
Nerve Growth Factor
67
Annu. Rev. Neurosci. 2001. 24:1217–281 Copyright c 2001 by Annual Reviews. All rights reserved NERVE GROWTH FACTOR SIGNALING, NEUROPROTECTION, AND NEURAL REPAIR Michael V Sofroniew Department of Neurobiology and Brain Research Institute, University of California Los Angeles, Los Angeles, California 90095-1763; e-mail: [email protected] Charles L Howe Department of Neurology and Neurological Sciences, Stanford University, Stanford, California 94305-5489; e-mail: [email protected] William C Mobley Department of Neurology and Neurological Sciences, Stanford University, Stanford, California 94305; e-mail: [email protected] Key Words neurotrophins, NGF, TrkA, p75 NTR , neurodegeneration, neuroregeneration, excitotoxicity, tyrosine kinase ■ Abstract Nerve growth factor (NGF) was discovered 50 years ago as a molecule that promoted the survival and differentiation of sensory and sympathetic neurons. Its roles in neural development have been characterized extensively, but recent findings point to an unexpected diversity of NGF actions and indicate that developmental effects are only one aspect of the biology of NGF. This article considers expanded roles for NGF that are associated with the dynamically regulated production of NGF and its receptors that begins in development, extends throughout adult life and aging, and involves a surprising variety of neurons, glia, and nonneural cells. Particular attention is given to a growing body of evidence that suggests that among other roles, endogenous NGF signaling subserves neuroprotective and repair functions. The analysis points to many interesting unanswered questions and to the potential for continuing research on NGF to substantially enhance our understanding of the mechanisms and treatment of neurological disorders. INTRODUCTION In mammals and other vertebrates, soluble peptide growth factors play essential roles in intercellular communication. They exert their effects by signaling through surface membrane receptors that interact with diverse types of intracellular second- messenger systems. In a sometimes surprising manner, many growth factors have been found to subserve a wide variety of functions by acting on many cell types at different stages of development or in adult life. 0147-006X/01/0621-1217$14.00 1217 Annu. Rev. Neurosci. 2001.24:1217-1281. Downloaded from arjournals.annualreviews.org by University of California - San Diego on 08/23/10. For personal use only.
-
Upload
teratos2025 -
Category
Documents
-
view
227 -
download
0
description
Información sobre el factor de crecimiento nervioso.
Transcript of Nerve Growth Factor

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
Annu. Rev. Neurosci. 2001. 24:1217–281Copyright c© 2001 by Annual Reviews. All rights reserved
NERVE GROWTH FACTOR SIGNALING,NEUROPROTECTION, AND NEURAL REPAIR
Michael V SofroniewDepartment of Neurobiology and Brain Research Institute, University of California LosAngeles, Los Angeles, California 90095-1763; e-mail: [email protected]
Charles L HoweDepartment of Neurology and Neurological Sciences, Stanford University, Stanford,California 94305-5489; e-mail: [email protected]
William C MobleyDepartment of Neurology and Neurological Sciences, Stanford University, Stanford,California 94305; e-mail: [email protected]
Key Words neurotrophins, NGF, TrkA, p75NTR, neurodegeneration,neuroregeneration, excitotoxicity, tyrosine kinase
■ Abstract Nerve growth factor (NGF) was discovered 50 years ago as a moleculethat promoted the survival and differentiation of sensory and sympathetic neurons. Itsroles in neural development have been characterized extensively, but recent findingspoint to an unexpected diversity of NGF actions and indicate that developmental effectsare only one aspect of the biology of NGF. This article considers expanded roles forNGF that are associated with the dynamically regulated production of NGF and itsreceptors that begins in development, extends throughout adult life and aging, andinvolves a surprising variety of neurons, glia, and nonneural cells. Particular attentionis given to a growing body of evidence that suggests that among other roles, endogenousNGF signaling subserves neuroprotective and repair functions. The analysis points tomany interesting unanswered questions and to the potential for continuing research onNGF to substantially enhance our understanding of the mechanisms and treatment ofneurological disorders.
INTRODUCTION
In mammals and other vertebrates, soluble peptide growth factors play essentialroles in intercellular communication. They exert their effects by signaling throughsurface membrane receptors that interact with diverse types of intracellular second-messenger systems. In a sometimes surprising manner, many growth factors havebeen found to subserve a wide variety of functions by acting on many cell typesat different stages of development or in adult life.
0147-006X/01/0621-1217$14.00 1217
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1218 SOFRONIEW ¥ HOWE ¥ MOBLEY
Nerve growth factor (NGF) was discovered 50 years ago as a molecule that regu-lates the survival and maturation of developing neurons in the peripheral nervoussystem (PNS) (Levi-Montalcini & Hamburger 1951, 1953), and ideas about thebiological role of NGF have been dominated by concepts that arose from studieson the differentiation and survival of young neurons. Until recently, the expecta-tion was that the biology of NGF would center around the classical target-derivedneurotrophic factor paradigm in which NGF released by postsynaptic targets actson presynaptic neurons to build or maintain functional contacts and enhance thefunction of well-defined neural circuits. Although this paradigm undoubtedly playsa critical role in both the PNS and central nervous system (CNS), it does not ap-pear to be the sole role for NGF actions. With the availability of tools that allowsensitive and specific measurements of mRNA and protein levels for NGF and itsreceptors, it has become apparent that NGF actions extend beyond the develop-mental period, beyond nerve cells, and even beyond the nervous system. Indeed,NGF and its receptors are produced throughout adult life and during aging bymany different cell types. The dynamically regulated expression of NGF and itsreceptors throughout adult life suggests multiple functions for NGF signaling,many of which are poorly understood. NGF and NGF receptor expression can beupregulated during the response to injury in both the PNS and CNS, and a grow-ing body of evidence suggests that among other roles, endogenous NGF signalingthrough both neurons and nonneuronal cells subserves neuroprotective functionsand facilitates neural repair.
