the upper premolars andmolars, and upper molars that are longer thanwide.
TYPE LOCALITY: Tapado (‘‘Main’’) Loca-lity, Abanico Formation, Rıo Tinguiriricavalley, east central Chile (Wyss et al., 1994).The Tapado (‘‘Main’’) Locality is located14 km northwest of the Tinguiririca Faunalocalities (see above; Wyss et al., 1994;Charrier et al., 1996; Flynn et al., 2003).Although fossils potentially referable to theTapado Fauna have been recovered fromexposures on both the north and south sidesof the Tinguiririca River (over a N-S distanceof more than 3 km), most specimens havebeen recovered from sites south of the river.Casual observation, and even previous map-ping (e.g., Klohn, 1957, map cross-section B)of the nearly continuous exposures of theAbanico Formation in the Tinguiririca valleygive the impression that the western horizonsproducing the Tapado Fauna are substantiallyhigher stratigraphically (many hundreds ofmeters) than the eastern deposits bearing theTinguiririca Fauna. Nevertheless, the TapadoFauna is undoubtedly older (probably Casa-
Fig. 8. Ignigena minisculus, new taxon, SGOPV 3168 (holotype), left maxillary fragment, partial zygoma,C-M3, left buccal view. Tapado Fauna.
Fig. 9. Ignigena minisculus, new taxon, SGOPV 3167 (referred specimen), left mandible, p1-p3, m1,occlusal view. Tapado Fauna.
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 11

mayoran SALMA) than the TinguiriricaFauna, demonstrating that low-angle faultinghas thrust (albeit cryptically) older strata overyounger within the Tinguiririca valley.Evidence suggesting a Casamayoran age forthe Tapado Fauna includes the didolodontidErnestokokenia, Notostylops, and possiblyEohyrax isotemnoides (Wyss et al., 1996;Flynn et al., 2005). A more definitive ageassignment awaits ongoing taxonomic andradioisotopic work.
KNOWN DISTRIBUTION: Known only fromthe Tapado Fauna, from the Tapado (‘‘Main’’)
Locality, south of the Rıo Tinguiririca; likelyCasamayoran in age.
DESCRIPTION
Mensural information for the two inter-atheriid specimens currently known from theTapado Fauna is presented in table 2.
UPPER DENTITION
The interatheriid occurring in the TapadoFauna compares closely with Johnbell hatcherifrom the Tinguiririca Fauna, and the mor-phological features of the dentition presentedabove for Johnbell hatcheri also pertain toIgnigena minisculus, except where noted be-low. The two forms differ significantly only inthe points noted in the diagnosis of Ignigenaminisculus. Also in contrast to Johnbell hatch-eri, the P4 of Ignigena minisculus showsa minor posterior cingulum high on the crown.A similar but more distinct cingulum occurson M1 and M2. (However, the holotype ofJohnbell hatcheri [SGOPV 3106] shows greaterwear than specimens of Ignigena minisculus;thus, the difference in posterior cingulumexpression may reflect wear rather thandiscrete, taxonomically informative characterdifferences.) The third upper molar in SGOPV3168 is in the process of erupting, with onlythe tips of the paracone and protocone havingemerged from the tooth crypt.
Fig. 10. Ignigena minisculus, new taxon, SGOPV 3167 (referred specimen), left mandible, p1–p3, m1, leftbuccal view. Tapado Fauna.
TABLE 2
Dental Measurements (in mm) for Upper and LowerJaws of Ignigena minisculus
Jaw
SGOPV 3167 SGOPV 3168
lower upper
AP P1 1.73 2.35
TR P1 0.83 1.73
AP P2 1.96 2.45
TR P2 — 2.22
AP P3 — 3.14
TR P3 — 2.69
AP P4 — 3.32
TR P4 — 3.20
AP M1 — 3.69
TR M1 — 3.45
AP M2 — 3.95
TR M2 — 3.72
12 AMERICAN MUSEUM NOVITATES NO. 3520

CRANIAL MATERIAL
The anterior root of the zygoma extendsfrom the posterior half of M2 to the middle ofP4 and is thus is not remarkably broad. Thezygoma is marked by a small descendingprocess. The palatine/maxillary suture of thepalate is curved (concave posteriorly); itsanterior end is even with P4.
LOWER DENTITION
Reference of lower dental material (SGPOV3167) to Ignigena minisculus is based on size(relative to the holotype SGPOV 3168) andsimilar suites of upper and lower dentalfeatures (as present in other basal interather-iids).
SGOPV 3167 preserves fragments of anunworn lower dentition, the anterior premolarportion of which displays some unusualmorphology for an interatheriid. The firsttwo premolars are unworn. The occlusaloutline of each of these teeth, which arenarrow, forms a broad ‘‘V’’, opening lingually.In lateral view, the two wings of the ‘‘V’’, eachof which is somewhat rounded, are separatedby a cleft at their juncture, giving the teeththe appearance of ‘‘Mickey Mouse ears’’.This shape is more emphasized on p1 thanon p2. The ‘‘V’’-shaped p1–2 seem to resemblethe morphology of these teeth in Notopithecusand Johnbell hatcheri, but the latter two taxaare not known from unworn specimens,complicating direct comparison with SGOPV3167.
Only the tip of the crown of p3 is preserved,but this tooth appears to have formeda thin crescent, convex labially. Other mor-phological details of this tooth are obscuredby breakage.
The m1 is partly exposed labially, showinga convex (labially) protolophid, a transverselyoriented metalophid, and a deep labial sulcusbetween the trigonid and talonid. The talonidis ovate in outline with a flattened posteriormargin. The well-defined labial sulcus on m1is similar to that observed in other basalinteratheriids, such as Notopithecus andJohnbell hatcheri (although some other smallnon-interatheriid typotheres also have thisfeature).
Antepithecus Ameghino, 1901
Antepithecus brachystephanus Ameghino, 1901
Figures 11–12
REFERRED SPECIMEN: SGOPV 3604: partialcranium, with palate bearing right and leftdP1-dP4, and right and left M1, and anerupting right M2, as well as portions of theright zygoma and rostrum.
LOCALITY AND DISTRIBUTION: Azufre Locality,Rıo Azufre valley, Abanico Formation,Chile. SGOPV 3604 represents part of a smallsuite of dental and postcranial materialrecovered in 1997 during helicopter accessand reconnaissance prospecting of remotewestern exposures of the Abanico Forma-tion along the Rıo Azufre, due north of theRıo Tinguiririca drainage and west of VolcanTinguiririca. Its taxonomic assignment indi-cates a Casamayoran age (likely ‘‘late’’Barrancan subage, of Cifelli, 1985). Extre-mely steep cliff exposures along the west sideof the north-south trending Rıo Azufreprecluded access to most of the thick, nearlyhorizontal stratigraphic sequence exposed inthe area during this brief exploratory trip. Allof the specimens recovered from the areawere collected in a large quebrada from floatblocks of uncertain provenance.
BACKGROUND: We refer SGOPV 3604from the Azufre Locality to Antepithecus(Ameghino, 1901). Antepithecus (1901) isa basal interatheriid from the CasamayoranSALMA; its dentition is nearly identicalto Notopithecus. Simpson (1967) provideda thorough treatment and revision ofAntepithecus, noting that its similarities withNotopithecus caused him to vacillate beforeultimately concurring with Ameghino (1901)that Antepithecus indeed represents a distincttaxon. Simpson (1967) noted, in his diagnosisfor Antepithecus, the following distinctionsof Antepithecus (relative to Notopithecus):‘‘slightly more brachydont, protocone andhypocone less united, cheek teeth moreelongate, horizontal mandibular ramus shal-lower’’ (Simpson, 1967: 96). With respect tothe greater elongation of the teeth inAntepithecus, Simpson stated elsewhere inhis discussion (pp. 77–78) that the ‘‘p3, p4,m1, and m2 are significantly longer’’ and forthe upper cheek teeth, ‘‘A. brachystephanus
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 13

averages longer than N. adapinus.’’ Simp-son provided t-test support for this sizedistinction for some of the teeth in the lowerdentition, but due to small sample size did notapply the statistical test to the upper denti-tions.
Other than these highlights, Simpson pro-vided a single detailed description of a denti-tion that served to characterize bothAntepithecus and Notopithecus. Antepithecusis not yet known from cranial material.Following is a description of SGOPV 3604,and a discussion of its diagnostic resemblances
to Antepithecus and differences fromNotopithecus and other basal interatheriids.Mensural information is presented in table 3.
DESCRIPTION: The specimen consists of theright and left sides of a heavily worn dP1-dP4series, recently erupted and unworn M1s, anda RM2 in the process of erupting. Alsoexposed are the palate, a portion of the rightzygoma, and a portion of the right rostrum(maxilla and premaxilla). Additional portionsof the rostrum may be preserved within thematrix of the sample, but have yet to beprepared.
Fig. 11. Antepithecus brachystephanus, SGOPV 3604 (referred specimen), palate, right and left dP1-dP4,right and left M1, erupting right M2, portions of the right zygoma and rostrum, occlusal view.Azufre Locality.
14 AMERICAN MUSEUM NOVITATES NO. 3520

