
Neurotransmitery - [email protected] RNDr. Alexander Kiss, DrSc....
285
Neurotransmitery Boris Mravec et al. Neurofyziologické laboratórium Ústavu patologickej fyziológie LFUK v Bratislave
Transcript of Neurotransmitery - [email protected] RNDr. Alexander Kiss, DrSc....

Neurotransmitery
Boris Mravec et al.
Neurofyziologické laboratórium Ústavu patologickej fyziológie
LFUK v Bratislave

Mravec B. et al.
Neurotransmitery
SAP, Bratislava 2007, 284 strán
Publikácia vznikla s finančným prispením
SAVOL - Spoločnosti autorov vedeckej a odbornej literatúry
Nadácie Slovenského plynárenského priemyslu, a.s.
Eli Lilly
Janssen-Cilag
Jazyková úprava PhDr. Helena Bernadičová
Sadzba Strelka
Tlač FABER 2007
c© SAP – Slovak Academic Press, spol. s r. o. 2007
c© Boris Mravec 2007
Všetky práva v zmysle platného „autorského zákonaÿpatria autorovi a vydavateľovi.
ISBN 80–8095–005–9

Recenzenti
Prof. MUDr. Jozef Maršala, DrSc.Neurobiologický ústav SAV, Košice
Doc. MUDr. Jaromír Mysliveček, PhD.Fyziologický ústav 1. Lekárskej fakulty Univerzity Karlovej, Praha

Autor ∼ editor
MUDr. Boris Mravec, PhD.Neurofyziologické laboratórium Ústavu patologickejfyziológie LFUK a Laboratórium funkčnej neuromorfológieÚstavu experimentálnej endokrinológie [email protected]
Autori ∼ prispievatelia
Doc. MUDr. Marián Bernadič, CSc.Ústav patologickej fyziológie LFUK, [email protected]
Prof. MUDr. Ivan Hulín, DrSc.Ústav patologickej fyziológie LFUK, [email protected]
RNDr. Alexander Kiss, DrSc.Laboratórium funkčnej neuromorfológie Ústavu experimentálnejendokrinológie SAV, [email protected]
RNDr. Richard Kvetňanský, DrSc.Laboratórium pre výskum stresu Ústavu experimentálnejendokrinológie SAV, [email protected]
MUDr. Barbara KukanováLaboratórium farmakologickej neuroendokrinológie Ústavuexperimentálnej endokrinológie SAV, [email protected]
MUDr. Ján Pečeňák, CSc.Psychiatrická klinika LFUK a FN, [email protected]

Predhovor
Až doteraz sme hovorili iba o uvoľňovaní látok z nervových zakon-
čení. To znamená, že uvoľnené neuronálne látky pochádzajú z nervo-
vých zakončení. Pre porozumenie podstaty neuronálnych funkcií je však
dôležité vedieť, čo presne si treba pod týmto uvoľňovaním predstaviť.
Existujú nasledujúce možnosti: buď sa tieto látky v pokojovom stave
nenachádzajú v nervových zakončeniach, vytvárajú sa iba v dôsledku
nervovej stimulácie a takto vytvorené difundujú, alebo sa vyskytujú už
aj v pokojovom stave, ale môžu difundovať iba po stimulácii. Pokiaľ ide
o tvorbu neuronálnych látok v nervoch, je isté, že tu môže prebiehať.
Otto Loewi, Nobelova prednáška, 1936
Dávnu túžbu vysvetliť procesy spojené s prenosom informácií medzijednotlivými neurónmi, ako aj medzi neurónmi a efektorovými bunkamisa podarilo čiastočne „naplniťÿ v prvej polovici 20. storočia. Experimen-tálne sa dokázal chemický základ týchto procesov. Zaviedol sa pojemneurotransmitery označujúci chemické látky, ktoré umožňujú komuniká-ciu medzi neurónmi. Zjednodušene možno povedať, že procesy spojenés ich syntézou, uvoľňovaním, účinkom a odstraňovaním sú „chémioumozguÿ, ktorá je spätá so širokým spektrom funkcií nervového sys-tému. Či už sú to najjednoduchšie senzorické vnemy, reflexné reakciealebo vedomie, všetky tieto procesy sú determinované činnosťou týchtochemických systémov. Narušenie neurotransmiterových systémov môžemať preto veľmi závažné dôsledky.Zavedenie sofistikovaných metodík umožnilo výrazný pokrok v po-
znaní jednotlivých „krokovÿ neurotransmisie. V slovenskej odbornej li-teratúre z posledných rokov nie je publikácia, ktorá by tieto poznatkyreflektovala. Preto sme sa rozhodli pripraviť monografiu, ktorá by savenovala procesom súvisiacim s neurotransmisiou.Na samom začiatku bol niekoľkostranový dokument, ktorý sa za-
oberal základnými charakteristikami neurotransmiterov. Na radu prof.MUDr. Ivana Hulína, DrSc., som však postupne začal jednotlivé častitohto zväzku rozširovať o nové kapitoly. Tak vznikol prvotný rukopis

s názvom Neurotransmitery. Ten už neobsahoval iba základné informá-cie o neurotransmisii, ale nachádzali sa v ňom aj kapitoly opisujúceúlohu jednotlivých neurotransmiterov vo fyziologických procesoch. Zatento výrazný posun od niekoľkostranového zväzku k rukopisu mono-grafie, ako aj za všestrannú podporu a cenné pripomienky úprimneďakujem profesorovi Hulínovi. Neskôr som oslovil kolegov z Lekárskejfakulty UK a z Ústavu experimentálnej endokrinológie SAV a s ich vý-razným prispením sa rukopis rozrástol o ďalšie kapitoly. Tak vzniklamonografia, v ktorej sa na základe najnovších poznatkov opisuje kom-plexnosť procesov spojených s neurotransmisiou za fyziologických, akoaj patologických podmienok. Po skompletizovaní rukopisu som požia-dal prof.MUDr. Jozefa Maršalu, DrSc., a doc.MUDr. Jaromíra Mysli-večka, PhD., o posúdenie rukopisu. Obaja moju prosbu prijali. V ob-lasti odbornej literatúry sa možno stretnúť s rôznymi typmi recenzií.Posudky recenzentov v odborných časopisoch, ktoré sú neoddeliteľnousúčasťou procesu akceptovania alebo odmietnutia rukopisu, často ok-rem hodnotenia obsahujú otázky, ktorých zodpovedanie autormi môževýrazne prispieť k vyššej kvalite publikácie. Pri posudzovaní rukopi-sov odborných textov sa však možno stretnúť aj s tým, že recenziaje často iba súhlasom, ktorý okrem pozitívneho ohodnotenia predlože-ného diela neobsahuje podnety a návrhy na zvýšenie kvality rukopisu.Recenzenti profesor Maršala a docent Mysliveček po detailnom preštu-dovaní rukopisu napísali okrem pozitívneho hodnotenia veľa podnetovpre doplnenie a upravenie rukopisu. Významne tým prispeli k výsled-nej podobe tejto monografie. Ich obdivuhodná dôslednosť je priliehavák vlastnej podstate „fungovaniaÿ neurotransmiterov a pre mňa je zavä-zujúcou skutočnosťou pre precíznosť v ďalšej práci.Nervový systém je tvorený obrovským počtom neurónov, podľa od-
hadov 1010–1011, ktoré navzájom komunikujú najmä prostredníctvomneurotransmiterov a neuromodulátorov. Rôznorodosť neurotransmite-rov je podkladom komplexnosti interakcií neurónov, ktorá umožňujeadekvátne spracovanie informácií v nervovom systéme.Pre pochopenie činnosti nervového systému je potrebné uvažovať
o centrálnom nervovom systéme ako o mimoriadne komplexnej štruk-túre. Pri snahe o pochopenie funkcií nervového systému často študujemeizolovane jednotlivé aspekty jeho činnosti. Príkladom je aj táto mono-grafia, ktorá sa venuje procesom prebiehajúcim v nervovom systéme,prenosu signálov prostredníctvom neurotransmiterov. Prenos signálov

neurotransmitermi predstavuje jeden zo základných procesov, ktorývšak neprebieha izolovane, ale je úzko spätý s ďalšími procesmi.Činnosť neurotransmiterových systémov je objasnená len čiastočne
a pre jej úplné pochopenie budú potrebné ďalšie experimentálne štúdie.Možno očakávať, že nové poznatky pozmenia náš pohľad na činnosťnervového systému a môžu prispieť aj k eventuálnemu pochopeniu ta-kých zložitých procesov, akými sú tie, ktoré sa zúčastňujú na vznikuľudského vedomia.V monografii je pre názornosť viacero schém, ktoré dopĺňajú text.
Komplexnosť neuronálnych systémov nedovoľuje zobraziť všetky ichzložky. Na základe súčasných poznatkov sú preto pre prehľadnosť zo-brazené iba podstatné súčasti opisovaných systémov.
Boris Mravec

Obsah
1 Štruktúra a činnosť neurónov . . . . . . . . . . . . . . . . 11Štruktúra neurónov . . . . . . . . . . . . . . . . . . . . . 11Prenos signálov v nervovom systéme . . . . . . . . . . . . 14
2 Neurotransmiterové systémy . . . . . . . . . . . . . . . . 16Komunikácia neurónov . . . . . . . . . . . . . . . . . . . 18Charakteristika neurotransmiterov . . . . . . . . . . . . . 20Základné skupiny neurotransmiterov . . . . . . . . . . . . 22Účinky neurotransmiterov . . . . . . . . . . . . . . . . . 33Nomenklatúra neurotransmiterových systémov mozgu . . . 35Neurotransmitery v periférii a v mozgu – úloha hematoen-cefalickej bariéry . . . . . . . . . . . . . . . . . . . . . 36Hematoencefalická bariéra . . . . . . . . . . . . . . . . 38Morfologické základy hematoencefalickej bariéry a štruk-túrne odlišnosti . . . . . . . . . . . . . . . . . . . . 38Funkčné charakteristiky hematoencefalickej bariéry,ktoré majú morfologický základ . . . . . . . . . . . . 39Hematolikvorová bariéra . . . . . . . . . . . . . . . . 39Likvoroencefalická bariéra . . . . . . . . . . . . . . . . 40Transportné mechanizmy hematoencefalickej bariéry . . 40Regionálne rozdiely hematoencefalickej bariéry – roz-diely vo vaskularizácii jednotlivých oblastí CNS . . . 41Ependýmové bunky . . . . . . . . . . . . . . . . . . . 41Cirkumventrikulárne orgány . . . . . . . . . . . . . . 41Ďalšie štruktúry mozgu s fenestrovanými kapilárami . . 43
Neurotransmitery z evolučného hľadiska . . . . . . . . . . 44
3 Malomolekulové neurotransmitery . . . . . . . . . . . . . 46Aminokyseliny . . . . . . . . . . . . . . . . . . . . . . . . 46Glutamát . . . . . . . . . . . . . . . . . . . . . . . . . 47Aspartát . . . . . . . . . . . . . . . . . . . . . . . . . 51Iné excitačné aminokyseliny . . . . . . . . . . . . . . . 52GABA (kyselina γ-aminomaslová) . . . . . . . . . . . . 53
6

Obsah 7
GHB (kyselina γ-hydroxymaslová) . . . . . . . . . . . 57Glycín . . . . . . . . . . . . . . . . . . . . . . . . . . 58
Acetylcholín a biogénne amíny . . . . . . . . . . . . . . . 60Acetylcholín . . . . . . . . . . . . . . . . . . . . . . . 61Katecholamíny . . . . . . . . . . . . . . . . . . . . . . 67Dopamín . . . . . . . . . . . . . . . . . . . . . . . . . 67Noradrenalín . . . . . . . . . . . . . . . . . . . . . . . 73Adrenalín . . . . . . . . . . . . . . . . . . . . . . . . . 77Sérotonín . . . . . . . . . . . . . . . . . . . . . . . . . 79Histamín . . . . . . . . . . . . . . . . . . . . . . . . . 82
Iné malomolekulové neurotransmitery . . . . . . . . . . . 85Puríny . . . . . . . . . . . . . . . . . . . . . . . . . . 85Neurosteroidy . . . . . . . . . . . . . . . . . . . . . . . 86Plynné neurotransmitery . . . . . . . . . . . . . . . . . 90Oxid dusnatý . . . . . . . . . . . . . . . . . . . . . . . 91Oxid uhoľnatý . . . . . . . . . . . . . . . . . . . . . . 94Sírovodík . . . . . . . . . . . . . . . . . . . . . . . . . 95Prostaglandíny . . . . . . . . . . . . . . . . . . . . . . 95Kanabinoidy . . . . . . . . . . . . . . . . . . . . . . . 96Iné neurotransmitery obsahujúce aminoskupinu . . . . . 97L-DOPA . . . . . . . . . . . . . . . . . . . . . . . . . 98Imidazolínový systém . . . . . . . . . . . . . . . . . . 99
4 Peptidové („veľkomolekulové“) neurotransmitery . . . . . 102Malomolekulové a peptidové neurotransmitery – spoločnásyntéza a uvoľňovanie z jedného neurónu . . . . . . . . 104
Vybrané peptidergické neurotransmitery . . . . . . . . . . 106Klasické neurohormóny CNS . . . . . . . . . . . . . . . 107Uvoľňujúce a inhibujúce hormóny hypotalamu . . . . . 109Deriváty proopiomelanokortínu (ACTH, MSH a iné) . . 113Endomorfíny 1 a 2 . . . . . . . . . . . . . . . . . . . . 115Tachykiníny (neurokiníny) . . . . . . . . . . . . . . . . 116Peptidy príbuzné s NPY . . . . . . . . . . . . . . . . . 117Peptidy zo skupiny sekretín-VIP-glukagón . . . . . . . 118Ďalšie neuropeptidy . . . . . . . . . . . . . . . . . . . 120

8 NEUROTRANSMITERY
5 Úloha neurotransmiterov v regulácii fyziologických procesov136Regulácia príjmu potravy . . . . . . . . . . . . . . . . . . 137Homeostáza telesných tekutín . . . . . . . . . . . . . . . . 146Bdenie a spánok . . . . . . . . . . . . . . . . . . . . . . . 147Učenie a pamäť . . . . . . . . . . . . . . . . . . . . . . . 152Autonómny nervový systém . . . . . . . . . . . . . . . . . 153Sympatikový nervový systém . . . . . . . . . . . . . . 154Parasympatikový nervový systém . . . . . . . . . . . . 157Enterický nervový systém . . . . . . . . . . . . . . . . 157Intrakardiálny nervový systém . . . . . . . . . . . . . . 159Nonadrenergický/noncholínergický (NANC) nervovýsystém . . . . . . . . . . . . . . . . . . . . . . . . . 159
Stresová reakcia . . . . . . . . . . . . . . . . . . . . . . . 161Špecificita stresovej reakcie . . . . . . . . . . . . . . . 161Neuronálne okruhy stresovej reakcie . . . . . . . . . . . 162Centrálna zložka stresovej reakcie . . . . . . . . . . . . 163Nucleus paraventricularis hypotalamu ako integračnécentrum stresovej reakcie . . . . . . . . . . . . . . . 163
Úloha centrálnych katecholamínergických oblastí v stre-sovej reakcii . . . . . . . . . . . . . . . . . . . . . . 166
Interakcie medzi CRH a katecholamínergickými neurónmi 167Úloha ďalších neurotransmiterov v stresovej reakcii . . . 168Periférna zložka stresovej reakcie . . . . . . . . . . . . 168Sympatikoadrenálny systém . . . . . . . . . . . . . . . 169Hypotalamo-hypofýzo-adrenokortikálny systém . . . . . 170Interakcie medzi sympatikoadrenálnym a hypotalamo-hypofýzo-adrenokortikálnym systémom . . . . . . . . 171
Nepriaznivé následky pôsobenia stresu . . . . . . . . . 171Bolesť . . . . . . . . . . . . . . . . . . . . . . . . . . . . 172Prenos nociceptívnych informácií do CNS . . . . . . . . 172Centrálna modulácia prenosu nociceptívnych informácií 173Neuropatická bolesť . . . . . . . . . . . . . . . . . . . 175
Vývin nervového systému . . . . . . . . . . . . . . . . . . 175Oxygenácia organizmu . . . . . . . . . . . . . . . . . . . 177Regulácia telesnej teploty . . . . . . . . . . . . . . . . . . 177Starnutie a nervový systém . . . . . . . . . . . . . . . . . 178

Obsah 9
6 Úloha neurotransmiterov pri ochoreniach nervového sys-tému . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180Neurodegeneratívne ochorenia . . . . . . . . . . . . . . . 181Parkinsonova choroba . . . . . . . . . . . . . . . . . . 182
Schizofrénia . . . . . . . . . . . . . . . . . . . . . . . . . 185Afektívne poruchy . . . . . . . . . . . . . . . . . . . . . . 190Depresia . . . . . . . . . . . . . . . . . . . . . . . . . 190
Psychiatrické ochorenia a neurogenéza . . . . . . . . . . . 196Látková závislosť . . . . . . . . . . . . . . . . . . . . . . 201Závislosť od alkoholu . . . . . . . . . . . . . . . . . . . 202
Epilepsia . . . . . . . . . . . . . . . . . . . . . . . . . . . 203Ischemické poškodenie CNS . . . . . . . . . . . . . . . . . 204
7 Úloha neurotransmiterov pri iných ochoreniach . . . . . . 207Hypertenzia . . . . . . . . . . . . . . . . . . . . . . . . . 207Zápalové procesy . . . . . . . . . . . . . . . . . . . . . . 209
Záver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
Literatúra . . . . . . . . . . . . . . . . . . . . . . . . . . . 212
Odkazy na webové stránky . . . . . . . . . . . . . . . . . . 268
Použité skratky . . . . . . . . . . . . . . . . . . . . . . . . 270
Zoznam tabuliek . . . . . . . . . . . . . . . . . . . . . . . 274
Zoznam obrázkov . . . . . . . . . . . . . . . . . . . . . . . 276
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 279

Štruktúra a činnosť neurónov
B. Mravec
Nervový systém obsahuje dva základné typy buniek – neurónya bunky glie (astrocyty, oligodendrocyty, mikrogliu, Schwannove bunkya iné). V prenose informácií majú kľúčové postavenie neuróny, aj keďglia nepredstavuje iba štruktúrnu, podpornú, či pasívnu zložku nervo-vého systému (Kandel a spol., 2000).
Štruktúra neurónov
Neurón obsahuje podobné intracelulárne zložky ako iné bunky. Typickýneurón sa skladá z tela bunky a výbežkov. Dendrity, axón a presynap-tické zakončenia sú štruktúry špecifické pre neurón (obr. 1.1). Aj keďmajú neuróny základné štruktúry podobné, na základe morfologickýchcharakteristík sa rozlišuje viac ako 50 typov neurónov (Kandel a spol.,2000).Bunkové telo (perikaryon) je metabolickým centrom neurónu. Obsa-
huje bunkové jadro, endoplazmatické retikulum, mitochondrie, Golgihoaparát, lyzozómy a iné bunkové organely. Z bunkového tela väčšinou vy-biehajú dva typy výbežkov, niekoľko krátkych dendritov a jeden dlhýaxón.Bunková membrána je miestom, v ktorom prebieha veľa dejov pod-
mieňujúcich činnosť neurónu (presun iónov iónovými kanálmi, pre-
11

12 NEUROTRANSMITERY
Obr. 1.1 Štruktúra neurónu. Intracelulárne štruktúry sú, okrem bunkového
jadra, pre prehľadnosť vynechané. Detail v hornej časti zobrazuje mechanizmusprenosu signálu chemickou synapsou (modifikované podľa Kandel a spol., 2000;
Petrovický, 2002).

1 Štruktúra a činnosť neurónov 13
nos informácií z okolia neurónu prostredníctvom aktivácie receptorovumiestnených v bunkovej membráne atď.).Bunkové jadro je veľké a zvyčajne má sférický tvar. Vonkajší obal
bunkového jadra niekedy kontinuálne prechádza do endoplazmatickéhoretikula.Nisslova substancia sa skladá z intracytoplazmatických bazofilných
štruktúr, ktoré sa rozvetvujú voľne v cytoplazme. Je typickou zložkouväčšiny neurónov. Nisslovu substanciu tvorí drsné endoplazmatické re-tikulum a voľné polyribozómy. Prebieha tu tvorba väčšiny proteínov,ktoré sú syntetizované v neuróne.Cytoskelet zabezpečuje v bunke špecifické funkcie a je hlavným vnú-
torným determinantom tvaru neurónov. Zúčastňuje sa aj na asymetric-kej distribúcii organel v cytoplazme. Cytoskelet tvoria tri hlavné vlák-nité štruktúry: mikrotubuly, neurofilamenty a aktínové mikrofilamenty.Mikrotubuly vytvárajú dlhé vláknité konštrukcie, ktoré vypĺňajú celýneurón. Majú kľúčovú úlohu pri vývoji a udržiavaní výbežkov neuró-nov. Neurofilamenty sú základnou štruktúrnou zložkou cytoskeletu ne-urónov. Vo veľkom množstve sa vyskytujú v axónoch. Neurofilamentysú v porovnaní s mikrotubulmi veľmi stabilnými štruktúrami. Mikrofi-lamenty sú najtenšou zložkou cytoskeletu. Vytvárajú relatívne krátkevlákna, ktoré sú však navzájom poprepájané, čím tvoria pomerne hustúsieť. Táto sieť má dôležitú úlohu v dynamických procesoch, akými súnapríklad pohyblivosť rastového kónusu počas rastu výbežkov neuró-nov a tvorba špecifickej konformácie presynaptickej a postsynaptickejoblasti neurónov. Mikrotubuly a aktínové mikrofilamenty umožňujúorganizovaný transport organel a proteínov v neuróne (zúčastňujú sanapríklad na axonálnom transporte neuropeptidov alebo enzýmov prebiosyntézu malomolekulových neurotransmiterov z tela bunky do ner-vových zakončení) (Maršala, 1985; Kandel a spol., 2000; Siegel a spol.,2006).Rozvetvujúce sa dendrity sú hlavnou oblasťou neurónu, ktorá pri-
jíma signály od iných nervových buniek (Jan a Jan, 2001).Axón, ktorý je väčšinou niekoľkonásobne dlhší ako dendrit, je hlav-
nou štruktúrou, ktorá vedie signál k ďalším neurónom. V oblasti od-stupu axónu z tela neurónu sa na ňom nachádza špecializovaná ob-lasť (axónový hrbolček nasledovaný iniciálnym segmentom), ktorá jemiestom vzniku akčného potenciálu. V blízkosti zakončenia sa axónrozdeľuje na niekoľko tenkých vetvičiek, ktoré vytvárajú presynaptické

14 NEUROTRANSMITERY
nervové zakončenia zodpovedné za komunikáciu s inými neurónmi. Pre-synaptické zakončenie axónu je väčšinou v blízkom kontakte s postsy-naptickou membránou neurónu, ktorá prijíma nervový signál. Presy-naptické nervové zakončenie a postsynaptická membrána spolu s pries-torom, ktorý ich oddeľuje, vytvárajú synapsu, v ktorej prebiehajú zlo-žité procesy prenosu signálov. Priestor medzi presynaptickým nervovýmzakončením a postsynaptickou membránou sa označuje ako synaptickáštrbina (Maršala, 1985) (obr. 1.1).
Prenos signálov v nervovom systéme
Nervový systém využíva na prenos signálov elektrochemické procesy,ktoré podmieňujú vznik akčného potenciálu. Akčný potenciál sa šíripo axóne ako vlna postupnej depolarizácie. Nervový vzruch podmie-nený akčným potenciálom, ktorý vzniká v iniciálnom segmente neurónu,predstavuje základnú informačnú jednotku v nervovom systéme. Šíri sabez výraznejšej zmeny amplitúdy akčného potenciálu. Preto sa v ner-vovom systéme nevyužíva amplitúdová, ale frekvenčná modulácia, keďje informácia zakódovaná do zmien frekvencie šíriacich sa akčných po-tenciálov.Prenos signálov medzi neurónmi sprostredkúvajú synapsy. Elek-
trické synapsy, ktorých je relatívne menej, umožňujú priamy prechodvlny depolarizácie (akčného potenciálu) z jedného neurónu na druhý.Naproti tomu prenos signálov chemickými synapsami, ktoré v nervo-vom systéme dominujú, vyžaduje zmenu elektrochemického signálu (vy-tvárajúceho akčný potenciál v presynaptickom neuróne) v synapse nachemický signál reprezentovaný uvoľneným neurotransmiterom. Neuro-transmiter následne indukuje na postsynaptickom neuróne vznik elek-trochemického signálu, ktorý ak je dostatočnej intenzity, umožní vznikakčného potenciálu.Pri prenose signálov medzi neurónmi platí princíp dynamickej pola-
rizácie, ktorý určuje to, že elektrický signál sa v nervovej bunke šíri ibajedným smerom. Signál sa šíri z prijímajúcich oblastí (väčšinou dendritya bunkové telo) k spúšťacej oblasti axónu (axónový hrbolček a iniciálnysegment). Odtiaľ sa akčný potenciál vedie jednosmerne po axóne až dooblastí presynaptických zakončení. Z presynaptického zakončenia sa ná-sledne uvoľňuje neurotransmiter, ktorý po naviazaní sa na receptory na

1 Štruktúra a činnosť neurónov 15
postsynaptickej membráne vyvoláva zmenu v činnosti postsynaptickéhoneurónu (Kandel a spol., 2000).Novšie poznatky však naznačujú, že v určitých prípadoch sa môže
signál prenášať aj dendritmi, ktoré môžu uvoľňovať neurotransmitery(väčšinou zo skupiny neuropeptidov) do svojho okolia (Ludwig a Pitt-man, 2003).Prenos a spracovanie signálov v nervovom systéme možno prirovnať
k hybridnému počítaču, ktorý využíva analógové aj digitálne signály,a preto ťaží z výhod oboch spôsobov prenosu a spracovania signálov.Kým intenzita analógových signálov varíruje v určitom spojitom roz-sahu, digitálne signály majú iba dva stavy „všetkoÿ (1) alebo „ničÿ (0).Dendrity integrujú informácie z tisícov synaptických vstupov.
Každý z týchto vstupov má malý vplyv na napätie v dendrite, ktorésa preto môže meniť kontinuálne v určitom rozmedzí. Dendrity pretospracúvajú informácie ako analógový počítač. Pretože analógové spra-covanie varíruje v určitom kontínuu hodnôt, je tento proces veľmi citlivýna informačný šum a náchylný na zmeny v amplitúde. Pri prenose ta-kéhoto typu signálov na väčšie vzdialenosti môže ľahko dôjsť k zánikusignálu. Preto nie je analógový systém vhodný na prenos informácií naväčšie vzdialenosti, ani na využitie vo veľkých systémoch.Výsledkom spracovania informácií v dendritoch je premena vstup-
ných analógových signálov na digitálne signály, ktoré reprezentujeakčný potenciál. V protiklade s analógovým systémom je digitálny sig-nál („všetkoÿ alebo „ničÿ) reprezentovaný akčným potenciálom, veľmivhodný na prenos informácií na veľké vzdialenosti, pretože nedochádzak jeho výraznejšiemu ovplyvneniu šumom ani k zmenám v jeho ampli-túde (Allman, 2000).

Neurotransmiterové systémy
B. Mravec
Zoskupenia neurónov v centrálnom nervovom systéme (CNS) možnoklasifikovať podľa rôznych kritérií. Klasický anatomický opis charak-terizuje mozgovú oblasť podľa dráh, ktoré z tejto oblasti vychádzajúa ktoré do nej prichádzajú. Toto hľadisko je základom deskriptívnejneuroanatómie. Ako príklad možno uviesť neuróny nucleus arcuatus,ktoré vysielajú niektoré axóny do oblasti eminentia mediana, čím tvo-ria tractus tuberoinfundibularis. Iným príkladom sú dlhé axóny vy-chádzajúce z motorických oblastí mozgovej kôry (z gyrus precentralis)k α-motoneurónom predných rohov miechy tvoriace tractus corticospi-nalis. Existujú aj iné kritériá na opis oblastí mozgu a ich spojov (napr.delenie na systém motorický a senzorický, limbický a ďalšie).Iná klasifikácia charakterizuje oblasti nervového systému podľa che-
mickej látky, ktorú určité zoskupenie neurónov syntetizuje a uvoľňujeako neurotransmiter (Kandel a spol., 2000). Neuróny syntetizujúce rov-naký neurotransmiter vytvárajú neurotransmiterové systémy. Naprí-klad zoskupenie neurónov, ktoré syntetizujú noradrenalín, tvorí norad-renalínergický systém neurónov. Noradrenalínergické neuróny spoločnes axónmi, ktoré vysielajú do jednotlivých oblastí mozgu, vytvárajú tzv.noradrenalínergický systém mozgu a miechy.V neuroanatómii sa neurotransmiterové systémy začali intenzívne
študovať od druhej polovice 20. storočia, keď Falck a Hillarp vypraco-
16

2 Neurotransmiterové systémy 17
vali fluorescenčnú histochemickú metódu (Falck a spol., 1962). Tátometóda umožnila detekciu monoamínov v mozgu. Následne došlo k in-tenzívnemu výskumu, ktorého výsledkom je podrobná charakterizáciadistribúcie neurónov na základe syntetizovaného neurotransmitera (pre-hľad používaných metodík pozri Windhorst a Johansson, 1999). Ďal-šou významnou skutočnosťou na poli výskumu neurotransmiterov bolozistenie, že mnohé biologicky aktívne peptidy, predtým známe ako hor-móny syntetizované periférnymi tkanivami (napr. v gastrointestinálnomtrakte), sú syntetizované aj neurónmi a slúžia ako neurotransmitery,resp. neuromodulátory (Ballesta a spol., 1985).Štruktúry nervového systému môžeme klasifikovať na základe ana-
tomických alebo funkčných vlastností. Takto klasifikované systémy CNSsú navzájom komplexne prepojené. Treba na to prihliadať aj pri členeníštruktúr CNS do rôznych neurotransmiterových systémov. Je dôležitéprihliadať aj na to, že na jeden neurón pôsobí väčšinou viacero rôznychneurotransmiterov. To vytvára podklad pre komplexnú sieť interakciív rámci neurotransmiterových systémov v nervovom systéme.Väčšina poznatkov o distribúcii neurónov tvoriacich neurotransmi-
terové systémy mozgu, ako aj o ich úlohe, sa získala z experimentov nalaboratórnych zvieratách (prevažne na hlodavcoch, potkanoch a my-šiach). Rovnaké, alebo podobné zoskupenia sa preukázali aj v ľudskommozgu (Maršala, 1985). Je však potrebné vyhnúť sa nekritickému zovše-obecňovaniu poznatkov o funkciách jednotlivých neurotransmiterovýchsystémov získaných z experimentov na laboratórnych zvieratách.Výskum zaoberajúci sa identifikáciou neurotransmiterov a ich fun-
kciami prebieha niekoľko desaťročí a v súčasnosti pribúda veľkou rých-losťou množstvo nových poznatkov. Sústavne sa identifikujú nové ne-uropeptidy, ktoré majú funkcie neurotransmiterov, resp. neuromodu-látorov. Štúdium neurotransmiterových systémov umožňuje pochopiťviaceré funkcie nervového systému a aspoň čiastočne porozumieť eti-opatogenéze tých ochorení nervového systému, pri ktorých dochádzak narušeniu činnosti neurotransmiterových systémov. Veľmi intenzívnesa skúma úloha jednotlivých neurotransmiterových systémov a ich vzá-jomných interakcií v etiopatogenéze ochorení CNS, ako sú napríkladafektívne poruchy, schizofrénia, Parkinsonova choroba a Alzheimerovachoroba. Bez poznania funkcie neurotransmiterov ťažko pochopiť prin-cípy farmakoterapie týchto ochorení.

18 NEUROTRANSMITERY
Komunikácia neurónov
V minulosti sa predpokladalo, že mozog je tvorený syncýtiom buniek,ktoré nie sú navzájom anatomicky ohraničené. Až neurónová teória(doktrína), ktorú vypracoval Ramón y Cajal na prelome 19. a 20. storo-čia, poukázala na to, že mozog sa skladá z veľkého počtu nervových bu-niek, ktoré sú navzájom oddelené (Sotelo, 2003; Bullock a spol., 2005).Preto sa do popredia dostala otázka, ako jednotlivé neuróny navzájomkomunikujú (Krnjevic, 2005).Ľudský mozog obsahuje podľa rôznych odhadov 10–100 miliárd
neurónov, pričom každý neurón môže priamo komunikovať priemernes 1–10 tisíc ďalšími neurónmi (Squire a spol., 2003). Pri prenose in-formácií v nervovom systéme majú dôležitú úlohu okrem neurónovaj bunky glie (prevažne astrocyty). Astrocyty plnia mnoho dôležitýchfunkcií, ktoré sú nevyhnutné pre normálnu komunikáciu medzi neurón-mi. Medzi tieto funkcie patrí podiel na odstraňovaní, resp. vychytávaníneurotransmiterov, modulácia synaptického prenosu, ako aj ohraničo-vanie synaptických formácií, čím zabraňujú nekontrolovanej difúzii neu-rotransmiterov (Araque a spol., 1999; Fields a Stevens-Graham, 2002;Essen, 2004).V súčasnosti je známych niekoľko odlišných typov komunikácie me-
dzi neurónmi (obr. 2.1):
• klasické chemické synapsy – nervový vzruch sa prenáša chemic-kými látkami (neurotransmitermi); difúzia neurotransmitera sy-naptickou štrbinou (širokou ∼ 20–40 nm) zapríčiňuje synaptickézdržanie v rozmedzí jednej až niekoľkých milisekúnd, prípadnesekúnd až minút,
• difúzia neurotransmiterov z varikozít (typu en passant) neuro-transmitery ovplyvňujú väčšie množstvo neurónov vo vzdialenosti∼ 2–100µm, synaptické zdržanie je niekoľko milisekúnd až minút(Vizi, 2000; Zoli a Agnati, 1996; Sykova, 2004, 2005),
• elektrická synapsa – priamy prechod elektrolytov z jedného neu-rónu do druhého charakterizovaný veľmi krátkym synaptickýmzdržaním (∼ 0,1ms), malou vzdialenosťou membrán neurónov(∼ 2–4 nm) a možnosťou obojsmerného prenosu vzruchov (Hor-muzdi a spol., 2004).

2 Neurotransmiterové systémy 19
Obr. 2.1 Schematické znázornenie jednotlivých typov komunikácie v nervovomsystéme. A – chemická synapsa; B – elektrická synapsa; C – juxtapozícia mem-
brán (elektrické potenciály umožňujú vzájomnú moduláciu excitability mem-brán neurónov, ktoré sa nachádzajú v tesnej blízkosti); D – extrasynaptické
uvoľňovanie (uvoľnený neurotransmiter difunduje v okolí synapsy a pôsobí na
receptory v jej blízkosti); E – uvoľnený neurotransmiter prechádza do cirkulácie(napr. hypotalamické liberíny a statíny); F – cirkulujúce látky môžu modifikovať
činnosť neurónov (E a F – prevažne v cirkumventrikulárnych orgánoch mozgu);G – uvoľnené neurotransmitery prechádzajú do mozgových komôr (V), v ktorých
cirkuluje cerebrospinálna tekutina, ktorá umožňuje pôsobenie neurotransmitera
vo vzdialenej oblasti nervového systému; H – synapsa typu en passant (nie jeutvorená klasická synaptická konformácia, uvoľnený neurotransmiter difunduje
do širokého okolia, a tak ovplyvňuje činnosť väčšieho počtu neurónov; modifi-
kované podľa Agnati a spol., 1995; Sykova, 2004).

20 NEUROTRANSMITERY
Významnou vlastnosťou elektrických synáps je to, že na základeveľmi rýchleho prenosu vzruchov sa zúčastňujú na procesochsynchronizácie a oscilácie v CNS (Zoidl a Dermietzel, 2002; pre-hľad pozri Connors a Long, 2004). Neuróny, ktoré vytvárajú elek-trické synapsy, môžu zároveň vytvárať aj klasické chemické sy-napsy. Takýto kombinovaný prenos vzruchov, na ktorom sa zú-častňujú chemické aj elektrické synapsy, je prítomný napríkladv mozgovej kôre (Hestrin a Galarreta, 2005),
• morfologické nálezy poukazujú na to, že veľké množstvo neurónov(aj buniek glie) vytvára tzv. primárne cilium; predpokladá sa, žeprimárne cilium môže slúžiť na nesynaptickú komunikáciu medzineurónmi, ktorá môže prispievať ku komplexnému spracovaniusignálov v nervovom systéme (Whitfield, 2004),
• hypotézy pokúšajúce sa vysvetliť procesy spojené s vedomímpredpokladajú aj prítomnosť extrémne rýchleho prenosu infor-mácií na väčšie vzdialenosti v CNS prostredníctvom kvantovýchdejov (bližšie pozri Reinis a spol., 2005).
Charakteristika neurotransmiterov
Jeden z prvých dôkazov chemického prenosu poskytol Otto Loewi vďakasvojmu dômyselnému experimentu s dvoma srdcami. Loewi dokázal po-mocou experimentu s inervovaným a denervovaným srdcom, že na regu-lácii činnosti srdca sa zúčastňuje chemická látka uvoľňovaná zo zakon-čení nervus vagus (acetylcholín). Za tento objav získal roku 1936 Nobe-lovu cenu. Bolo však potrebných ešte niekoľko desaťročí experimentálnejvedeckej práce, kým bola teória prenosu informácií v centrálnom ner-vovom systéme prostredníctvom chemických látok, neurotransmiterov,všeobecne prijatá (Bennett, 2000; Valenstein, 2002).Aby mohla byť chemická látka zaradená medzi neurotransmitery,
musí spĺňať niekoľko kritérií (Kandel a spol., 2000; Purves a spol., 2001;Squire a spol., 2003).Neurotransmiter musí byť syntetizovaný v neuróne. Neurón musí
disponovať potrebným syntetickým aparátom, ktorý zahŕňa enzýmy po-trebné pre biosyntézu neurotransmitera:
• enzýmy potrebné pre biosyntézu malomolekulových neurotrans-miterov (napr. katecholamínov, acetylcholínu, glutamátu) sú

2 Neurotransmiterové systémy 21
transportované do nervových zakončení axónovým transportom,čo umožňuje syntézu týchto neurotransmiterov priamo v nervo-vých zakončeniach,
• veľkomolekulové neurotransmitery, neuropeptidy, sú syntetizo-vané v oblasti bunkového tela vo forme inaktívnych prekurzorov,ktoré sú umiestnené do vezikúl a prenášané axonálnym transpor-tom do presynaptického zakončenia neurónu; počas axonálnehotransportu dochádza k odštiepenie častí reťazca a ďalším mo-difikáciám molekúl neuropeptidov, čím vznikajú aktívne formypeptidov.
Syntetizovaný neurotransmiter je uskladnený vo vezikulách v sy-naptickom zakončení neurónu. Na základe funkčných vlastností sa roz-lišujú tri základné zoskupenia vezikúl, v ktorých je neurotransmiteruskladnený:
• rýchlo uvoľniteľné vezikuly tvoria približne 1–2% všetkých vezi-kúl; umožňujú uvoľniť neurotransmiter ihneď po stimulácii pre-synaptického nervového zakončenia,
• recyklujúce vezikuly tvoria približne 5–20% vezikúl; uvoľňujú ne-urotransmiter po miernej fyziologickej stimulácii,
• rezervné vezikuly tvoria priemerne 80–90% vezikúl, slúžia akozásoba neurotransmitera, ktorý uvoľňujú iba pri intenzívnej sti-mulácii (Rizzoli a Betz, 2005).
Neurotransmiter sa uvoľňuje zo sekrečných vezikúl do synaptickejštrbiny po príchode akčného potenciálu do oblasti nervového zakon-čenia.Membrána postsynaptického neurónu musí obsahovať receptory,
na ktoré sa neurotransmiter uvoľnený z presynaptického nervového za-končenia viaže, a tak vyvoláva zmeny v činnosti neurónu. Väzbou napostsynapticky lokalizované receptory vyvoláva neurotransmiter odpo-veď postsynapticky lokalizovaného neurónu vo forme zvýšenia, resp.zníženia priepustnosti postsynaptickej membrány pre jednotlivé ióny.Receptory pre neurotransmiter sa môžu nachádzať aj na presynap-tickom nervovom zakončení.Označujú sa ako autoreceptory. Obsadenieautoreceptorov väčšinou zníži ďalšie uvoľňovanie neurotransmitera. Au-toreceptory preto zodpovedajú za negatívnu spätnoväzbovú reguláciu

22 NEUROTRANSMITERY
uvoľňovania neurotransmitera, čím zabraňujú jeho nadmernému uvoľne-niu do synaptickej štrbiny. Okrem toho sa na presynaptickej membránemôžu nachádzať aj receptory pre ďalšie neurotransmitery, čo vytvárapodklad pre komplexnosť interakcií medzi jednotlivými systémami ne-urotransmiterov.Musí byť prítomný mechanizmus, ktorý zabezpečuje inaktiváciu
uvoľneného neurotransmitera. Neurotransmiter môže byť v synaptic-kej štrbine degradovaný, napr. acetylcholín je biodegradovaný enzýmomacetylcholínesterázou na cholín a acetát. Iný mechanizmus predstavujevychytávanie neurotransmitera spätne do nervového zakončenia, z kto-rého bol uvoľnený (tzv. reuptake) alebo bunkami glie. Spätné vychy-távanie neurotransmitera predstavuje efektívny proces odstraňovanianeurotransmitera zo synaptického zakončenia, ktorý je energeticky vý-hodný, pretože spätne vychytaný neurotransmiter sa môže znova použiť(obr. 2.2).Ďalšou podmienkou je, že exogénne podaný neurotransmiter musí
vyvolať rovnakú odpoveď postsynaptického neurónu ako jeho endo-génne uvoľnenie.V súčasnosti sú však známe aj látky, ktoré sa aj napriek tomu, že ne-
spĺňajú všetky uvedené charakteristiky, zaraďujú medzi neurotransmi-tery, resp. neuromodulátory (napríklad plynné neurotransmitery, medziktoré sú zaradené NO, CO a H2S).
Základné skupiny neurotransmiterov
V súčasnosti je známych 50–100 látok z rôznych chemických skupín,ktoré spĺňajú niektoré alebo všetky kritériá potrebné pre zaradenie me-dzi neurotransmitery (Snyder a Ferris, 2000; Ramachandran, 2002).Jedna z klasifikácií rozdeľuje neurotransmitery podľa príslušnosti
k chemickým skupinám (Webster, 2001) (tab. 2.1).Zaradenie látok zo skupín steroidov, plynných látok a eikosanoidov
medzi neurotransmitery je podložené experimentálnymi poznatkami.Pre ich definitívne zaradenie medzi neurotransmitery bude však po-trebné ďalšie štúdium ich vlastností (Webster, 2001).Klasifikácia neurotransmiterov do rôznych skupín na základe vy-
braných charakteristík sa síce používa často, ale má skôr didaktickývýznam.

2 Neurotransmiterové systémy 23
Obr. 2.2 Schematické znázornenie základných charakteristík neurotransmi-terov. Enzýmy v nervovom zakončení syntetizujú neurotransmiter, ktorý je
uskladnený v sekrečných vezikulách. Po depolarizácii presynaptickej oblasti
vplyvom akčného potenciálu dochádza k vzostupu intracelulárnej koncentrácieCa2+. Zvýšená koncentrácia vápnika v presynaptickej oblasti vyvolá kaskádu
dejov, ktorých výsledkom je fúzia vezikúl obsahujúcich neurotransmiter s pre-synaptickou membránou neurónu, čím sa neurotransmiter uvoľní do synaptic-
kej štrbiny. Neurotransmiter pôsobí na postsynapticky lokalizované ionotropné
(i) alebo metabotropné (m) receptory, čím vyvoláva zmeny na postsynaptickejmembráne (depolarizáciu alebo hyperpolarizáciu), ale môže ovplyvniť aj intra-
celulárne deje, výsledkom ktorých môže byť napríklad zmena v génovej expresii
neurónu. Zároveň môže neurotransmiter pôsobiť aj na presynapticky lokalizo-vané autoreceptory. Pôsobenie neurotransmitera je ukončené jeho degradáciou
v synaptickej štrbine (napr. acetylcholín), alebo jeho spätným vychytaním dopresynaptického nervového zakončenia (napr. monoamíny) alebo bunkami glie
(napr. glutamát). Neurotransmiter spätne vychytaný do nervového zakončenia
je buď metabolizovaný, alebo môže byť po uskladnení v sekrečných vezikuláchznovu uvoľnený do synaptickej štrbiny.

24 NEUROTRANSMITERY
Chemická látka Príklad
Malomolekulové neurotransmitery
Ester cholínu acetylcholín (ACh)
Monoamíny
katecholamíny dopamín (DA)
noradrenalín (NA)
adrenalín (ADR)indoly sérotonín (5-hydroxytryptamín, 5-HT)
imidazoly histamín (Hist)
Aminokyseliny
acidické glutamát (Glu)
bázické γ-aminomaslová kyselina (GABA)
glycín (Gly)
Puríny adenozíntrifosfát (ATP)
adenozín
Steroidy pregnenalón
dehydroepiandrosterón
Plynné látky oxid dusnatý (NO)
oxid uhoľnatý (CO)
hydrogénsulfát (H2S)
Eikosanoidy prostaglandíny
Veľkomolekulové neurotransmitery
Peptidy napr. enkefalíny, substancia P, cholecystokinín
Tabuľka 2.1 Rozdelenie neurotransmiterov podľa príslušnosti k chemickýmskupinám.

2 Neurotransmiterové systémy 25
Podľa charakteru odpovede vyvolanej na postsynaptickom neurónemožno neurotransmitery rozdeliť na tri skupiny:
• excitačné aminokyseliny (napr. glutamát),
• inhibičné aminokyseliny (napr. GABA alebo glycín),
• modulačné (acetylcholín, noradrenalín, dopamín, adrenalín, séro-tonín a histamín).
Podľa veľkosti molekuly možno neurotransmitery rozdeliť na dveveľké skupiny:
• neurotransmitery s „malouÿ molekulovou hmotnosťou (napr. ami-nokyseliny, monoamíny) (tab. 2.2),
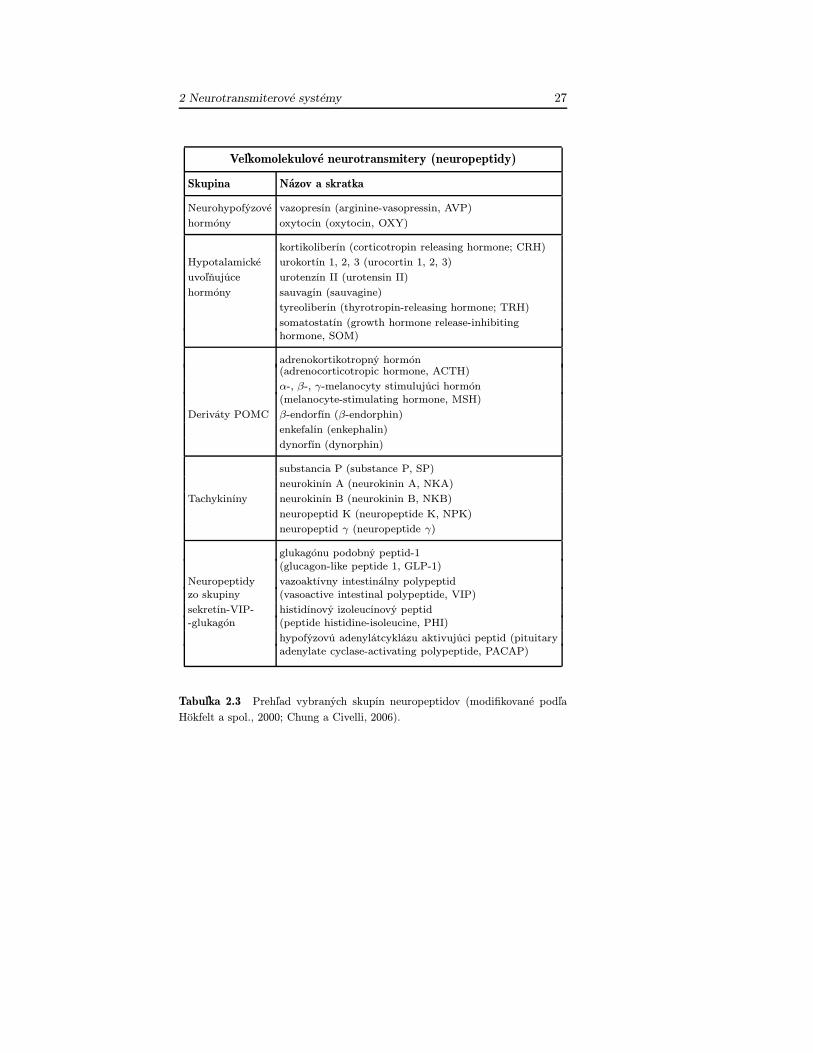
• neurotransmitery s „veľkouÿ molekulovou hmotnosťou (neuropep-tidy) (tab. 2.3).
Predpokladá sa, že ďalšiu skupinu látok, ktoré sa môžu zúčastňo-vať na procesoch zabezpečujúcich komunikáciu medzi neurónmi, tvoriachemokíny. Pozorovalo sa, že chemokíny okrem modulácie vývoja nervo-vého systému a imunitných funkcií ovplyvňujú aj excitabilitu neurónova synaptický prenos, čím by sa mohli zaradiť medzi neuromodulátory.Experimentálne údaje napríklad naznačujú, že môže dochádzať k hete-rológnej desenzitizácii medzi opioidmi a chemokínmi. Uvažuje sa o tom,že chemokíny môžu sprostredkúvať prenos signálov medzi gliovými bun-kami, medzi neurónmi, ako aj medzi neurónmi a gliovými bunkami (Ad-ler a Rogers, 2005; Adler a spol., 2006).Na prenos informácií využíva nervový systém široké spektrum neu-
rotransmiterov. Táto rôznorodosť neurotransmiterov umožňuje kom-plexnosť spracovania informácií v nervovom systéme. Pri prenose a spra-covaní informácií v nervovom systéme preto nedochádza k jednodu-chým, monotónnym a ľahko detegovateľným zmenám. Neurotransmi-tery vyvolávajú skôr zmeny v činnosti nervového systému, ktoré možnooznačiť ako vysokokomplexné.
Receptory pre neurotransmitery
Neurotransmitery uvoľnené z presynaptického nervového zakončenia di-fundujú do synaptickej štrbiny a viažu sa na receptory umiestnené na

26 NEUROTRANSMITERY
Malomolekulové neurotransmitery
Skupina Transmiter Molekulový vzorec
Ester cholínu acetylcholín
dopamín
noradrenalín
Biogénne amíny adrenalín
sérotonín
histamín
glutamát
Aminokyseliny GABA
glycín
Puríny adenozín
ATP
Tabuľka 2.2 Prehľad vybraných malomolekulových neurotransmiterov a ich
štruktúrnych vzorcov.
2 Neurotransmiterové systémy 27
Veľkomolekulové neurotransmitery (neuropeptidy)
Skupina Názov a skratka
Neurohypofýzové vazopresín (arginine-vasopressin, AVP)
hormóny oxytocín (oxytocin, OXY)
kortikoliberín (corticotropin releasing hormone; CRH)
Hypotalamické urokortín 1, 2, 3 (urocortin 1, 2, 3)
uvoľňujúce urotenzín II (urotensin II)
hormóny sauvagín (sauvagine)
tyreoliberín (thyrotropin-releasing hormone; TRH)
somatostatín (growth hormone release-inhibitinghormone, SOM)
adrenokortikotropný hormón(adrenocorticotropic hormone, ACTH)
α-, β-, γ-melanocyty stimulujúci hormón(melanocyte-stimulating hormone, MSH)
Deriváty POMC β-endorfín (β-endorphin)
enkefalín (enkephalin)
dynorfín (dynorphin)
substancia P (substance P, SP)
neurokinín A (neurokinin A, NKA)
Tachykiníny neurokinín B (neurokinin B, NKB)
neuropeptid K (neuropeptide K, NPK)
neuropeptid γ (neuropeptide γ)
glukagónu podobný peptid-1(glucagon-like peptide 1, GLP-1)
Neuropeptidy vazoaktívny intestinálny polypeptidzo skupiny (vasoactive intestinal polypeptide, VIP)
sekretín-VIP- histidínový izoleucínový peptid-glukagón (peptide histidine-isoleucine, PHI)
hypofýzovú adenylátcyklázu aktivujúci peptid (pituitaryadenylate cyclase-activating polypeptide, PACAP)
Tabuľka 2.3 Prehľad vybraných skupín neuropeptidov (modifikované podľaHökfelt a spol., 2000; Chung a Civelli, 2006).

28 NEUROTRANSMITERY
pokračovanie tabuľky 2.3
Veľkomolekulové neurotransmitery (neuropeptidy)
Skupina Názov a skratka
adrenomedulín (adrenomedullin, ADM)agouti príbuzný proteín (agouti-related protein, AGRP)amylín (amylin, islet amyloid polypeptide, AMY, IAPP)angiotenzín II (angiotensine II)apelín (apelin)endomorfín 1, 2 (endomorphin 1, 2)galanín (galanin, GAL)galanínu podobný peptid (galanin-like peptid, GALP)ghrelín (ghrelin)cholecystokinín (cholecystokinin, CCK)chromogranín A, B (chromogranin A, B),sekretoneurín (secretoneurin)kalcitonínu génovo príbuzný peptid(calcitonin gene-related peptide, CGRP)kokaínom a amfetamínom regulovaný transkript(cocaine- and amphetamine-regulated transcript, CART)melanín koncentrujúci hormón(melanin-concentrating hormone, MCH)
Iné metastín (metastin)neuropeptidy mozgový nátriuretický peptid (brain natriuretic peptide,
BNP)neuropeptid AF (neuropeptide AF, NPAF)neuropeptid B (neuropeptide B, NPB)neuropeptid EI (neuropeptide EI, NPEI)neuropeptid FF (neuropeptide FF, NPFF)neuropeptid GE (neuropeptide GE, NPGE)neuropeptid SF (neuropeptide SF, NPSF)neuropeptid S (neuropeptide S, NPS)neuropeptid tyrozín (neuropeptide tyrosine, NPY)neuropeptid W (neuropeptide W, NPW)neurotenzín (neurotensin, NT)nociceptín/ofranín FQ (nociceptin/orphanin FQ, N/OFQ)nocistatín (nocistatin)orexín/hypokretín (orexin/hypocretin, ORX/HCRT)prolaktín uvoľňujúci peptid (prolactin-releasing peptide,PrRP)prokineticín 2 (prokineticin 2, PK2)relaxín 3 (relaxin 3, RLX3)5-HT modulín (5-HT-moduline)

2 Neurotransmiterové systémy 29
postsynaptickej membráne neurónov (v niektorých prípadoch sa uvoľ-nené neurotransmitery môžu viazať aj na receptory umiestnené na pre-synaptickej membráne, tzv. autoreceptory).Receptory pre neurotransmitery možno rozdeliť do dvoch základ-
ných skupín, ktoré sa líšia hlavne rýchlosťou zmien iónovej priepustnostimembrány neurónov po ich obsadení.
• Ionotropné receptory – väzba neurotransmitera na ionotropnýreceptor vyvolá v prvom rade zmenu priepustnosti iónového ka-nála, ktorý je súčasťou receptora. Výsledkom je relatívne rýchlazmena polarity postsynaptickej membrány (obr. 2.3). Medzi neu-rotransmitery, ktoré sa viažu na ionotropné receptory, patria na-príklad excitačné a inhibičné aminokyseliny, ale aj acetylcholín.
• Metabotropné receptory – väzba neurotransmitera na meta-botropný receptor vyvoláva prostredníctvom G-proteínov a ná-sledne druhých poslov (tab. 2.4) kaskádu zmien, ktorých výsled-kom je zmena priepustnosti iónových kanálov s určitou latenciou(obr. 2.4). Neurotransmitery, ktoré pôsobia prostredníctvom me-tabotropných receptorov, sú napríklad biogénne amíny, acetyl-cholín, neuropeptidy, ale aj glutamát (Mysliveček a Trojan, 2002,2003).
Aktivácia ionotropných a metabotropných receptorov vyvoláva ok-rem zmien priepustnosti iónových kanálov a následných zmien pola-rity membrán neurónov aj ďalšie deje (od aktivácie proteínkináz až pozmeny na úrovni génovej expresie). Ide väčšinou o deje, ktoré prebie-hajú s určitou latenciou a často pretrvávajú aj dlhší čas v porovnaní sozmenami polarity membrán.Excitačné a inhibičné neuróny pôsobia prevažne prostredníctvom
uvoľňovania neurotransmiterov, ktoré reagujú s ionotropnými recep-tormi. Modulačné neuróny pôsobia prostredníctvom neurotransmite-rov, ktoré sa viažu na metabotropné receptory (Gu, 2002). Toto de-lenie však nie je striktné. Napríklad acetylcholín uvoľnený z pregan-gliových neurónov v autonómnych gangliách alebo z motorických ne-urónov predných rohov miechy v oblasti nervovosvalovej platničky poväzbe na ionotropné (nikotínové) receptory vyvoláva rýchlu excitáciupostgangliových autonómnych neurónov, alebo kontrakciu priečne pru-hovaného svalstva. V mozgu však môže acetylcholín väzbou na metabo-
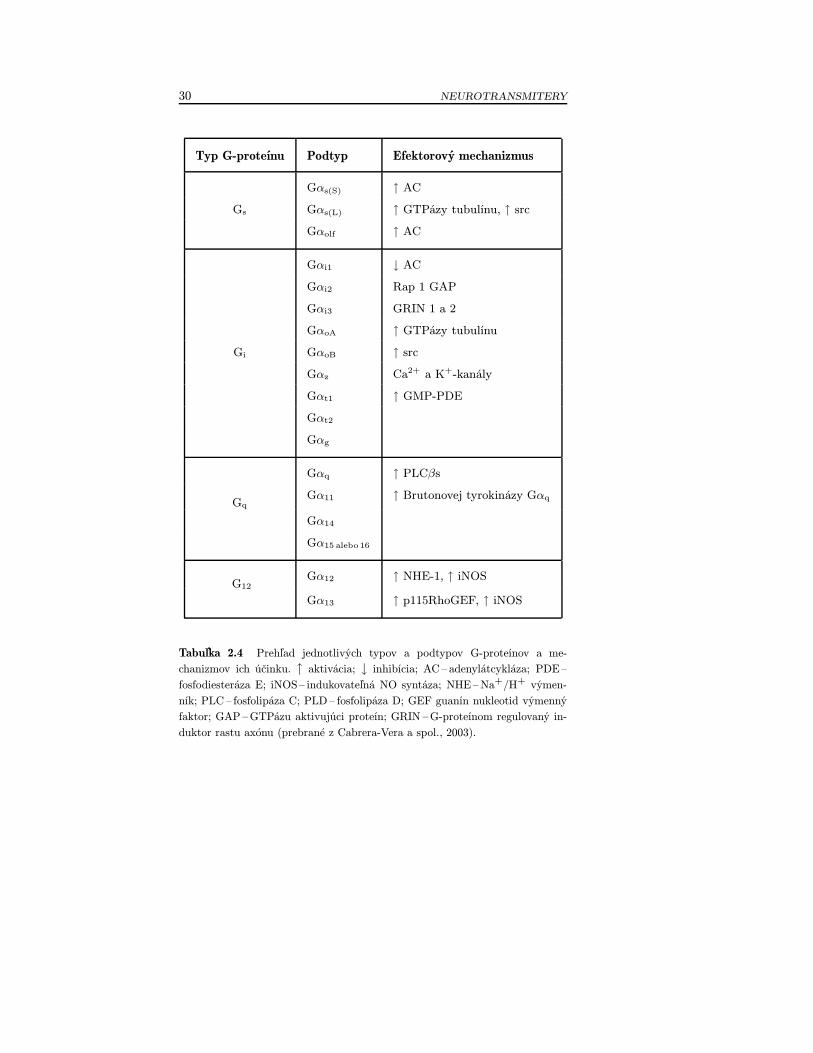
30 NEUROTRANSMITERY
Typ G-proteínu Podtyp Efektorový mechanizmus
Gαs(S) ↑ AC
Gs Gαs(L) ↑ GTPázy tubulínu, ↑ src
Gαolf ↑ AC
Gαi1 ↓ AC
Gαi2 Rap 1 GAP
Gαi3 GRIN 1 a 2
GαoA ↑ GTPázy tubulínu
Gi GαoB ↑ src
Gαz Ca2+ a K+-kanály
Gαt1 ↑ GMP-PDE
Gαt2
Gαg
Gαq ↑ PLCβs
GqGα11 ↑ Brutonovej tyrokinázy Gαq
Gα14
Gα15 alebo 16
G12Gα12 ↑ NHE-1, ↑ iNOS
Gα13 ↑ p115RhoGEF, ↑ iNOS
Tabuľka 2.4 Prehľad jednotlivých typov a podtypov G-proteínov a me-chanizmov ich účinku. ↑ aktivácia; ↓ inhibícia; AC – adenylátcykláza; PDE–
fosfodiesteráza E; iNOS– indukovateľná NO syntáza; NHE–Na+/H+ výmen-
ník; PLC– fosfolipáza C; PLD– fosfolipáza D; GEF guanín nukleotid výmennýfaktor; GAP–GTPázu aktivujúci proteín; GRIN –G-proteínom regulovaný in-
duktor rastu axónu (prebrané z Cabrera-Vera a spol., 2003).

2 Neurotransmiterové systémy 31
Obr. 2.3 Schematické znázornenie aktivácie ionotropného receptora.A) uzavretý iónový kanál; B) obsadenie ionotropného receptora vedie ku konfor-mačnej zmene, ktorej výsledkom je zmena priepustnosti pre ióny (modifikované
podľa Purves, 2004).
tropné (muskarínové) receptory vyvolávať pomalšiu, modulačnú zmenuna postsynaptickom neuróne.Okrem uvedených receptorov môžu niektoré neurotransmitery akti-
vovať intraneuronálne lokalizované receptory, resp. enzýmy. Príkladomsú plynné neurotransmitery, ktoré môžu uplatňovať svoj efekt väzbouna intracelulárne lokalizovaný enzým guanylátcyklázu (Guix a spol.,2005).Neurosteroidy môžu pôsobiť nielen prostredníctvom väzby na in-
tracelulárne receptory, ale aj prostredníctvom väzby na membránovéreceptory. Väzba neurosteroidov na intracelulárne receptory vyvolávazmeny, ktoré sa prejavujú až po určitom čase a môžu pretrvávať ajdlhšie. Môže ísť o tvorbu nových proteínov, napr. receptorov, iónovýchkanálov. Naproti tomu väzba na membránové receptory môže vyvolaťrelatívne rýchle zmeny v činnosti neurónov ovplyvnením aktivity re-ceptorov pre iné neurotransmitery (napr. GABAA-receptory). Predpo-

32 NEUROTRANSMITERY
Obr. 2.4 Schematické znázornenie aktivácie metabotropného receptora. A) ne-obsadený metabotropný receptor; B) obsadenie metabotropného receptora vedie
k aktivácii G-proteínu (zložený z podjednotiek α a βγ). Aktivácia G-proteínunásledne modifikuje intracelulárne procesy prostredníctvom druhých poslov. Vý-
sledkom týchto dejov môže byť nielen zmena excitability neurónu, ale aj ovplyv-
nenie génovej expresie. V uvedenom prípade väzba neurotransmitera s meta-botropným receptorom vedie k aktivácii Gs-proteínu, ktorý indukuje produkciu
cAMP enzýmom adenylátcyklázou (AC). Aktivácia proteínkinázy A (PKA) pro-
stredníctvom cAMP vedie k fosforylácii intracelulárnych molekúl. Fosforyláciaiónového kanála prostredníctvom PKA vedie k jeho uzavretiu (modifikované
podľa Kandel a spol., 2000).

2 Neurotransmiterové systémy 33
kladá sa, že rýchly účinok neurosteroidov môže byť sprostredkovaný ajreceptormi spriahnutými s G-proteínmi (Rupprecht a Holsboer, 1999;Norman a spol., 2004; Magnaghi a spol., 2006).
Účinky neurotransmiterov
Neurotransmitery môžu vyvolať veľké spektrum účinkov.
• Zmeny na postsynaptickej membráne:
– depolarizácia postsynaptickej membrány,
– hyperpolarizácia postsynaptickej membrány.
Depolarizácia alebo hyperpolarizácia membrány postsynaptickéhoneurónu patrí k základným prejavom účinku neurotransmiterov. Na zá-klade tohto efektu možno preto neurotransmitery deliť na excitačnéa inhibičné. Niektoré neurotransmitery však účinkujú tak, že prevažnemodulujú excitabilitu postsynaptickej membrány, a označujú sa akomodulačné.
• Zmeny génovej expresie v neuróne. Neurotransmitery môžu pro-stredníctvom druhých poslov indukovať fosforyláciu transkrip-čných proteínov. Fosforylované transkripčné faktory následne mo-difikujú génovú expresiu v neuróne. Výsledkom modifikovanej gé-novej expresie je syntéza nových proteínov (obr. 2.5) (Haddad,2005). Tento typ pôsobenia neurotransmiterov vyvoláva zmeny,ktoré pretrvávajú niekoľko dní až mesiacov. Takéto dlhodobézmeny sú pravdepodobne významné pre vývoj neurónov a pro-cesy tvorby dlhodobej pamäte. Neurotransmitery môžu induko-vať lokálnu syntézu proteínov v špecifických dendritických tŕňoch.Predpokladá sa, že je to mechanizmus, ktorý sa zúčastňuje nadlhodobých štruktúrnych a funkčných zmenách synáps (Kandela spol., 2000).
• Trofické účinky. Príkladom môže byť GABA, o ktorej sa predpo-kladá, že počas maturácie nervového systému ovplyvňuje prolife-ráciu, migráciu a diferenciáciu buniek nervového systému, matu-ráciu synáps a zánik buniek (Owens a Kriegstein, 2002).

34 NEUROTRANSMITERY
Obr. 2.5 Schematické znázornenie vplyvu neurotransmiterov na génovú
expresiu v neuróne. AC– adenylátcykláza; CREB– cAMP-response-element-
binding protein; CaM–kalmodulín; CaMKIV– kalmodulín dependentná ki-náza IV; TH– tyrozínhydroxyláza; BDNF–mozgový rastový faktor; CRH–
kortikoliberín; GluR1R –podjednotka receptora pre glutamát (modifikované
podľa Kandel a spol., 2000; Carlezon a spol., 2005).

2 Neurotransmiterové systémy 35
• Zmeny na presynaptickej membráne. Presynapticky lokalizo-vané receptory pre neurotransmiter, ktorý je uvoľňovaný z tohoistého presynaptického nervového zakončenia, sa označujú akopresynaptické autoreceptory. Ako presynaptické heteroreceptorysa označujú receptory lokalizované na presynaptickej membráne,ktoré však obsadzuje neurotransmiter uvoľňovaný z iného ner-vového zakončenia. Autoreceptory a heteroreceptory nachádza-júce sa na presynaptickom nervovom zakončení modulujú uvoľ-ňovanie neurotransmiterov z tohto nervového zakončenia. Obsa-denie týchto receptorov ich agonistami väčšinou spôsobuje zníže-nie uvoľňovania neurotransmitera. Aktivácia autoreceptorov uvoľ-neným neurotransmiterom preto pôsobí ako negatívna spätnáväzba, ktorá inhibuje ďalšie uvoľňovanie neurotransmiterov. Pre-synapticky lokalizované heteroreceptory umožňujú komplexnú in-terakciu medzi jednotlivými neurotransmiterovými systémami,ktorej výsledkom je komplikovaná, ale precízna regulácia uvoľ-ňovania neurotransmiterov v nervovom systéme (Langer, 1997).
Neurotransmitery zabezpečujú v nervovom systéme prenos signálov,kovalentné modifikácie proteínov a zmeny v génovej expresii neurónov.Okrem toho sú neurotransmitery zapojené aj v regulácii počtu iónovýchkanálov, denzity receptorov pre neurotransmitery, množstva a charak-teristík synáps a ďalších procesov. Všetky tieto účinky sú podkladomzapojenia neurotransmiterov do komplexných homeostatických dejovv nervovom systéme (Spitzer a spol., 2005).
Nomenklatúra neurotransmiterových systémov mozgu
Po preukázaní syntézy monoamínov (katecholamínov, sérotonínu, hista-mínu) v CNS sa v 50. rokoch 20. storočia (Twarog a Page, 1953; Rama-chandran, 2002) neuróny začali deliť do jednotlivých skupín podľa syn-tetizovaného neurotransmitera. Jednotlivé skupiny sa označujú podľahlavného neurotransmitera, ktorý neuróny syntetizujú, s pridaním prí-pony -ergický. Napríklad neuróny syntetizujúce dopamín tvoria dopa-mínergický systém, neuróny syntetizujúce sérotonín tvoria sérotoníner-gický systém atď.Na tomto mieste však treba podotknúť, že jeden neurón syntetizuje
väčšinou viac neurotransmiterov. Preto sú jednotlivé zoskupenia neuró-nov pomenované podľa ich hlavného neurotransmitera (napr. GABAer-

36 NEUROTRANSMITERY
gický, dopamínergický, sérotonínergický systém neurónov). To však ne-vylučuje možnosť, že v daných neurónoch je syntetizovaný aj iný neu-rotransmiter, príp. iné neurotransmitery.Väčšina neurotransmiterových systémov zahŕňa neuróny, ktoré sú
často lokalizované vo viacerých oblastiach mozgu. Zo začiatku sa častocharakterizovala lokalizácia neurónov syntetizujúcich určitý neurotrans-miter v danej oblasti použitím latinskej, resp. anglickej nomenklatúrymozgových oblastí, resp. jadier.Neuróny, ktoré syntetizujú určitý neurotransmiter v danej oblasti
nervového systému, však nie sú väčšinou zoskupené tak, že by sa na-chádzali striktne vo vymedzenej oblasti, resp. iba v jednom mozgovomjadre. Tieto neuróny sú skôr rozptýlené v určitom objeme mozgovéhotkaniva, čo zapríčiňuje, že sa nachádzajú vo viacerých anatomicky defi-novaných štruktúrach. Napríklad noradrenalínergické neuróny v dolnejčasti v zadnej oblasti predĺženej miechy sú rozptýlené tak, že sa nachá-dzajú nielen v nucleus tractus solitarii, ale aj v jeho okolí.Na základe potreby rozlišovať jednotlivé zoskupenia neurónov synte-
tizujúce určitý neurotransmiter sa zaviedlo označenie zložené z písmenaa číslice, ktoré jednoznačne charakterizuje skupinu neurónov syntetizu-júcu daný neurotransmiter (tab. 2.5). Napríklad vyššie uvedené neurónysyntetizujúce noradrenalín, ktoré sa nachádzajú v oblasti nucleus trac-tus solitarii a jeho okolí, sa označujú ako neuróny A2. Písmenom A saoznačujú neuróny syntetizujúce noradrenalín a číslica 2 charakterizujeich anatomickú lokalizáciu (Dahlstrom a Fuxe, 1964).
Neurotransmitery v periférii a v mozgu – úlohahematoencefalickej bariéryB.Mravec, I. Hulín
Neurotransmitery sa uvoľňujú v CNS aj v periférnom nervovomsystéme (PNS). PNS využíva predovšetkým acetylcholín, ale aj norad-renalín, ATP a ďalšie neurotransmitery. Neurotransmitery aj periférneuvoľňované hormóny, ktoré v CNS pôsobia ako neurotransmitery, môžuza určitých situácií dosahovať vysoké plazmatické hladiny. Príkladommôže byť stresová situácia, počas ktorej dochádza k výraznému uvoľne-niu noradrenalínu zo zakončení sympatikového nervstva aj k výraznémuuvoľneniu adrenalínu a v menšej miere aj noradrenalínu z drene nad-

2 Neurotransmiterové systémy 37
Syntetizovaný Označenie neurónov Označenie jednotlivýchneurotransmiter syntetizujúcich daný zoskupení neurónov
neurotransmiter
Sérotonín B B1 –B9
Dopamín A A8–A17
Noradrenalín A A1–A7
Adrenalín C C1 –C3
Acetylcholín CH CH1–CH6
Histamín E E1 –E5
Tabuľka 2.5 Označovanie skupín neurónov podľa typu neurotransmitera, ktorýsyntetizujú.
obličiek. Počas stresovej situácie môžu preto katecholamíny v plazmedosiahnuť niekoľkonásobne vyššiu koncentráciu v porovnaní s pokojo-vými hodnotami. V plazme sa teda nachádzajú látky, ktoré by mohlipôsobiť v CNS ako neurotransmitery. Tieto látky však väčšinou ne-ovplyvňujú činnosť neurónov CNS. Je to zapríčinené tým, že mozgovékapiláry spolu a astrocytmi vytvárajú tzv. hematoencefalickú bariéru(blood-brain-barrier; BBB) (Rubin a Staddon, 1999).BBB tvorí morfologické a funkčné rozhranie medzi systémovou cir-
kuláciou a nervovým tkanivom mozgu, ktoré reguluje zloženie extra-celulárnej tekutiny v CNS. Vytvára systém, ktorý chráni mozog prednepriaznivým vplyvom zmien v systémovej cirkulácii. BBB zodpovedáza udržiavanie nízkej extracelulárnej koncentrácie aminokyselín a pro-teínov v mozgu v porovnaní s krvou. Obmedzuje aj interakcie s imunit-ným systémom a ovplyvňuje prestup liekov z plazmy do mozgu. BBBreguluje prestup látok z mozgu do systémovej cirkulácie.