One of the major advances of molecular neuroscience in the past 25 years hasbeen to recognize that much of the cellular damage resulting from such CNSinsults as stroke, trauma, and degenerative disease may be caused by a limi-ted number of endogenously generated molecules with neurotoxic activities. Lesswell developed is the idea that endogenous mechanisms exist to provide neuropro-tection, and that endogenous molecules may be produced specifically to subserveneuroprotective signaling functions (Mattson 1997). For NGF to be viewed as aspecifically expressed, neuroprotective molecule with widespread activity in theCNS, several criteria must be fulfilled: (a) NGF and NGF receptor expressionmust occur in cellular compartments where it could influence the neural responseto injury; (b) NGF signaling should be able to influence cellular events involvedin the response to insults and injury; (c) NGF should exert protective effects; and(d) failure of NGF signaling should be associated with increased degenerationand vulnerability to injury. In this review, we consider evidence supporting thesecriteria and conclude that NGF does play a role in endogenous neuroprotection.
STRUCTURE, EXPRESSION, AND REGULATIONOF NGF AND ITS RECEPTORS
The NGF gene is located on human chromosome 1 and is expressed as two majorsplice variants (Edwards et al 1986, 1988). The mature, fully processed form ofbiologically active NGF appears to be similar in all tissues and consists of a dimer of
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1219
13-kDa polypeptide chains, each of which has three intrachain disulfide bridges.The crystal structure of NGF has been resolved (McDonald et al 1991). The NGFdimer has an elongated shape with a core, or “waist,” that is formed by twisted betasheets; the molecule also features a cysteine-knot motif, a reverse turn at one end(loop 3) and three beta-hairpin loops at the other (loops 1, 2, and 3). The aminoterminus of NGF is not defined in the crystal structure. An octapeptide derived fromthe NGF amino terminus has potent bradykinin-like activity (Taiwo et al 1991)and is normally produced in the mouse submandibular gland in response to stress,but whether it is found under physiological conditions in other tissues is unknown(Fahnestock et al 1991). NGF is part of the neurotrophin family of molecules, whichshare a high degree of structural homology and includes brain-derived neurotrophicfactors (BDNF), neurotrophin-3 (NT-3), and neurotrophin-4 (NT-4) (Butte et al1998; Ibanez 1994; Robinson et al 1995, 1999). Neurotrophins are found in bothmammals and lower vertebrates, and the neurotrophin homologues NT-6 and NT-7were recently cloned in fish (Gotz et al 1994, Lai et al 1998).
NGF has two known receptors, TrkA and p75NTR (Bothwell 1995, Kaplan &Miller 1997). TrkA is a single-pass transmembrane protein that serves as a receptortyrosine kinase (RTK) for NGF. NGF signaling through TrkA elicits many of theclassical neurotrophic actions ascribed to NGF (Loeb et al 1991). TrkA is a memberof the Trk gene family, which includes TrkB, the receptor for BDNF and NT-4, andTrkC, the receptor for NT-3 (Kaplan & Miller 1997). NGF activates only TrkA;NT-3 activates TrkA but only does so at much higher concentrations than does NGF.Two isoforms for TrkA exist that differ in their extracellular domain through theinclusion of six additional amino acids near the transmembrane domain of one ofthe variants (TrkAII). Inclusion of the insert appears to relax the specificity of TrkAactivation; NT-3 mediated signaling is markedly enhanced through this receptorisoform (Clary & Reichardt 1994). p75NTR is a transmembrane glycoprotein thatbinds all members of the neurotrophin family with approximately equal nanomolaraffinity. p75NTR regulates signaling through TrkA; in addition, as discussed below,NGF binding to p75NTR activates signaling pathways that are characteristic for thisreceptor (Casaccia-Bonnefil et al 1999; Dobrowsky et al 1994, 1995; Friedman &Greene 1999).
Recent findings for the three-dimensional structure of NGF bound to its TrkAreceptor provide a structural explanation for many of the results provided by mu-tagenesis studies (Wiesmann et al 1999). They show that NGF engages the TrkAsecond immunoglobulin (Ig)-like domain through two distinct patches (Wiesmannet al 1999). The first patch involves the four beta sheets that form the “waist” of theNGF molecule together with the first loop (residues 29–33); it includes NGF do-mains that show considerable homology with the other neurotrophins (Wiesmannet al 1999). It is likely that NGF and its neurotrophin family members engage eachof their Trk receptors through this patch. The second patch is formed by the aminoterminus of NGF, which in the NGF-TrkA structure is well defined (Wiesmannet al 1999). The lack of homology of the NGF amino terminus with that of otherneurotrophins suggests that the second patch serves to specify NGF binding toTrkA. As yet there is no three-dimensional structure for NGF binding to p75NTR.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1220 SOFRONIEW ¥ HOWE ¥ MOBLEY
Mutagenesis studies for NGF binding to p75NTR point to the importance of mostlydifferent domains (i.e. the first, third, and fourth loops and the carboxy-terminus)(Ibanez et al 1992, Ryden & Ibanez 1997, Urfer et al 1994) than those identifiedfor binding to TrkA. The findings suggest that NGF could bind to both TrkA andp75NTR simultaneously (Wiesmann et al 1999).
Both NGF and its receptors are produced during development, adult life, andaging by many cell types in the CNS and PNS, immune and inflammatory sys-tem, and various tissues. Given the wide range of neuronal and nonneuronal cellsthat have the potential to produce and/or respond to NGF, clues to the differentfunctions that might be played by NGF signaling have been obtained by exam-ining the expression of NGF and its receptors. During development, expressionof NGF by target cells is compatible with its role as a survival and maturationfactor for afferent neurons. In addition, as discussed in this section, a large bodyof evidence demonstrates that in response to numerous stimuli there is dynamicregulation of NGF and NGF receptor expression. It is interesting that NGF and/orits receptors are markedly upregulated by many cell types after tissue injury orinsult. Documenting the patterns for NGF and NGF receptor gene expression inspecific cells and tissues is required for documenting the plurality of NGF actionsand for interpreting their physiological significance.