The primary criterion for identifying thedentition as consisting of dP1-dP4, and M1-2,is the relative wear on each tooth. The anteriortooth is less worn than the succeeding three,while the two posterior teeth show almost nowear (the last one is only partly erupted). Thisis the expected pattern of wear for a dP1-4,M1-2 series in notoungulates, where theeruption sequence for upper cheekteeth isdP2, dP3, dP4, dP1, M1, M2, M3, P2, P3,P4 (M. Bond, personal commun.). Evidently,this specimen is a juvenile in which thepermanent premolars had yet to erupt.
The dP1 is worn, but still shows a prominentparacone and small parastyle. An ill-defined
anterior emargination occurs low on thetooth. Wear has carved a basin between theparacone and protocone. Enamel is visible onall sides of the tooth but is completely absenton the worn occlusal surface.
The dP2 is a larger version of dP1, but witha definite low anterior cingulum and a moredistinct parastyle.
The dP3 is likewise similar to the precedingtwo teeth, save for the presence of a smallmetacone.
The paracone and metacone of dP4 are ofequal height (this tooth is less worn than thefirst three); an anterior cingulum occurs high(dorsally) on the crown, while the protoconeand hypocone remain separate until the dorsaledge of the crown is reached (producinga lingual sulcus). A small cusp is present onthe ectoloph between the paracone andmetacone. The presence of this cusp definesa tiny basin on the ectoloph.
The nearly unworn M1 has a distinctparacone and parastyle with an inflectionbetween these two structures labially. Theparacone connects lingually with a less elevat-ed protocone. The tooth’s anterior marginbears a distinct cingulum low on the crown.The height of the metacone matches that ofthe paracone; strong undulation of the ecto-loph reflects the large size of these two cuspsand the inflection between them. The meta-cone connects lingually to the hypocone, the
Fig. 12. Antepithecus brachystephanus, SGOPV 3604 (referred specimen), right dP1-dP4, right M1,erupting right M2, portions of the right zygoma and rostrum, right buccal view. Azufre Locality.
TABLE 3
Dental Measurements (in mm) for Upper Jaw ofSGOPV 3604 (Antepithecus brachystephanus)
Jaw
SGOPV 3604
upper
AP P1 3.0
TR P1 2.6
AP P2 3.0
TR P2 3.5
AP P3 3.4
TR P3 3.6
AP P4 3.8
TR P4 4.3
AP M1 4.5
TR M1 4.8
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 15

latter of which equals the protocone in height(the two lingual cusps being lower than thelabial ones). The protocone and hypocone aredeeply divided; likely they would have mergedonly after heavy wear. A posterior cingulumsits low on the tooth. Enamel covers the entirecrown.
The only observable morphology on theerupting M2 is the emerging tip of theprotocone, and a portion of the loph connect-ing the protocone to the paracone, the latter ofwhich remains unerupted. Except for thecomplete lack of wear, the protocone andloph are identical to those seen on M1.
The palatine-maxilla suture is arcuate,reaching anteriorly to the middle of dP4. Themaxilla is excavated anteriorly of the orbit, theinfraorbital foramen being located above dP3.The zygoma bears a small descending processon its anteroventral border. In addition, itdisplays what we interpret as the suturebetween the maxilla and jugal. The latterelement is reduced and excluded from theorbit, a hallmark interatheriid synapomorphy(Riggs and Patterson, 1935).
DISCUSSION
With the modest amount of morphologyavailable for study, especially with most of thepreserved teeth being deciduous and withminimal occlusal wear on the only fullyerupted permanent teeth (L & R M1), theprecise taxonomic assignment of this specimenis tentative. SGOPV 3604 is clearly a basalinteratheriid (see next paragraph) and incomparison to early interatheriids, a numberof conditions distinguish this specimen fromNotopithecus and support its referral toAntepithecus, including brachydonty, well-separated hypocone and protocone, and size.In morphology and size SGOPV 3604 isextremely similar to Antepithecus brachyste-phanus, to which we assign the new materialfrom the Azufre Locality, Chile. Simpson(1967) provisionally recognized a second spe-cies of Antepithecus, A. innexus, distinguishingit solely by its wider molars (a primitivefeature, see below), but noted that it mayrepresent an extreme variant of Antepithecusbrachystephanus; if A. innexus is a validspecies, SGOPV 3604 differs from it in its
narrower molars (and possibly the deciduouspremolar features noted directly below).SGOPV 3604 does differ from specimens ofAntepithecus brachystephanus, one of only twoearly interatheriid taxa preserving deciduouspremolars (the other being Punapithecus min-ora), in having a more pronounced protoconeon dP1-4, resulting in teeth with longer lingualmargins, and thus less triangular in outline,than in A. brachystephanus (although theproportions of the M1 are very similar to thatof Antepithecus). We decline to name thematerial of Antepithecus from the AzufreLocality as a new species at this time, becauseit would be diagnosed solely on differences indeciduous premolars; these are only rarelypreserved, and their within-species variabilityis unknown. Recovery of additional specimensof the Chilean form (showing diagnosticdifferences in the permanent upper premolarsor upper or lower teeth not represented inSGOPV 3604), or the recovery of comparabledeciduous teeth of other taxa, may ultimatelywarrant recognition of a new species ofAntepithecus, one most closely related to A.brachystephanus among known species ofinteratheriids.
Following the phylogenetic analyses oftypothere relationships by Hitz (1995, 1997;see also Hitz et al., 2000), early divergingmembers of the minimally inclusive clade thatincludes Colbertia, Maxschlosseria, Brachy-stephanus, Tsamnichoria, Oldfieldthomasia,and Acropithecus are characterized by perma-nent upper molars that are significantly widerthan long. In Oldfieldthomasia debilitata, forexample, the upper molars are 30% wider thanlong. These dimensions impart an occlusalpattern that is likewise wider than long(following the very initial wear stage). Inother typotheres (interatheriids, hegetother-iids, mesotheriids, and archaeohyracids) theparacone/parastyle region expands anteriorly,producing upper molars that are equidimen-sional (or even longer than wide). SGOPV3604 has nearly equidimensional deciduouspremolars and M1 (the only permanent cheek-tooth erupted in the specimen). SGOPV 3604differs from mesotheriids, archaeohyracids,and hegetotheriids in being much smallerand very low crowned. The latter conditionalso distinguishes it from archaeopithecids.
16 AMERICAN MUSEUM NOVITATES NO. 3520

General dental morphology thus indicates thatSGOPV 3604 represents a basal interatheriid(given its brachydont condition and lack ofsynapomorphies of later diverging interather-iid clades). The tentative recognition that thejugal is excluded from the orbit substantiatesassignment to the Interatheriidae rather thanany other typothere subgroup. The presence ofan anterior cingulum on M1 of SGOPV 3604is shared by only three interatheriid taxa:Notopithecus, Antepithecus, and Punapithecus.The extremely small size of Punapithecusargues against referring the new Chileanmaterial to this taxon. Additionally, the M1of Punapithecus has a parastyle that projectsmore anteriorly, giving the tooth a differentaspect than SGOPV 3604, and the ectoloph ofthe M1 of Punapithecus is considerablysmoother. Although the M1 in SGOPV 3604is nearly the same size as that of Notopithecus,the brachydont nature of this tooth (hypso-donty index or HI 5 0.45), plus the well-separated protocone and hypocone, comparemore favorably to Antepithecus. Recognizingthe limited available diagnostic morphology,and the lack of recent taxonomic revisionsor comparisons of Notopithecus and Antepi-thecus, the balance of evidence suggests thatAntepithecus is the most reasonable assign-ment currently possible. AMNH 28701,Antepithecus brachystephanus, a juvenile spec-imen figured in Simpson (1967: 97) providesa useful comparison. The M1 of AMNH28701 closely resembles that of SGOPV3604, as do the deciduous premolars. Themost notable distinction between AMNH28701 and SGOPV 3604 is the more robustprotocone on dP1-4 of the Chilean specimen,resulting in teeth with longer lingual (andhence less triangular) margins, than in A.brachystephanus. Although this differencemight indicate a species-level distinction,lacking other diagnostic differences (particu-larly in the adult morphology or permanentdentition), and given the paucity of deciduousdentitions known for other basal interatheriidsand poor understanding of within-speciesvariability in deciduous premolar morpholo-gy, we choose not to recognize the AzufreLocality specimen as a new species.
SGOPV 3604 would thus seem to indicatea Casamayoran age (possibly the ‘‘late’’
Barrancan subage, Cifelli, 1985) for the stratain which it occurs along Rıo Azufre. Shouldthe discovery of additional specimens (pre-serving M3, permanent upper premolars orother anterior teeth, or lower dentitions) tiltthe taxonomic assignment of the Azufrematerial toward Notopithecus (unlikely, giventheir many morphological differences) ratherthan Antepithecus, the biochronologic impli-cations remain the same, as both Antepithecusand Notopithecus occur throughout approxi-mately the same temporal interval. Witha few (likely erroneous) exceptions, bothNotopithecus and Antepithecus are restrictedto the Casamayoran SALMA. Simpson(1967) listed Notopithecus as occurring withinthe Riochican, but Cifelli (1985) questionedthis assigment, identifying the suspect speci-mens as Notopithecinae indet. AlthoughSimpson (1967) and Marshall et al. (1983)both listed ?Notopithecus as occurring in theMustersan SALMA, these authors obviouslywere not convinced of this allocation. Thus,the range of specimens securely identifiable asNotopithecus and Antepithecus is restricted tothe Casamayoran. Moreover, both taxa arelimited to the Barrancan subage of theCasamayoran, the younger of the twosubages identified by Cifelli (1985) on thebasis of exposures at two classic localities, theGran Barranca and Canadon Vaca. Cifelli(1985) further divided the Barrancan subageinto ‘‘early’’ and ‘‘late’’ intervals. With oneexception (see below), Notopithecus appearsonly in the ‘‘early’’ Barrancan andAntepithecus in the ‘‘late’’ Barrancan. In theuppermost portion of Section 2 fromCanadon Vaca, Cifelli (1985) reported theisolated occurrence of Notopithecus, indicat-ing that the range of this taxon extends veryslightly into the preceding Vacan subage. Kayet al. (1999) used new radioisotopic datato correlate the Barrancan subage ofthe Casamayoran SALMA to two possiblepaleomagnetic chrons, giving an age rangeof 35.34–37.60 Ma for this section, and anentirely late Eocene age for the Casamayoran(as opposed to its traditional conception asearly Eocene, see Flynn and Swisher, 1995).Kay et al. (1999) did not directly address theage of the Vacan in the Canadon Vacasection.
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 17