38 NEUROTRANSMITERY
Hematoencefalická bariéra
Pod pojem hematoencefalická bariéra sa zahrnuje regulácia kinetikypresunu látok:
• z krvi do tkaniva CNS a späť,
• z krvi do mozgovomiechového likvoru a späť,
• z likvoru do nervového tkaniva a späť.
Na všetkých troch úrovniach ide o o špecifickú kinetiku. Túto sku-točnosť potvrdzujú rozdielne koncentrácie látok a rozdielna rýchlosť ichprestupu.
Morfologické základy hematoencefalickej bariéry a štruktúrneodlišnosti
• Medzi endotelovými bunkami sú pravé „tight junctionsÿ. Endo-telové bunky sú v mozgových kapilárach usporiadané tak, že ne-umožňujú prienik látok cez intercelulárne priestory.
• Mozgové kapiláry nemajú fenestrácie. Preto molekuly môžu pre-chádzať iba cez membrány endotélií.
• Okolo kapilár nie je voľný perikapilárny priestor. V tesnej blízkostikapilár astroglia vytvára súvislý val.
Endotel kapilár má veľmi nízku aktivitu transportných vezikúl a pi-nocytózy. Veľmi nízka endocytová a a exocytová aktivita je najmä naluminálnej membráne endotelu. V dôsledku tejto skutočnosti je v endo-telových bunkách znížené množstvo transportných vezikúl. Preto je ibaveľmi malý transport makromolekúl cez endotel mozgových kapilár.Súčasťou hematoencefalickej bariéry sú teda mozgové kapiláry so
špecifickou morfológiou, ktoré neumožňujú voľný priechod väčšiny lá-tok (Johnson a Gross, 1993). BBB vytvárajú tri bunkové elementy (en-dotelové bunky, výbežky astrocytov a pericyty) a bazálnu membránukapilár. Tesné spojenia (tight junctions) medzi endotelovými bunkamimozgových ciev vytvárajú bariéru, ktorá selektívne zabraňuje väčšinelátok nachádzajúcich sa v krvi vstupovať do mozgu. Malé lipofilné látky,ako O2 a CO2, difundujú voľne cez plazmatické membrány v smere ich

2 Neurotransmiterové systémy 39
koncentračného gradientu. Endotelové bunky mozgových kapilár obsa-hujú veľké množstvo špecifických transportérov, ktoré umožňujú regu-lovaný prenos látok do tkaniva mozgu (napr. glukózu, aminokyseliny,nukleotidy), ale aj opačným smerom, z mozgového tkaniva do systémo-vej cirkulácie (napr. P-glykoproteín, transportujúci xenobiotiká). Trans-portné systémy BBB zabezpečujú aj prenos niektorých neurotransmi-terov, ich prekurzorov, ako aj ich metabolitov cez hematoencefalickúbariéru (Ohtsuki, 2004). Prechod veľkých molekúl (napr. inzulín, lep-tín) cez BBB zabezpečuje od receptorov závislá endocytóza (Ballabha spol., 2004).
Funkčné charakteristiky hematoencefalickej bariéry, ktorémajú morfologický základ
• Asymetria funkcie luminálnej a abluminálnej membrány je príči-nou toho, že transport látok v smere mozog - krv je aktívnejší.
• Takmer nulový transport vysokomolekulových látok cez hema-toencefalickú bariéru podmieňuje prítomnosť tesných spojení(„tight junctionsÿ) medzi endotelovými bunkami kapilár v mozgu.Druhým faktorom takmer nulového transportu vysokomoleku-lových látok je absencia fenestrácií v kapilárach. Tieto látkysa môžu transportovať pinocytózou a transportnými vezikulami.Oba tieto procesy sú však iba minimálne.
• Transport nízkomolekulových látok cez medzibunkové kontaktyje iba minimálny. Transport látok z krvi do mozgu sa podobá natransmembránový transport alebo na transepitelový transport.
Hematolikvorová bariéra
Morfologický substrát hematolikvorovej bariéry sa týka procesov, ktoréprebiehajú najmä v plexus chorioideus.Kapiláry v plexus chorioideus sú permeabilné. Permeabilita je za-
bezpečená tým, že:
• endotelové bunky kapilár sú v tejto oblasti spojené kontaktmitypu gap junction,
• spojenie buniek je „permeabilnejšieÿ najmä pre nízkomolekulovélátky,

40 NEUROTRANSMITERY
• v stene kapilár sú fenestrácie. Kapiláry v plexus chorioideus súpodobné kapiláram v systémovej cirkulácii,
• vo vnútri endotelových buniek je veľa mikrovezikúl,
• transcelulárny transport je aktívnejší ako v iných mozgových ka-pilárach,
• obmedzenie permeability zabezpečuje intercelulárne spojenie me-dzi epitelovými bunkami plexus chorioideus.
Bunky plexus chorioideus sú vzájomne spojené medzibunkovýmikontaktmi typu tight junction spojenia sú permeabilnejšie ako tightjunction mozgových kapilár v iných oblastiach mozgu. Táto skutočnosťje významná najmä pre tvorbu likvoru v plexus chorioideus.
Likvoroencefalická bariéra
Existencia likvoroencefalickej bariéry je diskutabilná. Jediným znakomjej existencie sú relatívne úzke extracelulárne priestory medzi jednot-livými bunkami v CNS a medzi ich výbežkami. Nedostatok „morfolo-gických dôkazovÿ nemôže vyvrátiť existenciu kinetiky presunu látok natejto úrovni.
Transportné mechanizmy hematoencefalickej bariéry
Transport látok cez hematoencefalickú bariéru je zabezpečený viace-rými transportnými systémami.
• Difúzia a prestup cez iónové kanály. Takmer „pasívneÿ prestu-pujú cez hematoencefalickú bariéru lipoidné látky a niektoré maléneutrálne molekuly, ako sú O2, CO2 a voda. Prestup vody zabez-pečujú akvaporíny. Bez účasti aktívnych transportných systémovprestupujú malé ióny, napr. Na+ a Cl−.
• Aktívny transport má niektoré špecifiká. Prvým je stereošpeci-fickosť. To znamená, že napríklad D-glukóza je aktívne transpor-tovaná do mozgu, ale nie je transportovaná L-glukóza. Transportniektorých látok môže byť inhibovaný štruktúrne podobnými mo-lekulami. Pre niektoré látky existujú špecifické transportné sys-témy. Takýmito látkami sú D-glukóza, laktát, L-tyrozín, L-gluta-mát, L-arginín, adenín, adenozín a viaceré solúty.

2 Neurotransmiterové systémy 41
• Enzymatické cesty.Transport niektorých látok je čiastočne bloko-vaný enzýmami endotelových buniek. Mozgové kapiláry obsahujúmonoaminooxidázu a acetylcholínesterázu, ktoré rýchlo metabo-lizujú sérotonín a acetylcholín, alebo ich konvertujú na neaktívneformy. Enzymaticky sa inaktivujú aj enkefalíny. Enzymatická ba-riéra funguje veľmi účinne. Niektoré substráty sa transportujú ceztight junctions. Tým sa „obchádzajúÿ enzymatické cesty.
Regionálne rozdiely hematoencefalickej bariéry – rozdielyvo vaskularizácii jednotlivých oblastí CNS
Denzita kapilár je v šedej hmote 2–5-krát vyššia ako v bielej hmote. Ajv šedej hmote sú rozdiely vo vaskularizácii jednotlivých oblastí.
Ependýmové bunky
Tieto bunky tvoria výstelku komôr. Ependým na povrchu cirkumventri-kulárnych orgánov je pokrytý veľkým počtom riasiniek. V ependýme sú„zabudovanéÿ špecializované gliové bunky, ktoré sa označujú tanycyty.Tanycyty vysielajú dlhé výbežky do okolitého neuropilu.
Cirkumventrikulárne orgány
Hematoencefalická bariéra však nie je vytvorená úplne vo všetkých ob-lastiach CNS. Existujú oblasti s fenestrovanými kapilárami, ktoré umož-ňujú relatívne voľný prechod látok z cirkulácie do CNS a naopak. Tietooblasti vytvárajú tzv. cirkumventrikulárne orgány (CVOs) (obr. 2.6)(Ganong, 2000).Plexus chorioideus, subfornikálny orgán, organum vasculosum lami-
nae terminalis, eminentia mediana, epifýza, neurohypofýza a area po-strema majú odlišnú štruktúru kapilár v porovnaní s kapilárami CNS.Kapiláry sú fenestrované a endotelové bunky vytvárajú spojenia typugap junctions. Obsahujú veľa transportných vezikul. Fenestrované kapi-láry umožňujú prestup hormónov z krvi a umožňujú aj neurosekréciu.Na základe toho sa cirkumventrikulárne orgány rozdeľujú na (Cottrella Ferguson, 2004):
• senzorické, ktoré umožňujú monitorovať koncentráciu chemic-kých látok v krvi (organum subfornicale, organum vasculosumlaminae terminalis, area postrema),

42 NEUROTRANSMITERY
Obr. 2.6 Lokalizácia cirkumventrikulárnych orgánov v mozgu človeka (svetláfarba) a endokrinných žliaz mozgu s fenestrovanými kapilárami (tmavá farba).AP– area postrema; AH– adenohypofýza; EPI – epifýza; ME– eminentia me-
diana; NH–neurohypofýza; OVLT– organum vasculosum laminae terminalis;
SFO– organum subfornicale; SCO– organum subcommissurale (modifikovanépodľa Ganong, 2000).
• sekrečné, ktoré umožňujú prestup látok z CNS do cirkulácie (neu-rohypofýza, eminentia mediana),
• nezaradený (organum subcommissurale).
Príkladom látky, ktorej koncentrácia je v plazme monitorovaná pro-stredníctvom cirkumventrikulárnych orgánov, je angiotenzín II, ktorýsa zúčastňuje na modulácii tlaku krvi a homeostázy tekutín a elektro-lytov. Cirkulujúci angiotenzín II môže prostredníctvom cirkumventri-kulárnych orgánov modulovať procesy v CNS, ktoré sú spojené s re-guláciou objemu telesných tekutín a krvného tlaku. Je zaujímavé, žeangiotenzín II sa uvoľňuje aj neurónmi v CNS, ktoré sa zúčastňujúpráve na regulácii objemu telesných tekutín a krvného tlaku (Johnson

2 Neurotransmiterové systémy 43
a Gross, 1993; Dampne a spol., 2002). Veľmi významné je aj monito-rovanie plazmatických hladín hormónov, ktorých biosyntézu a sekréciureguluje hypotalamo-hypofýzový systém. Inými látkami, ktoré ovplyv-ňujú činnosť CNS prostredníctvom cirkumventrikulárnych orgánov, súcytokíny (Buller, 2001).Viaceré periférne syntetizované hormóny ovplyvňujú činnosť mozgu.
Významnou skupinou sú hormóny syntetizované v tráviacom a tuko-vom tkanive (napr. cholecystokinín, VIP, leptín), ktoré ovplyvňujú čin-nosť mozgových oblastí zapojených do modulácie príjmu potravy. Nie-ktoré z týchto oblastí (napr. nucleus arcuatus) sú umiestnené v blíz-kosti CVOs, čo umožňuje periférne syntetizovaným hormónom priamoovplyvňovať ich činnosť. Plocha ciev v CVOs je však pri porovnaní s cel-kovou plochou ciev v mozgu niekoľkonásobne menšia. Preto sa predpo-kladá, že periférne syntetizované hormóny do mozgu prechádzajú z väč-šej časti prostredníctvom transportných systémov BBB (Kastin a Pan,2000).Príkladom látok, ktoré prechádzajú prostredníctvom cirkumventri-
kulárnych orgánov do cirkulácie, sú hormóny neurohypofýzy (Fergusona Bains, 1996).Oproti iným chemickým látkam môžu steroidové látky prechádzať
voľne do CNS (existujú však mechanizmy, ktoré aktívne prenášajú ste-roidové látky späť do systémovej cirkulácie). Takto môžu periférnesyntetizované steroidy pôsobiť ako neuromodulátory v CNS (Ganong,2000).
Ďalšie štruktúry mozgu s fenestrovanými kapilárami
Okrem cirkumventrikulárnych orgánov sa v mozgu nachádzajú ďalšieštruktúry, ktoré obsahujú fenestrované kapiláry (obr. 2.6). Patria me-dzi ne adenohypofýza a epifýza, endokrinné žľazy, ktoré nie sú z on-togenetického hľadiska súčasťami mozgu (Ganong, 2005). Fenestrovanékapiláry uvedených endokrinných žliaz umožňujú prestup hypotalamic-kých regulačných hormónov a epifýzového melatonínu do systémovejcirkulácie.Hematoencefalická bariéra je dynamická štruktúra. Jej stabilita sa
zvyšuje pôsobením glukokortikoidov (Dietrich, 2004; Förster, 2005). Po-čas záťažových situácií môže však dôjsť k jej narušeniu (Skultetyovaa spol., 1998; Bhattacharjee a spol., 2001).

44 NEUROTRANSMITERY
Narušenie BBB sa môže zúčastňovať na vzniku viacerých ochorení,medzi ktoré patria napríklad mozgový edém a autoimunitné ochorenia(Edwards, 2001).
Neurotransmitery z evolučného hľadiska
Hlavne v minulosti sa pri štúdiu základných, alebo aj komplexnýchfunkcií nervového systému často ignoroval evolučný pohľad (Damasio,1997). Poznatky získané z porovnávacej anatómie a pohľad z evoluč-ného hľadiska môžu však významne uľahčiť pochopenie funkcie takéhokomplexného systému, akým je ľudský mozog. Poodhalenie procesovspojených s evolúciou mozgu je náplňou porovnávacej neuroanatómiev posledných viac ako 100 rokoch (Fritzsch, 1998).Neurotransmitery sa zákonite objavili so vznikom nervového sys-
tému. Neprekvapuje preto ich široký výskyt v živočíšnej ríši. Predpo-kladá sa, že v procesoch vzniku enzýmov pre biosyntézu malomoleku-lových neurotransmiterov mohol mať úlohu aj prenos génov pre tietoenzýmy z baktérií do živočíšnych buniek (Iyer a spol., 2004).O rozšírenosti neurotransmiterov v živočíšnej ríši svedčí aj to, že
v nervovom systéme hmyzu boli identifikované ako neurotransmiteryacetylcholín, dopamín, oktopamín, sérotonín, histamín, aminokyselinya rôzne neuropeptidy (bližšie pozri Osborne, 1996). Iným príkladomsú senzorické (respektíve senzori-motorické) bunky ektodermy chápa-diel jednoduchých živočíchov kmeňa mechúrnikov (napr. koraly, veter-nice), ktoré obsahujú katecholamíny. Rozšírenosť katecholamínov v žijú-cich organizmoch poukazuje na ich dlhú fylogenetickú históriu (Smeetsa González, 2000). Ďalším neurotransmiterom hojne sa vyskytujúcimv živočíšnej ríši je sérotonín. Predpokladá sa, že základná organizáciacentrálnej časti sérotonínergického nervového systému u stavovcov savyskytovala už v línii bezstavovcov (Hay-Schmidt, 2000).Okrem sledovania výskytu jednotlivých neurotransmiterov sa sle-
duje aj výskyt jednotlivých typov receptorov pre tieto neurotransmi-tery. Tak napríklad receptory pre opioidy (µ-receptory) sa objavili prav-depodobne ešte pred objavením sa stavovcov. To, že ich funkčné a far-makologické vlastnosti sú konzervované viac ako 400 miliónov rokov,svedčí o ich dôležitosti (Darlison a spol., 1997).V prípade neuropeptidov, ktoré pôsobia prostredníctvom viacerých
typov receptorov, sa predpokladá, že najskôr došlo ku génovým zme-

2 Neurotransmiterové systémy 45
nám, ktoré podmienili vznik viacerých typov neuropeptidov (pôsobiliiba na určitý počet receptorov) a až následne vznikli jednotlivé subtypyreceptorov (Darlison a Richter, 1999).Zaujímavý je pohľad na vznik ionotropných a metabotropných re-
ceptorov pre daný neurotransmiter. Napríklad pri receptoroch pre glu-tamát sa predpokladá, že nevznikli zo spoločného predchodcu. Zdása však, že väzbové miesto pre glutamát, ktoré sa vyskytuje na jehoionotropných, ako aj metabotropných receptoroch, je od spoločnéhopredchodcu odvodené. Tento predchodca bol základom pre vznik iono-tropných receptorov. Zároveň genetická rekombinácia génu pre väzbovémiesto pre glutamát spolu s génom pre receptor viazaný s G-proteínompravdepodobne vytvorila výsledný gén pre metabotropný receptorovýkomplex viažuci glutamát (Zakon, 2002).Bližšie pochopenie evolučného vývoja jednotlivých neurotransmite-
rových systémov umožnili hlavne molekulovobiologické metódy. Určeniesekvencie báz DNA génov kódujúcich neuropeptidy, enzýmy potrebnépre biosyntézu malomolekulových neurotransmiterov a receptory prejednotlivé neurotransmitery umožňuje na základe medzidruhového po-rovnávania určiť priebeh fylogenézy neurotransmiterových systémov.Tieto poznatky prispievajú k lepšiemu pochopeniu funkcie neurotrans-miterov v nervovom systéme.

Malomolekulové neurotransmitery
B. Mravec
Medzi prvé identifikované neurotransmitery patria acetylcholín, bio-génne amíny a niektoré aminokyseliny. Neskôr sa táto skupina malomo-lekulových neurotransmiterov rozrástla o puríny, neurosteroidy, kana-binoidy a ďalšie látky.
Aminokyseliny
Medzi neurotransmitery patria viaceré aminokyseliny (tab. 3.1). Na zá-klade charakteru odpovede vyvolanej na postsynaptickom neuróne sadelia na dve skupiny:
• excitačné aminokyseliny (glutamát, pravdepodobne aj aspartát),
• inhibičné aminokyseliny (GABA, glycín).
Zaradenie niektorých aminokyselín medzi neurotransmitery nará-žalo v minulosti na viaceré ťažkosti, ktoré vyplývali z toho, že skúmanéaminokyseliny sa vyskytujú v neurónoch nielen ako neurotransmitery,ale často aj ako stavebné zložky proteínov a produkty intermediárnehometabolizmu (prehľad histórie výskumu aminokyselín ako neurotrans-miterov pozri Bennett a Balcar, 1999). Dôkaz prítomnosti vysokoafinit-ných transportérov pre tieto látky umožnil určiť, ktoré neuróny využí-vajú určité aminokyseliny ako neurotransmitery (Webstel, 2001).
46

3 Malomolekulové neurotransmitery 47
Esenciálne Neesenciálne
arginín (Arg; R)# alanín (Ala; A)
histidín (His; H)# aspartát (Asp; D)†
izoleucín (Ile; I) asparagín (Asn; N)
leucín (Leu; L) cysteín (Cys; C)†#
lyzín (Lys; K) glutamín (Gln; Q)#
metionín (Met; M) glutamát (Glu, E)*#
fenylalanín (Phe; F)# glycín (Gly; G)∗
treonín (Thr; T) prolín (Pro; P)
tryptofán (Trp; W)# serín (Ser; S)#
valín (Val; V) tyrozín (Tyr; Y)#
Tabuľka 3.1 Prehľad esenciálnych a neesenciálnych aminokyselín nachádzajú-cich sa v ľudskom organizme (v zátvorke sú uvedené dva ekvivalenty medziná-rodných skratiek); ∗ neurotransmiter; † predpokladaný neurotransmiter;# aminokyselina zapojená v biosyntéze neurotransmiterov.
Excitačné a inhibičné aminokyseliny sa viažu prevažne na iono-tropné receptory. Prostredníctvom tejto väzby vyvolávajú relatívnerýchlu zmenu priepustnosti postsynaptickej membrány. Väzbou gluta-mátu alebo GABA na metabotropné receptory vyvolávajú pomalšiea spravidla aj dlhšie trvajúce reakcie.
Glutamát
Nález vysokých hladín voľného l-glutamátu v mozgu viedol k štúdiujeho úlohy v nervovom systéme. Na začiatku 50. rokov 20. storočia opí-sal Hayashi záchvatové stavy, ktoré vznikli po intracerebroventrikulár-nom, resp. intrakarotickom podaní glutamátu. O niečo neskôr preuká-zali Curtis a spol., že glutamát vyvoláva depolarizáciu a excitáciu neu-rónov CNS (Weinberg, 1999; Watkins, 2000). Pokusy s ionoforetickou

48 NEUROTRANSMITERY
Obr. 3.1 Schéma biosyntézy glutamátu (modifikované podľa Purvesa spol., 2001).
aplikáciou glutamátu viedli k jeho začleneniu medzi neurotransmitery(Krnjevic a Phillips, 1963). Roku 1971 sa zaviedla metodika na iden-tifikáciu glutamátergických neurónov založená na dôkaze prítomnostivysokoafinitného vychytávania glutamátu (Wofsey a spol., 1971).Glutamát uvoľnený z glutamátergických neurónov je hlavným exci-
tačným neurotransmiterom takmer vo všetkých oblastiach CNS. Odha-duje sa, že až 75% excitačného prenosu v mozgu je sprostredkovanéhoglutamátom a že 80–90% synáps v mozgu je glutamátergických. Pro-cesy spojené s glutamátergickou aktivitou (depolarizácia a repolarizáciaglutamátergických nervových zakončení) sa môžu zúčastňovať až 80%na celkovej energetickej spotrebe mozgu (Siegel a spol., 2006).Glutamát je neesenciálna aminokyselina, ktorá je syntetizovaná
v neurónoch z jeho prekurzorov. Predpokladá sa, že glutamát využí-vaný na neurotransmiterové funkcie je syntetizovaný prevažne z gluta-mínu (obr. 3.1) (Purves a spol., 2001). Uvoľnený glutamát pôsobí napostsynaptické a presynaptické receptory. Glutamát sa odstraňuje zosynaptickej štrbiny vysokoafinitným transportérom, ktorý sa nachádzav membránach presynaptického nervového zakončenia a v membránach

3 Malomolekulové neurotransmitery 49
okolitých buniek glie. Gliové bunky metabolizujú glutamát na gluta-mín, ktorý uvoľňujú do extracelulárnej tekutiny, odkiaľ sa transportujespäť do nervových zakončení (Gadea a Lopez-Colome, 2001a).
NMDA ↑gNa+ ↑[Ca2+ ]i
Ionotropné AMPA↑gNa+
Kainát
Trieda ImGlu R1
Gq/11↑PLC ↑DAG
↑[Ca2+ ]i ↑gK+
mGlu R5 ↑IP3
Metabotropné Trieda ImGlu R2
Gi/Go ↓AC ↓cAMP ↓[Ca2+ ]imGlu R3
mGlu R4
Trieda IIImGlu R6
Gi/Go ↓AC ↓cAMP ↓[Ca2+ ]imGlu R7
mGlu R8
Tabuľka 3.2 Klasifikácia receptorov pre glutamát a mechanizmy transdukciesignálu (Coutinho a Knopfel, 2002).
V CNS sa nachádzajú ionotropné aj metabotropné receptory preglutamát (tab. 3.2) (Bettler a Mulle, 1995; Pin a Duvoisin, 1995). Gluta-mát po naviazaní na ionotropné receptory zvyšuje prestup katiónov, čímsa zúčastňuje na excitácii postsynaptického neurónu (Danbolt, 2001;Purves a spol., 2001). AMPA a kainátové receptory sú jednoduchýmikatiónovými kanálmi, ktoré umožňuju prechod Na+ intracelulárne a K+
extracelulárne. NMDA-receptor má niekoľko špecifík. Ide o katiónovýkanál, ktorý však umožňuje presun relatívne veľkého množstva Ca2+.Funkciu NMDA-receptorov facilituje naviazanie glycínu a zdá sa, žepráve tento proces je veľmi dôležitý pre normálnu odpoveď tohto iono-tropného receptora na glutamát. Navyše je pri pokojovom membráno-vom potenciáli kanál NMDA-receptora blokovaný Mg2+. K odbloko-

50 NEUROTRANSMITERY
vaniu NMDA-receptora dochádza iba vtedy, keď je membrána neurónudepolarizovaná. Ionotropné glutamátové receptory sú tvorené viacerýmipodjednotkami. Boli identifikované štyri AMPA, päť kainátových a šesťNMDA-podjednotiek, pričom každá je kódovaná jedným génom. Meta-botropné receptory pôsobia prostredníctvom G-proteínov tak, že zvy-šujú intracelulárnu koncentráciu IP3 a DAG, alebo znižujú hladinu in-tracelulárneho cAMP. Metabotropné glutamátové receptory sú lokalizo-vané postsynapticky aj presynapticky. Sú široko distribuované v CNS,pričom sa predpokladá, že sú spojené aj so synaptickou plasticitou,hlavne v mozočku a v hipokampe (Ganong, 2005).
Obr. 3.2 Schematické znázornenie lokalizácie a dráh glutamátergických neuró-nov v mozgu človeka (modifikované podľa Feldman a Quenzer, 1984).
Medzi hlavné dráhy neurónov, ktoré uvoľňujú glutamát zo svojichsynaptických zakončení, patria (obr. 3.2):
• kortiko-spinálne, kortiko-striatické, kortiko-subtalamické, korti-ko-talamické, kortiko-nigrálne, kortiko-rubrálne, kortiko-pontín-ne, kortiko-bulbárne, kortiko-kortikálne (komisurálne),
• hipokampo-septálne, hipokampo-mamillárne,

3 Malomolekulové neurotransmitery 51
• bulbo-kortikálne, bulbo-talamické,
• subtalamo-pallidálne, subtalamo-nigrálne,
• olivo-cerebelárne (šplhavé vlákna),
• cerebelárne intrinsické spoje: z granulárnych buniek na Purky-ňove bunky,
• spino-talamické,
• talamo-kortikálne, talamo-striatické,
• somatosenzitívne a viscerosenzitívne dráhy (Petrovický, 2002).
Glutamátergické dráhy sú zapojené do regulácie širokého spektra fy-ziologických procesov prebiehajúcich v CNS. Medzi tieto procesy patrímotorická koordinácia, emočné a kognitívne procesy zahŕňajúce ukla-danie a vyvolávanie informácií z pamäte a učenie. Aj prenos prevaž-nej väčšiny senzorických informácií z receptorov prostredníctvom pe-riférnych nervov do CNS sprostredkúva práve glutamát (Siegel a spol.,2006). Príkladom je monosynaptická Ia-motoneurónová dráha, ktorá jehlavným zdrojom monosynaptických senzorických vstupov na motoneu-róny a má významnú úlohu vo vývoji miechových reflexov. AferentnéIa-vlákna svalových vretienok uvoľňujú v predných rohoch miechy glut-mát, ktorý pôsobí excitačne na motoneuróny (Glover, 2000).Predpokladá sa, že glutamátergické neuróny sa zúčastňujú v etiopa-
togenéze ochorení CNS, medzi ktoré patrí epilepsia, amyotrofická late-rálna skleróza, olivopontocereberálna atrofia a Huntingtonova chorea.Veľký význam sa pripisuje úlohe glutamátu v procesoch ischemickéhopoškodenia mozgu (Siegel a spol., 2006; Meldrum, 2000).
Aspartát
Aj keď sa aspartát nachádza v CNS a má excitačný vplyv na neuróny,nie je stále presne charakterizovaná lokalizácia a úloha neurónov využí-vajúcich aspartát ako neurotransmiter (Webster, 2001).Aspartát je syntetizovaný z kyseliny oxaloctovej a glutamátu enzý-
mom aspartátaminotransferázou (obr. 3.3). Nepreukázalo sa však, že bysa aspartát uskladňoval v synaptických vezikulách. S najväčšou pravde-podobnosťou sa preto uvoľňuje priamo z cytosólu nervových zakončení

52 NEUROTRANSMITERY
(Siegel a spol., 2006). Aspartát aktivuje NMDA-receptory, avšak naAMPA-receptory pravdepodobne nepôsobí (Dingledin a spol., 1999).
Obr. 3.3 Schéma biosyntézy aspartátu (modifikované podľa Palmadaa Centelles, 1998).
Predpokladá sa, že aspartát je neurotransmiterom niektorých in-terneurónov, neurónov v hipokampe a neurónov inervujúcich zrakovúkôru (Cotman a Monaghan, 1986; Tsumoto, 1990; Perez-Cerda a spol.,1996).
Iné excitačné aminokyseliny
Glutamát sa považuje za hlavný excitačný aminokyselinový neurotrans-miter v mozgu cicavcov. Predpokladá sa však, že aj iné aminokyselinymôžu spôsobiť excitáciu neurónov. O možnej úlohe aspartátu sa disku-tovalo v predchádzajúcom texte. Medzi ďalšie potenciálne neurotrans-

3 Malomolekulové neurotransmitery 53
mitery, resp. neuromodulátory s excitačným účinkom patria derivátyaminokyselín obsahujúce síru (obr. 3.4):
• deriváty glutamátu: l-homocysteín, l-homocysteínsulfát,
• deriváty aspartátu: l-cysteín, l-cysteínsulfát.
Obr. 3.4 Štruktúrne vzorce homocysteínu, cysteínu a ich derivátov (Shi a spol.,
2003).
Tieto látky pôsobia agonisticky na glutamátergické receptory rôz-nych neurónov. Preukázala sa aj ich syntéza a uvoľňovanie v CNS(Thompson a Kilpatrick, 1996).Uvažuje sa o možnej účasti homocysteínu, cysteínu a ich derivá-
tov v etiopatogenéze Alzheimerovej choroby a schizofrénie (Shi a spol.,2003).
GABA (kyselina γ-aminomaslová)
Podobne ako pri výskume, ktorý sa zaoberal úlohou glutamátu v CNS,sa v 50. a 60. rokoch 20. storočia začala študovať úloha kyseliny γ-ami-nomaslovej (γ-aminobutyric acid, GABA) ako inhibičného neurotrans-mitera v nervovom systéme stavovcov aj bezstavovcov. GABA bola pr-vým neurotransmiterom, pri ktorom sa preukázalo, že vyvoláva synap-tickú inhibíciu (Owens a Kriegstein, 2002; viac pozri sériu článkov v Bio-chemical Pharmacology, 2004; 68 (8) venovaných výskumu GABA).GABA je hlavným inhibičným neurotransmiterom v mozgu, mie-
che a sietnici. GABA je pravdepodobne aj neurotransmiterom v ANS

54 NEUROTRANSMITERY
a endokrinných orgánoch (Gladkevich a spol., 2006). GABA nie je esen-ciálnym metabolitom a nie je ani inkorporovaná do proteínov. Pretodôkaz jej prítomnosti v neuróne poukazuje na to, že neurón využívaGABA ako neurotransmiter (Purves a spol., 2001).
Obr. 3.5 Schéma biosyntézy GABA (modifikované podľa Purves a spol., 2001).
GABA je syntetizovaná z glutamátu enzýmom glutamátdekarboxy-lázou (obr. 3.5). Dôkaz prítomnosti enzýmu glutamátdekarboxylázy jepreto jedným z markerov GABAergických neurónov (Ribak a Roberts,1990). GABA je umiestnená v sekrečných vezikulách. Uvoľnená GABApôsobí na ionotropné alebo metabotropné receptory (tab. 3.3) (Owensa Kriegstein, 2002). Zvyšuje prechod Cl−-iónov do bunky, čím sa zú-častňuje na vzniku hyperpolarizácie postsynaptickej membrány. ÚčinokGABA je ukončený jednak spätným vychytávaním do neurónov, jednakvychytávaním okolitými gliovými bunkami (Gadea a Lopez-Colome,2001b; Purves a spol., 2001).V nezrelom nervovom systéme GABA väzbou na GABAergické re-
ceptory nevyvoláva hyperpolarizáciu, ale naopak depolarizáciu. To jespôsobené tým, že nezrelé neuróny obsahujú intracelulárne vyššiu kon-centráciu Cl− v porovnaní s extracelulárnym priestorom. Preto ot-vorenie iónových kanálov pre Cl− po obsadení ionotropných recepto-rov (GABAA a GABAC) neurotransmiterom GABA vyvolá prechodCl− extraneuronálne, čím dochádza k depolarizácii neurónu (Webster,2001). Až v dozretom nervovom systéme dochádza k takým zmenámv zastúpení iónových púmp a následne v iónovom zložení neurónov, ževäzba GABA na receptory vyvoláva hyperpolarizáciu postsynaptickejmembrány (Ben-Ari, 2002). GABAB metabotropné receptory sú spria-hnuté s G-proteínom. Aktivácia týchto receptorov vedie k zvýšeniu vo-

3 Malomolekulové neurotransmitery 55
IonotropnéGABAA
↑gCl−
GABAC
Metabotropné GABAB Gi/Go ↓AC ↓cAMP↑gK+
↓[Ca2+ ]i
Tabuľka 3.3 Klasifikácia receptorov pre GABA a mechanizmy transdukcie sig-nálu.
divosti pre K+ a k útlmu adenylátcyklázy a vytekania Ca2+, čo vediek hyperpolarizácii neurónov (Ganong, 2005)Väčšinou sa úloha neurotransmitera GABA spája s činnosťou in-
terneurónov lokálnych okruhov (napr. v bazálnych gangliách, v moz-govej kôre). Je však aj neurotransmiterom Purkyňových buniek mo-zočka, ktoré vysielajú dlhé axóny (obr. 3.6) (Gupta a spol., 2000; Purvesa spol., 2001). GABAergické neuróny sa nachádzajú aj v šedej hmotemiechy, s najvyšším výskytom v povrchových laminae I–III (Budai,2000).Niektoré GABAergické neuróny v mozgovej kôre vytvárajú okrem
chemických synáps zároveň aj elektrické synapsy. Chemické synapsysa tvoria medzi axónmi GABAergických neurónov a telami neurónov,kým elektrické synapsy sa tvoria medzi dendritickými výbežkami týchtoneurónov (Fukuda a Kosaka, 2000). Takéto usporiadanie poukazuje nakomplexnosť spracovania informácií v nervovom systéme. Pri štúdiuprocesov, na ktorých sa zúčastňujú chemické synapsy, preto nemožnoignorovať úlohu elektrických synáps.Medzi hlavné dráhy neurónov, ktoré uvoľňujú GABA zo svojich sy-
naptických zakončení, patria (obr. 3.6):
• striato-pallidárne, striato-nigrálne,
• pallido-retikulárne, pallido-talamické,
• nigro-tektálne, nigro-talamické,
• septo-hipokampálne,

56 NEUROTRANSMITERY
Obr. 3.6 Schematické znázornenie lokalizácie a dráh GABAergických neurónovv mozgu (modifikované podľa Feldman a Quenzer, 1984).
• intrinsické: cerebello-nukleárne (Purkyňove bunky-jadrá), intra-kortikálne, intratalamické, inhibičné bunky v mozgovom kmenia v mieche (Petrovický, 2002).
GABA je významne zapojená v regulácii excitability CNS. Predpo-kladá sa, že na úrovni miechy sa GABAergické neuróny zúčastňujú namodulácii monosynaptického aj polysynaptického prenosu signálov.Uvažuje sa, že GABAergické neuróny modulujú prenos nociceptívnychinformácií a presynapticky kontrolujú primárne aferentné vlákna typuAδ a Aβ, čím sa môžu zúčastňovať na presynaptickej selekcii aferent-ného toku informácií do CNS (Rudomin, 1999; Budai, 2000). Alkohol,barbituráty a benzodiazepíny modulujú činnosť GABAA-receptorov.Pri ich chronickom pôsobení dochádza k „downÿ-regulácii GABAA-receptorov.Pretože je GABA hlavným inhibičným neurotransmiterom v mozgu,
uvažuje sa, že narušenie GABAergického systému má významnú úlohuv etiopatogenéze neurologických a neuropsychiatrických ochorení. Pred-pokladá sa, že narušená činnosť GABAergických neurónov sa môže zú-častňovať v etiopatogenéze Huntingtonovej chorey, epilepsie, anxiety,

3 Malomolekulové neurotransmitery 57
tarditívnych dyskinéz, alkoholizmu, schizofrénie, porúch spánku, pre-menštruačnej dysfórie, Parkinsonovej choroby a mentálnej retardácie(Siegel a spol., 2006; Wong a spol., 2003).
GHB (kyselina γ-hydroxymaslová)
V metabolických procesoch v nervovom tkanive vzniká z GABA kyse-lina γ-hydroxymaslová (GHB) (obr. 3.7) (Snead a Gibbon, 2005). Pred-pokladá sa, že aj GHB môže v nervovom systéme plniť úlohu neuro-transmitera, resp. neuromodulátora. GHB účinkuje pravdepodobne nie-len prostredníctvom GABAergických receptorov, ale aj prostredníctvomšpecifického GHB-receptora spojeného s G-proteínom (Snead, 2000).Predpokladá sa, že GHB môže ovplyvňovať aktivitu viacerých neuro-transmiterových systémov (Wong a spol., 2003).Uvažuje sa o účasti GHB na etiopatogenéze epilepsie, porúch spánku
a látkovej závislosti (Wong a spol., 2003).
Obr. 3.7 Schéma biosyntézy GHB (modifikované podľa Snead a Gibbon, 2005).

58 NEUROTRANSMITERY
Glycín
V druhej polovici 60. rokov 20. storočia bolo na základe experimentál-nych poznatkov navrhnuté zaradenie glycínu medzi neurotransmitery(Siegel a spol., 2006).Glycín je štruktúrne najjednoduchšou aminokyselinou. Je zapojený
v mnohých metabolických dráhach, je esenciálnou súčasťou proteínov.Glycín sa preto nachádza vo všetkých oblastiach mozgu (Webster,2001).
Obr. 3.8 Schéma biosyntézy glycínu (modifikované podľa Purves a spol., 2001).
Metabolizmus glycínu v nervovom tkanive nie je presne charakte-rizovaný. Nie je jasné, či je glycín syntetizovaný de novo, alebo je vy-chytávaný neurónmi (obr. 3.8) (Webster, 2001). Glycín je umiestnenýv sekrečných vezikulách. Uvoľnený glycín sa viaže na ionotropný glycí-nový receptor (Lunch, 2004). Obsadenie glycínergických receptorov gly-cínom zvyšuje prechod Cl−-iónov do bunky, čo vedie k hyperpolarizáciipostsynaptickej membrány. Okrem glycínergických receptorov sa glycínviaže aj na NMDA-receptory, kde pôsobí ako ko-agonista glutamátu(sám glycín bez glutamátu NMDA-receptory neaktivuje). Kým väzbaglycínu na glycínergický receptor vyvoláva hyperpolarizáciu, jeho väzbana NMDA-receptor vedie k opačnému účinku, umožňuje excitáciu (Din-gledin a spol., 1999). Podobne ako pri predchádzajúcich neurotransmi-teroch aj účinok glycínu sa ukončuje jednak spätným vychytávanímdo neurónov, ako aj vychytávaním okolitými gliovými bunkami (Gadea

3 Malomolekulové neurotransmitery 59
Obr. 3.9 Schematické znázornenie lokalizácie glycínergických neurónovv mozgu (A) a v mieche (B).
a Lopez-Colome, 2001c; Purves a spol., 2001; Aragon a Lopez-Corcuera,2003).Glycín pôsobí ako inhibičný neurotransmiter interneurónov v moz-
govom kmeni (most a predĺžená miecha) a mieche (obr. 3.9) (prehľaddistribúcie glycínergických neurónov a ich axónov v mozgu potkana po-zri Rampon a spol., 1996). Aj keď sa uvádza, že neuróny využívajúceglycín ako neurotransmiter sú lokalizované vo vymedzených oblastiachcentrálneho nervového systému, novšie poznatky poukazujú na to, že

60 NEUROTRANSMITERY
synapsy glycínergických neurónov sa môžu nachádzať v mnohých ob-lastiach CNS (Legendre, 2001).Podobne ako pri GABA aj glycín môže v nezrelom CNS vyvolávať
excitáciu namiesto inhibície. Predpokladá sa, že niektoré inhibičné sy-napsy môžu uvoľňovať ako neurotransmitery spoločne glycín aj GABA(Legendre, 2001).Glycín je hlavným inhibičným neurotransmiterom v mieche. V mie-
che a mozgovom kmeni glycínergické interneuróny zabezpečujú inhi-bičnú spätnú väzbu, ktorá moduluje činnosť motoneurónov a koordinujeaktivitu spinálnych reflexov. Príkladom sú Renshawove neuróny, ktorésa významne zúčastňujú na regulácii spinálnych reflexov prostredníc-tvom recipročnej a rekurentnej inhibície činnosti α-motoneurónov pred-ných rohov miechy (Krnjevic, 1977; Webster, 2001). Glycín je aj neu-rotransmiterom sluchových dráh a niektorých neurónov sietnice. Glycí-nergický synaptický prenos sa preukázal aj v Golgiho bunkách mozočka(Aragon a Lopez-Corcuera, 2003).
Acetylcholín a biogénne amíny
Neuróny uvoľňujúce acetylcholín, noradrenalín, dopamín, adrenalín, sé-rotonín, alebo histamín majú určité spoločné črty. Tvoria zoskupe-nia buniek (nemusí však ísť vždy o kompaktné jadrá), ktoré sa na-chádzajú hlavne v mozgovom kmeni a medzimozgu. Tieto neuróny, ajkeď nie veľmi početné, inervujú prostredníctvom dlhých axónov, ktorésa bohato vetvia, rozsiahle podkôrové a hlavne kôrové oblasti mozgu(obr. 3.10).Neuróny uvoľňujúce ako neurotransmitery acetylcholín a biogénne
amíny majú v CNS prevažne modulačné funkcie (Gu, 2002). Pôso-bia hlavne prostredníctvom metabotropných receptorov, čím vyvolá-vajú dlhšie trvajúce zmeny priepustnosti postsynaptickej membrány,pričom tieto zmeny môžu pretrvávať niekoľkonásobne dlhšie ako zmenypozorované po väzbe neurotransmiterov na ionotropné receptory.Modulačné neuróny nie sú väčšinou zodpovedné za rýchle spraco-
vanie informácií, ale skôr modulujú činnosť nervového systému (hlavnemozgovej kôry). Táto modulácia potom v konečnom dôsledku zodpo-vedá za výsledný stupeň reaktivity nervového systému. Modulačné neu-róny sú zapojené (často nepriamo) do procesov spracovania informácií.Majú veľký význam napríklad v regulácii bdelosti alebo v emočných

3 Malomolekulové neurotransmitery 61
Obr. 3.10 Schéma zapojenia modulačných neurotransmiterov v regulácii čin-nosti nervového systému (modifikované podľa Gu, 2002).
reakciách. Pre tieto ich vlastnosti sú modulačné neuróny častým cieľomfarmakologickej intervencie u pacientov s psychiatrickými ochoreniami(Gu, 2002).Uvoľňovanie biogénnych amínov (ale aj niektorých iných neuro-
transmiterov) v mozgu má určitú podobnosť s uvoľňovaním neurotrans-miterov v autonómnom nervovom systéme. Axóny autonómnych nervovvytvárajú vo svojom priebehu v inervovanom tkanive sieť varikozít,tzv. boutons en passant, z ktorých sa neurotransmiter (noradrenalín,acetylcholín) uvoľňuje do okolia. Nie je však vytvorená štruktúra kla-sickej synapsy s presynaptickou a postsynaptickou membránou. Pretouvoľnený neurotransmiter difunduje do okolia a môže aktivovať väčšípočet receptorov umiestnených na bunkách inervovaného tkaniva. Ajv CNS vytvárajú axóny (prevažne amínergických neurónov) štruktúrypodobné boutons en passant. Predpokladá sa, že neurotransmiter uvoľ-nený z takýchto štruktúr môže ovplyvniť aktivitu väčšieho množstvaneurónov v ich okolí (obr. 3.11) (Vizi, 2000; Vizi a spol., 2004).
Acetylcholín
Acetylcholín bol prvým experimentálne identifikovaným neurotransmi-terom. Roku 1921 Otto Loewi ukázal, že elektrická stimulácia nervusvagus spôsobuje zmenu v činnosti srdca prostredníctvom uvoľnenia che-

62 NEUROTRANSMITERY
Obr. 3.11 Modulačné neurotransmitery sa môžu uvoľňovať z tzv. „variko-
zítÿ axónov (synapsy typu en passant). MA–monoamín; AR– autoreceptor;R– receptor (modifikované podľa Vizi, 2000).
mickej látky (acetylcholínu) z nervových zakončení nervus vagus (Zig-mond, 1999; Siegel a spol., 2006).Acetylcholín sa syntetizuje v nervovom zakončení z acetylkoenzý-
mu A a cholínu enzýmom cholínacetyltransferázou a je umiestnený vovezikulách (obr. 3.12). Enzým cholínacetyltransferáza sa vyskytuje špe-cificky v synapsách cholínergických neurónov. Jeho prítomnosť je pretoindikátorom cholínergických nervových zakončení. Cholín sa transpor-tuje z periférie krvným riečiskom do CNS, kde slúži na biosyntézu ace-tylcholínu, ako aj membránových fosfolipidov (Löffelholz, 1998; Web-ster, 2001).

3 Malomolekulové neurotransmitery 63
Obr. 3.12 Schéma syntézy a metabolizmu acetylcholínu (modifikované podľaPurves a spol., 2001).
Po uvoľnení do synaptickej štrbiny sa acetylcholín môže viazať naionotropné (nikotínové), alebo metabotropné (muskarínové) receptoryna postsynaptickej membráne. Okrem toho sa môže viazať aj na presy-napticky lokalizované autoreceptory (nikotínové, muskarínové), ktorýchaktivácia znižuje uvoľňovanie acetylcholínu z presynaptického nervo-vého zakončenia (tab. 3.4) (Caulfield a Birdsall, 1998; Lukas a spol.,1999; Paterson a Nordberg, 2000; Volpicelli a Levey, 2004; Sarter a Pa-rikh, 2005). Kým účinok väčšiny malomolekulových neurotransmite-rov sa ukončuje mechanizmom ich spätného vychytania do zakončeniapresynaptického neurónu, účinok acetylcholínu sa ukončuje jeho hyd-rolýzou prostredníctvom synapticky lokalizovaného enzýmu acetylcho-línesterázy (Purves a spol., 2001). Cholín sa spätne vychytáva do pre-synaptického zakončenia prostredníctvom vysokoafinitného cholínovéhotransportéra, pričom sa môže využiť na resyntézu acetylcholínu (Okudaa Haga, 2003).

64 NEUROTRANSMITERY
Ionotropné nACh ↑gNa+
„NepárneÿM1
↑IP3 ↑DAG
podtypyM3 Gq
↑GC ↑cGMP↓gK+
MetabotropnéM5
„Párneÿ M2Gi/Go ↓AC ↓cAMP ↓[Ca2+ ]i ↑gK+
podtypy M4
Tabuľka 3.4 Klasifikácia receptorov pre acetylcholín a mechanizmy transdukciesignálu.
Približne 10% všetkých neurónov uvoľňuje acetylcholín ako svoj pri-márny neurotransmiter. Acetylcholín sa uvoľňuje takmer na všetkýchúrovniach CNS. Okrem toho sa acetylcholín ako neurotransmiter uvoľ-ňuje z:
• α-motoneurónov v neuromuskulárnych spojeniach,
• pregangliových neurónov autonómneho nervového systému,
• postgangliových neurónov parasympatika (ako aj niektorýchpostgangliových neurónov sympatika).
Acetylcholín je preto neurotransmiterom všetkých neurónov, ktorévysielajú eferentné axóny z CNS na perifériu, motorických aj auto-nómnych neurónov (Webster, 2001).Neuróny syntetizujúce acetylcholín ako svoj neurotransmiter tvo-
ria v mozgu niekoľko ohraničených zoskupení (obr. 3.13). Cholínergickéneuróny sa nachádzajú v septe a diagonálnej oblasti (CH1-3), nucleusbasalis Meynerti (CH4), nucleus pedunculo-pontinus (CH5) a nucleuslatero-dorsalis (CH6) (Petrovický, 2002). Okrem toho sa cholínergickéneuróny nachádzajú aj v striate (nucleus caudatus, putamen) a moz-govej kôre (Oda a Nakanish, 2000). Medzi neuróny miechy, ktoré syn-tetizujú acetylcholín ako svoj neurotransmiter, patria α-motoneuróny

3 Malomolekulové neurotransmitery 65
Obr. 3.13 Schematické znázornenie lokalizácie hlavných zoskupení cholíner-gických neurónov a cholínergických dráh v mozgu (A) a mieche (B). CH1-3 –
septum a diagonálna oblasť, CH4 – ncl. basalis Meynerti, CH5 – ncl. pedunculo-
pontinus, CH6 – ncl. latero-dorsalis. A – cholínergická inervácia kôrových oblastí;B – cholínergické dráhy v bazálnych gangliách; C – inervácia predného mozgu
vychádzajúca z kmeňových cholínergických neurónov; D– cholínergická inervá-
cia mozočka; E – axóny α-motoneurónov; F – cholínergické pregangliové neurónyautonómneho nervového systému; G– cholínergické postgangliové neuróny pa-
rasympatika (modifikované podľa Petrovický, 2002).

66 NEUROTRANSMITERY
v predných rohoch miechy a pregangliové neuróny sympatika a para-sympatika lokalizované v bočných rohoch miechy.Acetylcholín uvoľnený z periférnych nervových zakončení pôsobí pre-
važne prostredníctvom nikotínových receptorov, čím vyvoláva rýchlezmeny na postsynaptickom segmente (napr. neuromuskulárne spojenie,autonómne gangliá). Podľa typu efektora možno rozdeliť nikotínové re-ceptory na muskulárny, gangliový a neuronálny podtyp (Buccafusco,2004).Acetylcholín uvoľnený z parasympatikových nervových zakon-
čení pôsobí hlavne prostredníctvom muskarínových receptorov (napr.v srdci, exokrinných žľazách). Periférne sú receptory M2 distribuovanéprevažne v srdci, M3 v žľazách, M4 v tkanive ostrovčekov a acinov pan-kreasu, M3 a M4 v hladkom svalstve (Ganong, 2005). V mozgu sa na-chádza prevaha muskarínových receptorov. M1-receptory sa nachádzajúhlavne v neokortexe, hipokampe a striate. M2-receptory sa nachádzajúv tých oblastiach predného mozgu, v ktorých sa začínajú ascendentnécholínergické dráhy. Kým M1-receptory sú lokalizované postsynapticky,M2-receptory sa často zúčastňujú na presynaptickej autoinhibícii (Web-ster, 2001).Cholínergické neuróny mozgu sú zapojené v regulácii bdenia a spán-
ku, motivácie a odmeňovania, pamäte a učenia. Cholínergické neu-róny miechy, ktoré vysielajú svoje axóny do periférie, riadia činnosťautonómneho nervového systému, ako aj činnosť kostrového svalstva(Webster, 2001). Na úrovni miechy a mozgu sa zúčastňujú muskarínovéreceptory (prevažne podtypy M2 a M4) na modulácii prenosu nocicep-tívnych informácií (Wess a spol., 2003).Komplexné narušenie cholínergického prenosu sa predpokladá u pa-
cientov s Alzheimerovou chorobou. Cholínergický prenos je narušený ajpri ochorení myastenia gravis a Lambertov-Eatonov myastenický syn-dróm. Motorická porucha pri myastenia gravis je podmienená tvorbouautoprotilátok proti nikotínovým receptorom nervovosvalovej platničky.Tieto protilátky obmedzujú prenos signálov zo synaptických zakon-čení α-motoneurónov v nervovosvalovej platničke, ktorý sprostredkúvaacetylcholín. Pri Lambertovom-Eatonovom myastenickom syndróme satvoria autoprotilátky proti podtypu Ca2+-kanálov lokalizovaných napresynaptickom nervovom zakončení v nervovosvalovej platničke. Tietoprotilátky preto zabraňujú vzniku depolarizácie presynaptickej mem-brány a následnému uvoľneniu acetylcholínu do synaptickej štrbiny

3 Malomolekulové neurotransmitery 67
(Webster, 2001; Siegel a spol., 2006). Cholínergický prenos je narušenýaj po pôsobení botulotoxínu. Botulotoxín blokuje synaptický prenosna nervovosvalovej platničke, čo vedie k svalovej paralýze. Blokuje ajsynaptický prenos z nervových zakončení v hladkom svalstve, čím spô-sobuje viscerálnu motorickú dysfunkciu (Purves, 2004). Uvedený úči-nok botulotoxínu na nervovosvalový prenos má aj terapeutické využitie(napr. liečba detskej mozgovej obrny) (Morton a spol., 2004).
Katecholamíny
Katecholamíny zahŕňajú tri endogénne syntetizované látky – dopamín,noradrenalín a adrenalín, ktoré slúžia ako hormóny aj ako neurotrans-mitery na periférii a v CNS. Syntetizujú sa z aminokyseliny tyrozín.Predpokladá sa, že tyrozín, z ktorého sa syntetizujú katecholamíny,vzniká premenou fenylalanínu v pečeni a cirkuláciou sa transportuje domozgu. Premena tyrozínu na DOPA, ktorú katalyzuje enzým tyrozín-hydroxyláza, je rýchlosť určujúcim krokom v biosyntéze katecholamínov(obr. 3.14, 3.15).Katecholamíny sú zapojené v regulácii veľkého množstva funkcií
CNS (Ramachandran, 2002). Označujú sa aj ako „The Nobel Chemi-calsÿ, pretože objavy spojené s dopamínom, noradrenalínom a adrena-línom boli odmenené niekoľkými Nobelovými cenami (Goldstein, 2001).
Dopamín
Roku 1951 sa dokázal výskyt dopamínu v tkanivách cicavcov (Goodall,1951). Predpokladalo sa, že dopamín prítomný v CNS je iba prekurzo-rom v biosyntéze noradrenalínu. Roku 1958 však Carlsson a spol. zis-tili, že v mozgu sa nachádza dopamín v oveľa vyššej koncentrácii, akoby sa očakávalo, ak by bol iba prekurzorom noradrenalínu. Prelom vovýskume dopamínergických neurónov (a katecholamínergických a séro-tonínergických neurónov) nastal roku 1962, keď Falck a Hillarp vyvinulihistofluorescenčnú metódu na detekciu katecholamínov a sérotonínuv nervovom tkanive (Falck a spol., 1962; Dahlstrom a Fuxe, 1999). Ná-sledne bola opísaná distribúcia dopamínu v mozgu mnohých druhov ži-vočíchov. Vysoká koncentrácia dopamínu v oblasti bazálnych ganglií po-ukazovala na to, že dopamín je ako neurotransmiter významne zapojenýv regulácii extrapyramídových motorických funkcií (Roe, 1999; 2001b).

68 NEUROTRANSMITERY
Obr. 3.14 Schéma biosyntézy katecholamínov.
Na základe výsledkov dlhoročného štúdia úlohy dopamínu v extrapyra-mídových funkciách a v etiopatogenéze neuropsychiatrických ochoreníbola roku 2000 udelená A.Carlssonovi Nobelova cena (Carlsson, 2001a).Dopamín vzniká z aminokyseliny tyrozínu. Biosyntéza dopamínu,
premena tyrozínu na DOPA, katalyzovaná enzýmom tyrozínhydroxylá-zou a premena DOPA na dopamín, katalyzovaná enzýmom dekarboxy-láxou L-aromatických aminokyselín, prebieha v cytoplazme (obr. 3.14,3.15). Syntetizovaný dopamín sa následne transportuje vezikulárnymmonoamínergickým transportérom do sekrečných vezikúl. Po depolari-zácii nervového zakončenia sa dopamín z vezikúl uvoľňuje do synaptickejštrbiny. Pôsobí na postsynaptické aj presynaptické dopamínergické me-tabotropné receptory, ktoré sa rozdeľujú do dvoch hlavných skupín,„D1-likeÿ a „D2-likeÿ (tab. 3.5). Účinok dopamínu sa ukončuje jeho

3 Malomolekulové neurotransmitery 69
Obr. 3.15 Schematické znázornenie noradrenalínergického nervového zakonče-nia. Enzýmy biosyntézy noradrenalínu, TH (tyrozínhydroxyláza), AADC (de-
karboxyláza aromatických aminokyselín) a DBH (dopamín-β-hydroxyláza) sa
syntetizujú v tele neurónu a axónový transport ich prenáša do oblasti nervo-vého zakončenia. Tieto enzýmy v nervovom zakončení syntetizujú z tyrozínu ne-
urotransmiter noradrenalín (NA), ktorý je uskladnený v sekrečných vezikulách.
Po depolarizácii presynaptickej oblasti vplyvom akčného potenciálu dochádzak vzostupu intracelulárnej koncentrácie Ca2+. Zvýšená koncentrácia vápnika
v presynaptickej oblasti vyvolá kaskádu dejov, ktorých výsledkom je fúzia ve-zikúl obsahujúcich neurotransmiter s presynaptickou membránou neurónu, čím
sa noradrenalín uvoľní do synaptickej štrbiny. Neurotransmiter pôsobí na post-
synapticky lokalizované receptory, čím vyvoláva zmeny v postsynaptickej ob-lasti neurónu. Zároveň môže pôsobiť aj na presynapticky lokalizované autore-
ceptory. Pôsobenie neurotransmitera je ukončené jeho spätným vychytaním do
presynaptického nervového zakončenia noradrenalínovým transportérom (NET;uptake-1) alebo do postsynaptického priestoru (uptake-2). Noradrenalín spätne
vychytaný do nervového zakončenia je buď metabolizovaný, alebo prostredníc-tvom vezikulárneho monoamínového transportéra (VMAT) prenesený do vezi-
kúl, čo umožňuje jeho ďalšie použitie.
enzymatickou premenou prostredníctvom extracelulárne lokalizovanéhoenzýmu katechol-O-metyltransferázy (COMT), ako aj spätným vychy-taním prostredníctvom dopamínového transportéra, ktorý premiestňuje

70 NEUROTRANSMITERY
dopamín späť do presynaptického nervového zakončenia. Spätne vychy-taný dopamín sa metabolizuje enzýmom monoaminooxidázou (MAO),ktorý sa nachádza na membránach mitochondrií (obr. 3.16) (Webster,2001).
„D1-like“D1A
Gs ↑AC ↑cAMP ↑[Ca2+ ]iD5
MetabotropnéD2D2S
↑gK+ ↓[Ca2+ ]iD2L
„D2-like“
D3
Gi/o ↓AC ↓cAMP
↓[Ca2+ ]iD4
Tabuľka 3.5 Klasifikácia receptorov pre dopamín a mechanizmy transdukcie
signálu. Predpokladá sa, že súčasná aktivácia receptorov D1 a D2 môže viesť
k aktivácii PLC a následnému vzostupu intracelulárnej koncentrácie Ca2+ (Leea spol., 2004).
Aj keď len približne jeden z každého milióna neurónov syntetizujedopamín ako svoj neurotransmiter, dopamín sa uvoľňuje takmer navšetkých úrovniach CNS. Dopamín slúži ako neurotransmiter vo viace-rých významných dráhach CNS. Na periférii moduluje dopamín hlavnečinnosť obličiek (Siegel a spol., 2006).Dopamínergické neuróny sa nachádzajú v oblastiach od pons až po
bulbus olfactorius (A8-16). Dopamínergické neuróny v strednom mozgutvoria tri zoskupenia (A8, A9 a A10) s neostrým ohraničením. Dopamí-nergické neuróny A9 sú lokalizované v oblasti substantia nigra, prevažnev časti pars compacta. Väčšina buniek skupiny A10 sa premieta do ven-trálnej tegmentálnej oblasti. Neuróny skupín A11-A15 sú lokalizovanév medzimozgu. A12 neuróny sa nachádzajú v nucleus arcuatus. V bul-bus olfactorius sa nachádzajú dopamínergické neuróny označované akoA16. Aj niektoré bunky sietnice využívajú dopamín ako neurotransmi-ter (A17). Dopamínergické neuróny vytvárajú niekoľko hlavných dopa-

3 Malomolekulové neurotransmitery 71
Obr. 3.16 Schéma metabolizmu dopamínu. COMT–katechol-O-metyltransfe-ráza; DOPAC– kyselina dihydroxyfenyloctová; MAO–monoaminooxidáza (mo-
difikované podľa Purves a spol., 2001; Webster, 2001).
mínergických dráh, ktoré možno na základe dĺžky axónov dopamíner-gických neurónov rozdeliť na ultrakrátke, stredné (intermediálne) a dlhé(obr. 3.17) (Maršala, 1985).Ultrakrátke dopamínergické dráhy: nachádzajú sa v sietnici medzi
vnútornou jadrovou a vnútornou plexiformnou vrstvou (A17) a v bulbusolfactorius (A16; periglomerulárne bunky) (Ganong, 2005).
Intermediárne dopamínergické dráhy:
• tuberoinfundibulárna dráha, ktorú tvoria axóny neurónov loka-lizovaných v nucleus arcuatus a nucleus periventricularis, ktoréinervujú oblasť eminentia mediana; dopamín uvoľnený z nervo-vých zakončení do portálnej cirkulácie hypotalomo-hypofýzovéhosystému pôsobí v adenohypofýze ako prolaktostatín (obr. 3.17D);
• incertohypotalamický systém: spája hypotalamus a laterálne sep-tálne jadrá,
• medulárna periventrikulárna skupina: bunky roztrúsených v o-kolí tretej a štvrtej komory (Ganong, 2005).

72 NEUROTRANSMITERY
Obr. 3.17 Schematické znázornenie lokalizácie dopamínergických neurónov
a dopamínergických dráh v mozgu. A8 – ncl. reticularis pontis oralis,A9 – substantia nigra, pars compacta, A10 – area ventralis tegmenti (Tsai),
A11 – ncl. premamillaris dorsalis, A12 – ncl. infundibularis (arcuatus), A13 – zona
incerta, A14 – area hypothalamica anterior, A15 – area preoptica, A16 – bulbusolfactorius; (A) – nigrostriatálna dráha; (B) –mezokortikálna dráha; (C) –mezo-
limbická dráha; (D) – tuberoinfundibulárna dráha; (E) – spinálne dráhy (modi-fikované podľa Petrovický, 2002).
Dopamínové systémy s dlhými axónmi:
• nigrostriatálna dráha, ktorú tvoria axóny neurónov substantianigra, pars compacta, sa zúčastňuje na regulácii motoriky pro-stredníctvom modulácie činnosti bazálnych ganglií (obr. 3.17A);
• mezokortikálna dráha, ktorú vytvárajú axóny neurónov area ven-tralis tegmenti, difúzne inervuje oblasti predného mozgu (napr.prefrontálny, cingulárny a peririnálny kortex) a zúčastňuje sa namodulácii kognitívnych procesov (obr. 3.17B);
• mezolimbická dráha, ktorú tvoria axóny neurónov area ventra-lis tegmenti, inervuje amygdalu, hipokampus, septum, nucleus

3 Malomolekulové neurotransmitery 73
accumbens a tuberculum olfactorium; zúčastňuje sa na reguláciiviacerých behaviorálnych funkcií, ako napríklad vnímanie príjem-ných pocitov a eufórie po aplikácii návykových látok, ako aj nadelúziách a halucináciách počas psychózy (obr. 3.17C);
• spinálna dráha, ktorá vychádza prevažne z A11-neurónov, iner-vuje hlavne zadné rohy miechy a zúčastňuje sa prevažnena descendentnej modulácii prenosu nociceptívnych informácií(obr. 3.17E) (Moore, 1987; Ganong, 2005; Chinta a Andersen,2005; Siegel a spol., 2006).
Dopamínergické neuróny sa zúčastňujú na procesoch motiváciea odmeňovania, konsolidácie pamäte a učenia, na regulácii sekréciehypotalamo-hypofýzového systému (napr. dopamín, ako prolaktosta-tín, inhibuje vyplavovanie prolaktínu), na regulácii motorických funk-cií a procesoch prenosu a spracovania nociceptívnych signálov (Wise,2004).Patologické narušenie dopamínergických buniek substantia nigra
a nedostatok dopamínu v oblasti bazálnych ganglií sa preukazuje u pa-cientov s Parkinsonovou chorobou. Predpokladá sa účasť dysfunkciedopamínergického systému v etiopatogenéze Huntingtonovej choroby,schizofrénie, depresie, Tourettovho syndrómu, látkovej závislosti, po-rúch príjmu potravy a porúch pozornosti (Nieoullon, 2002; Bozzia Borrelli, 2006).
Noradrenalín
Noradrenalín bol identifikovaný roku 1946 ako neurotransmiter v sym-patikovom nervovom systéme (von Euler, 1946). O dva roky neskôrAhlquist uviedol koncept α- a β-adrenergických receptorov (Ahlquist,1948). Roku 1954 Vogt rozšírila koncept noradrenalínu ako neurotrans-mitera dôkazom jeho prítomnosti v CNS (Ramachandran, 2002). Za ob-jasnenie procesov biosyntézy noradrenalínu získal Julius Axelrod roku1970 Nobelovu cenu (Axelrod, 1971).Noradrenalín sa syntetizuje z dopamínu nachádzajúceho sa v cy-
toplazme enzýmom dopamín-β-hydroxylázou, ktorý je lokalizovaný namembráne sekrečných vezikúl (obr. 3.15). Z vezikúl uvoľnený noradre-nalín pôsobí v synaptickom priestore na postsynaptické a presynap-tické adrenergické receptory, ktoré sa rozdeľujú do dvoch hlavných

74 NEUROTRANSMITERY
skupín (α a β), pričom sa ďalej delia na viacero podtypov (tab. 3.6)(Hein a Kobilka, 1995; Civantos Calzada a spol., 2001; Kirstein a In-sel, 2004). Účinok noradrenalínu je ukončený jeho enzymatickou pre-menou prostredníctvom extracelulárne lokalizovaného enzýmu COMT,ako aj spätným vychytaním prostredníctvom noradrenalínového trans-portéra, ktorý premiestňuje noradrenalín späť do presynaptického ner-vového zakončenia. Spätne vychytaný noradrenalín sa metabolizuje en-zýmom MAO (obr. 3.18) (Webster, 2001).
α1A
α1 α1B Gq/11↑PLC ↑DAG
↑[Ca2+ ]i
α1D↑IP3
α2A
Metabotropné α2 α2B Gi/Go ↓AC ↓cAMP ↑gK+ ↓[Ca2+]i
α2C
β1
β2 Gs ↑AC ↑cAMP ↑[Ca2+ ]i ↓gK+
β3
Tabuľka 3.6 Klasifikácia receptorov pre noradrenalín a adrenalín a mechanizmytransdukcie signálu.
Noradrenergické neuróny sa nachádzajú v predĺženej mieche a mos-te. Neuróny skupiny A1 sú lokalizované v dolnej časti predĺženej miechyv okolí nucleus funiculi lateralis a šíria sa dorzomediálnym smerom dolaterálnej časti retikulárnej formácie. Bunky A2 ležia v oblasti kaudál-nej časti nucleus tractus solitarii. Skupina A3 sa dokázala u potkanov,jej výskyt v mozgu primátov sa nepotvrdil. Neuróny A4 tvoria sube-pendýmový pruh, ktorý sa rozprestiera súbežne s vláknami pedunculuscerebellaris superior a pri prednom okraji sa spája so skupinou A6. Väč-šina noradrenalínu v mozgu sa syntetizuje v bunkách locus coeruleus(A6). Neuróny A5 sú roztrúsené v okolí nucleus nervi facialis a v okolí

3 Malomolekulové neurotransmitery 75
Obr. 3.18 Schéma metabolizmu noradrenalínu. COMT–katechol-O-metyl-
transferáza; MAO–monoaminooxidáza (modifikované podľa Purves a spol.,2001; Webster, 2001).
horného olivarného komplexu. Skupina neurónov A7 sa nachádza v ros-trálnej časti retikulárnej formácie v moste (Maršala, 1985). Noradre-nalín sa uvoľňuje takmer na všetkých úrovniach CNS a v mieche. Naperiférii sa noradrenalín syntetizuje v dreni nadobličiek a sympatiko-vých nervových zakončeniach (Siegel a spol., 2006).V mozgu možno rozlíšiť dve hlavné dráhy, ktoré vytvárajú axóny
noradrenalínergických neurónov, ventrálny a dorzálny noradrenergickýzväzok (obr. 3.19) („ventral and dorsal noradrenergic bundlesÿ):
• Axóny vytvárajúce dorzálny zväzok vychádzajú prevažne z neu-rónov locus coeruleus. Nervové dráhy dorzálneho zväzku v dience-fale vytvárajú tzv. „medial forebrain bundleÿ, ktorý sa zúčastňujena inervácii hypotalamu, hipokampu a telencefala.
• Axóny ventrálne lokalizovaných noradrenalínergických neurónovvytvárajú ventrálny zväzok, ktorý v strednom mozgu vytváracentrálny tegmentový trakt („central tegmental tractÿ), ktorý

76 NEUROTRANSMITERY
Obr. 3.19 Schematické znázornenie lokalizácie noradrenalínergických neurónova noradrenalínergických dráh v mozgu. A1 – ncl. reticularis lateralis et parvo-
cellularis; A2– ncl. tractus solitarii; A4 – substantia grisea centralis, pars cauda-
lis; A5 – ncl. reticularis parvocellularis, pars ventralis; A6 – locus coeruleus; A7 –ncl. reticularis parvocellularis; (A) – dorsal noradrenergic bundle; (B) – ventral
noradrenergic bundle; (C) –medial forebrain bundle; (D) – central tegmentaltract; (E) – spinálne dráhy (modifikované podľa Petrovický, 2002).
inervuje hypotalamus. Niektoré axóny prebiehajúce v centrálnomtegmentálnom trakte prechádzajú do „medial forebrain bundleÿ.
• Neuróny locus coeruleus a iných noradrenalínergických skupín(napr. A5 a A7) inervujú aj miechu a mozoček (Siegel a spol.,2006).
Noradrenalínergické neuróny sa zúčastňujú na regulácii procesovbdenia a spánku, pamäte a učenia, motorických funkcií, stresovejreakcie, bolesti a regulácii sekrécie hypotalamo-hypofýzového systému(Goldstein, 2001; Mravec a Kiss, 2004). Najvýznamnejšou oblasťoumodulujúcou uvedené procesy je locus coeruleus (podrobnejšie pozriAston-Jones a Cohen, 2005).

3 Malomolekulové neurotransmitery 77
Predpokladá sa, že dysregulácia činnosti neurónov locus coeruleussa môže zúčastňovať v etiopatogenéze ochorení CNS, medzi ktoré patriaporuchy pozornosti, spánku a bdelosti a afektívne poruchy (Berridgea Waterhouse, 2003).
Adrenalín
Roku 1895 Oliver a Schafer opísali pôsobenie „aktívnej zložkyÿ drenenadobličky (Garcia-Sainz, 1995). O dva roky neskôr Abel a Crawfordizolovali adrenalín a zaviedli pojem epinefrín. V rokoch 1901–1902 Ta-kamine a Aldrich izolovali adrenalín aj v kryštalickej forme a následnebola opísaná jeho štruktúra a zavedený pojem adrenalín (Davenport,1982; Parascandola a spol., 1987).Adrenalín sa syntetizuje z noradrenalínu. Noradrenalín sa transpor-
tuje z vezikúl do cytoplazmy a enzým fenyletanolamín-N-metyltransfe-ráza ho premieňa na adrenalín (obr. 3.14). Následne je adrenalín umiest-nený do sekrečných vezikúl. Predpokladá sa, že uvoľnenie, pôsobenie nareceptory, ako aj ukončenie účinku adrenalínu prebieha podobne ako prinoradrenalíne (obr. 3.20) (Flatmark, 2000).
Obr. 3.20 Schéma metabolizmu adrenalínu. COMT– katechol-O-metyltrans-feráza (modifikované podľa Goldstein, 2001).
Hlavným zdrojom adrenalínu v organizme sú chrómafinné bunkydrene nadobličiek. Adrenalín uvoľnený z nadobličiek do cirkulácie sti-muluje adrenergické receptory buniek jednotlivých orgánov (tab. 3.6)(Siegel a spol., 2006).