Peripheral Nervous System and Peripheral Tissues
NGF Receptor Expressing Cells Sympathetic neurons and small diameter peri-pheral sensory neurons that mediate nociception, the first identified NGF-respon-sive neurons, express both TrkA and p75NTR during development and in the adult(Ruit et al 1990, Verge et al 1989). Most, if not all,α-motor neurons, whose cellbodies reside in the CNS and send projections through peripheral nerve to muscletargets, transiently express p75NTR during the phase of axon elongation that occursin development; expression is downregulated to undetectable levels in adults butreturns after peripheral nerve injury (Ernfors et al 1989, Wood et al 1990). AmongPNS glial cells, Schwann cells in peripheral nerve express p75NTR during devel-opment. In the normal adult, p75NTR expression is reduced to levels that are onlyone percent of those seen during development (Heumann et al 1987b). Schwanncells markedly upregulate p75NTR in response to the loss of contact with axonsthat follows axotomy, to local tissue injury, or if stimulated with inflammatorycytokines (Heumann et al 1987b, Lemke & Chao 1988, Mirsky & Jessen 1999,Taniuchi et al 1988). Expression patterns for NGF receptors in the PNS suggestthat distinct functions are carried out during development, normal adult life, andfollowing injury.
NGF-Producing Cells Nonneuronal target cells of sympathetic and sensory neu-rons throughout the body produce NGF during development. These include targetsin the skin (e.g. keratinocytes and melanocytes), vascular and other smooth musclecells, and various endocrine tissues, such as the testis and ovary, pituitary, thyroidand parathyroid, and exocrine salivary (e.g. submandibular) glands. Most of these
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1221
cells continue to produce NGF during adult life and modulate NGF productionin response to stimuli (reviewed by Levi-Montalcini et al 1995, 1996). In sometissues, including skin and viscera such as the bladder, experimental evidence sug-gests that NGF production is markedly upregulated after injury or in response totissue inflammation or injury, but the NGF-producing cell types have not yet beencharacterized (Dmitrieva et al 1997, McMahon et al 1995, Mendell et al 1999).Among PNS glia, immature Schwann cells and satellite cells produce NGF dur-ing development (Mirsky & Jessen 1999). In adults, mature myelinating Schwanncells downregulate NGF expression to undetectable levels, but after nerve injury,reactive and dedifferentiated Schwann cells markedly upregulate NGF productionin vivo; in vitro, NGF expression by Schwann cells is upregulated by cytokines andother inflammatory mediators (Lindholm et al 1987, Mirsky & Jessen 1999). Asfor its receptors, the patterns for NGF expression suggest roles that extend be-yond development and beyond its classical role as a target-derived neurotrophicfactor.
Central Nervous System
NGF Receptor Expressing Cells p75NTR gene expression in the CNS is wide-spread, especially during development. In addition to both major populations offorebrain cholinergic neurons, p75NTR mRNA and protein are found in a num-ber of developing neuronal populations in both the brain and brainstem (Longoet al 1993). p75NTR expression is more restricted in the adult, and several popu-lations, including cholinergic neurons of the caudate-putamen and cranial nervenuclei of the brainstem, show markedly reduced or no expression (Koh & Higgins1991). Cerebellar Purkinje neurons, hippocampal pyramidal neurons, and retinalganglion neurons also downregulate expression to undetectable levels in adultsbut reexpress p75NTR after injury (Brann et al 1999, Eckenstein 1988, Mart´ınez-Murillo et al 1998, Yamashita et al 1999b). The majority of p75NTR-expressingneurons do not also express TrkA, but developing horizontal cells and amacrinecells of the retina express TrkA and potentially p75NTR (Karlsson et al 1998),whereas cholinergic neurons of the septal-basal forebrain complex express bothTrkA and p75NTR during development and throughout adult life (Holtzman et al1992). It is interesting that expression of TrkA, but not of p75NTR, in these neuronsis significantly decreased in aged animals (Cooper et al 1994, Hasen¨ohrl et al 1997)and is particularly reduced in aged patients with Alzheimer’s disease (Mufson et al1997). Expression of both TrkA and p75NTR in forebrain neurons is upregulatedby NGF (Gage et al 1989, Holtzman et al 1992). Adult cholinergic neurons ofthe extended striatal complex (caudate, putamen, accumbens, etc) express onlyTrkA; however, p75NTR is upregulated to detectable levels, and TrkA expressionis increased by local tissue injury or NGF infusions (Gage et al 1989, Holtzmanet al 1995). Adult neurons that express TrkA, but not p75NTR, are found in thethalamic paraventricular nuclei, rostral and intermediate subnuclei of the interpe-duncular nucleus, and various other brain regions (Holtzman et al 1995, Veneroet al 1994), and also in the spinal cord in regions associated with regulation of the
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1222 SOFRONIEW ¥ HOWE ¥ MOBLEY
autonomic outflow (Michael et al 1997). TrkA mRNA has been detected in CNSregions where its cellular localization is yet to be established. Some hippocampalpyramidal neurons may also express very low levels of TrkA (Cellerino 1995), anda recent immunocytochemical study points to the presence of TrkA and p75NTR
proteins in pyramidal cells of the somatosensory cortex of the mature rat (Pitts &Miller 2000). If confirmed, these results would contribute significantly to our un-derstanding of NGF production and actions in the CNS. As detection methodsincrease in sensitivity, it is likely that other NGF receptor-expressing neurons willbe identified in the CNS.
Among glial cells, light microscopic studies show that CNS astrocytes in vivorarely stain for p75NTR (P Belichenko & WC Mobley, unpublished observations).However, as many as one fifth of astrocytes in the dentate gyrus were immunore-active for p75NTR in a recent immuno-EM study (Dougherty & Milner 1999).This result suggests that very low levels of p75NTR are present in many matureastrocytes. p75NTRand, more controversially, TrkA are also expressed by astrocytesin vitro, particularly after exposure to NGF or inflammatory cytokines (Hutton et al1992, Hutton & Perez-Polo 1995, Kumar et al 1993, Semkova & Krieglstein 1999).A detailed analysis of NGF receptor expression by reactive astrocytes after CNSinjury would provide information for detailing the actions of neurotrophins in theCNS. Astrocytes are not alone in expressing NGF receptors. Oligodendrocytesexpress p75NTR (Casaccia-Bonnefil et al 1996, Kumar et al 1993). Microglia havethe capacity to express p75NTR and TrkA, and expression levels are modulated byinflammatory stimuli, such as cytokines and bacterial lipopolysaccharide (Elkabeset al 1998). The diversity of NGF receptor expression in the CNS is at least asgreat as that in the PNS and suggests that NGF signaling mediates many differentfunctions.