INTERATHERIID PHYLOGENY ANDTHE RELATIONSHIPS OF THE
CHILEAN BASAL INTERATHERIIDS
Several cladistic studies of the Inter-atheriidae have been completed in recent years(Cifelli, 1993; Hitz, 1997; Reguero, 1999;Reguero et al., 1996, 2003b), with varyinglevels of success with respect to resolution ofinterathere phylogenetic relationships. Cifelli’swork, based almost exclusively on dentalcharacters, identified epidemic homoplasy inthe group; the single resolved node stemmingfrom his study was one demarcating theInteratheriinae from Notopithecus. Cifelli(1993) suggested, as in previous papers(Cifelli, 1985; MacFadden et al., 1985,Marshall et al., 1986), the probable existenceof two monophyletic groups within theInteratheriinae: ‘‘Interatherium and allies’’and ‘‘Protypotherium and allies’’. Cifelli alsohighlighted four provisional synapomorphiesfor Interatheriidae: maxilla excluding jugalfrom orbit (sensu Riggs and Patterson, 1935),I1-C bifid, I2-P1 transversely compressed,lower molars bilobate.
Hitz (1997; the phylogeny stemming there-from is presented in Hitz et al., 2000) proposeda more highly resolved interatheriid phylogenybased on an analysis of 44 dental and cranialcharacters across 17 taxa. This more compre-hensive level of sampling, and the degree ofresolution achieved, motivated a phylogeneticdefinition (node-based) for the nameInteratheriinae to accompany the list ofsynapomorphies diagnostic of the clade towhich this name was associated (Hitz et al.,2000).
Reguero et al. (2003b) presented a phyloge-netic analysis of Interatheriidae, focusingmainly on interatheriine taxa (Notopithecusrepresenting the only basal interatheriid),building on the studies of Reguero et al.(1996) and Reguero (1999). Reguero et al.(2003b) achieved well-resolved relationshipswithin Interatheriinae, although differingsomewhat from those proposed by Hitz(1997), Hitz et al. (2000), and herein.Possible sources of these discrepancies arediscussed below.
Here we present an analysis of interatheriidsbased on the data set used in Hitz (1997)augmented with the information available for
five additional taxa: 1) Punapithecus, a basalinteratheriid from northwestern Argentina(Lopez and Bond, 1995); 2) Proargyrohyrax,a recently described interatheriine (Hitzet al., 2000); 3) Antepithecus brachystephanus;4) SGOPV 3604 (treated in isolation fromAntepithecus and Notopithecus); and5) Eopachyrucos (the diagnosis of which hasbeen recently emended [Hitz et al., 2000],including a recently described species E.ranchoverdensis [Reguero et al., 2003b]) (seetable 4 for a list of taxa in the analysis).Colbertia, an oldfieldthomasiid, served as theoutgroup. The character list and data matrix(appendix 1) included 43 dental and cranialcharacters across 21 taxa. The analysis, usingPAUP 4.0 (Swofford, 2002), yielded 623equally parsimonious trees of 70 steps (branchand bound search, all characters unordered,equally weighted; for individual trees: CI 50.74, RI 5 0.86). All characters discussedbelow are numbered as in the charactermatrix.
TABLE 4
Taxa Studied for Phylogenetic Analysis
Archaeophylus patrius Ameghino, 1897
Plagiarthrus clivus (5 Argyrohyrax proavus) Ameghino,
1897
Progaleopithecus tourneri (5 Argyrohyrax proavunculus)
Ameghino, 1904
Protypotherium australe Ameghino, 1887 (see also
Sinclair, 1909)
Protypotherium praerutilum Ameghino, 1887 (see also
Sinclair, 1909)
Interatherium robustum Ameghino, 1887 (see also Sinclair,
1909)
Interatherium extensum Ameghino, 1887 (see also Sinclair,
1909)
Cochilius volvens Ameghino, 1902
Cochilius fumensis Simpson, 1932
Miocochilius anamopodus Stirton, 1953
Punapithecus minor Reguero et al., 1996
Santiagorothia chiliensis Hitz et al., 2000
Proargyrohyrax curanderensis Hitz et al., 2000
Notopithecus adapinus Ameghino, 1897
Transpithecus obentus Ameghino, 1901
Guilielmoscottia plicifera Ameghino, 1901
Colbertia magellanica Price and Paula Couto, 1950
Eopachyrucos plicifera (Ameghino, 1901)
Eopachyrucos ranchoverdensis (Reguero et al., 2003b)
Salla new taxon A (informal taxon, Hitz, 1997)
Salla new taxon B (informal taxon, Hitz, 1997)
18 AMERICAN MUSEUM NOVITATES NO. 3520

Several nomenclatural issues regardingthe names Plagiarthrus Ameghino, 1896,Argyrohyrax Ameghino, 1897, and Pro-galeopithecus Ameghino, 1904, require clarifi-cation. All recent sources agree that thename Argyrohyrax is a junior synonym ofPlagiarthrus (Patterson, 1952; Marshall et al.,1986; Reguero, 1999; Reguero et al., 2003b),as suggested initially by Loomis (1914) andSimpson (1932). This synonymy was docu-mented most concretely by Patterson’s (1952)observation that Argyrohyrax proavus (thetype species of Argyrohyrax) is based on anupper dentition belonging to Plagiarthrusclivus (the type species of Plagiarthrus).Following Hitz (1997) and Hitz et al. (2000),we use the name Plagiarthrus to refer tospecimens of Plagiarthrus clivus as well asthose previously referred to Argyrohyraxproavus.
Secondly, Patterson (1952; repeated byMarshall et al., 1986) suggested that thetype specimen of Progaleopithecus tourneriAmeghino, 1904, probably represents a por-tion of the same specimen upon whichAmeghino based Argyrohyrax proavunculus,rendering Progaleopithecus a junior synonymof Argyrohyrax. This synonymy was acceptedby Hitz (1997) and Hitz et al. (2000), whereinspecies previously placed in Progaleopithecus(sensu Ameghino) were transferred toArgyrohyrax.
Lastly, Reguero (1999) and Reguero et al.(2003b) further revised interatheriid nomen-clature, although some of their recommenda-tions are at odds with the points justdiscussed. While recognizing the general syn-onymy of Argyrohyrax and Plagiarthrus, theseauthors regard Plagiarthrus as having priority(over Argyrohyrax), and recognize proavuncu-lus as a distinct species within Plagiarthrus—proposing the new combination Plagiarthrusproavunculus Reguero 1999. Although unableto examine specimens of Argyrohyrax proa-vunculus directly, we note that in Ameghino’soriginal descriptions (1897) of Argyrohyraxproavus and Argyrohyrax proavunculus hestated that A. proavunculus is merely a smallerversion of A. proavus and otherwise is verysimilar in morphology. Accepting A. proavusas a junior synonym of Plagiarthrus clivusjustifies the new combination Plagiarthrus
proavunculus (Reguero, 1999) for ‘‘A. proa-vunculus’’. Reguero (1999) and Reguero et al.(2003b) also suggested, based on the presenceof diagnostic characters (bicolumnar lowerincisors, rounded [?molar] trigonids; Regueroet al., 2003b), that Progaleopithecus shouldcontinue to be recognized as distinct, ratherthan as a synonym of Argyrohyrax proavun-culus as suggested previously by Patterson(1952) and Marshall et al. (1986). Havingexamined specimens of Progaleopithecus tour-neri firsthand, we concur that there aresufficient diagnostic characters to justifycontinued recognition of this taxon. Here weuse the name Plagiarthrus to refer toPlagiarthrus clivus (5 Argyrohyrax proavus)and Plagiathrus proavunculus, while Pro-galeopithecus refers to P. tourneri, which isdistinct from Plagiathrus proavunculus (5Argyrohyrax proavunculus).
The differences between the maximallyparsimonious hypothesis of relationships iden-tified here and the results from our previousanalyses (Hitz, 1997; Hitz et al., 2000)primarily center on decreased resolution ofthe basal part of the entire tree. Three of thetaxa added to the current analysis are veryfragmentary with respect to the charactermatrix; for example, SGOPV 3604 is only13% complete, Punapithecus is 46% complete,and Proargyrohyrax is 50% complete. Nonepreserves a significant amount of cranialmaterial. Novacek (1989, 1992a, 1992b) docu-mented that missing data contributed to treeinstability insofar as greater numbers ofmaximally parsimonious trees resulted fromthe inclusion of taxa with considerablemissing data. The missing data associatedwith SGOPV 3604, Punapithecus, andProargyrohyrax seem to account for thedecreased resolution from our earlier results.We have opted to employ the completecharacter matrix rather than exclude fragmen-tary taxa that potentially could act as ‘‘wild-cards’’ (Nixon and Wheeler, 1992) or other-wise obscure phylogenetic relationships, forseveral reasons. First, the correlation betweenmissing data and ‘‘wildcard behavior’’ is notwell demonstrated (Kearney and Clark, 2003).Second, the results of our analysis, althoughmore polytomous than in the analysis by Hitz(1997) still identify many well-supported
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 19