78 NEUROTRANSMITERY
Okrem drene nadobličiek sa adrenalín syntetizuje aj bunkami CNS.Neuróny, ktoré syntetizujú adrenalín, sú lokalizované v dolnej časti moz-gového kmeňa (obr. 3.21). Väčšina adrenalínergických dráh sa pravdepo-dobne tvorí axónmi C1-neurónov. Ascendentné adrenalínergické dráhyvychádzajúce z neurónov C1, C2 a C3 inervujú prevažne locus coeru-leus, periakvaduktálnu šedú hmotu a hypotalamus. Tieto dráhy pre-biehajú prevažne vo ventrálnom noradrenalínergickom zväzku. Descen-dentné adrenalínergické dráhy sa pravdepodobne zúčastňujú na pria-mej inervácii sympatikových pregangliových neurónov, čo vytvára ana-tomický podklad pre možnú reguláciu aktivity sympatikoadrenálnehosystému adrenalínergickými neurónmi mozgového kmeňa (Goldstein,2001).
Obr. 3.21 Schematické znázornenie lokalizácie noradrenalínergických neu-rónov a noradrenalínergických dráh v mozgu. C1 – ncl. reticularis lateralis
et parvocellularis; C2 – ncl. tractus solitarii; C3 – ncl. prepositus hypoglossi.A – ascendentné adrenalínergické dráhy; B– spinálne dráhy (modifikované podľa
Petrovický, 2002).
Adrenalín sa v CNS vyskytuje iba v nízkych koncentráciách. Je topodmienené tým, že adrenalínergických neurónov je v mozgu relatívnemálo len niekoľko tisíc (Gai a spol., 1993). Ascendentná adrenalínergická

3 Malomolekulové neurotransmitery 79
inervácia hypotalamu sa zúčastňuje na prenose informácií o pôsobenístresorov do hypotalamu, čím sa významne zapája do neuroendokrinnejstresovej odpovede (Goldstein, 2001).Predpokladá sa, že adrenalínergická inervácia sympatikových pre-
gangliových neurónov vychádzajúca z C1-neurónov sa zúčastňuje namodulácii činnosti sympatikoadrenálneho systému hlavne počas záťa-žových situácií (Schreihofer a spol., 2000).
Sérotonín
Roku 1948 Rapport a spol. izolovali z krvných doštičiek látku, ktoráspôsobovala vazokonstrikciu. Na základe tejto vlastnosti bola označenáako sérotonín (Rapport a spol., 1948). Roku 1953 Twarog a Page do-kázali výskyt sérotonínu aj v CNS (Twarog a Page, 1953).Sérotonín (5-hydroxytryptamín, 5-HT) sa syntetizuje z aminoky-
seliny tryptofán nachádzajúcej sa v potrave. V prvom kroku bio-syntézy vzniká z tryptofánu pôsobením enzýmu tryptofánhydroxylázy5-hydroxytryptofán. Účinkom enzýmu dekarboxylázy L-aromatickýchaminokyselín z neho vzniká sérotonín, ktorý je umiestnený v sekrečnýchvezikulách (obr. 3.22). Uvoľnený sérotonín pôsobí na postsynaptické ajpresynaptické sérotonínergické receptory. Jeho účinok sa ukončuje čin-nosťou sérotonínového transportéra, ktorý sérotonín premiestňuje späťdo presynaptického nervového zakončenia. Sérotonín sa metabolizujeenzýmom MAO. Enzým COMT sa nezúčastňuje na metabolizme séro-tonínu (Webster, 2001).Periférne sa sérotonín nachádza okrem neurónov a enterochróma-
finných buniek GIT aj v trombocytoch a žírnych bunkách (Kim a Ca-milleri, 2000). Periférne uvoľnený sérotonín však neprechádza hema-toencefalickou bariérou (podobne ako neprechádzajú ani iné biogénneamíny pôsobiace ako neurotransmitery). Sérotonín sa uvoľňuje takmerna všetkých úrovniach CNS. V mozgu sa nachádza približne 1 milión sé-rotonínergických neurónov. Každý sérotonínergický neurón ovplyvňujepribližne 500 000 neurónov. Sérotonín pôsobí prostredníctvom ionotrop-ných a metabotropných receptorov (tab. 3.7) (Barnes a Sharp, 1999).Receptory pre sérotonín majú najväčšie množstvo podtypov spomedzivšetkých receptorov pre monoamíny (Hannon a Hoyer, 2002).Sérotonínergické neuróny sa nachádzajú v ohraničených oblastiach
mozgového kmeňa (obr. 3.23). Väčšina neurónov syntetizujúcich séroto-

80 NEUROTRANSMITERY
Obr. 3.22 Schéma syntézy a metabolizmu sérotonínu. MAO-A–monoamino-oxidáza A (modifikované podľa Purves a spol., 2001).
nín sa nachádza v mediálnej a paramediálnej pozdĺžnej zóne. Séroto-nínergické neuróny sú lokalizované v relatívne ohraničených oblastiach,v nuclei raphes (Maršala, 1985; Siegel a spol., 2006).Axóny sérotonínergických neurónov vytvárajú ascendentné aj des-
cendentné dráhy: ventrálna ascendentná dráha, dorzálna ascendentná

3 Malomolekulové neurotransmitery 81
5-HT3A
Ionotropné 5-HT3 5-HT3B ↑gNa+ ↑gK+
5-HT3C
5-HT1A ↑gK+
5-HT1B
5-HT1 5-HT1D Gi/o ↓cAMP
5-HT1E
5-HT1F
5-HT2A
Metabotropné 5-HT2 5-HT2B Gq/11 ↑PLC ↓gK+ ↑[Ca2+ ]i
5-HT2C
5-HT4 Gs ↑cAMP
5-HT5A Gi/o ? ↓cAMP
5-HT55-HT5B ?
5-HT6 Gs ↑cAMP
5-HT7 Gs ↑cAMP
Tabuľka 3.7 Klasifikácia receptorov pre sérotonín a mechanizmy transdukciesignálu.
dráha, cerebelárna dráha, descendentná propriobulbárna dráha, bulbo-spinálne dráhy (Maršala, 1985).Sérotonínergické neuróny sa zúčastňujú na regulácii procesov bde-
nia a spánku, cirkadiánnych rytmov, bolesti, príjmu potravy, sexuál-neho správania, neurotrofných procesoch (maturácia a vývoj mozgu– synaptická plasticita, dĺžka dendritov, cytoskeletálna architektúra)(Azmitia, 1999). Činnosť sérotonínergických neurónov modulujú via-

82 NEUROTRANSMITERY
Obr. 3.23 Schematické znázornenie lokalizácie sérotonínergických neu-rónov a sérotonínergických dráh v mozgu. B1 – ncl. raphealis pallidus; B2 – ncl.
raphealis parvus; B3 – ncl. raphealis magnus; B4 – ncl. raphealis pallidus;
B5 – ncl. raphealis pontis; B6 – ncl. raphealis dorsalis; B7 – ncl. raphealis dorsalis;B8 – ncl. linearis caudalis; B9 – ncl. pontis oralis (modifikované podľa Petrovický,
2002).
ceré neuropeptidy (napr. substancia P, CRH) (Valentino a Commons,2005).Patologické narušenie sérotonínergického systému sa predpokladá
napríklad pri depresívnej poruche, anxiete a migréne (Ressler a Ne-meroff, 2000).
Histamín
V 50. rokoch 20. storočia sa začal študovať vplyv antihistamínergickejaktivity neuroleptík a antidepresív na behaviorálne funkcie. V druhejpolovici 70. rokov bol opísaný histamínergický systém mozgu (Schwartz,1975; Schwartz a spol., 1976; Parsons a Ganellin, 2006).Histamín sa syntetizuje z aminokyseliny histidín enzýmom histidín-
dekarboxylázou (obr. 3.24). V súčasnosti sú známe štyri histamínergické

3 Malomolekulové neurotransmitery 83
receptory H1–H4. V mozgu bola dokázaná prítomnosť H1–H3 recepto-rov (tab. 3.8) (Hill a spol., 1997; Parsons a Ganellin, 2006).
Obr. 3.24 Schéma syntézy a metabolizmu histamínu. MAO-B–monoamino-oxidáza B (modifikované podľa Purves a spol., 2001, Webster, 2001).
H1 Gq/11 ↑PLC ↑DAG ↑IP3 ↑[Ca2+]i
H2 Gs ↑AC ↑cAMP ↑PKA
MetabotropnéH3 Gi/Go ↑AC ↑cAMP ↓[Ca2+]i
H4 Gi/Go ↑AC ↑cAMP
Tabuľka 3.8 Klasifikácia receptorov pre histamín a mechanizmy transdukciesignálu (modifikované podľa Haas a Panula, 2003; de Esch a spol., 2005).

84 NEUROTRANSMITERY
Podobne ako iné biogénne amíny aj histamín sa uvoľňuje takmer navšetkých úrovniach CNS. V CNS sa nachádzajú histamínergické neu-róny v ohraničenej oblasti hypotalamu. Na periférii histamín obsahujúžírne bunky a bazofily (Webster, 2001).Histamínergický systém mozgu tvorí skupina neurónov v nucleus tu-
beromamillaris hypotalamu, ktoré vysielajú axóny takmer do všetkýchoblastí CNS. Predpokladá sa, že axóny histamínergických neurónov vy-tvárajú dve ascendentné a najmenej jednu descendentnú dráhu, ktoréinervujú oblasti mozgu a miechy (obr. 3.25) (Siegel a spol., 2006).
Obr. 3.25 Schematické znázornenie lokalizácie histamínergických neurónov
a histamínergických dráh v mozgu. E – ncl. tuberomamillaris; A – ascendentnédráhy inervujúce predný mozog; B – descendentné dráhy inervujúce mozgový
kmeň a mozoček; C – spinálne dráhy (modifikované podľa Petrovický, 2002).
Histamínergický systém má dôležitú úlohu v regulácii cyklov spá-nok – bdenie, energetickej a endokrinnej homeostáze, v regulácii te-lesnej teploty, v modulácii synaptickej plasticity a učenia, v procesochspracovania nociceptívnych signálov, príjme potravy a v regulácii sek-récie hypotalamo-hypofýzového systému (Passani a spol., 2000; Browna spol., 2001; Haas a Panula, 2003; Siegel a spol., 2006). Predpokladá

3 Malomolekulové neurotransmitery 85
Obr. 3.26 Štruktúrne vzorce ATP a adenozínu (Webster, 2001).
sa, že histamín má aj dôležitú úlohu v inhibícii vzniku záchvatovýchstavov vo vyvíjajúcom sa mozgu (Yokoyama, 2001).Uvažuje sa, že histamínergický systém sa môže zúčastňovať v etiopa-
togenéze niektorých ochorení CNS, ako sú napr. Alzheimerova chorobaa schizofrénia (Fernandez-Novoa a Cacabelos, 2001).
Iné malomolekulové neurotransmitery
Okrem excitačných a inhibičných aminokyselín, acetylcholínu a biogén-nych amínov neuróny využívajú na prenos signálov aj iné látky s maloumolekulou. Do tejto skupiny sa zaraďujú aj niektoré chemické látky,ktoré nespĺňajú všetky charakteristiky klasických neurotransmiterov(napr. oxid uhoľnatý, oxid dusnatý, hydrogénsulfát) (Snyder a Ferris,2000; Boehning a Snyder, 2003).
Puríny
Puríny, ATP a adenozín (obr. 3.26) majú centrálnu úlohu v energetic-kom metabolizme organizmov. Pravdepodobne preto sa úloha purínovako neurotransmiterov rozpoznala až neskôr (Siegel a spol., 2006).ATP a produkt jeho metabolizmu, adenozín, sa uvoľňujú niektorými
neurónmi prevažne ako kotransmitery (kotransmiter – látka uvoľňovaná

86 NEUROTRANSMITERY
z neurónu spolu s iným neurotransmiterom). Puríny sa napríklad uvoľ-ňujú z autonómnych neurónov (inervujúcich vas deferens, močový me-chúr, svalstvo srdca), z nervových pletení gastrointestinálneho traktua niektorými neurónmi CNS (Kandel a spol., 2000).Receptory pre ATP sa rozdeľujú na ionotropné (P2X) a metabo-
tropné (P2Y) (Khakh a spol., 2001; North, 2002). Doteraz bolo opísa-ných sedem podtypov P2X a najmenej päť podtypov P2Y receptorov(tab. 3.9) (Ralevic a Burnstock, 1998; Illes a Ribeiro, 2004).Adenozín pôsobí prostredníctvom adenozínových receptorov A1,
A2A, A2B alebo A3, ktoré sa nachádzajú aj v mozgu. Všetky adenozí-nové receptory sú spojené s G-proteínmi (tab. 3.10) (Fredholm a spol.,2001).ATP a adenozín majú pravdepodobne modulačný vplyv na synap-
tický prenos. Adenozín pôsobí vo všeobecnosti ako inhibičný neuromo-dulátor (Dunwiddie a Masino, 2001). ATP je pravdepodobne neuro-transmiterom zodpovedným za prenos signálov z chemických senzoric-kých neurónov informujúcich prevažne o koncentrácii CO2 a pH (Spyera spol., 2004). Predpokladá sa, že adenozín sa zúčastňuje na reguláciiprietoku krvi v mozgu (Phillis, 2004).
Neurosteroidy
Je známe, že steroidové látky syntetizované v endokrinných orgánoch(napr. ováriá, nadobličky) môžu ovplyvňovať činnosť neurónov. V po-sledných desaťročiach sa však ukázalo, že aj priamo v mozgu dochádzak syntéze steroidových látok, ktoré môžu pôsobiť ako neurotransmitery,resp. neuromodulátory. Prekurzorom biosyntézy neurosteroidov je po-dobne ako aj pri iných steroidových látkach cholesterol (obr. 3.27). Ste-roidy syntetizované v nervovom systéme sa označujú ako neurosteroidy(Compagnone a Mellon, 2000).Podľa miesta syntézy sa steroidy rozdeľujú do troch skupín.
• syntetizované periférnymi endokrinnými žľazami:
– kortikosterón
– aldosterón
– estradiol
– testosterón

3 Malomolekulové neurotransmitery 87
P2X1
P2X2
P2X3
Ionotropné P2X4 ↑gNa+ ↑gK+ ↑[Ca2+ ]i
P2X5
P2X6
P2X7
P2Y1 Gq/11 Gi ↑PLC
P2Y2 Gq/11 Gi/o ↑PLC
Metabotropné P2Y4 Gq/11 Gi ↑PLC
P2Y6 Gq/11 ↑PLC
P2Y11 Gs ↑AC↑cAMP
Tabuľka 3.9 Klasifikácia receptorov pre ATP a mechanizmy transdukcie sig-nálu (modifikované podľa Ralevic a Burnstock, 1998).
A1 Gi/o/Gq↓AC ↓cAMP
↑PLC
Metabotropné A2A2A Gs ↑AC ↑cAMP
A2B Gs ↑AC ↑cAMP
A3 Gi/o ↓AC ↓cAMP
Tabuľka 3.10 Klasifikácia receptorov pre adenozín a mechanizmy transdukciesignálu (modifikované podľa Ralevic a Burnstock, 1998).

88 NEUROTRANSMITERY
Obr. 3.27 Schéma biosyntézy neurosteroidov. 3β-HSD=3β – hydroxy-steroiddehydrogenáza; 3α-HSOR=3α – hydroxysteroidoxidoreduktáza; 17β-
HSOR=17β-hydroxysteroidoxidoreduktáza (modifikované podľa Schumachera spol., 2000).
• syntetizované na periférii aj v CNS:
– progesterón
– tetrahydroprogesterón (allopregnenolón)
• syntetizované v CNS:
– dehydroepiandrosterón (Webster, 2001).
Neurosteroidy sa zaraďujú k neuromodulátorom. Neurosteroidy ne-účinkujú primárne prostredníctvom klasických nukleárnych receptorovpre steroidové hormóny, ale prostredníctvom modifikácie činnosti recep-torov pre iné neurotransmitery (tab. 3.11) (Belelli a Lambert, 2005).Experimentálne údaje poukazujú na to, že neurosteroidy majú vý-
znamnú úlohu v modulácii veľkého spektra činností CNS:

3 Malomolekulové neurotransmitery 89
Receptor Steroid
3α,5α-tetrahydroprogesterón
3α,5β-tetrahydroprogesterón
GABAA 3α,5α-tetrahydrodeoxykortikosterón
pregnenolón sulfát
dehydroepiandrosterón sulfát
progesterónnACh
3α,5α-tetrahydroprogesterón
progesterónGlyR
pregnenolón sulfát
estradiol (17α a β)
progesterón
5-HT3 testosterón
3α,5α-tetrahydroprogesterón
pregnenolón sulfát
17β-estradiol
pregnanolón sulfátNMDA
pregnanolón hemisukcinát
pregnenolón sulfát
AMPA pregnenolón sulfát
pregnenolón sulfát
Kainate R 17β-estradiol
progesterón
Oxytocín R progesterón
dehydroepiandrosterón sulfát
σ1 pregnenolón sulfát
progesterón
Tabuľka 3.11 Prehľad receptorov pre neurotransmitery, ktorých činnosť je mo-dulovaná neurosteroidmi (modifikované podľa Rupprecht a Holsboer, 1999).

90 NEUROTRANSMITERY
• modulácia činnosti receptorov pre iné neurotransmitery(tab. 3.11),
• neuroprotekcia, indukcia rastu neuritov, dendritických tŕňov a sy-naptogenéza, zlepšenie pamäte (pregnenolón, dehydroepiandro-sterón), modulácia spánku,
• predpokladá sa účasť neurosteroidov v procesoch únavy/vyčerpa-nia počas tehotnosti, premenštruačnom syndróme, postpartálnejdepresii, epilepsii a depresii,
• hypnotický, anestetický, anxiolytický, antikonvulzívny, antipsy-chotický, antidepresívny efekt (Rupprecht a Holsboer, 1999).
• Uvažuje sa o tom, že neurosteroidové látky môžu tvoriť novúskupinu liečiv ovplyvňujúcich rôzne funkcie nervového systému(Melcangi a Panzica, 2006).
Plynné neurotransmitery
Poznatky získané v poslednom období naznačujú, že molekuly plynovoxidu dusnatého (NO), oxidu uhoľnatého (CO) a hydrogénsulfátu (H2S)by mohli v nervovom systéme plniť funkciu neurotransmiterov, resp.neuromodulátorov (Kimura, 2002). NO, CO ani H2S však nespĺňajúvšetky charakteristiky klasických neurotransmiterov. Tieto neurotrans-mitery možno charakterizovať na základe ich vlastností nasledovne:
• sú to malé molekuly plynov,
• voľne prechádzajú biologickými membránami (z miesta svojejsyntézy difundujú do okolia; môžu sa uvoľňovať z postsynaptic-kého neurónu a pôsobiť v presynaptickom nervovom zakončení,čo je v protiklade k spôsobu, ktorým pôsobia klasické neurotrans-mitery),
• ich účinok nezávisí od membránových receptorov (nevyvolávajúzmeny v aktivite neurónov väzbou na membránové receptory, aleskôr zásahom do intracelulárnych dejov),
• sú endogénne enzymaticky syntetizované a ich produkcia je regu-lovaná,

3 Malomolekulové neurotransmitery 91
• neuróny obsahujúce enzýmy potrebné pre biosyntézy plynnýchneurotransmiterov nie sú distribuované difúzne, ale skôr v defi-novaných oblastiach CNS,
• majú dobre definovateľné špecifické funkcie vo fyziologických kon-centráciách (napr. modulácia synaptického prenosu),
• ich bunkový efekt môže, ale aj nemusí byť sprostredkovaný pro-stredníctvom druhých poslov; má však vždy špecifický bunkovýa molekulový cieľ (napr. NO a CO aktivujú KCa-kanály v plaz-matickej membráne buď priamo, alebo prostredníctvom cGMPdráhy) (Snyder a Ferris, 2000; Wang, 2002).
• Tieto „neštandardnéÿ vlastnosti molekúl NO, CO a H2S sa zú-častňujú na tom, že sa plynné neurotransmitery môžu zúčast-ňovať na komplexných interakciách prebiehajúcich v nervovomsystéme (Boehning a Snyder, 2003).
Oxid dusnatý
NO je syntetizovaný z l-arginínu enzýmom NO-syntázou (NOS)(obr. 3.28). V mozgu bola najvyššia koncentrácia NOS detegovaná v mo-zočku a hipokampe. Prekvapujúco nízka koncentrácia je v hypotalame(Moncada a spol., 1997).Na regulácii syntézy NO sa zúčastňuje glutamát, v menšej miere aj
iné neurotransmitery (napr. acetylcholín). Väzba glutamátu na NMDA-receptory aktivuje priamo, ako aj prostredníctvom iónov Ca2+ NOS,čo vedie k zvýšenej syntéze NO. NO difunduje z miesta syntézy dookolitých neurónov a stimuluje solubilnú guanylátcyklázu. Výsledkomje zvýšenie množstva sekundárneho posla, cGMP, v neuróne (Snydera Ferris, 2000).NO sa pravdepodobne zúčastňuje na procesoch spojených s vývo-
jom mozgu, tvorbou pamäťových stôp a reguláciou správania prostred-níctvom modulácie synaptickej plasticity. Počas ontogenézy nervovéhosystému je NO zapojený v procesoch modulujúcich arborizáciu dendri-tov. Predpokladá sa aj úloha NO v prenose nociceptívnych informácií,v procesoch poškodenia, ale aj protekcie neurónov (Inglis a spol., 1998).Experimentálne údaje poukazujú na to, že NO v oblasti nucleus pa-
raventricularis a supraopticus hypotalamu môže zasahovať do regulácie

92 NEUROTRANSMITERY
Obr. 3.28 Schéma biosyntézy NO z l-arginínu (modifikované podľa Moncadaa spol., 1997).
činnosti neuroendokrinného, ako aj autonómneho nervového systému(Costa a spol., 1996). Viacero nálezov svedčí o zapojení NO v regu-lácii sekrécie GnRH z hypotalamu. NO pravdepodobne pôsobí tlmivona činnosť HPA osi. Napríklad inhibuje stimulačný vplyv IL-1, vazo-presínu a zápalu na aktivitu HPA osi. NO týmto pôsobí prozápalovo,pretože obmedzuje uvoľňovanie protizápalovo pôsobiacich kortikoste-roidov z kôry nadobličiek. Predpokladá sa, že aj sekréciu prolaktínua rastového hormónu reguluje NO, pričom pravdepodobne moduluje ichsekréciu pôsobením priamo v adenohypofýze. Moduláciou hypotalamo-vého portálneho prietoku môže NO ovplyvňovať aj sekréciu vazopresínaa oxytocínu (Haddad a spol., 2002).Viaceré pozorovania poukazujú aj na zapojenie NO v modulácii
behaviorálnych reakcií. Predpokladá sa, že NO ovplyvňuje motivačnésprávania zahŕňajúce sexuálne a agresívne správania a správanie spo-jené s príjmom potravy. NO ovplyvňuje aj učenie a pamäť (Haddada spol., 2002).

3 Malomolekulové neurotransmitery 93
Experimentálne pozorovania poukazujú na to, že NO moduluje pre-nos signálov na všetkých úrovniach nervového systému, ktoré sa zú-častňujú na regulácii činnosti kardiovaskulárneho systému. NO modu-luje procesy spojené s barorecepciou, centrálnym spracovaním signá-lov (napr. v nucleus tractus solitarii, nucleus paraventricularic), pre-nosom signálov sympatikovými a parasympatikovými kardiálnymi ner-vami, ako aj na úrovni vodivého systému srdca (Paton a spol., 2002).Na inervácii ciev sa okrem klasických zložiek autonómneho sys-
tému zúčastňuje aj tzv. NANC-systém. Ako neurotransmitery NANC-systému slúžia viaceré neuropeptidy, ale aj NO. Práve nitrergická iner-vácia má veľmi významnú úlohu v regulácii cievneho tonusu. NO uvoľ-nený z nitrergických nervových zakončení aktivuje solubilnú guanylyl-cyklázu, čím dochádza k tvorbe cGMP v hladkom svalstve ciev a ich ná-slednej relaxácii. Nitrergické nervy inervujú takmer všetky cievy v tele(Toda a Okamura, 2003). NO uvoľnený z nervových zakončení v cor-pora cavernosae a v cievach penisu sa zúčastňuje na erekcii (Barananoa spol., 2001).NO ako významný neurotransmiter NANC-systému ovplyvňuje čin-
nosť hladkého svalstva vnútorných orgánov. V gastrointestinálnom sys-téme pôsobí NO uvoľnený z nervových zakončení myenterického plexuinhibične na činnosť hladkého svalstva. Niektoré nálezy poukazujú ajna spoločnú biosyntézu NO a CO v neurónoch inervujúcich gastro-intestinálny trakt, pričom tieto plynné neurotransmitery pôsobia akokotransmitery (Xue a spol., 2000).Uvažuje sa o tom, že NO môže predstavovať integrujúcu mole-
kulu, ktorá spája procesy aktivované „klasickouÿ chemickou transmi-siou s transmisiou z monoamínergických synáps typu en passant (Kissa Vizi, 2001).Úloha NO v etiopatogenéze ochorení CNS nie je jasná, ale pred-
pokladá sa účasť na autoimunitnom poškodení CNS (skleróza multi-plex, amyotrofická laterálna skleróza) a v etiopatogenéze Parkinsono-vej, Huntingtonovej a Alzheimerovej choroby (Esplugues, 2002). Expe-rimentálne údaje poukazujú aj na zapojenie NO v procesoch spojenýchs ischemickým poškodením neurónov (Snyder a spol., 1998).

94 NEUROTRANSMITERY
Oxid uhoľnatý
CO vzniká za fyziologických podmienok za účasti enzýmu hemoxyge-názy 2, ktorá štiepi porfyrínový reťazec hému na biliverdín, z ktoréhonásledne vzniká enzymatickou premenou bilirubín (obr. 3.29). Experi-mentálne údaje poukazujú na to, že syntéza CO je aktivovaná kaskádouprocesov, ktoré nasledujú po vstupe Ca2+ do neurónu. Hemoxygenáza 2je aktivovaná jednak priamo iónmi Ca2+, jednak kalmodulínom, ktorýnaviazal Ca2+, ktoré do stimulovaného neurónu vstúpilo. UvoľňovanieCO je teda dôsledkom príchodu akčného potenciálu do nervového za-končenia neurónu, z ktorého sa plynný neurotransmiter, resp. neuromo-dulátor uvoľňuje. Syntéza a uvoľňovanie CO preto odráža neurónálnuaktivitu (Wickelgren, 2003). Podobne ako NO aj CO aktivuje enzýmguanylátcyklázu, čo vedie k zvýšenej syntéze cGMP ako druhého posla(Snyder a Ferris, 2000).
Obr. 3.29 Schéma biosyntézy CO z hému (modifikované podľa Snyder a Ba-ranano, 2001).
Predpokladá sa, že CO je významný neuromodulátor v čuchovomsystéme, kde reguluje množstvo cGMP v neurónoch. V gastrointestinál-nom trakte CO, ktorý sa syntetizuje v myenterickom plexe, modulujeintestinálnu motilitu (Snyder a spol., 1998). Dokázalo sa, že CO je po-trebný pre normálnu ejakuláciu u myší (Snyder a Ferris, 2000; Wickel-gren, 2003). Prostredníctvommodulácie uvoľňovania vazopresínu sa COmôže zúčastňovať aj na regulácii neuroendokrinnej stresovej odpovede(Navarra a spol., 2000).

3 Malomolekulové neurotransmitery 95
Sírovodík
H2S sa syntetizuje z cysteínu enzýmom cystatión-β-syntázou (obr. 3.30)(Eto a Kimura, 2002). Predpokladá sa, že H2S pôsobí podobne akoiné plynné mediátory v nervovom systéme ako neuromodulátor (Wang,2002; Kamoun, 2004).
Obr. 3.30 Schéma biosyntézy H2S z cysteínu (modifikované podľa Wang,
2002).
Experimentálne zistenia poukazujú na to, že H2S vo fyziologickýchkoncentráciách zvyšuje neurotransmiterové procesy spojené s aktiváciouNMDA-receptorov a facilituje indukciu dlhodobej potenciácie v hipo-kampe, pravdepodobne prostredníctvom cAMP (Kimura, 2000). Pred-pokladá sa, že H2S sa zúčastňuje aj na modulácii činnosti HPA osi, čímmôže modulovať stresovú odpoveď (Navarra a spol., 2000).Uvažuje sa, že u pacientov s Alzheimerovou chorobou dochádza
k zníženiu aktivity cystatión-β-syntázy. Poukazuje na to nález výraznenižších koncentrácií H2S v mozgoch pacientov s Alzheimerovou choro-bou. Predpokladá sa, že zníženie tvorby H2S sa zúčastňuje na kogni-tívnom deficite, ktorý sprevádza Alzheimerovu chorobu (Eto a spol.,2002).
Prostaglandíny
Prostaglandíny sú metabolity kyseliny arachidónovej (obr. 3.31). Zara-denie prostaglandínov medzi neurotransmitery naráža na niekoľko ťaž-kostí. Aj keď sa nachádzajú v nízkom, ale detegovateľnom množstve

96 NEUROTRANSMITERY
v CNS, zdá sa, že sú skôr syntetizované v závislosti od aktuálnych po-trieb nervového tkaniva, ako by mali byť preformované a uložené dozásoby. Problémom je aj ich veľmi krátky biologický polčas (Webster,2001).Predpokladá sa, že prostaglandíny môžu participovať na regulácii
procesov spánku a bdenia, v centrálnej regulácii telesnej teploty, ale ajv modulácii dlhodobých potenciálov zapojených v pamäťových proce-soch a v procesoch neurotoxicity (Webster, 2001).
Obr. 3.31 Schéma biosyntézy prostaglandínov (modifikované podľa Narumiyaa spol., 1999).
Kanabinoidy
Kanabinoidy syntetizované v organizme (endokanabinoidy) sú derivátykyseliny arachidónovej konjugované s etanolamínom alebo glycerolom(obr. 3.32). Endogénne kanabinoidy sú signálnymi molekulami, ktoré

3 Malomolekulové neurotransmitery 97
sa objavili už vo včasnom evolučnom vývoji. Majú dôležité regulačnéfunkcie v organizme u všetkých stavovcov. Endokanabinoidy boli de-tegované v mozgu, plazme a periférnych tkanivách. Kanabinoidy sav mozgu viažu na kanabinoidné receptory CB1 a CB2, ktoré sú spo-jené s G-proteínom (tab. 3.12) (Howlett a spol., 2002).
Obr. 3.32 Štruktúrne vzorce endogénnych agonistov a exogénneho agonistu
kanabinoidných receptorov (Rodriguez de Fonseca a spol., 2004).
Predpokladá sa, že systém neurónov, ktoré uvoľňujú endokanabi-noidy, zasahuje do regulácie odmeňovania, motivácie a emočnej home-ostázy (Rodriguez de Fonseca a spol., 2005). Okrem toho sa uvažujeo tom, že v hipokampe sa zúčastňujú na modulácii učenia a pamäte,prostredníctvom bazálnych ganglií ovplyvňujú motorickú aktivitu a mo-dulujú príjem potravy (Irving a spol., 2002).
Iné neurotransmitery obsahujúce aminoskupinu
Okrem klasických monoamínov (napr. dopamín, sérotonín, histamín) sapredpokladá, že aj ďalšie molekuly obsahujúce aminoskupinu môžu pl-

98 NEUROTRANSMITERY
CB1 Gi/o ↓AC ↓cAMP
Metabotropné CB2 Gi/o ↓AC ↓cAMP
non-CB1/CB2 Gi/o ? ↓AC ↓cAMP ?
Tabuľka 3.12 Klasifikácia receptorov pre kanabinoidy a mechanizmy trans-dukcie signálu. CB1-receptory môžu za špecifických podmienok aktivovať aj
Gs-proteín, a to vtedy, keď sú súčasne aktivované ďalšie Gi/o-receptory (modi-
fikované podľa Mackie a Stella, 2006).
niť funkcie neurotransmiterov. Medzi tieto látky patrí DOPA a tzv. sto-pové amíny tryptamín, fenyletylamín, tyramín a oktopamín (obr. 3.33)(Webster, 2001).Stopové amíny, endogénne metabolity biogénnych amínov sa nachá-
dzajú v nervovom systéme v nízkych koncentráciách. Ukázalo sa však, žeich exogénne podanie vyvoláva zmeny v aktivite nervového systému. Dl-hodobo sa uvažuje aj o ich možnej účasti na etiopatogenéze niektorýchochorení CNS (napr. depresia, schizofrénia). Až v poslednom obdobívšak boli identifikované v CNS pre stopové amíny receptory spriahnutés G-proteínom (Lindemann a Hoener, 2005).Zaujímavou molekulou, potenciálnym neurotransmiterom, resp.
neuromodulátorom je D-serín (aminokyseliny sú u vyšších organizmovprevažne v L-konfigurácii). Predpokladá sa, že pôsobí prostredníctvomNMDA-receptorov (Snyder a Kim, 2000; Wolosker a spol., 2002). Po-dobne ako pri D-seríne sa predpokladá, že aj D-aspartát by mohol slúžiťako neurotransmiter, resp. neuromodulátor (Boehning a Snyder, 2003).
L-DOPA
Experimentálne údaje poukazujú na to, že aj L-DOPA (obr. 3.14, 3.33),prekurzor dopamínu, je potenciálnym neurotransmiterom, resp. neuro-modulátorom (Misu a spol., 1996).Predpokladá sa, že L-DOPA pôsobí ako neurotransmiter primárnych
nervových zakončení vedúcich informácie z baroreceptorov do nucleus

3 Malomolekulové neurotransmitery 99
Obr. 3.33 Schéma biosyntézy DOPA, tryptamínu, fenyletylamínu, oktopamínu(A) a tyramínu (B) (modifikované podľa Webster, 2001).
tractus solitarii (Misu a spol., 2003). Okrem toho sa predpokladá, že saL-DOPA môže zúčastňovať na procesoch oneskoreného zániku neurónovv striate a CA1 oblasti hipokampu po pôsobení prechodnej ischémie(Misu a spol., 2003).
Imidazolínový systém
Ukázalo sa, že klonidín (látka s antihypertenzívnym účinkom) sa viažeokrem centrálnych α2-receptorov aj na tzv. imidazolínové väzbovémiesta (Eglen a spol., 1998). V súčasnosti sa rozlišujú tri imidazo-línové väzbové miesta na základe špecificity ligandov a subcelulárnej

100 NEUROTRANSMITERY
lokalizácie. Najlepšie sú charakterizované I1-receptory, údaje týkajúcesa I2-väzbových miest sú kontroverznejšie. I3-väzbové miesta, pri kto-rých sa predpokladá, že sa zúčastňujú na regulácii sekrécie inzulínuz β-buniek pankreasu, boli identifikované iba nedávno (Morgan a Chan,2001). I1-receptory sa nachádzajú v rôznych častiach CNS. Nachá-dzajú sa v oblastiach mozgového kmeňa, ktoré sa zúčastňujú na re-gulácii krvného tlaku, ako aj v locus coeruleus (Hudson, 2000). Naj-vyššia koncentrácia I1-receptorov je v striate, pallide, gyrus dentatus,amygdale a substantia nigra. Predpokladá sa, že I2-väzbové miesta samôžu nachádzať na enzýme monoaminooxidáza, ktorý sa zúčastňujena biodegradácii monoamínov. Ako o endogénnych ligandoch viažu-cich sa na imidazolínové receptory, resp. väzbové miesta, sa uvažujeo agmatíne (vzniká dekarboxyláciou arginínu), tryptamíne, harmane,kyseline ribotidimidazol-4-octovej a kyseline ribozidimidazol-4-octovej(obr. 3.34) (Reis a Regunathan, 2000; Head, 2003; Dardonville a Rozas,2004; Prell a spol., 2004). Experimentálne údaje poukazujú na anti-depresívny, anxiolytický, protizápalový, antiproliferačný a neuroprotek-tívny účinok agmatínu. Či sú však všetky tieto účinky sprostredkovanéväzbou agmatínu na imidazolínové receptory alebo väzbou na iný typreceptorov, nie je zatiaľ známe (Holt, 2003).
Obr. 3.34 Štruktúrne vzorce endogénnych ligandov pre imidazolínové recep-tory (Hudson, 2000; Prell a spol., 2004).

3 Malomolekulové neurotransmitery 101
Viaceré experimentálne údaje poukazujú na pozitívny vplyv imi-dazolínov v liečbe závislosti. Klonidín je efektívny napríklad v liečbezávislosti od nikotínu a opiátov (Kleber, 2003; Gourlay a spol., 2004).Predpokladá sa, že uvedený efekt klonidínu môže byť sprostredkovanýaktiváciou imidazolínových a adrenergických receptorov vo VTA, ob-lasti kľúčovo zapojenej v procesoch vzniku závislosti (Georges a Aston-Jones, 2003).

Peptidové („veľkomolekulovéÿ)
neurotransmitery
B. Mravec
Prenos signálov prostredníctvom molekúl peptidov je charakteris-tický pre endokrinný systém. Ako prvé boli charakterizované jednotlivéneuropeptidy ako hormóny hypofýzy a periférnych tkanív (napr. v gas-trointestinálnom trakte) (Rehfeld, 1998). Neskôr sa dokázalo, že mnohopeptidov spĺňa charakteristiky neurotransmiterov. V súčasnosti je zná-mych viac ako 50 rôznych neuropeptidov (tab. 2.3).Termín neuropeptidy v jeho súčasnom význame prvýkrát použili
roku 1971 na pomenovanie fragmentov peptidových hormónov, ktorésíce stratili hormonálnu aktivitu, ale boli schopné ovplyvniť správanie.Moderná definícia hovorí, že neuropeptid je peptid prítomný v ner-vovom systéme, ktorý má biologické účinky, ak sa podáva exogénne(Klavdieva, 1995).Vo výskume neuropeptidov mali významnú úlohu práce Ramona
y Cajala, ktorý opísal nervové dráhy z jadier hypotalamu do zadnéholaloka neurohypofýzy. Poznatky, ktoré vyplynuli z obsiahlej práce, ktorúuskutočnili Geoffrey Harris, ako aj iní vedci v 50. rokoch 20. storočia,preukázali, že funkciu adenohypofýzy reguluje hypotalamus prostred-níctvom tzv. hypotalamických faktorov. Na konci 60. a na začiatku
102

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 103
70. rokov sa dokázalo, že hypotalamické faktory sú peptidy. Významnýprelom vo výskume neuropeptidov znamenali nálezy dokazujúce, že neu-róny hypotalamu syntetizujúce vazopresín a oxytocín, okrem neuro-hypofýzy, vysielajú axóny aj do extrahypotalamických oblastí nervo-vého systému. Ďalším dôležitým objavom bolo zistenie, že neuropep-tidy a klasické neurotransmitery (napr. noradrenalín) sú kolokalizo-vané v tých istých neurónoch. Navyše sa ukázalo, že syntéza niektorých(napr. opioidových peptidov) prebieha nielen v mozgu, ale aj v perifér-nych tkanivách (Landgraf a Neumann, 2004).Peptidové neurotransmitery spĺňajú viacero (niektoré všetky) kri-
térií potrebných na ich zaradenie medzi neurotransmitery. V porov-naní s malomolekulovými neurotransmitermi sú však rozdiely v biosyn-téze a uvoľňovaní peptidových neurotransmiterov. Medzi charakteris-tiky peptidových neurotransmiterov patrí (Kandel a spol., 2000):
• sú syntetizované v tele neurónu, kde sa nachádza potrebný synte-tický aparát (malomolekulové neurotransmitery sú syntetizovanév nervovom zakončení),
• jeden gén môže kódovať viacero typov neuropeptidov, ktorévznikajú na základe rozdielneho enzymatického spracovania pro-duktu tohto génu (napr. neuropeptidy zo skupiny proopiomela-nokortínov),
• sú prítomné v tkanivách v omnoho nižších koncentráciách v po-rovnaní s klasickými neurotransmitermi,
• je potrebná vyššia frekvencia vzruchov, aby došlo k ich uvoľneniuz neurónov,
• enzymatickou konverziou neuropeptidov (napr. opioidov, tachy-kinínov, CGRP) môžu vznikať fragmenty so zachovanou, alebomodifikovanou biologickou aktivitou (Hallberg a Nyberg, 2003).
Novšie pozorovania naznačujú, že neuropeptidy sa môžu v mozguuvoľňovať často z dendritov. Neuropeptidy potom môžu difundovať dookolia a pôsobiť aj na neuróny vzdialené od miesta ich uvoľnenia (Morrisa Ludwig, 2004; Ludwig a Leng, 2006).Neuropeptidy pôsobia ako neurotransmitery, resp. neuromodulátory
v rôznorodých nervových okruhoch. Vyplavenie neuropeptidov a obsa-

104 NEUROTRANSMITERY
denie ich receptorov nevyvoláva väčšinou rýchle, ale skôr pomalé a dlh-šie trvajúce zmeny v excitabilite a aktivite neurónov, ktoré sa môžuprejaviť aj zmenou v génovej expresii neurónu, ktorá môže byť pod-kladom dlhodobých morfologických a funkčných zmien v nervovom sys-téme. Keďže sú neuropeptidy zapojené do modulácie rôznorodých funk-cií, skúma sa aj ich význam v etiopatogenéze ochorení nervového sys-tému (Ressler a Nemeroff, 2001; Swaab, 2004). Medzi ochorenia, v kto-rých etiopatogenéze sa predpokladá účasť neuropeptidov, patria napr.látková závislosť, depresia, záchvatové stavy, poruchy príjmu potravya poruchy spánku (Hokfelt a spol., 2003).Experimentálne údaje poukazujú na to, že peptidové neurotrans-
mitery okrem prenosu signálov môžu mať aj dôležitú úlohu pri proce-soch maturácie nervového systému a regenerácie poškodených neurónov(Palkovits, 1995; Strand, 2000).
Malomolekulové a peptidové neurotransmitery –spoločná syntéza a uvoľňovanie z jedného neurónu
V súčasnosti sa akceptuje, že jeden neurón môže využívať ako neuro-transmitery súčasne viacero chemických látok. V jednom neuróne tedamôžu byť viaceré neurotransmitery:
• syntetizované – koexpresia a kosyntéza,
• uskladnené v nervovom zakončení – kolokalizácia,
• uvoľňované z nervového zakončenia – kotransmisia (Burnstock,1976; Davanger, 1996).
Výskyt viacerých neurotransmiterov a ich kolokalizácia v neurónevšak nemusí nevyhnutne znamenať, že sa aj spoločne uvoľňujú z jednejsynapsy (bližšie pozri Marder, 1999).Príkladom kolokalizácie sú peptidergické a malomolekulové neuro-
transmitery, ktoré sa môžu syntetizovať v tom istom neuróne (Hokfelta spol., 1987). Zvyčajne jeden neurón uvoľňuje jeden malomolekulovýneurotransmiter a súčasne jeden alebo viac neuropeptidov (tab. 4.1).Ak je syntetizovaných viac neuropeptidov v jednom neuróne, ide o pep-tidy, ktoré väčšinou vznikajú z jedného prekurzorového polypeptidu.Jedným zo základných rozdielov medzi peptidmi a klasickými (malo-molekulovými) neurotransmitermi je však to, že kým malomolekulové

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 105
neurotransmitery sa syntetizujú priamo v synaptickom zakončení, neu-ropeptidy sa syntetizujú v tele neurónu a až potom sa transportujú dosynaptických zakončení (Bean a spol., 1994).
dopamíncholecystokinín
enkefalín
somatostatín
noradrenalínenkefalín
neurotenzín
neuropeptid Y
adrenalín enkefalín
substancia P
sérotonín TRH
enkefalín
VIP
acetylcholínenkefalín
neurotenzín
somatostatín
GABAsomatostatín
cholecystokinín
glutamátsubstancia P
CGRP
CRH vazopresín
Tabuľka 4.1 Prehľad najčastejšie sa vyskytujúcej kolokalizácie neurotransmi-terov (modifikované podľa Stahl, 2000).
V prípade uvoľňovania kotransmiterov z jednej synapsy sa pozoro-valo, že pri nižších frekvenciách stimulácie neurónu sa uvoľňujú pre-važne malomolekulové neurotransmitery, kým pri intenzívnejšej stimu-

106 NEUROTRANSMITERY
lácii charakterizovanej vyššou frekvenciou vzruchov sa uvoľňujú aj pep-tidové neurotransmitery (Siegel a spol., 2006).Pri kotransmisii pôsobia uvoľnené neuropeptidy na činnosť okolitých
neurónov väčšinou neuromodulačne (Nusbaum a spol., 2001).Okrem kotransmisie malomolekulového neurotransmitera a neuro-
peptidov sa hromadia dôkazy svedčiace aj o spoločnom uvoľňovanídvoch malomolekulových neurotransmiterov z jedného neurónu. Expe-rimentálne zistenia poukazujú na to, že ATP sa uvoľňuje spolu s ace-tylcholínom alebo noradrenalínom zo sympatikových nervových zakon-čení. Uvažuje sa aj o uvoľňovaní ATP spolu s acetylcholínom z cho-línergických neurónov v striate. Zaujímavým pozorovaním je spoločnéuvoľňovanie klasického excitačného (glutamát) a inhibičného (GABA)neurotransmitera z neurónov tzv. machových (mossy) vlákien v hipo-kampe. Predpokladá sa, že táto „atypickáÿ kombinácia neurotransmi-terov má významnú úlohu počas vývinu nervového systému. Uvažujesa, že koexpresia glutamátu a GABA môže zabezpečovať adekvátnevytvorenie neuronálnych okruhov a ovplyvniť expresiu a funkciu recep-torov pre glutamát a GABA. V dospelom mozgu môže takáto kombi-nácia neurotransmiterov inhibovať procesy spojené s hyperexcitabilitouv CNS (Gutierrez, 2005). Cholínergické neuróny v bazálnej časti pred-ného mozgu zasa pravdepodobne uvoľňujú okrem acetylcholínu aj glu-tamát. Viacero pozorovaní poukazuje na kotransmisiu glutamátu spolus monoamínmi. Na základe týchto pozorovaní sa preto uvažuje aj o mož-nej účasti narušenej kotransmisie malomolekulových neurotransmiterovpri viacerých ochoreniach nervového systému (Trudeau, 2004).
Vybrané peptidergické neurotransmiteryB.Mravec, A.Kiss
Neuropeptidy sú biologicky aktívne látky produkované nervovýmibunkami. Funkčne spĺňajú úlohu neurohormónov, neurotransmiterova neuromodulátorov. Pôsobia nielen v CNS, ale aj na periférii.Peptidy, ktoré môžu pôsobiť ako neurotransmitery, resp. neuromo-
dulátory, predstavujú veľkú skupinu látok, ktorá sa člení podľa príbuz-nosti do niekoľkých podskupín (tab. 2.3). V porovnaní s malomolekulo-vými neurotransmitermi možno očakávať, že prevažná väčšina v budúc-nosti identifikovaných neurotransmiterov bude patriť práve do skupiny

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 107
neuropeptidov. Pretože sa stále rozpoznávajú nové neuropeptidy, časo-pis Peptides pravidelne uverejňuje prehľadové články, ktoré sumarizujúnovoobjavené neuropeptidy a ich funkcie (Zadina a spol., 1986; Ahmeda spol., 1994; Pan a spol., 1999; McLay a spol., 2001). V nasledujúcomtexte sú zhrnuté údaje o vybraných neuropeptidoch s dôrazom na ichfunkcie v nervovom systéme.
Klasické neurohormóny CNS
Vazopresín a oxytocín
Vazopresín (arginine-vasopressin, AVP) a oxytocín (OXY) sa skla-dajú z 9 aminokyselinových zvyškov a sú typickými neurohormónmi,ktoré sú produkované v hypotalame. Obidva hormóny sú syntetizo-vané v dvoch výrazných konglomeráciách hypotalamických neurónov(nucleus paraventricularis hypothalami a nucleus supraopticus hypot-halami) a niekoľkých malých skupinkách neurónov, tzv. akcesórnychjadrách hypotalamu. Tieto neuróny vysielajú axóny, ktoré vytvárajútzv. hypotalamo-neurohypofýzovú neurosekretorickú dráhu, ktorej úlo-hou je presun OXY a AVP z hypotalamu do neurohypofýzy. Z neuro-hypofýzy sa oba hormóny vyplavujú na adekvátny podnet do krvnéhoriečiska, ktoré ich transportuje k cieľovým orgánom (McEwen, 2004).Ich spätnému účinku na tkanivo mozgu bráni mozgovo-cievna bariéra.
Vazopresín
Vazopresín označovaný aj ako argipresín alebo antidiuretický hormón(ADH) je ľudský hormón, ktorý sa vylučuje z neurohypofýzy hlavnepočas hypovolémie a zvýšenej osmolality krvnej plazmy. Pôsobí v ob-ličkách, kde znižuje vylučovanie vody zvýšením jej spätnej resorpcie.To vedie k zadržiavaniu vodu v organizme a spôsobuje tvorbu koncen-trovaného moču. Týmto mechanizmom AVP chráni organizmus preddehydratáciou. Podnet na vylúčenie vazopresínu počas hypovolémie jesprostredkovaný tlakovými receptormi ciev, kým osmotické zmeny krv-nej plazmy registrujú aj samy neuróny nucleus paraventricularis hypo-thalami (PVN) a nucleus supraopticus hypothalami (SON). Vazopresínúčinkuje prostredníctvom troch receptorov V1a, V1b a V2.

108 NEUROTRANSMITERY
Sekréciu AVP ovplyvňuje množstvo faktorov – napr. etylalkohola kofeín redukujú jeho sekréciu, kým angiotenzín jeho sekréciu stimu-luje.Väčšina vazopresínu cirkulujúceho v krvi pochádza z magnocelulár-
nych neurónov PVN a SON neurohypofýzy. Niektoré z nádorov môžu se-cernovať aj vazopresín. AVP, ktorý účinkuje lokálne, pochádza z parvo-celulárnych neurónov PVN, ktorý sa transportuje krátkymi portálnymicievami stopky hypofýzy do adenohypofýzy, kde reguluje v súčinnostis kortikoliberínom uvoľňovanie ACTH do krvného obehu. AVP je pretopopri CRH druhým stresovým hormónom, ktorý sa vylučuje v rôznychstresových situáciách (Holmes a spol., 2003). AVP sa týmto zúčast-ňuje na regulácii činnosti „hypotalamo-hypofýzo-adrenokortikálnejÿosi. Okrem toho sa AVP produkuje v parvocelulárnych neurónoch v ďal-ších oblastiach mozgu, medzi ktoré patria corpus amygdaloideum a nuc-leus interstitialis striae terminalis. Neuróny v týchto oblastiach uvoľňujúAVP ako neurotransmiter v rozličných častiach mozgu.AVP pôsobiaci v CNS má mnoho funkcií, napríklad účasť na for-
movaní krátkodobej a dlhodobej pamäte, na modulácii reakcií agresi-vity, na regulácii krvného tlaku a telesnej teploty. Ovplyvňuje sociálnesprávanie a vylučuje sa aj počas sexuálnej aktivity a zúčastňuje sa naudržiavaní partnerských vzťahov a vzniku agresivity v rámci samčíchkonkurenčných vzťahov (Goodson a Bass, 2001). AVP prítomný v nuc-leus suprachiasmaticus hypothalami je zapojený do modulácie cirka-diánnych rytmov.
Oxytocín
Oxytocín pôsobí okrem periférie aj centrálne a vylučuje sa v rôznychoblastiach mozgu, kde má úlohu typického neurotransmitera. OXY syn-tetizovaný v parvocelulárnych neurónoch má v porovnaní s OXY se-cernovaným magnocelulárnymi neurónmi širšie spektrum účinkov. Vý-znamná je úloha OXY v modulácii sociálneho správania u zvierat aju človeka. OXY výrazne posilňuje rodičovský inštinkt a ovplyvňuje ajsexuálne správanie. Aplikácia OXY do mozgových komôr spôsobujespontánnu erekciu a urýchľuje pohyb spermií (Gimpl a Fahrenholz,2001). U samíc stimuluje pôrod a laktáciu (Schreiber, 1992). Má ajantistresové účinky a na rozdiel od stresovej reakcie „fight or flightÿ(boj alebo útek) pôsobí pozitívne a upevňuje priateľské vzťahy medzi

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 109
jedincami (Kosfeld a spol., 2005). OXY znižuje hladiny stresových hor-mónov, hlavne kortikosterónu (resp. kortizolu u človeka). Na druhejstrane zvyšuje toleranciu bolesti a strachu. Niektoré pozorovania na-značujú, že OXY redukuje toleranciu návykových látok, ako sú napr.opiáty, kokaín, alkohol, a redukuje aj abstinenčný syndróm (Kovacsa spol., 1998).Na periférii sa oxytocínové receptory nachádzajú u niektorých hlo-
davcov aj v srdci a počas embryogenézy podporujú diferenciáciu kardi-omyocytov (Paquin a spol., 2002; Jankowski, 2004).Parvocelulárne neuróny nucleus paraventricularis hypothalami,
ktoré syntetizujú oxytocín a vazopresín, inervujú autonómne nervovécentrá mozgového kmeňa a miechy. Tým sa OXY a AVP neuróny hy-potalamu zúčastňujú na regulácii činnosti autonómneho nervového sys-tému (McEwen, 2004; Kiss a Mikkelsen, 2005).
Uvoľňujúce a inhibujúce hormóny hypotalamu
Kortikoliberín a príbuzné neuropeptidy
Kortikotropín uvoľňujúci hormón (kortikoliberín, corticotrophin relea-sing hormone, CRH, CRF), urokortín, urotenzín a sauvagín patria dospoločnej rodiny neuropeptidov (Lovejoy a Balment, 1999; Bale a Vale,2004).Kortikoliberín, peptid zložený z 42 aminokyselinových zvyškov, je
primárnym stresovým hormónom mozgu a zodpovedá za aktiváciu stre-sového systému, t.j. hypotalamo-hypofýzo-adrenokortikálnej osi (hypo-thalamic-pituitary-adrenal axis, HPA axis). Je to systém, ktorý stresomvyvolané senzorické signály spracúvané rôznymi oblasťami transformujeprostredníctvom HPA osi na stresovú odpoveď príslušnej kvality a kvan-tity. CRF sa viaže na dva podtypy receptorov CRF1 a CRF2, ktoré súspriahnuté s G-proteínom (Hauger a spol., 2003).
Kľúčovými elementmi HPA osi sú:
• nucleus paraventricularis hypothalami, ktorého parvocelulárneneuróny syntetizujú CRH spoločne s vazopresínom,
• adenohypofýza, kde synergickým účinkom CRH a AVP dochá-dza k zvýšenej syntéze a sekrécii adrenokortikotropného hormónu(ACTH),

110 NEUROTRANSMITERY
• kôra nadobličky, kde ACTH reguluje vyplavovanie glukokortiko-idov do periférnej cirkulácie.
Vyplavené glukokortikoidy spätne ovplyvňujú (inhibujú) sekréciuCRH a AVP, ako aj sekréciu ACTH. Transport CRH a AVP do adeno-hypofýzy je sprostredkovaný portálnymi cievami. Terminály PVN neu-rónov dosadajú na portálne cievy v oblasti eminentia mediana (na dnetretej mozgovej komory), kde uvoľňujú CRH a AVP, ktoré sa krvnýmriečiskom cez stopku hypofýzy dopravujú k ACTH-bunkám adenohypo-fýzy. ACTH, uvoľnený z buniek adenohypofýzy, sa transportuje krvnýmriečiskom do nadobličky, kde stimuluje biosyntézu a sekréciu glukokorti-koidov. Glukokortikoidy pôsobia prostredníctvom glukokortikoidovýchreceptorov vo väčšine tkanív organizmu. Počas stresu sa účinok glu-kokortikoidov výrazne prejavuje v hipokampe, primárnom nadradenomcentre HPA osi. Za normálnych podmienok hladiny kortizolu u človekakolíšu, t.j. ráno sú veľmi vysoké, postupne sa znižujú, neskoro večeropäť narastajú a o polnoci sú opäť nízke. Trvalé zvýšenie hladín glu-kokortikoidov má neurodegeneratívny účinok na neuróny hipokampu.Degenerácia hipokampu vedie k poškodeniu pamäti a strate predchá-dzajúcich skúseností so zvládaním stresovej reakcie, čo môže zapríčiniťneadekvátnu reakciu organizmu na stres.K významným poznatkom patrí aj to, že pri niektorých psychiatric-
kých ochoreniach (afektívne choroby) preukazuje HPA os zvýšenú ak-tivitu spôsobenú hyperaktivitou CRH-neurónov v PVN. Predklinickéa klinické štúdie ukázali, že za vznik abnormálnej aktivity HPA osizodpovedajú genetické aj environmentálne faktory. Okrem PVN za zvý-šenie aktivity HPA osi však zodpovedajú aj CRH-neuróny tých oblastímozgu, ktoré spracúvajú informácie týkajúce sa behaviorálnych reakcií.Niektoré parvocelulárne neuróny nucleus paraventricularis vysie-
lajú axóny do oblastí mozgového kmeňa a miechy. Tieto neuróny sapravdepodobne zúčastňujú na regulácii činnosti autonómneho nervo-vého systému. K ďalším hypotalamickým oblastiam, ktoré syntetizujúCRF, patria area preoptica medialis, nucleus dorsomedialis, nucleus ar-cuatus, posteriórny hypothalamus a nuclei mamillares. Okrem nucleusparaventricularis sa CRF-neuróny (ako aj CRF-receptory) nachádzajúaj v ďalších oblastiach mozgu, ktoré sú zapojené do stresovej odpo-vede, medzi ktoré patria napríklad amygdala, laterálne septum, locuscoeruleus a rafeálne jadrá v mozgovom kmeni. Aj niektoré neuróny neo-kortexu syntetizujú CRF ako svoj neurotransmiter. Ide predovšetkým

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 111
o bipolárne interneuróny lokalizované prevažne v I. a IV. vrstve mozgo-vej kôry, s najvyšším výskytom v prefrontálnej, inzulárnej a cingulárnejoblasti (Davis a spol., 2002).Predpokladá sa, že narušená činnosť CRH-neurónov sa vyskytuje
u pacientov s depresiou, anxietou, látkovou závislosťou, syndrómomdráždivého čreva, ďalej u pacientov so zápalmi a dysfunkciou kardiovas-kulárneho systému (Holmes a spol., 2003; Bale a Vale, 2004; Heinrichsa Koob, 2004).
Urokortín
V mozgu cicavcov sa nachádzajú okrem CRF aj ďalšie tri príbuzné pep-tidy - urokortín 1, 2 a 3, ktoré prejavujú vysokú afinitu nielen k CRF1,ale hlavne k CRF2-receptorom (Hauger a spol., 2003). Účinky urokor-tínov sú v mnohých prípadoch podobné účinkom CRH. No v prípadeich anorexigenického efektu je urokortín účinnejší. Štúdium anatomickejdistribúcie preukázalo, že neuróny syntetizujúce urokortín sú väčšinoulokalizované v iných oblastiach CNS v porovnaní s neurónmi synteti-zujúcimi CRH. To poukazuje na určitú odlišnosť v úlohe urokortínuv porovnaní s CRH. Predpokladá sa, že urokortín je zapojený v regu-lácii príjmu potravy, ovplyvňuje motoriku, stresovú reakciu a anxietu(Dautzenberg a Hauger, 2002; Reul a Holsboer, 2002). Na periférii uro-kortín pravdepodobne ovplyvňuje činnosť srdca (Skelton a spol., 2000;Latchman, 2002).
Urotenzín II
Urotenzín II je veľmi účinná vazokonstrikčná látka. Špecifický ľudskýurotenzínový receptor identifikovali roku 1999 (Ames a spol., 1999).Tieto receptory boli opísané v mnohých oblastiach CNS a endokrin-ných tkanivách. Predpokladá sa, že urotenzín II moduluje činnosť týchoblastí nervového systému, ktoré sa zúčastňujú na regulácii aktivitysympatikoadrenálneho systému. Aj urotenzín II pravdepodobne stimu-luje HPA-systém a má úlohu v mnohých patofyziologických procesoch,ako sú napr. ateroskleróza, kardiálna hypertrofia, pulmonárna hyper-tenzia a nádorová proliferácia (Maguire a Davenport, 2003; Richardsa Charles, 2004).

112 NEUROTRANSMITERY
Rastový hormón uvoľňujúci hormón
Rastový hormón uvoľňujúci hormón (growth hormone releasing hor-mone, GHRH) sa skladá zo 44 aminokyselinových zvyškov. Tvorí sav neurónoch nucleus arcuatus hypothalami. GHRH je uvoľňovaný z ner-vových zakončení týchto neurónov v oblasti eminentia mediana dohypotalamo-hypofýzového portálneho systému, ktorým sa transportujedo adenohypofýzy. Tam stimuluje produkciu a sekréciu rastového hor-mónu. GHRH je secernovaný v pravidelných pulzoch a indukuje sekréciurastového hormónu tiež pulzového charakteru. Centrálny efekt GHRHnie je známy.
Somatostatín (hormón inhibujúci uvoľňovanie rastového hormónu)
Somatostatín (growth hormone release inhibiting hormone, SOM) pred-stavujú dva neuropeptidy zložené zo 14 a 28 aminokyselinových zvyš-kov. Somatostatín je zapojený do regulácie vyplavovania rastového hor-mónu z adenohypofýzy, inhibície rastu nádorov, kognitívnych procesov,spánku a motorickej aktivity (Vezzani a Hoyer, 1999).V CNS je somatostatín produkovaný v hypotalame, a to neuro-
endokrinnými neurónmi nucleus periventricularis hypothalami. Tietoneuróny projikujú do oblasti eminentia mediana, odkiaľ sa somatosta-tín uvoľňuje z nervových zakončení do hypotalamo-hypofýzovej portál-nej cirkulácie a krvou transportuje do adenohypofýzy. V adenohypo-fýze pôsobí ako inhibítor sekrécie somatotropných buniek. Somatosta-tínové neuróny hypotalamu sú pod kontrolou cirkulujúceho rastovéhohormónu, ktorý podobne ako glukokortikoidy spätnoväzbovo regulujeaktivitu týchto neurónov.Somatostatín produkujú aj iné populácie neurónov v mozgu, ako sú
napr. nucleus arcuatus, hipokampus a nucleus tractus solitarii, ktoréinervujú rôzne mozgové oblasti s vysokou koncentráciou somatostatí-nových receptorov. Somatostatín má aj úlohu v inhibičnej moduláciihipokampových nervových okruhov. Predpokladá sa preto, že somato-statín môže mať úlohu v kontrole parciálnych záchvatových stavov (Bi-naschi a spol., 2003). Na periférii je somatostatín secernovaný bunkamipankreasu.Somatostatín sa klasifikuje ako inhibičný hormón, ktorého hlavné
účinky zahŕňajú okrem inhibície sekrécie rastového a tyreoidového hor-mónu aj supresiu sekrécie gastrointestinálnych hormónov, ako sú gas-

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 113
trín, cholecystokinín, sekretín, motilín, vazoaktívny intestinálny poly-peptid, gastrický inhibičný polypeptid a enteroglukagón. Inhibuje ajsekréciu hormónov pankreasu, inzulínu a glukagónu. Somastostatín zni-žuje rýchlosť vyprázdňovania obsahu žalúdka a redukuje kontrakciuhladkého svalstva a prietok krvi v gastrointestinálnom trakte.
Tyreoliberín
Tyreoliberín (thyreotropin releasing hormone, TRH) bol pôvodne iden-tifikovaný ako hypotalamový neuropeptid, ktorý stimuluje uvoľňovanietyreotropného hormónu z adenohypofýzy. Predpokladá sa, že okremtejto funkcie účinkuje v CNS aj ako neurotransmiter, resp. neuromodu-látor. Pozorovalo sa, že TRH mení excitabilitu neurónov v mozgovomkmeni a mieche. Receptory pre TRH, ako aj TRH obsahujúce nervovévlákna a zakončenia sa identifikovali v oblasti spinálnych sympatiko-vých pregangliových neurónov. Predpokladá sa preto, že TRH uvoľňo-vaný v mieche ovplyvňuje činnosť sympatikového nervového systému(Kolaj a spol., 1997).
Deriváty proopiomelanokortínu (ACTH, MSH a iné)
Do skupiny peptidov vznikajúcich z proopiomelanokortínu (POMC)patria adrenokortikotropný hormón (ACTH), melanocyty stimulujúcihormón (MSH) a opioidy (Raffin-Sanson a spol., 2003). Predpokladása, že intenzita syntézy POMC odráža aj energetický stav organizmu(Wynne a spol., 2005).Ako melanokortíny sa označujú deriváty POMC, medzi ktoré pat-
ria ACTH, α-MSH, β-MSH a γ-MSH. α-MSH a ACTH ovplyvňujúpigmentáciu kože. ACTH okrem toho moduluje steroidogenézu. MSHje zapojený v procesoch regulácie príjmu potravy. Predpokladá sa ajúčasť MSH na regulácii reprodukčných procesov a regulácii kardiovas-kulárnych funkcií. Potenciálne veľmi významným môže byť protizápa-lový účinok α-MSH. Centrálne podanie α-MSH je schopné inhibovať ajzápalové procesy prebiehajúce na periférii organizmu (Starowicz a Prze-wlocka, 2003; Seeley a spol., 2004).

114 NEUROTRANSMITERY
Opioidy
V organizme sa tvoria tri druhy opioidových neuropeptidov: endorfíny,enkefalíny a dynorfíny. Aj keď sa tieto látky označujú ako opioidy, ichchemická štruktúra sa od opiátov líši. Opioidové neuropeptidy môžuúčinkovať ako neurotransmitery alebo neuromodulátory, ktoré modifi-kujú účinok iných neurotransmiterov v CNS. Opioidy môžu napríkladzmeniť elektrické vlastnosti neurónov natoľko, že sa stanú rezistentnýmivoči excitácii.Endorfíny sú endogénne opioidy s analgetickými vlastnosťami. Sú
produktom génu, ktorý kóduje veľkú prekurzorovú molekulu proopio-melanokortín. POMC je exprimovaný v hypofýze, ako aj v nucleusarcuatus hypothalami (Akil a spol., 1988). K endorfínom patria α-,β- a γ-endorfíny, z ktorých najznámejší je β-endorfín zúčastňujúci sahlavne na tlmení bolesti. β-endorfín sa z hypofýzy uvoľňuje do krvi, alejeho periférny účinok nie je presne známy. Centrálny účinok periférneuvoľneného endorfínu je minimálny vzhľadom na jeho nepriechodnosťmozgovo-cievnou bariérou. Behaviorálne efekty β-endorfínu sú spôso-bené účinkami hypotalamického β-endorfínu, ktorý pôsobí na mnohýchmiestach mozgu a miechy.Enkefalíny sa môžu vyskytovať v dvoch formách ako metionínový
enkefalín (met-enkephalin) a ako leucínový enkefalín (leu-enkephalin).Neuróny produkujúce enkefalíny majú rozsiahlejší výskyt v mozgu akoneuróny syntetizujúce endorfíny. Enkefalíny sa syntetizujú v hypota-lame, limbických štruktúrach, bazálnych gangliách, mozgovom kmeni,zadných rohoch miechy, periférnych nervoch a nadobličke.Dynorfíny predstavujú najúčinnejšie opioidy. Vyskytujú sa na mno-
hých miestach v mozgu, ako aj periférneho nervového systému. Ichúčinky sú mnohoraké. Dynorfíny regulujú bolesť na úrovni miechy,ovplyvňujú proces príjmu potravy na úrovni hypotalamu a v súčinnostis inými opioidmi regulujú činnosť kardiovaskulárneho systému. Aj počasbolesti spoluúčinkujú pri inhibícii motility gastrointestinálneho traktu.Opioidy všeobecne ovplyvňujú vnímanie bolesti a môžu vyvolať
eufóriu. Príjemné pocity vznikajúce pri jedení čokolády, usmievaní, ma-sáži, meditáciách, počúvaní obľúbenej hudby a dokonca aj príjemné po-city počas orgazmu sa pripisujú práve účinkom uvoľnených endorfínov.Neuróny, ktoré obsahujú rozličné opioidy, sú distribuované v roz-
dielnych oblastiach mozgu. Napríklad neuróny syntetizujúce enkefalíny

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 115
sú lokalizované v šedej hmote v okolí akvaduktu, limbických oblastiacha okolí autonómnych oblastí hypotalamu a mozgového kmeňa. Tietoneuróny vysielajú prevažne krátke axóny, čím ovplyvňujú činnosť oko-litých neurónov. Neuróny syntetizujúce β-endorfín sú lokalizované pre-važne v hypotalame a mozgovom kmeni (nucleus tractus solitarii). Dy-norfín syntetizujúce neuróny sú lokalizované prevažne v hypotalame.Dynorfín a β-endorfín syntetizujúce neuróny vysielajú dlhé axóny na-príklad do hypofýzy, mozgového kmeňa a miechy (Froehlich, 1997). Dy-norfín a enkefalín pôsobia aj ako kotransmitery GABAergických pro-jekčných neurónov v striate (Steiner a Gerfen, 1998).Opioidy sa viažu na 5 základných typov opioidových receptorov,
µ (µ1 a µ2), κ, δ, σ a ε. Endomorfíny (endomorfín 1, endomorfín 2)majú najvyššiu afinitu k µ-opioidovým receptorom v porovnaní so všet-kými známymi opioidmi. Opioidové receptory sú lokalizované pre- ajpost-synapticky.Opioidy sú zapojené v procesoch vzniku látkovej závislosti, percep-
cie bolesti, kognitívnych procesoch, afektívnych reakciách a motorike,ako aj v centrálnej kontrole endokrinných funkcií a modulácii streso-vej odpovede (Reisine, 1995; Zadina a spol., 1999). Majú aj modulačnúúlohu v regulácii činnosti neuronálnych okruhov v predĺženej mieche,ktoré kontrolujú sympatikový tonus a krvný tlak (Guyenet a spol.,2002).
Endomorfíny 1 a 2
Okrem endogénnych opioidových peptidov obsahujúcich Try-Gly-Gly-Phe ako domény pre enkefalíny, endorfíny a dynorfíny existujú ďalšienedávno identifikované peptidy s doménami Tyr-Pro-Phe/Trp opísa-nými ako endomorfín 1 a endomorfín 2. Endomorfín 1 (Tyr-Pro-Trp-Phe-NH2), ako aj endomorfín 2 boli izolované z hovädzieho aj ľudskéhomozgu. Oba peptidy sa viažu s opioidovým µ-receptorom. AktivovanímG-proteínov pôsobia na činnosť gastrointestinálnej motility, antinoci-ceptívne, ďalej pôsobia na vaskulárny systém a pamäť. Ukazuje sa, žeagonisty opioidov so sekvenciou aminokyselín endomorfínov Tyr-Pro-Phe/Trp sú látkami s perspektívnym využitím v liečbe rôznych druhovbolestí (Okada a spol., 2002).

116 NEUROTRANSMITERY
Tachykiníny (neurokiníny)
Rodina tachykinínov zahrňuje niekoľko biologicky aktívnych peptidov,ako sú substancia P (SP), neurokinín A (NKA), neurokinín B (NKB),neuropeptid K (NPK) a neuropeptid γ (predĺžená forma NKA), ktorémajú spoločnú karboxyterminálovú sekvenciu aminokyselinových zvyš-kov Phe-X-Gly-Leu-Met-NH2 (Severini a spol., 2002). Vyskytujú sau stavovcov aj u bezstavovcov. U človeka boli opísané 3 druhy tachy-kinínov, ktoré kódujú dva gény. Jeden kóduje prekurzor, ktorý je pod-kladom pre syntézu SP a NKA, a druhý kóduje prekurzor umožňujúcisyntézu NKB.Tachykiníny vyvolávajú rýchlu kontrakciu hladkého svalstva, na zá-
klade čoho bol odvodený aj ich názov (Snijdelaar a spol., 2000; Beau-jouan a spol., 2004). Pôsobia prostredníctvom receptorov NK1, NK2a NK3. Tachykiníny sa nachádzajú v centrálnom aj periférnom nervo-vom systéme, kde účinkujú ako excitačné neurotransmitery (Pennefat-her a spol., 2004). Predpokladá sa, že tachykiníny modulujú uvoľňova-nie neurotransmiterov z dopamínergických, cholínergických a noradre-nalínergických nervových zakončení. Tachykiníny majú široké spektrumúčinkov v mozgu aj na periférii, kde ovplyvňujú kardiovaskulárny, ge-nitourinárny, respiračný a gastrointestinálny systém. Predpokladá sa ajzapojenie tachykinínov v etiopatogenéze neurodegeneratívnych ocho-rení, depresií a anxiety (Khawaja a Rogers, 1996).Substancia P (SP) je najznámejší tachykinín – peptid zložený z 11
aminokyselinových zvyškov. Viacero prelomových objavov týkajúcichsa štúdia neuropeptidov je spojených práve so SP. Už roku 1932 sadokázal jej výskyt v gastrointestinálnom trakte, ako aj v mozgu. Jeprvým neuropeptidom, ktorého receptor už bol klonovaný. Aj prvouklinicky účinnou látkou, ktorá pôsobí prostredníctvom neuropeptido-vého receptora v CNS, je antagonista substancie P (Hokfelt a spol.,2001b). Substancia P sa viaže na tachykinínový NK1-receptor spria-hnutý s G-proteínom (Snijdelaar a spol., 2000).V CNS sa účinok SP spája s mnohými procesmi, ako sú poruchy
nálady, strach, stres, neurogenéza, neurotoxicita, vracanie a prenos in-formácií z nociceptorov (Holmes a spol., 2003). Predpokladá sa aj účasťSP v zápalových procesoch, v gastrointestinálnych a respiračných pro-cesoch. Je aj potenciálnym vazodilatátorom.

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 117
Substancia P sa nachádza napríklad aj v zadných rohoch miechy(prevažne v oblastiach, kde sa nachádzajú zakončenia axónov primár-nych aferentných neurónov), ako aj v ďalších oblastiach CNS, ktoré súzapojené do prenosu nociceptívnych signálov, medzi ktoré patria šedáhmota v okolí akvaduktu, nucleus raphe magnus, nucleus reticularisgigantocellularis a talamus (Snijdelaar a spol., 2000).Substancia P sa vyskytuje aj v neurónoch syntetizujúcich mono-
amíny. Odhaduje sa, že približne 50% sérotonínergických neurónovv nucleus raphe dorsalis syntetizuje aj SP.Neurokinín A, neurokinín B a neuropeptid K sú schopné modulo-
vať aktivitu hypotalamo-hypofýzového systému. Všetky 3 neuropeptidyaktivujú tento systém v určitej koncentračnej závislosti. Účinok NKBna sekréciu CRH je však malý. NKB je účinnejší ako NKA a účinokoboch týchto neuropeptidov na ACTH je zablokovateľný antagonistamiCRH-receptorov. Tieto reakcie po podaní NKA a NPK jasne svedčiao ich potenciálnom vplyve nielen na centrálnu, ale aj na periférnu zložkuHPA-systému.
Peptidy príbuzné s NPY
Neuropeptid Y (NPY) sa skladá z 36 aminokyselín a je jeden z naj-rozšírenejších neuropeptidov v mozgu so širokou paletou fyziologickýchúčinkov (Colmers a Bahh, 2003). Patrí do rodiny pankreatických poly-peptidov, ktorá zahŕňa aj pankreatický polypeptid (PP) a peptid YY(PYY). NPY sa zúčastňuje na mnohých regulačných procesoch týka-júcich sa cirkadiánnej rytmicity, sexuálnych funkcií, anxiety, stresovejreakcie, periférnej vaskulárnej rezistencie, kontraktility srdca, reguláciechuti do jedla, príjmu potravy a karbohydrátovej preferencie a regulá-cie množstva telesného tuku a vzniku obezity. Predpokladá sa, že kon-centrácia NPY v hypotalame odráža nutričný stav organizmu (Wynnea spol., 2005).NPY sa vyskytuje v CNS aj v autonómnom nervovom systéme.
NPY sa syntetizuje v mnohých oblastiach mozgu, medzi ktoré patrialocus coeruleus, hypotalamus, corpus amygdalae, hipokampus, nucleusaccumbens a neokortex. Hlavnou oblasťou hypotalamu, ktorá synteti-zuje NPY, je nucleus arcuatus. V mozgu účinkuje hlavne ako neuro-transmiter. NPY je syntetizovaný ako kotransmiter v neurónoch synte-tizujúcich noradrenalín, GABA, somatostatín a agouti-related peptid.