NGF-Producing Cells NGF is produced in the CNS during development andthroughout adult life. NGF-producing cells are present in the cortical target regionsof basal forebrain cholinergic neurons. Most such cells are neurons, including pyra-midal neurons, though glial cells are occasionally found to contain NGF (Pitts &Miller 2000). In the hippocampal formation, pyramidal and dentate granule neu-rons express NGF, as do subpopulations of GABAergic interneurons (French et al1999, Gall & Isackson 1989, Pascual et al 1998). These neurons also serve astargets of cholinergic innervation. In striatum, NGF is produced by a subpopula-tion of small interneurons (Bizon et al 1999). NGF expression in hippocampus isregulated by neuronal activity; increases are caused by glutamatergic and choliner-gic neurotransmission, and decreases are caused by GABAergic neurotransmission(Berzaghi et al 1993, Knipper et al 1994, French et al 1999). Neuronal NGF ex-pression in vivo is markedly upregulated by seizures, forebrain ischemia, markedhypoglycemia, and tissue injury (Gall & Isackson 1989, Lindvall et al 1994, Zafraet al 1991). Studies in vivo and in vitro indicate that cerebral insults influence NGFgene expression via excitatory amino acid neurotransmission as well as throughother pathways (Lindvall et al 1994).
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1223
Among glial cells, NGF is produced throughout the CNS by astrocytes andmicroglia, and NGF expression in both cell types is markedly upregulated bylocal tissue injury, inflammation, cytokines, and bacterial lipopolysaccharide (bothin vivo and in vitro) (Arendt et al 1995, Elkabes et al 1996, Heese et al 1998,Micera et al 1998, Yoshida & Gage 1992). In astrocytes, NGF expression is alsoupregulated by fibroblast growth factor, interleukin-1, glutamate agonists, reactiveoxygen species, high potassium, ischemia, and traumatic brain injury (Abiru et al1998; Friedman et al 1996; Goss et al 1998; Gottlieb & Matute 1999; Pechanet al 1992, 1993; Strauss et al 1968; Yoshida & Gage 1991). The data for NGFexpression in the uninjured brain are largely consistent with a role for NGF intarget-derived trophic support. Increased NGF levels in the injured CNS suggestthat astrocytes and microglial cells could serve as local sources of NGF for injuredneurons and other NGF responsive cell types.
Immune and Inflammatory System
In recent years, a great deal of interest has focused on NGF and NGF receptorgene expression in cells of the immune and inflammatory system. Several typesof bone marrow-derived leukocytes have the capacity to express TrkA, includingmast cells, CD4+ T lymphocytes, B lymphocytes, monocytes, and macrophages;follicular dendritic cells and B lymphocytes express p75NTR (Labouyrie et al 1997,Levi-Montalcini et al 1996, Torcia et al 1996). Many of the same types of leukocytesalso have the capacity to express NGF. These include mast cells, monocytes andmacrophages, T lymphocytes (CD3+ and CD4+ T cells), and B lymphocytes(Lambiase et al 1997, Leon et al 1994, Levi-Montalcini et al 1996, Mizuma et al1999, Torcia et al 1996). Both NGF and NGF receptor expression are dynamicallyregulated in leukocytes such that expression is increased by inflammatory andother stimuli as well as in activated cells (Barouch et al 2000, Lambiase et al 1997,Levi-Montalcini et al 1996, Mizuma et al 1999, Torcia et al 1996). A previouslyunexpected role for NGF in immune and inflammatory functions is suggested bythese findings.
NGF SIGNALING MECHANISMS
Cellular responses to NGF are elicited through binding and activation of its re-ceptors, TrkA and p75NTR (Bothwell 1995). NGF signaling is now recognizedas being broad based, dynamically regulated, and context dependent. Numerousintracellular signaling cascades are triggered by NGF receptor activation, and thereis evidence for convergence of, and direct interactions between, NGF signaling andsignaling triggered by other molecules. Studies on the intracellular signaling cas-cades triggered by NGF have relied heavily on in vitro models using primary cellcultures or cell lines, in particular the rat pheochromocytoma cell line PC12. Infact, many of the signaling cascades discussed in the following section have only
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1224 SOFRONIEW ¥ HOWE ¥ MOBLEY
been delineated in PC12 cells. However, the insights gained from analysis of suchcell culture models are useful in the context of an instructive role for further inves-tigation of NGF signaling within neurons and other neural cells. Likewise, studiesof NGF signaling have largely focused on developmental processes, such as neu-ronal differentiation and neurite outgrowth, but information about NGF signalingmechanisms in other contexts, such as degeneration, death, and neuroprotection isincreasingly available.
NGF Signaling Through TrkA
TrkA Activation TrkA, a single transmembrane-spanning polypeptide chainmember of the receptor tyrosine kinase (RTK) superfamily, was initially dis-covered as an oncogenic fusion protein isolated from human colon carcinoma(Martin-Zanca et al 1986a,b). Genetic analysis revealed that in normal cells theproto-oncogene encoded a 140-kDa glycosylated protein containing an extracel-lular region comprised of several immunoglobulin-like binding domains, a short,single transmembrane domain, and an intracellular domain encoding a tyrosinekinase (Martin-Zanca et al 1989). Following its initial discovery in 1986, the re-ceptor remained an “orphan receptor” until 1991, when it was discovered that NGFevoked a rapid tyrosine phosphorylation of endogenous TrkA in PC12 cells andof exogenous TrkA in transfected fibroblasts (Kaplan et al 1991a,b; Klein et al1991). Furthermore, TrkA was found to elicit signaling cascades necessary for thebiological responses of PC12 cells and neurons to NGF. Upon binding of NGF toTrkA, the receptor is subjected to a series of events that characterize RTK signal-ing. These include receptor dimerization and transphosphorylation of activationloop tyrosines leading to activation of kinase activity, followed by autophosphory-lation of tyrosines outside of the activation loop (Cunningham et al 1997). Theseautophosphorylation sites serve as binding sites for specific signaling proteins andadaptors such as PLCγ and Shc. Subsequent phosphorylation and activation ofaccessory proteins lead to the generation of a cascade of receptor-independentsignaling pathways (Greene & Kaplan 1995).