clades. Moreover, as it happens, some of thetaxa we are most interested in placing phylo-genetically (e.g., SGOPV 3604) are also amongthe most fragmentary.
An important contrast between the resultsof the present analysis and those presented byHitz (1997) is that Santiagorothia chiliensis (asa consequence of adding new taxa to theanalysis) is no longer resolved as the soleproximal outgroup to all other previouslyrecognized interatheriines. This raises somequestions with respect to the phylogeneticdefinition for the name Interatheriinae (sensuHitz et al., 2000) considered below.
A strict consensus tree (fig. 6) identifiesa clade (node 1 in fig. 6) that we equate withInteratheriidae. Monophyly of this clade issupported by many unambiguous synapomor-phies: I2-C laterally compressed (sensu Cifelli,1993) (character 2: character state 1), jugalexcluded from orbit by maxilla (sensu Cifelli,1993; Riggs and Patterson, 1935) (character30: character state 1), nasofrontal sutureanterior of orbit margin (character 31: char-acter state 1), descending process on zygoma(character 33: character state 1 or 2), zygomaangled steeply at juncture with face (character36: character state 1 or 2), narrow muzzle(character 37: character state 1), glenoid fossadeeply excavated and narrow (character 41:character state 1), postglenoid process sharpand narrow and separating fossa from meatus(character 42: character state 1 or 2). Twoadditional features identified by Cifelli (1993)(I1-C bifid, lower molars bilobate) as charac-terizing Interatheriidae are less widely distrib-uted and therefore are not regarded as synapo-morphic for all interatheriids in this analysis.
Numerous clades within Interatheriidae aresupported by unambiguous synapomorphies.(There exist additional, potential synapomor-phies diagnosing the nodes in fig. 6—beyondthose listed. Nevertheless, these are consideredequivocal, as at least one basal taxon of theclade that the character potentially diagnoses,and the nearest outgroup of that clade, cannotbe scored.) Of note is the clade (the polytomyof node 4 in fig. 6) that is roughly equivalentto—aside from including Proargyrohyrax—the Interatheriinae (sensu Hitz et al., 2000,p. 3, defined as: ‘‘the clade stemming from themost recent ancestor of Santiagorothia chilien-
sis and Interatherium plus all of its descen-dants’’). Following the phylogenetic definitionof Hitz et al. (2000), Proargyrohyrax is thuspresently considered a member of theInteratheriinae. Should future work resolvethe polytomy and place Proargyrohyrax asan outgroup to the least inclusive cladecontaining Santiagorothia chiliensis andInteratherium, then Proargyrohyrax would nolonger be an interatheriine, by definition (Hitzet al., 2000). Thus, new results might yieldchanges in the phylogenetic placement of taxa(Proargyrohyrax in this case), without per-turbing in the least a stable definition of thewidely used name Interatheriinae. Three sub-groups within the above clade warrant discus-sion. The first (node 5 in fig. 6), includingInteratherium, Cochilius, and Plagiarthrus, isdiagnosed by talonids on p2–4 subequal orlarger than the trigonids (character 23: char-acter state 3). The second (node 6 in fig. 6) isan unresolved tritomy of Protypotherium, Pro-galeopithecus tournoueri, and Miocochilius,diagnosed by a distinct third lobe on m3(character 28: character state 1) and a narrowanterior border on the lower premolarsand molars (character 29: character state 1).These two subgroups appear to correspondto the ‘‘Interatherium and allies’’ and the‘‘Protypotherium and allies’’ of Cifelli (1993).A clade (node 2 in fig. 6) relevant to thesystematic discussions in this paper is the leastinclusive one containing Interatheriinae (sensuHitz et al., 2000) plus Eopachyrucos, Johnbellhatcheri, and Ignigena minisculus; it is di-agnosed by two synapomorphies: M1–3 para-cone/parastyle inflection is reduced or absent(character 12: character state 1), and uppermolars longer than wide (character 17: char-acter state 1).
In a second analysis, all trees #71 steps (i.e.,one step more than the maximally parsimoni-ous) were retained, and a strict consensus treederived from them. This process yielded12,548 equally parsimonious trees. Severalclades of note persist in this consensus tree:the pairing of Interatherium and Cochilius,a clade consisting of Interatheriinae (sensuHitz et al., 2000) plus Proargyrohyrax andEopachyrucos (node 3, see fig. 13), and a cladeconsisting of Johnbell hatcheri and Ignigenaminisculus plus the previous clade (node 2, see
20 AMERICAN MUSEUM NOVITATES NO. 3520

fig. 13). The former clade (node 3) is the onlyone to persist in a third analysis, in which alltrees #72 steps were retained (126,998).
We next compare our results with those ofReguero et al. (2003b) to the extent possiblegiven differences between the datasets (fig. 14).
Reguero et al. (2003b) used 29 characters (22dental; seven mandibular, cranial, and post-cranial) across 12 taxa. Ten of the taxa in theiranalysis were interatheriines, and one wasa basal interatheriid. They presented a singleconsensus of their most parsimonious trees, but
Fig. 13. Upper cladogram is a consensus of all trees 71 steps or shorter. The labeled nodes are those thatpersist compared to the consensus tree of 70 steps and shorter (fig. 6). The lower cladogram is a consensus oftrees 72 steps and shorter, with persisting nodes labeled.
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 21

they did not run additional analyses retainingtrees of greater length. By contrast, we em-ployed 43 characters (29 dental; 14 mandibular,cranial, and postcranial) across 21 taxa (11interatheriines, 9 basal interatheriids, and 1outgroup). Reviewing the characters inReguero et al. (2003b), we determined 18 ofthem as roughly coincident with characters inour dataset, leaving 11 characters not corre-sponding to any of ours. Conversely, 25characters in our dataset have no equivalentsin Reguero et al. (2003b). The main differencebetween the consensus trees presented inReguero et al. (2003b) and this study is thetopology of the clades within the Interatheriinae(sensu Hitz et al., 2000). Reguero et al. (2003b)placed Plagiarthrus and Progaleopithecus inmore basal positions within Interatheriinaethan did our results.
Both studies identify Eopachyrucos ashaving diverged prior to the appearance of
the interatheriine (sensu Hitz et al., 2000)common ancestor. Reguero et al. (2003b)identified Proargyrohyrax as the nearestoutgroup to Interatheriinae (sensu Hitz etal., 2000), whereas in the current analysisProargyrohyrax falls in a basal interatheriinetritomy. Although Eopachyrucos was consid-ered an interatheriine by both Hitz et al.(2000) and Reguero et al. (2003b), based onits very high-crowned cheekteeth, it cannotbe considered such any more, given that itnow lies as an outgroup to the clade linkedto the name Interatheriinae, sensu Hitz et al.(2000). The shifting taxonomy of Eopa-chyrucos presents an opportunity to discussthe usefulness of applying phylogenetic defi-nitions to groups the understanding of whosephylogenetic relationships is in flux, andwhy a phylogenetic definition is of parti-cular utility in the case of the nameInteratheriinae.
Fig. 14. Interathere phylogeny after Reguero et al. (2003b; fig. 8). Reguero et al (2003b) presented twodifferent phylogenies using two different analytic techniques but the topologies of the trees are identical.
22 AMERICAN MUSEUM NOVITATES NO. 3520

A potential criticism of the phylogeneticdefinition for Interatheriinae (sensu Hitz et al.,2000) is that it is now at odds with thedistribution of one of the characters common-ly used to diagnose interatheriines tradition-ally, high-crowned cheekteeth (Cifelli, 1985).We explore an alternative to our earlierphylogenetic definition (Hitz et al., 2000),one possibly better conforming to classicalusage, concluding, however, that our originalnode-based definition is as robust as anyconceivable alternative and that its continueduse enhances the stability of interatheretaxonomy.
An alternative to a node-based phylogeneticdefinition (as that of Hitz et al., 2000) wouldbe an apomorphy-based definition (deQueiroz and Gauthier, 1990, 1992), withhypsodonty being the logical candidate asthe specifying apomorphy. A hypotheticalapomorphy-based phylogenetic definitionmight read, ‘‘the clade stemming from themost recent common ancestor of all inter-atheres marked by hypsodonty’’. The namewould refer to a clade encompassing allinterathere taxa historically considered to beinteratheriines (those that are hypsodont),plus it would be in alignment with moderatelyrobust nodes in both the phylogenetic analysesof Reguero et al. (2003b) and this study (node3 in fig. 6). [An extension of this examplewould be to use any of the diagnosticcharacters associated with the clade namedInteratheriinae.]
We perceive two weaknesses with thisalternative, the first being that hypsodontyis ultimately a gradational designation.Hypsodonty indices vary continuously fromvery low to very high; it does not occur indiscrete states.
The second, more fundamental, problem isthat with the broad sampling of interatheres inour analysis, we observe that the morphologi-cal contrast between what have traditionallybeen termed interatheriines and their nearestoutgroups has been lessened, with the con-tinuing recognition of successive outgroupsforming a ‘‘comb’’ (e.g., nodes 2, 3, and 4 infig. 6). Morphological ‘‘jumps’’ along thiscomb are small (i.e., nodes with significantlymore supporting character changes than pre-ceding and following nodes). Indeed, the
characters identified by Hitz et al. (2000) asdiagnostic of Interatheriinae are now spreadacross nodes 2, 3, and 4 (fig. 6). Santiagorothiachiliensis, Proargyrohyrax, and Eopachyrucos,all of which have been recently described (oremended), may be construed as morphologi-cally intermediate between basal interatheriidsand classic interatheriines such as Inter-atherium and Protypotherium. This is wellillustrated by the consensus of trees 71 stepsor shorter (fig. 13), which identifies the formerthree taxa as sharing a unique commonancestry with (but being placed outside of )the clade containing all other interatheriines.In this case, new discoveries clearly documentthat traits once thought to arise coincidentwith the origin of ‘‘interatheriines’’ (possiblyas a correlated suite of adaptations to grazingor rapid environmental change) actually wereacquired sequentially or in a mosaic fashionacross a longer time span, and only appearedclustered because of substantial taxonomicand temporal sampling gaps. In fact, recoveryof morphologically intermediate taxa is pre-cisely the expected outcome of sampling newtemporal intervals or previously poorly un-derstood portions of the phylogeny (recentdiscoveries of a long series of transitionalfeathered dinosaurs are an appropriate anal-ogy).
One could argue that the name Intera-theriinae has been used historically to refer totaxa clumping at one end of a morphologicalspectrum ranging from early, small, brachy-dont basal interatheriids to later, larger,hypsodont or hypselodont forms, but withouta clear demarcation of the taxonomic ‘‘di-viding point’’. The increasingly blurred‘‘boundary’’ between traditional ‘‘interather-iines’’ and their proximal outgroups highlightsthe many clades available to chose from incrafting a phylogenetically defined version ofthat name.
In the interest of stability, and in theabsence of a stronger alternative, here we optto leave the phylogenetic definition ofInteratheriine (sensu Hitz et al., 2000) un-changed. To modify the definition to betterconform with the traditional delineation be-tween basal interatheres and interatheriines isto chase an historical ideal that, in light of
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 23