118 NEUROTRANSMITERY
NPY účinkuje prostredníctvom heterogénnych receptorov spriahnutýchs G-proteínom.Vmozgu bolo identifikovaných 5 podtypov NPY-receptorov (Y1–Y5).
Y2-receptory môžu účinkovať ako presynaptické autoreceptory, ktorýchobsadenie znižuje ďalšie uvoľňovanie NPY (Wynne a spol., 2005). U člo-veka sú početné fyziologické účinky tohto neuropeptidu sprostredkú-vané receptormi typu Y1, Y2, Y4 a Y5. Centrálne aktívne antagonistyreceptorov Y1 a Y5 majú potenciálny terapeutický význam v liečbeobezity. Agonistické a antagonistické ligandy pre receptory typu Y2sú perspektívnymi antikonvulznými, antidepresívnymi a analgetickýmilátkami.Predpokladá sa účasť NPY v záchvatových procesoch, vzniku lát-
kovej závislosti a v afektívnych poruchách (Holmes a spol., 2003; Ped-razzini a spol., 2003; Heilig, 2004).Peptid YY 3-36 (PYY3-36) obsahuje 34 aminokyselinových zvyš-
kov, pričom mnohé z nich sa nachádzajú v rovnakej pozícii, ako je tov prípade neuropeptidu Y. Účinky peptidu YY3-36 v porovnaní s ne-uropeptidom Y sú však často opačné, teda peptid YY3-36 pôsobí akoinhibítor príjmu potravy a jeho obsah sa zvyšuje v tráviacom traktehneď po jedle. Množstvo secernovaného peptidu v tráviacom trakte nieje konštantné, ale narastá so stúpajúcou hodnotou prijatých kalórií.Supresia chuti do jedla spôsobená PYY3-36 je pomalšia v porovnaní
s cholecystokinínom, ale rýchlejšia v porovnaní s leptínom. PYY3-36pôsobí na úrovni hypotalamu ako supresor chuti do jedla, kým v pan-krease zvyšuje exokrinnú sekréciu tráviacich štiav a v žlčníku stimulujesekréciu žlče.
Peptidy zo skupiny sekretín-VIP-glukagón
Glukagónu podobné peptidy 1 a 2
Glukagónu podobné peptidy 1 (GLP-1) a 2 (GLP-2) sú deriváty prepro-glukagónovej molekuly a sú zapojené do mnohých periférnych procesovspojených s homeostázou glukózy, príjmom potravy, vyprázdňovanímobsahu žalúdka a sekréciou inzulínu. Preproglukagón bol identifikovanýaj v nervových bunkách mozgu v nucleus tractus solitarii. GLP-1 ner-vové dráhy a GLP-1-receptory sa vyskytujú najmä v mediálnom hypo-talame. Centrálne aplikovaný GLP-1 stimuluje vo veľkej miere neurónyv nucleus supraopticus hypothalami, nucleus paraventricularis hypo-

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 119
thalami a nucleus amygdaloideus centralis a v menšej miere nucleusarcuatus. Injekcia GLP-1 priamo do nucleus paraventricularis hypotha-lami vyvoláva anorexiu, ale nevyvoláva chuťovú averziu. Predpokladása, že GLP-1 má špecifickú úlohu v regulácii príjmu potravy a tekutínaj v termoregulácii a regulácii krvného tlaku (Blazquez a spol., 1998;Acuna-Goycolea a van den Pol, 2004).GLP-2 sa vyskytuje v CNS v kolokalizácii s GLP-1. Najvyššie kon-
centrácie GLP-2 sa zistili vo ventrálnej časti nucleus dorsomedialishypothalami. Na rozdiel od rozsiahleho výskytu GLP-1-receptorovejmRNA sa však GLP-2-receptorová mRNA vyskytuje hlavne v kom-paktnej časti nucleus dorsomedialis hypothalami, ktorého neuróny súvýrazne aktivované po centrálnom podaní GLP-2. GLP-2 aplikovanýdo bočných mozgových komôr spôsobuje signifikantný pokles v príjmepotravy. Zatiaľ nie je jasné, aký majú tieto peptidy vplyv na procesyspojené so vznikom obezity (Tang-Christensen a spol., 2001).
Vazoaktívny intestinálny polypeptid (VIP) a histidínovýizoleucínový peptid (PHI)
Peptidy VIP a PHI lokalizované u človeka v subpopuláciách neuró-nov nucleus paraventricularis hypothalami sú pravdepodobne zapo-jené do regulácie akútnej sekrécie prolaktínu (PRL) v podmienkachstresu. Tento proces je však kontrolovaný aj sérotonínergickým systé-mom s neurónmi v nucleus raphae dorsalis, ktoré vysielajú axóny dohypotalamu. Na druhej strane je akútna sekrécia prolaktínu počas lak-tácie pravdepodobne len čiastočne pod kontrolou VIP/PHI-peptidov.Okrem hypotalamických zložiek zapojených do regulácie sekrécie PRLbol opísaný aj výskyt VIP-buniek v hypofýze. Táto skupina VIP-buniekreaguje výhradne na zmeny stavu tyreoidovej osi a je stimulovaná hy-potyreózou. Fyziologický význam VIP/PHI-peptidov hypofýzového pô-vodu je zatiaľ u človeka nejasný (Reichlin, 1988).VIP/PHI-peptidy sú syntetizované aj vo ventrolaterálnej časti hy-
potalamického jadra nucleus suprachiasmaticus, pričom sú oba peptidykolokalizované v tých istých neurónoch (Card a spol., 1988). Predpo-kladá sa, že sa zúčastňujú na regulácii cirkadiánnych procesov.V posledných rokoch sa pripisuje VIP aj neuroprotektívny vplyv.
Pre jeho protizápalové, protiapoptotické a neurotrofické vplyvy sa uva-

120 NEUROTRANSMITERY
žuje o využití VIP v liečbe niektorých neurodegeneratívnych ochorení,ako je napr. Parkinsonova choroba (Gonzalez-Rey a spol., 2005).
Hypofýzovú adenylátcyklázu stimulujúci polypeptid
Hypofýzovú adenylátcyklázu stimulujúci polypeptid (pituitary adeny-late cyclase-activating polypeptide, PACAP) bol pôvodne izolovanýz ovčieho mozgu roku 1989, ako hormón, ktorý aktivuje adenylátcyklázuv hypofýze. Tento neuropeptid existuje v dvoch formách, PACAP38 zlo-žený z 38 aminokyselinových zvyškov a PACAP27 zložený z 27 amino-kyselinových zvyškov. Predpokladá sa, že PACAP pôsobí v centrálnomaj periférnom nervovom systéme. Uvažuje sa o jeho zapojení v regulá-cii hypotalamickej neurosekrécie, homeostatickej kontrole cirkadiánnychrytmov, v modulácii správania, pamäťových procesov a učenia. Okremtoho má pravdepodobne neuroprotektívny vplyv (anti-apoptotický úči-nok) a je zapojený v odpovedi na poškodenie a zápal, ako aj v ontoge-netických a diferenciačných procesoch v nervovom systéme (Waschek,2002; Zhou a spol., 2002).
Ďalšie neuropeptidy
Adrenomedulín
Ľudský adrenomedulín (adrenomedullin, ADM) je peptid zložený z 52aminokyselinových zvyškov pôvodne izolovaný z feochromocytómu.Patrí do skupiny peptidov, ktoré zahŕňajú amylín, kalcitonín a CGRP(Wimalawansa, 1997).ADM je syntetizovaný v nadobličkách, obličkách, pľúcach, srdci, sle-
zine, tenkom čreve, slinných žľazách a mozgu. Podanie ADM vyvolávana periférii vazodilatáciu a v srdci pôsobí pozitívne inotropne a chrono-tropne. Predpokladá sa účasť ADM na regulácii elektrolytového meta-bolizmu účinkom na centrálne a periférne štruktúry. ADM vyvoláva in-hibíciu sekrécie aldosterónu z nadobličiek a v obličkách zvyšuje diurézua nátriurézu (Serrano a spol., 2002; Taylor a Samson, 2002; Beltowskia Jamroz, 2004).Neuróny syntetizujúce ADM sa nachádzajú vo všetkých oblastiach
CNS. Hlavným miestom lokalizácie ADM neurónov sú však oblasti zú-častňujúce sa na regulácii činnosti autonómneho nervového systému –nucleus interstitialis striae terminalis, septum, corpus amygdaloideum,

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 121
hypotalamus (napr. nucleus paraventricularis hypothalami) a subforni-kálny orgán. V mozgovom kmeni ADM syntetizujú neuróny locus co-eruleus, šedá hmota v okolí akvaduktu, nucleus parabrachialis, nucleiraphes, nucleus tractus solitarii, ventrolaterálna predĺžená miecha, nuc-leus dorsalis nervi vagi, nucleus ambiguus a area postrema.Lokalizácia neurónov syntetizujúcich ADM, receptorov pre ADM,
ako aj experimentálne údaje poukazujú na to, že ADM pravdepodobnemoduluje autonómne a neuroendokrinné funkcie (Shan a spol., 2003).Predpokladá sa, že aj ADM sa zúčastňuje na modulácii cirkulácie krviv mozgu v reakcii na ischémiu a má úlohu pri vývoji mozgovo-cievnejbariéry (Hinson a spol., 2000; Kis a spol., 2003). Uvažuje sa, že v srdcisyntetizovaný ADM môže mať úlohu v patofyziológii hypertenzie, srd-covej ischémie, hypertrofie a zlyhaní srdca (Cheung a spol., 2004; Nis-hikimi a Matsuoka, 2005).
Agouti podobný proteín
Agouti podobný proteín (agouti-related peptide, AGRP) je zložený zo132 aminokyselinových zvyškov a je syntetizovaný v neurónoch nuc-leus arcuatus v hypotalame v kolokalizácii s neuropeptidom Y (Haskell-Luevano a spol., 1999). Účinok AGRP je sprostredkovaný melanokortí-novými receptormi MCR3 a MCR4 (Ollmann a spol., 1997). AGRP jevýznamne zapojený do procesov regulujúcich príjem potravy a pri cen-trálnej infúzii stimuluje príjem potravy u hlodavcov a primátov (Rossia spol., 1998; Haskell-Luevano a spol., 1999; Wilson a spol., 1999). Patrímedzi najúčinnejšie orexigénne neuropeptidy, ktoré zvyšujú chuť dojedla a znižujú metabolický výdaj. AGRP mRNA sa zvyšuje pri ne-dostatku energie, keď je inhibovaná aj reprodukčná os (Hahn a spol.,1998).
Amylín
Amylín (AMY) je polypeptid pankreatického pôvodu zložený z 37 ami-nokyselinových zvyškov. Bol identifikovaný v štruktúrach mozgu ajv periférnom nervovom systéme. Bunky produkujúce amylín sa na-šli vo viacerých štruktúrach mozgu, ako sú vestibulárne, cerebelárne,trapezoidné a kochleárne jadrá a mezencefalické jadro trigeminálnehonervu. AMY obsahujúce bunkové telá boli opísané v laterálnych, gigan-tocelulárnych a magnocelulárnych jadrách retikulárnej formácie. Mnohé

122 NEUROTRANSMITERY
z nich sú pravdepodobne regulované katecholamínergickými neurónmi.Početné AMY-imunoreaktívne nervové vlákna sa vyskytujú v tractustrigeminalis spinalis, nucleus tractus solitarii a area postrema. Pred-pokladá sa, že amylín môže ovplyvňovať niektoré senzorické procesy(D’Este a spol., 2000, 2001). Centrálna aj periférna aplikácia AMYvyvoláva u potkanov anorektické správanie. Uvažuje sa, že jeho úlohav regulácii príjmu potravy by mohla byť sprostredkovaná prostredníc-tvom histamínergického systému.
Angiotenzín II
Angiotenzín II je peptid zložený z 8 aminokyselín, ktorý môže pôso-biť systémovo (cirkulujúci) a lokálne na okolité bunky tkanív v miestejeho syntézy (tkanivový). Cirkulujúci angiotenzín II má široké spek-trum účinkov, medzi ktoré patria napríklad regulácia cievnej rezisten-cie, modulácia sekrécie aldosterónu, ako aj na modulácii činnosti CNS.Tkanivový angiotenzín II sa významne zúčastňuje na modulácii činnostivnútorných orgánov (napr. srdce, obličky a mozog). Ovplyvňuje aj rasta proliferáciu buniek (Ferguson a spol., 2001). Z klinického hľadiska jevýznamný prospešný účinok antagonistov receptorov pre angiotenzínII, ktorý sa využíva napríklad na liečbu hypertenzie a na redukciu po-škodenia obličiek pri stavoch spojených s proteinúriou (Appel a Appel,2004).Cirkulujúci angiotenzín ovplyvňuje prostredníctvom cirkumventri-
kulárnych orgánov činnosť CNS (Ganong, 2000). Okrem periférie bolasyntéza angiotenzínu, ako aj prítomnosť receptorov AT1, AT2 a AT4dokázaná aj v mozgu (Allen a spol., 1998; de Gasparo a spol., 2000).Predpokladá sa, že angiotenzín syntetizovaný v mozgu pôsobí prostred-níctvom AT-receptorov ako neurotransmiter alebo neuromodulátor ner-vových dráh, ktoré sa zúčastňujú na regulácii krvného tlaku, na udr-žiavaní homeostázy tekutín a iónov a zasahuje aj do termoregulácie.Uvažuje sa aj o jeho účasti v regulácii syntézy proteínov, uvoľňovaniaprostaglandínov, regulácii syntézy a uvoľňovania hypofýzových hormó-nov, ako aj v pamäťových procesoch a procesoch učenia (Fitzsimons1998; Ferguson a spol., 2001; McKinle a spol., 2001, 2003).

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 123
Apelín
Apelíny môžu byť zložené z 12, 13 a 36 aminokyselinových zvyškov.Peptidy zo skupiny apelínov sú syntetizované hlavne v periférnychtkanivách (napr. pľúca, srdce, obličky, cievy, mliečna žľaza). Účinkujúprostredníctvom jedného receptora APJ spojeného s G-proteínom. Ichhlavný fyziologický význam nebol zatiaľ určený. Prítomnosť neurónovsyntetizujúcich apelín sa dokázala v nucleus paraventricularis hypotha-lami, nucleus supraopticus hypothalami, nucleus suprachiasmaticus hy-pothalami a v akcesórnych jadrách hypotalamu potkana (Reaux a spol.,2001; Brailoiu a spol., 2002). V týchto jadrách je apelín kolokalizovanýs vazopresínom a predpokladá sa, že lokálne inhibuje sekréciu vazopre-sínergických neurónov, a tak pôsobí opačne ako vazopresín, teda diure-ticky (Llorens-Cortes a spol., 2005). Výrazné apelín-pozitívne imunore-aktívne dráhy a nervové výbežky boli identifikované aj v oblasti eminen-tia mediana, v stopke hypofýzy a prednom a zadnom laloku hypofýzy.Predpokladá sa, že apelíny sú zapojené v regulácii kardiovaskulárnychfunkcií, v udržiavaní homeostázy tekutín a modulácii imunitných fun-kcií (Kleinz a Davenport, 2005).
Galanín a „galanin-likeÿ peptid
Galanín (GAL), peptid obsahujúci 29–30 aminokyselín, je syntetizovanýv noradrenalínergických neurónoch locus coeruleus a sérotonínergickýchneurónoch nucleus raphe dorsalis, ako aj v hypotalame (napr. nucleusparaventricularis). Dokázalo sa, že galanín pôsobí ako inhibičný modu-látor. V súčasnosti sú známe tri podtypy receptorov pre galanín (GAL1,GAL2 a GAL3), ktoré sú spojené s G-proteínom.Výsledky z experimentálnych pozorovaní naznačujú, že galanín
môže ovplyvňovať kognitívne procesy, príjem potravy, sexuálne sprá-vanie, záchvaty, regeneráciu nervových vlákien, nociceptívne procesya stresovú odpoveď (Holmes a spol., 2003).Okrem galanínu bol v mozgu identifikovaný aj štruktúrne podobný
peptid, ktorý patrí do rovnakej rodiny neurotransmiterov a označujesa ako galanin-like peptide (GALP). GALP sa viaže na receptory pregalanín (Gundlach a spol., 2001). Neuróny syntetizujúce GALP komu-nikujú s ďalšími neuropeptidergickýmu neurónmi, ktoré sú zapojené doregulácie príjmu potravy. Predpokladá sa, že GALP ovplyvňuje aj re-produkčné funkcie prostredníctvom vplyvu na gonadotropín uvoľňujúci

124 NEUROTRANSMITERY
hormón (GnRH) syntetizujúce neuróny, čím môže vytvárať spojeniemedzi metabolickými a reprodukčnými procesmi (Cunningham, 2004;Gottsch a spol., 2004; Kageyama a spol., 2005).
Ghrelín
Ghrelín je hormón zložený z 28 aminokyselinových zvyškov, ktorý jeprodukovaný prevažne bunkami výstelky žalúdka a len v malom množ-stve je syntetizovaný v hypotalame (v nucleus arcuatus hypothalami).Je to typický gastrointestinálny hormón, ktorého hladiny stúpajú predpríjmom potravy, teda v stave hladu, a klesajú po príjme potravy, resp.nasýtení. Je tzv. „signalizátorom hladuÿ na úrovni hypotalamu a máopačné účinky ako leptín, PYY3-36 a obestatín. Tieto neuropeptidyvyvolávajú pocit nasýtenosti, keď sú v cirkulácii prítomné vo vysokýchhladinách. Ghrelín stimuluje aj sekréciu rastového hormónu z adenohy-pofýzy. Má aj úlohu v regulácii spánku, termoregulácii a v moduláciičinnosti autonómneho nervového systému (Rosicka a spol., 2002; Kor-bonits a spol., 2004; Davenport a spol., 2005).Ghrelínové receptory sa nachádzajú hlavne v membránach neuró-
nov nucleus arcuatus hypothalami a nucleus ventromedialis hypotha-lami. Ghrelín má významnú úlohu v neurotrofických procesoch hlavnev hipokampe a je esenciálny pre kognitívnu adaptáciu na meniace sa en-vironmentálne podmienky a v procesoch spojených s učením. Pacientis anorexiou mávajú vysoké hladiny ghrelínu. Hladiny ghrelínu sú vy-soké aj pri kachexii indukovanej nádormi (Garcia a spol., 2005). Bypa-sová operácia žalúdka redukuje aj hladiny ghrelínu (Cummings a spol.,2002).Štúdie na zvieratách preukázali, že ghrelín vstupuje do hipokampu
z krvného riečiska a následne môže stimulovať procesy učenia a pamäte.Vzhľadom na túto situáciu sa zdá, že učenie môže byť najefektívnejšiepočas dňa s prázdnym žalúdkom a vysokými hladinami ghrelínu. Z tejistej prekurzorovej molekuly mRNA, z ktorej sa syntetizoval ghrelín, sasyntetizuje aj neuropeptid nazvaný obestatín. Obestatín, ktorý objaviliroku 2005, sa skladá z 23 aminokyselinových zvyškov. Jeho účinky súopačné v porovnaní s účinkami ghrelínu (Zhang a spol., 2005). Pred-pokladá sa, že z jednej prekurzorovej molekuly (peptid obsahujúci 117aminokyselín) sa odštiepi jedna molekula ghrenínu a obestatínu, kto-rých účinok by sa mal navzájom rušiť. Ide však o neaktívne molekuly,

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 125
ktoré sa stávajú aktívnymi až po ďalšej chemickej modifikácii. Pred-pokladá sa preto, že práve v tomto procese aktivácie molekuly môžupôsobiť regulačné mechanizmy.
Cholecystokinín
Cholecystokinín (CCK) je peptid gastrointestinálneho systému a bolprvým peptidom, ktorý priamo podporil koncept obojsmernej peptider-gickej komunikácie medzi gastrointestinálnym traktom a CNS. CCK mániekoľko foriem, CCK58, CCK 33 a CCK8, pričom CCK8 je najaktív-nejší. CCK pôsobí na dva typy receptorov – CCKA a CCKB spriahnutés G-proteínmi (Alexander a spol., 2006).CCK je secernovaný mukózou duodenálnej časti tenkého čreva, keď
chymus bohatý na tuky alebo bielkoviny opúšťa žalúdok a vstupuje doduodéna. CCK reguluje uvoľňovanie pankreatických enzýmov, ako súlipáza, amyláza, trypsín a chymotrypsín a zároveň vyvoláva kontrakciužlčníka a relaxáciu Glissonovho sfinktera. Pôsobí ako supresor hladu.Ako neuropeptid syntetizovaný v CNS nastavuje úroveň sýtosti pro-
stredníctvom CCK-receptorov. Predpokladá sa, že CCK syntetizovanýv nervovom systéme je zapojený nielen do regulácie príjmu potravy,ale aj do pamäťových procesov. Uvažuje sa aj o jeho účasti pri vznikuanxiety (Hansen a Nielsen, 2001). Aplikovanie CCK človeku spôsobujenauzeu a anxietu. Prítomnosť CCK v nervových zakončeniach a teláchneurónov v zadných rohoch miechy poukazuje na jeho úlohu v pre-nose senzorických informácií (Hokfelt a spol., 2001a). Navyše sa uvažujeo možných neuroprotektívnych účinkoch CCK, ako aj o jeho modulač-nom vplyve na činnosť HPA-systému (Beinfeld, 2001). Má významnúúlohu aj v tolerancii na drogy, ako sú morfium a heroín, a zodpovedá ajza vznik svalovej bolesti v prípade nútenej abstinencie opioidov. ÚčinkyCCK sú veľmi variabilné v rámci toho istého druhu. Napríklad u potka-nov CCK redukuje hlad len u mladých jedincov, menej účinný je u star-ších zvierat a ešte menej účinný u samíc. Jeho chuť do jedla potlačujúciefekt je u obéznych potkanov takmer zanedbateľný (Fink a spol., 1998).
Chromograníny
Chromograníny sú polypeptidy široko distribuované v centrálnom ner-vovom systéme. Sú uskladnené v denzných vezikulách nervových za-končení. Rozlišujú sa chromogranín A, chromogranín B a sekretoneurín

126 NEUROTRANSMITERY
(Pirker a spol., 2001). Predpokladá sa účasť chromogranínov v etiopa-togenéze temporálnej epilepsie a Alzheimerovej choroby (Kandlhofera spol., 2000; Lechner a spol., 2004).Sekretoneurín obsahuje 33 aminokyselinových zvyškov. Je to pep-
tid výrazne fylogeneticky konzervovaný, ktorý sa vyskytuje u cicav-cov, plazov, vtákov, obojživelníkov a rýb. Je exprimovaný v endokrin-ných, neuroendokrinných a nervových tkanivách. Prekurzorovou mole-kulou je látka označovaná sekretogranín II, ktorá je zapojená v mo-dulácii dopamínergického prenosu v striate, regulácii činnosti sympati-kového systému, ale aj v modulácii zápalových procesov (Wiedermann,2000). Za patologických podmienok sa sekretoneurín produkuje aj v ne-endokrinných tkanivách, ako sú svaly alebo pneumocyty aj v tumo-roch epitelových tkanív. Sekretoneurín sa zúčastňuje aj na uvoľňovanídopamínu z preparátov striata a ovplyvňuje rast neuritov v cerebele.Prispieva k neurogénnemu zápalu a môže mať úlohu pri ischemickýchochoreniach v CNS (Fischer a spol., 2005).
Kalcitonínovému génu príbuzný peptid
Kalcitonínovému génu príbuzný peptid (calcitonin gene-related peptide,CGRP) patrí do rodiny kalcitonínových biologicky aktívnych pepti-dov spolu s kalcitonínom, amylínom, a adrenomedulínom. Skladá saz 37 aminokyselín a vyskytuje sa v dvoch odlišných formách, CGRPαa CGRPβ. Obom peptidom sa pripisujú podobné funkcie a oba peptidysa vyskytujú v centrálnom aj periférnom nervovom systéme. Pôsobiaprostredníctvom 2 typov receptorov CGRP1 a CGRP2. Predpokladása, že CGRP má úlohu v prenose signálov v mieche (napr. prenos no-ciceptívnych a motorických informácií). Okrem toho sa uvažuje o jehoúčasti v pamäťových procesoch, regulácii príjmu potravy a v modu-lácii uvoľňovania rastového a luteinizačného hormónu. CGRP má vý-znamný vplyv na aktivitu kardiovaskulárneho systému. Predpokladá saaj jeho zapojenie v indukcii hypertermie, katalepsie a migrény (Edvins-son a Uddman, 2004; Dobolyi a spol., 2005).
Kokaínom a amfetamínom regulovaný transkript
Neuropeptid označovaný ako kokaínom a amfetamínom regulovanýtranskript (cocaine- and amphetamine-regulated transcript, CART) je

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 127
produkovaný neurónmi hypotalamu. Prítomnosť CART mRNA sa ur-čila v mnohých oblastiach hypotalalamu, napríklad nucleus supraopti-cus, paraventricularis, periventricularis, arcuatus, area hypothalamicalateralis. Detegoval sa však aj mimo hypotalamu – v zona incerta.CART je kolokalizovaný s ďalšími neuropeptidmi, ako sú vazopresín,oxytocín, galanín a somatostatín (Vrang a spol., 1999). Predpokladása, že neuróny syntetizujúce CART sú zapojené do centrálnej reguláciepríjmu potravy (Vrang a spol., 1999) a do regulácie aktivity HPA osi(Vrang a spol., 2002)
Melanín koncentrujúci hormón
Melanín koncentrujúci hormón (melanin-concentrating hormone, MCH)je syntetizovaný hlavne v neurónoch laterálneho hypotalamu a subta-lamu, ktoré inervujú rozsiahle oblasti mozgu (Knigge a spol., 1996).MCH je pravdepodobne zapojený do viacerých procesov spojených s re-guláciou príjmu potravy, ako je modulácia chuti do jedla, správania spo-jeného s vyhľadávaním potravy, pohybom žuvacích svalov a kontrolouenergetickej rovnováhy (Kawano a spol., 2002). Okrem toho sa pred-pokladá účasť MCH v modulácii reprodukčných procesov a stresovejreakcie (Griffond a Baker, 2002). Z prekurzora pre MCH môže byť syn-tetizovaný aj neuropeptid EI a neuropeptid GE, ktorých funkcia nie jepresne charakterizovaná (Borowsky a spol., 2002).
Mozgový nátriuretický peptid
Atriálny nátriuretický peptid (ANP) objavili roku 1983. Dokázalo sa,že ANP sa uvoľňuje z pravej predsiene srdca počas jej dilatácie. Neskôrsa dokázal výskyt peptidu podobného ANP vo viacerých oblastiach hy-potalamu. Tento peptid bol označený ako mozgový nátriuretický peptid(brain natriuretic peptide, BNP). BNPergické neuróny sa rozprestierajúrostrálne od organum subfornicale a organum vasculosum laminae ter-minalis až po nucleus paraventricularis hypothalami. BNPergické neu-róny sa zúčastňujú na regulácii vodného hospodárstva a príjmu soli,ako aj na modulácii pocitu smädu. BNPergické neuróny vysielajú svojeaxóny do oblasti eminentia mediana a zadného laloka hypofýzy. BNP samôže preto uvoľniť do predného laloka hypofýzy, ako aj do systémovejcirkulácie (McCann a spol., 1997). Viaceré údaje poukazujú na to, že

128 NEUROTRANSMITERY
nátriuretické peptidy môžu slúžiť ako markery zlyhania srdca (Dousta spol., 2004).
Neuropeptid B
Neuropeptid B (NPB) je len nedávno identifikovaný ligand pre GPR7a GPR8 receptory spriahnuté s G-proteínom. GPR7-receptory sa vysky-tujú v rôznych oblastiach mozgu, hlavne v hypotalame, čo naznačuje, žeNPB môže mať významnú úlohu v regulácii rôznych neuroendokrinnýchprocesov regulovaných hypotalamom. Reguluje aj prenos signálov vede-ných k adenohypofýze, ale priamo na úrovni adenohypofýzy nepôsobí,aspoň nie, pokiaľ ide o reguláciu vyplavovania prolaktínu, rastovéhohormónu alebo adrenokortikotropného hormónu (Dun a spol., 2005).NPB je faktor, ktorý má tiež úlohu v zápalových procesoch.V hypotalame boli NPB bunky identifikované v area preoptica me-
dialis, nucleus preopticus ventromedialis, area retrochiasmatica, nuc-leus paraventricularis hypothalami, nucleus supraopticus hypothalami,hypothalamic accessory nuclei, nucleus periventricularis hypothalami,area hypothalamica lateralis, nucleus dorsomedialis hypothalami, areahypothalamica posterior a zona incerta. Menší výskyt NPB bunkovýchtiel sa nachádza v nucleus arcuatus hypothalami a hustá spleť nervo-vých vlákien v eminentia mediana.V strednom mozgu boli NPB telá opísané v substantia nigra, nucleus
paranigralis, area tegmentalis ventralis, nucleus interfascicularis a nucleiraphae. Výraznú NPB imunoreaktivitu preukazovali aj neuróny v nuc-leus Edinger-Westphal.
Neuropeptid FF
Neuropeptid FF (NPFF), neuropeptid AF (NPAF) a neuropeptid SFsú príbuzné neuropeptidy, o ktorých sa predpokladá, že sú zapojenév regulácii širokého spektra funkcií CNS (Vilim a spol., 1999). NPFF,spoločne s NPAF sa vyskytuje vo vysokej koncentrácii v zadnom la-loku hypofýzy, mieche, hypotalame a mozgovom kmeni. Predpokladása účasť NPFF v pamäťových procesoch, regulácii činnosti autonóm-neho a endokrinného nervového systému (Panula a spol., 1996; Jha-mandas a spol., 2001). NPFF sa zúčastňuje aj na modulácii prenosunociceptívnych informácií na úrovni miechy a mozgu pravdepodobne aj

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 129
prostredníctvom interakcie s opioidovým systémom (Roumy a Zajac,1998; Panula a spol., 1999; Mollereau a spol., 2005).
Neuropeptid S
Neuropeptid S (NPS) je nedávno identifikovaný bioaktívny peptid.Predpokladá sa, že je zapojený v modulácii procesov spánku a bde-nia, ale aj anxiózneho správania. NPS zvyšuje bdelosť a potláča všetkyštádiá spánku. Hlavným miestom syntézy NPS sú oblasti nachádzajúcesa v blízkosti locus coeruleus, štruktúry zapojenej do regulácie spánkua bdenia, ako aj anxiózneho správania (Koob a Greenwell, 2004; Rein-scheid a Xu, 2005a, 2005b).
Neuropeptid W
Neuropeptid W (NPW) je endogénny ligand viažuci sa v CNS na recep-tory GPR7 and GPR8 spojené s G-proteínom. Vyskytuje sa v dvochformách – ako peptid obsahujúci 23 (NPW-23) alebo 30 (NPW-30) ami-nokyselinových zvyškov. Obe formy neuropeptidov pripravených synte-ticky sa viažu a aktivujú oba typy receptorov v podobnej koncentrácii(Shimomura a spol., 2002). GPR-7 mRNA bola detegovaná v hypota-lame a hipokampe (Fujii a spol., 2002). Intracerebroventrikulárne poda-nie NPW-23 zvyšuje u potkanov príjem potravy. Predpokladá sa účasťNPW v regulácii uvoľňovania prolaktínu, rastového hormónu, moduláciipríjmu potravy, regulácii činnosti neuroendokrinného systému a modu-lácii stresovej odpovede (Shimomura a spol., 2002; Baker a spol., 2003;Mondal a spol., 2003; Niimi a Murao, 2005).
Neurotenzín
Neurotenzín (NT) je peptid obsahujúci 13 aminokyselinových zvyškov.Predpokladá sa, že je zapojený v regulácii činnosti nervového aj endo-krinného systému (Dobner, 2005).NT preukazuje výraznú heterogénnu distribúciu. Najvyššie hladiny
NT sa zistili v hypotalame hlavne jeho posteriórnej časti aj v mamilár-nej oblasti. Aj substantia nigra, ventrálna tegmentálna oblasť, centrálnejadro amygdalárneho komplexu, dorzálny hipokampus, nucleus accum-bens, septum a globus pallidus obsahujú pomerne významné množstvoNT. Telá neurónov obsahujúcich NT sa nachádzajú takmer vo všetkých

130 NEUROTRANSMITERY
uvedených oblastiach. NT je kolokalizovaný s mnohými inými látkami,ako sú napr. dopamín a CCK v dopamínergických neurónoch ventrálnejtegmentálnej oblasti a vyskytuje sa aj v dopamínergických neurónochnucleus arcuatus hypothalami. V mozgu bolo opísaných niekoľko sys-témov NT dráh. Projekcie neurónov nucleus arcuatus hypothalami dooblasti eminentia mediana zabezpečujú uvoľňovanie NT do portálnehosystému hypofýzy. Ďalšie dráhy boli opísané z centrálneho jadra amyg-daly do parabrachiálneho jadra, zo subikula do hipokampu a z nucleustractus solitarii do nucleus accumbens atď. Najdetailnejšie je charak-terizovaná mezolimbickokortikálna NT dráha vedúca z ventrálnej teg-mentálnej oblasti do frontálneho kortexu a nucleus accumbens.Uvažuje sa, že neuróny uvoľňujúce neurotenzín ako neurotransmi-
ter modulujú činnosť buniek syntetizujúcich gonadoliberíny, dopamín,somatostatín a kortikoliberín (Rostene a Alexander, 1997). NT sa pova-žuje za regulátor sekrécie luteinizačného hormónu a prolaktínu. Pred-pokladá sa, že neurotenzín môže mať úlohu v mechanizmoch účinkuantipsychotík a psychostimulačných látok (Dobner a spol., 2003). Pod-poruje to aj fakt, že neuróny syntetizujúce neurotenzín sú lokalizovanév oblastiach, o ktorých sa predpokladá, že sú zapojené v etiopatogenézeschizofrénie (Kinkead a spol., 1999). Študuje sa aj interakcia medzi neu-rotenzínom a dopamínergickým systémom, ktorého narušená činnosť jepodkladom klinických prejavov Parkinsonovej choroby a schizofrénie(Binder a spol., 2001; Seutin, 2005). Hromadia sa dôkazy o úlohe ne-urotenzínu v stresom indukovanej analgézii (Dobner, 2005). Situáciuvšak komplikuje fakt, že mnohé účinky NT závisia od faktorov, akosú pohlavie, spôsob a miesto aplikácie, psychický stav, alebo spoločnáaplikácia iných látok.
Nociceptín/orfanín (N/OFQ)
Nociceptín (N/OFQ) je peptid zložený zo 17 aminokyselinových zvyš-kov, ktorý je podobný dynorfínu A. Bol identifikovaný roku 1995 (Hein-richer, 2003). Je endogénnym ligandom pre receptor 1 (ORL 1, dnesoznačovaný ako NOP), ktorý nie je citlivý na naloxon a je podobnýopioidovému receptoru 1. Hoci štruktúra N/OFQ sa veľmi podobá dy-norfínu A, neinteraguje s jeho receptormi. Predpokladá sa, že N/OFQ jezapojený v modulácii prenosu nociceptívnych informácií a v procesochmodulovaných opioidmi a ovplyvňovaní pamäťových procesov a učenia

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 131
(Noda a spol., 2000; Meis, 2003). Okrem toho sa predpokladá, že noci-ceptín je zapojený v procesoch centrálnej regulácie príjmu potravy a žepôsobí ako neuromodulátor v gastrointestinálnom nervovom systéme(Osinski a Brown, 2000; Olszewski a Levine, 2004).Nociceptín sa vyskytuje v likvore aj v plazme. Nedávne štúdie uká-
zali, že nociceptínový transkript je u človeka exprimovaný aj bunkamiimunitného systému a bol detegovaný aj v iných periférnych orgánoch,vrátane pečene. Vysoké koncentrácie nociceptínu sa zistili v plazme osôbs akútnymi a chronickými bolesťami a Wilsonovou chorobou. Zdá sa, žeaj nádorové procesy výrazne zvyšujú plazmatické hladiny nociceptínu,ako sa to pozorovalo v prípade cirhózy pečene spojenej s primárnym he-patocelulárnym karcinómom, ale bez prítomnosti nociceptívneho dráž-denia.
Nocistatín
Nocistatín je bioaktívny peptid, ktorý vzniká z rovnakej molekuly akonociceptín. Nociceptín a nocistatín vznikajú z toho istého preprohor-mónu rozličným spracovaním prekurzorovej mRNA. Preto profil ichsplicingových molekúl (na základe rozdielu v „zostrihuÿ mRNA) pod-mieňuje ich fyziologické účinky. Predpokladá sa, že aj účinnosť morfí-novej analgézie nocistatínu závisí od produktu splicingového procesu.Uvažuje sa, že nocistatín spolu s nociceptínom majú významnú úlohuv prenose nociceptívnych informácií pri neuropatickej bolesti (Martina spol., 1998). Nocistatín má pravdepodobne významnú úlohu aj v pa-mäťových procesoch a v procesoch učenia (Okuda-Ashitaka a Ito, 2000).
Orexín
Roku 1998 dve skupiny vedcov (de Lecea a spol. a Sakurai a spol.)nezávisle opísali nový druh neuropeptidov, ktorý jedna skupina po-menovala ako hypokretíny (HCRT) a druhá ako orexíny (ORX). Ideo identické neuropeptidy, ktorých syntéza prebieha v neurónoch vysky-tujúcich sa v laterálnom hypotalame v oblastiach nucleus perifornicalisa v dorzálnom, laterálnom a posteriórnom hypotalame. No napriek ichanatomicky obmedzenému výskytu, ako aj ich relatívne malému počtu,ORX/HCRT neuróny inervujú rozsiahle oblasti CNS vrátane talamu,subtalamu, substantia nigra, rapheálnych jadrier, locus coeruleus, ven-trálnej tegmentálnej oblasti, medulárnej retikulárnej formácie, nucleus

132 NEUROTRANSMITERY
tractus solitarii a iných oblastí mozgového kmeňa (Peyron a spol., 1998).Okrem CNS sa orexínergické bunky nachádzajú aj v gastrointestinál-nom trakte, pankrease a nadobličkách (Kirchgessner, 2002; Kukkonena spol., 2002).Rozlišuje sa ORX A (HCRT 1) a ORX B (HCRT 2), pričom ORX A
tvorí 33 a orexín B 28 aminokyselinových zvyškov. Pôsobia prostredníc-tvom receptorov OXR1 a OXR2. Typ receptorov OXR1 sa vyskytuje ajv membránach hypotalamických neurónov produkujúcich vazopresín,oxytocín, somatostatín, vazoaktívny intestinálny polypeptid, kortiko-liberín, neuropeptid Y, AGRP, POMC, CART, MCH a ORX/HCRT(Backberg a spol., 2002).Predpokladá sa, že orexín syntetizujúce neuróny ovplyvňujú chuť
do jedla a príjem potravy. Svedčí o tom recipročné prepojenie medziHCRT-neurónmi a hypotalamickými neurónmi syntetizujúcimi neuro-peptid Y a skutočnosť, že HCRT-neuróny exprimujú leptínové receptory(Horvath a spol., 1999). Inzulínom indukovaná hypoglykémia zvyšujemnožstvo prekurzorových molekúl pre HCRT a podanie leptínu zni-žuje hladiny HCRT. Komorové podanie HCRT aktivuje príjem potravya zvyšuje metabolizmus (Sweet a spol., 1999).Experimentálne práce poukazujú na spojitosť medzi dysfunkciou
orexínergického systému a vznikom narkolepsie. Narkolepsia je charak-teristická ospanlivosťou a kataplexiou (rýchly pokles svalového tonusupočas náhle vznikajúcich silných emócií). Hoci kľúčové anatomické ele-menty zodpovedné za vznik kataplexie sú zrejmé, neuróny zodpovednéza náhlu ospanlivosť počas narkolepsie sa zatiaľ neidentifikovali. Do-kázalo sa však, že redukovaná excitácia noradrenergických neurónovv locus coeruleus, sérotonínergických neurónov v nuclei raphes, dopamí-nergických neurónov v hypotalame a histamínergických neurónov v po-steriórnom hypotalame zodpovedá za redukciu bdelosti a vyúsťuje doútlmu.Orexín pravdepodobne moduluje aj činnosti spojené s hypotalamic-
kou kontrolou funkcií predného laloka hypofýzy, ako aj s kontrolu auto-nómnych funkcií (Beuckmann a Yanagisawa, 2002; Ferguson a Samson,2003; Samson a spol., 2005). Orexín môže ovplyvňovať činnosť sympa-tikového systému (a tým aj kardiovaskulárneho systému) prostredníc-tvom modifikácie činnosti neurónov nucleus paraventricularis hypotha-lami (Shirasaka a spol., 2002). Niektoré zistenia naznačujú, že neurónylaterálneho hypotalamu, ktoré syntetizujú orexín, sa zúčastňujú na re-

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 133
Neuropeptid ReceptorTransdukčný
mechanizmus
VazopresínV1a, V1b Gq/11V2 Gs
Oxytocín OT-R Gq/11, Gi/o
Kortikoliberín; Urokortín 1, 2, 3; Sauvagín CRFR1, CRFR2 Gs
Urotenzín II UT-R Gq/11
Tyreoliberín TRH-R Gq/11
Somatostatín sst1–sst5 Gi
Adrenokortikotropný hormón MC2 Gs
α-, β-, γ-melanocyty stimulujúci hormón MC1–MC5 Gs
β-endorfín; enkefalín; dynorfín δ, κ, µ Gi/o
Substancia P; neurokinín A, B; NK1–NK3 Gq/11neuropeptid K, γ
Glukagónu podobný peptid 1 GLP-1-R Gs
Vazoaktívny intestinálny polypeptid; VPAC1, VPAC2, Gshistidínový izoleucínový peptid; PAC1hypofýzovú adenylátcyklázu aktivujúcipeptid
Adrenomedulín AM-R Gs
Agouti príbuzný proteín antagonistaMC-receptorov
Amylín AMY-R Gs
Angiotenzín IIAT1 Gq/11AT2 Tyr&Ser/Thr
fosfatázy
Apelín APJ Gi/o
Endomorfín 1, 2 µ Gi/o
Galanín; galanínu podobný peptidGAL1, GAL3 Gi/oGAL2 Gi/o, Gq/11
Ghrelín GHS-R1a Gq/11
Tabuľka 4.2 Klasifikácia receptorov pre neuropeptidy a mechanizmy trans-
dukcie signálu. ↑ – receptor nie je známy; ↓ – zvýšenie koncentrácie; zníženieaktivity (modifikované podľa Alexander a spol., 2006).
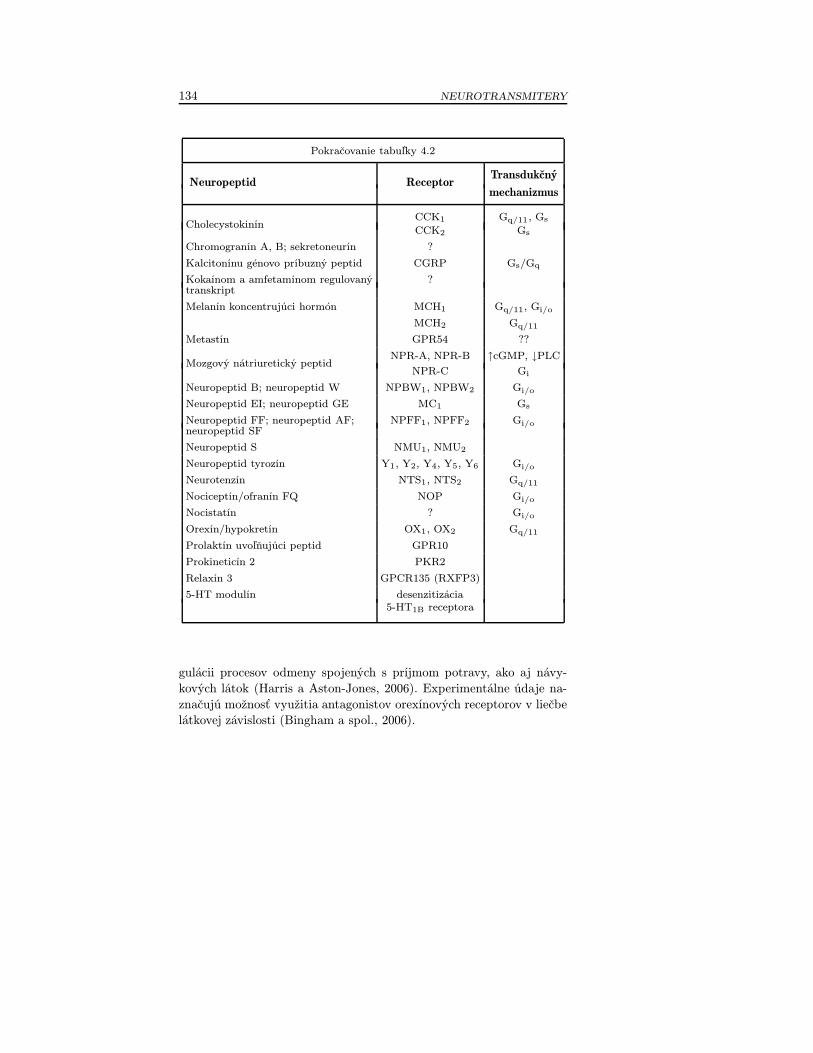
134 NEUROTRANSMITERY
Pokračovanie tabuľky 4.2
Neuropeptid ReceptorTransdukčný
mechanizmus
CholecystokinínCCK1 Gq/11, GsCCK2 Gs
Chromogranín A, B; sekretoneurín ?
Kalcitonínu génovo príbuzný peptid CGRP Gs/Gq
Kokaínom a amfetamínom regulovaný ?transkript
Melanín koncentrujúci hormón MCH1 Gq/11, Gi/o
MCH2 Gq/11
Metastín GPR54 ??
Mozgový nátriuretický peptidNPR-A, NPR-B ↑cGMP, ↓PLC
NPR-C Gi
Neuropeptid B; neuropeptid W NPBW1, NPBW2 Gi/o
Neuropeptid EI; neuropeptid GE MC1 Gs
Neuropeptid FF; neuropeptid AF; NPFF1, NPFF2 Gi/oneuropeptid SF
Neuropeptid S NMU1, NMU2
Neuropeptid tyrozín Y1, Y2, Y4, Y5, Y6 Gi/o
Neurotenzín NTS1, NTS2 Gq/11
Nociceptín/ofranín FQ NOP Gi/o
Nocistatín ? Gi/o
Orexín/hypokretín OX1, OX2 Gq/11
Prolaktín uvoľňujúci peptid GPR10
Prokineticín 2 PKR2
Relaxín 3 GPCR135 (RXFP3)
5-HT modulín desenzitizácia5-HT1B receptora
gulácii procesov odmeny spojených s príjmom potravy, ako aj návy-kových látok (Harris a Aston-Jones, 2006). Experimentálne údaje na-značujú možnosť využitia antagonistov orexínových receptorov v liečbelátkovej závislosti (Bingham a spol., 2006).

4 Peptidové („veľkomolekulovéÿ) neurotransmitery 135
Prolaktín uvoľňujúci peptid
Prolaktín uvoľňujúci peptid (prolactin-releasing peptide, PrRP) bolspočiatku charakterizovaný ako peptid regulujúci vyplavovanie prolak-tínu z adenohypofýzy. Preukázalo sa však, že PrRP vyvoláva vypla-vovanie prolaktínu iba u samíc potkanov počas estru (Ibata a spol.,2000). Neskoršie štúdie však poukázali na jeho zapojenie v reguláciimetabolickej homeostázy, stresovej odpovede, kardiovaskulárnych regu-láciách, modulácii sekrécie gonadotropínov a rastového hormónu a re-gulácii spánku (Sun a spol., 2005).
5-HT-modulín
5-HT-modulín (5-HT-moduline) je endogénny neuropeptid, ktorý selek-tívne a alostericky interaguje s 5-HT1B-receptormi. Neviaže sa však nato isté miesto receptora ako sérotonín. Jeho väzba na 5-HT1B-receptoryspôsobuje zmeny v ich konformácii. Týmto spôsobom blokuje účinkysérotonínu, resp. vedie k ich desenzitizácii, a preto pôsobí ako antago-nista 5-HT1B-receptorov. Na druhej strane 5-HT-modulín prostredníc-tvom presynaptických 5-HT1B-autoreceptorov zvyšuje sekréciu séroto-nínu. Sérotonínergická aktivita je sprostredkovaná najmenej 14 druhmisérotonínergických receptorov a 5-HT1B-receptory patria medzi najvý-znamnejšie z nich. Takto 5-HT-modulín môže pôsobiť ako veľmi dôležitýa účinný lokálny regulátor sérotonínergickej aktivity. Je lokalizovanýv sérotonínergických nervových termináloch, kde inhibuje sekréciu, aleaj biosyntézu sérotonínu. Receptory pre 5-HT-modulín vystupujú ajako heteroreceptory lokalizované v nesérotonínergických termináloch,kde inhibujú sekréciu korešpondujúcich neurotransmiterov (acetylcho-lín, GABA, noradrenalín).5-HT-modulín sa uvoľňuje najmä v tých oblastiach mozgu, ktoré
sú bohaté na 5-HT1B-receptory. Akútny reštrikčný stres zvyšuje sekré-ciu 5-HT-modulínu u potkanov. Deaktivovanie 5-HT-modulínu špecific-kými protilátkami bráni vzniku anxiety u myší. Preto existuje predpo-klad, že látky schopné inhibovať tento peptid by mohli mať protistresovéa anxiolytické vlastnosti. Pretože zvýšená sérotonínergická aktivita jespojená s vývojom psychiatrických ochorení, ako sú depresia, agresivitaa anxieta, uvažuje sa o tom, že látky inhibujúce aktivitu 5-HT-modulínuby mohli predstavovať nové antidepresíva a anxiolytiká (Massot a spol.,1998; Grimaldi a spol., 1999).

Úloha neurotransmiterov v regulácii
fyziologických procesov
B. Mravec
V procesoch prenosu a spracovania signálov nervovým systémomvyužíva organizmus dva základné mechanizmy. Prvým mechanizmomje prenos signálov medzi neurónmi prostredníctvom chemických látok.Druhým mechanizmom je prenos informácií prostredníctvom elektric-kých synáps. Na mnohých miestach v CNS však neuróny využívajú naprenos informácií chemické aj elektrické synapsy (Valenstein, 2005).Chemický prenos signálov prostredníctvom neurotransmiterov, resp.neuromodulátorov prevláda. Dá sa povedať, že správna činnosť väčšinyprocesov prebiehajúcich v nervovom systéme je podmienená precíznoureguláciou:
• syntézy, uvoľňovania a inaktivácie neurotransmiterov,
• syntézou, aktiváciou a inaktiváciou ich receptorov,
• intracelulárnych procesov prenosu signálov na úrovni cytoplazmy,alebo až na úrovni bunkového jadra.
Nervový systém je komplexný systém, v ktorom jednotlivé neuro-transmitery navzájom komplexne interagujú. Každá funkcia je preto
136

5 Úloha neurotransmiterov v regulácii fyziologických procesov 137
podmienená interakciou neurónov syntetizujúcich rôzne neurotransmi-tery. Na regulácii činností v nervovom systéme sa zúčastňujú interakciemedzi jednotlivými štruktúrami. Opis funkcií neurotransmiterov v re-gulácii vybraných fyziologických procesov sa preto zameriava na najdô-ležitejšie zložky neurotransmiterových systémov v súlade so súčasnýmipoznatkami. To však neznamená, že iné neurotransmiterové systémy sanezúčastňujú na regulácii opisovaných fyziologických dejov.
Regulácia príjmu potravyA.Kiss, B.Mravec
Hlad je pocit, ktorým nervový systém vyšších živočíchov upozorňujena potrebu príjmu potravy. Po uspokojení tejto potreby sa dostavuje po-cit sýtosti. V mozgu človeka existuje komplex oblastí zodpovedných zatvorbu a prenos týchto pocitov. Počas svojej existencie človek postupneracionalizoval príjem potravy a postupne nahradil primárny inštinktobživy za spoločenskú zvyklosť. Tým sa človek oddelil od ostatných ži-vočíchov a stal špecifickým druhom, ktorý dokáže prijímať potravu ajbez pocitu hladu, ale aj dobrovoľne hladovať.Udržiavanie energetickej rovnováhy je komplexne regulovaný pro-
ces, na ktorom sa zúčastňujú neuróny syntetizujúce široké spektrumneurotransmiterov (Kalra a spol., 1999; Berthoud 2002).Neuronálna kontrola a regulácia príjmu potravy a výdaja energie
spočíva v troch úzko prepojených procesoch: vstup – integrácia – vý-stup. Vstup predstavuje prísun informácií z periférie do mozgu, a tohumorálnou a nervovou cestou prostredníctvom signálnych látok, akosú inzulín, ghrelín, leptíny. Experimentálne práce ukázali, že mozog nieje schopný bez týchto signálov samočinne regulovať energetické pomeryv organizme. Integrácia je veľmi zložitý proces, v ktorom majú vý-znamnú úlohu mnohé nervové okruhy vychádzajúce z viacerých oblastímozgu, ktoré spracúvajú množstvo informácií. Predpokladá sa, že hlav-ným regulačným centrom pre reguláciu príjmu potravy a výdaj energieje hypotalamus (Kalra a spol., 1999; Wilding, 2002). Výstup je odpo-veď regulačného centra smerom k periférii, pričom cieľom výstupu jeaktivácia takých regulačných a kompenzačných mechanizmov na peri-férii, aby sa v daných podmienkach dosiahol čo najoptimálnejší stavenergetických pomerov v organizme (obr. 5.1).

138 NEUROTRANSMITERY
Obr. 5.1 Schematické znázornenie periférnych a centrálnych zložiek zúčast-
ňujúcich sa na regulácii príjmu potravy. Periférne syntetizované látky (napr.inzulín, leptín, ghrelín, CCK) ovplyvňujú činnosť CNS prostredníctvom dvoch
ciest – humorálnej a neuronálnej. Cirkulujúce hormóny účinkujú v oblastiach
cirkumventrikulárnych orgánov (eminentia mediana – ME, area postrema –AP). Senzorické zakončenia a neuróny v ich blízkosti (napr. neuróny nucleus
arcuatus, NARC) spracúvajú informácie z periférie o nutričnom stave v or-
ganizme. Nervové zakončenia nervus vagus obsahujú receptory pre niektorégastrointestinálne hormóny, čo umožňuje rýchly prenos informácií z gastroin-
testinálneho a tukového tkaniva do CNS neuronálnou cestou. Na spracovaníinformácií o nutričnom stave sa zúčastňujú neuróny CNS, ktoré syntetizujú
aj orexigenické a anorexigenické peptidy. AHL– laterálna hypotalamická ob-
lasť; APF–perifornikálna oblasť; NTS – nucleus tractus solitarii; GN– ganglionnodosum; GIT – gastrointestinálny trakt; NDM–nucleus dorsomedialis; NVM–
nucleus ventromedialis; OXM–oxyntomodulín; PVN–nucleus paraventricula-
ris; NAc – nucleus accumbens; skratky hormónov pozri v tab. 2.3; prerušovanáčiara – prenos signálu systémovou cirkuláciou; bodkovaná čiara – prenos signálu
k lokálnym zakončeniam nervus vagus (modifikované podľa Korner a Leibel,2003; Wynne a spol., 2005).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 139
Zo 40 štruktúr hypotalamu má významnú úlohu v regulácii príjmupotravy a výdaja energie najmenej 5 oblastí: nucleus arcuatus, nucleusparaventricularis, nucleus ventromedialis, nucleus dorsomedialis a late-rálny hypotalamus (Elmguist a spol., 1999; Kalra a spol., 1999; Bert-houd 2002). Tieto oblasti obsahujú neuróny, ktoré na základe účinkuna reguláciu príjmu potravy možno rozdeliť na dve veľké skupiny, ore-xigenické a anorexigenické. Orexigenické neuróny syntetizujú orexige-nicky pôsobiace látky, ktoré zvyšujú príjem potravy, a tým aj telesnúhmotnosť. Anorexigenické neuróny syntetizujú anorexigenické látky,ktorá pôsobia opačne. Orexigénne neuróny produkujú širokú paletu bio-logicky aktívnych látok, ako sú neuropeptid Y (NPY), agouti-príbuznýpeptid (AGRP), hypokretíny/orexíny (HCRT/ORX), melanín koncen-trujúci hormón (MCH), galanín (GAL), dynorfín (DYN), β-endorfín(β-END), GABA, noradrenalín a adrenalín. Anorexigénne neuróny syn-tetizujú napr. α-melanocyty stimulujúci hormón (α-MSH), kokaínoma amfetamínom regulovaný transkript (CART), kortikoliberín (CRH,CRF), neurotenzín (NT), urokortín III, cholecystokinín (CCK), gluka-gónu podobný peptid (GLP-1) (Rowland a spol., 1996; Schwartz a spol.,2000; Sainsbury a spol., 2002; Palkovits, 2003; Ramo a spol., 2005).Distribúcia týchto látok v uvedených jadrách hypotalamu, charakterich vzájomnej komunikácie a ovplyvňovanie periférnymi signalizačnýmilátkami, ako aj ich komunikácia s inými štruktúrami mozgu predstavujemozaiku vysokodynamických funkčných vzťahov, ktoré tvoria podstaturegulácie príjmu potravy a výdaja energie (obr. 5.1) (Kalra a spol., 1999;Berthoud, 2002).Signály informujúce mozog o fyziologickom stave energetickej ho-
meostázy prichádzajú z viacerých periférnych orgánov (Kalra a spol.,1999; Berthoud, 2002). Peptidové hormóny syntetizované v gastroin-testinálnom trakte sa významnou mierou zúčastňujú na regulácii pro-cesov spojených s príjmom potravy tým, že modulujú činnosť neuró-nov v CNS, s výrazným vplyvom na činnosť hypotalamických štruktúr(Rehfeld, 1998; Richard a Baraboi, 2004). Pocit nasýtenosti signalizujegastrointestinálny peptid YY (PYY3−36), ktorého úlohou je zastaveniepríjmu potravy. Naopak ghrelín, syntetizovaný z rovnakej prekurzorovejmolekuly mRNA ako PYY, signalizuje znížením svojich plazmatickýchhladín vyprázdnený žalúdok a zvýšenými hladinami naplnený žalúdok.Ghrelín je teda látka, ktorá aktuálne reaguje na dva opačné stavy ob-jemu žalúdka. Ghrelín zvyšuje chuť do jedla, pričom jeho hladiny sú

140 NEUROTRANSMITERY
ráno vyššie ako večer. Plazmatickú hladinu ghrelínu zvyšuje hlad a hy-poglykémia a naopak jeho hladiny znižuje nasýtenosť, hyperglykémiaa obezita (Solomon a spol., 2005). Leptín, ktorý je produkovaný adipo-cytmi, signalizuje stav zásob energie a ich vyrovnanosť (Zhang a spol.,1994). Účinok leptínu nie je rýchly, ale prejavuje sa až po 8–10 hodi-nách. Neinhibuje epizodický príjem potravy, len znižuje objem prija-tej potravy počas tejto epizódy. Leptín pôsobí na mnohých miestachhypotalamu s výrazným efektom v nucleus arcuatus (Kalra a spol.,1998). Obézni ľudia majú obyčajne vysoké hladiny leptínu. Inzulín jeprodukovaný β-bunkami pankreasu a je veľmi významne zapojený dokontroly chuti do jedla a poskytuje CNS informácie o hladinách plazma-tickej glukózy. Zmeny plazmatických hladín inzulínu sa odrážajú aj najeho koncentrácii v likvore. Keď sa hladiny inzulínu zvýšia, dochádzav mozgu k redukcii príjmu potravy. Inzulín teda pôsobí inhibične nacentrálne orexigénne dráhy, inhibuje účinok NPY, pričom jeho účinokje veľmi rýchly (Sahu a spol., 1992, 1995). V mnohých situáciách dochá-dza k synergistickým efektom uvedených hormónov. Napríklad u zvie-rat trpiacich podvýživou znížené hladiny inzulínu signalizujú v NARCodblokovanie pocitu nasýtenia sprostredkovaného CCK, čím sa docielipríjem väčšieho množstva potravy. U obéznych zvierat bude naopak in-zulín tou istou cestou zvyšovať účinok CCK pre pocit nasýtenosti, čímvedie k obmedzeniu príjmu potravy. V súčasnosti sa zdá, že endogénnyCCK pôsobí počas prijímania potravy tak, že limituje množstvo pri-jatej potravy. Existujú dôkazy pre úlohu CCK v patofyziológii porúchv príjme potravy, napr. pri vzniku bulímie (Woods a Gibbs, 1989).Periférne hormóny a neuropeptidy regulujúce príjem potravy možno
vo všeobecnosti rozdeliť na pomaly a rýchlo pôsobiace. Pomaly pôso-biace látky nie sú viazané na presný druh potravinovej zložky a účin-kujú dlhodobo, ako napr. inzulín a leptín, ktoré sú secernované do krviproporcionálne k množstvu tuku a energetických látok v cirkulácii. Sta-bilne pôsobia inhibične na príjem potravy a stimulačne na výdaj energie.Rýchlo pôsobiace látky, ako cholecystokinín (CCK) a ghrelín, účinkujúkrátkodobo a špecifickejšie na určité druhy zložiek potravy. Sú secer-nované v gastrointestinálnom trakte počas príjmu potravy, signalizujúpocit „naplneniaÿ, čo vedie k ukončenie príjmu potravy. Periférne pro-dukované hormóny inzulín, leptín a ghrelín patria medzi významné pe-riférne signalizačné látky, ktoré humorálnou cestou a prostredníctvomsvojich receptorov účinkujú priamo na nervové bunky hypotalamu a zú-

5 Úloha neurotransmiterov v regulácii fyziologických procesov 141
častňujú sa na regulácii príjmu potravy a výdaja energie (Kalra a spol.,1999).Jedna z najvýznamnejších mozgových oblastí slúžiaca na kontrolu
energetického metabolizmu a reguláciu chuti do jedla je nucleus ar-cuatus (NARC). Anatomická lokalizácia NARC bilaterálne po bokochbázy III. mozgovej komory nad eminentia mediana, t.j. oblasťou bezmozgovo-cievnej bariéry, je strategicky veľmi výhodná pre príjem sig-nálov z krvného riečiska (Banks a spol., 1996). Leptín, rastový hormón,glukokortikoidy, ghrelín, inzulín a metabolity ako glukóza môžu pretoprostredníctvom svojich receptorov priamo pôsobiť na neuróny NARC(Kalra a spol., 1999; Berthoud, 2002). Okrem toho sa v NARC nachá-dza veľké množstvo neurónov produkujúcich orexigénne a anorexigénnepeptidy, ktoré majú bohaté prepojenie na všetky ostatné hypotalamickéjadrá. Dôležité aferentácie okrem intrahypotalamických štruktúr do-stáva NARC aj z extrahypotalamických oblastí mozgu, ako sú nucleusparabrachialis a nucleus tractus solitarii (Guan a spol., 2001).V NARC sú dva významné typy neurónov, ktoré kontrolujú príjem
potravy. Jeden typ pôsobí ako akcelerátor, druhý ako inhibítor. Akce-leračné neuróny produkujú NPY, inhibičné αMSH. Neuróny produku-júce NPY syntetizujú zároveň aj ďalší hormón AGRP, ktorý blokujereceptory pre αMSH (Broberger a spol., 1998). Neuróny exprimujúceNPY/AGRP, ktoré reagujú napr. na zníženie hmotnosti, môžu zvýšiťpríjem potravy dvoma spôsobmi – zvýšením produkcie a aktivity neuró-nov produkujúcich NPY, alebo blokovaním receptorov pre αMSH. Obatieto peptidy sú potenciálnymi anabolickými peptidmi a ich aplikácia domozgových komôr vyvoláva hyperfágiu, znížený výdaj energie a zvyšo-vanie telesnej hmotnosti. NPY aj AGRP neuróny syntetizujú receptorypre inzulín aj leptín (Mercer a spol., 1996). Podanie týchto hormónovlokálne do NARC znižuje syntézu NPY aj AGRP. NPY mRNA sa zvy-šuje počas hladu, ale exogénne podanie leptínu potláča jeho nadbytočnúprodukciu. Periférne zmeny v energetických pomeroch signalizované domozgu leptínom a inzulínom inhibujú NPY/AGRP neuróny. U hlodav-cov napr. endogénne hladiny NPY v NARC-PVN systéme vrcholia kukoncu dňa, keď sa začína ich nočná aktivita, resp. keď obyčajne za-čínajú potkany prijímať potravu. NPY/AGRP syntetizujúce neurónyvýznamne projikujú hlavne do nucleus paraventricularis a laterálnehohypotalamu (Berthoud, 2002).

142 NEUROTRANSMITERY
Druhou dvojicou peptidov aktivovaných signálmi z tukového tka-niva (leptínom) sú αMSH/CART, ktoré tiež obsahujú mnoho leptíno-vých receptorov a sú potenciálnymi katabolickými peptidmi (Cheunga spol., 1997). Ich podanie do III. mozgovej komory znižuje príjem po-travy, zvyšuje výdaj energie a znižuje telesnú hmotnosť. Vo všeobecnostisa dá povedať, že počas aktivácie NPY/AGRP neurónov dochádza k tl-meniu αMSH/CART a naopak, pričom v oboch prípadoch ide o úči-nok leptínu (Wilding 2002). αMSH/CART, ako aj NPY/AGRP syn-tetizujúce neuróny projikujú hlavne do oblasti laterálneho hypotalamua inervujú HCRT aj MCH neuróny (Elias a spol., 1998). Malá skupinkaαMSH/CART syntetizujúcich neurónov lokalizovaných v prednej častiNARC projikuje k autonómnym oblastiam predĺženej miechy a miechy.Nucleus paraventricularis (PVN) má heterogénne bunkové zlože-
nie. Magnocelulárne neuróny PVN, syntetizujúce vazopresín a oxyto-cín, projikujú do neurohypofýzy. Parvocelulárne neuróny syntetizujúceCRH a vazopresín projikujú k portálnemu systému eminentia mediana,ktorý zásobuje krvou adenohypofýzu. Niektoré parvocelulárne neurónyPVN obsahujú oxytocín a projikujú do kaudálnej časti mozgu k centrámparasympatikového nervového systému a do miechy k pregangliovýmneurónom sympatikového nervového systému (Swanson a Sawchenko,1980). Prostredníctvom týchto dráh môžu neuróny PVN priamo ovplyv-ňovať sekréciu hormónov z pankreasu a drene nadobličiek aj somatomo-torickú aktivitu, ktorá je spojená so správaním sa v čase príjmu potravy.Neurotransmisia sprostredkovaná parvocelulárnym oxytocínom v moz-govom kmeni a mieche má supresívny účinok na chuť do jedla. Leptínaktivuje PVN neuróny. PVN môže sprostredkúvať mnoho signálov lep-tínu, pretože dostáva priame projekcie z neurónov NACR senzitívnychna leptín (Berthoud 2002).Z funkčného hľadiska je PVN dodnes najpodrobnejšie opísaná štruk-
túra hypotalamu. PVN je zapojený v rôznych endokrinných, autonóm-nych a imunitných procesov a dostáva aferentácie takmer zo všetkýchoblastí mozgu. Z hľadiska regulácie energetickej homeostázy k naj-významnejším projekciám patria tie, ktoré prichádzajú z laterálnehohypotalamu, noradrenergickej oblasti locus coeruleus, sérotonínergic-kej oblasti nucleus raphe dorsalis a mozgového kmeňa. Množstvo úda-jov v odbornej literatúre poukazuje na stimulačný účinok noradrena-línu a inhibičný účinok sérotonínu na neuróny PVN (Smith a spol.,1999). Všetky druhy orexigénnych peptidov vrátane NPY, GAL, HCRT,

5 Úloha neurotransmiterov v regulácii fyziologických procesov 143
GABA, noradrenalínu a adrenalínu stimulujú chuť do jedla a naopakCRH a leptín pôsobia inhibične. Z hľadiska regulácie energetickej ho-meostázy sa zdá, že PVN je integračné miesto mnohých interoreceptív-nych informácií a zúčastňuje sa prevažne na integrácii prijatých endok-rinných a autonómnych signálov a má kľúčovú úlohu pri regulácii príjmupotravy v rôznych stresových podmienkach (Imaki a spol., 1992).Nucleus ventromedialis (NVM) je pomerne rozsiahla štruktúra hy-
potalamu s mnohými funkčnými podoblasťami, ktoré však dodnes niesú presne anatomicky ohraničené a funkčne charakterizované. Mnohorokov sa táto štruktúra mylne považovala za centrum sýtosti. Neurónydorzomediálnej časti NVM sú bohaté hlavne na inzulínové a leptínovéreceptory, ktoré sprostredkúvajú signály z tukového tkaniva (Van Dijka spol., 1996). Je však zaujímavé, že ani orexigénne, ani anorexigénneneuróny sa v tejto oblasti nedetegovali, pričom však selektívna léziatiel neurónov v tejto oblasti (bez porušenia dráh) vedie k hyperfágii.Aj lokálne podanie mnohých látok, ako NPY, GAL, GABA, β-END,stimuluje príjem potravy, kým leptín ho inhibuje. Na základe týchtopozorovaní sa usudzuje, že NVM je hlavne receptívna oblasť pre viacerésignálne molekuly, ktoré sú zapojené v regulácii chuti do jedla (Kalraa spol., 1999; Berthoud 2002).NVM prijíma aferentáciu hlavne z amygdaly, laterálneho hypota-
lamu a mediálnych jadier hypotalamu. Má prepojenie aj na štruktúrymozgového kmeňa. Mnohé neuróny, lokalizované hlavne v jeho dorzálnejčasti, obsahujú leptínové receptory, pričom časť týchto buniek projikujek neurónom PVN a subPVN, t.j. oblasti nachádzajúcej sa bezprostrednepod PVN (Elmquist a spol., 1998). Na rozdiel od ostatných hypotala-mických štruktúr neuróny NVM projikujú hlavne do oblasti retikulár-nej formácie a periakvaduktálnej šedej hmoty. Bohaté projekcie vysielaNVM aj k ostatným mediálne (ale nie laterálne) a periventrikulárne ulo-ženým hypotalamických jadrám, čo svedčí hlavne o jeho intrahypotala-mickej modulácii (Canteras a spol., 1994). Hoci eferentné dráhy NVMsmerom k NARC neboli identifikované, neurónové projekcie obsahujúceNPY, CART a β-END boli z NARC do NVM jasne opísané. Dôležité jehlavne spojenie NVM-NDM-NPV, ktorého prerušenie na úrovni NVMvedie k hyperfágii.Nucleus dorsomedialis (NDM) má približne rovnakú veľkosť ako
NVM a anatomicky sa nachádza nad ňou. Vyradenie NDM znižuje chuťdo jedla, ale v menšej miere ako vyradenie NVM. NDM sa považuje za

144 NEUROTRANSMITERY
dôležitú oblasť regulácie príjmu potravy a za oblasť, v ktorej dochádzak interakcii medzi NPY a leptínom. Najcitlivejším miestom NDM naleptín je jeho dorzomediálna podoblasť (Elmquist a spol., 1998). Tátooblasť prijíma rozsiahlu aferentáciu z okolitých jadier vrátane NARC,NVM, NPV a laterálneho hypotalamu (Thompson a Swanson, 1998).Projikujú k nej aj neuróny z mozgového kmeňa, ale aj štruktúry reti-kulárnej formácie a periakvaduktálnej šedej hmoty. V kaudálnej častijadra mnohé neuróny obsahujú leptínové receptory a tieto neuróny pro-jikujú priamo do PVN. V jeho strednej, t.j. kompaktnej časti mnohéneuróny produkujú NPY a obsahujú CCK-A receptory (Lewis a spol.,1993). Potkany s mutáciou týchto receptorov sú hyperfágické a obézne.CCK inhibuje expresiu NPY v NDM, a tým zároveň znižuje aj prí-jem potravy pri veľkej strate energie. Naopak intracerebroventrikulárnepodanie GAL stimuluje NPY-neuróny v NDM a vedie aj k stimuláciichuti do jedla (Kuramochi a spol., 2006). Zdá sa teda, že NDM môževýznamne prispievať k signalizácii počas situácií vedúcich k zvýšenýmnárokom na energiu (napr. počas vývoja, laktácie a intenzívneho cviče-nia).Area hypothalamica lateralis (AHL) anucleus perifornicalis. AHL
je rozsiahla oblasť vyplňujúca laterálny hypotalamus, ktorá je charak-teristická tým, že ňou prechádza množstvo ascendentných a descen-dentných nervových dráh, ako aj významný zväzok dráh – fasciculusmedialis prosencephali (medial forebrain bundle). Významné sú hlavneprojekcie vedúce do predného mozgu, medzimozgu a mozgového kmeňa.Vyradenie tejto oblasti spôsobuje dočasnú afágiu a akinézu (Berthoud,2002). Správanie sa zvierat po veľkej lézii pripomína parkinsonizmus,ktorý je spôsobený dysfunkciou ascendentných dopamínergických dráhstredného mozgu. Neuróny zapojené do regulácie energetického meta-bolizmu produkujú melanín koncentrujúci hormón (MCH) (Bittencourta spol., 1992). Nedostatok leptínu zvyšuje hladiny MCH mRNA. Intra-cerebroventrikulárna aplikácia MCH zvyšuje príjem potravy a nadpro-dukcia MCH vedie k obezite a inzulínovej rezistencii. Zdá sa, že MCHmá mnohostranný a integračný účinok týkajúci sa hlavne behaviorál-nych aspektov prijímania potravy. Druhý významný hormón, ktorý saprodukuje v tejto oblasti, je hypokretín/orexín (Sakurai a spol., 1998).Neuróny produkujúce hypokretín/orexín okupujú oblasť okolo fornixu.Aplikovanie MCH aj hypokretínu stimuluje príjem potravy. Naopak hla-dovanie zvyšuje hladiny mRNA pre oba peptidy (Qu a spol., 1996; Sa-

5 Úloha neurotransmiterov v regulácii fyziologických procesov 145
kurai a spol., 1998). Laterálny hypotalamus prijíma významné NPYer-gické projekcie z NARC. Okrem rôznych projekcií sa v AHL nachádzajúaj glukózareceptívne a glukózasenzitívne neuróny, ktoré môžu priamomonitorovať glykemické zmeny v cirkulácii (Oomura a spol., 1984).Nucleus suprachiasmaticus (NSCH). Aj keď toto jadro nie je ty-
pickou štruktúrou, ktorá spracúva periférne signály týkajúce sa energe-tickej homeostázy, je zapojené do integrácie mnohých procesov spoje-ných s prijímaním potravy v rámci cirkadiánnych alebo cirkaanuálnychrytmov (Nagai, 1978; Kalra a spol., 1999). Potkany prijímajú 85–90%potravy v noci. Človek sa riadi aj sociálnymi zvyklosťami alebo indivi-duálnymi potrebami, kedy prijímať potravu. Vyradenie tejto štruktúryvyvoláva poruchu v časovom naprogramovaní príjmu potravy.Nucleus tractus solitarii (NTS) a area postrema (AP). NTS je
miestom, ktoré dostáva nervovú signalizáciu hlavne zo žalúdka pro-stredníctvom nervus vagus. Tak napríklad CCK prostredníctvom tohtonervu prenáša informáciu nasýtenosti do mozgu. Táto signalizácia jerýchlejšia ako humorálna, ale nie je monosynaptická. NTS slúži akoprepojovacia stanica pre viscerálne informácie prichádzajúce z perifériek iným oblastiam mozgu vrátane hypotalamu (Berthoud, 2002).V súčasnosti sa ukazuje, že kaudálne štruktúry mozgu, ako NTS
a AP, nie sú len senzormi pre viscerálne a chuťové vnemy, ale majú úlohuaj regulačného charakteru. Zistilo sa, že pri prerušení kontaktu medzipredným mozgom/hypotalamom a štruktúrami mozgového kmeňa ne-dochádza ku kompletnému vyradeniu príjmu potravy (musí sa všakpotrava vložiť priamo do ústnej dutiny, aby mohla byť prijatá) (Grilla Kaplan, 1994). Okrem toho je NTS štruktúra, v ktorej sa nachádzajúokrem NARC aj neuróny syntetizujúce POMC. V tejto oblasti je ajveľký počet MC4-receptorov (Mountjoy a spol., 1994). Navyše apliká-cia agonistu pre MC4-receptory do NTS moduluje príjem potravy a te-lesnú hmotnosť podobným spôsobom, ako keď sa podá do hypotalamu.Podobne urokortín podaný do NTS tlmí príjem potravy. Značná podob-nosť medzi hypotalamom a NTS/AP je aj v schopnosti štruktúr tejtooblasti detegovať množstvo cirkulujúcich hormónov a iných látok v krvi.Takto sú informácie o hormónoch gastrointestinálneho traktu priamomonitorované aj v tejto oblasti mozgu. Aj prítomnosť glukózasenzi-tívnych neurónov v NTS pripomína fenotypovú podobnosť s neurónmihypotalamu (Ritter a spol., 2000). Okrem toho priame a nepriame napo-jenia NTS s parasympatikovými a sympatikovými neurónmi vytvárajú