Ras Pathway Tyrosines 490 and 785 are two autophosphorylation targets thatare transphosphorylated following TrkA kinase activation (Loeb et al 1994,Middlemas et al 1994, Stephens et al 1994). Shc, an adaptor protein that is critical toactivation of the Ras signaling cascade (Figure 1) binds to phosphorylated tyrosine490 (Basu et al 1994, Obermeier et al 1994). Following binding and phosphoryla-tion of Shc, the Grb2-Sos complex binds to phospho-Shc via an SH2 interaction(Rozakis-Adcock et al 1992), thereby bringing Sos into proximity to membrane-associated Ras and activating the MAP kinase signaling cascade. Sos is a RasGTP exchange factor that promotes the transition from inactive Ras-GDP to activeRas-GTP (McCormick 1994). Ras is targeted to the plasma membrane via farne-sylation (Casey 1995) and resides at the plasma membrane in an inactive,GDP-bound state. Upon recruitment of Sos to the membrane, Ras is activated by
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1225
exchange of GDP for GTP (McCormick 1994). Ras then recruits the serine-threonine kinase C-Raf to the plasma membrane (Marshall 1994, Van Aelst et al1993, Wood et al 1992). In PC12 cells, Raf family members (Jaiswal et al 1994,Oshima et al 1991, Traverse & Cohen 1994) mediate NGF signaling by phos-phorylating and thereby activating the dual-specificity MAP kinase kinase MEK1at serine 217 and serine 221 (Jaiswal et al 1994, Lange-Carter & Johnson 1994,Vaillancourt et al 1994). MEK1 activation leads to the phosphorylation of twomembers of the MAP kinase family, extracellular signal-related kinases 1 and 2(Erk 1/2) (Crews et al 1992, Crews & Erikson 1992). Erk1/2 are phosphorylated onthreonine 202 and tyrosine 204 by MEK1 (Payne et al 1991), leading to activationand translocation of Erk1/2 into the nucleus (Chen et al 1992). Erk1/2 are proline-directed serine-threonine kinases that phosphorylate several substrates, includingElk-1 (Miranti et al 1995). Phosphorylation of Elk-1 at serine 383 and serine389 stimulates its interaction with the transcription factor serum response factor(SRF) and with the CAGGAT binding site of the serum response element (SRE)within thec-fosgene (Gille et al 1995, Hill et al 1993, Mueller & Nordheim 1991,Treisman 1992).c-fos is an immediate early gene that is rapidly transcribed inresponse to many extracellular stimuli, including NGF, and is an early componentof a series of transcriptional events necessary for initiation and maintenance ofdifferentiation (Ginty et al 1994, Greenberg et al 1986, Sheng & Greenberg 1990).
Additional transcription factors contribute to the regulation of c-fos transcrip-tion in response to NGF signaling. The cAMP regulatory element binding protein(CREB) is a transcription factor that binds to a site called the CRE, or cAMPresponse element, within thec-fos promoter (Berkowitz et al 1989). NGF sig-naling leads to the phosphorylation of CREB at serine 133 via a Ras-dependentmechanism (Ginty et al 1994). This allows CREB to interact with SRF and Elk-1(Bonni et al 1995, Ramirez et al 1997), possibly via the transcriptional coactivatorprotein CREB binding protein (CBP), which binds to phosphorylated serine 133in CREB (Chrivia et al 1993). CBP also binds to SRF (Ramirez et al 1997) andElk-1 family members (Janknecht et al 1993). CREB may also play an impor-tant role in transcriptional regulation of several NGF-specific delayed responsegenes, including theVGF gene. Mutation of the CREB binding site within theVGF gene significantly reduced NGF-inducedVGF transcription (Hawley et al1992). It is interesting thatVGF transcription may require the cooperation ofCREB with an as yet unidentified transcription factor product of an immediateearly gene. CREB is persistantly phosphorylated at serine 133 for several hoursafter an initial NGF stimulus, and this may permit accumulated immediate earlygene proteins to interact with activated CREB. In contrast, EGF stimulation, whichdoes not lead toVGF transcription, only transiently phosphorylates CREB, suchthat by the time sufficient immediate early gene product is present, activated CREBmay no longer be available to cooperatively stimulateVGF transcription (Bonniet al 1995). This may be one mechanism by which NGF and EGF activate differenttranscriptional programs leading to either differentiation or proliferation (Marshall1994).
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.