recently described ‘‘intermediate’’ taxa, nolonger exists.
There are several consequences of thisaction. First, the character diagnosis of thenode associated with the name Interatheriinaein Hitz et al. (2000) changes to reflect thepresent analysis (distinctly bilobed p3-4 withpersistent labial and lingual sulci; maxillaexcluded from the superior orbital border byan anteriorly projecting sliver of frontal;auditory bulla lapped posteriorly onto theparaoccipital process—the latter two charac-ters are provisional, as proximal outgroups tothe clade in question cannot be scored forthese characters). Second, Eopachyrucos, pre-viously considered an interatheriine (Hitz etal., 2000; see also Reguero et al., 2003b)based on the phylogeny and diagnosis of theclade accepted at that time, is currently seenas falling outside the clade to which thatname is tied. Our conception of the clade’smembership has thus changed slightly sincethe phylogenetic definition was first pro-posed.
Our current phylogenetic results, and thoseof other studies (e.g., Cifelli, 1993; Reguero etal., 1996, 2003b; Hitz, 1997), underscore thestrong support for the monophyly ofInteratheriidae, regardless of the precise listof synapomorphies diagnosing it. In thesephylogenies, Interatheriidae is consistentlydiagnosed by the apomorphic sandwichingof the jugal between the maxilla and squamo-sal, plus the exclusion of the jugal from theorbit, unusual features not seen in othernotoungulates and that have long been usedto assign taxa to the Interatheriidae. Otherconditionally synapomorphic attributes arelisted above and in Cifelli (1993). As the nameInteratheriidae has not been defined phyloge-netically, we do so here.
The unique configuration of the zygoma ininteratheres offers a convenient basis fordefining the name of this clade of notoungu-lates via an apomorphy-based phylogeneticdefinition. Defining the name Interatheriidaein this manner maximizes its congruence withhistorical usage. In apomorphy-based defini-tions the appearance of a particular characterprovides the means of stipulating the specificclade/ancestor to which a given name is linked.Interatheriidae is hereby defined as the no-
toungulate clade stemming from the ancestordisplaying a splintlike jugal on the zygomaticarch, wherein this element is ‘‘sandwiched’’between the maxilla and squamosal andexcluded from the orbit.
All cladistic analyses of the Interatheriidaeto date (e.g., Hitz, 1997; Reguero et al., 2003b;this study) highlight the paraphyletic nature ofthe Notopithecinae. Ameghino (1897) coinedthe name Notopithecidae to encompass solelyNotopithecus adapinus, a taxon he consideredancestral to ‘‘prosimians’’ and other primates.More credibly, within the same paper he alsorecognized the close relationship of Noto-pithecidae to the Interatheriidae (5 ‘‘Proty-potheriidae’’). Although the proposed link ofNotopithecus to Primates has long been out-moded, Riggs and Patterson (1935) persua-sively confirmed the affiliation of Notopithecusand interatheriids by virtue of the uniqueconfiguration of the jugal mentioned pre-viously. Attempting to highlight this anatomi-cal finding, Simpson (1945) proposed thesubfamily Notopithecinae (within Intera-theriidae) to encompass Notopithecus, Antepi-thecus, Transpithecus, and Guilielmoscottia—to emphasize their lack of derived featuressuch as hypsodont teeth, a taxonomic group-ing that remains in wide usage to this day (e.g.,McKenna and Bell, 1997).
Simpson (1967) provided a comprehensivesystematic treatment of Notopithecinae. His‘‘definition’’ (diagnosis—in current terminol-ogy) of the group listed a mix of cranial anddental characters, some being primitive fea-tures that also are typical of other typotheres(such as oldfieldthomasiids and archaeopithe-cids), while others are more likely synapo-morphic for Interatheriidae. No synapomor-phies uniting ‘‘notopithecines’’ to the exclu-sion of other interatheriids have yet beenidentified. We propose that non-interatheriineinteratheriids are more appropriately termed‘‘basal interatheriids’’ as this phrasing lacksany connotation of the monophyly of thosetaxa. The name Notopithecinae (and all of itsderivatives) may someday be useful if anyinteratheriid species prove to be more closelyrelated to Notopithecus than to other inter-atheriids. In such a case, this vintage namemight be linked to this newly recognizedclade.
24 AMERICAN MUSEUM NOVITATES NO. 3520

DISCUSSION
The two new taxa described here, Johnbellhatcheri and Ignigena minisculus, and thespecimen referred to Antepithecus brachyste-phanus, are derived from four geographicallydistant faunas (Tinguiririca, Tapado, Azufre,Cachapoal), adding to the burgeoning di-versity of fossil mammals known from theAbanico Formation of central Chile. Thiswork provides new information about thesystematics of basal interatheriids, and bio-chronological age estimates for localities out-side of the original Abanico Formation fossilmammal discoveries in the Portozuelo ElFierro area near Termas del Flaco in the RıoTinguiririca valley.
As one of the youngest known basalinteratheriids, Johnbell hatcheri underscoresthe transitional nature of the TinguiriricaFauna, wherein ‘‘archaic’’ Paleogene taxaand later diverging clades co-occur (Wyss etal., 1994; Flynn et al., 2003). The occurrenceof Johnbell hatcheri at the Rıo CachapoalLocality (Cachapoal Fauna) indicates a tenta-tive Tinguirirican (Flynn et al., 2003) ageassignment for these deposits, consistent withpreliminary evidence provided by other faunalelements (Flynn and Wyss 2004) and geo-logical data (Charrier et al., 1996, 1997, inprogress).
The age of the Tapado Fauna is somewhatless certain, but the presence of a basalinteratheriid, Ignigena minisculus, is compati-ble with a Casamayoran age estimate (Wyss etal., 1994, 1996; Flynn et al., 2005) for thislocality based on other taxa.
Our referral of a specimen from the AzufreLocality to Antepithecus brachystephanus sug-gests a Casamayoran age for these deposits,and is the first report of this taxon outside ofPatagonia. Fieldwork in the Rıo Azufrespecifically targeted western exposures of theAbanico Formation, the structural regionfrom which the oldest fauna known previously(Tapado Fauna, Rıo Tinguiririca valley) isderived, in an attempt to uncover even olderfaunas, thereby better constraining the age ofinitiation of basin subsidence. Because thisprospecting trip had an auxiliary educationalobjective (see acknowledgments), we chose toexplore a region that had never previouslyyielded fossils, making an explicit prediction
about the age of the unit in this region. Basedon our experience in the Rıo Tinguiriricavalley, we used aerial photographs to tracelarge-scale lithologic units from that valley tothe Rıo Azufre valley. This led to the pre-diction that the targeted strata would likelybe correlative with, or somewhat older than,the Tapado Fauna (, Casamayoran in age;and older than the youngest known faunas[Tinguirirican SALMA] cropping out at theeastern end of the Tinguiririca valley). TheCasamayoran age indicated by SGOPV 3604(Antepithecus brachystephanus) confirms thatprediction. Radioisotopic dating and addi-tional paleontological collecting from the RıoAzufre region will help refine the geochronol-ogy of some of the oldest Abanico Formationstrata currently known.
The two new basal interatheriid taxa fromthe Abanico Formation are substantiallysmaller than forms known from Patagonia(fig. 15). The diminutive nature of theseAndean forms is shared uniquely withPunapithecus minora, the smallest interatheriidknown, recently described from Mustersanage localities in the Salta and CatamarcaProvinces of northern Argentina (Lopez andBond, 1993, 1995; Alonso et al., 1988). Allthree of these taxa are from latitudes wellnorth (Tinguiririca ,35u S; Punapithecus 5,25–26u S) of those yielding other basalinteratheriids such as Notopithecus, Trans-pithecus, and Guilielmoscottia, which areknown solely from Patagonian localities
Fig. 15. Bivariate plot of basal interatheriids(excluding Eopachyrucos) using upper first molardimensions.
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 25