146 NEUROTRANSMITERY
anatomický predpoklad pre priame zapojenie tejto oblasti do modulá-cie činnosti mnohých periférnych orgánov zapojených do procesu príjmua výdaja energie.Nutričné procesy organizmu sú značne dynamické a variabilné pro-
cesy, ktoré sú nepretržite monitorované a modulované centrálnym ner-vovým systémom. V súčasnosti veľmi rýchlo pribúdajú poznatky o pe-riférnych a centrálnych zložkách tohto zložitého systému a ich vzájom-ných vzťahoch. Vzhľadom na veľkú dynamiku faktorov zúčastňujúcichsa na procesoch spojených s príjmom potravy a výdajom energie všakzostáva ešte mnoho neobjasneného (Blundell a spol., 1993).
Homeostáza telesných tekutínB.Mravec
V regulácii homeostázy telesných tekutín má nervový systém integ-račnú úlohu. S tým súvisí aj zapojenie veľkého množstva neurotransmi-terov v regulácii uvedených procesov (Palkovits, 1984).Nervový systém veľmi precízne monitoruje osmolalitu a zloženie
plazmy. V týchto procesoch majú významnú úlohu hypotalamickéneuróny a cirkumventrikulárne orgány, organum subfornicale a orga-num vasculosum laminae terminalis (OVLT) (obr. 2.6). Tieto oblasti,v ktorých cievy a okolité tkanivo nevytvárajú mozgovo-cievnu barié-ru, priamo monitorujú osmolalitu plazmy a koncentráciu angiotenzínuII. Eferentné nervové dráhy z OVLT inervujú nucleus paraventricula-ris a nucleus supraopticus hypotalamu. Tieto dráhy sa zúčastňujú naaktivácii sekrécie vazopresínu z hypotalamických jadier počas situácií,keď sú osmolalita plazmy a plazmatické hladiny angiotenzínu II zvýšené(McKinley a spol., 2001a, 2001b).Ďalší mechanizmus v regulácii homeostázy telesných tekutín vy-
užíva neuróny syntetizujúce atriálny nátriuretický peptid (ANP). Tietoneuróny, ktoré sa nachádzajú v nucleus paraventricularis, ako aj v ros-trálnej časti tretej mozgovej komory (tzv. „anteroventral third ventri-cular regionÿ, ktorý zahŕňa OVLT), vysielajú axóny do oblasti emi-nentia mediana a do neurohypofýzy. Neuróny týchto oblastí sú akti-vované počas situácií charakterizovaných zväčšenou náplňou krvnéhoriečiska. To vedie k aktivácii karotických, aortových a renálnych baro-receptorov a následnej stimulácii neurónov v nucleus tractus solitarii

5 Úloha neurotransmiterov v regulácii fyziologických procesov 147
(NTS). Neuróny NTS aktivujú noradrenergické neuróny v locus coeru-leus a pravdepodobne aj sérotonínergické neuróny v nuclei raphe. Tietomonoamínergické neuróny vysielajú axóny do hypotalamu, kde aktivujúcholínergické interneuróny. Cholínergické neuróny stimulujú hypotala-mické neuróny syntetizujúce ANP. Uvoľnenie ANP vedie k aktiváciikaskády procesov, ktorých výsledkom je inhibícia príjmu vody a solí. Natýchto procesoch sa zúčastňuje zvýšené vyplavovanie oxytocínu a blo-káda účinku angiotenzínu II (obr. 5.2) (McCann a spol., 1997).
Bdenie a spánok
Spánok a bdenie predstavujú stavy, v ktorých majú viaceré štruktúrycentrálneho nervového systému rozdielnu aktivitu. Táto aktivita môžebyť významne ovplyvnená senzorickou stimuláciou.Pacemakerom biologických rytmov v organizme je nucleus supra-
chiasmaticus (NSCH), ktorý je synchronizovaný s cyklom deň/noc pro-stredníctvom priamych neuronálnych signálov zo špecializovaných foto-receptorov sietnice. Na molekulárnej úrovni sa na regulácii cirkadián-nych rytmov zúčastňujú periodické zmeny v transkripčnej aktivite gé-nov per a cry, ktoré sú modulované proteínmi CLOCK a BMAL1 (Greena Menaker, 2001).NSCH obsahuje neuróny, ktoré syntetizujú niekoľko rôznych neuro-
transmiterov (napr. vazopresín, angiotenzín II, GABA, VIP). NeurónyNSCH inervujú oblasti hypotalamu, ktoré sa významne zúčastňujú naregulácii spánku a bdenia, reprodukčných cyklov a nálady. Inerváciabazálnych oblastí predného mozgu a talamu určuje stav bdelosti. Iner-vácia epifýzy neurónmi NSCH reguluje syntézu a uvoľňovanie melato-nínu, ktorý je významným faktorom modulujúcim cirkadiánne rytmyv organizme (Webster a Stanford, 2001).V regulácii procesov spánku a bdenia majú významnú úlohu neu-
róny lokalizované v retikulárnej formácii mozgového kmeňa a neurónyv hypotalame. Dokázalo sa, že stimulácia oblastí predného hypotalamuvyvoláva spánok, kým stimulácia neurónov v mozgovom kmeni, ktorévytvárajú tzv. ascendentný retikulárny aktivačný systém (ARAS), in-dukuje bdelosť.Medzi neurotransmitery, o ktorých sa predpokladá, že sa zúčastňujú
na regulácii bdenia a spánku, patria hlavne modulačné neurotransmi-tery (acetylcholín, histamín, noradrenalín a sérotonín) (obr. 5.3). Neu-

148 NEUROTRANSMITERY
Obr. 5.2 Neurotransmiterové systémy, ktoré sa zúčastňujú na regulácii ho-
meostázy telesných tekutín. Osmolalita plazmy a cirkulujúci angiotenzín (AII)
aktivujú prostredníctvom nervových zakončení v oblasti cirkumventrikulárnychorgánov (CVO) vazopresínergické neuróny (AVP), ktoré uvoľňujú vazopresín
v neurohypofýze (NH). Informácie z baroreceptorov sú vedené do nucleus trac-tus solitarii (NTS). Neuróny NTS modulujú činnosť noradrenalínergických (NA)
neurónov locus coeruleus (LC) a sérotonínergických (5-HT) neurónov nuclei
raphe (NR). Monoamínergické neuróny následne regulujú činnosť neurónov syn-tetizujúcich atriálny nátriuretický peptid (ANP) priamo, alebo prostredníctvom
cholínergických neurónov (ACh). Neuróny syntetizujúce ANP ovplyvňujú čin-
nosť oxytocínergických neurónov (OXY). Zvýšená aktivácia baroreceptorov ve-die k uvoľneniu OXY a ANP z neurohypofýzy (NH) do systémovej cirkulácie.
Neuróny syntetizujúce AII modulujú aj procesy spojené s príjmom tekutín.
GN– ganglion nodosum (modifikované podľa McCann a spol., 1997).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 149
Obr. 5.3 Schematické znázornenie zoskupení neurónov modulujúcich procesyspánku a bdenia. ORX– orexín syntetizujúce neuróny pôsobia excitačne, prav-
depodobne aj na základe toho, že inervujú zoskupenia modulačných neuro-transmiterov v mozgovom kmeni aj v hypotalame. VLPO–neuróny ventrola-
terálnej preoptickej oblasti syntetizujúce GABA a galanín pôsobia inhibične naviacero zoskupení neurónov. TM–histamínergické neuróny ventrálnej prema-
milárnej oblasti; BF – cholínergické neuróny bazálneho predného mozgu; A10 –
dopamínergické neuróny ventrálnej tegmentálnej oblasti; LC – locus coeruleus,obsahuje noradrenalínergické neuróny; DR– sérotonínergické neuróny nucleus
raphe dorsalis; TP – cholínergické neuróny mozgového kmeňa; PRF– retikulárna
formácia mostu obsahujúca cholínergické receptory (modifikované podľa Mignota spol., 2002).
róny, ktoré syntetizujú tieto neurotransmitery, sa vyznačujú niekoľkýmišpecifikami:
• nie sú v mozgu veľmi početné,
• tvoria zoskupenia, ktoré sú relatívne ohraničené,
• vysielajú axóny do takmer všetkých oblastí centrálneho nervo-vého systému,

150 NEUROTRANSMITERY
• ich axóny často vytvárajú veľké množstvo kolaterál,
• môžu uvoľňovať neurotransmitery aj prostredníctvom synápstypu en passant, čím pôsobia na veľké množstvo neurónov v okolísynapsy.
Takéto štruktúrne charakteristiky vytvárajú predpoklad na to, že ajrelatívne malý počet neurónov syntetizujúcich modulačné neurotrans-mitery môže modulovať také globálne procesy v nervovom systéme,akými sú spánok a bdenie (tab. 5.1).
Zoskupenie neurónovNeurotransmiter
Stav aktivityv mozgovom kmeni neurónov
Bdenie
Cholínergické neuróny Acetylcholín Aktívne
Locus coeruleus Noradrenalín Aktívne
Nuclei raphe Sérotonín Aktívne
Nucleus talamotuberalis Histamín Aktívne
Non-REM spánok
Cholínergické neuróny Acetylcholín Znížená aktivita
Locus coeruleus Noradrenalín Znížená aktivita
Nuclei raphe Sérotonín Znížená aktivita
REM spánok
Cholínergické neuróny Acetylcholín Aktívne
Nuclei raphe Sérotonín Neaktívne
Koniec REM spánku
Locus coeruleus Noradrenalín Aktívne
Tabuľka 5.1 Prehľad účasti monoamínergických neurónov v regulácii bdeniaa spánku (modifikované podľa Purves, 2004).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 151
Acetylcholín. Neuróny syntetizujúce acetylcholín sa nachádzajúv oblastiach, ktoré sú prepojené na ARAS, ako aj na talamus. Ros-trálna skupina cholínergických neurónov je aktívnejšia v bdelom staveako v spánku. Kaudálne lokalizované cholínergické neuróny sa považujúza súčasť ARAS. Predpokladá sa, že tieto neuróny sa môžu zúčastňovaťna tvorbe špecifických vzruchov, ktoré možno zaznamenať počas REMfázy spánku. Zdá sa teda, že cholínergické neuróny majú nielen úlohuv modulácii bdelosti, ale aj v procesoch „podobným bdelostiÿ počasREM fázy spánku (Webster a Stanford, 2001).Histamín. Histamín môže vyvolávať excitačnú aj inhibičnú odpoveď
neurónov CNS. Z pozorovaní efektu antagonistov histamínových recep-torov sa však zdá, že v CNS prevažuje excitačný efekt. Tento predpokladpodporuje aj zistenie, že neuróny v oblasti zadného hypotalamu (ob-lasť, kde sa nachádzajú aj histamínergické neuróny) sú aktívne v bdelomstave a počas spánku je ich aktivita znížená (Webster a Stanford, 2001).Noradrenalín. Všeobecne sa akceptuje, že noradrenalín uvoľňovaný
v CNS zvyšuje behaviorálnu aktivitu a bdelosť. Tento predpoklad jezaložený na pozorovaniach stimulačného vplyvu látok, ktoré zvyšujúuvoľňovanie noradrenalínu v CNS (napr. amfetamín). Zníženie obsahunoradrenalínu a jeho uvoľňovania v mozgu (napr. po podaní rezerpínu)vyvoláva naopak psychomotorické spomalenie. Neuróny locus coeruleus(tvoria najväčšie zoskupenie noradrenergických neurónov v mozgu) súaktívnejšie v bdelom stave ako počas non-REM fázy spánku. Ich akti-vita je výrazne zvýšená aj počas stresových situácií, na zvládnutie kto-rých je nevyhnutná dostatočná úroveň bdelosti (Webster a Stanford,2001).Sérotonín. Prvé experimentálne pozorovania naznačovali, že séroto-
nín je potrebný na indukciu spánku. Priama stimulácia sérotonínergic-kých neurónov, ako aj podanie prekurzorov sérotonínu však zvyšovalibdelosť. Možno preto predpokladať, že tento neurotransmiter je vý-znamne zapojený v procesoch bdenia, ako aj spánku. Zdá sa, že uvoľ-ňovanie sérotonínu je zvýšené v bdelom stave a znižuje sa počas spánku(Webster a Stanford, 2001).Okrem malomolekulových neurotransmiterov má významnú úlohu
v regulácii bdenia a spánku aj neuropeptid orexín. Orexín uplatňujesvoj vplyv pravdepodobne prostredníctvom modulácie činnosti histamí-nergických, sérotonínergických, noradrenalínergických a iných neurónov(obr. 5.3) (Huang a spol., 2001). Predpokladá sa, že dysfunkcia orexíner-

152 NEUROTRANSMITERY
gického systému sa zúčastňuje napríklad na vzniku narkolepsie (Miedaa Yanagisawa, 2002).
Učenie a pamäť
Jednou z fascinujúcich komplexných funkcií mozgu je jeho schopnosťukladať informácie (vštiepivosť) a možnosť ich vyvolania (výbavnosť).Ako učenie sa označuje proces, ktorý na základe spracovania novýchinformácií nervovým systémom umožňuje modifikovať správanie sa or-ganizmu. Pamäť umožňuje kódovať, uchovávať a vyvolávať získané in-formácie (Purves, 2004).Neurobiologické procesy učenia a pamäte sa začali študovať v re-
latívne jednoduchom organizme u morského mäkkýša (Aplysia). Prisledovaní obranného reflexu sa dokázalo, že významnú úlohu má neuro-transmiter sérotonín. Pri krátkodobej senzitizácii (trvajúcej niekoľkominút až hodín) sérotonín aktiváciou sérotonínových receptorov indu-koval kaskádu procesov, ktorých výsledkom bola kovalentná modifiká-cia proteínov. Opakovaná stimulácia viedla k dlhodobej senzitizácii,ktorú zapríčinila zmena v génovej expresii neurónu, ktorej výsledkombol vznik nových synáps (Kandel, 2001). Za tieto objavy získal roku2000 Eric R.Kandel Nobelovu cenu (Benešová a Höschl, 2005).Ďalší výskum u stavovcov ukázal, že na vzniku pamäte sa význam-
nou mierou zúčastňuje glutamát (obr. 5.4). Sledoval sa vznik dlhodo-bých potenciálov (LTP) v hipokampe, o ktorom sa predpokladá, že ještruktúrou podmieňujúcou konsolidáciu dlhodobej pamäte. Preukázalasa významná úloha pôsobenia neurotransmitera glutamátu, prostred-níctvom ionotropných glutamátergických receptorov pri vzniku LTP.Vyplavenie glutamátu aktivuje AMPA-receptory neurónov hipokampu,čo vedie k depolarizácii postsynaptickej membrány. Depolarizácia zaprí-čiňuje okrem iného aj uvoľnenie Mg2+ z NMDA-receptorov, čo umožnítýmto receptorom reagovať na nasledujúcu stimuláciu zmenou ich prie-pustnosti pre ióny Ca2+. Opakované vyplavenie glutamátu preto môžepodmieniť aktiváciu NMDA-receptorov a výrazne zvýšiť intracelulárnukoncentráciu Ca2+. To vedie ku kaskáde procesov, ktorých výsledkomje zmena v génovej expresii neurónov a novotvorba, resp. modifikáciaexistujúcich synáps (Kandel, 2001).Na procesy spojené s pamäťou a učením však nemožno nahliadať
ako na výsledok pôsobenia iba glutamátu, resp. sérotonínu. Aj keď je

5 Úloha neurotransmiterov v regulácii fyziologických procesov 153
Obr. 5.4 Schematické znázornenie úlohy glutamátu v procesoch tvorby dlho-dobých potenciálov (modifikované podľa Jedlička a spol., 2002).
aktivácia glutamátergických receptorov jedným z kľúčových krokov, navzniku pamätovej stopy sa zúčastňuje interakcia veľkého počtu neuro-transmiterových systémov. Významným sa javí napríklad aj pôsobenieplynného neurotransmitera NO (Inglis a spol., 1998).
Autonómny nervový systém
Autonómny nervový systém (ANS) sa tradične rozdeľuje na sympati-kovú a parasympatikovú časť. Okrem toho možno rozlišovať aj ente-rický, intrakardiálny a nonadrenergický/noncholínergický nervový sys-tém.Základom regulácie činnosti ANS sú pregangliové neuróny lokali-
zované v predĺženej mieche a laterálnych rohoch miechy. Sympatikovéa parasympatikové pregangliové neuróny vysielajú svoje axóny do au-

154 NEUROTRANSMITERY
tonómnych ganglií, kde sa nachádzajú postgangliové neuróny, ktorépriamo inervujú jednotlivé tkanivá. Sympatikové pregangliové neurónyvysielajú relatívne krátke axóny k sympatikovým postgangliovým ne-urónom, ktoré sa nachádzajú v paravertebrálnych a prevertebrálnychgangliách. Parasympatikové pregangliové neuróny vysielajú dlhé axónyku gangliám obsahujúcim postgangliové neuróny, ktoré sa nachádzajúv blízkosti vnútorných orgánov, alebo priamo v ich tkanivách. Axónypostgangliových neurónov netvoria klasické synapsy, ale vytvárajú mno-hopočetné varikozity (synapsy typu en passant), ktoré uvoľňujú ne-urotransmiter do okolia. Uvoľnený mediátor difunduje na relatívneveľkú vzdialenosť, čím môže pôsobiť na väčší počet efektorových bu-niek (obr. 5.5) (Maršala, 1985). Okrem klasických neurotransmiterovuvoľňujú autonómne nervy aj viaceré kotransmitery, ktoré modulujúsynaptický prenos (tab. 5.2).
Sympatikový nervový systém
Sympatikové pregangliové neuróny sú lokalizované v intermediolaterál-nom stĺpci torakolumbálnej miechy. Tieto neuróny sú inervované inter-neurónmi miechy aj descendentnými nervovými dráhami. Interneurónyprenášajú informácie z descendentných dráh, ako aj z periférnych recep-torov (Maršala, 1985). Medzi mozgové oblasti, ktoré priamo inervujúsympatikové pregangliové neuróny, sa zaraďuje nucleus paraventricula-ris hypothalami, kaudálna časť nuclei raphe, noradrenalínergická oblasťA5, ventromediálna a rostrálna ventrolaterálna oblasť predĺženej mie-chy (obsahuje aj adrenalínergické C1-neuróny) (Strack a spol., 1989a,1989b; Gilbey a Spyer; 1993; Sved a spol., 2001; Kerman a spol., 2003).Novšie práce poukazujú aj na priamu inerváciu sympatikových pre-gangliových neurónov z Barringtonovho jadra, ventrálnej časti locussubcoeruleus, laterálneho hypotalamu a neurónov lokalizovaných v bie-lej hmote horných segmentov cervikálnej miechy (Cano a spol., 2000,2001; Seyedabadi a spol., 2006). Medzi neurotransmitery, o ktorých sapredpokladá, že sa zúčastňujú na prenose informácií k sympatikovýmpregangliovým neurónom, patria monoamíny (adrenalín, noradrenalín,sérotonín), peptidy (substanciu P, TRH, metenkefalín, oxytocín, vazo-presín, CGRP, neuropeptid Y) a aminokyseliny (glutamát, GABA, gly-cín) (obr. 5.5) (Dun a spol., 1992, 1993; Hosoya a spol., 1995; Pilowsky

5 Úloha neurotransmiterov v regulácii fyziologických procesov 155
A
Mediátor Receptor Miesto účinku
GnRH GnRH receptor sympatikové gangliá
Dopamín D1, D2, D3, D5 sympatikové gangliá
ATP P-receptory (purínergické) hladké svalstvotráviaceho traktu,močového mechúra(sympatikus)
VIP VIP-receptory postgangliové vláknaparasympatika
B
ACTH cholecystokinín
angiotenzín II luliberín
endorfíny neuropeptid Y
enkefalíny neurotenzín
bombezín sérotonín
GABA somatostatín
galanín substancia P
gastrín žalúdok inhibujúci peptid
Tabuľka 5.2 Modulátory autonómneho nervového systému s vyhranenou loka-lizáciou (A) a prehľad ďalších modulátorov autonómneho nervového systému (B)
(modifikované podľa Mysliveček, 2003).

156 NEUROTRANSMITERY
Obr. 5.5 Schematické znázornenie sympatikových (A) a parasympatikových
(B) dráh. DRG–ganglion zadných rohov miechy; NG– ganglion nodosum;
SPrN– sympatikový pregangliový neurón; SPoN– sympatikový postgangliovýneurón; PSPrN– sympatikový pregangliový neurón; PSPoN–parasympatikový
postgangliový neurón; SIF – small intensely fluorescent cells; Glu – glutamát;IN – interneurón; OXY– oxytocín; AVP–vazopresín; NA–noradrenalín; 5-HT–
sérotonín; DA–dopamín; KA–katecholamíny; NPY–neuropeptid Y.
a spol., 1995; Sun, 1995; Llewellyn-Smith a spol., 1997; Teclemariam-Mesbah a spol., 1997; Stornetta a spol., 2004; Coote, 2005).Sympatikové pregangliové neuróny inervujúce postgangliové neu-
róny sympatikových ganglií uvoľňujú ako neurotransmiter acetylcho-lín, ktorý pôsobí na gangliové nikotínové receptory a modulačne pôso-biace muskarínové receptory (Janig, 2005). Okrem toho sa v sympa-tikových gangliách nachádzajú aj tzv. small intensely fluorescent cells(SIF-bunky), ktoré predstavujú interneuróny, resp. modulačné neuróny,ktoré ovplyvňujú synaptický prenos medzi pregangliovými a postgan-gliovými neurónmi sympatikového nervového systému prostredníctvomuvoľňovania katecholamínov (obr. 5.5A) (Chiba, 1977).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 157
Sympatikové postgangliové neuróny, inervujúce vnútorné orgánya tkanivá, uvoľňujú z nervových zakončení ako hlavný neurotransmiternoradrenalín; okrem toho uvoľňujú aj kotransmitery ATP a neuropep-tid Y (Lundberg a spol., 1990; Westfall a spol., 2002; Pablo Huidobro-Toro a spol., 2004). Určitou výnimkou je sympatiková inervácia potnýchžliaz, kde ako neurotransmiter vystupuje acetylcholín. Dokázalo sa však,že zakončenia sympatikových axónov inervujúcich potné žľazy obsahujúaj enzýmy nevyhnutné pre syntézu katecholamínov. Je preto možné, žetáto špecifická inervácia je zmiešaná (Weihe a spol., 2005).
Parasympatikový nervový systém
Parasympatikové pregangliové neuróny sú lokalizované v predĺženejmieche a v laterálnom stĺpci sakrálnej miechy. Podobne ako sympa-tikové pregangliové neuróny sú aj tieto neuróny inervované interneu-rónmi predĺženej miechy a spinálnej miechy, ako aj descendentnýminervovými dráhami (Maršala, 1985; Shefchyk, 2001). Medzi neurotrans-mitery, ktoré sa zúčastňujú na prenose informácií k parasympatikovýmpregangliovým neurónom, pravdepodobne patria glutamát, acetylcho-lín, GABA, katecholamíny, sérotonín a viaceré neuropeptidy (napr. ga-lanín, TRH) (obr. 5.5B) (Nemoto a spol., 1995; Krowicki, 1996; Wanga spol., 2001; Kaneko a spol., 2002; Xu a spol., 2005).Parasympatikové pregangliové neuróny inervujúce postgangliové ne-
uróny parasympatikových ganglií uvoľňujú ako neurotransmiter acetyl-cholín, ktorý pôsobí na gangliové nikotínové receptory. Muskarínové re-ceptory parasympatikových ganglií majú v tomto prenose modulačnúfunkciu (Myers, 2001).Parasympatikové postgangliové neuróny, inervujúce vnútorné or-
gány a tkanivá, uvoľňujú z nervových zakončení acetylcholín, ktorý pô-sobí na muskarínové receptory.
Enterický nervový systém
Tvoria ho neuronálne siete nachádzajúce sa v stenách tráviaceho traktuod ezofágu až po anus, ako aj v slinných žľazách, pankrease a žlčníku.Tvorí ho veľký počet neurónov lokalizovaných v stene gastrointestinál-neho traktu, ktorých činnosť je modulovaná parasympatikovou a sym-patikovou inerváciou (obr. 5.6) (Hansen, 2003). Na inervácii gastroin-testinálneho traktu sa zúčastňujú aj nitrergické (NO uvoľňujúce) nervy

158 NEUROTRANSMITERY
(Toda a Herman, 2005). Neuróny v gastrointestinálnom trakte synte-tizujú viacero neurotransmiterov. Interneuróny syntetizujú napríkladacetylcholín, sérotonín, GABA, CGRP, VIP, somatostatín a NO (Sche-mann a Neunlist, 2004).
Obr. 5.6 Schéma nervových okruhov tvoriacich enterický nervový systém.DRG–ganglion zadných rohov miechy; GN– ganglion nodosum; PSPr – axón
parasympatikového pregangliového neurónu; SG – sympatikové ganglion; SPr –
axón sympatikového pregangliového neurónu; SPo – axón sympatikového po-stgangliového neurónu; MP–plexus myentericus; SMP–plexus submucosus.
Šípky znázorňujú smer šírenia nervových vzruchov (modifikované podľa Hol-zer a spol., 2001).
Zaujímavý je mechanizmus prenosu senzorických informácií, na kto-rom sa zúčastňujú enterické endokrinné (enterochrómafinné) bunky,ktoré sa nachádzajú v intestinálnej mukóze. Enterochrómafinné bunkyodpovedajú na zmeny v chemickom zložení látok nachádzajúcich sav lúmene čreva uvoľnením neuroaktívnych mediátorov cholecystokinínu

5 Úloha neurotransmiterov v regulácii fyziologických procesov 159
a sérotonínu. Tieto mediátory aktivujú receptory zakončení primárnychaferentných nervov v stene gastrointestinálneho traktu, čo umožní pre-nos informácií do CNS (Raybould, 2002).
Intrakardiálny nervový systém
Komplexnosť regulačných mechanizmov sa uplatňuje aj v precíznomriadení činnosti srdca. Činnosť srdca je regulovaná autonómnym ner-vovým systémom, hormonálne aj prostredníctvom vnútorných regulač-ných mechanizmov. Na autonómnej inervácii srdca sa zúčastňujú ex-trakardiálne aj intrakardiálne lokalizované neuróny (obr. 5.7). Kým súextrakardiálne neuróny a nervové dráhy relatívne dobre opísané, ažv posledných desaťročiach sa začala rozpoznávať rôznorodosť populá-cií intrakardiálne lokalizovaných neurónov a komplexnosť ich nervovýchokruhov (Randall, 2000; Kukanova a Mravec, 2006). Hlavným neuro-transmiterom parasympatikových postgangliových neurónov je acetyl-cholín. Okrem toho viacero populácií neurónov má enzymatickú vý-bavu, ktorá je potrebná pre syntézu biogénnych amínov a rozličnýchneuropeptidov. SIF-bunky v srdci pravdepodobne syntetizujú dopamína sérotonín. Predpokladá sa, že niektoré z veľkobunkových neurónovsyntetizujú noradrenalín a adrenalín (Slavikova a spol., 2003). V in-trakardiálnych neurónoch sa dokázala aj prítomnosť enzýmov pre bio-syntézu histamínu (Singh a spol., 1999). Okrem biogénnych amínov sav neurónoch srdca dokázala aj prítomnosť rôznych neuropeptidov, akosú napr. neuropeptid Y, vazoaktívny intestinálny polypeptid, peptidodvodený od génu pre kalcitonín a tachykiníny (Steele a spol., 1994;Slavikova, 1997; Slavíková a spol., 2005).
Nonadrenergický/noncholínergický (NANC) nervový systém
Moduluje činnosť hladkého svalstva viacerých orgánových systémov(Bennett, 1997). NANC-systém využíva viaceré neurotransmitery.V respiračnom trakte pôsobia excitačne substancia P a neurokinín A,inhibične účinkuje vazoaktívny intestinálny peptid, neuropeptid Y a NO(Widdicombe, 1998). NANC systém sa zúčastňuje aj na procesoch spo-jených s erekciou. Významnú úlohu tu má nitrergická, ale pravdepo-dobne aj peptidergická (VIP, NPY) inervácia corpus cavernosum, ktorej

160 NEUROTRANSMITERY
Obr. 5.7 Schéma motorickej a senzorickej inervácie srdca sympatikovým
a parasympatikovým nervovým systémom (ľavá časť schémy) a detail ner-vových okruhov v srdcových gangliách (pravá časť schémy). A– parapara-
sympatikový postgangliový neurón; B – sympatikový „postgangliovýÿ neurón;C– axón vychádzajúci z neurónu lokalizovaného v inom srdcovom gangliu;
D– interneurón lokálnych nervových okruhov; E – axón inervujúci iné srdcové
ganglion; F – senzorický neurón; DRG–ganglion zadných rohov miechy; GN–ganglion nodosum; PSPr – axón parasympatikového pregangliového neurónu;
PSPo – axón parasympatikového postgangliového neurónu; SG– sympatikové
ganglion; SPr – axón sympatikového pregangliového neurónu; SPo – axón sympa-tikového postgangliového neurónu; SrG– srdcové ganglion; P – viscerosenzorické
dráhy prebiehajúce v nervus vagus; S – viscerosenzorické dráhy prebiehajúcev sympatikových nervoch. Senzitívne neuróny (F) vysielajú svoje axóny aj do
zadných rohov miechy (Verrier a Antzelevitch, 2004), pre prehľadnosť neznázor-
nené. Šípky znázorňujú smer šírenia nervových vzruchov (modifikované podľaArmour, 1999; Randall a spol., 2003; Nozdrachev a spol., 2003).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 161
narušená činnost sa môže zúčastňovať na vzniku erektilnej dysfunkcie(Andersson, 2000; Toda a spol., 2005).
Stresová reakciaB.Mravec, R.Kvetňanský
Aj keď sa pojmy stres a stresom podmienené ochorenia používajúveľmi často, stále neexistuje všeobecne akceptovaná definícia stresu. Tovedie k nejednoznačnosti pojmu stres a nedostatočnému vymedzeniuochorení, na vzniku ktorých sa môže stres zúčastňovať (Pacak, 2000).Stresor sa definuje ako podnet, ktorý vyvoláva stresovú reakciu. Nie-kedy však môže ako stresor pôsobiť paradoxne nedostatok podnetov(Vigaš, 1985). Stresory sú vonkajšie alebo vnútorné podnety, ktoré spô-sobujú také narušenie homeostázy, ktoré prekročí bežné kompenzačnémechanizmy. Následne dochádza k aktivácii stresovej reakcie. Aj keďsa na stresovej reakcii zúčastňuje veľké množstvo systémov, dva z nich,sympatikoadrenálny a hypotalamo-hypofýzo-adrenokortikálny systém,sú kľúčové (Miller a O’Callaghan, 2002; Nicolaidis, 2002).
Špecificita stresovej reakcie
Na zavedení pojmu stres a na jeho širokej popularizácii sa významnoumierou zaslúžil Hans Selye. Definoval stres ako nešpecifickú odpoveďna akúkoľvek záťaž pôsobiacu na organizmus (Selye, 1950, 1974). Vy-chádzal z pozorovaní, že vystavenie experimentálnych zvierat rôznymstresorom viedlo k triáde patologických zmien – zväčšeniu nadobličiek,vzniku gastrointestinálnych ulcerácií a involúcii týmusu (Selye, 1936).Veľké množstvo neskorších štúdií však preukázalo, že odpoveď na pôso-benie stresora je relatívne špecifická. Táto špecificita sa prejavuje špe-cificitou neuroendokrinných reakcií na pôsobenie stresorov. Príkladomsú nasledujúce situácie:
• pôsobenie chladu prednostne zvyšuje plazmatické hladiny norad-renalínu, kým hladiny adrenalínu sa významnejšie nemenia,
• hypoglykémia a hypoxia predstavujú podnety, ktoré vedú k vý-raznému vyplaveniu adrenalínu, pričom hladiny noradrenalínu savýznamnejšie nemenia,

162 NEUROTRANSMITERY
• hemorágia spôsobuje špecifické zvýšenie plazmatických hladínACTH (Pacak a spol., 1998).
Tieto pozorovania viedli k prehodnoteniu Selyeho doktríny nešpe-cifickosti stresovej reakcie. V súčasnosti sa akceptuje, že stresová reak-cia je špecifická reakcia, ktorej priebeh a miera aktivácie jednotlivýchneuroendokrinných systémov zúčastňujúcich sa na odpovedi závisí odvyvolávajúceho činiteľa, špecifického stresora.Stresory možno klasifikovať podľa viacerých kritérií. Jedna z klasi-
fikácií (Myslivecek a Kvetnansky, 2006) rozlišuje:
• fyzikálne stresory (teplo, chlad, žiarenie, hluk, vibrácie), častovyvolávajú bolesť,
• chemické (hypoglykémia, oxidačný stres, otravy, hypoxia),
• psychologické stresory, ktoré odzrkadľujú odpoveď na pôsobeniepodnetov, ktoré už v minulosti vyvolali averzívnu reakciu (obme-dzenie príjmu potravy, spánková deprivácia, emočný stres, depre-sia),
• sociálne stresory ako napríklad nezamestnanosť,
• stresory narušujúce kardiovaskulárnu a metabolickú homeostázu(hemorágia, svalová práca, ortostáza),
• ovplyvňujúce viaceré systémy (imobilizácia, restrain, bolesť, hla-dovanie a smäd, zmeny v osvetlení, nútené plávanie).
Z časového hľadiska sa rozlišujú akútne (jednoduché, intermitentnéa časovo ohraničene pôsobiace) a chronické (intermitentné, dlhodoboalebo stále pôsobiace) stresory. Pri pôsobení stresorov na organizmus jevýznamným faktorom aj ich intenzita (Pacak a Palkovits, 2001).
Neuronálne okruhy stresovej reakcie
Aktivácia mechanizmov stresovej odpovede vedie k zmenám v činnostiviacerých neuronálnych okruhov, ktoré ovplyvňujú činnosť CNS (pod-mieňujú zmeny v správaní sa organizmu), ako aj činnosť periférie (napr.zmeny vo vyplavovaní glukokortikoidov a katecholamínov). Všetky tietozmeny majú za cieľ zvýšiť schopnosť organizmu zachovať homeostázu,

5 Úloha neurotransmiterov v regulácii fyziologických procesov 163
a tým zvýšiť šance na prekonanie stresovej situácie (Tsigos a Chrousos,2002).Stresová reakcia je výsledkom činnosti systému, ktorý ma centrálnu
a periférnu časť (obr. 5.8, 5.9).
Centrálna zložka stresovej reakcie
Podnety z vonkajšieho alebo vnútorného prostredia organizmu, ktorévyvolávajú stresovú reakciu, aktivujú veľké množstvo neurónov mozgu.V spracovaní stresových informácií sú zastúpené neuróny z rôznych ob-lastí CNS, ktoré syntetizujú široké spektrum neurotransmiterov (Ver-metten a Bremner, 2002a). Významnú úlohu majú neuróny syntetizu-júce kortikoliberín, noradrenalín a adrenalín (Roth a spol., 1982; Char-mandari a spol., 2005). Centrálna časť stresovej odpovede je výsledkomčinnosti neurónov lokalizovaných prevažne v hypotalame a mozgovomkmeni:
• parvocelulárne neuróny nucleus paraventricularis (PVN) synteti-zujúce kortikoliberín (CRH),
• magnocelulárne neuróny PVN syntetizujúce vazopresín,
• noradrenergické neuróny predĺženej miechy a mosta (LC/NE sys-tém).
Nucleus paraventricularis hypotalamu ako integračné centrumstresovej reakcie
Jednotlivé funkcie, ako napríklad učenie, pamäť, spracovanie zrakovýchinformácií, regulácia kardiovaskulárneho systému a iné, sa v CNS zabez-pečujú neuronálnymi okruhmi, ktorých zložky sa väčšinou široko dis-tribuujú. Niekedy však možno identifikovať oblasti, v ktorých sa „pre-línajúÿ viaceré, často aj funkčne odlišné neuronálne okruhy. Jednouz takýchto oblastí je zoskupenie neurónov v oblasti hypotalamu, ktorása označuje ako nucleus paraventricularis. PVN je „uzlovýmÿ bodomviacerých systémov, ktoré sa zúčastňujú na regulácii stresovej reakcie(Pacak, 2000).Na základe morfologických kritérií sa PVN rozdeľuje na magnocelu-
lárne, mediocelulárne a parvocelulárne podoblasti. Neuróny PVN synte-tizujú veľké množstvo rôznych neurotransmiterov (napr. angiotenzín II,

164 NEUROTRANSMITERY
Obr. 5.8 Schéma prenosu informácií o pôsobení stresorov. Kognitívne spracú-vané podnety (anticipačné stresory) pôsobia prostredníctvom zrakových, slucho-vých, chuťových, čuchových alebo statokinetických receptorov. Sú spracované
napr. v senzorických (SCx), prefrontálnych (PFCx), orbitofrontálnych (OFCx)
oblastiach mozgovej kôry a v amygdale (Am). Fyzikálne a metabolické podnetypôsobia prostredníctvom somatických a viscerálnych nervových zakončení. In-
formácie z týchto nervových zakončení sú vedené do oblastí mozgového kmeňa
(napr. nucleus tractus solitarii –NTS; locus coeruleus – LC) a následne do vyš-ších centier (napr. hypotalamus, talamus a mozgová kôra). Senzorické cirkum-
ventrikulárne orgány (OVLT – organum vasculosum laminae terminalis; SFO–organum subfornicale; AP – area postrema) umožňujú monitorovať zmeny v che-
mickom zložení plazmy. GN– ganglion nodosum, DRG–ganglion zadných rohov
miechy.

5 Úloha neurotransmiterov v regulácii fyziologických procesov 165
Obr. 5.9 Neuroendokrinná odpoveď v dôsledku pôsobenia stresového podnetu.Spracovanie anticipačných, fyzikálnych a metabolických stresorov ovplyvňuje
činnosť autonómnych oblastí v hypotalame a mozgovom kmeni. Aktivácia ne-
urónov nucleus paraventricularis hypotalamu vedie k uvoľneniu kortikoliberínu(CRH) z ich nervových zakončení v oblasti eminentia mediana. Následne sa uvoľ-
ňuje ACTH z adenohypofýzy, ktorý stimuluje syntézu a sekréciu glukokortiko-
idov v kôre nadobličiek. Aktivácia hypotalamických a kmeňových autonómnychoblastí stimuluje činnosť sympatikových pregangliových neurónov (SPN) v in-
termediolaterálnom stĺpci miechy. Následne dochádza k uvoľneniu katecholamí-nov z drene nadobličiek a sympatikových nervových zakončení. AG–nadoblička;
SG– sympatikové ganglion (modifikované podľa Vermetten a Bremner, 2002b).

166 NEUROTRANSMITERY
CCK, CRH, dynorfín, enkefalín, galanín, NO, oxytocín, TRH, vazopre-sín) (de Greef a spol., 1992; Riva a spol., 1999; Palkovits, 2000; Nylena spol., 2001). Predpokladá sa, že v reakcii na rôznorodé stresory majúvýznačnú úlohu neuróny PVN syntetizujúce CRH, vazopresín a oxyto-cín (Pacak, 2000).Mediálne lokalizované parvocelulárne neuróny PVN, ktoré synteti-
zujú CRH, vysielajú axóny do oblasti eminentia mediana. V tejto oblastiuvoľnený CRH vstupuje do portálneho hypotalamo-hypofýzového ciev-neho systému a v adenohypofýze stimuluje uvoľňovanie ACTH. CRH-neuróny PVN vysielajú axóny aj do autonómnych centier v predĺže-nej mieche a spinálnej mieche, čím sa zúčastňujú na regulácii činnostisympatikoadrenálneho systému, regulácii metabolizmu a ovplyvňovaníbehaviorálnych funkcií (Swanson a spol., 1986).Magnocelulárne neuróny PVN syntetizujú vazopresín a oxytocín.
Axóny týchto neurónov uvoľňujú vazopresín a oxytocín v neurohypo-fýze do systémovej cirkulácie, prostredníctvom ktorej sa transportujú doperiférnych tkanív, ktorých činnosť ovplyvňujú. Aj vazopresín sa zúčast-ňuje na regulácii sekrécie ACTH z adenohypofýzy. Parvocelulárne neu-róny PVN syntetizujúce vazopresín a oxytocín inervujú aj autonómneoblasti predĺženej miechy a spinálnej miechy. Predpokladá sa, že tietodva neurotransmitery sa zúčastňujú na modulácii činnosti autonómnychpregangliových neurónov, čím môžu ovplyvňovať aktivitu autonómnychnervov (McEwen, 2004). Uvedené anatomické charakteristiky umožňujúneurónom PVN regulovať činnosť oboch základných efektorových systé-mov stresovej reakcie, hypotalamo-hypofýzo-adrenokortikálneho a sym-patikoadrenálneho systému.Neuróny PVN prijímajú senzorické (vrátane viscerálnych) infor-
mácie, nociceptívne informácie a informácie prostredníctvom cirkum-ventrikulárnych orgánov (Herman a spol., 2003). Na základe takéhotoanatomicko-funkčného usporiadania môžu neuróny PVN monitorovaťaktuálne podmienky vnútorného aj vonkajšieho prostredia organizmua v prípade ich významných zmien začať adekvátnu reakciu.
Úloha centrálnych katecholamínergických oblastí v stresovejreakcii
Stresové podnety zvyšujú aktivitu noradrenalínergických a adrenalíner-gických neurónov v mozgovom kmeni. Možno však pozorovať rozdiely

5 Úloha neurotransmiterov v regulácii fyziologických procesov 167
v intenzite aktivácie medzi jednotlivými katecholamínergickými oblas-ťami (Pacak a spol., 1998; Pacak a Palkovits, 2001). Výsledkom stre-sovej aktivácie katecholamínergických oblastí je zvýšené uvoľňovaniekatecholamínov v mozgu. Kým neuróny locus coeruleus uvoľňujú nor-adrenalín v rozsiahlych oblastiach CNS, adrenalín sa uvoľňuje hlavnev hypotalame (Stanford, 1995; Palkovits, 1999). PVN je jednou z pro-minentných oblastí, v ktorej sú počas stresu katecholamíny zvýšeneuvoľňované (Pacak a spol., 1992, 1993, 1995a, 1995b; Palkovits a spol.,1992, 1999).Katecholamíny uvoľnené v oblasti amygdaly počas stresových situ-
ácií zlepšujú konsolidáciu pamäte, čo má význam pre budúcu reakciuorganizmu na daný stresor, resp. situáciu vedúcu k jeho pôsobeniu. Akvšak dôjde k nadmernému uvoľneniu katecholamínov v kôrových oblas-tiach (napr. prefrontálnom kortexe), môže to mať za následok kogni-tívnu dysfunkciu. Jedným z príkladov môže byť tzv. „off-lineÿ fenoménzapríčinený masívnym uvoľnením katecholamínov v prefrontálnom kor-texe počas pôsobenia stresu. Dochádza k inhibícii činnosti prefrontál-neho kortexu, ktorý je významne zapojený v rozhodovacích a plánova-cích procesoch. Preto počas intenzívneho stresu môže dôjsť k inhibíciivyšších kognitívnych funkcií a prevahe „primitívnejšíchÿ podkôrovýchreakcií (Arnsten, 1998; Arnsten a Li, 2005).Pretože sa predpokladá účasť dysfunkcie dopamínergického systému
v etiopatogenéze neuropsychiatrických ochorení, venuje sa pozornosť ajúlohe dopamínergických neurónov v stresovej reakcii. Predpokladá sa,že mezokortikálny dopamínergický systém je viac senzitívny na pôso-benie stresu ako mezolimbický/nigrostriátový systém (Flugge a spol.,2004).
Interakcie medzi CRH a katecholamínergickými neurónmi
Vzájomné interakcie medzi neurónmi syntetizujúcimi noradrenalína adrenalín na jednej strane a neurónmi syntetizujúcimi CRH na stranedruhej umožňujú počas pôsobenia stresorov vypracovanie adekvátnejstresovej odpovede (Lehnert a spol., 1998; Dayas a spol., 2001).Glukokortikoidy ovplyvňujú činnosť neurónov locus coeruleus, čím
priamo na centrálnej úrovni modulujú činnosť katecholamínergickéhosystému mozgu. Na druhej strane neuróny locus coeruleus inervujúCRH-neuróny nucleus paraventricularis, čím sa zúčastňujú na regulá-

168 NEUROTRANSMITERY
cii činnosti HPA osi (Pacak a spol., 1996). Tieto interakcie zabezpe-čujú koordináciu činnosti HPA osi a sympatikoadrenálneho systému naúrovni centrálneho nervového systému (Dunn a spol., 2004). Predpo-kladá sa, že dysfunkcia v interakciách medzi neurónmi syntetizujúcimiCRH a monoamíny môže byť jedným z faktorov, ktorý sa zúčastňujena etiopatogenéze afektívnych porúch.
Úloha ďalších neurotransmiterov v stresovej reakcii
Okrem uvedených neurotransmiterových systémov sú v stresovej odpo-vedi zapojené aj iné neurotransmitery. Pozornosť sa venuje štúdiu účastisérotonínu v stresovej odpovedi. Boli opísané zmeny v syntéze a uvoľ-ňovaní sérotonínu po vystavení organizmu stresovému podnetu. Údajetýkajúce sa týchto zmien sú však často protichodné. Údaje o účasti sé-rotonínergickej transmisie v regulácii stresovej odpovede a vplyv antide-presív na túto reguláciu spolu zo zmenami stresovej odpovede u depre-sívnych jedincov naznačujú možnú účasť alterovanej stresovej odpovedev etiopatogenéze depresie (Dunčko a Ježová, 2002; Flugge a spol., 2004).Histamínergické neuróny sú aktívne počas situácií sprevádzaných
aktiváciou sympatikoadrenálneho systému. Receptory H1 a H2 modu-lujú uvoľňovanie „stresových hormónovÿ CRH a vazopresínu z hypota-lamu. Aj viacero stresorov, ako napríklad dehydratácia alebo hypogly-kémia, zvyšujú uvoľňovanie histamínu v prefrontálnom kortexe (Fluggea spol., 2004).V stresovej reakcii pravdepodobne majú dôležitú úlohu okrem CRH
aj iné neuropeptidy. Uvažuje sa o úlohe endogénnych opioidov, neu-rotenzínu, neuropeptidu Y, vazopresínu, oxytocínu, angiotenzínu IIa iných (Jezova a spol., 1998; Vermetten a Bremner, 2002a).
Periférna zložka stresovej reakcie
Periférnu časť systému zúčastňujúceho sa na stresovej odpovedi tvoria:
• sympatikoadrenálny systém,
• periférna časť HPA-systému (Engelmann a Ludwig, 2004; Char-mandari a spol., 2005).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 169
Sympatikoadrenálny systém
Regulačné centrum sympatikoadrenálneho systému tvorí viacero moz-gových štruktúr, z ktorých k najvýznamnejším patria hypotalamickéoblasti (napr. nucleus paraventricularis, laterálny hypotalamus). Počaspôsobenia emočných stresových podnetov je činnosť hypotalamu modu-lovaná prevažne descendentnými dráhami z kôrových oblastí a limbic-kého systému. Naproti tomu pri pôsobení fyzikálnych stresorov je čin-nosť hypotalamu modulovaná prevažne ascendentnými dráhami z moz-gového kmeňa (hlavne z monoamínergických neurónov). Hypotalamusnásledne prostredníctvom descendentých dráh vedúcich do mozgovéhokmeňa a miechy modifikuje činnosť autonómnych oblastí (v mozgo-vom kmeni je to napr. C1-skupina neurónov syntetizujúcich adrenalín,v mieche intermediolaterálny stĺpec, v ktorom sa nachádzajú telá sym-patikových pregangliových neurónov) (Palkovits, 1999).Sympatikoadrenálny systém sa anatomicky a funkčne delí na sympa-
tikoneurálnu zložku a adrenomedulárnu hormonálnu zložku. Aktiváciasympatikoneurálnej zložky vedie k uvoľneniu noradrenalínu zo zakon-čení sympatikových nervov v inervovaných tkanivách. Zvýšená aktivitaadrenomedulárneho systému spôsobuje vyplavenie adrenalínu a v men-šej miere aj noradrenalínu z chrómafinných buniek drene nadobličiekdo systémovej cirkulácie.Katecholamíny uvoľnené počas pôsobenia stresu z drene nadobli-
čiek a sympatikových nervových zakončení ovplyvňujú činnosť väčšinyorgánov tela. Katecholamíny zvyšujú silu kontrakcií srdca, frekvenciuakcie srdca a vyvolávajú aj vazokonstrikciu. Výrazná aktivácia sympati-koadrenálneho systému preto počas stresu spôsobuje zvýšenie krvnéhotlaku prostredníctvom zvýšenia periférnej cievnej rezistencie a zvýše-ného srdcového výdaja. Pri aktivácii sympatikoneurálneho systému do-chádza k redistribúcii krvi so zvýšením prekrvenia svalstva a zníženýmprekrvením splanchnických oblastí a kože. Stresom aktivovaný sym-patikoadrenálny systém zapríčiňuje aj rozšírenie zreníc (mechanizmus,ktorý uľahčuje sledovanie situácie v blízkom okolí), zvyšuje produk-ciu potu (uľahčuje odvádzanie tepla z organizmu do okolia), spôsobujepiloerekciu, inhibuje vylučovanie sodíka v obličkách a moduluje tonuskostrového svalstva. Dochádza aj k zvýšenej agregabilite krvných doš-tičiek (ako protektívny mechanizmus v prípade poranenia). Katechola-míny ovplyvňujú aj energetický metabolizmus stimuláciou glykogeno-

170 NEUROTRANSMITERY
lýzy v pečeni a lipolýzy v tukovom tkanive. Katecholamíny uvoľnenéna periférii počas pôsobenia stresových podnetov majú v konečnom dô-sledku za cieľ pripraviť organizmus na vykonanie intenzívnej svalovejčinnosti na základe redistribúcie a zvýšenej oxygenácie krvi, ako aj do-stupnosti energetických substrátov.Periférne uvoľnené katecholamíny neprechádzajú mozgovo-cievnou
bariérou, preto nemôžu priamo ovplyvniť činnosť neurónov centrálnehonervového systému. V dôsledku pôsobenia stresu sa však uvoľňujú ka-techolamíny z monoamínergických neurónov mozgového kmeňa, ktorézabezpečujú primeranú reakciu na stresový podnet ovplyvnením bde-losti, pozornosti, pamäte a vnímania bolesti (Mravec a Kiss, 2004).
Hypotalamo-hypofýzo-adrenokortikálny systém
Stres vyvoláva aktiváciu HPA-systému, ktorá má za následok zvýše-nie plazmatických hladín glukokortikoidov. Podnety, ktoré vyvolávajúaktiváciu HPA-systému, pôsobia aj prostredníctvom katecholamínergic-kých, sérotonínergických a pravdepodobne aj cholínergických zoskupeníneurónov. Parvocelulárne neuróny v nucleus paraventricularis predsta-vujú oblasť, ktorá integruje stresovú odpoveď v mozgu. Tieto neurónyuvoľňujú kortikoliberín, ktorý sprostredkuje uvoľnenie adrenokortiko-tropného hormónu z kortikotropných buniek adenohypofýzy. Adreno-kortikotropný hormón uvoľnený do systémovej cirkulácie indukuje ste-roidogenézu v kôre nadobličiek a zvyšuje plazmatické hladiny glukokor-tikoidov (Ganong a spol., 1974; Herman a spol., 2003).Zvýšené plazmatické hladiny glukokortikoidov ovplyvňujú činnosť
väčšiny tkanív, ale v porovnaní s katecholamínmi je nástup ich účinkupomalší. Glukokortikoidy stimulujú glukoneogenézu, glykogenogenézua pôsobia proteokatabolicky. Glukokortikoidmi indukované procesy glu-koneogenézy a glykogenogenézy majú za cieľ prispieť k zachovaniuenergetickej rovnováhy narušenej vplyvom procesov indukovaných ka-techolamínmi v počiatočných fázach stresovej reakcie. Glukokortiko-idy ovplyvňujú aj funkciu imunitného systému svojím protizápalovýma imunosupresívnym vplyvom. Predpokladá sa, že dlhodobo zvýšenéplazmatické hladiny glukokortikoidov pri pôsobení chronického stresu samôžu zúčastňovať na vzniku gastrointestinálnych lézií, znížení aktivitysympatikoadrenálneho systému a zvýšenej senzitivite kardiovaskulár-

5 Úloha neurotransmiterov v regulácii fyziologických procesov 171
neho systému na katecholamíny (Sapolsky a spol., 2000; Charmandaria spol., 2004).Glukokortikoidy na rozdiel od katecholamínov prechádzajú mozgo-
vo-cievnou bariérou. V mozgu pôsobia na glukokortikoidové, ale aj mi-neralokortikoidové receptory. Predpokladá sa, že chronický stres môžepráve prostredníctvom centrálneho pôsobenia glukokortikoidov viesťk zmenám v činnosti centrálneho nervového systému, ktoré sa môžuprejaviť aj narušením psychických procesov. Experimentálne údaje po-ukazujú na negatívny vplyv glukokortikoidov na plasticitu neurónov,ako aj na neurogenézu. Predpokladá sa, že znížená plasticita neurónov,pozorovaná hlavne v hipokampe, môže pôsobiť ako jeden z etiopato-genetických faktorov pri vzniku neuropsychiatrických ochorení (Hoschla Hajek, 2001).
Interakcie medzi sympatikoadrenálnym a hypotalamo-hypofý-zo-adrenokortikálnym systémom
Hypotalamo-hypofýzo-adrenokortikálny a sympatikoadrenálny systémnepredstavujú izolované systémy, ale sú anatomicky a funkčne prepo-jené a počas stresu môžu interagovať na rôznych úrovniach. Napríkladhormóny HPA-systému ovplyvňujú aktivitu sympatikoadrenálneho sys-tému na úrovni mozgu aj drene nadobličiek.
Nepriaznivé následky pôsobenia stresu
Na jednej strane akútny stres spúšťa dôležité regulačné mechanizmy,ktoré pomáhajú organizmu prekonať kritické situácie; na druhej stranevšak môže mať pôsobenie dlhodobých, či intenzívnych stresorov ne-priaznivý vplyv na organizmus, čo sa môže prejaviť vznikom viacerýchochorení (Esch a spol., 2002; Charney a Manji, 2004).Predpokladá sa, že dysregulácia stresovej reakcie podmienená chro-
nickým pôsobením stresových situácií sa zúčastňuje na vzniku afektív-nych ochorení, anxiety, anorexie, syndróm inzulínovej rezistencie spo-jený s dyslipidémiou, narušenej činnosti gastrointestinálneho systému.Okrem toho sa predpokladá, že chronický stres sa zúčastňuje na patoge-néze kardiovaskulárnych (hypertenzia, ateroskleróza), autoimunitnýcha nádorových ochorení, ako aj na zvýšenej vnímavosti organizmu nainfekcie (Chrousos a Gold, 1998; DaSilva, 1999; Kiecolt-Glaser a spol.,

172 NEUROTRANSMITERY
2002). Chronické zvýšenie hladín glukokortikoidov podmienené pretr-vávajúcim stresom môže nepriaznivo ovplyvniť štruktúru a funkciu hi-pokampu, čo môže zapríčiniť pamäťové a kognitívne dysfunkcie. Príkla-dom ochorenia, pri ktorom sa predpokladá priama súvislosť s pôsobe-ním intenzívneho stresu, je posttraumatické stresové ochorenie (post-traumatic stress disorder) (Vanitallie, 2002).
BolesťB.Mravec, M. Bernadič
Bolesť možno definovať ako nepríjemný senzorický alebo emočný po-cit spojený s aktuálnym alebo potenciálnym poškodením tkanív. Veľminepríjemný vnem spojený s pôsobením bolestivých podnetov je výsled-kom komplexných mechanizmov, ktoré navzájom interagujú na všet-kých úrovniach nervového systému – od periférie, cez zadné rohy mie-chy až po najvyššie centrá v mozgovej kôre. Bolesť sa líši od klasickýchzmyslových vnemov, pretože sa vyznačuje mnohotvárnosťou (má diskri-minačnú zložku, vyvoláva afektívne, autonómne aj motorické reakcie)a variabilitou (Craig, 2003).Bolesť je väčšinou vyvolaná aktiváciou špecifických nociceptorov
(nociceptívna bolesť). Na základe lokalizácie aktivovaných nociceptorovmožno bolesť rozdeľovať na somatickú a viscerálnu. Bolesť však môževzniknúť aj pri poškodení senzorických nervových vlákien, alebo akonásledok poškodenia štruktúr CNS (neuropatická bolesť). Z časovéhohľadiska sa rozlišuje bolesť akútna a chronická. Líšia sa etiológiou, me-chanizmom a patofyziológiou. Akútna bolesť má ochrannú úlohu a far-makoterapeuticky sa dá relatívne dobre ovplyvniť. Naproti tomu chro-nická bolesť pôsobí maladaptačne a jej terapia je problematická (Millan,1999; Bernadič, 2004).
Prenos nociceptívnych informácií do CNS
Prvým krokom v prenose nociceptívnych informácií je aktivácia špeciali-zovaných senzorických receptorov (nociceptorov) periférnych C-vlákiena Aδ-vlákien. Tieto receptory zahŕňajú mechanoreceptory, chemorecep-tory a termoreceptory. V prípade neuropatickej bolesti však nezávisí odaktivácie periférnych nociceptorov (Besson a Chaouch, 1987; Millan,1999; Bernadič, 2004). Myelinizované Aδ vlákna zodpovedajú za rýchly

5 Úloha neurotransmiterov v regulácii fyziologických procesov 173
prenos tzv. prvej bolesti. Nemyelinizované C-vlákna sa zúčastňujú napomalom prenose tzv. sekundárnej bolesti (Furst, 1999).Periférne senzorické vlákna prenášajú nociceptívne informácie do
zadných rohov miechy (hlavne laminae I a II, ale aj III–V) alebo dosenzorických jadier hlavových nervov. V prenose nociceptívnych infor-mácií medzi primárnymi aferentnými vláknami a neurónmi zadných ro-hov miechy má významnú úlohu glutamát pôsobiaci na ionotropné, akoaj na metabotropné glutamátergické receptory. Experimentálne údajepoukazujú na to, že medzi ďalšie neurotransmitery zapojené v prenosenociceptívnych informácií z periférie do CNS patria napr. substancia P,CCK, CGRP, NO, prostaglandíny, puríny (Millan, 1999; Budai, 2000;Zubrzycka a Janecka, 2000; Webster, 2001). Prenos nociceptívnych in-formácií zo zadných rohov miechy ascendentne k štruktúram mozgu za-bezpečuje niekoľko nervových dráh, ktoré uvoľňujú viacero neurotrans-miterov. Predpokladá sa, že hlavným neurotransmiterom pre prenosz tractus spinothalamicus do talamu, ako aj z tractus spinomesence-phalicus do neurónov šedej hmoty v okolí akvaduktu je glutamát a inéexcitačné aminokyseliny. Uvažuje sa aj o účasti niektorých excitačnepôsobiacich neuropeptidov (substancia P, CCK) (obr. 5.10) (Millan,1999).
Centrálna modulácia prenosu nociceptívnych informácií
Prenos nociceptívnych informácií je modulovaný viacerými oblasťamiCNS, ktoré využívajú široké spektrum neurotransmiterov. Rozoznávajúsa descendentné systémy, ktoré inhibujú alebo facilitujú prenos nocicep-tívnych signálov do mozgu. Neuróny týchto descendentných dráh syn-tetizujú neurotransmitery aminokyseliny, acetylcholín, biogénne amíny(noradrenalín, dopamín, sérotonín), ako aj neuropeptidy (β-endorfíny,dynorfín). Medzi centrálne štruktúry, ktoré modulujú synaptický prenosv zadných rohoch miechy, patria mozgová kôra (frontálna, parietálna,gyrus cinguli), hypotalamus (napr. nucleus paraventricularis, nucleustuberomamillaris, nucleus arcuatus), nucleus parabrachialis, nucleustractus solitarii, sérotonínergické a noradrenalínergické neuróny moz-gového kmeňa a šedá hmota v okolí akvaduktu (Millan, 2002).V nociceptívnych procesoch môže jeden typ neurotransmitera pôso-
biť inhibične a zároveň aj facilitačne. Výsledný účinok neurotransmitera

174 NEUROTRANSMITERY
závisí od miesta, kde je neurotransmiter uvoľnený, ako aj od toho, ktorýpodtyp receptorov aktivuje (obr. 5.10).
Obr. 5.10 Prehľad neurotransmiterov zúčastňujúcich sa na prenose no-ciceptívnych informácií a ich modulácii. ADN– adenozín; AVP–vazopresín;
CB– kanabinoidy; DRG–gangliá zadných rohov miechy, ININ - inhibičné
interneuróny; EXIN – excitačné interneuróny; PN–projekčné neuróny;CCK– cholecystokinín; SP – substancia P; Glu – glutamát; DYN–dynorfín;
MC–melanokortín; 5-HT– sérotonín; NA- noradrenalín; DA–dopamín; Hist –histamín; GAL– galanín; EM– endomorfín; ACh – acetylcholín; Gly – glycín;
ENK– enkefalín; NPFF–neuropeptid FF; OFQ– orfanín FQ; β-END–β-endor-
fín; OXY– oxytocín (modifikované podľa Millan, 2002).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 175
Neuropatická bolesť
Neuropatická bolesť vzniká poškodením nociceptívnych nervových dráh.Môže preukazovať relatívne vysokú rezistenciu proti farmakoterapii.Ako príklad možno uviesť opioidy, ktoré sú veľmi účinné v inhibíciibolesti, ktorá vzniká v dôsledku poškodenia tkanív (napr. pooperačnábolesť). Pri neuropatickej bolesti spojenej s poranením nervov sú všakopioidy často neúčinné. Predpokladá sa, že neuropatická bolesť je vý-sledkom dlhodobej centrálnej senzitizácie alebo hyperexcitability neu-rónov v zadných rohoch miechy, čím dochádza k zvýšenému prenosunociceptívnych signálov do CNS. V procesoch centrálnej senzitizáciepravdepodobne majú dôležitú úlohu viaceré neurotransmitery (napr.glutamát účinkujúci prostredníctvom NMDA-receptorov). Predpokladása, že v etiopatogenéze neuropatickej bolesti majú významnú úlohu ajneuropeptidy nociceptín a nocistatín (Martin a spol., 1998).
Vývin nervového systému
Neurotransmitery a neuromodulátory neplnia iba úlohu molekúl zúčast-ňujúcich sa na prenose informácií v nervovom systéme, ale zúčastňujúsa aj na modulácii diferenciácie a rastu nervového tkaniva počas on-togenézy. Experimentálne údaje poukazujú na to, že neurotransmiterysa zúčastňujú na regulácii morfogenetických dejov, ako sú proliferácia,rast, migrácia, diferenciácia a prežívanie neurálnych prekurzorových bu-niek (Nguyen a spol., 2001).Experimentálne údaje naznačujú, že noradrenalínergické axóny lo-
cus coeruleus a cholínergické axóny bazálneho predného mozgu modu-lujú tvorbu neuronálnych spojov počas ontogenézy nervového systému.Uvažuje sa, že aktivita týchto modulačných systémov závisí od vonkaj-ších aj vnútorných vplyvov pôsobiacich na organizmus. Výsledná orga-nizácia nervového systému je potom výsledkom interakcie genetickýchfaktorov a okolitého prostredia, na ktorej sa zúčastňujú aj neurotrans-miterové systémy (Singer, 1986).Noradrenergický systém sa zúčastňuje na regulácii vývoja Cajal-
Retzius buniek, ktoré sú prvými neurónmi, ktoré vznikajú v oblas-tiach budúcej mozgovej kôry. Tieto bunky pravdepodobne tvoria pod-klad pre migráciu novovznikajúcich neurónov a vrstvovité usporiada-nie mozgovej kôry. Predpokladá sa, že dopamínergické neuróny sú vý-

176 NEUROTRANSMITERY
znamne zapojené v regulácii vývinu motorických a kognitívnych okru-hov mozgu. Sérotonínergické neuróny majú úlohu morfogénu v nervo-vom systéme. Pravdepodobne sa zúčastňujú na regulácii diferenciácieneurónov v prednom mozgu (Herlenius a Lagercrantz, 2001).Na význam monoamínov v procesoch vývoja nervového systému po-
ukazuje aj skutočnosť, že ich syntéza sa začína v embryonálnom tkaniveešte pred diferenciáciou zárodočných buniek na neuróny. Na perifériikatecholamíny modulujú činnosť srdca pred tým, ako sa vyvinie jehosympatiková inervácia (Ebert a Thompson, 2001). Monoamíny vo vyví-jajúcom sa nervovom systéme môžu ovplyvňovať aj formovanie spojenímedzi neurónmi typu „gap junctionÿ (Rorig a Sutor, 1996).Acetylcholín pravdepodobne moduluje vývin nervových štruktúr
v gangliách zadných rohov miechy. Predpokladá sa, že acetylcholín mo-duluje vývin axónov ovplyvňovaním ich mobility a rastu. Uvažuje saaj o tom, že acetylcholín môže ovplyvňovať interakcie medzi neurónmia okolitou gliou počas vývinu CNS (Biagioni a Tata, 2000).Predokladá sa, že aj plynný neurotransmiter oxid dusnatý sa vý-
znamne zúčastňuje na procesoch spojených s ontogenézou nervovéhosystému. Experimentálne údaje poukazujú na modulačnú úlohu NOv procesoch spojených s arborizáciou dendritov (Inglis a spol., 1998).Neurotransmitery môžu vo vyvíjajúcom sa mozgu vyvolávať odlišnú
odpoveď neurónov v porovnaní s tou, aká sa pozoruje vo vyzretom ner-vovom systéme. GABA a glycín predstavujú v maturovanom nervovomsystéme hlavné inhibičné neurotransmitery. Vo vyvíjajúcom sa mozgupôsobia však excitačne, pretože nezrelé neuróny majú vysokú intrace-lulárnu koncentráciu Cl−. Obsadenie receptorov pre GABA a glycínpreto vedie k prestupu iónov Cl− extracelulárne, čo vedie k depolarizá-cii membrány neurónu (Herlenius a Lagercrantz, 2004).Predpokladá sa, že okrem uvedených neurotransmiterov a neuro-
modulátorov sa na procesoch spojených s reguláciou vývoja nervovéhosystému zúčastňujú aj kalcium-viažuce proteíny. Zistilo sa napríklad,že expresia paralbumínu v mieche počas ontogenézy nervového systémupreukazuje časové a priestorové zmeny (Clowry a spol., 1997). Zmenyv expresii parvalbumínu primárnymi senzorickými neurónmi sa môžezúčastňovať na formovaní reflexných okruhov (Clowry a spol., 2000).Predpokladá sa, že narušenie činnosti neurotransmiterových systé-
mov počas prenatálneho vývoja CNS sa môže zúčastňovať na vznikuviacerých ochorení nervového systému (Levitt a spol., 1998).

5 Úloha neurotransmiterov v regulácii fyziologických procesov 177
Oxygenácia organizmu
Kyslík predstavuje esenciálnu molekulu pre organizmy. Bunky nervo-vého systému sú obzvlášť citlivé na nedostatok kyslíka (Bunn a Poy-ton, 1996). Preto živočíchy disponujú sofistikovanými systémami, ktorézabezpečujú precízne monitorovanie oxygenácie tkanív (Nattie, 1999).V procesoch monitorovania oxygenácie tkanív sú významne zapo-
jené neuronálne aj neneuronálne bunky, ktoré syntetizujú monoamíny.Tak napr. bunky v glomus caroticum, ktoré vytvárajú periférny che-mosenzorický orgán, využívajú na prenos signálu aj katecholamíny(Oomori a spol., 1994; Nurse, 2005). Uvažuje sa aj o možnom mo-nitorovaní oxygenácie krvi chrómafinnými bunkami drene nadobličiek(Mravec, 2005). Na monitorovaní oxygenácie sa zúčastňujú okrem peri-férnych aj centrálne monoamínergické bunky. Patria medzi ne noradre-nalínergické, sérotonínergické a histamínergické neuróny (Haxiu a spol.,2001).Zdá sa teda, že monoamínergické bunky v organizme vytvárajú
sieť senzorov, ktoré monitorujú oxygenáciu periférnych tkanív a CNS.Takéto redundantné usporiadanie senzorov umožňuje zabezpečiť adek-vátnu reakciu na hypoxiu aj v prípade, že niektoré zo senzorov bynemohli plniť svoju funkciu. Ďalším dôvodom zapojenia monoamíner-gických buniek môžu byť práve monoamíny, ktoré počas hypoxickýchsituácií uvoľňujú. Monoamíny sa významne zúčastňujú na zvládnutí hy-poxickej situácie. Periférne uvoľnené katecholamíny z drene nadobličiekumožňujú organizmu uskutočniť adekvátnu motorickú reakciu potrebnúna odstránenie hypoxickej situácie. Centrálne uvoľnené monoamíny sazasa zúčastňujú na primeranom spracovaní informácií a zodpovedajú-com stupni reaktivity CNS. Aj monoamínergické neuróny sa zúčastňujúna koordinácii respiračnej aktivity, činnosti sympatikového a parasym-patikového nervového systému a adekvátnom prísune krvi do svalstva(Ballantyne a Scheid, 2001; Haxhiu, 2001).
Regulácia telesnej teploty
Telesnú teplotu reguluje integrovaná sieť neuronálnych spojení medzineurónmi hypotalamu, limbického systému, retikulárnej formácie, mie-chy a sympatikových ganglií. Hlavné termoregulačné centrum je loka-lizované v hypotalame (Boulant, 2000). Na modulácii telesnej teploty

178 NEUROTRANSMITERY
sa zúčastňujú viaceré zoskupenia neurónov, ktoré uvoľňujú celé spek-trum neurotransmiterov. Jeden z prvých experimentov, ktorý preukázalvplyv chemickej látky, ktorá patrí medzi neurotransmitery, demonštro-val prechodnú, ale intenzívnu hypotermiu po podaní adrenalínu do moz-gových komôr (Myers, 1984). Výsledky ďalšieho výskumu naznačujú, žev termoregulačných procesoch majú významnú úlohu dopamín, norad-renalín a sérotonín (Cuddy, 2004). Predpokladá sa aj účasť niektorýchneuropeptidov. Okrem dobre známeho vplyvu TRH na termoregulačnéprocesy sa uvažuje aj o účasti ACTH, α-melanocyty stimulujúceho hor-mónu, vazopresínu, cholecystokinínu a opioidov (Myers, 1984; Szelenyi,2001; Richmond, 2003; Tatro a Sinha, 2003; Bodnar a Klein, 2004).Vazopresín a α-melanocyty stimulujúci hormón (spoločne s glukokorti-koidmi) sú na základe ich vplyvu na termoreguláciu označované ako en-dogénne antipyretiká. Tieto látky sú schopné inhibovať zvýšenie telesnejteploty, ktoré vyvoláva pôsobenie pyrogénov. αMSH je jedno z najúčin-nejších endogénnych antipyretík (Starowicz a Przewlocka, 2003).Podobne ako striedanie spánku a bdenia aj telesná teplota má cir-
kadiánny rytmus. Predpokladá sa, že na vzájomnej koordinácii regulá-cie teploty a stavu bdelosti sa zúčastňujú spoločné neurotransmiterovésystémy. Neprekvapuje preto zistenie, že v regulácii telesnej teploty súpravdepodobne zapojené aj neurotransmitery regulujúce spánok a bde-nie. Okrem uvedeného sérotonínu je to aj histamín a orexín (Yoshimichia spol., 2001).Viacero experimentálnych nálezov poukazuje na význam cholíner-
gickej transmisie v regulácii telesnej teploty. Predpokladá, sa, že mus-karínové receptory nachádzajúce sa v termoregulačných centrách hy-potalamu sa zúčastňujú na regulácii telesnej teploty. Podporuje to ajzistenie, že systémové podanie centrálne účinkujúcich muskarínovýchagonistov vyvoláva výrazný pokles telesnej teploty (Wess a spol., 2004).Zistilo sa, že kým stimulácia, ktorá zvyšuje extracelulárnu koncetráciuacetylcholínu v nucleus paraventricularis hypothalami, vedie k zníženiutelesnej teploty, zvýšená cholínergická stimulácia v nucleus supraopticushypothalami telesnú teplotu zvyšuje (Takahashi a spol., 2001).
Starnutie a nervový systém
Starnutie organizmu cicavcov sprevádzajú významné štruktúrne a funk-čné zmeny takmer všetkých orgánov a systémov, pričom nervový systém

5 Úloha neurotransmiterov v regulácii fyziologických procesov 179
nie je výnimkou (Raz, 2001). Neuróny môžu na tieto zmeny reagovaťadaptáciou, alebo môžu podľahnúť kaskáde neurodegeneratívnych pro-cesov.Starnutie sa prejavuje aj zmenami v činnosti neurotransmiterových
systémov. Za jeden z najvýznamnejších prejavov zmien v aktivite gluta-mátergického systému sa pokladá pokles počtu NMDA-receptorov po-čas starnutia. Predpokladá sa, že starnutie môže podmieňovať aj na-rušenie interakcií medzi glutamátom a inými neurotransmitermi (napr.GABA, dopamín) (Segovia a spol., 2001).Dostupné údaje poukazujú na to, že narušenie neurotransmitero-
vých systémov môže byť aspoň čiastočne zodpovedné za niektoré beha-viorálne abnormality súvisiace so starnutím. Viac-menej mierne naruše-nie neurochemických parametrov však počas starnutia ostro kontrastujes výrazným narušením pozorovaným pri ochoreniach CNS. Preto sa vovšeobecnosti usudzuje, že neexistuje jednoznačná závislosť medzi naru-šením neurotransmiterových systémov v dôsledku starnutia a behavi-orálnym deficitom. Predpokladá sa, že neurochemický deficit vznikajúcipočas starnutia sa môže zúčastňovať aj na znížení neuroplasticity. Tomôže limitovať schopnosť CNS reagovať adaptívne na farmakologickéintervencie. Starnutie môže znížiť aj regeneračnú schopnosť CNS priochoreniach postihujúcich nervový systém (Strong, 1998).