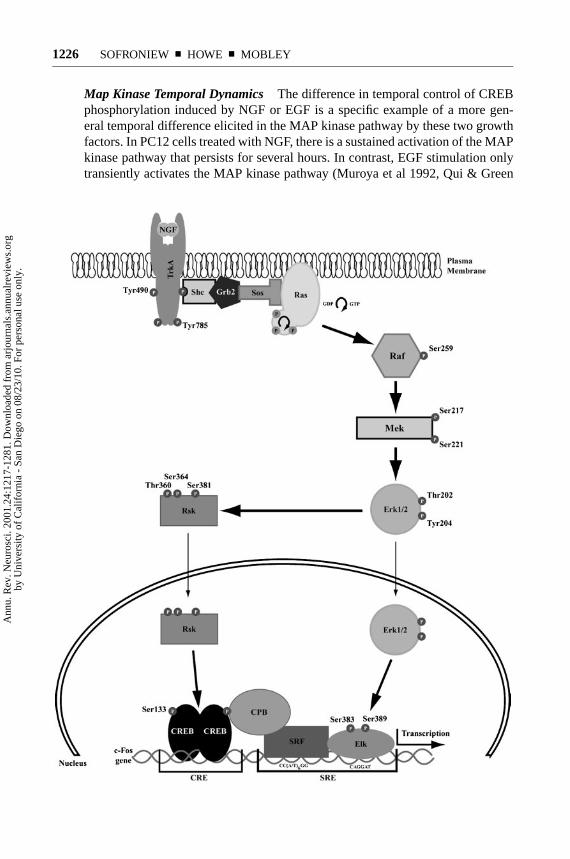
P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1226 SOFRONIEW ¥ HOWE ¥ MOBLEY
Map Kinase Temporal Dynamics The difference in temporal control of CREBphosphorylation induced by NGF or EGF is a specific example of a more gen-eral temporal difference elicited in the MAP kinase pathway by these two growthfactors. In PC12 cells treated with NGF, there is a sustained activation of the MAPkinase pathway that persists for several hours. In contrast, EGF stimulation onlytransiently activates the MAP kinase pathway (Muroya et al 1992, Qui & Green
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1227
1992, Traverse et al 1992), suggesting that the temporal dynamics of Erk1/2 ac-tivation may account for a differentiative versus proliferative signaling outcome.One explanation for how two RTKs linked to very similar signaling pathwaysmight induce such very different MAP kinase activation kinetics requires a bet-ter understanding of the specific isoforms of certain adaptor proteins utilized inthese cascades. For example, while both NGF and EGF appear to utilize the clas-sic Shc/Grb2/Sos/Ras/C-Raf/MEK pathway to activate Erk, NGF also utilizes anaccessory route to Erk activation that utilizes Gab2/CrkL/C3G/Rap1/B-Raf/MEK(Figure 1). This second pathway, which may be unique to NGF signaling, pro-motes sustained activation of Erk1/2 (CB Wu, CF Lai, WC Mobley, submittedfor publication; York et al 1998). The persistant Erk activation that follows NGFstimulation of the Rap1 pathway may induce expression of immediate early geneproteins that interact with activated CREB, induce transcription of novel delayedresponse genes, or both. Rap1 signaling through MAP kinase does not regulateall aspects of differentiation, nor can one exclude a role for Ras. Expression ofa mutant Rap that blocks sustained Erk activation in response to NGF does notblock neurite outgrowth in PC12 cells (York et al 1998). On the other hand, com-plete inhibition of Erk activation, either by pharmacological inhibition of MEK ortransfection with a dominant-interfering MEK mutant, does block NGF-inducedneurite outgrowth (Cowley et al 1994, Pang et al 1995), and inhibition of Ras ac-tivity by microinjection of a Ras-neutralizing antibody also blocks differentiation(Hagag et al 1986). Thus, Ras-dependent signaling is apparently important forNGF-induced differentiation. It is likely that some early event triggered by a Ras-and C-Raf-mediated activation of the Erk pathway is necessary for priming thecell to respond to the later and sustained activation of Erk by the Rap1 and B-Rafpathway.
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 1 The Ras-MAP kinase cascade downstream from TrkA. Following phosphorylationof tyrosine 490 within TrkA, Shc is recruited to the receptor via either an SH2- or phosphotyrosine-binding domain-based interaction. Consequently, Shc is bound by the Grb2-Sos complex. Recruit-ment of Sos to the membrane brings it into proximity of Ras, where it functions as a GTP-exchangefactor, activating Ras. Activated Ras recruits and activates Raf. Raf is a serine-threonine kinasethat phosphorylates the MAP kinase kinase MEK on 2 serines. This phosphorylation event initiatesactivity of the dual-specificity kinase, leading to activation of the MAP kinases Erk1/2 via phos-phorylation of threonine 202 and tyrosine 204. Phosphorylated Erk1/2 then participate in at leasttwo cascades. Erk1/2 may translocate into the nucleus, where they phosphorylate the transcriptionfactor Elk-1, or they may phosphorylate the kinase Rsk. Phosphorylation of Elk-1 allows it tointeract with the accessory transcription factor SRF, after which it binds to the serum responseelement (SRE) within thec-fospromoter region and contributes to initiation of transcription. Phos-phorylation of Rsk leads to its nuclear translocation and consequent phosphorylation of CREB onserine 133. Phosphorylated CREB is bound by the transcriptional coactivator protein CPB, whichalso binds to the SRF-Elk complex, creating an extended transcriptional factor complex that leadsto c-fostranscription.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1228 SOFRONIEW ¥ HOWE ¥ MOBLEY
Rsk Pathway A further level of control of NGF-induced immediate early genetranscription and translation comes from parallel activation of the Rsk pathwaydownstream from Ras. The Rsk serine-threonine kinase was originally isolated asa 90-kDa cell-cycle regulated kinase that phosphorylated the S6 protein of the 40Sribosomal subunit (Erikson & Maller 1991, Erikson et al 1991). This p90 kinase(ribosomal S6 kinase, hence Rsk) was itself found to be regulated by serine-threonine phosphorylation, and Erk1/2 were subsequently identified as the kinasesresponsible for this regulatory phosphorylation (Sturgill et al 1988, Zhao et al1996). The Rsk family is comprised of Rsk1, Rsk2, and Rsk3, each showing uniquepatterns of tissue expression (Moller et al 1994, Zhao et al 1995). Rsk2 was iden-tified as a Ras-dependent protein kinase that phosphorylates CREB on serine 133(Ginty et al 1994, Xing et al 1996), thereby regulating its transcriptional activation.Rsk family members are also involved in phosphorylation of the estrogen receptor-α, IκBα/NFκB, and c-fos (Ghoda et al 1997, Joel et al 1998, Schouten et al 1997,Xing et al 1996). Rsks also bind to the transcriptional coactivator CBP (Nakajimaet al 1996) and phosphorylate several members of the ribosomal complex(Angenstein et al 1998). Sos, a substrate for Rsk, appears to be negatively reg-ulated by Rsk kinase activity, suggesting that Rsk activation downstream from ac-tivation of Erk1/2 may feed back to truncate Ras signaling (Douville & Downward1997). Recently, all three members of the Rsk family were found to be activatedby NGF in PC12 cells, and all were able to phosphorylate CREB at serine 133(Xing et al 1998). Hence, the Ras pathway is able to regulate c-fos induction byusing a parallel and cooperative pathway in which Erk phosphorylation of Elk-1converges upon Rsk phosphorylation of CREB (Xing et al 1996). Thus, the Erkpathway is marked by both divergent and convergent signaling, in which an earlydivergence at the level of Shc versus Gab2 can control the temporal dynamics ofErk activation, and convergence at the level of Elk-1 and CREB regulation of c-foscan control gene transcription and protein translation.