(e.g., Gran Barranca south of Lake ColhueHuapi, ,46u S). Although only three suchdiminutive basal interatheriids are currentlyknown, it is tempting to posit that a provincialbiogeographic pattern existed during thePaleogene, with smaller basal interatheriidsto the north and larger forms to the south. Arelated possibility, amenable to testing withfuture discoveries, is the possibility of clinalvariation (large to small body size; Bergman’sRule) from Patagonia to central Chile tonorthern Argentina. Our phylogenetic analysisof Interatheriidae does not identify a separateclade of basal interatheriids diagnosed bysmall size, indicating perhaps that small sizeappeared convergently in these geographicallydisjunct species, potentially in response tosimilar environmental pressures. Alter-natively, depending on how existing poly-tomies are resolved, very small size mighthave characterized part of the main inter-atheriid stem lineage.
There appears to be a temporal componentto the distribution of small-bodied taxa aswell, given their restriction to Mustersan andTinguirirican SALMA deposits, whereassomewhat larger basal forms are found inlocalities of Casamayoran through Mustersanage, and large-bodied interatheriines occuronly in Tinguirirican and younger sites.
Our work here does not fully resolveuncertainty about whether Antepithecus andNotopithecus are distinct, but it does bear onthe issue. The morphological differences be-tween these two taxa are slight, withAntepithecus differing from Notopithecus inonly a single character state in our charactermatrix (table 2: character 13: character state 1:Antepithecus has a lingual sulcus on the uppermolars that persists longer than inNotopithecus [scored as ‘‘0’’], due to moreprominent hypocone and protocone inAntepithecus), placing Antepithecus withina clade including all interatheres exceptNotopithecus. Comparison beyond the denti-tion is hindered by lack of cranial materialfor Antepithecus. Based on these results, weconcur with Simpson’s (1967) view that thesetaxa, although very similar, are nonethelessdistinct. SGOPV 3604 scored identically toAntepithecus in our data matrix and thusis positioned at the same polytomy as
Antepithecus on our consensus tree. Al-though this provides no positive evidence ofspecial relationship between Antepithecus andSGOPV 3604 (i.e., a pairing of these taxa tothe exclusion of other interatheres), it does notpreclude this possibility either. Since SGOPV3604 from the Azufre Locality cannot bedistinguished from Antepithecus brachystepha-nus in any morphological attributes, weconservatively refer this specimen to thatCasamayoran taxon.
The phylogenetic analysis presented heresubstantiates the existence of a clade encom-passing Santiagorothia chiliensis plus all otherpreviously recognized interatheriines (exceptEopachyrucos), validating the phylogeneticnaming of that clade (Hitz et al, 2000). Twoclades roughly equating to the ‘‘Interatheriumand allies’’ and ‘‘Protypotherium and allies’’suggested by Cifelli (1985) occur in ourconsensus tree of 70 steps. These two cladescollapse, however, in the consensus treeresulting from trees #71 steps, arguing thatis it probably premature to name themformally.
CONCLUSION
The work detailed here may be summarizedby these concluding points:
N Two new basal interatheriid taxa (non-interatheriine interatheriids) are recognizedfrom the Andean Main Range of centralChile, Johnbell hatcheri and Ignigena miniscu-lus. Johnbell hatcheri derives from the samehorizons of the Abanico Formation as thoseproducing the Tinguiririca Fauna (Tingui-rirican SALMA), near Termas del Flaco.Ignigena minisculus also derives from theAbanico Formation in the Rıo Tinguiriricavalley (Tapado Fauna, ,14 km W of Termasdel Flaco) but from older parts of thestratigraphic section, and is estimated topertain to the Casamayoran SALMA.
N We also refer SGOPV 3451, a maxillaryfragment from the Rıo Cachapoal drainage,100 km N of the fossiliferous strata in theTinguiririca River valley, to Johnbell hatcheri.This is first time an Andean fossil mammalfrom from outside of the type area of theTinguiririca Fauna (upper Rıo Tinguiriricavalley) has been referred to as a member of
26 AMERICAN MUSEUM NOVITATES NO. 3520

that fauna at the species level. The occurrenceof Johnbell hatcheri at the Rıo CachapoalLocality suggests a tentative Tinguirirican(Flynn et al., 2003) age for the CachapoalFauna and at least part of these deposits.
N We refer a specimen recovered fromwestern exposures of the Abanico Formationalong the Rıo Azufre (Azufre Fauna), imme-diately north of the Rıo Tinguiririca drainageand west of Volcan Tinguiririca, to thebasal interatheriid Antepithecus brachystepha-nus (SGOPV 3604). Antepithecus brachyste-phanus has previously been reported only fromPatagonian deposits of the ‘‘late’’ Barrancansubage of the Casamayoran SALMA.
N We provide the first phylogenetic defini-tion of the name Interatheriidae, applying anapomorphy-based definition (‘‘the notoungu-late clade stemming from the ancestor dis-playing a splint-like jugal on the zygomaticarch, wherein this element is ‘sandwiched’between the maxilla and squamosal andexcluded from the orbit’’) to best preservehistorical conceptions of this major notoun-gulate clade. Basal interatheriids form a highlypectinate paraphyletic assemblage relative tointeratheriines; hence, we eschew the name‘‘Notopithecinae’’ in favor of the informalphrase ‘‘basal interatheriids’’, which has theadvantage of lacking phylogenetic connota-tions of monophyly. Both of the new taxa,Johnbell hatcheri and Ignigena minisculus,are shown to be closely related to (but arenot members of) the Interatheriinae (as de-fined by Hitz et al., 2000), but they themselvesdo not appear to constitute a monophyleticgroup. Discovery of these two new taxasignificantly increases the known diversityof basal interatheriids, and their incorpora-tion into phylogenetic analyses enhances un-derstanding of character transformationswithin this important herbivorous ‘‘ungulate’’clade.
N Both new taxa are smaller than allpreviously known interatheres except Puna-pithecus. Johnbell hatcheri, Ignigena miniscu-lus, and Punapithecus occur only in localitieswell north of the Patagonian deposits thathave yielded all other basal interatheriidsknown. This pattern may reflect minor geo-graphic provincialism or clinal variation, withsmaller-bodied basal interatheriids inhabiting
more northerly latitudes than their larger-bodied relatives, until the Tinguririrican.Following the Tinguirircan, all interatheretaxa are larger-bodied.
ACKNOWLEDGMENTS
We acknowledge the financial support ofthe National Science Foundation (DEB9020213, 9318126, 9317943), the NationalGeographic Society (3932–88), the Depart-ment of Geological Sciences, University ofCalifornia, Santa Barbara (graduate studentresearch grant), the American Museum ofNatural History (collections study grant), andthe Field Museum of Natural History (visitingscholar funds). Our work has had thestrong backing of the Museo Nacional deHistoria Natural and the Consejo deMonumentos Nacionales, Santiago, Chile.Daniel Frassinetti and Marıa Eliana Ramırezhave been particularly instrumental in facili-tating that support. Numerous colleaguesparticipated in the fieldwork, discussions,and earlier publications essential to thegeneration of this study, including MichaelNovacek, Mark Norell, Malcolm McKenna,Carl Swisher, Daniel Frassinetti, DanielBryant, Andres Charrier, Jacques Christinet,Gina Gould, Jin Meng, William Simpson,Claudio Veloso, and Franyo Zapatta.Reynaldo Charrier has been our close col-league and collaborator on virtually all aspectsof our work in the Chilean Andes, and weespecially acknowledge him for discovery ofthe Azufre Antepithecus specimen described inthis paper. Exploration of the Rıo Azufrevalley, undertaken as part of our long-termresearch investigations in this region, wasdone as an episode of Bill Kurtis’ NewExplorers television series highlighting thiswork, with support from the US NationalScience Foundation (ISE/BIO supplement toDEB 9317943 to JJF). Further thanks go tothe Department of Geology and staff at theField Museum, for their long-term supportand especially for the exceptional preparationwork on the Tinguiririca, Tapado, and Azufrefossils by Andrew Lehman, Bob Masek, SteveMcCarroll, and Bill Simpson. We thankMarlene Donnelly for the exceptional draw-ings. We also thank Mariano Bond, Darin
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 27

Croft, and Marcelo Reguero for their help andexpertise on notoungulates, Rosendo Pascualfor access to comparative collections at theMuseo de La Plata, and Darin Croft and ananonymous referee for thorough reviewsand helpful critique. JF was assisted bya John S. Guggenheim Memorial Founda-tion fellowship during preparation of thismanuscript.
REFERENCES
Alonso, R.N., W.D. Berman, M. Bond, A.A.Carlini, R. Pascual, and M.A. Reguero. 1988.Vertebrados Paleogenos de la Puna Austral:sus aportes a la evolucion biogeographica.V Jornadas Argentinas Paleontologıa deVertebrados Resumenes: 38–39. La Plata.
Ameghino, F. 1882. Catalogo de la provincia deBuenos Aires en la Exposicion ContinentaleSud-America, March, 1882.
Ameghino, F. 1887. Observaciones generales sobreel orden de mamıferos estinguidos Sud-Americanos. Llamados, toxodontes (Toxo-dontia) y sinopsis de los generos y especieshasta ahora conocidos. Anales Museo de LaPlata Buenos Aires, vol. 6: xxxii + 1027 pp.,atlas, 98 plates.
Ameghino, F. 1896. Notas sobre cuestiones degeologıa y paleontologıa Argentinas. Boletindel Instituto Geographica Argentino 17: 92.
Ameghino, F. 1897. Mammiferes cretaces del’Argentine (Deuxieme contribution a la con-naissance de la faune mammalogique descouches a Pyrotherium. Boletin del InstitutoGeographica Argentino 8: 406–429, 431–531.
Ameghino, F. 1901. Notices preliminaires sur desongules nouveau des terrains cretaces dePatagonie. Boletin Academie NacionalCiencias Cordoba 16: 350–426.
Ameghino, F. 1902. Premiere contribution a laconnaissance de la faune mammalogique descouches a Colpodon. Boletin AcademieNacional Ciencias. Cordoba 17: 71–138.
Ameghino, F. 1904. Nuevas especies de mamıferos,cretaceos y terciaros de la Republica Argentina.Anales de la Sociedad Cientifica Argentino 57:162–175: 58: 35–41, 56–71, 182–192, 225–290.
Bond, M., G. Lopez, and M. Reguero. 1996.‘‘Astraponoteen plus superieure’’ of Ameghino:another interval in the Paleogene record ofSouth America. Journal of Vertebrate Pale-ontology 16(3, suppl.): 23A.
Bond, M., M. Reguero, G. Lopez, A.A. Carlini, F.Goin, R.H. Madden, M.G. Vucetich, and R.F.Kay. 1997a. The ‘‘Astraponoteen plus super-
ieure’’ (Paleogene) in Patagonia. Ameghiniana34(4): 533.
Bond, M., G. Lopez, and M. Reguero. 1997b.Rocas Bayas, una localidad fosilıferaPaleogena de la Provincia de Rıo Negro,Republica Argentina. Ameghiniana 34(4): 533.
Charrier, R., A.R. Wyss, M.A. Norell, J.J. Flynn,M.J. Novacek, M.C. McKenna, C.C. Swisher,III, D. Frasinetti, and P. Salinas. 1990.Hallazgo de mamıferos fosiles del TerciarioInferior en el sector de Termas del Flaco,Cordillera Principal, Chile Central: implica-ciones paleontologicas, estratigraphicas y tec-tonicas. Segundo Simposio sobre le Tercario deChile, Concepcion, Actas: 73–84.
Charrier, R., A.R. Wyss, J.J. Flynn, C.C. Swisher,III, M.A. Norell, F. Zapatta, M.C. McKenna,and M.J. Novacek. 1996. New evidence oflate Mesozoic-early Cenozoic evolution ofthe Chilean Andes in the upper TinguiriricaValley (35u S), central Chile. Journal ofSouth American Earth Sciences 9(5/6): 393–422.
Charrier, R., O. Baeza, S. Elgueta, J.J. Flynn, P.Gans, S.M. Kay, N. Munoz, A.R. Wyss, and E.Zurita. 2002. Evidence for Cenozoic extension-al basin development and tectonic inversion inthe southern central Andes, Chile (33u–36uS.L.). Journal of South American EarthSciences 15(1): 117–139.
Charrier, R., M. Bustamante, D. Comte, S.Elgueta, J.J. Flynn, N. Iturra, N. Munoz, M.Pardo, R. Thiele, and A.R. Wyss. 2005. TheAbanico extensional basin: Regional extension,chronology of tectonic inversion, and relationto shallow seismic activity and Andean uplift.Neues Jahrbuch fur Geologie und Palaon-tologie, Abhandlungen 236(1–2): 43–77.
Cifelli, R.L. 1985. South American ungulateevolution and extinction. In F.G. Stehli andS.D. Webb (editors), The great Americaninterchange: 249–266. New York: Plenum.
Cifelli, R.L. 1993. Phylogeny of native SouthAmerican ungulates. In F.S. Szalay, M.J.Novacek, and M.C. McKenna (editors),Mammal phylogeny, placentals, 195–216. NewYork: Springer-Verlag.
Croft, D.A., M. Bond, J.J. Flynn, M. Reguero, andA.R. Wyss. 2003. Large archaeohyracids(Typotheria, Notoungulata) from centralChile and Patagonia, including a revision ofArchaeotypotherium. Fieldiana (Geology),n.ser. 49: 1–38.
De Queiroz, K., and J. Gauthier. 1990. Phylo-geny as a central principle in taxonomy:phylogenetic definitions of taxon names.Systematic Zoology 39: 307–322.
28 AMERICAN MUSEUM NOVITATES NO. 3520