Úloha neurotransmiterovpri ochoreniach nervového systému
B. Mravec, B. Kukanová, J. Pečeňák
Poznatky o funkciách neurotransmiterových systémov umožnili as-poň čiastočne pochopiť etiopatogenézu niektorých neurologických, neu-ropsychiatrických a neurodegeneratívnych ochorení. Významným prí-spevkom v tomto procese bolo odhalenie mechanizmu účinku niektorýchliekov účinných pri liečbe rôznych psychických a neurologických porúch(napr. neuroleptiká, antidepresíva). Výsledkom je cielený vývoj novýchliekov, či liekových skupín. Látky používané na liečbu neuropsychiatric-kých ochorení účinkujú prostredníctvom zásahu do procesov spojenýchso syntézou, uvoľňovaním, spätným vychytávaním, biodegradáciou neu-rotransmiterov, ako aj ich priamym pôsobením na receptory (Carlsson,1999). V súčasnosti sa vo výskume etiopatogenézy neuropsychiatrickýchochorení sústreďuje stále viac pozornosti na úlohu neurotransmiterov,resp. neuromodulátorov zo skupiny neuropeptidov. Neuropeptidy častovystupujú ako kotransmitery, ktoré modulujú efekt klasických (malo-molekulových) neurotransmiterov (Holsboer, 2003).K aspoň čiastočnému objasneniu mechanizmov vzniku neuropsy-
chiatrických ochorení prispievajú do značnej miery experimenty na la-boratórnych zvieratách. Súčasné poznatky, často získané práve z ani-málnych experimentov, naznačujú možné mechanizmy vzniku týchto
180

6 Úloha neurotransmiterov pri ochoreniach nervového systému 181
ochorení a smery ďalšieho výskumu. Od komplexného pochopenia eti-opatogenézy a patofyziologických procesov, ktoré sa zúčastňujú navzniku a udržiavaní neuropsychiatrických ochorení, sme však stálevzdialení. Preto v nasledujúcom texte budú uvedené pri jednotlivýchochoreniach aj informácie, ktoré majú často charakter hypotéz. Ichsprávnosť môže určiť až ďalší výskum a predovšetkým overenie novýchpoznatkov v klinickej praxi.
Neurodegeneratívne ochoreniaB.Mravec
Neurodegeneratívne ochorenia tvoria veľkú skupinu ochorení. Preniektoré z nich je charakteristický akcelerovaný zánik skupín neuró-nov, ktoré syntetizujú určitý neurotransmiter. Napríklad pri Alzheime-rovej chorobe dochádza k výraznému zániku cholínergických neurónov;pri Parkinsonovej chorobe zanikajú prednostne dopamínergické neuróny(Mattson a spol., 2002).Veľká pozornosť sa venuje štúdiu vekom podmieneného poklesu poč-
tu dopamínergických neurónov v substantia nigra a poklesu dopamíner-gickej inervácie striata (Palmer a Remosky, 1993). Kým pri narodenísubstantia nigra obsahuje približne 400 tisíc dopamínergických neuró-nov, vo veku 60 rokov je to priemerne iba 250 tisíc. U pacientov s Parkin-sonovou chorobou je tento úbytok dopamínergických buniek zrýchlený– vo veku 60 rokov je ich iba 60–120 tisíc (obr. 6.1). Predpokladá sa,že na zvýšenej vulnerabilite neurónov niektorých neurotransmiterovýchsystémov (prevažne dopamínergického) sa zúčastňujú excitotoxické me-chanizmy (Raz, 2001).Pokles počtu neurónov sa týka aj noradrenalínergických neurónov
v locus coeruleus. So zvyšujúcim sa vekom sa však pozoroval vzostupaktivity monoaminooxidázy. Predpokladá sa, že zvýšená biodegradáciamonoamínov sa môže zúčastňovať na znížení množstva noradrenalínuaj sérotonínu a viesť k zmenám v činnosti neuroendokrinných systé-mov hypotalamu, ako aj striedania spánku a bdenia. Na kognitívnomdeficite pozorovanom v súvislosti zo starnutím a Alzheimerovou cho-robou sa pravdepodobne významne zúčastňuje zmenená činnosť cho-línergického systému. Štúdie preukázali, že v súvislosti zo starnutímu pacientov s Alzheimerovou chorobou dochádza napríklad k zmenám

182 NEUROTRANSMITERY
Obr. 6.1 Úbytok počtu dopamínergických neurónov v oblasti substantia nigra(u pacientov s Parkinsonovou chorobou dochádza k ich rýchlejšiemu úbytku)
(Roth a Růžička, 1998).
v transporte cholínu, uvoľňovaní actylcholínu a zmenám génovej ex-presie nikotínových aj muskarínových receptorov (Terry a Buccafusco,2003). U pacientov s Alzheimerovou chorobou sa dokázal pokles poč-tu cholínergických neurónov v oblastiach bazálneho predného mozgu(Rehman a Mason, 2001).
Parkinsonova choroba
Parkinsonova choroba je neurodegeneratívne ochorenie prejavujúce samotorickými poruchami, medzi ktoré patria tremor, bradykinéza, rigi-dita a poruchy posturálnych funkcií (Grafton, 2004). Výskum etiopato-genézy Parkinsonovej choroby predstavuje jeden z najlepších príkladovuplatnenia experimentálne získaných poznatkov v liečbe narušenej fun-kcie CNS. Pozorovanie vplyvu farmakologicky navodenej deplécie dopa-mínu spolu so zistením signifikantného poklesu počtu dopamínergickýchneurónov v oblasti substantia nigra pri autopsii v mozgoch pacientovs Parkinsonovou chorobou viedli k zavedeniu liečby tohto ochorenia

6 Úloha neurotransmiterov pri ochoreniach nervového systému 183
pomocou prekurzora dopamínu – látky L-DOPA (Kordower a Goetz,1999; Roe, 1999).Predpokladá sa, že idiopatická forma Parkinsonovej choroby je za-
príčinená pôsobením genetických aj environmentálnych vplyvov. Vý-sledkom je zánik dopamínergických neurónov v substantia nigra, ktorývedie ku klinickým symptómom ochorenia (Steece-Collier a spol., 2002).Uvažuje sa, že na zániku dopamínergických neurónov v substantia nigrasa zúčastňujú toxické deriváty dopamínu, kyslíkové radikály, exogénnetoxické látky a iné faktory, ktoré vedú cez kaskádu procesov k apoptózetýchto buniek (Maguire-Zeiss a spol., 2005). Na degenerácii dopamíner-gických buniek sa pravdepodobne zúčastňuje aj interakcia medzi do-pamínom a α-synukleínom, narušená činnosť ubikvitínového systémupodmienená alterovaným proteínom parkinom, ako aj narušenie čin-nosti mitochondriálneho komplexu I (Steece-Collier a spol., 2002; Daw-son a Dawson, 2003). V etiopatogenéze Parkinsonovej choroby a inýchneurodegeneratívnych ochorení sa uvažuje aj o pôsobení NO. Predpo-kladá sa, že NO sa môže zúčastňovať na narušení mitochondriálnehotransportného systému elektrónov, čo môže zapríčiniť alteráciu energe-tického metabolizmu neurónov (Carreras a spol., 2004; Duncan a Hea-les, 2005).Klinické symptómy Parkinsonovej choroby sú podmienené zrých-
leným zánikom pigmentových buniek v substantia nigra, ktoré synte-tizujú neurotransmiter dopamín (obr. 6.1). Následkom toho dochádzak deficitu dopamínu v oblasti bazálnych ganglií (obr. 6.2). Okrem bu-niek v oblasti substantia nigra dochádza u pacientov s Parkinsonovouchorobou k neurodegeneratívnym zmenám aj v amygdale, substantia in-nominata, locus coeruleus (noradrenalínergické neuróny), VTA (dopa-mínergické neuróny), nuclei raphe (sérotonínergické neuróny), nucleusdorsalis nervi vagi a intermediolaterálnom stĺpci miechy (nachádzajúsa tam sympatikové pregangliové neuróny) (Lang a Lozano, 1998). Ažv poslednom období sa ukázalo, že pri Parkinsonovej chorobe dochádzaaj k zániku sympatikových zakončení inervujúcich srdce, čo môže pod-mieňovať autonómnu dysreguláciu činnosti kardiovaskulárneho systému(Goldstein a spol., 2000, 2005).Zachovaná receptorová odpoveď na dopamín pri Parkinsonovej
chorobe je predpokladom úspešnej liečby prekurzorom dopamínu(L-DOPA), dopamínergickými agonistami, látkami zvyšujúcimi uvoľňo-vanie dopamínu a inhibítormi enzýmov zúčastňujúcimi sa na biodegra-

184 NEUROTRANSMITERY
Obr. 6.2 Schéma zobrazujúca okruhy bazálnych ganglií, ktorých činnosť je
u pacientov s Parkinsonovou chorobou narušená. Zánikom dopamínergických
neurónov v substantia nigra dochádza k dysregulácii týchto okruhov. Hrúbkačiar vyjadruje aktivitu danej nervovej dráhy. SNc – substantia nigra, pars com-
pacta; SNr – substantia nigra, pars reticularis; GPi – globus pallidus internus;
GPe – globus pallidus externus; STN–nucleus subthalamicus; PPN–nucleus pe-dunculopontinus; CM–nucleus centromedianus thalami; VA–nucleus ventra-
lis anterior thalami; VL – nucleus ventralis lateralis thalami; D1R, D2R–dopa-mínergické receptory (modifikované podľa Siegel a spol., 2006).
dácii dopamínu. Cieľom všetkých týchto postupov je zvýšiť množstvodopamínu v synapsách, resp. zvýšiť stimuláciu postsynaptických do-pamínergických receptorov v oblasti bazálnych ganglií (Colcher, 2002).Pretože v bazálnych gangliách pri deficite dopamínu dochádza k relatív-

6 Úloha neurotransmiterov pri ochoreniach nervového systému 185
nej prevahe pôsobenia cholínergického systému, používajú sa na liečbuParkinsonovej choroby aj anticholínergiká (Ježová, 1998). Okrem far-makologickej liečby sa využívajú ablačné zásahy (vyradenie časti talamualebo globus pallidus) a hĺbková elektrická stimulácia (v oblasti nuc-lei thalamici, globus pallidus internus, nucleus subthalamicus). Expe-rimentálne sa uskutočňuje aj transplantácia dopamínergických buniek(William, 2001).Okrem Parkinsonovej choroby sa rozpoznáva aj parkinsonovský syn-
dróm, ktorý môže byť zapríčinený postsynaptickým defektom na úrovnidopamínergických receptorov v corpus striatum (napr. po vaskulár-nom, toxickom alebo poúrazovom poškodení). Špecifickým prípadomv rámci sekundárneho parkinsonovského syndrómu je blokáda dopa-mínergických receptorov navodená látkami s neuroleptickým účinkom(Höschl a spol., 2004).
SchizofréniaJ.Pečeňák, B.Mravec
Schizofrénia je psychotické ochorenie, ktoré zasahuje celú osobnosťpostihnutého človeka s dominujúcimi prejavmi v oblasti vnímania, mys-lenia, emotivity, vôle a konania. Pre schizofréniu je charakteristická nie-len mnohotvárnosť psychopatogických prejavov, ale aj rôznorodý prie-beh a reakcia na liečbu. Ochorenie je klinicky charakterizované dvomahlavnými skupinami symptómov. Medzi pozitívne symptómy zaraďu-jeme halucinácie, bludy a dezorganizované myslenie a konanie. Nega-tívne symptómy sú menej nápadné, ale veľmi dôležité pre prognózuochorenia. Patrí sem napr. nedostatok vôľovej aktivity, sociálna izo-lácia a zanedbávanie starostlivosti o seba. Dôležité sú však aj ďalšiepríznaky. Narušenie kognitívnych funkcií (kognitívny syndróm), medziktoré patrí napr. porucha pracovnej pamäte, poruchy tzv. výkonovýchfunkcií, alebo narušenie plynulosti reči, považujú v súčasnosti niektoríautori za najdôležitejšiu súčasť ochorenia. Aj často sa vyskytujúce de-presívne príznaky majú dôležitý klinický význam (Pečeňák a spol., 2005;Coyle, 2006).Predpokladá sa, že schizofrénia je výsledkom multifaktoriálneho pô-
sobenia genetických a environmentálnych faktorov (Miyamoto a spol.,2003; Sawa a Snyder, 2003). V mozgoch pacientov so schizofréniou sa

186 NEUROTRANSMITERY
zistili viaceré anatomicko-histologické abnormality, medzi ktoré patrínapríklad redukcia počtu neurónov a objemu mozgu, malé lézie v ob-lasti amygdaly, hipokampu a prefrontálnej kôry a zväčšenie mozgovýchkomôr (Crow, 1980; Webster, 2001). Funkčné vyšetrovacie metódy pre-ukázali abnormality v činnosti funkčných okruhov zahrňujúcich pre-frontálny kortex, talamus, limbické štruktúry a cerbelum (Andreasena spol., 1999; Coyle, 2006).Predpokladá sa, že pri schizofrénii dochádza k narušeniu viacerých
neurotransmiterových systémov (napr. dopamínergického, sérotoníner-gického, noradrenalínergického, glutamátergického) (Tsai a Coyle, 2002;Wong a Van, 2003).Pri štúdiu etiopatogenézy schizofrénie sa spomedzi neurotransmi-
terových systémov najväčšia pozornosť sústredila na úlohu dopamínu.Podnetom pre formuláciu tzv. dopamínovej teórie schizofrénie bolo po-zorovanie, ktoré ukázalo, že neuroleptiká s antidopamínergickým účin-kom potláčajú psychotické symptómy. Túto teóriu podporovalo aj pozo-rovanie výskytu extrapyramídového syndrómu, ktorý vznikal ako ved-ľajší nežiaduci účinok neuroleptickej liečby. Extrapyramídový syndrómbol prejavom narušenej regulácie motoriky v dôsledku blokády dopamí-nergickej neurotransmisie v okruhoch bazálnych ganglií. Ďalším pod-kladom uvedenej teórie bola indukcia psychotických symptómov popodaní agonistov dopamínu (napr. amfetamínu). Pôvodne sa predpo-kladalo, že etiopatogenetickým podkladom schizofrénie je globálna hy-peraktivita dopamínergického systému v mozgu. V súčasnosti sa všakuvažuje o tom, že dochádza k dopamínergickej hypofunkcii v oblastiprefrontálnej mozgovej kôry a k dopamínergickej hyperfunkcii v pod-kôrových oblastiach (obr. 6.3) (Meyer-Lindenberg a spol., 2002). Podľatejto hypotézy významnú úlohu majú dopamínergická mezokortikálnadráha a mezolimbická dráha. Predpokladá sa, že kým hyperaktivita do-pamínergickej transmisie v mezolimbickej dráhe vedie k prejavom po-zitívnych symptómov, hypoaktivita v mezokortikálnej dráhe má vzťahk negatívnym symptómom. Hoci údaje o obsahu dopamínu v mozgupacientov so schizofréniou nie sú konzistentné, na jeho úlohu v etiopa-togenéze schizofrénie poukazujú viaceré štúdie. Údaje získané z expe-rimentov, v ktorých sa podával rádioaktívne značený antagonista do-pamínových receptorov, naznačujú, že u pacientov so schizofréniou jedopamínergický systém skutočne v oblasti bazálnych ganglií aktívnejšív porovnaní s kontrolnou skupinou (Abi-Dargham a spol., 2000).

6 Úloha neurotransmiterov pri ochoreniach nervového systému 187
Obr. 6.3 Prefrontálny-striatový model schizofrénie. Predpokladá sa, že naru-šená činnosť prefrontálnych glutamátergických neurónov vedie k strate fyzio-
logickej inhibície dopamínergickej aktivity v subkortikálnych oblastiach (napr.v striate). Hrúbka čiar vyjadruje aktivitu danej nervovej dráhy. Glu – glutamát-
ergické neuróny; GABA–GABAergické neuróny; MCDA–mezokortikálne do-pamínergické neuróny; MLDA–mezolimbické dopamínergické neuróny; PPT–
pedunkulo-pontínne glutamátergické neuróny (modifikované podľa Laruelle
a spol., 2003; Winterer a Weinberger, 2004).
Predpokladá sa, že na dysfunkcii dopamínergického systému prischizofrénii sa môže zúčastňovať aj zmenená rýchlosť metabolizmu do-pamínu. Dopamín uvoľnený do synaptickej štrbiny sa spätne vychytávadopamínovým transportérom (DAT) do nervového zakončenia, z kto-rého sa pôvodne uvoľnil, a následne je metabolizovaný prostredníc-tvom enzýmu monoaminooxidázy (MAO). Extraneuronálne je dopamínmetabolizovaný prostredníctvom MAO a katechol-O-metyltransferázy(COMT) na kyselinu homovanilínovú. V prefrontálnom kortexe na roz-diel od mezencefalických oblastí DAT chýba. Preto dopamín uvoľnenýv prefrontálnom kortexe nie je spätne vychytávaný dopamínergickýminervovými zakončeniami, ale ostáva v synaptickej štrbine a difunduje

188 NEUROTRANSMITERY
do okolia, kde je metabolizovaný a deaktivovaný prostredníctvom en-zýmu COMT. Predpokladá sa, že v etiopatogenéze schizofrénie môžemať úlohu aj polymorfizmus génu, ktorý kóduje COMT. Jedna takátobodová mutácia (polymorfizmus Val→Met v pozícii 108/158 sekven-cie peptidu) spôsobuje, že enzým COMT preukazuje až štvornásobnevyššiu aktivitu. Dôsledkom je zrýchlený metabolizmus dopamínu v pre-frontálnej kôre a pokles jeho koncentrácie. Zdá sa však, že rozdiel v ak-tivite COMT závislý od bodovej mutácie nezostáva obmedzený len naprefrontálnu kôru, kde je význam aktivity COMT podstatný. Mecha-nizmom nepriameho ovplyvnenia smerujúceho z mozgovej kôry do pod-kôrových štruktúr prostredníctvom GABAergických neurónov dochá-dza aj k ovplyvneniu dopamínergického systému v mezencefale, čo samanifestuje zvýšenou aktivitou dopamínergickej transmisie v podkô-rových oblastiach. Tieto nálezy významne menia predstavu o primár-nom zdroji „schizofrenogénnejÿ dopamínergickej transmisie s pôvodnepredpokladaným primárnym zdrojom v mezencefale (mezokortikálnadráha). Predpokladá sa, že primárna dysfunkcia je lokalizovaná v pre-frontálnej kôre, ktorá zodpovedá za kognitívne poruchy, ktoré sú podľaniektorých názorov v centre symptomatiky schizofrénie (Akil a spol.,2003; Perlman a spol., 2004).Experimentálne údaje poukazujú na to, že zmeny v koncentrácii
dopamínu v synaptických štrbinách majú za následok na jednej straneexcesívnu stimuláciu dopamínergických D2-receptorov v subkortikál-nych oblastiach a na druhej strane zníženú stimuláciu D1-receptorovv prefrontálnej kôre. Znížená stimulácia D1-receptorov vedie k ich zvý-šenej denzite v prefrontálnej mozgovej kôre (Abi-Dargham, 2004).V etiopatogenéze schizofrénie sa uvažuje aj o úlohe interakcií medzi
dopamínergickým a glutamátergickým systémom. Predpokladá sa, žepráve narušený glutamátergický prenos prostredníctvom subpopuláciehypofunkčných NMDA-receptorov v prefrontálnej kôre sa môže zúčast-ňovať na vzniku funkčných abnormalít dopamínergického systému po-zorovaných u pacientov so schizofréniou (obr. 6.3) (Goff a Coyle, 2001;Laruelle a spol., 2003).Ďalším monoamínom, pri ktorom sa predpokladá účasť v etiopa-
togenéze schizofrénie, je sérotonín. Sérotonínová hypotéza schizofrénievychádza z pozorovaní efektu halucinogénov (LSD, meskalín), ktoréuplatňujú svoj centrálny efekt prostredníctvom 5-HT2-receptorov. Ďal-ším podporným argumentom tejto hypotézy je fakt, že väčšina nových

6 Úloha neurotransmiterov pri ochoreniach nervového systému 189
antipsychotík má výraznú afinitu k sérotonínovým receptorom. Okremtoho má sérotonín regulačný inhibičný vplyv na dopamínergické neu-róny. Pozornosť sa venuje aj štúdiu interakcií medzi sérotonínergickýma glutamátergickým systémom pri schizofrénii (Aghajanian a Marek,2000).Na etiopatogenéze schizofrénie má pravdepodobne podiel aj cholí-
nergický systém, pretože ovplyvňuje funkcie CNS, ktoré sú pri tomtoochorení narušené. Účasť cholínergického systému v etiopatogenéze schi-zofrénie sa predpokladá aj na základe pôsobenia antipsychotika kloza-pínu, ktorého jeden z dokázaných mechanizmov účinku je sprostredko-vaný práve cholínergickými muskarínovými receptormi (Tandon, 1999).Štúdie mozgového tkaniva získaného pri autopsii pacientov s diagnos-tikovanou schizofréniou preukázali znížené množstvo nikotínových re-ceptorov v niektorých oblastiach mozgu. Niektoré štúdie naznačujú, žeinhalácia nikotínu zvyšuje množstvo nikotínových receptorov (Breesea spol., 2000). Vzťah medzi zvýšeným výskytom fajčenia u pacientovso schizofréniou tak môže mať nielen sociálne a kulturálne pozadie,ale môže byť aj biologicky podmienený (de Leon, 1999). Ďalšie štú-die poukazujú na možnú účasť kanabinoidových receptorov v patofy-ziológii schizofrénie. Predpokladá sa, že fajčenie marihuany môže zvý-šiť riziko prepuknutia ochorenia u predisponovaných osôb (Hollister,1998; Henquet a spol., 2005). V etiopatogenéze schizofrénie sa skúmaaj možná účasť GABA a neuropeptidov (napr. β-endorfínu, cholecysto-kinínu, neurotenzínu, somatostatínu, neuropeptidu Y) (De Wied a Sigli,2002; Höschl a spol., 2004; Kinkead a Nemeroff, 2004).Dopamínová teória schizofrénie je základným teoretickým predpo-
kladom pre farmakologickú liečbu schizofrénie, no na koncepčnej úrovnisa v súčasnosti prechádza od dopamínovej teórie schizofrénie k multi-faktoriálnemu pohľadu. Uvažuje sa o komplexných interakciách medzimonoamínergickým, glutamátergickým, GABAergickým a inými neuro-transmiterovými systémami. Predpokladá sa, že primárne narušenie sanemusí týkať žiadneho z uvedených neurotransmiterových systémov, aleže môže ísť o oveľa globálnejšiu poruchu. Uvažuje sa o možnom naru-šení migrácie neurónov a tvorby spojení medzi neurónmi počas ontoge-nézy nervového systému. Pozorované neurotransmiterové abnormalityby potom mohli byť len sekundárnym následkom vývinových porúchCNS (Carlsson a spol., 2001; Sawa a Snyder, 2002; Freedman, 2003).

190 NEUROTRANSMITERY
Afektívne poruchyB.Kukanová, J.Pečeňák, B.Mravec
Poruchy nálady (afektívne poruchy) sú závažné, často chronickéa niekedy život ohrozujúce ochorenia. Zahŕňajú skupinu porúch, ktorésa vyznačujú patologickou náladou a periodickým priebehom. Súčasnépoznatky ešte nevytvárajú dostatočné podklady pre rozdelenie afektív-nych porúch podľa mechanizmu ich vzniku. Klasifikácia afektívnych po-rúch sa neustále vyvíja a opiera sa o klinickú symptomatológiu a priebe-hové charakteristiky. Dve hlavné skupiny afektívnych porúch tvoria uni-polárna depresia (jednotlivá fáza alebo rekurentná depresia) a bipolárnaafektívna porucha, pri ktorej sa vyskytujú aj manické, či hypomanickéfázy (MKCH 10, pozri literatúra). Narastá množstvo poznatkov, na zá-klade ktorých sa uvažuje, že v etiopatogenéze afektívnych porúch majúúlohu genetické faktory, rôznorodé stresory, narušenie biologických ryt-mov, imunoendokrinné alterácie, poruchy neurotransmiterových systé-mov (neurotransmitery, receptory, postreceptorové, transportné, mem-bránové mechanizmy a iné) (Siegel a spol., 2006).
Depresia
Depresia zahŕňa viacero porúch, ktorých zjednocujúcim príznakom jeprítomnosť tzv. depresívneho syndrómu. Depresívny syndróm možnocharakterizovať ako stav s patologickou depresívnou náladou sprevádza-nou somatickými a vegetatívnymi príznakmi a zmenou psychomotoric-kého tempa (väčšinou jeho spomalenie). V rámci somatických príznakovdepresívneho syndrómu sa vyskytujú aj poruchy spánku (hyposomnia ajhypersomnia), poruchy príjmu potravy (najmä nechutenstvo) a stratalibida. Prítomnosť ranného pesima (depresia je najvýraznejšia ráno)a skorého prebúdzania sú charakteristické pre tzv. endogénnu formudepresie, táto forma však nie je v súčasných klasifikačných systémochjednoznačne vymedzená. Popri depresívnej nálade je typickým preja-vom depresívnej poruchy strata záujmov a potešenia z obvyklých čin-ností. Prítomné sebaobviňovanie môže nadobudnúť až charakter bludua môže sa vyskytovať popri iných psychotických príznakoch (haluci-nácie a ďalšie typy bludov). Pri závažnejšej depresii je vysoké rizikosamovražedného konania (Češková, 2001; Höschl a spol., 2004).

6 Úloha neurotransmiterov pri ochoreniach nervového systému 191
Depresívna porucha sa môže vyskytnúť ako reakcia na závažnú ži-votnú udalosť alebo ako endogénna porucha, pri ktorej sa predpokladázmena v činnosti neuronálnych systémov a iné endogénne mechanizmyvzniku. Rozlišuje sa aj tzv. symptomatická depresia, ktorá je sprie-vodným znakom niektorých telesných ochorení (napr. pri nádorovomochorení), depresia pri organickom postihnutí mozgu (napr. po trau-matickom alebo zápalovom poškodení) a farmakogénna depresia (podlhodobom užívaní neuroleptík, hormonálnej antikoncepcie). Depresiasa môže vyskytovať aj ako súčasť klinického obrazu iných psychiatric-kých porúch (napr. pri mentálnej anorexii, alkoholizme, pri porucháchosobnosti a pod.) (Höschl a spol., 2004).Aj napriek intenzívnemu výskumu v priebehu posledných desaťročí
zostávajú príčiny a mechanizmus vzniku depresie dosiaľ neobjasnené.Uvažuje sa, že na etiopatogenéze depresie sa zúčastňuje veľké množ-stvo faktorov (Hasler a spol., 2004; Nemeroff a Vale, 2005). Pozornosťsa venuje prevažne štúdiu genetických faktorov, abnormalít neurotrans-miterových systémov, endokrinného a imunitného systému, ako aj mož-ného vplyvu zmenenej neurogenézy a neuropatologickým abnormalitám(Licinio a Wong, 1999; Harrison, 2002; Taylor a spol., 2005).Klinické sledovania vedľajších účinkov látok rezerpínu a iproniazidu
zo začiatku 50. rokov 20. storočia naznačili možnú súvislosť medzi de-presiou a monoamínergickými neurónmi v CNS. Rezerpín, ktorý sa po-užíval na liečbu hypertenzie, ale aj ako neuroleptikum, vyvolával u nie-ktorých pacientov výraznú depresiu. Experimentálne štúdie na labora-tórnych zvieratách preukázali, že po podaní rezerpínu u nich dochádzak inaktivite a sedácii, teda k príznakom poukazujúcim na určitú ana-lógiu s depresívnymi symptómami u ľudí. Predpokladalo sa, že rezer-pín vyvoláva nežiaduce vedľajšie príznaky depléciou zásob monoamínovv nervových zakončeniach a následným obmedzením prenosu signálovmonoamínergickými neurónmi v mozgu. Iproniazid, ktorý sa používalna liečbu tuberkulózy, naopak vyvolával u pacientov eufóriu. Následnesa dokázal jeho terapeutický efekt pri liečbe depresie. Experimentálneštúdie zistili, že iproniazid ireverzibilne inhibuje činnosť enzýmu MAO.Inhibícia MAO iproniazidom má za následok zníženú degradáciu mo-noamínov, čo vedie k zvýšeniu množstva monoamínov v sekrečných ve-zikulách, z ktorých môžu byť vo väčšom množstve uvoľnené do synap-tickej štrbiny. Roku 1958 bol objavený imipramín látka, ktorá preuka-zovala účinnosť pri liečbe depresie. Imipramín blokuje spätné vychy-

192 NEUROTRANSMITERY
távanie monoamínov, predovšetkým noradrenalínu, zo synaptickej štr-biny. To vedie k predĺženému a zvýšenému pôsobenie uvoľneného nor-adrenalínu, čo sa považovalo za mechanizmus antidepresívneho účinkuimipramínu. Na základe týchto pozorovaní sa formovala monoamínováhypotéza depresie. Táto hypotéza predpokladala, že na vzniku depresiesa zúčastňuje funkčný deficit monoamínergickej transmisie (sérotonínu,noradrenalínu a dopamínu) v určitých mozgových štruktúrach (Kafkaa spol., 1998; Ressler a Nemeroff, 2000; Malhi a spol., 2005). Uvažujesa preto, že medzi štruktúry, ktorých dysfunkcia sa môže zúčastňovaťna etiopatogenéze depresie, patria oblasti s najväčšou akumuláciou mo-noamínergických neurónov v mozgu (locus coeruleus a nuclei raphe).Noradrenalínergické neuróny v locus coeruleus, ako aj sérotonínergickéneuróny v nuclei raphe modulujú rozsiahle oblasti nervového systémua zároveň môžu vzájomne ovplyvňovať svoju aktivitu (Ressler a Neme-roff, 1999, 2000). Takéto usporiadanie umožňuje monoamínovým systé-mom modulovať veľké množstvo funkcií, pričom narušenie niektorýchz nich sa pozoruje práve pri depresii.Hlavným nedostatkom monoamínovej teórie je však fakt, že aj keď
látky zo skupiny antidepresív ovplyvňujú monoamínergickú transmisiurelatívne rýchlo, nástup ich antidepresívnych účinkov sa prejavuje poniekoľkých dňoch až týždňoch (Garattini a Samanin, 1988; Hindmarch,2001). Predpokladá sa preto, že antidepresívne pôsobiace látky nemu-sia uplatňovať svoj klinický efekt iba priamym ovplyvňovaním koncen-trácie monoamínov v synaptickej oblasti, ale že ich efekt je výsled-kom komplexných zmien v monoamínergickom prenose v mozgu a neu-roplastických procesoch (obr. 6.4). Podanie antidepresív môže akútnezvýšiť množstvo monoamínov v synaptickej štrbine buď inhibíciou ichspätného vychytávania, inhibíciou ich biodegradácie v neurónoch, alebozvýšením ich uvoľňovania v dôsledku blokády α2-autoreceptorov a he-teroreceptorov na monoamínergických nervových zakončeniach. Pred-pokladá sa však, že takéto relatívne rýchle zvýšenie koncentrácie mo-noamínov v synapsách môže viesť k dlhodobým adaptačným zmenám,ktorých výsledkom je desenzitizácia inhibičných autoreceptorov a he-teroreceptorov, medzi ktoré patria receptory α2 a 5-HT1B, ako aj so-matodendriticky lokalizované 5-HT1A-receptory nachádzajúce sa v rôz-nych mozgových oblastiach. Desenzitizácia týchto inhibičných recepto-rov môže viesť k zvýšenej aktivite centrálnych monoamínergických sys-témov, čo sa dáva do súvislosti s terapeutickým účinkom antidepresív

6 Úloha neurotransmiterov pri ochoreniach nervového systému 193
(Elhwuegi, 2004). Predpokladá sa, že za terapeutický efekt antidepresívmôže zodpovedať aj zlepšenie plasticity a odolnosti neurónov na základezmien génovej expresie, ktorá vedie k tvorbe nových proteínov.Uvažuje sa tiež o tom, že v etiopatogenéze depresie môžu mať určitú
úlohu aj iné neurotransmiterové systémy (napr. acetylcholín, GABA,glutamát, tachykiníny, neuropeptid Y). Vo fázach klinického skúšaniasú látky ovplyvňujúce neurokinínové receptory NK1 a NK2 a agomela-tín, látka, ktorá agonisticky pôsobí na melatonínových receptoroch, sadostala do fázy schvaľovacieho procesu pre klinické použitie.Na účasť cholínergického systému v etiopatogenéze depresie pouka-
zuje výskyt depresívnych zmien nálady po podaní cholinomimetík pa-cientom s afektívnymi poruchami alebo rodinnou anamnézou depresie.Predpokladá sa, že zvýšená cholínergická aktivita by sa mohla zúčast-ňovať na skrátení REM fázy spánku u depresívnych pacientov. Aj keďviacero antidepresív blokuje muskarínové receptory, novšie antidepre-síva túto vlastnosť nemajú. Ani biochemické nálezy týkajúce sa účasticholínergickej transmisie v etiopatogenéze depresie nie sú konzistentné.Aj keď sa vyskytujú symptómy úzkosti u depresívnych pacientov
často, nie sú k dispozícii ucelené poznatky, ktoré by poukazovali navzťah medzi GABAergickým systémom a depresiou.O možnej účasti glutamátergického systému v etiopatogenéze depre-
sie je iba obmedzené množstvo klinických údajov. Údaje však poukazujúna to, že viacero antidepresív vyvoláva adaptačné zmeny vo väzbe li-gandov na glutamátergické NMDA-receptory, ako aj zmenu v množstvemRNA pre NMDA-receptory. Vyšetrenie magnetickou spektroskopioupreukázalo zmenené množstvo glutamátu v mozgu pacientov s depre-siou. Keďže bunky glie majú významnú úlohu pri odstraňovaní gluta-mátu zo synaptickej štrbiny, abnormality týchto buniek sa môžu zú-častňovať na etiopatogenéze depresie prostredníctvom porušenej čin-nosti glutamátergického systému (Kugaya a Sanacora, 2005). Pozoro-valo sa, že podanie kompetitívnych a funkčných antagonistov NMDA-receptorov vyvoláva antidepresívny efekt v animálnych experimentoch.Na možný význam tachykinínov v etiológii depresie poukazujú via-
ceré klinické pozorovania. Zistilo sa, že podanie antagonistov substan-cie P pri migrenóznej bolesti ovplyvňuje u pacientov aj náladu. Ďal-šie štúdie, v ktorých sa sledovali koncentrácie substancie P v plazmea cerebrospinálnej tekutine, ako aj efekt antagonistov receptorov pretachykiníny, nepriniesli jednoznačné výsledky. Tachykiníny sa nachá-

194 NEUROTRANSMITERY
Obr. 6.4 Schematické znázornenie noradrenalínergického a sérotonínergic-
kého synaptického prenosu, ich vzájomných interakcií a mechanizmov účinku
antidepresív. Miesta účinku antidepresív: A – inhibícia spätného vychytáva-nia monoamínov; B– inhibícia biodegradácie monoamínov enzýmom MAO-A;
C– blokáda presynaptických inhibičných autoreceptorov; D – stimulácia post-synaptických receptorov; E – aktivácia prenosu signálov a kaskády druhých
poslov. MAO-A–monoaminooxidáza A; VMAT–vezikulárny transportér pre
monoamíny; NET– transportér umožňujúci spätné vychytávanie noradrena-línu; 5-HT– transportér umožňujúci spätné vychytávanie sérotonínu; 5-HIAA–
kyselina 5-hydroxyindoloctová; DHPG– 3,4-dihydroxyfenylglykol (modifikované
podľa Leonard, 2003; Mann, 2005).

6 Úloha neurotransmiterov pri ochoreniach nervového systému 195
dzajú v amygdale, o ktorej sa predpokladá, že je kritickou štruktúrouregulujúcou emócie. Nachádzajú sa aj v oblastiach s intenzívnou mo-noamínergickou inerváciou, čo môže vytvárať podklad pre modulačnývplyv tachykinínov na monoamínergickú transmisiu (Stahl, 2000; Herp-fer a Lieb, 2005; McLean, 2005).Predpokladá sa, že v etiopatogenéze depresie môže mať úlohu aj
neuropeptid Y. Jeho význam v etiopatogenéze ochorenia spočíva prav-depodobne v modulácii monoamínergického prenosu, ktorú neuropep-tid Y ako kotransmiter monoamínov navodzuje (Harro a Oreland, 2001).Už pred viac ako 40 rokmi sa pozorovalo, že štatisticky významný
počet pacientov s diagnózou depresie preukazuje zvýšené plazmatickéhladiny kortizolu a jeho metabolitov. Klinické pozorovania pouka-zujú na častý výskyt depresie u pacientov s Cushingovou chorobou.Tieto pozorovania spoločne s poznatkami o zvýšenej syntéze a uvoľ-ňovaní glukokortikoidov počas stresových situácií viedli k štúdiu úlohyHPA-systému v patofyziológii depresie (Arborelius a spol., 1999; Gil-lespie a Nemeroff, 2005). U časti depresívnych pacientov sa zistili od-lišnosti denného rytmu ACTH a kortizolu, zvýšené hladiny kortizoluv moči a plazme, zvýšené koncentrácie CRH v mozgovo-miechovej te-kutine a zväčšený objem nadobličiek (Carroll a spol., 1976; Nemeroffa spol., 1984, 1992; Halbreich a spol., 1985; Deuschle a spol., 1997).Predpokladá sa, že u pacientov s depresiou sa môže zvyšovať aktivitaHPA-systému, na ktorej sa môže zúčastňovať narušenie negatívnej spät-nej väzby. U niektorých pacientov s depresiou sa dokázala nedostatočnásupresia vyplavovania CRH po podaní dexametazónu, a tým nedosta-točné následné zníženie hladiny kortizolu. Klinické využitie tohto ná-dejného laboratórneho funkčného testu (DST – dexametazónový supre-sívny test) pre diagnostiku depresie sa však neuplatnilo pre jeho ne-dostatočnú senzitivitu a špecificitu. Uvažuje sa, že niektoré antidepre-síva uplatňujú svoj efekt aj prostredníctvom ovplyvnenia činnosti HPA-systému. Experimentálne výsledky poukazujú na to, že antidepresívamôžu znížiť aktivitu transportéra pre steroidy lokalizovaného v mem-bránach neurónov a buniek tvoriacich hematoencefalickú bariéru. Tomôže viesť k zvýšenej dostupnosti glukokortikoidov v CNS a následnek inhibícii činnosti HPA-systému (Pariante a spol., 2004). Na regulá-cii činnosti HPA-systému sa významne zúčastňujú aj monoamínergickéneuróny CNS. Uvažuje sa preto, že na dysfunkcii činnosti HPA-systémupozorovanej pri depresii sa môže zúčastňovať zmenená činnosť mono-

196 NEUROTRANSMITERY
amínergických systémov mozgu, resp. narušená interakcia medzi mono-amínergickými systémami a CRH-neurónmi nucleus paraventricularishypotalamu.Aj keď sa na regulácii emočných pochodov zúčastňuje veľké množ-
stvo mozgových oblastí, poznatky o neuronálnych okruhoch zapojenýchv regulácii nálady a ich dysfunkcií pri depresii sú iba čiastkové. To ostrokontrastuje s neuropatologickými nálezmi pri iných neuropsychiatric-kých ochoreniach (napr. Parkinsonova, Huntingtonova a Alzheimerovachoroba), pri ktorých sa identifikovali patologické lézie v špecifickýchoblastiach CNS. Predpokladá sa, že za symptomatológiu depresie zod-povedá narušená činnosť viacerých mozgových oblastí (obr. 6.5). Tietopredpoklady podporujú štúdie u pacientov využívajúce zobrazovacietechniky, ktoré ukázali zmeny v prietoku krvi (ako markera aktivitynervového tkaniva) vo viacerých oblastiach mozgu, medzi ktoré pat-ria prefrontálny a cingulárny kortex, hipokampus, striatum, amygdalaa talamus. Anatomické štúdie mozgov pacientov s depresiou ukázali ab-normality v mnohých z uvedených mozgových oblastí (Nestler a spol.,2002). Okrem toho sa v štúdiu etiopatogenézy depresie venuje pozor-nosť aj génu pre sérotonínový transportér. Polymorfizmus uvedenéhogénu môže mať význam pri rozvoji depresie aj v odpovedi na liečbuantidepresívami (Smits a spol., 2004; Kendler a spol., 2005).Súčasné hypotézy pokúšajúce sa vysvetliť etiopatogenézu depresie
predpokladajú, že ide o ochorenie, ktoré nie je manifestáciou jednodu-chého zvýšenia alebo zníženia koncentrácie určitých neurotransmiterovv mozgu. Poruchy, ktoré zapríčiňujú toto ochorenie, majú pravdepo-dobne komplexný charakter. Okrem narušenej činnosti neurotransmi-terových systémov sa predpokladá aj porucha v neuronálnej plasticitea v procesoch neurogenézy v dospelom mozgu (Siever a spol., 1985).
Psychiatrické ochorenia a neurogenézaB.Kukanová, B.Mravec
Väčšina tkanív v ľudskom organizme obsahuje multipotentné kme-ňové bunky, ktoré sa môžu zúčastňovať na obnovovaní tkanivovýchštruktúr počas života jedinca. Donedávna sa mozgové tkanivo v tomtosmere považovalo za výnimku. Predpokladalo sa, že neurogenéza pre-bieha len v prenatálnom a skorom postnatálnom vývoji. Novšie práce

6 Úloha neurotransmiterov pri ochoreniach nervového systému 197
Obr. 6.5 Štruktúry a dráhy CNS, o ktorých sa predpokladá, že ich dysfunkciasa môže zúčastňovať na etiopatogenéze depresie. Locus coeruleus (LC) a nuc-lei raphe (NR) modulujú činnosť limbického systému, ktorý ovplyvňuje afek-
tívne funkcie. Stres a iné aktivačné podnety vedú k aktivácii LC aj prostredníc-
tvom CRH syntetizovaného neurónmi v nucleus paraventricularis hypotalamua v amygdale. Činnosť CRH-neurónov, ktoré inervujú LC, je inhibovaná vply-
vom sérotonínu uvoľneného zo zakončení axónov z neurónov NR. Neuróny NR sapravdepodobne zúčastňujú na vzniku tolerancie na averzívne situácie a znižujú
stresovú odpoveď amygdaly a nucleus striae terminalis (BNST). Ďalšie limbické
oblasti vrátane hipokampu sú modulované monoamínergickými neurónmi, pri-čom noradrenalín sa zúčastňuje na adekvátnom vytvorení pamäťovej stopy na
averzívne podnety, sérotonín pôsobí opačne. Dorzálny aj ventrálny/orbitálny
prefrontálny kortex (PFC) sú významne zapojené do pamäťových procesov sú-visiacich s úzkostnými situáciami a toleranciou averzívnych podnetov. Aktivácia
HPA-systému vedie k zvýšeniu plazmatických hladín kortizolu. Kortizol na jed-nej strane inhibuje činnosť CRH-neurónov modulujúcich sekréciu ACTH z ade-
nohypofýzy, na druhej strane však zvyšuje aktivity CRH-neurónov inervujúcich
LC (modifikované podľa Ressler a Nemeroff, 2000; Gold a Chrousos, 2002).

198 NEUROTRANSMITERY
však naznačujú, že k tvorbe nových neurónov dochádza aj počas životadospelého jedinca (Mori a spol., 2005; Mendez-Otero a spol., 2005).Aj keď sa prvé štúdie dokumentujúce neurogenézu v dospelom moz-
gu cicavcov objavili už v 60. rokoch 20. storočia, iba v poslednom desať-ročí sa začala všeobecne akceptovať. Predpokladá sa, že tvorba novýchneurónov prebieha v subventrikulárnej zóne bočných komôr a v sub-granulárnej zóne gyrus dentatus. V subventrikulárnej zóne u dospelýchhlodavcov sa nachádzajú kmeňové bunky, ktorých neuroblastové po-tomstvo migruje cez rostrálny migratórny prúd za účelom vytvoreniafunkčných interneurónov v bulbus olfactorius. V gyrus dentatus sa novéneuróny tvoria v subgranulárnej zóne, kde vytvárajú granulárne bunkyparticipujúce na kognitívnych a pravdepodobne aj emočných procesoch.V súčasnosti sa predpokladá, že kmeňové bunky, z ktorých môžu vznik-núť nové neuróny v dospelom mozgu, môžu pochádzať z radiálnej glie(Lie a spol., 2004; Mirescu a Gould, 2004).Predpokladá sa, že poruchy v neurogenéze u dospelého jedinca môžu
mať úlohu v etiopatogenéze viacerých ochorení CNS (obr. 6.6) (Manjia spol., 2001; Reif a spol., 2006). Na animálnych modeloch depresie, akoaj v hipokampe depresívnych pacientov sa ukázal zvýšený výskyt dege-neratívnych procesov a zníženie neurogenézy (Malberg, 2004). Ďalšímdôležitým poznatkom je zistenie, že vystavenie organizmu stresu, ako ajvysoké hladiny glukokortikoidov môžu negatívne zasahovať do procesovneurogenézy a neuroplasticity a môžu viesť k atrofii v hipokampovýchštruktúrach (Lee a spol., 2002). Dokázalo sa, že dlhodobé podávanie an-tidepresív z rôznych farmakologických skupín zvyšuje hipokampovú ne-urogenézu u experimentálnych zvierat (Malberg a spol., 2000). Uvažujesa o tom, že alterácia neurogenézy pozorovaná pri psychiatrických ocho-reniach súvisí s narušením tvorby BDNF (Brain-Derived NeurotrophicFactor) – faktora podporujúceho rast nových neurónov v mozgu. U dep-resívnych pacientov sa preukázalo zníženie hladín BDNF. Stresové pod-nety zapríčiňujú aj zníženie génovej expresie BDNF v hipokampe u po-tkanov (Smith a spol., 1995; Karege a spol., 2002; Hashimoto a spol.,2004; Hayley a spol., 2005).Imunohistochemické štúdie zamerané na kvantifikáciu proliferácie
buniek mozgového kmeňa uskutočnené post mortem na mozgoch pa-cientov so schizofréniou naznačujú, že redukovaná neurogenéza sa môžezúčastňovať aj na etiopatogenéze tohto ochorenia (Reif a spol., 2006).

6 Úloha neurotransmiterov pri ochoreniach nervového systému 199
Obr. 6.6 Schematické znázornenie faktorov, ktoré ovplyvňujú neuroplasticitua odolnosť neurónov. Genetické a vývinové faktory, opakované epizódy afektív-
nych ochorení a cerebrovaskulárne ochorenia sa môžu zúčastňovať na narušeníodolnosti neurónov a môžu viesť k ich atrofii až zániku. Stres a depresia pravde-
podobne negatívne ovplyvňujú neuroplasticitu a bunkovú odolnosť prostredníc-
tvom viacerých mechanizmov, ktoré zahŕňajú zníženie množstva BDNF, zvýše-nie glutamátergickej transmisie prostredníctvom NMDA a iných glutamátergic-
kých receptorov a redukciou energetickej kapacity bunky. BDNF–mozgový ras-tový faktor; ROS – reaktívne formy kyslíka; GR– glukokortikoidové receptory;
CREB– cAMP odpovedajúci viažuci proteín; MAR– receptory pre monoamíny;
PKA–proteínkináza A (modifikované podľa Manji a spol., 2001; D’Sa a Duman,2002).

200 NEUROTRANSMITERY
Na regulácii neurogenézy sa zúčastňujú okrem rastových faktorov(BDNF, vascular endothelial growth factor) aj hormóny (estrogén, tes-tosterón, prolaktín, tyroxín), ako aj viaceré neurotransmitery (gluta-mát, sérotonín, noradrenalín, dopamín, acetylcholín, β-endorfín) a inélátky (napr. NO) (obr. 6.6) (Hagg, 2005). Glutamát má pravdepodobneobzvlášť dôležitú úlohu v novotvorbe neurónov z kmeňových buniek.Predpokladá sa, že liečivá so selektívnym účinkom na glutamátové re-ceptory môžu predstavovať nový trend v terapii neurodegeneratívnychochorení, ako napr. Parkinsonova, Alzheimerova a Huntingtonova cho-roba.Významnú úlohu v neurogenéze dospelého nervového systému má
pravdepodobne aj GABA. Predpokladá sa, že GABA môže sprostred-kúvať informácie o aktivite nervových okruhov v blízkosti novovznik-nutého neurónu, čím umožní jeho správne začlenenie do týchto okru-hov. Fyziologické a patologické podnety, ktoré interferujú s GABAer-gickou signalizáciou, môžu proces začlenenia novovzniknutých neurónovdo nervových okruhov vyzretého CNS významne ovplyvniť (Ge a spol.,2006).Zmeny, ktoré sa týkajú plasticity v nervovom systéme, možno roz-
členiť na tri základné úrovne: neurónovú, interneuronálnu a synaptickú.Na neurónovej úrovni dochádza neustále k zániku, ako aj novotvorbeneurónov. Na úrovni interneuronálnych spojení možno pozorovať stáleprebiehajúcu prestavbu spojení medzi neurónmi, keď jednotlivé synapsymôžu zanikať a iné vznikať. Na úrovni synáps dochádza k ich posilňo-vaniu alebo oslabovaniu. Regulácia procesov neurogenézy, neuroplas-ticity a posilňovania/oslabovania synaptických spojov u dospelého je-dinca je komplexná a je pravdepodobne podmienená veľkým počtom en-dogénnych aj exogénnych faktorov. Možno predpokladať, že narušenietýchto procesov v zmysle zmeny pomerov novotvorba/zánik neurónov,tvorba/zánik interneuronálnych spojení a posilňovanie/oslabovanie silysynaptických spojení sa zúčastňuje ako jeden z etiopatogenetických fak-torov na vzniku niektorých neuropsychiatrických ochorení.Pochopenie uvedených mechanizmov môže napomôcť objasneniu
etiopatogenézy niektorých ochorení nervového systému a môže byť pod-kladom pre nové trendy v terapii týchto chorôb.

6 Úloha neurotransmiterov pri ochoreniach nervového systému 201
Látková závislosťB.Kukanová, B.Mravec
Látkovú závislosť možno charakterizovať ako chronické relapsujúceochorenie prejavujúce sa nutkavým užívaním látky, neschopnosťou ob-medziť jej príjem a objavením sa abstinenčných príznakov pri prerušeníjej príjmu. Látková závislosť podmieňuje zmenu v činnosti neurotrans-miterových systémov v skupine nervových štruktúr, medzi ktoré patriaventrálna tegmentálna oblasť, nucleus accumbens, amygdala, talamusa prefrontálny kortex. Medzi neurotransmitery, o ktorých sa predpo-kladá, že sú zapojené v procesoch látkovej závislosti, patria napríkladdopamín, sérotonín, GABA, glutamát, opioidové peptidy a CRH (Kooba spol., 1998).Predpokladá sa, že v procesoch látkovej závislosti majú významnú
úlohu dopamínergické neuróny nachádzajúce sa vo ventrálnej tegmen-tálnej oblasti (označované ako VTA alebo A10 neuróny). Tieto neurónyvysielajú axóny aj do nucleus accumbens, kde uvoľňujú dopamín (súčasťmezolimbických dopamínergických dráh).Transportéry pre monoamíny, ako napríklad dopamínový transpor-
tér (DAT) a vezikulárny monoamínový transportér 2 (VMAT-2), sa vý-znamne zúčastňujú na regulácii množstva dopamínu v mozgu. Zmenyv transporte dopamínu môžu viesť aj k vzniku látkovej závislosti. Nie-ktoré látky vyvolávajú vznik závislosti práve prostredníctvom ovplyv-nenia činnosti dopamínových transportérov (DAT, VMAT-2). Na zá-klade mechanizmu ovplyvňovania činnosti dopamínových transporté-rov sa rozlišujú psychostimulanciá, ktoré blokujú vychytávanie dopa-mínu („blokátory vychytávaniaÿ), a tie, ktoré zvyšujú jeho uvoľňova-nie („uvoľňovačeÿ). Jednotlivé psychostimulanciá však môžu pôsobiťovplyvňovaním oboch uvedených mechanizmov. Medzi psychostimu-lanciá, ktoré blokujú spätné vychytávanie dopamínu, patrí napríkladkokaín a metylfenidát. Amfetamín a jemu podobné psychostimulan-ciá účinkujú hlavne prostredníctvom stimulácie uvoľňovania dopamínuz nervových zakončení (Riddle a spol., 2005).Nikotín sa viaže priamo na nikotínové receptory dopamínergických
neurónov a následne zvyšuje aktivitu mezolimbických neurónov. µ-re-ceptory sa nachádzajú vo vysokej hustote na presynaptických zakon-čeniach GABAergických interneurónov inervujúcich VTA. Stimuláciatýchto receptorov inhibuje uvoľňovanie GABA, čo vedie k dezinhibí-

202 NEUROTRANSMITERY
cii dopamínergických neurónov VTA. Pravdepodobne aj alkohol vyvo-láva niektoré svoje účinky prostredníctvom modulácie aktivity neuró-nov VTA (obr. 6.7) (Davis a spol., 2002).
Obr. 6.7 Kľúčové štruktúry nervového systému, o ktorých sa predpokladá, že sazúčastňujú na vzniku látkovej závislosti. Vyznačené sú miesta, na ktorých prav-
depodobne pôsobia jednotlivé látky vyvolávajúce závislosť. VTA– area ventralistegmenti, Tsai; DA–dopamínergické neuróny; GABA–GABAergické neuróny
(modifikované podľa Davis a spol., 2002).
Užívanie návykových látok vedie okrem krátkodobých efektov (zme-ny v aktivite neurónov, uvoľňovanie neurotransmiterov atď.) aj k dl-hodobým zmenám, ktoré sú zapríčinené modifikáciou génovej expresieneurónov a syntézou nových proteínov. Predpokladá sa, že takéto dl-hodobé zmeny môžu byť podkladom pre vznik dlhodobej patologickejasociačnej pamäte, ktorá môže podmieňovať nutkavé užívanie látok.Predpokladá sa, že tieto zmeny sú aj podkladom oneskoreného relapsu,ktorý je pre látkovú závislosť charakteristický (Davis a spol., 2002).
Závislosť od alkoholu
Etiopatogenéza závislosti od alkoholu je stále nedostatočne charakte-rizovaná. Mechanizmus účinku alkoholu je nešpecifický, pretože alko-hol môže ovplyvňovať veľké spektrum neurotransmiterových systémov.Viaceré experimentálne údaje poukazujú na to, že alkohol potencujeneurotransmisiu viazanú na GABAA-receptory a redukuje excitačnú

6 Úloha neurotransmiterov pri ochoreniach nervového systému 203
neurotransmisiu viazanú na NMDA-glutamátergické receptory. To môžespôsobiť zvýšenie inhibičných a útlm excitačných pochodov v nervovomsystéme, čo môže mať za následok inhibíciu činnosti CNS charakteris-tickú pre intoxikáciu alkoholom (Stahl, 2000). Tolerancia na alkoholmôže viesť k zvýšenej senzitivite NMDA-receptorov. Vynechanie užíva-nia alkoholu má potom za následok hyperexcitabilný stav, ktorý môževyvolať záchvaty, delírium a excitotoxicitou podmienený zánik neuró-nov (Tsai a Coyle, 1998). Predpokladá sa, že aj ovplyvnenie činnostidopamínergického, noradrenenalínergického, sérotonínergického a opi-oidového systému sa zúčastňuje na vzniku závislosti od alkoholu (Kafkaa spol., 1998; Addolorato a spol., 2005). Na úlohu opioidového systémupoukazuje pozorovanie zníženého výskytu relapsu pri závislosti od al-koholu po podaní naloxonu (blokátor opioidových receptorov). V tejtoindikácii sa ukázal byť účinný v humánnych štúdiách aj blokátor gluta-mátergickej neurotransmisie (Bodnar a spol., 2004).Predpokladá sa aj určitá asociácia medzi poruchou neuroendokrin-
nej regulácie a závislosťou od alkoholu. V tomto smere sa uvažuje hlavneo účasti HPA-systému. Zistila sa korelácia medzi nízkym CRH a ten-denciou k relapsu po akútnej detoxikácii. Predpokladá sa, že aj leptína β-endorfín, systémy nepriamo modulujúce HPA-systém, sa môžu zú-častňovať na etiopatogenéze závislosti od alkoholu (Kiefer a spol., 2004).Niektoré animálne štúdie poukazujú na možnú súvislosť medzi závis-
losťou od alkoholu a neuropeptidom galanínom. Uvažuje sa, že existujepozitívna spätná väzba medzi príjmom alkoholu a špecifickým hypota-lamickým systémom neurónov syntetizujúcich galanín, ktorá môže pris-pievať k motivácii konzumovať excesívne množstvo alkoholu, a tak pris-pieť k vzniku alkoholovej závislosti (Lewis, 2005). Medzi ďalšie neuro-peptidy, o ktorých sa predpokladá, že majú úlohu v etiopatogenéze al-koholovej závislosti, patria urokortín I a neuropeptid Y (Cowen a spol.,2004).
EpilepsiaB.Mravec
Epilepsia je chronické záchvatové ochorenie nervového systému,ktoré vzniká nadmernou synchrónnou a udržiavanou výbojovou akti-vitou skupiny neurónov. Tieto neuróny vytvárajú epileptické ohnisko.

204 NEUROTRANSMITERY
Predpokladá sa, že na vzniku tohto ohniska alterovanej neuronálnejaktivity sa zúčastňujú:
• zmeny vo vnútorných vlastnostiach a excitabilite neurónov oh-niska,
• redukcia fyziologickej inhibičnej kontroly, ktorú zabezpečujeGABA,
• zvýšenie excitačného spriahnutia medzi neurónmi.
Vo výskume etiopatogenézy epilepsie sa dlho študovala úloha GABAako hlavného inhibičného neurotransmitera. Predpokladalo sa, že defektGABAergického systému (pokles počtu GABAergických neurónov) zod-povedá za vznik záchvatov. Niektoré sledovania poukázali na špecifickýdefekt v excitabilite GABAergických neurónov. U pacientov s epilepsioupostihujúcou temporálny lalok mozgovej kôry sa zistil pokles počtu tzv.„chandelier cellsÿ, ktoré predstavujú najvýznamnejšie kôrové GABAer-gické inhibičné interneuróny. Predpokladá sa, že normálna variabilitav počte týchto neurónov môže vysvetliť predispozíciu niektorých osôbna vznik epilepsie v porovnaní so zvyškom populácie (DeFelipe, 1999).Novšie poznatky dokázali heterogenitu populácií neurónov syntetizujú-cich GABA, pričom tieto neuróny obsahujú rôzne kotransmitery (napr.neuropeptid Y, somatostatín). Úloha týchto neurotransmiterov v eti-opatogenéze epilepsie však zatiaľ nie je objasnená (Davis a spol., 2002).V súčasnosti sa zdá, že na etiopatogenéze epilepsie sa zúčastňujú ok-
rem dysfunkcie viacerých vlastností GABAergického systému aj iné ab-normality. Predpokladá sa, že na zmenách v inhibičných a excitačnýchmechanizmoch pozorovaných pri epilepsii sa zúčastňuje aj glutamát,acetylcholín, monoamíny a adenozín (Dalby a Mody, 2001; Webster,2001; Cossart a spol., 2005; Magloczky a Freund, 2005).
Ischemické poškodenie CNSB.Mravec
Nervové tkanivo je extrémne citlivé na nedostatok kyslíka. Len nie-koľko minút trvajúca ischémia môže viesť k ireverzibilnému poškodeniuCNS. Jednotlivé oblasti mozgu sú rozdielne citlivé na ischémiu. Najcit-livejšie sú neuróny mozgovej kôry, podkôrové oblasti sú relatívne rezis-tentnejšie (Lipton, 1999).

6 Úloha neurotransmiterov pri ochoreniach nervového systému 205
Obr. 6.8 Schéma procesov aktivovaných glutamátom počas ischémie neurónov.Deplécia energetických zdrojov vedie k zvýšenému uvoľneniu glutamátu, ako aj
k blokáde jeho vychytávania bunkami glie. Intenzívna a dlhodobá stimulácia
glutamátových receptorov (AMPAR, NMDAR, kainateR) vedie k akumuláciiiónov Na+ a Ca2+ v neuróne a aktivácii kaskády procesov, ktoré môžu viesť
k zániku neurónu (modifikované podľa Won a spol., 2002).
Ischémia CNS vyvoláva kaskádu procesov, ktoré môžu viesť k zá-niku neurónov. Medzi základné mechanizmy ischemického poškodenianeurónov patrí:
• narušenie činnosti iónových púmp, ktoré vedie k intraneuróno-vej akumulácii Ca2+, Na+ a Cl−, čo následne zapríčiňuje vznikintracelulárneho edému,

206 NEUROTRANSMITERY
• nadmerná tvorba reaktívnych foriem kyslíka a dusíka počas reper-fúzie, ktoré vedú k štruktúrnemu a funkčnému poškodeniu neu-rónov,
• excesívne vyplavenie glutamátu, ktoré podmieňuje prehĺbenie in-tracelulárnej akumulácie Ca2+ a Na+ (Brott a Bogousslavsky,2000; Sugawara a spol., 2004).
Významným faktorom, ktorý môže podmieňovať rozsah ischemic-kého, resp. ischemicko-reperfúzneho poškodenia CNS, je excitotoxicképôsobenie neurotransmitera glutamátu. Prerušenie krvného zásobeniamozgu zapríčiňuje nedostatočný prísun kyslíka a glukózy, substrátovnevyhnutných pre tvorbu energie. Narušenie energetického metabo-lizmu vedie k zvýšenému presynaptickému uvoľneniu glutamátu násled-kom depolarizácie bunkovej membrány a aktivácie napäťovo riadenýchCa2+-kanálov. Ischémia má za následok aj zníženie vychytávania glu-tamátu bunkami glie, čo vedie k ešte výraznejšej akumulácii glutamátuv synaptickej štrbine. Nadmerná a dlhodobá aktivácia ionotropnýchglutamátergických receptorov vedie k výraznej akumulácii Ca2+ a Na+
v postsynaptickom segmente neurónu (obr. 6.8). To zapríčiňuje nielenvznik bunkového edému (ktorý je ešte potencovaný porušenou činnosťouiónových púmp), ale aj aktiváciu enzýmov závislých od Ca2+ a následnéireverzibilné poškodenie neurónov (Won a spol., 2002).Uvažuje aj o zapojení NO v ischemickom poškodení CNS. Predpo-
kladá sa, že v dôsledku zvýšenia extraneuronálnej koncentrácie gluta-mátu počas ischémie následne stúpa aj koncentrácia NO v nervovomtkanive. Niektoré experimentálne práce poukazujú na to, že práve vy-soké koncentrácie NO sa môžu významne zúčastňovať na ischemickompoškodení neurónov (Snyder a spol., 1998).

Úloha neurotransmiterovpri iných ochoreniach
B. Mravec
Experimentálne údaje poukazujú na to, že narušená činnosť neuro-transmiterových systémov môže mať určitú úlohu aj v etiopatogenézeiných ako neurologických a neuropsychiatrických porúch. Medzi tietoďalšie ochorenia možno zaradiť napríklad hypertenziu a niektoré zápa-lové ochorenia.
Hypertenzia
Na modulácii tlaku krvi sa významnou mierou zúčastňuje rostrálna ven-trolaterálna oblasť predĺženej miechy (RVLM), ktorej neuróny vysielajúaxóny k sympatikovým spinálnym pregangliovým neurónom lokalizova-ným v intermediolaterálnych stĺpcoch torakolumbálnej miechy. Sympa-tikové pregangliové neuróny následne modulujú aktivitu sympatikovýchnervov a drene nadobličky (obr. 7.1).Činnosť neurónov RVLM je modulovaná hypotalamom, stredným
mozgom a inhibičnými oblasťami predĺženej miechy (Sapru, 2002).U hypertenzných zvierat sa pozorovali funkčné zmeny CNS prevažnev hypotalame a predĺženej mieche. Funkčná alterácia jednotlivých hypo-talamických jadier môže mať za následok zvýšenie alebo zníženie tlaku
207

208 NEUROTRANSMITERY
Obr. 7.1 Kľúčové štruktúry predĺženej miechy zúčastňujúce sa na reguláciikrvného tlaku. RVLM– rostrálna ventrolaterálna predĺžená miecha; CVLM–
kaudálna ventrolaterálna predĺžená miecha; NTS – nucleus tractus solitarii;
IML– intermediolaterálny stĺpec miechy (obsahuje sympatikové pregangliovéneuróny); SG – sympatikové ganglion; GN– ganglion nodosum (modifikované
podľa Dampney a spol., 2003).
krvi prostredníctvom modifikácie činnosti sympatikového systému. Jed-notlivé hypotalamické jadrá sú navzájom bohato poprepájané, pričomkomunikujú aj s množstvom iných oblastí CNS. Na jednej strane neu-róny hypotalamu inervujú oblasti stredného mozgu, predĺženej miechya sympatikové pregangliové neuróny. Na druhej strane hypotalamus pri-jíma informácie zo stredného mozgu a predĺženej miechy, z chemorecep-torov a tlakových receptorov, ako aj z iných viscerálnych receptorov.Takéto anatomické usporiadanie umožňuje hypotalamu precízne regu-lovať krvný tlak.Regulácia krvného tlaku sa zabezpečuje komplexným systémom,
ktorý zahŕňa okrem hypotalamu aj veľké množstvo ďalších nervových

7 Úloha neurotransmiterov pri iných ochoreniach 209
štruktúr. Preto je pravdepodobné, že pri vzniku hypertenzie dochádzak alterácii v činnosti viacerých neuronálnych okruhov. Predpokladá sa,že pri hypertenzii dochádza v hypotalame, ako aj iných oblastiach ner-vového systému k zmenám vo funkcii viacerých neurotransmiterovýchsystémov. Uvažuje sa o alterácii nielen klasických neurotransmiterov(katecholamíny, acetylcholín, sérotonín, histamín, GABA), plynnéhoneurotransmitera NO, ale aj neuropeptidov (angiotenzín II, nátriure-tický peptid, vazopresín, neuropeptid Y, opioidy, TRH, tachykiníny,CRH). Aj keď sa čiastočné zmeny v týchto neurotransmiterových systé-moch pozorujú v celom hypotalame, zdá sa, že najvýznamnejšie zmenysú v prednom hypotalame. Predpokladá sa, že neurotransmiterová dys-funkcia sa môže zúčastňovať na zvýšenej aktivite sympatikového sys-tému, čo môže viesť k zvýšeniu krvného tlaku (de Wardener, 2001).
Zápalové procesy
Nervový systém nielenže monitoruje, ale môže aj modulovať priebeh zá-palových procesov prebiehajúcich v organizme (Dantzer a spol., 2000).Informácie o prebiehajúcom zápalovom procese sa prenášajú do CNSdvoma cestami:
• solubilné mediátory zápalu (cytokíny, chemokíny) prostredníc-tvom cirkumventrikulárnych orgánov prenášajú informácie doCNS,
• nervovými dráhami, hlavne prostredníctvom nervus vagus (Stern-berg, 1997; Goehler a spol., 2000).
Aj modulácia zápalu prebieha dvoma cestami:
• aktivácia HPA-systému vedie k vyplaveniu ACTH, ktorý regu-luje syntézu a vyplavovanie glukokortikoidov z kôry nadobličiek;glukokortikoidy modulujú priebeh zápalového procesu;
• nervovými dráhami prebiehajúcimi hlavne v nervus vagus(Berczi, 1997; Bernik, 2002).
Ukázalo sa, že aktivácia protizápalových dráh v nervus vagus zvy-šuje vyplavovanie acetylcholínu. Acetylcholín uvoľnený v tkanivách pro-stredníctvom α7-podjednotky nikotínových receptorov inhibuje činnosť

210 NEUROTRANSMITERY
imunitných buniek, čo je podstatou protizápalového pôsobenia nervusvagus (Tracey, 2002; Pavlov a spol., 2003). Viaceré experimentálneúdaje naznačujú, že na aktivácii protizápalového pôsobenia nervus va-gus sa zúčastňujú melanokortíny, ktoré sú syntetizované neurónmi CNSa viažu sa na melanokortínové receptory v mozgu (Guarini a spol.,2003).

Záver
Neurotransmitery zabezpečujú v nervovom systéme rôznorodé funkcieod prenosu signálov, kovalentných modifikácií proteínov, zmien v géno-vej expresii neurónov až po reguláciu počtu iónových kanálov, denzityreceptorov pre neurotransmitery, moduláciu množstva a charakteristíksynáps a podobne. Neurotransmitery preto majú v nervovom systémevýznamné homeostatické funkcie. Veľmi významné je toto pôsobeniepočas ontogenézy nervového systému. Narušenie komplexných neuro-transmiterových systémov sa preto môže zúčastňovať na vzniku ocho-rení nervového systému, ktoré majú neraz veľmi závažný vplyv na životpacienta.Poznanie funkcie neurotransmiterových systémov je podkladom pre
lepšie pochopenie funkcie nervového systému, ako aj pre ďalšie poodha-lenie mechanizmov zúčastňujúcich sa na etiopatogenéze psychiatrickýchochorení. Umožňuje aspoň čiastočne pochopiť aj mechanizmy farmako-terapie týchto ochorení.Aj keď poznáme veľké množstvo neurotransmiterov, je viac ako
pravdepodobné, že ich počet v budúcnosti narastie. Možno predpokla-dať, že prevažne pôjde o neurotransmitery zo skupiny neuropeptidov.Očakáva sa, že novoidentifikované neurotransmiterové systémy môžubyť cieľom farmakologickej intervencie zameranej na liečbu širokéhospektra neuropsychiatrických ochorení.Pri poznávaní významu jednotlivých neurotransmiterov, ako aj ich
vzájomných interakcií sa síce výrazne pokročilo, ale naše vedomosti tvo-ria stále iba fragmenty, ktorých zloženie do uceleného obrazu si vyžiadaešte veľké množstvo experimentálnej práce a klinického výskumu.
211

Literatúra
Abi-Dargham A, Rodenhiser J, Printz D, Zea-Ponce Y, Gil R, KegelesLS, Weiss R, Cooper TB, Mann JJ, Van Heertum RL, Gorman JM,Laruelle M. Increased baseline occupancy of D2 receptors by dopa-mine in schizophrenia. Proc Natl Acad Sci USA 2000; 97: 8104-8109.
Abi-Dargham A. Do we still believe in the dopamine hypothesis? Newdata bring new evidence. Int J Neuropsychopharmacol 2004; 7: S1-S5.
Acuna-Goycolea C, van den Pol A. Glucagon-like peptide 1 excites hy-pocretin/orexin neurons by direct and indirect mechanisms: implica-tions for viscera-mediated arousal. J Neurosci 2004; 24: 8141-8152.
Addolorato G, Leggio L, Abenavoli L, Gasbarrini G. Alcoholism Treat-ment Study Group. Neurobiochemical and clinical aspects of cravingin alcohol addiction: a review. Addict Behav 2005; 30: 1209-1224.
Adler MW, Geller EB, Chen X, Rogers TJ. Viewing Chemokines asa Third Major System of Communication in the Brain. AAPS 2006;7: E865-E870.
Adler MW, Rogers TJ. Are chemokines the third major system in thebrain? J Leukoc Biol 2005; 78: 1204-1209.
Aghajanian GK, Marek GJ. Serotonin model of schizophrenia: emergingrole of glutamate mechanisms. Brain Res Brain Res Rev 2000; 31:302-312.
Agnati LF, Zoli M, Stromberg I, Fuxe K. Intercellular communicationin the brain: wiring versus volume transmission. Neuroscience 1995;69: 711-726.
Ahlquist RP. A study of the adrenotropic receptors. Am J Physiol 1948;153: 586-600.
Ahmed B, Kastin AJ, Banks WA, Zadina JE. CNS effects of peptides:a cross-listing of peptides and their central actions published in thejournal Peptides, 1986-1993. Peptides 1994; 15: 1105-1155.
212

Literatúra 213
Akil M, Kolachana BS, Rothmond DA, Hyde TM, Weinberger DR, Kle-inman JE. Catechol-O-methyltransferase genotype and dopamine re-gulation in the human brain. J Neurosci 2003; 23: 2008-2013.
Alexander SP, Mathie A, Peters JA. Guide to receptors and channels,2nd edition. Br J Pharmacol 2006; 147: S1-168.
Allen AM, Moeller I, Jenkins TA, Zhuo J, Aldred GP, Chai SY, Men-delsohn FA. Angiotensin receptors in the nervous system. Brain ResBull 1998; 47: 17-28.
Allman J. Evolving brains. Scientific American Library, New York,2000, p. 1-225.
Ames RS, Sarau HM, Chambers JK, Willette RN, Aiyar NV, RomanicAM, Louden CS,Foley JJ, Sauermelch CF, Coatney RW, Ao Z, DisaJ, Holmes SD, Stadel JM, Martin JD, Liu WS, Glover GI, WilsonS, McNulty DE, Ellis CE, Elshourbagy NA, Shabon U, Trill JJ, HayDW, Ohlstein EH, Bergsma DJ, Douglas SA. Human urotensin-II isa potent vasoconstrictor and agonist for the orphan receptor GPR14.Nature 1999; 401: 282-286.
Andersson KE. Neurotransmitters: central and peripheral mechanisms.Int J Impot Res 2000; 12: S26-S33.
Andreasen NC, Nopoulos P, O’Leary DS, Miller DD, Wassink T, FlaumM. Defining the phenotype of schizophrenia: cognitive dysmetria andits neural mechanisms. Biol Psychiatry 1999; 46: 869-870.
Appel GB, Appel AS. Angiotensin II receptor antagonist: role in hyper-tension, cardiovascular disease, and renoprotection. Prog CardiovascDis 2004; 47: 105-115.
Aragon C, Lopez-Corcuera B. Structure, function and regulation of gly-cine neurotransporters. Eur J Pharmacol 2003; 479: 249-262.
Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses:glia, the unacknowledged partner. Trends Neurosci 1999; 22: 208-215.
Arborelius L, Owens MJ, Plotsky PM, Nemeroff CB. The role ofcorticotropin-releasing factor in depression and anxiety disorders.J Endocrinol 1999; 160: 1-12.
Armour JA. Myocardial ischemia and the cardiac nervous system. Car-diovasc Res 1999; 41: 41-54.