Src and PKC Pathways Convergence of control over the MAP kinase pathwaymay also occur between Ras, PKC, and Src. Src is a member of a large familyof nonreceptor protein tyrosine kinases that share significant sequence homology.This family includes Fyn, Yes, Yrk, Blk, Fgr, Hck, Lck, Lyn, Frk/Rak, and Iyk/Bsk(Brown & Cooper 1996, Cance et al 1994, Lee et al 1994a, Thomas & Brugge1997, Thuveson et al 1995, Welch & Maridonneau-Parini 1997). Src kinases reg-ulate a wide range of cellular events, ranging from cell proliferation, cytoskeletalalterations, and differentiation, to survival, adhesion, and migration. RTKs interactwith Src kinases and use them to transduce several signaling pathways (Erpel &Courtneidge 1995). Involvement of Src or an Src family member in NGF-mediateddifferentiative signaling was first proposed when it was discovered that infectionof PC12 cells with the oncogenic form of Src recapitulated the neurite outgrowthinduced by NGF (Alema et al 1985). Further analysis showed that neutralizationof Ras by microinjection of anti-Ras antibodies blocked the neuritogenic effects ofboth Src and NGF (Hagag et al 1986, Kremer et al 1991). In contrast, neutralization
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1229
of Src activity by antibody microinjection did not block neurite outgrowth inducedby infection with oncogenic Ras (Bar-Sagi & Feramisco 1985, Kremer et al 1991,Noda et al 1985) but did inhibit NGF-induced neuritogenesis. It also caused re-traction of established neurites induced by NGF or FGF treatment (Kremer et al1991). Finally, both oncogenic Src and oncogenic Ras are able to “prime” PC12cells, such that subsequent NGF treatment elicits a more rapid and robust neurito-genesis than NGF treatment of unprimed cells (Thomas et al 1991). It is interestingthat oncogenic Src activated the N-terminal c-jun kinase (JNK), a member of theMAP kinase family, without activating Erk1/2 (Kuo et al 1997). Hence, one pos-sible explanation for the role that both Src and Ras play in differentiation is thatthey control the activity of a common MEK family member that is upstream ofboth Erk1/2 and JNK (Ellinger-Ziegelbauer et al 1997, Lewis et al 1998). Thismodel is compatible with data showing that pharmacological inhibition of MEKin PC12 cells abrogated neurite outgrowth in response to NGF (Pang et al 1995).MEK activity is also regulated by several PKC isoforms (Berra et al 1993, 1995;Schonwasser et al 1998, van Dijk et al 1997), and overexpression of either PKCι orPKCζ resulted in enhanced NGF-induced neurite outgrowth and enhanced NGF-induced JNK activation (Wooten et al 1999), while inhibition of atypical PKCisoforms blocked NGF-induced activation of JNK (Wooten et al 1999). PI3 ki-nase is also implicated in signaling to JNK, as NGF-induced JNK activation wasimpaired by either wortmannin or LY294002, and overexpression of PI3 kinaseresulted in neurite outgrowth and JNK activation in the absence of Erk activation(Kobayashi et al 1997). Thus, a signaling cascade including Src, PI3 kinase, PKC,and JNK appears to be involved in neurite outgrowth and differentiative signalingand may either complement or parallel the Ras-Raf-MEK-Erk1/2 cascade.
Signaling through Src, PI3 kinase, PKC, and JNK may also play a role in cellsurvival signaling. Overexpression of either Src or PKCι enhanced PC12 cell sur-vival in serum-free conditions, and both increased the activation of the transcriptionfactor NFκB (Wooten et al 2000, 1999), apparently via JNK signaling. Moreover,inhibition of Src or atypical PKC isoforms promoted cell death (Seibenhener et al1999, Wooten et al 2000). Likewise, inhibition of PI3 kinase activity blocked cellsurvival and reduced NGF-induced NFκB activation (Wooten et al 2000). Thesefindings are compatible with data showing that activation of NFκB promotes cellsurvival and resistance to apoptosis, and that NGF induction of NFκB is primarilydependent on signaling through the JNK pathway (Wooten et al 2000). Thus, bothdifferentiative and survival signaling may be controlled in part by a signaling unitthat includes Src, PI3 kinase, and PKC.
PI3 Kinase Pathway PI3 kinase and Src are also implicated in survival signal-ing via the common substrate Akt, a serine-threonine kinase also known as proteinkinase B (PKB), or related to A and C protein kinase (RAC-PK). Akt is regulatedby growth factor and serum factor signaling through PI3 kinase (Alessi et al 1996;Andjelkovic et al 1996; Burgering & Coffer 1995; Franke et al 1995, 1997; Klippelet al 1997). PI3 kinase is a heterodimer composed of an 85-kDa regulatory subunit
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1230 SOFRONIEW ¥ HOWE ¥ MOBLEY
and a 110-kDa catalytic subunit. Activation of the kinase involves binding of theregulatory subunit either directly or via adaptors to activated RTKs. This interac-tion with the cytoplasmic domain of an RTK results in recruitment of the 110-kDacatalytic subunit to the plasma membrane, where it can interact with and phos-phorylate membrane phosphoinositides. Such phosphorylation results in the pro-duction of PI-3,4-P2 and PI-3,4,5-P3. Akt interacts with PI-3,4-P2 or PI-3,4,5-P3,and with the 3-phosphoinositide-dependent kinase (PDK1). PDK1 contains apleckstrin homology domain that binds PI-3,4-P2 or PI-3,4,5-P3, and this bind-ing is necessary to permit PDK1 to phosphorylate and activate Akt (Alessi et al1997a,b; Cohen et al 1997; Stephens et al 1998; Stokoe et al 1997). Hence, TrkAsignaling via PI3 kinase presumably signals to generate 3-phosphoinositides thatbind PDK1 and induce the activation of Akt. PDK1 phosphorylates the activa-tion loop of several other serine-threonine kinases, including certain isoforms ofPKC (Chou et al 1998, Le Good et al 1998), suggesting that PI3 kinase-mediatedgeneration of 3-phosphoinositides may also control differentiative or survival sig-naling via PKC activation.