De Queiroz, K., and J. Gauthier. 1992.Phylogenetic taxonomy. Annual Review ofEcological Systematics 23: 449–480.
Flynn, J.J., D.A. Croft, R. Hitz, and A.R. Wyss.2005. The Tapado Fauna (?CasamayoranSALMA), Abanico Formation, TinguiriricaValley, Central Chile. Journal of VertebratePaleontology 25(3, suppl.): 57A–58A.
Flynn, J.J., M.A. Norell, C.C. Swisher, III, andA.R. Wyss. 1991. Pre-Deseadan, post-Mustersan mammals from central Chile: anupdate. Journal of Vertebrate Paleontology11(3, suppl.): 29A.
Flynn, J.J., and C.C. Swisher, III. 1995. CenozoicSouth American Land Mammal Ages: correla-tion to global geochronologies. In Geo-chronology, time scales and global stratigraph-ic correlation, SEPM Special PublicationNo. 54: 319–333. Tulsa, Oklahoma.
Flynn, J.J., and A.R. Wyss. 1999. New marsupialsfrom the Eocene-Oligocene transition of theAndean Main Range, Chile. Journal ofVertebrate Paleontology 19(3): 533–549.
Flynn, J.J., and A.R. Wyss. 2004. A polydolopinemarsupial skull from the Cachapoal Valley,Andean Main Range, Chile. Bulletin of theAmerican Museum of Natural History 285:80–92.
Flynn, J.J., A.R. Wyss, R. Charrier, and C.C.Swisher, III. 1995. An Early Miocene anthro-poid skull from the Chilean Andes. Nature 373:603–607.
Flynn, J.J., A.R. Wyss, D.A. Croft, and R.Charrier. 2003. The Tinguiririca Fauna, Chile:biochronology, paleoecology, biogeography,and a new earliest Oligocene South AmericanLand Mammal ‘‘Age’’. Palaeogeography, Pala-eoclimatology, Palaeoecology 195: 229–259.
Goin, F.J., and A.M. Candela. 1997. NewPatagonian marsupials from Ameghino’s‘‘Astraponoteen Plus Superieure’’ (Post-Mustersan/Pre-Deseadan Age). Journal ofVertebrate Paleontology 17(3, suppl.) [un-num-bered insertion to abstract volume].
Hitz, R. 1995. Typothere (Notoungulata) phyloge-ny and proposed taxonomic revisions. Journalof Vertebrate Paleontology 15(3, suppl.): 34.
Hitz, R. 1997. Contributions to South Americanmammalian paleontology: new interatheres(Notoungulata) from Chile and Bolivia,Typothere (Notoungulata) phylogeny, andPaleosols from the Late Oligocene Salla beds.Ph.D. dissertation, University of California,Santa Barbara.
Hitz, R.B., M.A. Reguero, A.R. Wyss, andJ.J. Flynn. 2000. New interatheriines(Interatheriidae, Notoungulata) from the
Paleogene of central Chile and southernArgentina. Fieldiana Geology, n.ser. 42: 1–26.
Kay, R.F., B.J. MacFadden, R.H. Madden, H.Sandeman, and F. Anaya. 1998. Revised age ofthe Salla beds, Bolivia, and its bearing on theage of the Deseadan South American LandMammal ‘‘Age’’. Journal of VertebratePaleontology 18: 189–199.
Kay, R.F., R. Madden, M., M.G. Vucetich, A.A.Carlini, M.M. Mazzoni, G.H. Re, M. Heizler,and H. Sandeman. 1999. Revised age of theCasamayoran South American Land Mammal‘Age’: climatic and biotic implications.Proceedings of the National Academy ofSciences (US) 96(23): 13235–13240.
Kearny, M., and J.M. Clark. 2003. Problems due tomissing data in phylogenetic analyses includingfossils: a critical review. Journal of VertebratePaleontology 23(2): 263–274.
Loomis, F.B. 1914. The Deseado Formation ofPatagonia. Concord, NH: Rumford Press,232 pp.
Lopez, G., and M. Bond. 1993. Consideracionessobre los ungulados Paleogenos de Antofagastade la Sierra (Catamarca, Argentina).Ameghiniana 30(3): 321–356.
Lopez, G., and M. Bond. 1995. Un nuevonotopithecine (Notoungulata, Typotheria) delTerciario Inferior de la Puna, Argentina. StudiaGeologica Salmanticensia 31: 87–99.
MacFadden, B.J., K.E. Campbell, Jr, R.L. Cifelli,O. Siles, N.M. Johnson, C.W. Maeser, andP.K. Zeitler. 1985. Magnetic polarity stratigra-phy and mammalian fauna of the Deseadan(Late Oligocene-Early Miocene) Salla beds ofnorthern Bolivia. Journal of Geology 93(3):223–250.
Marshall, L.G., and R.L. Cifelli. 1990. Analysis ofchanging diversity patterns in Cenozoic LandMammal Age faunas, South America.Palaeovertebrata, Montpellier 19(4): 169–210.
Marshall, L.G., R.L. Cifelli, R.E. Drake, and G.Curtiss. 1986. Vertebrate paleontology geology,and geochronology of the Tapera de Lopez andScarritt Pocket, Chubut Province, Argentina.Journal of Paleontology 60(4): 920–951.
Marshall, L.G., R. Hoffstetter, and R. Pascual.1983. Mammals and stratigraphy: geochronol-ogy of the continental mammal-bearingTertiary of South America. Paleovertebrata,Memoire Extraordinaire 1983: 1–93.
McKenna, M.C., and S.K. Bell. 1997. Classificationof mammals above the species level. New York:Columbia University Press.
Nixon, K.C., and Q.D. Wheeler. 1992. Extinctionand the origin of species. In M.J. Novacek andQ.D. Wheeler (editors), Extinction and phylog-
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 29