214 NEUROTRANSMITERY
Arnsten AF, Li BM. Neurobiology of executive functions: catecholamineinfluences on prefrontal cortical functions. Biol Psychiatry 2005; 57:1377-1384.
Arnsten AF. The biology of being frazzled. Science 1998; 280: 1711-1712.
Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance.Annu Rev Neurosci 2005; 28:403-450.
Axelrod J. Noradrenaline: fate and control of its biosynthesis. Science1971; 173: 598-606.
Azmitia EC. Serotonin neurons, neuroplasticity, and homeostasis of ne-ural tissue. Neuropsychopharmacology 1999; 21: 33S-45S.
Backberg M, Hervieu G, Wilson S, Meister B. Orexin receptor-1 (OX-R1) immunoreactivity in chemically identified neurons of the hypot-halamus: focus on orexin targets involved in control of food and waterintake. Eur J Neurosci 2002; 15: 315-328.
Baker JR, Cardinal K, Bober C, Taylor MM, Samson WK. Neuropep-tide W acts in brain to control prolactin, corticosterone, and growthhormone release. Endocrinology 2003; 144: 2816-2821.
Bale TL, Vale WW. CRF and CRF receptors: role in stress responsivityand other behaviors. Annu Rev Pharmacol Toxicol 2004; 44: 525-557.
Ballabh P, Braun A, Nedergaard M. The blood-brain barrier: an over-view: structure, regulation, and clinical implications. Neurobiol Dis2004; 16: 1-13.
Ballantyne D, Scheid P. Central chemosensitivity of respiration: a briefoverview. Respir Physiol 2001; 129: 5-12.
Ballesta J, Bloom SR, Polak JM. Distribution and localization of regu-latory peptides. Crit Rev Clin Lab Sci 1985; 22: 185-218.
Banks WA, Kastin AJ, Huang W, Jaspan JB, Maness LM. Leptin entersthe brain by a saturable system independent of insulin. Peptides1996; 17: 305-311.
Baranano DE, Ferris CD, Snyder SH. Atypical neural messengers.Trends Neurosci 2001; 24: 99-106.
Barnes NM, Sharp T. A review of central 5-HT receptors and theirfunction. Neuropharmacology 1999; 38: 1083-1152.

Literatúra 215
Bean AJ, Zhang X, Hokfelt T. Peptide secretion: what do we know?FASEB J 1994; 8: 630-638.
Beaujouan JC, Torrens Y, Saffroy M, Kemel ML, Glowinski J. A 25 yearadventure in the field of tachykinins. Peptides 2004; 25: 339-357.
Beinfeld MC. An introduction to neuronal cholecystokinin. Peptides2001; 22: 1197-1200.
Belelli D, Lambert JJ. Neurosteroids: endogenous regulators of theGABA(A) receptor. Nat Rev Neurosci 2005; 6: 565-575.
Beltowski J, Jamroz A. Adrenomedullin-what do we know 10 years sinceits discovery? Pol J Pharmacol 2004; 56: 5-27.
Ben-Ari Y. Excitatory actions of gaba during development: the natureof the nurture. Nat Rev Neurosci 2002; 3: 728-739.
Benešová O, Höschl C. Nobelova cena za fyziologii a medicínu 2000.Psychiatrické centrum Praha 2005, 145 pp.
Bennett MR, Balcar VJ. Forty years of amino acid transmission in thebrain. Neurochem Int 1999; 35: 269-280.
Bennett MR. Non-adrenergic non-cholinergic (NANC) transmission tosmooth muscle: 35 years on. Prog Neurobiol 1997; 52: 159-195.
Bennett MR. The concept of transmitter receptors: 100 years on. Ne-uropharmacology 2000; 39: 523-546.
Berczi I. Pituitary hormones and immune function. Acta Paediatr 1997;423: 70-75.
Bernadič M. Patofyziológia bolesti. In: Regulované zvýšenie telesnejteploty a patofyziológia bolesti. Bernadič M, Hulín I, Uhliar R, Štvr-tinová V, Ferenčík M (Eds), Faber: Bratislava, 2004; pp. 35-74.
Bernik TR, Friedman SG, Ochani M, DiRaimo R, Ulloa L, Yang H,Sudan S, Czura CJ, Ivanova SM, Tracey KJ. Pharmacological stimu-lation of the cholinergic antiinflammatory pathway. J Exp Med 2002;195: 781-788.
Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic sys-tem: modulation of behavioral state and state-dependent cognitiveprocesses. Brain Res Brain Res Rev 2003; 42: 33-84.
Berthoud HR. Multiple neural system controlling food intake and bodyweight. Neurosci Beh Rev 2002; 26: 393-428.

216 NEUROTRANSMITERY
Besson JM, Chaouch A. Peripheral and spinal mechanisms of nocicep-tion. Physiol Rev 1987; 67: 67-186.
Bettler B, Mulle C. Review: neurotransmitter receptors. II. AMPA andkainate receptors. Neuropharmacology 1995; 34: 123-139.
Beuckmann CT, Yanagisawa M. Orexins: from neuropeptides to energyhomeostasis and sleep/wake regulation. J Mol Med 2002; 80: 329-342.
Bhattacharjee AK, Nagashima T, Kondoh T, Tamaki N. The effects ofthe Na(+)/Ca(++) exchange blocker on osmotic blood-brain barrierdisruption. Brain Res 2001; 900: 157-162.
Biagioni S, Tata AM, De Jaco A, Augusti-Tocco G. Acetylcholine synt-hesis and neuron differentiation. Int J Dev Biol 2000; 44: 689-697.
Binaschi A, Bregola G, Simonato M. On the role of somatostatin inseizure control: clues from the hippocampus. Rev Neurosci 2003; 14:285-301.
Binder EB, Kinkead B, Owens MJ, Nemeroff CB. Neurotensin and do-pamine interactions. Pharmacol Rev 2001; 53: 453-486.
Bingham MJ, Cai J, Deehan MR. Eating, sleeping and rewarding: orexinreceptors and their antagonists. Curr Opin Drug Discov Dev 2006;9: 551-559.
Bittencourt JC, Presse F, Arias C, Peto C, Vaughan J, Nahon JL, ValeW, Sawchenko PE. The melanin-concentrating hormone system ofthe rat brain: an immuno- and hybridization histochemical charac-terization. J Comp Neurol 1992; 319: 218-245.
Blazquez E, Alvarez E, Navarro M, Roncero I, Rodriguez-Fonseca F,Chowen JA, Zueco JA. Glucagon-like peptide-1 (7-36) amide as a no-vel neuropeptide. Mol Neurobiol 1998; 18: 157-173.
Blundell JE, Lawton CL, Hill AJ. Mechanisms of appetite control andtheir abnormalities in obese patients. Horm Res 1993; 39: 72-76.
Bodnar RJ, Klein GE. Endogenous opiates and behavior: 2003. Peptides2004; 25: 2205-2256.
Boehning D, Snyder SH. Novel neural modulators. Annu Rev Neurosci2003; 26: 105-131.
Borowsky B, Durkin MM, Ogozalek K, Marzabadi MR, DeLeon J, LaguB, Heurich R, Lichtblau H, Shaposhnik Z, Daniewska I, BlackburnTP, Branchek TA, Gerald C, Vaysse PJ, Forray C. Antidepressant,

Literatúra 217
anxiolytic and anorectic effects of a melanin-concentrating hormone-1 receptor antagonist. Nat Med 2002; 8: 825-830.
Boulant JA. Role of the preoptic-anterior hypothalamus in thermore-gulation and fever. Clin Infect Dis 2000; 31: S157-S161.
Brailoiu GC, Dun SL, Yang J, Ohsawa M, Chang JK, Dun NJ. Apelin-immunoreactivity in the rat hypothalamus and pituitary. NeurosciLett 2002; 327: 193-197.
Breese CR, Lee MJ, Adams CE, Sullivan B, Logel J, Gillen KM, MarksMJ, Collins AC, Leonard S. Abnormal regulation of high affinity ni-cotinic receptors in subjects with schizophrenia. Neuropsychophar-macology 2000; 23: 351-364.
Broberger C, De Lecea L, Sutcliffe JG, Hokfelt T. Hypocretin/orexin-and melanin-concentrating hormone-expressing cells form distinctpopulations in the rodent lateral hypothalamus: relationship to theneuropeptide Y and agouti gene-related protein systems. J CompNeurol 1998; 402: 460-474.
Brott T, Bogousslavsky J. Treatment of acute ischemic stroke. N EnglJ Med 2000; 343: 710-722.
Brown RE, Stevens DR, Haas HL. The physiology of brain histamine.Prog Neurobiol 2001; 63: 637-672.
Buccafusco JJ. Neuronal nicotinic receptor subtypes: defining therape-utic targets. Mol Interv 2004; 4: 285-295.
Budai D. Neurotransmitters and receptors in the dorsal horn of thespinal cord. Acta Biol Szeged 2000; 44: 21-38.
Buller KM. Role of circumventricular organs in pro-inflammatorycytokine-induced activation of the hypothalamic-pituitary-adrenalaxis. Clin Exp Pharmacol Physiol 2001; 28: 581-589.
Bullock TH, Bennett MV, Johnston D, Josephson R, Marder E, FieldsRD. Neuroscience. The neuron doctrine, redux. Science 2005; 310:791-793.
Bunn HF, Poyton RO. Oxygen sensing and molecular adaptation tohypoxia. Physiol Rev 1996; 76: 839-885.
Burnstock G. Do some nerve cells release more than one transmitter?Neuroscience 1976; 1: 239-248.

218 NEUROTRANSMITERY
Cabrera-Vera TM, Vanhauwe J, Thomas TO, Medkova M, Preininger A,Mazzoni MR, Hamm HE. Insights into G protein structure, function,and regulation. Endocr Rev 2003; 24: 765-781.
Cano G, Card JP, Rinaman L, Sved AF. Connections of Barrington’snucleus to the sympathetic nervous system in rats. J Auton NervSyst 2000; 79: 117-128.
Cano G, Sved AF, Rinaman L, Rabin BS, Card JP. Characterization ofthe central nervous system innervation of the rat spleen using viraltransneuronal tracing. J Comp Neurol 2001; 439: 1-18.
Canteras NS, Simerly RB, Swanson LW. Organization of projectionsfrom the ventromedial nucleus of the hypothalamus: A Phaseolusvulgaris-leucoagglutinin study in the rat. J Comp Neurol 1994; 348:41-79.
Card JP, Fitzpatrick-McElligott S, Gozes I, Baldino FJr. Localizationof vasopressin-, vasoactive intestinal polypeptide-, peptide histidineisoleucine-and somatostatin-mRNA in rat suprachiasmatic nucleus.Cell Tissue Res 1988; 252: 307-315.
Carlezon WA Jr, Duman RS, Nestler EJ. The many faces of CREB.Trends Neurosci 2005; 28: 436-445.
Carlsson A, Waters N, Holm-Waters S, Tedroff J, Nilsson M, CarlssonML. Interactions between monoamines, glutamate, and GABA inschizophrenia: new evidence. Annu Rev Pharmacol Toxicol 2001; 41:237-260.
Carlsson A. A half-century of neurotransmitter research: impact on ne-urology and psychiatry. Nobel lecture Biosci Rep 2001a; 21: 691-710.
Carlsson A. A paradigm shift in brain research. Science 2001b; 294:1021-1024.
Carlsson A. Birth of neuropsychopharmacology - impact on brain rese-arch. Brain Res Bull 1999; 50: 363.
Carreras MC, Franco MC, Peralta JG, Poderoso JJ. Nitric oxide, com-plex I, and the modulation of mitochondrial reactive species in bio-logy and disease. Mol Aspects Med 2004; 25: 125-139.
Carroll BJ, Curtis GC, Davies BM, Mendels J, Sugerman AA. Urinaryfree cortisol excretion in depression. Psychol Med 1976; 6: 43-50.

Literatúra 219
Caulfield MP, Birdsall NJ. International Union of Pharmacology. XVII.Classification of muscarinic acetylcholine receptors. Pharmacol Rev1998; 50: 279-290.
Civantos Calzada B, Aleixandre de Artinano A. Alpha-adrenoceptorsubtypes. Pharmacol Res 2001; 44: 195-208.
Clowry GJ, Arnott GA, Clement-Jones M, Fallah Z, Gould S, WrightC. Changing pattern of expression of parvalbumin immunoreactivityduring human fetal spinal cord development. J Comp Neurol 2000;423: 727-735.
Clowry GJ, Fallah Z, Arnott G. Developmental expression of parval-bumin by rat lower cervical spinal cord neurones and the effect ofearly lesions to the motor cortex. Brain Res Dev Brain Res 1997;102: 197-208.
Colcher A. Management of Parkinson’s disease. Expert Rev Neurothe-rapeutics 2002; 2: 97-104.
Colmers WF, El Bahh B. Neuropeptide Y and Epilepsy. Epilepsy Curr2003; 3: 53-58.
Compagnone NA, Mellon SH. Neurosteroids: biosynthesis and functionof these novel neuromodulators. Front Neuroendocrinol 2000; 21: 1-56.
Connors BW, Long MA. Electrical synapses in the mammalian brain.Annu Rev Neurosci 2004; 27: 393-418.
Coote JH. A role for the paraventricular nucleus of the hypothalamusin the autonomic control of heart and kidney. Exp Physiol 2005; 90:169-173.
Cossart R, Bernard C, Ben-Ari Y.Multiple facets of GABAergic neuronsand synapses: multiple fates of GABA signalling in epilepsies. TrendsNeurosci 2005; 28: 108-115.
Costa A, Poma A, Navarra P, Forsling ML, Grossman A. Gaseous trans-mitters as new agents in neuroendocrine regulation. J Endocrinol1996; 149: 199-207.
Cotman CW, Monaghan DT. Anatomical organization of excitatoryamino acid receptors and their properties. Adv Exp Med Biol 1986;203: 237-252.

220 NEUROTRANSMITERY
Cottrell GT, Ferguson AV. Sensory circumventricular organs: centralroles in integrated autonomic regulation. Regul Pept 2004; 117: 11-23.
Coutinho V, Knopfel T. Metabotropic glutamate receptors: electricaland chemical signaling properties. Neuroscientist 2002; 8: 551-561.
Cowen MS, Chen F, Lawrence AJ. Neuropeptides: implications for al-coholism. J Neurochem 2004; 89: 273-285.
Crow TJ. Molecular pathology of schizophrenia. More than one diseaseprocess? Br Med J 1980; 289: 66-68.
Cuddy ML. The effects of drugs on thermoregulation. AACN Clin Issues2004; 15: 238-253.
Cummings DE, Weigle DS, Frayo RS, Breen PA, Ma MK, Dellinger EP,Purnell JQ. Plasma Ghrelin Levels after Diet-Induced Weight Lossor Gastric Bypass Surgery. Engl J Med 2002; 346: 1623-1630.
Cunningham MJ. Galanin-like peptide as a link between metabolismand reproduction. J Neuroendocrinol 2004; 16: 717-723.
Češková E. Afektivní poruchy. In: Psychiatrie. Raboch J, Zvolský P(Eds). Galén, Praha, 2001, pp. 253-274.
Dahlstrom A, Fuxe K. Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstrationof monoamines in the cell bodies of brain stem neurons. Acta PhysiolScand 1964; 232: 1-55.
Dahlstrom A, Fuxe K. The autonomic nervous system and the histo-chemical fluorescence method for the microscopical localization ofcatecholamines and serotonin. Brain Res Bull 1999; 50: 365-367.
Dalby NO, Mody I. The process of epileptogenesis: a pathophysiologicalapproach. Curr Opin Neurol 2001; 14: 187-192.
Damasio AR. Exploring the minded brain. The tanner lectures on hu-man values. University of Michigan 1997; 167-188.
Dampney RA, Coleman MJ, Fontes MA, Hirooka Y, Horiuchi J, Li YW,Polson JW, Potts PD, Tagawa T. Central mechanisms underlyingshort- and long-term regulation of the cardiovascular system. ClinExp Pharmacol Physiol 2002; 29: 261-268.
Dampney RA, Polson JW, Potts PD, Hirooka Y, Horiuchi J. Functionalorganization of brain pathways subserving the baroreceptor reflex:

Literatúra 221
studies in conscious animals using immediate early gene expression.Cell Mol Neurobiol 2003; 23: 597-616.
Danbolt NC. Glutamate uptake. Prog Neurobiol 2001; 65: 1-105.
Dantzer R, Konsman JP, Bluthe RM, Kelley KW. Neural and humoralpathways of communication from the immune system to the brain:parallel or convergent? Auton Neurosci 2000; 85: 60-65.
Dardonville C, Rozas I. Imidazoline binding sites and their ligands: anoverview of the different chemical structures. Med Res Rev 2004; 24:639-661.
Darlison MG, Greten FR, Harvey RJ, Kreienkamp HJ, Stuhmer T,Zwiers H, Lederis K, Richter D. Opioid receptors from a lower ver-tebrate (Catostomus commersoni): sequence, pharmacology, couplingto a G-protein-gated inward-rectifying potassium channel (GIRK1),and evolution. Proc Natl Acad Sci USA 1997; 94: 8214-8219.
Darlison MG, Richter D. Multiple genes for neuropeptides and theirreceptors: co-evolution and physiology. Trends Neurosci 1999; 22:81-88.
DaSilva J. Sex hormones and glucocorticoids: interactions with the im-mune system. Ann NY Acad Sci 1999; 816: 102-118.
Dautzenberg FM, Hauger RL. The CRF peptide family and their re-ceptors: yet more partners discovered. Trends Pharmacol Sci 2002;23: 71-77.
Davanger S. Colocalization of amino acid signal molecules in neuronsand endocrine cells. Anat Embryol (Berl) 1996; 194: 1-12.
Davenport AP, Bonner TI, Foord SM, Harmar AJ, Neubig RR, Pin JP,Spedding M, Kojima M, Kangawa K. International Union of Pharma-cology. LVI. Ghrelin receptor nomenclature, distribution, and func-tion. Pharmacol Rev 2005; 57: 541-546.
Davenport H W. Epinephrin(e). The Physiologist 1982; 25: 76-82.
Davis KL, Charney D, Coyle JT, Nemeroff C (Eds). Neuropsychophar-macology: The Fifth Generation of Progress. Lippincott Williams &Wilkins, Philadelphia, 2002; 2080 pp.
Dawson TM, Dawson VL. Molecular pathways of neurodegeneration inParkinson’s disease. Science 2003; 302: 819-822.

222 NEUROTRANSMITERY
Dayas CV, Buller KM, Day TA. Medullary neurones regulate hypot-halamic corticotropin-releasing factor cell responses to an emotionalstressor. Neuroscience 2001; 105: 707-719.
de Esch IJ, Thurmond RL, Jongejan A, Leurs R. The histamine H4receptor as a new therapeutic target for inflammation. Trends Phar-macol Sci 2005; 26: 462-469.
de Gasparo M, Catt KJ, Inagami T, Wright JW, Unger T. Internati-onal union of pharmacology. XXIII. The angiotensin II receptors.Pharmacol Rev 2000; 52: 415-472.
de Greef WJ, Rondeel JM, van Haasteren GA, Klootwijk W, Visser TJ.Regulation of hypothalamic TRH production and release in the rat.Acta Med Austriaca 1992; 19: 77-79.
de Leon J, Dadvand M, Canuso C, White AO, Stanilla JK, SimpsonGM. Schizophrenia and smoking: an epidemiological survey in a statehospital. Am J Psychiatry 1995; 152: 453-455.
de Wardener HE. The hypothalamus and hypertension. Physiol Rev2001; 81: 1599-1658.
De Wied D, Sigling HO. Neuropeptides involved in the pathophysiologyof schizophrenia and major depression. Neurotox Res 2002; 4: 453-468.
Dean B, Sundram S, Bradbury R, Scarr E, Copolov D. Studies on[3H]CP-55940 binding in the human central nervous system: regionalspecific changes in density of cannabinoid-1 receptors associated withschizophrenia and cannabis use. Neuroscience 2001; 103: 9-15.
Dean B. Understanding the pathology of schizophrenia: recent advancesfrom the study of the molecular architecture of postmortem CNStissue. Postgrad Med J 2002; 78: 142-148.
DeFelipe J. Chandelier cells and epilepsy. Brain 1999; 122: 1807-1822.
D’Este L, Casini A, Wimalawansa SJ, Renda TG. Immunohistochemicallocalization of amylin in rat brain stem. Peptides 2000; 21: 1743-1749.
D’Este L, Wimalawansa SJ, Renda TG. Distribution of amylin-immuno-reactive neurons in the monkey hypothalamus and their relationshipswith the histaminergic system. Arch Histol Cytol 2001; 64: 295-303.
Deuschle M, Schweiger U, Weber B, Gotthardt U, Korner A, Schmider J,Standhardt H, Lammers CH, Heuser I. Diurnal activity and pulsati-lity of the hypothalamus-pituitary-adrenal system in male depressed

Literatúra 223
patients and healthy controls. J Clin Endocrinol Metab 1997; 82:234-238.
Dietrich JB. Endothelial cells of the blood-brain barrier: a target forglucocorticoids and estrogens? Front Biosci 2004; 9: 684-693.
Dingledine R, Borges K, Bowie D, Traynelis SF. The glutamate receptorion channels. Pharmacol Rev 1999; 51: 7-61.
Dinulescu DM, Cone RD. Agouti and agouti-related protein: analogiesand contrasts. J Biol Chem 2000; 275: 6695-6698.
Dobner PR, Deutch AY, Fadel J. Neurotensin: dual roles in psychosti-mulant and antipsychotic drug responses. Life Sci 2003; 73: 801-811.
Dobner PR. Multitasking with neurotensin in the central nervous sys-tem. Cell Mol Life Sci 2005; 62: 1946-1963.
Dobolyi A, Irwin S, Makara G, Usdin TB, Palkovits M. Calcitonin gene-related peptide-containing pathways in the rat forebrain. J CompNeurol 2005; 489: 92-119.
Doust JA, Glasziou PP, Pietrzak E, Dobson AJ. A systematic reviewof the diagnostic accuracy of natriuretic peptides for heart failure.Arch Intern Med 2004; 164: 1978-1984.
D’Sa C, Duman RS. Antidepressants and neuroplasticity. Bipolar Di-sord 2002; 4: 183-194.
Dun NJ, Karczmar AG, Wu SY, Shen E. Putative transmitter systemsof mammalian sympathetic preganglionic neurons. Acta NeurobiolExp (Wars) 1993; 53: 53-63.
Dun NJ, Wu SY, Shen E, Miyazaki T, Dun SL, Ren C. Synaptic mecha-nisms in sympathetic preganglionic neurons. Can J Physiol Pharma-col 1992; 70: S86-S91.
Dun SL, Brailoiu GC, Mizuo K, Yang J, Chang JK, Dun NJ. Neuro-peptide B immunoreactivity in the central nervous system of the rat.Brain Res 2005; 1045: 157-163.
Duncan AJ, Heales SJ. Nitric oxide and neurological disorders. MolAspects Med 2005; 26: 67-96.
Dunčko R, Ježová D. Stresová odpoveď a serotonínergická transmisiavo vzťahu k patogenéze a farmakoterapii afektívnych porúch. Psy-chiatrie 2002; 6: 10-16.

224 NEUROTRANSMITERY
Dunn AJ, Swiergiel AH, Palamarchouk V. Brain circuits involvedin corticotropin-releasing factor-norepinephrine interactions duringstress. Ann NY Acad Sci 2004; 1018: 25-34.
Dunwiddie TV, Masino SA. The role and regulation of adenosine in thecentral nervous system. Annu Rev Neurosci 2001; 24: 31-55.
Ebert SN, Thompson RP. Embryonic epinephrine synthesis in the ratheart before innervation: association with pacemaking and conduc-tion tissue development. Circ Res 2001; 88: 117-124.
Edvinsson L, Uddman R. Neurobiology in primary headaches. BrainRes Brain Res Rev 2005; 48: 438-456.
Edwards RH. Drug delivery via the blood-brain barrier. Nat Neurosci2001; 4: 221-222.
Eglen RM, Hudson AL, Kendall DA, Nutt DJ, Morgan NG, WilsonVG, Dillon MP. ’Seeing through a glass darkly’: casting light onimidazoline ’I’ sites. Trends Pharmacol Sci 1998; 19: 381-390.
Elhwuegi AS. Central monoamines and their role in major depression.Prog Neuropsychopharmacol Biol Psychiatry 2004; 28: 435-451.
Elias CF, Saper CB, Maratos-Flier E, Tritos NA, Lee C, Kelly J, TatroJB, Hoffman GE, Ollmann MM, Barsh GS, Sakurai T, Yanagisawa M,Elmquist JK. Chemically defined projections linking the mediobasalhypothalamus and the lateral hypothalamic area. J Comp Neurol1998; 402: 442-459.
Elmquist JK, Ahima RS, Elias CF, Flier JS, Saper CB. Leptin activatesdistinct projections from the dorsomedial and ventromedial hypot-halamic nuclei. Proc Natl Acad Sci USA 1998; 95: 741-746.
Engelmann M, Ludwig M. The activity of the hypothalamo-neurohypophysial system in response to acute stressor exposure: ne-uroendocrine and electrophysiological observations. Stress 2004; 7:91-96.
Esch T, Stefano GB, Fricchione GL, Benson H. Stress in cardiovasculardiseases. Med Sci Monit 2002; 8: RA93-RA101.
Esplugues JV. NO as a signalling molecule in the nervous system. Br JPharmacol 2002; 135: 1079-1095.
Eto K, Asada T, Arima K, Makifuchi T, Kimura H. Brain hydrogensulfide is severely decreased in Alzheimer’s disease. Biochem BiophysRes Commun 2002; 293: 1485-1488.

Literatúra 225
Eto K, Kimura H. A novel enhancing mechanism for hydrogen sulfide-producing activity of cystathionine beta-synthase. J Biol Chem 2002;277: 42680-42685.
Falck B, Hillarp NA, Thieme G, Torp A. Fluorescence of catecholaminesand related compounds condensed with formaldehyde. J HistochemCytochem 1962; 10: 348-354.
Feldman RS, Quenzer LF. Fundamentals of Neuropsychopharmacology.Sinauer Associates, Sunderland 1984, pp. 1-508.
Ferguson AV, Bains JS. Electrophysiology of the circumventricular or-gans. Front Neuroendocrinol 1996; 17: 440-475.
Ferguson AV, Samson WK. The orexin/hypocretin system: a criticalregulator of neuroendocrine and autonomic function. Front Neuro-endocrinol 2003; 24: 141-150.
Ferguson AV, Washburn DL, Latchford KJ. Hormonal and neurotrans-mitter roles for angiotensin in the regulation of central autonomicfunction. Exp Biol Med (Maywood) 2001; 226: 85-96.
Fernandez-Novoa L, Cacabelos R. Histamine function in brain disorders.Behav Brain Res 2001; 124: 213-233.
Fields RD, Stevens-Graham B. New insights into neuron-glia commu-nication. Science 2002; 298: 556-562.
Fillion G. Potential of 5-HT-moduline as a drug target for affectivedisorders. Curr Opin Investig Drugs 2000; 1: 104-109.
Fink H, Rex A, Voits M, Voigt JP. Major biological actions of CCK -a critical evaluation of research findings. Exp Brain Res 1998; 123:77-83.
Fischer-Colbrie R, Kirchmair R, Kahler CM, Wiedermann CJ, Saria A.Secretoneurin: a new player in angiogenesis and chemotaxis linkingnerves, blood vessels and the immune system. Curr Protein Pept Sci2005; 6: 373-385.
Fišar Z. Biochemické hypotézy afektivních poruch. Galén, Praha 1998;103 pp.
Fitzsimons JT. Angiotensin, thirst, and sodium appetite. Physiol Rev1998; 78: 583-686.
Flatmark T. Catecholamine biosynthesis and physiological regulationin neuroendocrine cells. Acta Physiol Scand 2000; 168: 1-17.

226 NEUROTRANSMITERY
Flugge G, Van Kampen M, Mijnster MJ. Perturbations in brain mono-amine systems during stress. Cell Tissue Res 2004; 315: 1-14.
Förster C.Glucocorticoid regulation of blood brain barrier permeability.Physiology News 2005; 61: 34-35
Fredholm BB, IJzerman AP, Jacobson KA, Klotz KN, Linden J. Interna-tional Union of Pharmacology. XXV. Nomenclature and classificationof adenosine receptors. Pharmacol Rev 2001; 53: 527-552.
Freedman R. Schizophrenia. N Engl J Med 2003; 349: 1738-1749.
Fritzsch B. Of mice and genes: evolution of vertebrate brain develop-ment. Brain Behav Evol 1998; 52: 207-217.
Froehlich JC. Opioid peptides. Alcohol Health Res World 1997; 21: 132-136.
Fujii R, Yoshida H, Fukusumi S, Habata Y, Hosoya M, Kawamata Y,Yano T, Hinuma S, Kitada C, Asami T, Mori M, Fujisawa Y, FujinoM. Identification of a neuropeptide modified with bromine as anendogenous ligand for GPR7. J Biol Chem 2002; 277: 34010-34016.
Fukuda T, Kosaka T. The dual network of GABAergic interneuronslinked by both chemical and electrical synapses: a possible infras-tructure of the cerebral cortex. Neurosci Res 2000; 38: 123-130.
Furst S. Transmitters involved in antinociception in the spinal cord.Brain Res Bull 1999; 48: 129-141.
Gadea A, Lopez-Colome AM. Glial transporters for glutamate, glycineand GABA I. Glutamate transporters. J Neurosci Res 2001a; 63:453-460.
Gadea A, Lopez-Colome AM. Glial transporters for glutamate, glycine,and GABA: II. GABA transporters. J Neurosci Res 2001b; 63: 461-468.
Gadea A, Lopez-Colome AM. Glial transporters for glutamate, glycine,and GABA III. Glycine transporters. J Neurosci Res 2001c; 64: 218-222.
Gai WP, Geffen LB, Denoroy L, Blessing WW. Loss of C1 and C3epinephrine-synthesizing neurons in the medulla oblongata in Par-kinson’s disease. Ann Neurol 1993; 33: 357-367.
Ganong WF, Alpert LC, Lee TC. ACTH and the regulation of adreno-cortical secretion. N Engl J Med 1974; 290: 1006-1011.

Literatúra 227
Ganong WF. Circumventricular organs: definition and role in the re-gulation of endocrine and autonomic function. Clin Exp PharmacolPhysiol 2000; 27: 422-427.
Ganong WF. Synaptický a junkční přenos. In: Ganong WF (Ed). Přeh-led lékařské fyziologie. 20. vyd. Galén, Praha 2005: 87-120.
Garattini S, Samanin R. Biochemical hypotheses on antidepressantdrugs: a guide for clinicians or a toy for pharmacologists? PsycholMed 1988; 18: 287-304.
Garcia JM, Garcia-Touza M, Hijazi RA, Taffet G, Epner D, Mann D,Smith RG, Cunningham GR, Marcelli M. Active ghrelin levels andactive to total ghrelin ratio in cancer-induced cachexia. J Clin En-docrinol Metab 2005; 90: 2920-2926.
Garcia-Sainz JA. Adrenaline and its receptors: one hundred years ofresearch. Arch Med Res 1995; 26: 205-212.
Ge S, Goh EL, Sailor KA, Kitabatake Y, Ming GL, Song H. GABA re-gulates synaptic integration of newly generated neurons in the adultbrain. Nature 2006; 439: 589-593.
Gilbey MP, Spyer KM. Essential organization of the sympathetic ner-vous system. Baillieres Clin Endocrinol Metab 1993; 7: 259-278.
Gillespie CF, Nemeroff CB. Hypercortisolemia and depression. Psycho-som Med 2005; 67: S26-S28.
Gimpl G, Fahrenholz F. The oxytocin receptor system: structure, func-tion, and regulation. Physiol Rev 2001; 81: 629-683.
Gladkevich A, Korf J, Hakobyan VP, Melkonyan KV. The peripheralGABAergic system as a target in endocrine disorders. Auton Ne-urosci 2006; 124: 1-8.
Glover JC. Development of specific connectivity between premotor ne-urons and motoneurons in the brain stem and spinal cord. PhysiolRev 2000; 80: 615-647.
Goehler LE, Gaykema RP, Hansen MK, Anderson K, Maier SF, WatkinsLR. Vagal immune-to-brain communication: a visceral chemosensorypathway. Auton Neurosci 2000; 85: 49-59.
Goff DC, Coyle JT. The emerging role of glutamate in the pathophy-siology and treatment of schizophrenia. Am J Psychiatry 2001; 158:1367-1377.

228 NEUROTRANSMITERY
Gold PW, Chrousos GP. Organization of the stress system and itsdysregulation in melancholic and atypical depression: high vs lowCRH/NE states. Mol Psychiatry 2002; 7: 254-275.
Goldstein DS, Eldadah BA, Holmes C, Pechnik S, Moak J, Saleem A,Sharabi Y. Neurocirculatory abnormalities in Parkinson disease withorthostatic hypotension: independence from levodopa treatment. Hy-pertension 2005; 46: 1333-1339.
Goldstein DS, Holmes C, Li ST, Bruce S, Metman LV, Cannon RO. 3rd.Cardiac sympathetic denervation in Parkinson disease. Ann InternMed 2000; 133: 338-347.
Goldstein DS. The Autonomic Nervous System in Health and Disease.New York Basel, Marcel Dekker 2001; 618 pp.
Gonzalez-Rey E, Chorny A, Fernandez-Martin A, Varela N, DelgadoM. Vasoactive intestinal peptide family as a therapeutic target forParkinson’s disease. Expert Opin Ther Targets 2005; 9: 923-929.
Goodall M. Studies of adrenaline and noradrenaline in mammalian he-art and suprarenals. Acta Physiol Scand Suppl 1951; 24: 7-51.
Goodson JL, Bass AH. Social behavior functions and related anatomicalcharacteristics of vasotocin/vasopressin systems in vertebrates. BrainRes Brain Res Rev 2001; 35: 246-265.
Gottsch ML, Clifton DK, Steiner RA. Galanin-like peptide as a link inthe integration of metabolism and reproduction. Trends EndocrinolMetab 2004; 15: 215-221.
Grafton ST. Contributions of functional imaging to understanding par-kinsonian symptoms. Curr Opin Neurobiol 2004; 14: 715-719.
Greenough A, Cole G, Lewis J, Lockton A, Blundell J. Untangling theeffects of hunger, anxiety, and nausea on energy intake during intra-venous cholecystokinin octapeptide (CCK-8) infusion. Physiol Behav1998; 65: 303-310.
Griffond B, Baker BI. Cell and molecular cell biology of melanin-concentrating hormone. Int Rev Cytol 2002; 213: 233-277.
Grill HJ, Kaplan JM. Caudal brain stem participates in the distributedneural control of feeding. In: Handbook of behavioral neurobiology,Vol. 10. Stricker EM (Ed). New York: Plenum Pres 1990, p. 125-149.

Literatúra 229
Grimaldi B, Bonnin A, Fillion MP, Prudhomme N, Fillion G. 5-Hydroxytryptamine-moduline: a novel endogenous peptide involvedin the control of anxiety. Neuroscience 1999; 93: 1223-1225.
Gu Q. Neuromodulatory transmitter systems in the cortex and theirrole in cortical plasticity. Neuroscience 2002; 111: 815-835.
Guan JL, Saotome T, Wang QP, Funahashi H, Hori T, Tanaka S, ShiodaS. Orexinergic innervation of POMC-containing neurons in the ratarcuate nucleus. Neuroreport 2001; 12: 547-551.
Guarini S, Altavilla D, Cainazzo MM, Giuliani D, Bigiani A, MariniH, Squadrito G, Minutoli L, Bertolini A, Marini R, Adamo EB, Ve-nuti FS, Squadrito F. Efferent vagal fibre stimulation blunts nuclearfactor-kappaB activation and protects against hypovolemic hemorr-hagic shock. Circulation 2003; 107: 1189-1194.
Guix FX, Uribesalgo I, Coma M, Munoz FJ. The physiology and pat-hophysiology of nitric oxide in the brain. Prog Neurobiol 2005; 76:126-152.
Gundlach AL, Burazin TC, Larm JA. Distribution, regulation and roleof hypothalamic galanin systems: renewed interest in a pleiotropicpeptide family. Clin Exp Pharmacol Physiol 2001; 28: 100-105.
Gupta A, Wang Y, Markram H. Organizing principles for a diversityof GABAergic interneurons and synapses in the neocortex. Science2000; 287: 273-278.
Gutierrez R. The dual glutamatergic-GABAergic phenotype of hippo-campal granule cells. Trends Neurosci 2005; 28: 297-303.
Guyenet PG, Stornetta RL, Schreihofer AM, Pelaez NM, Hayar A, Ai-cher S, Llewellyn-Smith IJ. Opioid signalling in the rat rostral ven-trolateral medulla. Clin Exp Pharmacol Physiol 2002; 29: 238-242.
Haas H, Panula P. The role of histamine and the tuberomamillary nuc-leus in the nervous system. Nat Rev Neurosci 2003; 4: 121-130.
Haddad JJ, Saade NE, Safieh-Garabedian B. Cytokines and neuro-immune-endocrine interactions: a role for the hypothalamic-pituitary-adrenal revolving axis. J Neuroimmunol 2002; 133: 1-19.
Haddad JJ. N-methyl-d-aspartate (NMDA) and the regulation ofmitogen-activated protein kinase (MAPK) signaling pathways: A re-volving neurochemical axis for therapeutic intervention? Prog Ne-urobiol 2005; 77: 252-282.

230 NEUROTRANSMITERY
Hagg T. Molecular regulation of adult CNS neurogenesis: an integratedview. Trends Neurosci 2005; 28: 589-595.
Hahn TM, Breininger JF, Baskin DG, Schwartz MW. Coexpression ofAgrp and NPY in fasting-activated hypothalamic neurons. Nat Ne-urosci 1998; 1: 271-272.
Halbreich U, Asnis GM, Shindledecker R, Zumoff B, Nathan RS. Cortisolsecretion in endogenous depression. I. Basal plasma levels. Arch GenPsychiatry 1985; 42: 904-908.
Hallberg M, Nyberg F. Neuropeptide conversion to bioactive fragments-an important pathway in neuromodulation. Curr Protein Pept Sci2003; 4: 31-44.
Hannon J, Hoyer D. Serotonin receptors and systems: endless diversity?Acta Biol Szeged 2002; 46: 1-12.
Hansen MB. The enteric nervous system I: organisation and classifica-tion. Pharmacol Toxicol 2003; 92: 105-113.
Hansen TV, Nielsen FC. Regulation of neuronal cholecystokinin genetranscription. Scand J Clin Lab Invest Suppl 2001; 234: 61-67.
Harrison PJ. The neuropathology of primary mood disorder. Brain2002; 125: 1428-1449.
Harro J, Oreland L. Depression as a spreading adjustment disorder ofmonoaminergic neurons: a case for primary implication of the locuscoeruleus. Brain Res Brain Res Rev 2001; 38: 79-128.
Hashimoto K, Shimizu E, Iyo M. Critical role of brain-derived neurot-rophic factor in mood disorders. Brain Res Brain Res Rev 2004; 45:104-114.
Haskell-Luevano C, Chen P, Li C, Chang K, Smith MS, Cameron JL,Cone RD. Characterization of the neuroanatomical distribution ofagouti-related protein immunoreactivity in the rhesus monkey andthe rat. Endocrinology 1999; 140: 1408-1415.
Hasler G, Drevets WC, Manji HK, Charney DS. Discovering endophe-notypes for major depression. Neuropsychopharmacology 2004; 29:1765-1781.
Hauger RL, Grigoriadis DE, Dallman MF, Plotsky PM, Vale WW, Daut-zenberg FM. International Union of Pharmacology. XXXVI. Currentstatus of the nomenclature for receptors for corticotropin-releasingfactor and their ligands. Pharmacol Rev 2003; 55: 21-26.

Literatúra 231
Haxhiu MA, Tolentino-Silva F, Pete G, Kc P, Mack SO. Monoaminergicneurons, chemosensation and arousal. Resp Physiol 2001; 129: 191-209.
Hayley S, Poulter MO, Merali Z, Anisman H. The pathogenesis of clini-cal depression: stressor- and cytokine-induced alterations of neurop-lasticity. Neuroscience 2005; 135: 659-678.
Hay-Schmidt A. The evolution of the serotonergic nervous system. ProcBiol Sci 2000; 267: 1071-1079.
Head GA. Agmatine and imidazoline systems-fourth international sym-posium: 9-11 April 2003, San Diego, CA, USA. IDrugs 2003; 6: 544-547.
Heilig M. The NPY system in stress, anxiety and depression. Neuro-peptides 2004; 38: 213-224.
Hein L, Kobilka BK. Adrenergic receptor signal transduction and regu-lation. Neuropharmacology 1995; 34: 357-366.
Heinricher MM. Orphanin FQ/nociceptin: from neural circuitry to be-havior. Life Sci 2003; 73: 813-822.
Heinrichs SC, Koob GF. Corticotropin-releasing factor in brain: a rolein activation, arousal, and affect regulation. J Pharmacol Exp Ther2004; 311: 427-440.
Heinz A. The dopamine hypothesis of schizophrenia. New findings foran old theory. Nervenarzt 2000; 71: 54-57.
Henquet C, Murray R, Linszen D, van Os J. The environment and schi-zophrenia: the role of cannabis use. Schizophr Bull 2005; 31: 608-612.
Herlenius E, Lagercrantz H. Development of neurotransmitter systemsduring critical periods. Exp Neurol 2004; 190: S8-S21.
Herlenius E, Lagercrantz H. Neurotransmitters and neuromodulatorsduring early human development. Early Hum Dev 2001; 65: 21-37.
Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, OstranderMM, Choi DC, Cullinan WE. Central mechanisms of stress in-tegration: hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocrinol 2003; 24: 151-180.
Herpfer I, Lieb K. Substance P receptor antagonists in psychiatry: ra-tionale for development and therapeutic potential. CNS Drugs 2005;19: 275-293.

232 NEUROTRANSMITERY
Hestrin S, Galarreta M. Electrical synapses define networks of neocor-tical GABAergic neurons. Trends Neurosci 2005; 28: 304-309.
Heuser I, Yassouridis A, Holsboer F. The combined dexametha-sone/CRH test: a refined laboratory test for psychiatric disorders.J Psychiatr Res 1994; 28: 341-356.
Hill SJ, Ganellin CR, Timmerman H, Schwartz JC, Shankley NP, YoungJM, Schunack W, Levi R, Haas HL. International Union of Pharma-cology. XIII. Classification of histamine receptors. Pharmacol Rev1997; 49: 253-278.
Hindmarch I. Expanding the horizons of depression: beyond the mono-amine hypothesis. Hum Psychopharmacol 2001; 16: 203-218.
Hinson JP, Kapas S, Smith DM. Adrenomedullin, a multifunctionalregulatory peptide. Endocr Rev 2000; 21: 138-167.
Hokfelt T, Bartfai T, Bloom F. Neuropeptides: opportunities for drugdiscovery. Lancet Neurol 2003; 2: 463-472.
Hokfelt T, Broberger C, Xu ZQ, Sergeyev V, Ubink R, Diez M. Neuro-peptides - an overview. Neuropharmacology 2000; 39: 1337-1356.
Hokfelt T, Holmberg K, Shi TJ, Broberger C. CCK-ergic mechanismsin sensory systems. Scand J Clin Lab Invest Suppl 2001a; 234: 69-74.
Hokfelt T, Millhorn D, Seroogy K, Tsuruo Y, Ceccatelli S, Lindh B,Meister B, Melander T, Schalling M, Bartfai T a spol. Coexistenceof peptides with classical neurotransmitters. Experientia 1987; 43:768-780.
Hokfelt T, Pernow B, Wahren J. Substance P: a pioneer amongst ne-uropeptides. J Intern Med 2001b; 249: 27-40.
Hollister LE. Health aspects of cannabis: revisited. Int J Neuropsychop-harmacol. 1998; 1: 71-80
Holmes A, Heilig M, Rupniak NM, Steckler T, Griebel G. Neuropep-tide systems as novel therapeutic targets for depression and anxietydisorders. Trends Pharmacol Sci 2003; 24: 580-588.
Holsboer F. The role of peptides in treatment of psychiatric disorders.J Neural Transm Suppl 2003; 17-34.
Holt A. Imidazoline binding sites on receptors and enzymes: emergingtargets for novel antidepressant drugs? J Psychiatry Neurosci 2003;28: 409-414.

Literatúra 233
Holzer P, Schicho R, Holzer-Petsche U, Lippe IT. The gut as a neuro-logical organ. Wien Klin Wschr 2001; 113: 647-660.
Hormuzdi SG, Filippov MA, Mitropoulou G, Monyer H, Bruzzone R.Electrical synapses: a dynamic signaling system that shapes the ac-tivity of neuronal networks. Biochim Biophys Acta 2004; 1662: 113-137.
Hoschl C, Hajek T. Hippocampal damage mediated by corticosteroids-aneuropsychiatric research challenge. Eur Arch Psychiatry Clin Ne-urosci 2001; 251: 81-88.
Höschl C, Libiger J, Švestka J (Eds). Psychiatrie. Tigis, Praha, 2004;884 pp.
Hosoya Y, Matsukawa, Okado N, Sugiura Y, Kohno K. Oxytocinergicinnervation to the upper thoracic sympathetic preganglionic neuronsin the rat. A light and electron microscopical study using a combinedretrograde transport and immunocytochemical technique. Exp BrainRes 1995; 107: 9-16.
Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, Fel-der CC, HerkenhamM, Mackie K, Martin BR, Mechoulam R, PertweeRG. International Union of Pharmacology. XXVII. Classification ofcannabinoid receptors. Pharmacol Rev 2002; 54: 161-202.
Huang ZL, QuWM, Li WD, Mochizuki T, Eguchi N, Watanabe T, UradeY, Hayaishi O. Arousal effect of orexin A depends on activation of thehistaminergic system. Proc Natl Acad Sci USA 2001; 98: 9965-9970.
Hudson A. Imidazoline receptors. Tocris Reviews 2000; 12: 1-5.
Hungs M, Fan J, Lin L, Lin X, Maki RA, Mignot E. Identification andfunctional analysis of mutations in the hypocretin (orexin) genes ofnarcoleptic canines. Genome Res 2001; 11: 531-539.
Charmandari E, Kino T, Chrousos GP. Glucocorticoids and their acti-ons: an introduction. Ann NY Acad Sci 2004; 1024: 1-8.
Charmandari E, Tsigos C, Chrousos G. Endocrinology of the stress res-ponse. Annu Rev Physiol 2005; 67: 259-284.
Charney DS, Manji HK. Life stress, genes, and depression: multiple pat-hways lead to increased risk and new opportunities for intervention.Sci STKE 2004; 2004: 1-11.
Charney DS. Neuroanatomical circuits modulating fear and anxiety be-haviors. Acta Psychiatr Scand Suppl 2003; 417: 38-50.

234 NEUROTRANSMITERY
Chen X, Jhee KH, Kruger WD. Production of the neuromodulator H2Sby cystathionine beta-synthase via the condensation of cysteine andhomocysteine. J Biol Chem 2004; 279: 52082-52086.
Chiba T. Monoamine-containing paraneurons in the sympathetic gan-glia of mammals. Arch Histol Jpn 1977; 40: 163-176.
Chinta SJ, Andersen JK. Dopaminergic neurons. Int J Biochem CellBiol 2005; 37: 942-946.
Chrousos GP, Gold PW. A healthy body in a healthy mind-and viceversa-the damaging power of „uncontrollableÿ stress. J Clin EndocrinMetabol 1998; 83: 1842-1845.
Ibata Y, Iijima N, Kataoka Y, Kakihara K, Tanaka M, Hosoya M, Hi-numa S. Morphological survey of prolactin-releasing peptide and itsreceptor with special reference to their functional roles in the brain.Neurosci Res 2000; 38: 223-230.
Illes P, Ribeiro JA. Neuronal P2 receptors of the central nervous system.Curr Top Med Chem 2004; 4: 831-838.
Imaki T, Shibasaki T, Hotta M, Demura H. Early induction of c-fosprecedes increased expression of corticotropin- releasing factor mes-senger ribonucleic acid in the paraventricular nucleus after immobi-lization stress. Endocrinology 1992; 131: 240-246.
Inglis FM, Furia F, Zuckerman KE, Strittmatter SM, Kalb RG. The roleof nitric oxide and NMDA receptors in the development of motorneuron dendrites. J Neurosci 1998; 18: 10493-10501.
Irving AJ, Rae MG, Coutts AA. Cannabinoids on the brain. Sci WordJ 2002; 2: 632-648.
Iyer LM, Aravind L, Coon SL, Klein DC, Koonin EV. Evolution ofcell-cell signaling in animals: did late horizontal gene transfer frombacteria have a role? Trends Genet 2004; 20: 292-299.
Jan YN, Jan LY. Dendrites. Genes Dev 2001; 15: 2627-2641.
Janig W. Non-nicotinic transmission in autonomic ganglia revisited-animportant physiological function? J Physiol 2005; 566: 1-2.
Jankowski M, Danalache B, Wang D, Bhat P, Hajjar F, MarcinkiewiczM, Paquin J, McCann SM, Gutkowska J. Oxytocin in cardiac onto-geny. Proc Natl Acad Sci USA 2004; 101: 13074-13079.

Literatúra 235
Jedlička P, Beňušková Ľ, Mačáková J, Ostatníková D. Molekulové me-chanizmy učenia a pamäte. In: Patofyziológia. 6. vyd. Hulin I (Ed).Slovak Academic Press, Bratislava 2002; pp. 1183-1199.
Jessen KR. Glial cells. Int J Biochem Cell Biol 2004; 36: 1861-1867.
Jezova D, Ochedalski T, Kiss A, Aguilera G. Brain angiotensin II mo-dulates sympathoadrenal and hypothalamic pituitary adrenocorticalactivation during stress. J Neuroendocrinol 1998; 10: 67-72.
Ježová D. Antiparkinsoniká. In: Farmakológia a klinická farmakológianervového systému. Kriška M, Ježová D, Kořínková V (Eds), Askle-pios, Bratislava 1998; pp. 96-103.
Jhamandas JH, Jhamandas A, Harris KH. New central projections ofneuropeptide FF: colateral branching pathways in the brain stemand hypothalamus in the rat. J Chem Neuroanat 2001; 21: 171-179.
Johnson AK, Gross PM. Sensory circumventricular organs and brainhomeostatic pathways. FASEB J 1993; 7: 678-686.
Kafka J a spol. (Eds). Psychiatria. Osveta, Martin, 1998; 255 pp.
Kageyama H, Takenoya F, Kita T, Hori T, Guan JL, Shioda S. Galanin-like peptide in the brain: effects on feeding, energy metabolism andreproduction. Regul Pept 2005; 126: 21-26.
Kalra SP, Dube MG, Pu S, Xu B, Horvath TL, Kalra PS. Interactingappetite-regulating pathways in the hypothalamic regulation of bodyweight. Endocr Rev 1999; 20: 68-100.
Kalra SP, Xu B, Dube MG, Moldawer LL, Martin D, Kalra PS. Leptinand ciliary neotropic factor (CNTF) inhibit fasting induced suppres-sion of luteinizing hormone release in rats: role of neuropeptide Y.Neurosci Lett 1998; 240: 45-49.
Kamoun P. Endogenous production of hydrogen sulfide in mammals.Amino Acids 2004; 26: 243-254.
Kandel ER, Schwartz JH, Jessell TM (Eds). Principles of neural scien-ces. McGraw-Hill, New York, 2000; 1414 pp.
Kandel ER. The molecular biology of memory storage: a dialogue bet-ween genes and synapses. Science 2001; 294: 1030-1038.
Kandlhofer S, Hoertnagl B, Czech T, Baumgartner C, Maier H, Novak K,Sperk G. Chromogranins in temporal lobe epilepsy. Epilepsia 2000;41: S111-S114.

236 NEUROTRANSMITERY
Kaneko H, Tache Y, Kusugami K. Importance of medullary thyrotropin-releasing hormone in brain-gut circuits regulating gastric integrity:preclinical studies. J Gastroenterol 2002; 37: 128-132.
Karege F, Perret G, Bondolfi G, Schwald M, Bertschy G, Aubry JM.Decreased serum brain-derived neurotrophic factor levels in majordepressed patients. Psychiatry Res 2002; 109: 143-148.
Kastin AJ, Pan W. Dynamic regulation of leptin entry into brain bythe blood-brain barrier. Regul Pept 2000; 92: 37-43.
Kawano H, Honma S, Honma A, Horie M, Kawano Y, Hayashi S.Melanin-concentrating hormone neuron system: the Wide Web thatcontrols the feeding. Anat Sci Int 2002; 77: 149-160.
Kendler KS, Kuhn JW, Vittum J, Prescott CA, Riley B. The interactionof stressful life events and a serotonin transporter polymorphism inthe prediction of episodes of major depression: a replication. ArchGen Psychiatry 2005; 62: 529-535.
Kerman IA, Enquist LW, Watson SJ, Yates BJ. Brain stem substratesof sympatho-motor circuitry identified using trans-synaptic tracingwith pseudorabies virus recombinants. J Neurosci 2003; 23: 4657-4666.
Khakh BS, Burnstock G, Kennedy C, King BF, North RA, Seguela P,Voigt M, Humphrey PP. International union of pharmacology. XXIV.Current status of the nomenclature and properties of P2X receptorsand their subunits. Pharmacol Rev 2001; 53: 107-118.
Khawaja AM, Rogers DF. Tachykinins: receptor to effector. Int J Bio-chem Cell Biol 1996; 28: 721-738.
Kiecolt-Glaser JK, Robles TF, Heffner KL, Loving TJ, Glaser R. Psycho-oncology and cancer: psychoneuroimmunology and cancer. Ann On-col 2002; 13: 165-169.
Kiefer F, Wiedemann K. Neuroendocrine pathways of addictive beha-viour. Addict Biol 2004; 9: 205-212.
Kim DY, Camilleri M. Serotonin: a mediator of the brain-gut connec-tion. Am J Gastroenterol 2000; 95: 2698-2709.
Kimura H. Hydrogen sulfide as a neuromodulator. Mol Neurobiol 2002;26: 13-19.
Kimura H. Hydrogen sulfide induces cyclic AMP and modulates theNMDA receptor. Biochem Biophys Res Commun 2000; 267: 129-133.

Literatúra 237
Kinkead B, Binder EB, Nemeroff CB. Does neurotensin mediate theeffects of antipsychotic drugs? Biol Psychiatry 1999; 46: 340-351.
Kinkead B, Nemeroff CB. Neurotensin, schizophrenia, and antipsychoticdrug action. Int Rev Neurobiol 2004; 59: 327-349.
Kirchgessner AL. Orexins in the brain-gut axis. Endocr Rev 2002; 23:1-15.
Kirstein SL, Insel PA. Autonomic nervous system pharmacogenomics:a progress report. Pharmacol Rev 2004; 56: 31-52.
Kis B, Abraham CS, Deli MA, Kobayashi H, Niwa M, Yamashita H,Busija DW, Ueta Y.Adrenomedullin, an autocrine mediator of blood-brain barrier function. Hypertens Res 2003; 26: S61-S70.
Kiss A, Mikkelsen JD. Oxytocin - anatomy and functional assignments:a minireview. Endocrin Regul 2005; 39: 97-105.
Kiss A. Hypotalamo-hypofýzo-adrenokortikálna os: organizácia, dyna-mické aspekty a vplyv stresu na aktivitu CRH neurónu. Bratisl LekListy 1999; 100: 611-624.
Kiss JP, Vizi ES. Nitric oxide: a novel link between synaptic and non-synaptic transmission. Trends Neurosci 2001; 24: 211-215.
Klavdieva MM. The history of neuropeptides 1. Front Neuroendocrinol1995; 16: 293-321.
Kleinz MJ, Davenport AP. Emerging roles of apelin in biology andmedicine. Pharmacol Ther 2005; 107: 198-211.
Knigge KM, Baxter-Grillo D, Speciale J, Wagner J. Melanotropic pep-tides in the mammalian brain: the melanin-concentrating hormone.Peptides 1996; 17: 1063-1073.
Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K. Gh-relin is a growth-hormone-releasing acylated peptide from stomach.Nature 1999; 402: 656-660.
Kolaj M, Shefchyk SJ, Renaud LP. Two conductances mediatethyrotropin-releasing-hormone-induced depolarization of neonatalrat spinal preganglionic and lateral horn neurons. J Neurophysiol1997; 78: 1726-1729.
Koob GF, Greenwell TN. Neuropeptide S: a novel activating anxiolytic?Neuron 2004; 43: 441-442.

238 NEUROTRANSMITERY
Koob GF, Sanna PP, Bloom FE. Neuroscience of addiction. Neuron1998; 21: 467-476.
Korbonits M, Goldstone AP, Gueorguiev M, Grossman AB. Ghrelin –a hormone with multiple functions. Front Neuroendocrinol 2004; 25:27-68.
Kordower JH, Goetz CG. The first miracle in neurodegenerative disease:the discovery of oral levodopa. Brain Res Bull 1999; 50: 377-378.
Korner J, Leibel RL. To eat or not to eat - how the gut talks to thebrain. N Engl J Med 2003; 349: 926-928.
Kosfeld M, Heinrichs M, Zak PJ, Fischbacher U, Fehr E. Oxytocin inc-reases trust in humans. Nature 2005; 435: 673-676.
Kovacs GL, Sarnyai Z, Szabo G. Oxytocin and addiction: a review.Psychoneuroendocrinology 1998; 23: 945-962.
Krnjevic K, Phillis JW. Iontophoretic studies of neurones in the mam-malian cerebral cortex. J Physiol 1963; 165: 274-304.
Krnjevic K, Puil E, Werman R. GABA and glycine actions on spinalmotoneurons. Can J Physiol Pharmacol 1977; 55: 658-669.
Krnjevic K. From ’soup physiology’ to normal brain science. J Physiol2005; 569: 1-2.
Krowicki ZK. Role of selected peptides in the vagal regulation of gastricmotor and endocrine pancreatic function. J Physiol Pharmacol 1996;47: 399-409.
Kugaya A, Sanacora G. Beyond monoamines: glutamatergic function inmood disorders. CNS Spectr 2005; 10: 808-819.
Kukanova B, Mravec B. Complex intracardiac nervous system. BratislLek Listy 2006; 107: 45-51.
Kukkonen JP, Holmqvist T, Ammoun S, Akerman KE. Functions of theorexinergic/hypocretinergic system. Am J Physiol Cell Physiol 2002;283: C1567-C1591.
Kuramochi M, Onaka T, Kohno D, Kato S, Yada T. Galanin-like pep-tide stimulates food intake via activation of neuropeptide Y neuronsin the hypothalamic dorsomedial nucleus of the rat. Endocrinology2006; 147: 1744-1752.
Landgraf R, Neumann ID. Vasopressin and oxytocin release within thebrain: a dynamic concept of multiple and variable modes of neuro-peptide communication. Front Neuroendocrinol 2004; 25: 150-176.

Literatúra 239
Lang AE, Lozano AM. Parkinson’s disease. First of two parts. N EnglJ Med 1998; 339: 1044-1053.
Langer SZ. 25 years since the discovery of presynaptic receptors: presentknowledge and future perspectives. Trends Pharmacol Sci 1997; 18:95-99.
Laruelle M, Kegeles LS, Abi-Dargham A. Glutamate, dopamine, andschizophrenia: from pathophysiology to treatment. Ann NY AcadSci 2003; 1003: 138-158.
Latchman DS. Urocortin. Int J Biochem Cell Biol 2002; 34: 907-910.
LeDoux J. Fear and the brain: where have we been, and where are wegoing? Biol Psychiatry 1998; 44: 1229-1238.
Lee AL, Ogle WO, Sapolsky RM. Stress and depression: possible linksto neuron death in the hippocampus. Bipolar disorders 2002; 4: 117-128.
Lee SP, So CH, Rashid AJ, Varghese G, Cheng R, Lanca AJ, O’DowdBF, George SR. Dopamine D1 and D2 receptor Co-activation gene-rates a novel phospholipase C-mediated calcium signal. J Biol Chem2004; 279: 35671-35678.
Legendre P. The glycinergic inhibitory synapse. Cell Mol Life Sci 2001;58: 760-793.
Lehnert H, Schulz C, Dieterich K. Physiological and neurochemical as-pects of corticotropin-releasing factor actions in the brain: the roleof the locus coeruleus. Neurochem Res 1998; 23: 1039-1052.
Lechner T, Adlassnig C, Humpel C, Kaufmann WA, Maier H, Reinstad-ler-Kramer K, Hinterholzl J, Mahata SK, Jellinger KA, MarksteinerJ. Chromogranin peptides in Alzheimer’s disease. Exp Gerontol 2004;39: 101-113.
Leonard BE. Drug treatment of depression. In: Fundamentals of psy-chopharmacology. Third edition. Leonard BE (Ed), John Wiley &Sons, Chichester 2003; pp. 153-192.
Levitt P, Reinoso B, Jones L. The critical impact of early cellular envi-ronment on neuronal development. Prev Med 1998; 27: 180-183.
Lewis DE, Shellard L, Koeslag DG, Boer DE, McCarthy HD, Mc KibbinPE, Russell JC, Williams G. Intense exercise and food restrictioncause similar hypothalamic neuropeptide Y increases in rats. Am JPhysiol 1993; 264: E279-E284.

240 NEUROTRANSMITERY
Lewis MJ, Rada P, Johnson DF, Avena NM, Leibowitz SF, Hoebel BG.Galanin and alcohol dependence: neurobehavioral research. Neuro-peptides 2005; 39: 317-321.
Licinio J, Wong ML. The role of inflammatory mediators in the bio-logy of major depression: central nervous system cytokines modu-late the biological substrate of depressive symptoms, regulate stress-responsive systems, and contribute to neurotoxicity and neuropro-tection. Mol Psychiatry 1999; 4: 317-327.
Lie DC, Song H, Colamarino SA, Ming GL, Gage FH. Neurogenesis inthe adult brain: new strategies for central nervous system diseases.Annu Rev Pharmacol Toxicol 2004; 44: 399-421.
Lindemann L, Hoener MC. A renaissance in trace amines inspired bya novel GPCR family. Trends Pharmacol Sci 2005; 26: 274-281.
Lipton P. Ischemic cell death in brain neurons. Physiol Rev 1999; 79:1431-1568.
Llewellyn-Smith IJ, Cassam AK, Krenz NR, Krassioukov AV, WeaverLC. Glutamate- and GABA-immunoreactive synapses on sympathe-tic preganglionic neurons caudal to a spinal cord transsection in rats.Neuroscience 1997; 80: 1225-1235.
Loffelholz K. Brain choline has a typical precursor profile. J PhysiolParis 1998; 92: 235-239.
Lovejoy DA, Balment RJ. Evolution and physiology of the corticotropin-releasing factor (CRF) family of neuropeptides in vertebrates. GenComp Endocrinol 1999; 115: 1-22.
Ludwig M, Leng G. Dendritic peptide release and peptide-dependentbehaviours. Nat Rev Neurosci 2006; 7: 126-136.
Ludwig M, Pittman QJ. Talking back: dendritic neurotransmitter rele-ase. Trends Neurosci 2003; 26: 255-261.
Lukas RJ, Changeux JP, Le Novere N, Albuquerque EX, Balfour DJ,Berg DK, Bertrand D, Chiappinelli VA, Clarke PB, Collins AC, DaniJA, Grady SR, Kellar KJ, Lindstrom JM, Marks MJ, Quik M, TaylorPW, Wonnacott S. International Union of Pharmacology. XX. Cur-rent status of the nomenclature for nicotinic acetylcholine receptorsand their subunits. Pharmacol Rev 1999; 51: 397-401.

Literatúra 241
Lundberg JM, Franco-Cereceda A, Lacroix JS, Pernow J. NeuropeptideY and sympathetic neurotransmission. Ann NY Acad Sci 1990; 611:166-174.
Lynch JW. Molecular structure and function of the glycine receptorchloride channel. Physiol Rev 2004; 84: 1051-1095.
Mackie K, Stella N. Cannabinoid receptors and endocannabinoids: evi-dence for new players. AAPS J 2006; 8: 298-306.
Magloczky Z, Freund TF. Impaired and repaired inhibitory circuits inthe epileptic human hippocampus. Trends Neurosci 2005; 28: 334-340.
Magnaghi V, Ballabio M, Consoli A, Lambert JJ, Roglio I, Melcangi RC.GABA receptor-mediated effects in the peripheral nervous system:A cross-interaction with neuroactive steroids. Mol Neurosci 2006; 28:89-102.
Maguire JJ, Davenport AP. Is urotensin-II the new endothelin? Br JPharmacol 2002; 137: 579-588.
Maguire-Zeiss KA, Short DW, Federoff HJ. Synuclein, dopamine andoxidative stress: co-conspirators in Parkinson’s disease? Brain ResMol Brain Res 2005; 134: 18-23.
Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressanttreatment increases neurogenesis in adult rat hippocampus. J Ne-urosci 2000; 20: 9104-9110.
Malberg JE. Implications of adult hippocampal neurogenesis in anti-depressant action. J Psychiatry Neurosci 2004; 29: 196-205.
Malhi GS, Parker GB, Greenwood J. Structural and functional modelsof depression: from sub-types to substrates. Acta Psychiatr Scand2005; 111: 94-105.
Manji HK, Drevets WC, Charney DS. The cellular neurobiology of dep-ression. Nat Med 2001; 7: 541-547.
Mann JJ. The medical management of depression. N Engl J Med 2005;353: 1819-1834.
Marder E. Neural signalling: Does colocalization imply cotransmission?Curr Biol 1999; 9: R809-R811.
Maršala J. Systematická a funkčná neuroanatómia. Osveta, Martin1985; 784 pp.

242 NEUROTRANSMITERY
Martin WJ, Malmberg AB, Basbaum AI. Pain: nocistatin spells relief.Curr Biol 1998; 8: R525-R527.
Massot O, Rousselle JC, Grimaldi B, Cloez-Tayarani I, Fillion MP, Plan-tefol M, Bonnin A, Prudhomme N, Fillion G. Molecular, cellular andphysiological characteristics of 5-HT-moduline, a novel endogenousmodulator of 5-HT1B receptor subtype. Ann NY Acad Sci 1998; 861:174-182.
Mattson MP, Chan SL, Duan W. Modification of brain aging and ne-urodegenerative disorders by genes, diet, and behavior. Physiol Rev2002; 82: 637-672.
McCann SM, Franci CR, Favaretto AL, Gutkowska J, Antunes-Rodrigues J. Neuroendocrine regulation of salt and water metabo-lism. Braz J Med Biol Res 1997; 30: 427-441.
McEwen BB. General introduction to vasopressin and oxytocin: struc-ture/metabolism, evolutionary aspects, neural pathway/receptor dis-tribution, and functional aspects relevant to memory processing. AdvPharmacol 2004; 50: 1-50, 655-708.
McKinley MJ, Albiston AL, Allen AM, Mathai ML, May CN, McAl-len RM, Oldfield BJ, Mendelsohn FA, Chai SY. The brain renin-angiotensin system: location and physiological roles. Int J BiochemCell Biol 2003; 35: 901-918
McKinley MJ, Allen AM, Mathai ML, May C, McAllen RM, OldfieldBJ, Weisinger RS. Brain angiotensin and body fluid homeostasis. JpnJ Physiol 2001a; 51: 281-289.
McKinley MJ, Allen AM, May CN, McAllen RM, Oldfield BJ, Sly D,Mendelsohn FA. Neural pathways from the lamina terminalis influen-cing cardiovascular and body fluid homeostasis. Clin Exp PharmacolPhysiol 2001b; 28: 990-992.
McLay RN, Pan W, Kastin AJ. Effects of peptides on animal and humanbehavior: a review of studies published in the first twenty years ofthe journal Peptides. Peptides 2001; 22: 2181-2255.
McLean S. Do substance P and the NK1 receptor have a role in dep-ression and anxiety? Curr Pharm Des 2005; 11: 1529-1547.
Meis S. Nociceptin/orphanin FQ: actions within the brain. Neuroscien-tist 2003; 9: 158-168.

Literatúra 243
Melcangi RC, Panzica GC. Neuroactive steroids: Old players in a newgame. Neuroscience 2006; 138: 733-739.
Meldrum BS. Glutamate as a neurotransmitter in the brain: review ofphysiology and pathology. J Nutr 2000; 130: 1007S-1015S.
Mendez-Otero R, Zaverucha-do-Valle C, Gubert F, Freitas GR, SantiagoMF. Regulation and function of neurogenesis in the adult vertebratebrain. Braz J Med Biol Res 2005; 38: 1553-1559.
Mercer JG, Hoggard N, Williams LM, Lawrence CB, Hannah LT, Mor-gan PJ, Trayhurn P. Coexpression of leptin receptor and prepro-neuropeptide Y mRNA in arcuate nucleus of mouse hypothalamus.J Neuroendocrinol 1996; 8: 733-735.
Meyer-Lindenberg A, Miletich RS, Kohn PD, Esposito G, Carson RE,Quarantelli M, Weinberger DR, Berman KF. Reduced prefrontal ac-tivity predicts exaggerated striatal dopaminergic function in schi-zophrenia. Nat Neurosci 2002; 5: 267-271.
Mieda M, Yanagisawa M. Sleep, feeding, and neuropeptides: roles oforexins and orexin receptors. Curr Opin Neurobiol 2002; 12: 339-345.
Mignot E, Taheri S, Nishino S. Sleeping with the hypothalamus: emer-ging therapeutic targets for sleep disorders. Nat Neurosci 2002; 5:1071-1075.
Millan MJ. Descending control of pain. Prog Neurobiol 2002; 66: 355-474.
Millan MJ. The induction of pain: an integrative review. Prog Neurobiol1999; 57: 1-164.
Miller DB, O’Callaghan JP. Neuroendocrine aspects of the response tostress. Metabolism 2002; 51: 5-10.
Mirescu C, Gould E. Stem Cells in the Adult Brain. In: Handbook ofStem Cells. Vol.2 Adult and Fetal Stem Cells. Weissman IL, MeltonDA, Verfaillie CM, Moore MAS, Blau HM, West MD, Thomas ED,Lanza RP (Eds), Academic Press 2004; pp.219-224.
Misu Y, Goshima Y, Ueda H, Okamura H. Neurobiology of L-DOPAer-gic systems. Prog Neurobiol 1996; 49: 415-454.
Misu Y, Kitahama K, Goshima Y. L-3,4-Dihydroxyphenylalanine asa neurotransmitter candidate in the central nervous system. Phar-macol Ther 2003; 97: 117-137.

244 NEUROTRANSMITERY
Miyamoto S, LaMantia AS, Duncan GE, Sullivan P, Gilmore JH, Lie-berman JA. Recent advances in the neurobiology of schizophrenia.Mol Interv 2003; 3: 27-39.
MKCH 10. Mezinárodní klasifikace nemocí. 10. revize. Duševní poru-chy a poruchy chování. Popisy klinických příznaku a diagnostickávodítka. (Přel. z angl. orig.). Praha, Psychiatrické centrum 1996.282 s. „Zprávyÿ č. 102.; Diagnostic and Statistical Manual of Men-tal Disorders. Fourth edition. Text Revison. Washington, AmericanPsychiatric Association, 2000.
Mollereau C, Roumy M, Zajac JM. Opioid-modulating peptides: me-chanisms of action. Curr Top Med Chem 2005; 5: 341-355.
Moncada S, Higgs A, Furchgott R. International Union of PharmacologyNomenclature in Nitric Oxide Research. Pharmacol Rev 1997; 49:137-142.
Mondal MS, Yamaguchi H, Date Y, Shimbara T, Toshinai K, ShimomuraY, Mori M, Nakazato M. A role for neuropeptide W in the regulationof feeding behavior. Endocrinology 2003; 144: 4729-4733.
Moore KE. Interactions between prolactin and dopaminergic neurons.Biol Reprod 1987; 36: 47-58.
Morgan NG, Chan SL. Imidazoline binding sites in the endocrine panc-reas: can they fulfil their potential as targets for the development ofnew insulin secretagogues? Curr Pharm Des 2001; 7: 1413-1431.
Mori T, Buffo A, Gotz M. The novel roles of glial cells revisited: thecontribution of radial glia and astrocytes to neurogenesis. Curr TopDev Biol 2005; 69: 67-99.
Morris JF, Ludwig M. Magnocellular dendrites: prototypic recei-ver/transmitters. J Neuroendocrinol 2004; 16: 403-408.
Morton RE, Hankinson J, Nicholson J. Botulinum toxin for cerebralpalsy; where are we now? Arch Dis Child 2004; 89: 1133-1137.
Mountjoy KG, Mortrud MT, Low MJ, Simerly RB, Cone RD. Loca-lization of the melanocortin-4 receptor (MC4-R) in neuroendocrineand autonomic control circuits in the brain. Mol Endocrinol 1994; 8:1298-1308.
Mravec B, Kiss A. Katecholamíny mozgu: stručná anatómia a účasťv stresovej reakcii a regulácii kardiovaskulárnej činnosti. Cesk Fysiol2004; 53: 102-116.

Literatúra 245
Mravec B. A new focus on interoceptive properties of adrenal medulla.Auton Neurosci 2005; 120: 10-17.
Myers AC. Transmission in autonomic ganglia. Respir Physiol 2001;125: 99-111.
Myers RD. Neurochemistry of thermoregulation. Physiologist 1984; 27:41-46.
Myslivecek J, Kvetnansky R. The effects of stress on muscarinic recep-tors. Heterologous receptor regulation: yes or no? Auton AutacoidPharmacol 2006; 26: 235-251.
Mysliveček J, Trojan S.. Regulace přenosu signálu spřaženými s G pro-teiny (desenzitizace, fodforylace, down-regulace). Psychiatrie 2002;6: 55-60.
Mysliveček J, Trojan S. Regulace receptorových funkcí v CNS. Čas Lékčes 2003; 149: 338-348.
Mysliveček J. Základy neurověd. Nakladatelství Triton, Praha 2003; 348pp.
Nagai K, Nishio T, Nakagawa H, Nakamura S, Fukuda Y. Effect of bi-lateral lesions of the suprachiasmatic nuclei on the circadian rhythmof food-intake. Brain Res 1978; 142: 384-399.
Narumiya S, Sugimoto Y, Ushikubi F. Prostanoid receptors: structures,properties, and functions. Physiol Rev 1999; 79: 1193-1226.
Nattie E. CO2 brain stem chemoreceptors and breathing. Progr Neuro-biol 1999; 59: 299-331.
Navarra P, Dello Russo C, Mancuso C, Preziosi P, Grossman A. Gaseousneuromodulators in the control of neuroendocrine stress axis. AnnNY Acad Sci 2000; 917: 638-646.
Nemeroff CB, Krishnan KR, Reed D, Leder R, Beam C, Dunnick NR.Adrenal gland enlargement in major depression. A computed tomog-raphic study. Arch Gen Psychiatry 1992; 49: 384-387.
Nemeroff CB, Vale WW. The neurobiology of depression: inroads totreatment and new drug discovery. J Clin Psychiatry 2005; 66: S5-S13.
Nemeroff CB, Widerlov E, Bissette G, Walleus H, Karlsson I, EklundK, Kilts CD, Loosen PT, Vale W. Elevated concentrations of CSFcorticotropin-releasing factor-like immunoreactivity in depressed pa-tients. Science 1984; 226: 1342-1344.