eny, 119–143. New York: Columbia UniversityPress.
Novacek, M. 1989. Higher mammal phylogeny:a morphological-molecular synthesis. In B.Fernholm, K. Bremer, and H. Jornvall (edi-tors), The hierarchy of life: 421–435.Amsterdam: Elsevier Press.
Novacek, M. 1992a. Fossils, topologies, missingdata, and the higher level phylogeny ofeutherian mammals. Systematic Biology 41(1):58–73.
Novacek, M. 1992b. Fossils as critical data forphylogeny. In M.J. Novacek and Q.D. Wheeler(editors), Extinction and phylogeny: 46–88.New York: Columbia University Press.
Novacek, M.J., A.R. Wyss, D. Frassinetti, and P.Salinas. 1989. A new ?Eocene mammal faunafrom the Andean Main Range. Journal ofVertebrate Paleontology 9(3, suppl.): 34A.
Pascual, R. 1965. Un nuevo Condylarthra(Mammalia) de edad Casamayorense de Pasode los Indios (Chubut, Argentina). Brevesconsideraciones sobre la edad Casamayorense.Ameghiniana 4: 57–65.
Pascual, R., P. Bondesio, M.G. Vucetich, G.J.Scillato Yane, M. Bond, and E.P. Tonni. 1984.Vertebrados fosiles cenozoicos. Capıtulo II. IXCongreso Geologico Argentino, San Carlos deBariloche. Relatorio: 439–461.
Patterson, B. 1952. Catalogo de los Typotheria delColhuehuapiano y Deseadiano. Unpublishedmanuscript on file, Seccion Paleontologıa(Vertebrados), Museo Argentino de CienciasNaturales ‘‘Bernadino Rivadavia’’. BuenosAires, pp. 1–4.
Price, L.I., and C. Paula Couto. 1950. Vertebradosterrestres do Eoceno na bacia calcaria deItaboraı, Brasil. Anais Do Segundo Con-gresso Panamericano de Engenharia deMinas e Geologia, Rio de Janeiro, 3: 149–173(Dated 1946 but not published until 1950).
Reguero, M. 1999. El problema de las relacionessistematicas y filogeneticas de los Typotheria yHegetotheria (Mammalia, ?Notoungulata):analisis de los taxones de Patagonia de laEdad-mamıfero Deseadense (Oligoceno). Ph.D.dissertation, Universidad de Buenos Aires,Buenos Aires.
Reguero, M., M. Bond, and G. Lopez. 1996.Campanorco inauguralis (Typotheria, Noto-ungulata): an approach to the phylogeny ofthe Typotheria. Journal of VertebratePaleontology 16(3, suppl.): 59A.
Reguero, M.A., D.A. Croft, J.J. Flynn, and A.R.Wyss. 2003a. Small archaeohyracids fromChubut, Argentina and central Chile: implica-tions for trans-temporal Andean correlation.Fieldiana: Geology, n.ser. 48: 1–17.
Reguero, M.A., M. Ubilla, and D. Perea. 2003b. Anew species of Eopachyrucos (Mammalia,Notoungulata, Interatheriidae) from the LateOligocene of Uruguay. Journal of VertebratePaleontology 23(2): 445–457.
Riggs, E.S., and B. Patterson. 1935. Descriptions ofsome notoungulates from the Casamayor(Notostylops) beds of Patagonia. Proceedingof the American Philosophical Society 75:163–215.
Simpson, G.G. 1932. New or little-known ungulatesfrom the Pyrotherium and Colpodon beds ofPatagonia. American Museum Novitates 576:1–13.
Simpson, G.G. 1945. The principles of classificationand a classification of mammals. Bulletin of theAmerican Museum of Natural History 85:1–350.
Simpson, G.G. 1967. The beginning of the age ofmammals in South America. Part 2. Bulletin ofthe American Museum Natural History 137:1–260.
Sinclair, W.J. 1909. Typotheria of the Santa Cruzbeds. In W.B. Scott (editor), Reports of thePrinceton University expeditions to Patagonia,1896–1899, 4: 1–110.
Stirton, R.A. 1953. A new genus of interathere fromthe Miocene of Columbia. University ofCalifornia Publications in Geological Sciences29: 265–348.
Swofford, D.L. 2002. PAUP*. PhylogeneticAnalysis Using Parsimony (*and OtherMethods). Version 4. Sunderland, MA:Sinauer Associates.
Wyss, A.R., R. Charrier, and J.J. Flynn. 1996.Fossils mammals as a tool in Andean stratig-raphy: dwindling evidence of Late Cretaceousvolcanism in the south central Main Range.Paleobios 17(2–4): 13–27.
Wyss, A.R., and J.J. Flynn. 2004. A polydolopinemarsupial skull from the Cachapoal Valley,Andean Main Range, Chile. AmericanMuseum Novitates 285: 80–92.
Wyss, A.R., J.J. Flynn, M.A. Norell, C.C. Swisher,III, R. Charrier, M.J. Novacek, and M.C.McKenna. 1993. South America’s earliestrodent and recognition of a new interval ofmammalian evolution. Nature 365: 434–437.
Wyss, A.R., J.J. Flynn, M.A. Norell, C.C. Swisher,M.J. Novacek, M.C. McKenna, and R.Charrier. 1994. Paleogene mammals from theAndes of central Chile: a preliminary taxo-nomic, biostratigraphic, and geochronologicassessment. American Museum Novitates3098: 1–31.
Wyss, A.R., M.A. Norell, J.J. Flynn, M.J.Novacek, R. Charrier, M.C. McKenna, C.C.Swisher, III, D. Frassinetti, and M. Jin. 1990.
30 AMERICAN MUSEUM NOVITATES NO. 3520

A new early Tertiary mammal fauna fromcentral Chile: implications for Andean stratig-raphy and tectonics. Journal of VertebratePaleontology 10(4): 518–522.
Wyss, A.R., M.A. Norell, M.J. Novacek, and J.J.Flynn. 1992. New ?early Tertiary localitiesfrom the Chilean Andes. Journal of Verte-brate Paleontology 12(3, suppl.): 61A.
APPENDIX 1
DESCRIPTION OF CHARACTERS IN PHYLOGENETIC ANALYSIS AND DATA MATRIX
1. I1 size: larger than other anterior teeth (0); greatly enlarged (1)2. I2-C shape: I2 laterally compressed, I3-C ovoid/conical (0); all laterally compressed (1)3. C size: large (0); reduced (1)4. I2-3 size: subequal (0); reduced, with I3 smaller than I2 (1)5. I2-C exterior face: convex or vague anterior swelling (0); distinct vertical anteroexternal
ridge (1)6. P1 vertical anteroexternal ridge: no ridge (0); ridge present (1)7. P2-4 parastyle/paracone inflection: shallow (0); deep (1)8. P3-4 fossettes and internal sulcus: anterior, posterior, and lingual fossette moderately to
strongly persistent (0); lingual remains open as a sulcus, others disappear rapidly with wear (1)9. P2-4 metacone ectoloph: moderate to strong metacone ectoloph column (0); metacone
ectoloph very low or almost flat (1)10. P3-4 dimensions: W . L or with mild anteroexternal expansion (0); L . W, rectangular
shape (1)11. P2-4 cingulum: anterior cingulum low on crown, posterior about midway up crown (0);
anterior cingulum absent, posterior high on crown, merging early in wear (1)12. M1-3 paracone/parastyle inflection: distinct but shallow (0); absent or reduced (1)13. M1-2 lingual sulcus and fossettes: anteroexternal, posteroexternal and lingual fossettes
present (0); moderately persistent lingual sulcus, other fossettes of varying longevity (1);completely persistent lingual sulcus, fossettes absent or very rapidly disappear with wear (2)
14. M3 hypocone: absent or diminutive (0); developed (1)15. M1-3 metacone ectoloph: distinct (0); very low or flat (1)16. M1-3 anterior cingulum: present and low on crown (0); absent (1)17. M1-3 dimensions: W . L or about equidimensional (0); L . W, rectangular shape (1)18. Crown height in posterior Ps and Ms: brachydont (0); hypsodont (1); hypseledont (2)19. i3 size: i3 . i2 (0); reduced (1)20. i1-3 shape: two shallow lingual grooves (0); i1-2 bifid labial and lingually (1)21. c size: larger than i1-3 (0); subequal to i1-3 (1); smaller than i1-2 or missing (2)22. c-p1 diastema: absent (0); present (1)23. p2-4 talonid development and relative size: p2 no talonid, p3 very small, p4 present but
smaller than trigonid (0); p2-3 distinct talonids but small, p4 smaller or subequal to trigonid (1);p2-4 display reduced talonids (2); p2 subequal to larger, p3-4 larger talonids (3)
24. p3-4 crown morphology: narrow labial sulcus, more posteriorly oriented lingual sulcus (0);two lobes, well defined labial and lingual sulci opposite one another (1)
25. Relative size of m1-3 trigonid and talonid: m1 talonid subequal to or slightly smaller thantrigonid, m2 talonid subequal to or slightly larger to trigonid, m3 talonid larger than trigonid (0);all talonids larger than trigonids (1)
26. m1-3 crown morphology: not bilobed, lingual sulcus broad and extends posteriorly (0);bilobed, persistent labial and lingual sulci directly opposite one another (1)
27. p3-4 molarization: p4 approaching molariform, p3 considerably smaller than the molars(0); p4 molariform, p3 enlarged but smaller than molars (1)
28. m1-3 trigonid shape: anterior margin broad (0); anterior margin short, producinga triangular appearing lobe (1)
29. m3 third lobe: small (0); larger with a distinct labial groove (1)
2006 HITZ ET AL.: NEW BASAL INTERATHERIIDS, CHILE 31

30. Role of jugal in zygoma: included in orbit (0); excluded from orbit by maxilla (1)31. Position of nasofrontal suture: flush or posterior to anterior margin of orbit (0); well
anterior to anterior margin of orbit (1)32. Nasofrontal suture shape: rounded, nasal projecting into frontal (0); nearly straight
transversely (1); nasal trends posteriorly from the median (2); nasal trends posteriorly from themedian but is jagged with small posteriorly projecting processes (3)
33. Descending process on zygoma: none (0); small (1); large (2)34. Dorsal-posterior maxillary process: dorsal process present but is not excluded from orbit
rim (0); dorsal process present and is excluded from orbit margin by an anteriorly projectingsliver of frontal (1)
35. Rostrum length: rostrum short compared to anteroposterior length of skull, ,35% of totalcranium length (0); modest lengthening of rostrum, $35% of total cranium length (1)
36. Angle of zygoma with face, at attachment site: low angle (#45u) (0); steep angle (.45u and,90u) (1); perpendicular (90u)(2)
37. Muzzle shape and dental arcade: ventrally broader than dorsally, posterior broader thananterior, dental arcade forms a ‘‘triangle’’ (0); muzzle is narrower (ventral portion no broaderthan dorsal) and cheektooth rows are convex labially (1)
38. Dorsal-posterior process on external premaxillary: present (0); absent (1)39. Palatine notch position: reaches mid to anterior M3 region (0); reaches posterior M3 region
(1)40. Posterior bulla: adjacent to paraoccipital process (0); posterior bulla laps up onto
paraoccipital process (1)41. Glenoid fossa: shallow, wide mediolaterally (0); deeply excavated and narrow (1)42. Postglenoid process and foramen: broad process with uncompressed foramen; glenoid fossa
encroaches medioposteriorly on bulla/meatus (0); process is sharper and narrower separatingfossa from meatus (1); process severely compressed as is foramen (2)
43. Size, as estimated from molar dimensions: moderate to large (M1 AP and TR dimensionsboth $4 mm) (0); very small (M1 AP and TR dimensions both ,4 mm) (1)
32 AMERICAN MUSEUM NOVITATES NO. 3520


