246 NEUROTRANSMITERY
Nemoto T, Konno A, Chiba T. Synaptic contact of neuropeptide-andamine-containing axons on parasympathetic preganglionic neuronsin the superior salivatory nucleus of the rat. Brain Res 1995; 685:33-45.
Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM.Neurobiology of depression. Neuron 2002; 34: 13-25.
Nguyen L, Rigo JM, Rocher V, Belachew S, Malgrange B, Rogister B,Leprince P, Moonen G. Neurotransmitters as early signals for centralnervous system development. Cell Tissue Res 2001; 305: 187-202.
Nicolaidis S. A hormone-based characterization and taxonomy of stress:possible usefullness in management. Metabolism 2002; 51: 31-36.
Nieoullon A. Dopamine and the regulation of cognition and attention.Prog Neurobiol 2002; 67: 53-83.
Niimi M, Murao K. Neuropeptide W as a stress mediator in the hypot-halamus. Endocrine 2005; 27: 51-54.
Nishikimi T, Matsuoka H. Cardiac adrenomedullin: its role in cardiachypertrophy and heart failure. Curr Med Chem Cardiovasc HematolAgents 2005; 3: 231-242.
Noda Y, Mamiya T, Manabe T, Nishi M, Takeshima H, Nabeshima T.Role of nociceptin systems in learning and memory. Peptides 2000;21: 1063-1069.
Norman AW, Mizwicki MT, Norman DP. Steroid-hormone rapid actions,membrane receptors and a conformational ensemble model. Nat RevDrug Discov 2004; 3: 27-41.
North RA. Molecular physiology of P2X receptors. Physiol Rev 2002;82: 1013-1067.
Nozdrachev AD, Fateev MM, Jiminez B, Morales MA. Circuits andprojections of cat stellate ganglion. Arch Med Res 2003; 34: 106-115.
Nurse CA. Neurotransmission and neuromodulation in the chemosen-sory carotid body. Auton Neurosci 2005; 120: 1-9.
Nusbaum MP, Blitz DM, Swensen AM, Wood D, Marder E. The rolesof co-transmission in neural network modulation. Trends Neurosci2001; 24: 146-154.
Nylen A, Skagerberg G, Alm P, Larsson B, Holmqvist B, Andersson KE.Nitric oxide synthase in the hypothalamic paraventricular nucleus of

Literatúra 247
the female rat; organization of spinal projections and coexistencewith oxytocin or vasopressin. Brain Res 2001; 908: 10-24.
Oda Y, Nakanishi I. The distribution of cholinergic neurons in the hu-man central nervous system. Histol Histopathol 2000; 15: 825-834.
Ohtsuki S. New aspects of the blood-brain barrier transporters; its phy-siological roles in the central nervous system. Biol Pharm Bull 2004;27: 1489-1496.
Okada Y, Tsuda Y, Bryant SD, Lazarus LH. Endomorphins and relatedopioid peptides. Vitam Horm 2002; 65: 257-279.
Okuda T, Haga T. High-affinity choline transporter. Neurochem Res2003; 28: 483-488.
Okuda-Ashitaka E, Ito S. Nocistatin: a novel neuropeptide encoded bythe gene for the nociceptin/orphanin FQ precursor. Peptides 2000;21: 1101-1109.
Olszewski PK, Levine AS. Minireview: Characterization of influence ofcentral nociceptin/orphanin FQ on consummatory behavior. Endoc-rinology 2004; 145: 2627-2632.
Oomori Y, Nakaya K, Tanaka H, Iuchi H, Ishikawa K, Satoh Y, Ono K.Immunohistochemical and histochemical evidence for the presence ofnoradrenaline, serotonin and gamma-aminobutyric acid in chief cellsof the mouse carotid body. Cell Tissue Res 1994; 278: 249-254.
Oomura Y, Yoshimatsu H. Neural network of glucose monitoring sys-tem. J Auton Nerv Syst 1984; 10: 359-372.
Osborne RH. Insect neurotransmission: neurotransmitters and their re-ceptors. Pharmacol Ther 1996; 69: 117-142.
Osinski MA, Brown DR. Orphanin FQ/nociceptin: a novel neuromodu-lator of gastrointestinal function? Peptides 2000; 21: 999-1005.
Owens DF, Kriegstein AR. Is there more to GABA than synaptic inhi-bition? Nat Rev Neurosci 2002; 3: 715-727.
Pablo Huidobro-Toro J, Veronica Donoso M. Sympathetic co-transmission: the coordinated action of ATP and noradrenaline andtheir modulation by neuropeptide Y in human vascular neuroeffectorjunctions. Eur J Pharmacol 2004; 500: 27-35.
Pacak K, Armando I, Fukuhara K, Kvetnansky R, Palkovits M, KopinIJ, Goldstein DS. Noradrenergic activation in the paraventricular

248 NEUROTRANSMITERY
nucleus during acute and chronic immobilization stress in rats: an invivo microdialysis study. Brain Res 1992; 589: 91-96.
Pacak K, Baffi JS, Kvetnansky R, Goldstein DS, Palkovits M. Stressor-specific activation of catecholaminergic systems: implications forstress-related hypothalamic-pituitary-adrenocortical responses. AdvPharmacol 1998; 42: 561-564.
Pacak K, McCarty R, Palkovits M, Kopin IJ, Goldstein DS. Effects ofimmobilization on in vivo release of norepinephrine in the bed nucleusof the stria terminalis in conscious rats. Brain Res 1995; 688: 242-246.
Pacak K, Palkovits M, Kopin IJ, Goldstein DS. Stress-induced nore-pinephrine release in the hypothalamic paraventricular nucleus andpituitary-adrenocortical and sympathoadrenal activity: in vivo mic-rodialysis studies. Front Neuroendocrinol 1995; 16: 89-150.
Pacak K, Palkovits M, Kvetnansky R, Kopin IJ, Goldstein DS. Stress-induced norepinephrine release in the paraventricular nucleus of ratswith brain stem hemisections: a microdialysis study. Neuroendocri-nology 1993; 58: 196-201.
Pacak K, Palkovits M, Yadid G, Kvetnansky R, Kopin IJ, Goldstein DS.Heterogeneous neurochemical responses to different stressors: a testof Selye’s doctrine of nonspecificity. Am J Physiol 1998; 275: R1247-R1255.
Pacak K, Palkovits M. Stressor specificity of central neuroendocrineresponses: implications for stress-related disorders. Endocr Rev 2001;22: 502-548.
Pacak K. Stressor-specific activation of the hypothalamic-pituitary-adrenocortical axis. Physiol Res 2000; 49: S11-S17.
Palkovits M, Baffi JS, Pacak K. The role of ascending neuronal pat-hways in stress-induced release of noradrenaline in the hypothalamicparaventricular nucleus of rats. J Neuroendocrinol 1999; 11: 529-539.
Palkovits M, Mezey E, Skirboll LR, Hokfelt T. Adrenergic projectionsfrom the lower brain stem to the hypothalamic paraventricular nuc-leus, the lateral hypothalamic area and the central nucleus of theamygdala in rats. J Chem Neuroanat 1992; 5: 407-415.
Palkovits M. Hypothalamic regulation of food intake. Ideggyogy Sz2003; 56: 288-302.

Literatúra 249
Palkovits M. Interconnections between the neuroendocrine hypothala-mus and the central autonomic system. Geoffrey Harris MemorialLecture, Kitakyushu, Japan, October 1998. Front Neuroendocrinol1999; 20: 270-295.
Palkovits M. Neuropeptide messenger plasticity in the CNS neuronsfollowing axotomy. Mol Neurobiol 1995; 10: 91-103.
Palkovits M. Role of the central nervous system neuropeptides in bodyfluid homeostasis. J Physiol (Paris) 1984; 79: 428-431.
Palkovits M. Stress-induced expression of co-localized neuropeptides inhypothalamic and amygdaloid neurons. Eur J Pharmacol 2000; 405:161-166.
Palmada M, Centelles JJ. Excitatory amino acid neurotransmission.Pathways for metabolism, storage and reuptake of glutamate inbrain. Front Biosci 1998; 3: 701-718.
Palmer AM, DeKosky ST.Monoamine neurons in aging and Alzheimer’sdisease. J Neural Transm Gen Sect 1993; 91: 135-159.
Pan W, Kastin AJ, Banks WA, Zadina JE. Effects of peptides: a cross-listing of peptides and their central actions published in the journalPeptides from 1994 through 1998. Peptides 1999; 20: 1127-1138.
Panula P, Aarnisalo AA, Wasowicz K. Neuropeptide FF, a mammalianneuropeptide with multiple functions. Prog Neurobiol 1996; 48: 461-487.
Panula P, Kalso E, Nieminen M, Kontinen VK, Brandt A, PertovaaraA. Neuropeptide FF and modulation of pain. Brain Res 1999; 848:191-196.
Paquin J, Danalache BA, Jankowski M, McCann SM, Gutkowska J.Oxytocin induces differentiation of P19 embryonic stem cells to car-diomyocytes. Proc Natl Acad Sci USA 2002; 99: 9550-9555.
Parascandola J, Appel TA, Gilbert Daniel L. A century of Americanphysiology: based on an exhibit commemorating the centennial of theAmerican Physiological Society at the National Library of Medicine.1987; pp. 1-21.
Pariante CM, Thomas SA, Lovestone S, Makoff A, Kerwin RW. Do an-tidepressants regulate how cortisol affects the brain? Psychoneuro-endocrinology 2004; 29: 423-447.

250 NEUROTRANSMITERY
Parsons ME, Ganellin CR. Histamine and its receptors. Br J Pharmacol2006; 147: S127-S135.
Passani MB, Bacciottini L, Mannaioni PF, Blandina P. Central his-taminergic system and cognition. Neurosci Biobehav Rev 2000; 24:107-113.
Paterson D, Nordberg A. Neuronal nicotinic receptors in the humanbrain. Prog Neurobiol 2000; 61: 75-111.
Paton JF, Kasparov S, Paterson DJ. Nitric oxide and autonomic controlof heart rate: a question of specificity. Trends Neurosci 2002; 25: 626-631.
Pavlov VA, Wang H, Czura CJ, Friedman SG, Tracey KJ. The choliner-gic anti-inflammatory pathway: a missing link in neuroimmunomo-dulation. Mol Med 2003; 9: 125-134.
Pečeňák J a spol. Kapitoly o schizofrénii. Osveta, Martin, 2005; 196 pp.
Pedrazzini T, Pralong F, Grouzmann E. Neuropeptide Y: the universalsoldier. Cell Mol Life Sci 2003; 60: 350-377.
Pennefather JN, Lecci A, Candenas ML, Patak E, Pinto FM, MaggiCA. Tachykinins and tachykinin receptors: a growing family. Life Sci2004; 74: 1445-1463.
Perez-Cerda F, Martinez-Millan L, Matute C. Anatomical evidence forglutamate and/or aspartate as neurotransmitters in the geniculo-,claustro-, and cortico-cortical pathways to the cat striate cortex.J Comp Neurol 1996; 373: 422-432.
Perlman WR, Weickert CS, Akil M, Kleinman JE. Postmortem investi-gations of the pathophysiology of schizophrenia: the role of suscep-tibility genes. J Psychiatry Neurosci 2004; 29: 287-293.
Petrovický P. Anatomie s topografií a klinickými aplikacemi. III svazek.Neuroanatomie, smyslová ústrojí a kůže. Osveta, Martin, 2002; 542pp.
Phillis JW. Adenosine and adenine nucleotides as regulators of cerebralblood flow: roles of acidosis, cell swelling, and KATP channels. CritRev Neurobiol 2004; 16: 237-270
Piet R, Vargova L, Sykova E, Poulain DA, Oliet SH. Physiological con-tribution of the astrocytic environment of neurons to intersynapticcrosstalk. Proc Natl Acad Sci USA 2004; 101: 2151-2155.

Literatúra 251
Pilowsky PM, Llewellyn-Smith IJ, Minson JB, Arnolda LF, ChalmersJP. Substance P and serotonergic inputs to sympathetic pregangli-onic neurons. Clin Exp Hypertens 1995; 17: 335-344.
Pin JP, Duvoisin R. The metabotropic glutamate receptors: structureand functions. Neuropharmacology 1995; 34: 1-26.
Pirker S, Czech T, Baumgartner C, Maier H, Novak K, Furtinger S,Fischer-Colbrie R, Sperk G. Chromogranins as markers of alteredhippocampal circuitry in temporal lobe epilepsy. Ann Neurol 2001;50: 216-226.
Powrozek TA, Sari Y, Singh RP, Zhou FC. Neurotransmitters and Sub-stances of Abuse: Effects on Adult Neurogenesis. Curr Neurovasc Res2004; 1: 251-260.
Prell GD, Martinelli GP, Holstein GR, Matulic-Adamic J, WatanabeKA, Chan SL, Morgan NG, Haxhiu MA, Ernsberger P. Imidazoleace-tic acid-ribotide: an endogenous ligand that stimulates imidazol(in)ereceptors. Proc Natl Acad Sci USA 2004; 101: 13677-13682.
Purves D. Complex brain functions. In: Purves D, Augustine GJ, Fitz-patrick D, Hall WC, Lamantia AS, McNamara JO, Williams SM(Eds). Neuroscience. Sunderland; Sinauer Associated, 2004; 773 pp.
Qu D, Ludwig DS, Gammeltoft S, Piper M, Pelleymounter MA, CullenMJ, Mathes WF, Przypek R, Kanarek R, Maratos-Flier E. A role formelanin-concentrating hormone in the central regulation of feedingbehaviour. Nature 1996; 380: 243-247.
Raffin-Sanson ML, de Keyzer Y, Bertagna X. Proopiomelanocortin,a polypeptide precursor with multiple functions: from physiology topathological conditions. Eur J Endocrinol 2003; 149: 79-90.
Raisman G. An urge to explain the incomprehensible: Geoffrey Harrisand the Discovery of the Neural Control of the Pituitary Gland.Annu Rev Neurosci 1997; 20: 533-566.
Ralevic V, Burnstock G. Receptors for purines and pyrimidines. Phar-macol Rev 1998; 50: 413-492.
Ramachandran VS (Ed). Encyclopedia of the Human Brain. AcademicPress, Vol 3, 2002; 2616 pp.
Ramos EJ, Meguid MM, Campos AC, Coelho JC. Neuropeptide Y,alpha-melanocyte-stimulating hormone, and monoamines in food in-take regulation. Nutrition 2005; 21: 269-279.

252 NEUROTRANSMITERY
Rampon C, Luppi PH, Fort P, Peyron C, Jouvet M. Distribution ofglycine-immunoreactive cell bodies and fibers in the rat brain. Ne-uroscience 1996; 75: 737-755.
Randall DC, Brown DR, McGuirt AS et al. Interactions within the in-trinsic cardiac nervous system contribute to chronotropic regulation.Am J Physiol Regul Integr Comp Physiol 2003; 285: 1066-1075.
Randall DC. Towards an understanding of the function of the intrinsiccardiac ganglia. J Physiol 2000; 528: 406.
Rapport MM, Green AA, Page IH. Serum Vasoconstrictor (Serotonin)IV. Isolation and Characterization. J Biol Chem 1948; 176: 1243-1251.
Raybould HE. Visceral perception: sensory transduction in visceral af-ferents and nutrients. Gut 2002; 51: i11-i14.
Raz N. Ageing and the brain. In Encyclopedia of Life Sciences. London,UK: Macmillan Reference Ltd. 2001; pp. 1-6.
Reaux A, De Mota N, Skultetyova I, Lenkei Z, El Messari S, Gallatz K,Corvol P, Palkovits M, Llorens-Cortes C. Physiological role of a novelneuropeptide, apelin, and its receptor in the rat brain. J Neurochem2001; 77: 1085-1096.
Rehfeld JF. The new biology of gastrointestinal hormones. Physiol Rev1998; 78: 1087-1108.
Rehman HU, Masson EA. Neuroendocrinology of ageing. Age Ageing2001; 30: 279-287.
Reif A, Fritzen S, Finger M, Strobel A, Lauer M, Schmitt A, Lesch KP.Neural stem cell proliferation is decreased in schizophrenia, but notin depression. Mol Psychiatry 2006; 11: 514-522.
Reichlin S. Neuroendocrine significance of vasoactive intestinal poly-peptide. 1988; 527: 431-449.
Reinis S, Holub RF, Smrz P. Kvantová hypotéza činnosti mozkua vědomí. Cesk Fysiol 2005; 54: 26-31.
Reinscheid RK, Xu YL. Neuropeptide S and its receptor: a newly de-orphanized G protein-coupled receptor system. Neuroscientist 2005a;11: 532-538.
Reinscheid RK, Xu YL. Neuropeptide S as a novel arousal promotingpeptide transmitter. FEBS J 2005b; 272: 5689-5693.

Literatúra 253
Reis DJ, Regunathan S. Is agmatine a novel neurotransmitter in brain?Trends Pharmacol Sci 2000; 21: 187-193.
Reisine T. Opiate receptors. Neuropharmacology 1995; 34: 463-472.
Ressler KJ, Nemeroff CB. Peptides and Psychiatry. In: InternationalEncyclopedia of the Social & Behavioral Sciences. Smelser NJ, BaltesPB (Eds.); 2001; pp. 11189-11195.
Ressler KJ, Nemeroff CB. Role of norepinephrine in the pathophysiologyand treatment of mood disorders. Biol Psychiatry 1999; 46: 1219-1233.
Ressler KJ, Nemeroff CB. Role of serotonergic and noradrenergic sys-tems in the pathophysiology of depression and anxiety disorders.Depress Anxiety 2000; 12: 2-19.
Reul JM, Holsboer F. Corticotropin-releasing factor receptors 1 and 2in anxiety and depression. Curr Opin Pharmacol 2002; 2: 23-33.
Ribak CE, Roberts RC. GABAergic synapses in the brain identifiedwith antisera to GABA and its synthesizing enzyme, glutamate de-carboxylase. J Electron Microsc Tech 1990; 15: 34-48.
Richard D, Baraboi D. Circuitries involved in the control of energyhomeostasis and the hypothalamic-pituitary-adrenal axis activity.Treat Endocrinol 2004; 3: 269-277.
Richards AM, Charles C. Urotensin II in the cardiovascular system.Peptides 2004; 25: 1795-1802.
Richmond CA. The role of arginine vasopressin in thermoregulationduring fever. J Neurosci Nurs 2003; 35: 281-286.
Ritter S, Dinh TT, Zhang Y. Localization of hindbrain glucoreceptivesites controling food intake and blood glucose. Brain Res 2000; 856:37-47.
Riva C, Eggli P, Felix D, Mosimann R, Imboden H. Hypothalamic ac-cessory nuclei and their relation to the angiotensinergic and vasop-ressinergic systems. Regul Pept 1999; 83: 129-133.
Rizzoli SO, Betz WJ. Synaptic vesicle pools. Nat Rev Neurosci 2005; 6:57-69.
Rodriguez de Fonseca F, Del Arco I, Bermudez-Silva FJ, Bilbao A, Cip-pitelli A, Navarro M. The endocannabinoid system: physiology andpharmacology. Alcohol Alcohol 2005; 40: 2-14.

254 NEUROTRANSMITERY
Roe DL. The discovery of dopamine’s physiological importance. BrainRes Bull 1999; 50: 375-376.
Rorig B, Sutor B. Regulation of gap junction coupling in the developingneocortex. Mol Neurobiol 1996; 12: 225-249.
Rosicka M, Krsek M, Jarkovska Z, Marek J, Schreiber V. Ghrelin -a new endogenous growth hormone secretagogue. Physiol Res 2002;51: 435-441.
Rossi M, KimMS, Morgan DG, Small CJ, Edwards CM, Sunter D, Abus-nana S, Goldstone AP, Russell SH, Stanley SA, Smith DM, YagaloffK, Ghatei MA, Bloom SR. A C-terminal fragment of agouti-relatedprotein increases feeding and antagonizes the effect of á-melanocytestimulating hormone in vivo. Endocrinology 1998; 139: 4428-4431.
Rostene WH, Alexander MJ. Neurotensin and neuroendocrine regula-tion. Front Neuroendocrinol 1997; 18: 115-173.
Roth J, Růžička E. Nemoci extrapyramidového systému. Stručný kli-nický přehled. Sanofi, Praha 1998; pp. 1-57.
Roth KA, Mefford IM, Barchas JD. Epinephrine, norepinephrine, do-pamine and serotonin: differential effects of acute and chronic stresson regional brain amines. Brain Res 1982; 239: 417-424.
Roumy M, Zajac JM. Neuropeptide FF, pain and analgesia. Eur J Phar-macol 1998; 345: 1-11.
Rowland NE, Morien A, Li BH. The physiology and brain mechanismsof feeding. Nutrition 1996; 12: 626-639.
Rubin LL, Staddon JM. The cell biology of the blood-brain barrier.Annu Rev Neurosci 1999; 22: 11-28.
Rudomin P. Presynaptic selection of afferent inflow in the spinal cord.J Physiol Paris 1999; 93: 329-347.
Rupprecht R, Holsboer F. Neuroactive steroids: mechanisms of ac-tion and neuropsychopharmacological perspectives. Trends Neurosci1999; 22: 410-416.æ
Sahu A, Dube MG, Phelbs CP, Sninski CA, Kalra PS, Kalra SP. Insulinand insulin-like growth factor II suppress neuropeptide Y releasefrom the nerve terminals in the paraventricular nucleus: a putativehypothalamic site for energy homeostasis. Endocrinology 1995; 136:5718-5724.

Literatúra 255
Sahu A, Sninski CA,Phelbs CP, Dube MG, Kalra PS, Kalra SP. Neuro-peptide Y release from the paraventricular nucleus increases in as-sociation with hyperphagia in streptozotocin-induced diabetic rats.Endocrinology 1992; 131: 2979-2985.
Sainsbury A, Cooney GJ, Herzog H. Hypothalamic regulation of energyhomeostasis. Best Pract Res Clin Endocrinol Metab 2002; 16: 623-637.
Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, TanakaH, Williams SC, Richarson JA, Kozlowski GP, Wilson S, Arch JR,Buckingham RE, Haynes AC, Carr SA, Annan RS, McNulty DE, LiuWS, Terrett JA, Elshourbagy NA, Bergsma DJ, Yanagisawa M. Ore-xins and orexin receptors: a family of hypothalamic neuropeptidesand G protein-coupled receptors that regulate feeding behavior. Cell1998; 92: 1 page following 696.
Samson WK, Taylor MM, Ferguson AV. Non-sleep effects of hypocre-tin/orexin. Sleep Med Rev 2005; 9: 243-252.
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influ-ence stress responses? Integrating permissive, suppressive, stimula-tory, and preparative actions. Endocr Rev 2000; 21: 55-89.
Sapru HN. Glutamate circuits in selected medullo-spinal areas regula-ting cardiovascular function. Clin Exp Pharmacol Physiol 2002; 29:491-496.
Sarter M, Parikh V. Choline transporters, cholinergic transmission andcognition. Nat Rev Neurosci 2005; 6: 48-56.
Sawa A, Snyder SH. Schizophrenia: diverse approaches to a complexdisease. Science 2002; 296: 692-695.
Sawa A, Snyder SH. Schizophrenia: neural mechanisms for novel thera-pies. Mol Med 2003; 9: 3-9.
Seeley RJ, Drazen DL, Clegg DJ. The critical role of the melanocortinsystem in the control of energy balance. Annu Rev Nutr 2004; 24:133-149.
Segovia G, Porras A, Del Arco A, Mora F. Glutamatergic neurotrans-mission in aging: a critical perspective. Mech Ageing Dev 2001; 122:1-29.
Selye H. A syndrome produced by diverse nocuous agents. Nature 1936;138: 32.

256 NEUROTRANSMITERY
Selye H. Stress. Canada Acta, Montreal, 1950.
Selye, H. Stress without Distress. New York: Lippincott & Crowell Pub-lishers; 1974.
Serrano J, Alonso D, Fernandez AP, Encinas JM, Lopez JC, Castro-Blanco S, Fernandez-Vizarra P, Richart A, Santacana M, UttenthalLO, Bentura ML, Martinez-Murillo R, Martinez A, Cuttitta F, Rod-rigo J. Adrenomedullin in the central nervous system. Microsc ResTech 2002; 57: 76-90.
Seutin V. Dopaminergic neurones: much more than dopamine?Br J Pharmacol. 2005; 146: 167-169.
Severini C, Improta G, Falconieri-Erspamer G, Salvadori S, ErspamerV. The tachykinin peptide family. Pharmacol Rev 2002; 54: 285-322.
Seyedabadi M, Li Q, Padley JR, Pilowsky PM, Goodchild AK. A novelpressor area at the medullo-cervical junction that is not dependenton the RVLM: efferent pathways and chemical mediators. J Neurosci2006; 26: 5420-5427.
Shan J, Stachniak T, Jhamandas JH, Krukoff TL. Autonomic and ne-uroendocrine actions of adrenomedullin in the brain: mechanisms forhomeostasis. Regul Pept 2003; 112: 33-40.
Shefchyk SJ. Sacral spinal interneurones and the control of urinary blad-der and urethral striated sphincter muscle function. J Physiol 2001;533: 57-63.
Shi Q, Savage JE, Hufeisen SJ, Rauser L, Grajkowska E, Ernsberger P,Wroblewski JT, Nadeau JH, Roth BL. L-homocysteine sulfinic acidand other acidic homocysteine derivatives are potent and selectivemetabotropic glutamate receptor agonists. J Pharmacol Exp Ther2003; 305: 131-142.
Shimomura Y, Harada M, Goto M, Sugo T, Matsumoto Y, Abe M,Watanabe T, Asami T, Kitada C, Mori M, Onda H, Fujino M. Iden-tification of neuropeptide W as the endogenous ligand for orphanG-protein-coupled receptors GPR7 and GPR8. J Biol Chem 2002;277: 35826-35832.
Shirasaka T, Kunitake T, Takasaki M, Kannan H. Neuronal effects oforexins: relevant to sympathetic and cardiovascular functions. RegulPept 2002; 104: 91-95.

Literatúra 257
Schemann M, Neunlist M. The human enteric nervous system. Neuro-gastroenterol Motil 2004; 16: 55-59.
Schreiber V. 12. Fyziológia žliaz s vnútornou sekréciou. In: Fyziológia2. Trojan S a spol. (Eds). Osveta, Martin, 1992; pp. 429-469.
Schreihofer AM, Stornetta RL, Guynet PG. Regulation of sympathe-tic tone and arterial pressure by rostral ventrolateral medulla afterdepletion of C1 cells in rat. J Physiol 2000; 529: 221-236.
Schumacher M, Akwa Y, Guennoun R, Robert F, Labombarda F, Desar-naud F, Robel P, De Nicola AF, Baulieu EE. Steroid synthesis andmetabolism in the nervous system: trophic and protective effects.J Neurocytol 2000; 29: 307-326.
Schwartz JC, Barbin G, Garbarg BM, Pollard H, Rose C, VerdiereM. Neurochemical evidence for histamine acting as a transmitterin mammalian brain. Adv Biochem Psychopharmacol 1976; 15: 111-126.
Schwartz JC. Histamine as a transmitter in brain. Life Sci 1975; 17:503-517.
Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG. Centralnervous system control of food intake. Nature 2000; 404: 661-671.
Siegel GJ, Albers RW, Brady ST, Price DL (Eds). Basic neurochemistry:Molecular, cellular, and medical aspects. Elsevier Academic Press,San Diego, 2006; 1016 pp.
Siever LJ, Davis KL. Overview: toward a dysregulation hypothesis ofdepression. Am J Psychiatry 1985; 142: 1017-1031.
Singer W. The brain as a self-organizing system. Eur Arch PsychiatryNeurol Sci 1986; 236: 4-9.
Singh S, Johnson PI, Javed A, Gray TS, Lonchyna VA, Wurster RD.Monoamine- and histamine-synthesizing enzymes and neurotrans-mitters within neurons of adult human cardiac ganglia. Circulation1999; 99: 411-419.
Skelton KH, Owens MJ, Nemeroff CB. The neurobiology of urocortin.Regul Pept 2000; 93: 85-92.
Skultetyova I, Tokarev D, Jezova D. Stress-induced increase in blood-brain barrier permeability in control and monosodium glutamate-treated rats. Brain Res Bull 1998; 45: 175-178.

258 NEUROTRANSMITERY
Slavíková J, Kuncová J, Chottová-Dvořáková M, Reischig J. Neadre-nergní necholinergní nervová regulace srdeční činnosti: morfologiea funkce peptidergní inervace srdce savců. Ces Fysiol 2005; 54: 70-77.
Slavikova J, Kuncova J, Reisching J, Dvorakova M. Catecholaminergicneurons in the rat intrinsic cardiac nervous system. Neurochem Res2003; 28: 593-598.
Slavikova J. Distribution of peptide-containing neurons in the develo-ping rat right atrium, studied using immunofluorescence and confocallaser scanning. Neurochem Res 1997; 22: 1013-1021.
Smeets WJAJ, González A. Catecholamine systems in the brain of ver-tebrates: new prospectives through a comparative approach. BrainRes Rev 2000; 33: 308-379.
Smith BK, York DK, Bray GA. Activation of hypothalamic serotoninreceptors reduced intake of dietary fat and protein but not carbo-hydrate. Am J Physiol 1999; 277: R802-R811.
Smith MA, Makino S, Kvetnansky R, Post RM. Stress and glucocorti-coids affect the expression of brain-derived neurotrophic factor andneurotrophin-3 mRNAs in the hippocampus. J Neurosci 1995; 15:1768-1777.
Smits KM, Smits LJ, Schouten JS, Stelma FF, Nelemans P, Prins MH.Influence of SERTPR and STin2 in the serotonin transporter geneon the effect of selective serotonin reuptake inhibitors in depression:a systematic review. Mol Psychiatry 2004; 9: 433-441.
Snead OC 3rd, Gibson KM.Gamma-hydroxybutyric acid. N Engl J Med2005; 352: 2721-2732.
Snead OC 3rd. Evidence for a G protein-coupled gamma-hydroxybu-tyric acid receptor. J Neurochem 2000; 75: 1986-1996.
Snijdelaar DG, Dirksen R, Slappendel R, Crul BJ. Substance P. EurJ Pain 2000; 4: 121-135.
Snyder SH, Baranano DE. Heme oxygenase: a font of multiple messen-gers. Neuropsychopharm 2001; 25: 294-298.
Snyder SH, Ferris CD.Novel neurotransmitters and their neuropsychiat-ric relevance. Am J Psychiatry 2000; 157: 1738-1751.

Literatúra 259
Snyder SH, Jaffrey SR, Zakhary R. Nitric oxide and carbon monoxide:parallel roles as neural messengers. Brain Res Brain Res Rev 1998;26: 167-175.
Snyder SH, Kim PM. D-amino acids as putative neurotransmitters: fo-cus on D-serine. Neurochem Res 2000; 25: 553-560.
Solomon A, De Fanti BA, Martinez JA. Peripheral ghrelin participatesin glucostatic feeding mechanisms and in the anorexigenic signallingmediated by CART and CRF neurons. Nutr Neurosci 2005; 8: 287-295.
Sotelo C. Viewing the brain through the master hand of Ramon y Cajal.Nat Rev Neurosci 2003; 4: 71-77.
Spitzer NC, Borodinsky LN, Root CM. Homeostatic activity-dependentparadigm for neurotransmitter specification. Cell Calcium 2005; 37:417-423.
Spyer KM, Dale N, Gourine AV. ATP is a key mediator of central andperipheral chemosensory transduction. Exp Physiol 2004; 89: 53-59.
Squire LR, Bloom FE, McConnell SK, Roberts JL, Spitzer NC, ZigmondMJ (Eds). Fundamental Neuroscience. Academic Press, San Diego,2003; 1426 pp.
Stahl SM. Essential Psychopharmacology: Neuroscientific Basis andPractical Applications. Second edition. Stahl SM (Ed). CambridgeUniversity Press, New York 2000; 601 pp.
Stanford SC. Central noradrenergic neurones and stress. PharmacolTher 1995; 68: 297-342.
Starowicz K, Przewlocka B. The role of melanocortins and their recep-tors in inflammatory processes, nerve regeneration and nociception.Life Sci 2003; 73: 823-847.
Steece-Collier K, Maries E, Kordower JH. Etiology of Parkinson’s dise-ase: Genetics and environment revisited. Proc Natl Acad Sci USA2002; 99: 13972-13974.
Steele PA, Gibbins IL, Morris JL, Mayer B. Multiple populations ofneuropeptide-containing intrinsic neurons in the guinea-pig heart.Neuroscience 1994; 62: 241-250.
Steiner H, Gerfen CR. Role of dynorphin and enkephalin in the regula-tion of striatal output pathways and behavior. Exp Brain Res 1998;123: 60-76.

260 NEUROTRANSMITERY
Sternberg EM. Neural-immune interactions in health and disease. J ClinInvest 1997; 100: 2641-2647.
Stornetta RL, McQuiston TJ, Guyenet PG. GABAergic and glyciner-gic presympathetic neurons of rat medulla oblongata identified byretrograde transport of pseudorabies virus and in situ hybridization.J Comp Neurol 2004; 479: 257-270.
Strack AM, Sawyer WB, Hughes JH, Platt KB, Loewy AD. A generalpattern of CNS innervation of the sympathetic outflow demonstratedby transneuronal pseudorabies viral infections. Brain Res 1989a; 491:156-162.
Strack AM, Sawyer WB, Platt KB, Loewy AD. CNS cell groups re-gulating the sympathetic outflow to adrenal gland as revealed bytransneuronal cell body labeling with pseudorabies virus. Brain Res1989b; 491: 274-296.
Strand FL. David and Goliath - the slingshot that started the neuro-peptide revolution. Eur J Pharmacol 2000; 405: 3-12.
Strong R. Neurochemical changes in the aging human brain: implica-tions for behavioral impairment and neurodegenerative disease. Ge-riatrics 1998; 53: S9-S12.
Sugawara T, Fujimura M, Noshita N, Kim GW, Saito A, Hayashi T, Na-rasimhan P, Maier CM, Chan PH. Neuronal death/survival signalingpathways in cerebral ischemia. NeuroRx 2004; 1: 17-25.
Sun B, Fujiwara K, Adachi S, Inoue K. Physiological roles of prolactin-releasing peptide. Regul Pept 2005; 126: 27-33.
Sun MK. Central neural organization and control of sympathetic ner-vous system in mammals. Prog Neurobiol 1995; 47: 157-233.
Sved AF, Cano G, Card JP. Neuroanatomical specificity of the circuitscontrolling sympathetic outflow to different targets. Clin Exp Phar-macol Physiol 2001; 28: 115-119.
Swaab DF. Neuropeptides in hypothalamic neuronal disorders. Int RevCytol 2004; 240: 305-375.
Swanson LW, Sawchenko PE, Lind RW. Regulation of multiple peptidesin CRF parvocellular neurosecretory neurons: implications for thestress response. Prog Brain Res 1986; 68: 169-190.

Literatúra 261
Swanson LW, Sawchenko PE. Paraventricular nucleus: a site for theintegration of neuroendocrine and autonomic mechanisms. Neuroen-docrinology 1980; 31: 410-417.
Sykova E. Extrasynaptic volume transmission and diffusion parametersof the extracellular space. Neuroscience 2004; 129: 861-876.
Sykova E. Glia and volume transmission during physiological and pat-hological states. J Neural Transm 2005; 112: 137-147.
Szelenyi Z. Cholecystokinin and thermoregulation-a minireview. Pepti-des 2001; 22: 1245-1250.
Takahashi A, Ishimaru H, Ikarashi Y, Kishi E, Maruyama Y. Oppositeregulation of body temperature by cholinergic input to the paraven-tricular nucleus and supraoptic nucleus in rats. Brain Res 2001; 909:102-111.
Tandon R. Cholinergic aspects of schizophrenia. Br J Psychiatry Suppl1999; 37: 7-11.
Tang-Christensen M, Vrang N, Larsen PJ. Glucagon-like peptide con-taining pathways in the regulation of feeding behaviour. Int J ObesRelat Metab Disord 2001; 25: S42-S47.
Tatro JB, Sinha PS. The central melanocortin system and fever. AnnNY Acad Sci 2003; 994: 246-257.
Taylor C, Fricker AD, Devi LA, Gomes I. Mechanisms of action of an-tidepressants: from neurotransmitter systems to signaling pathways.Cell Signal 2005; 17: 549-557.
Taylor MM, Samson WK. Adrenomedullin and the integrative physi-ology of fluid and electrolyte balance. Microsc Res Tech 2002; 57:105-109.
Teclemariam-Mesbah R, Kalsbeek A, Buijs RM, Pevet P. Oxytocin in-nervation of spinal preganglionic neurons projecting to the superiorcervical ganglion in the rat. Cell Tissue Res 1997; 287: 481-486.
Terry AV Jr, Buccafusco JJ. The cholinergic hypothesis of age and Alz-heimer’s disease-related cognitive deficits: recent challenges and theirimplications for novel drug development. J Pharmacol Exp Ther2003; 306: 821-827.
Thompson GA, Kilpatrick IC. The neurotransmitter candidature of sul-phur-containing excitatory amino acids in the mammalian centralnervous system. Pharmacol Ther 1996; 72: 25-36.

262 NEUROTRANSMITERY
Thompson RH, Swanson LW. Organization of inputs to the dorsomedialnucleus of the hypothalamus: reexamination with Fluorogold andPHAL in the rat. Brain Res Brain Res Rev 1998; 27: 89-118.
Toda N, Ayajiki K, Okamura T. Nitric oxide and penile erectile function.Pharmacol Ther 2005; 106: 233-266.
Toda N, Herman AG. Gastrointestinal function regulation by nitrergicefferent nerves. Pharmacol Rev 2005; 57: 315-338.
Toda N, Okamura T. The pharmacology of nitric oxide in the peripheralnervous system of blood vessels. Pharmacol Rev 2003; 55: 271-324.
Tormasiova M, Feketeova E. Narkolepsia - novšie poznatky v patoge-néze, diagnostika a liečba. Medicínsky Monitor, 2005, 2: 13-17.
Tracey KJ. The inflammatory reflex. Nature 2002; 420: 853-859.
Trudeau LE. Glutamate co-transmission as an emerging concept in mo-noamine neuron function. J Psychiatry Neurosci 2004; 29: 296-310.
Tsai G, Coyle JT. Glutamatergic mechanisms in schizophrenia. AnnuRev Pharmacol Toxicol 2002; 42: 165-179.
Tsai G, Coyle JT. The role of glutamatergic neurotransmission in thepathophysiology of alcoholism. Annu Rev Med 1998; 49: 173-184.
Tsigos C, Chrousos GP. Hypothalamic-pituitary-adrenal axis, neuroen-docrine factors and stress. J Psychosom Res 2002; 53: 865-871.
Tsumoto T. Excitatory amino acid transmitters and their receptors inneural circuits of the cerebral neocortex. Neurosci Res 1990; 9: 79-102.
Twarog BM, Page IH. Serotonin content of some mammalian tissuesand urine and a method for its determination. Am J Physiol 1953;175: 157-161.
Valenstein ES. The discovery of chemical neurotransmitters. Brain Cogn2002; 49: 73-95.
Valentino RJ, Commons KG. Peptides that fine-tune the serotonin sys-tem. Neuropeptides 2005; 39: 1-8.
Van Dijk G, Thiele TE, Donahey JC, Campfield LA, Smith FJ, Burn P,Bernstein IL, Woods SC, Seeley RJ. Central infusions of leptin andGLP-1-(7-36) amide differentially stimulate c-FLI in the rat brain.Am J Physiol 1996; 271: R1096-R1100.

Literatúra 263
Vanitallie TB. Stress: a risk factor for serious illness. Metabolism 2002;51: 40-45.
Vermetten E, Bremner JD. Circuits and systems in stress. I. Preclinicalstudies. Depress Anxiety 2002a; 15: 126-147.
Vermetten E, Bremner JD. Circuits and systems in stress. II. Applicati-ons to neurobiology and treatment in posttraumatic stress disorder.Depress Anxiety 2002b; 16: 14-38.
Verrier RL, Antzelevitch C. Autonomic aspects of arrhythmogenesis:the enduring and the new. Curr Opin Cardiol 2004; 19: 2-11.
Vezzani A, Hoyer D. Brain somatostatin: a candidate inhibitory role inseizures and epileptogenesis. Eur J Neurosci 1999; 11: 3767-3776.
Vigaš M. Stres. In: Neuroendokrinná reakcia v strese u človeka. VigašM (Ed). Veda, Bratislava, 1985; pp. 13-21.
Vilim FS, Aarnisalo AA, Nieminen ML, Lintunen M, Karlstedt K, Kon-tinen VK, Kalso E, States B, Panula P, Ziff E. Gene for pain mo-dulatory neuropeptide NPFF: induction in spinal cord by noxiousstimuli. Mol Pharmacol 1999; 55: 804-811.
Vizi ES, Kiss JP, Lendvai B. Nonsynaptic communication in the centralnervous system. Neurochem Int 2004; 45: 443-451.
Vizi ES. Role of high-affinity receptors and membrane transporters innonsynaptic communication and drug action in the central nervoussystem. Pharmacol Rev 2000; 52: 63-89.
Volpicelli LA, Levey AI. Muscarinic acetylcholine receptor subtypes incerebral cortex and hippocampus. Prog Brain Res 2004; 145: 59-66.
von Euler US. A specific sympathomimetic ergone in adrenergic nervefibres (sympathin) and its relation to adrenaline and nor-adrenaline.Acta Physiol Scand 1946; 12: 73-97.
Vrang N, Larsen PJ, Clausen JT, Kristensen P. Neurochemical characte-rization of hypothalamic cocaine- amphetamine-regulated transcriptneurons. J Neurosci 1999; 19: RC5.
Vrang N, Larsen PJ, Kristensen P. Cocaine-amphetamine regulatedtranscript (CART) expression is not regulated by amphetamine.Neuroreport 2002; 13: 1215-1218.
Wallukat G. The beta-adrenergic receptors. Herz 2002; 27: 683-690.

264 NEUROTRANSMITERY
Wang J, Irnaten M, Neff RA, Venkatesan P, Evans C, Loewy AD, Met-tenleiter TC, Mendelowitz D. Synaptic and neurotransmitter activa-tion of cardiac vagal neurons in the nucleus ambiguus. Ann NY AcadSci 2001; 940: 237-246.
Wang R. Two’s company, three’s a crowd: can H2S be the third endo-genous gaseous transmitter? FASEB J 2002; 16: 1792-1798.
Waschek JA. Multiple actions of pituitary adenylyl cyclase activatingpeptide in nervous system development and regeneration. Dev Ne-urosci 2002; 24: 14-23.
Watkins JC. l-glutamate as a central neurotransmitter: looking back.Biochem Soc Trans 2000; 28: 297-309.
Webster RA (Ed). Neurotransmitters, drugs and brain function. JohnWiley & Sons, Ltd, Chichester, 2001; 534 pp.
Weihe E, Schutz B, Hartschuh W, Anlauf M, Schafer MK, Eiden LE.Coexpression of cholinergic and noradrenergic phenotypes in humanand nonhuman autonomic nervous system. J Comp Neurol 2005; 492:370-379.
Weinberg RJ. Glutamate: an excitatory neurotransmitter in the mam-malian CNS. Brain Res Bull 1999; 50: 353-354.
Wess J, Duttaroy A, Gomeza J, Zhang W, Yamada M, Felder CC, Ber-nardini N, Reeh PW. Muscarinic receptor subtypes mediating cen-tral and peripheral antinociception studied with muscarinic receptorknockout mice: a review. Life Sci 2003; 72: 2047-2054.
Wess J. Muscarinic acetylcholine receptor knockout mice: novel pheno-types and clinical implications. Annu Rev Pharmacol Toxicol 2004;44: 423-450.
Westfall DP, Todorov LD, Mihaylova-Todorova ST. ATP as a cotrans-mitter in sympathetic nerves and its inactivation by releasable enzy-mes. J Pharmacol Exp Ther 2002; 303: 439-444.
Whitfield JF. The neuronal primary cilium - an extrasynaptic signalingdevice. Cell Signal 2004; 16: 763-767.
Wickelgren I. Society for Neuroscience meeting. CO gas joins brain sig-naling team. Science 2003; 302: 1320-1323.
Widdicombe JG. Autonomic regulation. i-NANC/e-NANC. Am J Res-pir Crit Care Med 1998; 158: S171-S175.

Literatúra 265
Wiedermann CJ. Secretoneurin: a functional neuropeptide in health anddisease. Peptides 2000; 21: 1289-1298.
Wilding JP. Neuropeptides and appetite control. Diabet Med 2002; 19:619-627.
William JM. Surgical treatment of Parkinson’s disease: ablation, stimu-lation and transplantation. Expert Rev Neurother 2001; 1: 239-246.
Wilson BD, Bagnol D, Kaelin CB, Ollmann MM, Gantz I, Watson SJ,Barsh GS. Physiological and anatomical circuitry between Agouti-related protein and leptin signaling. Endocrinology 1999; 140: 2387-2397.
Wimalawansa SJ. Amylin, calcitonin gene-related peptide, calcitonin,and adrenomedullin: a peptide superfamily. Crit Rev Neurobiol 1997;11: 167-239.
Windhorst U, Johansson H. Modern techniques in neuroscience rese-arch. Springer, New York, 1999; 1325 pp.
Winterer G, Weinberger DR. Genes, dopamine and cortical signal-to-noise ratio in schizophrenia. Trends Neurosci 2004; 27: 683-690.
Wise RA. Dopamine, learning and motivation. Nat Rev Neurosci 2004;5: 483-494.
Wofsey AR, Kuhar MJ, Snyder SH. A unique synaptosomal fraction,which accumulates glutamic and aspartic acids, in brain tissue. ProcNatl Acad Sci USA 1971; 68: 1102-1106.
Wolosker H, Panizzutti R, De Miranda J. Neurobiology through thelooking-glass: D-serine as a new glial-derived transmitter. NeurochemInt 2002; 41: 327-332.
Won SJ, Kim DY, Gwag BJ. Cellular and molecular pathways of ische-mic neuronal death. J Biochem Mol Biol 2002; 35: 67-86.
Wong AH, Van Tol HH. Schizophrenia: from phenomenology to neuro-biology. Neurosci Biobehav Rev 2003; 27: 269-306.
Wong CG, Bottiglieri T, Snead OC. 3rd. GABA, gamma-hydroxybutyricacid, and neurological disease. Ann Neurol 2003; 54: S3-S12.
Woods SC, Gibbs J. The regulation of food intake by peptides Ann NYAcad Sci 1989; 575: 236-243.
Wynne K, Stanley S, McGowan B, Bloom S. Appetite control. J En-docrinol 2005; 184: 291-318.

266 NEUROTRANSMITERY
Xu ZQ, Zheng K, Hokfelt T. Electrophysiological studies on galanin ef-fects in brain-progress during the last six years. Neuropeptides 2005;39: 269-275.
Xue L, Farrugia G, Miller SM, Ferris CD, Snyder SH, Szurszewski JH.Carbon monoxide and nitric oxide as coneurotransmitters in the en-teric nervous system: evidence from genomic deletion of biosyntheticenzymes. Proc Natl Acad Sci USA 2000; 97: 1851-1855.
Yildiz BO, Suchard MA, Wong ML, McCann SM, Licinio J. Alterationsin the dynamics of circulating ghrelin, adiponectin, and leptin inhuman obesity. Proc Natl Acad Sci USA 2004; 101:10434-10439.
Yokoyama H. The role of central histaminergic neuron system as ananticonvulsive mechanism in developing brain. Brain Dev 2001; 23:542-547.
Yoshimichi G, Yoshimatsu H, Masaki T, Sakata T. Orexin-A regulatesbody temperature in coordination with arousal status. Exp Biol Med(Maywood) 2001; 226: 468-476.
Zadina JE, Banks WA, Kastin AJ. Central nervous system effects ofpeptides, 1980-1985: a cross-listing of peptides and their central ac-tions from the first six years of the journal Peptides. Peptides 1986;7: 497-537.
Zadina JE, Martin-Schild S, Gerall AA, Kastin AJ, Hackler L, Ge LJ,Zhang X. Endomorphins: novel endogenous mu-opiate receptor ago-nists in regions of high mu-opiate receptor density. Ann NY AcadSci 1999; 897: 136-144.
Zakon HH. Convergent evolution on the molecular level. Brain BehavEvol 2002; 59: 250-261.
Zhang JV, Ren PG, Avsian-Kretchmer O, Luo CW, Rauch R, KleinC, Hsueh AJW. Obestatin, a peptide encoded by the ghrelin gene,opposes ghrelin’s effects on food intake. Science 2005; 310: 996-999.
Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM.Positional cloning of the mouse obese gene and its human homologue.Nature 1994; 372: 406-407.
Zhou CJ, Shioda S, Yada T, Inagaki N, Pleasure SJ, Kikuyama S. PA-CAP and its receptors exert pleiotropic effects in the nervous systemby activating multiple signaling pathways. Curr Protein Pept Sci2002; 3: 423-439.

Literatúra 267
Zigmond MJ. Otto Loewi and the demonstration of chemical neuro-transmission. Brain Res Bull 1999; 50: 347-348.
Zoidl G, Dermietzel R. On the search for the electrical synapse: a glim-pse at the future. Cell Tissue Res 2002; 310: 137-142.
Zoli M, Agnati LF.Wiring and volume transmission in the central ner-vous system: the concept of closed and open synapses. Prog Neurobiol1996; 49: 363-380.
Zubrzycka M, Janecka A. Substance P: transmitter of nociception (Mi-nireview). Endocr Regul 2000; 34: 195-201.

Odkazy na webové stránky
Neurotransmitery
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=Books
• Basic Neurochemistry. Molecular, Cellular and Medical As-pects. Sixth Edition. Siegel GJ, Agranoff BW, Albers RW, FisherSK, Uhler MD (Eds.), Philadelphia, Lippincott Williams & Wil-kins, 1999.
• Neuroscience. Second Edition. Purves D, Augustine GJ, Fitz-patrick D, Lawrence K, LaMantia AS, McNamara JO, WilliamsSM. (Eds), Sunderland (MA), Sinauer Associates, 2001.
http://www.acnp.org/modules.php?name=Content&pa=showpage&pid=39
• Neuropsychopharmacology: The Fifth Generation of Progress.Davis KL, Charney D, Coyle JT, Nemeroff C (Eds.), Lippincott,Williams & Wilkins, Philadelphia, 2002.
http://www.benbest.com/science/anatmind/anatmd10.htmlhttp://www.neurotransmitter.nethttp://en.wikipedia.org/wiki/Neurotransmitterhttp://www.psy.plym.ac.uk/year1/Bbb.htmhttp://www.chemistry.emory.edu/justice/chem190j/
Látková závislosť
http://www.a1b2c3.com/drugs/index.htmhttp://www.addictionscience.net/
268

Odkazy na webové stránky 269
Parkinsonova choroba
http://www.scienceden.com/mbiology/research/parkinsonshttp://www.parkinson.org/site/pp.asp?c=9dJFJLPwB&b=71354http://www.nlm.nih.gov/medlineplus/parkinsonsdisease.html
Schizofrénia
http://www.schizophrenia.com/index.htmlhttp://www.nlm.nih.gov/medlineplus/schizophrenia.html

Použité skratky
5-HIAA kyselina 5-hydroxyindoloctová5-HT sérotonín (5-hydroxytryptamín)AADC dekarboxyláza aromatických aminokyselínAC adenylátcyklázaACTH adrenokortikotropný hormónADH antidiuretický hormón (vazopresín)ADM adrenomedulínADR adrenalínAGRP agouti príbuzný proteín
(agouti-related protein)AH adenohypofýzaAHL area hypothalamica lateralisACh acetylcholínAMPA kyselina α-amino-3-hydroxy-5-metyl-4-izoxazole-
propionováAMY amylínANP atriálny nátriuretický peptidANS autonómny nervový systémAP area postremaARAS ascendentný retikulárny aktivačný systémATP adenozíntrifosfátAVP vazopresín (arginín vazopresín)BBB hematoencefalická bariéra (blood-brain-barrier)BDNF mozgový rastový faktorBNP mozgový nátriuretický peptid
(brain natriuretic peptide)β-END β-endorfínCaM kalmodulínCaMKIV kalmodulín dependentná kináza IVcAMP cyklický adenozínmonofosfát
270

Použité skratky 271
CART kokaín-amfetamín regulovaný transkript(cocaine-amphetamine-regulated transcript)
CCK cholecystokinínCGRP kalcitonínovému génu príbuzný peptid
(calcitonin gene-related peptide)CNS centrálny nervový systémCO oxid uhoľnatýCOMT katechol-O-metyltransferázaCREB cAMP-response-element-binding proteinCRH kortikoliberín (CRF)CVOs cirkumventrikulárne orgányDA dopamínDAG diacylglycerolDAT dopamínový transportérDBH dopamín-β-hydroxylázaDOPA dihydroxyfenylalanínDOPAC kyselina dihydroxyfenyloctováDYN dynorfínEPI epifýzaGABA kyselina γ-aminomaslová (γ-aminobutyric acid)GAL galanínGALP galanin-like peptideGHB kyselina γ-hydroxymaslová (γ-hydroxybutyric acid)GHRH rastový hormón uvoľňujúcí hormónGIT gastrointestinálny traktGLP-1 glukagónu podobný peptid 1GLP-2 glukagónu podobný peptid 2Glu glutamátGly glycínGN ganglion nodosumGnRH gonadotropín uvoľňujúci hormónH2S hydrogénsulfátHCRT hypokretínHist histamínHPA os hypotalamo-hypofýzo-adrenokortikálna os
(hypothalamo-pituitary adrenocortical axis)IP3 inozitol-3-fosfát

272 NEUROTRANSMITERY
LC locus coeruleusLTP dlhodobý potenciálMAO monoaminooxidázaME eminentia medianaMCH melanín koncentrujúci hormónMSH melanocyty stimulujúci hormónN/OFQ nociceptín/orfanín FQNA noradrenalínNANC nonadrenergic/noncholinergic (nervový systém)NARC nucleus arcuatusNDM nucleus dorsomedialisNET noradrenalínový transportérNH neurohypofýzaNKA neurokinín ANKB neurokinín BNPK neurokinín KNMDA N-metyl-d-aspartátNO oxid dusnatýNPAF neuropeptid AFNPB neuropeptid BNPEI neuropeptid EINPFF neuropeptid FFNPGE neuropeptid GENPSF neuropeptid SFNPS neuropeptid SNPW neuropeptid WNPY neuropeptid YNSCH nucleus suprachiasmaticusNT neurotenzínNTS nucleus tractus solitariiNVM nucleus ventromedialisORX orexínOVLT organum vasculosum laminae terminalisOXY oxytocínPACAP hypofýzovú adenylátcyklázu stimulujúci polypeptid
(pituitary adenylate cyclase-activating polypeptide)PHI histidínový izoleucínový peptid

Použité skratky 273
PKA proteínkináza APNMT fenyletanolamín-N-metyltransferázaPNS periférny nervový systémPOMC proopiomelanokortínPP pankreatický polypeptidPRL prolaktínPrRP prolaktín uvoľňujúci peptidPVN nucleus paraventricularisPYY peptid YYRVLM rostrálna ventro-laterálna oblasť predĺženej miechySCO organum subcommissuraleSFO organum subfornicaleSIF cells „small intensely fluorescent cellsÿSOM somatostatínSON nucleus supraopticusSP substancia PTH tyrozínhydroxylázaTRH tyreoliberín; tyreotropín uvoľňujúci hormónVIP vazoaktívny intestinálny polypeptidVTA ventrálna tegmentálna area

Zoznam tabuliek
2.1 Rozdelenie neurotransmiterov podľa príslušnosti k chemic-kým skupinám . . . . . . . . . . . . . . . . . . . . . . . . 24
2.2 Prehľad vybraných malomolekulových neurotransmiterova ich štruktúrnych vzorcov . . . . . . . . . . . . . . . . . 26
2.3 Prehľad vybraných skupín neuropeptidov . . . . . . . . . . 272.4 Prehľad jednotlivých typov a podtypov G-proteínov a me-
chanizmov ich účinku . . . . . . . . . . . . . . . . . . . . 302.5 Označovanie skupín neurónov podľa typu neurotransmitera,
ktorý syntetizujú . . . . . . . . . . . . . . . . . . . . . . . 37
3.1 Prehľad esenciálnych a neesenciálnych aminokyselín nachá-dzajúcich sa v ľudskom organizme . . . . . . . . . . . . . 47
3.2 Klasifikácia receptorov pre glutamát a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 49
3.3 Klasifikácia receptorov pre GABA a mechanizmy transduk-cie signálu . . . . . . . . . . . . . . . . . . . . . . . . . . 55
3.4 Klasifikácia receptorov pre acetylcholín a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 64
3.5 Klasifikácia receptorov pre dopamín a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 70
3.6 Klasifikácia receptorov pre noradrenalín a adrenalín a me-chanizmy transdukcie signálu . . . . . . . . . . . . . . . . 74
3.7 Klasifikácia receptorov pre sérotonín a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 81
3.8 Klasifikácia receptorov pre histamín a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 83
3.9 Klasifikácia receptorov pre ATP a mechanizmy transdukciesignálu . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
3.10 Klasifikácia receptorov pre adenozín a mechanizmy trans-dukcie signálu . . . . . . . . . . . . . . . . . . . . . . . . 87
274

Zoznam tabuliek 275
3.11 Prehľad receptorov pre neurotransmitery, ktorých činnosť jemodulovaná neurosteroidmi . . . . . . . . . . . . . . . . 89
3.12 Klasifikácia receptorov pre kanabinoidy a mechanizmytransdukcie signálu . . . . . . . . . . . . . . . . . . . . . 98
4.1 Prehľad najčastejšie sa vyskytujúcej kolokalizácie neuro-transmiterov . . . . . . . . . . . . . . . . . . . . . . . . 105
4.2 Klasifikácia receptorov pre neuropeptidy a mechanizmytransdukcie signálu . . . . . . . . . . . . . . . . . . . . . 133
5.1 Prehľad účasti monoamínergických neurónov v reguláciibdenia a spánku . . . . . . . . . . . . . . . . . . . . . . . 150
5.2 Modulátory autonómneho nervového systému s vyhranenoulokalizáciou (A) a prehľad ďalších modulátorov autonóm-neho nervového systému (B) . . . . . . . . . . . . . . . . 155

Zoznam obrázkov
1.1 Štruktúra neurónu . . . . . . . . . . . . . . . . . . . . . . 12
2.1 Schematické znázornenie jednotlivých typov komunikáciev nervovom systéme . . . . . . . . . . . . . . . . . . . . . 19
2.2 Schematické znázornenie základných charakteristík neuro-transmiterov . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.3 Schematické znázornenie aktivácie ionotropného receptora 312.4 Schematické znázornenie aktivácie metabotropného receptora 322.5 Schematické znázornenie vplyvu neurotransmiterov na gé-
novú expresiu v neuróne . . . . . . . . . . . . . . . . . . . 342.6 Lokalizácia cirkumventrikulárnych orgánov v mozgu človeka
a endokrinných žliaz mozgu s fenestrovanými kapilárami . 42
3.1 Schéma biosyntézy glutamátu . . . . . . . . . . . . . . . 483.2 Schematické znázornenie lokalizácie a dráh glutamátergic-
kých neurónov v mozgu človeka . . . . . . . . . . . . . . 503.3 Schéma biosyntézy aspartátu . . . . . . . . . . . . . . . . 523.4 Štruktúrne vzorce homocysteínu, cysteínu a ich derivátov . 533.5 Schéma biosyntézy GABA . . . . . . . . . . . . . . . . . 543.6 Schematické znázornenie lokalizácie a dráh GABAergických
neurónov v mozgu . . . . . . . . . . . . . . . . . . . . . 563.7 Schéma biosyntézy GHB . . . . . . . . . . . . . . . . . . 573.8 Schéma biosyntézy glycínu . . . . . . . . . . . . . . . . . 583.9 Schematické znázornenie lokalizácie glycínergických neuró-
nov v mozgu a v mieche . . . . . . . . . . . . . . . . . . 593.10 Schéma zapojenia modulačných neurotransmiterov v regu-
lácii činnosti nervového systému . . . . . . . . . . . . . . 613.11 Uvoľňovanie modulačných neurotransmiterov z „varikozít“
axónov . . . . . . . . . . . . . . . . . . . . . . . . . . . . 623.12 Schéma syntézy a metabolizmu acetylcholínu . . . . . . . 63
276

Zoznam obrázkov 277
3.13 Schematické znázornenie lokalizácie hlavných zoskupení cho-línergických neurónov a cholínergických dráh v mozgu a mie-che . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
3.14 Schéma biosyntézy katecholamínov . . . . . . . . . . . . . 683.15 Schematické znázornenie noradrenalínergického nervového
zakončenia . . . . . . . . . . . . . . . . . . . . . . . . . . 693.16 Schéma metabolizmu dopamínu . . . . . . . . . . . . . . 713.17 Schematické znázornenie lokalizácie dopamínergických ne-
urónov a dopamínergických dráh v mozgu . . . . . . . . . 723.18 Schéma metabolizmu noradrenalínu . . . . . . . . . . . . 753.19 Schematické znázornenie lokalizácie noradrenalínergických
neurónov a noradrenalínergických dráh v mozgu . . . . . . 763.20 Schéma metabolizmu adrenalínu . . . . . . . . . . . . . . 773.21 Schematické znázornenie lokalizácie noradrenalínergických
neurónov a noradrenalínergických dráh v mozgu . . . . . . 783.22 Schéma syntézy a metabolizmu sérotonínu . . . . . . . . . 803.23 Schematické znázornenie lokalizácie sérotonínergických ne-
urónov a sérotonínergických dráh v mozgu . . . . . . . . . 823.24 Schéma syntézy a metabolizmu histamínu. . . . . . . . . . 833.25 Schematické znázornenie lokalizácie histamínergických neu-
rónov a histamínergických dráh v mozgu. . . . . . . . . . 843.26 Štruktúrne vzorce ATP a adenozínu . . . . . . . . . . . . 853.27 Schéma biosyntézy neurosteroidov . . . . . . . . . . . . . 883.28 Schéma biosyntézy NO z l-arginínu . . . . . . . . . . . . 923.29 Schéma biosyntézy CO z hému . . . . . . . . . . . . . . . 943.30 Schéma biosyntézy H2S z cysteínu . . . . . . . . . . . . . 953.31 Schéma biosyntézy prostaglandínov . . . . . . . . . . . . 963.32 Štruktúrne vzorce endogénnych agonistov a exogénneho ago-
nistu kanabinoidových receptorov . . . . . . . . . . . . . 973.33 Schéma biosyntézy DOPA, tryptamínu, fenyletylamínu, ok-
topamínu a tyramínu . . . . . . . . . . . . . . . . . . . . 993.34 Štruktúrne vzorce endogénnych ligandov pre imidazolínové
receptory . . . . . . . . . . . . . . . . . . . . . . . . . . 100
5.1 Schematické znázornenie periférnych a centrálnych zložiekzúčastňujúcich sa na regulácii príjmu potravy . . . . . . . 138
5.2 Neurotransmiterové systémy, ktoré sa zúčastňujú na regulá-cii homeostázy telesných tekutín . . . . . . . . . . . . . . 148

278 NEUROTRANSMITERY
5.3 Schematické znázornenie zoskupení neurónov modulujúcichprocesy spánku a bdenia . . . . . . . . . . . . . . . . . . 149
5.4 Schematické znázornenie úlohy glutamátu v procesochtvorby dlhodobých potenciálov . . . . . . . . . . . . . . . 153
5.5 Schematické znázornenie sympatikových a parasympatiko-vých dráh . . . . . . . . . . . . . . . . . . . . . . . . . . 156
5.6 Schéma nervových okruhov tvoriacich enterický nervový sys-tém . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
5.7 Schéma motorickej a senzorickej inervácie srdca sympatiko-vým a parasympatikovým nervovým systémom a detail ner-vových okruhov v srdcových gangliách . . . . . . . . . . . 160
5.8 Schéma prenosu informácií o pôsobení stresorov . . . . . . 1645.9 Neuroendokrinná odpoveď v dôsledku pôsobenia stresového
podnetu . . . . . . . . . . . . . . . . . . . . . . . . . . . 1655.10 Prehľad neurotransmiterov zúčastňujúcich sa na prenose no-
ciceptívnych informácií a ich modulácii . . . . . . . . . . 174
6.1 Úbytok počtu dopamínergických neurónov v oblasti substan-tia nigra u pacientov s Parkinsonovou chorobou . . . . . . 182
6.2 Schéma zobrazujúca okruhy bazálnych ganglií, ktorých čin-nosť je u pacientov s Parkinsonovou chorobou narušená . . 184
6.3 Prefrontálny-striatový model schizofrénie . . . . . . . . . 1876.4 Schematické znázornenie noradrenalínergického a sérotoní-
nergického synaptického prenosu, ich vzájomných interakciía miesta účinku antidepresív . . . . . . . . . . . . . . . . 194
6.5 Schematické znázornenie štruktúr a dráh CNS, o ktorýchsa predpokladá, že ich dysfunkcia sa môže zúčastňovaťna etiopatogenéze depresie . . . . . . . . . . . . . . . . . 197
6.6 Schematické znázornenie faktorov, ktoré ovplyvňujú neuro-plasticitu a odolnosť neurónov . . . . . . . . . . . . . . . 199
6.7 Kľúčové štruktúry nervového systému, o ktorých sa predpo-kladá, že sa zúčastňujú na vzniku látkovej závislosti . . . . 202
6.8 Schéma procesov aktivovaných glutamátom počas ischémieneurónov . . . . . . . . . . . . . . . . . . . . . . . . . . 205
7.1 Kľúčové štruktúry predĺženej miechy zúčastňujúce sa na re-gulácii krvného tlaku . . . . . . . . . . . . . . . . . . . . 208

Index
A
A1-A7 37, 74A8-A16 37, 70AADC 68Acetylcholín 20, 60, 61, 151,
209biosyntéza 62distribúcia neurónov 64funkcie 66receptory 63Acetylcholínesteráza 63ACTH 109, 113Adenozín 85ADH 107Adrenalín 77biosyntéza 77distribúcia neurónov 78funkcie 79receptory 77Adrenomedulín 120Afektívne poruchy 77, 110, 190Agouti príbuzný proteín 121Agmatín 100Akčný potenciál 14Alzheimerova choroba 53, 66,
93, 181Aminokyseliny 46
Amylín 121Anandamín 97Angiotenzín 42, 122Apelín 122Area hypothalamica lateralis
144Area postrema 41, 145Area ventralis tegmenti (Tsi)
70, 201Aspartát 51ATP 85, 157Atriálny nátriuretický peptid
146Autonómny nervový systém 64,
153
B
B1-B9 37, 82Barbituráty 56Benzodiazepíny 56Bdenie 147BDNF 198Biogénne amíny 60Bolesť 56, 73, 114, 172centrálna modulácia 173neuropatická 172, 175nociceptívna 172Botulotoxín 67
279

280 NEUROTRANSMITERY
C
C1-C3 37, 78, 169Cirkumventrikulárne orgány 41CRH 109, 163Cysteín 53Cysteínsulfát 53Cytokíny 209
D
Depresia 82, 190DOPA 98, 183funkcie 98Dopamín 67biosyntéza 68distribúcia neurónov 70funkcie 73receptory 68Dopamín-β-hydroxyláza 68Dorzálny noradrenergický zvä-
zok 75Dráhamezokortikálna 71mezolimbická 71nigrostriatálna 71tuberoinfundibulárna 71Dynorfín 114
E
E1-E4 37Eminentia mediana 41, 71Endomorfíny 115Endorfíny 114Enkefalíny 114Enterický nervový systém 157Epilepsia 51, 57, 203Excitotoxicita 205
F
Fenyletanolamín-N-metyltransferáza68
Fenyletylamín 98
G
G-proteíny 30GABA 53, 176biosyntéza 54distribúcia neurónov 55funkcie 56receptory 54Galanín 123Galanin-like peptid 123Ghrelín 123GHRH 112Glia 11funkcie 18Glukagónu podobné peptidy
118Glukokortikoidy 20, 110, 170Glutamát 47biosyntéza 48distribúcia neurónov 50funkcie 51receptory 49Glycín 58biosyntéza 58distribúcia neurónov 59funkcie 60receptory 58
H
Harman 100Hemato-encefalická bariéra 36

Index 281
Histamín 82, 151biosyntéza 82distribúcia neurónov 84funkcie 84receptory 82Histidínový izoleucínový peptid
119Homeostáza tekutín 146Homocysteín 53Homocysteínsulfát 53Huntingtonova choroba 51, 57,
73, 93Hypertenzia 207Hypofýzovú adenylátcyklázu
aktivujúci peptid119
Hypokretín 131Hypotalamo-hypofýzo-adreno-
kortikálna os 92,170, 195
Hypoxia 177
CH
CH1-CH6 37Chemokíny 25Cholecystokinín 124Cholín 62Chrómafinné bunky 77Chromagraníny 125
I
Imidazolíny 99funkcie 100väzbové miesta 99Imipramín 191
Intrakardiálny nervový systém159
Inzulín 140Ischémia 51, 93, 204
K
Kalcitonínu génovo príbuznýpeptid 126
Kanabinoidy 96funkcie 97Katecholamíny 67, 169Koexpresia 104Kokaínom a amfetamínom
regulovaný trans-kript 126
Kolokalizácia 104Kotransmisia 85, 104Kosyntéza 104Kyselina
γ-hydroxymaslová 57ribotidimimidazol-4-octová
100ribozidimimidazol-4-octová
100
L
Lambertov-Eatonov myastenic-ký syndróm 66
Leptín 140Locus coeruleus 74, 151, 163LTP 152
M
Melanín koncentrujúci hormón127

282 NEUROTRANSMITERY
Membránapresynaptická 15postsynaptická 15, 21, 35Motoneuróny 64Migréna 82Mozgový nátriuretický peptid
127MSH 113Myasthenia gravis 66
N
Nadoblička 77, 110, 120Narkolepsia 132, 152Neurogenéza 196Neurón 11axón 13cytoskelet 13dendrity 13jadro 13membrána 11Nisslova substancia 13Renshawov 60telo 11Neuropeptidy 102funkcie 103charakteristiky 103Neurosteroidy 86funkcie 88Nonadrenergický/noncholínergický
nervový systém159
Noradrenalín 73, 151biosyntéza 73distribúcia neurónov 74receptory 73Nervus vagus 61, 145, 209
Neurodegeneratívne ochorenia181
Neurogenéza 196Neurokinín A 115Neurokinín B 115Neuropeptid B 127Neuropeptid FF 128Neuropeptid K 115Neuropeptid S 128Neuropeptid W 129Neuropeptid Y 117, 157Neuropeptid YY 139Neuropeptid γ 115Neuropeptidy 27, 103funkcie 136charakteristika 103receptory 133Neurotenzín 129Neurotransmiteryevolúcia 44charakteristika 20inaktivácia 22kritériá 20malomolekulové 20, 24, 26, 46plynné 90účinky 33, 136veľkomolekulové 21, 24, 27, 102Neurotransmiterové systémyklasifikácia 16, 22, 24, 25, 35Nociceptín/orfanín 130Nocistatín 131Noladín ester 97Nuclei raphe 80Nucleus arcuatus 71, 141Nucleus dorsomedialis 143Nucleus paraventricularis 71,
91, 108, 109, 142,163

Index 283
Nucleus perifornicalis 144Nucleus suprachiasmaticus 145Nucleus supraopticus 91, 108Nucleus tractus solitarii 145Nucleus tuberomamillaris 84Nucleus ventromedialis 143
O
Obestatín 124Obezita 140Oktopamín 98Opioidy 113, 173Orexín 131, 151Organum subfornicale 41Organum vasculosum laminae
terminalis 41, 146Oxid dusnatý 90, 91, 157, 205biosyntéza 91funkcie 91Oxid uhoľnatý 90, 94biosyntéza 94funkcie 94Oxytocín 107
P
Pamäť 152Parasympatikový nervový sys-
tém 64, 157Parkinsonova choroba 73, 93,
181, 182Peptid YY 118Prenos signálov 14, 18Príjem potravy 97, 137anorexigenické neuróny 139orexigenické neuróny 139Proopiomelanokortín 113
Prolaktín uvoľňujúci peptid 132Prostaglandíny 95funkcie 96Psychostimulanciá 201Puríny 85
R
Receptory 21, 25AMPA 49, 152ionotropné 29kainát 49metabotropné 29NMDA 49, 58, 91, 152, 175,
193postsynaptické 21presynaptické 21Rezerpín 191RVLM 207
S
Sauvagín 109Sekretoneurín 125Sérotonín 79, 151biosyntéza 79distribúcia neurónov 79funkcie 81receptory 79Schizofrénia 53, 185SIF-bunky 156, 159Sírovodík 90, 95biosyntéza 95funkcie 95Somatostatín 112Spánok 147non-REM 150REM 150

284 NEUROTRANSMITERY
Starnutie 178Stopové amíny 97Stres 76, 161klasifikácia 162následky 171neuronálne okruhy 162špecificita 161Substantia nigra 70, 182Substancia P 115Sympatikoadrenálny systém
78, 169Sympatikový nervový systém
64, 154centrálna regulácia 154Synapsaelektrická 14, 18, 55en passant 18, 61, 150, 154chemická 14, 18štrbina 18
T
Tachykiníny 115Telesná teplota 177Transportérdopamínový 201vezikulárny monoamínový 201Tryptamín 98Tyramín 98Tyreoliberín 113
Tyrozínhydroxyláza 68
U
Učenie 152Urokortín 109, 111Urotenzín 109, 111
V
Vazoaktívny intestinálny poly-peptid 119
Vazopresín 107Vedomie 20Ventrálny noradrenergický zvä-
zok 75Vezikuly 21Virodhamín 97Vývin nervového systému 175
Z
Zápal 209Závislosť 115, 132, 201alkoholizmus 202Zlyhanie srdca 127
5
5-HT-modulín 135

















![PROBLEMATIKA VYUŽITIA BROWNFIELDS V RÁMCI FUNKČNEJ ... · mi CLARINET a CABERNET [11, 12] zaoberajúcimi sa analýzou troch skupín faktorov v rámci funkčnej intraurbán-nej](https://static.fdocuments.net/doc/165x107/5e2fc44abf54d0613871988c/problematika-vyuitia-brownfields-v-rmci-funkoenej-mi-clarinet-a-cabernet.jpg)