Mediation of TrkA survival signaling by PI3 kinase is indicated by the resultsof experiments showing that two inhibitors of PI3 kinase activity, wortmannin andLY294002, induce apoptosis in PC12 cells and sympathetic neurons supportedby NGF (Crowder & Freeman 1998, Yao & Cooper 1995). The role of Akt inregulation of cell survival downstream from PI3 kinase is suggested by the factthat overexpression of Akt in primary cultures of cerebellar neurons or sympa-thetic neurons provides protection against death induced by serum withdrawal orinhibition of PI3 kinase, while expression of dominant-interfering forms of Aktblocked NGF-mediated survival (Crowder & Freeman 1998, Dudek et al 1997).The mechanism by which Akt mediates survival is unclear, though Akt has beenreported to bind and phosphorylate Bad, a member of the Bcl-2 family of proteins(Figure 2) (Datta et al 1997, del Peso et al 1997). Phosphorylation of Bad preventsit from binding the anti-apoptotic Bcl-2 family members Bcl-2 and Bcl-XL (Zhaet al 1996), shifting the cell to contain more Bcl-2 homodimers than Bcl-2/Baxheterodimers. The Bcl family is composed of two groups of proteins, one thatpromotes cell survival and includes Bcl-2 and Bcl-XL, and the other that promotescell death and includes Bad and Bax (Boise et al 1995, Kroemer 1997, Steller1995). The members of the Bcl family form homo- and heterodimers, and thebalance of each dimer within the cell is considered to regulate the maintenanceof survival or the induction of death. In the absence of phosphorylation of Badon serine 112 and serine 136, Bad signals to promote cell death, apparently byforming heterodimers with Bcl-XL. Formation of these heterodimers leads to thegeneration of Bax homodimers. Homodimerization of Bax induces its transloca-tion into mitochondria and insertion into the mitochondrial membrane (Gross et al1998). There it leads to altered mitochondrial membrane potential via ion chan-nel formation and to generation of cytotoxic reactive oxygen species (Xiang et al1996). In contrast, the phosphorylation of Bad promotes cell survival by inducingan interaction between Bad and the 14-3-3 protein. This interaction effectively
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.
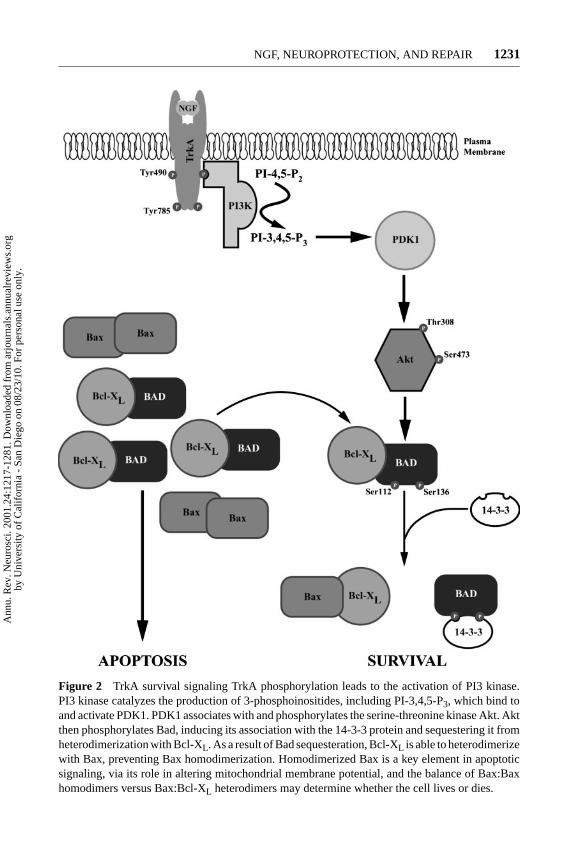
P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1231
Figure 2 TrkA survival signaling TrkA phosphorylation leads to the activation of PI3 kinase.PI3 kinase catalyzes the production of 3-phosphoinositides, including PI-3,4,5-P3, which bind toand activate PDK1. PDK1 associates with and phosphorylates the serine-threonine kinase Akt. Aktthen phosphorylates Bad, inducing its association with the 14-3-3 protein and sequestering it fromheterodimerization with Bcl-XL. As a result of Bad sequesteration, Bcl-XL is able to heterodimerizewith Bax, preventing Bax homodimerization. Homodimerized Bax is a key element in apoptoticsignaling, via its role in altering mitochondrial membrane potential, and the balance of Bax:Baxhomodimers versus Bax:Bcl-XL heterodimers may determine whether the cell lives or dies.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1232 SOFRONIEW ¥ HOWE ¥ MOBLEY
sequesters Bad from any interaction with Bcl-XL, keeping the balance of Bcl-XL/Bax heterodimers high and preventing Bax homodimerization (Zha et al 1996).Hence, TrkA survival signaling involves PI3 kinase-mediated activation of Akt andthe consequent maintenance of Bcl-XL/Bax heterodimers. Src is also implicated inthe activation of Akt via a mechanism that involves PI3 kinase and SHP-2 (Dattaet al 1996, Hakak et al 2000). This interaction may explain the finding, presentedabove, that inhibition of Src promotes cell death, and it suggests that additionalcomplexity may exist in the mechanism by which TrkA signaling induces cellsurvival.
TrkA activation may be linked to the phosphotidylinositol 3-kinase (PI3 kinase)pathway via binding of Grb2 and the Grb2-associated binder-1 (Gab1) protein totyrosine 490. Gab1 was initially identified as a Grb2-associated protein in a humanglial tumor expression library and was also identified in a yeast 2-hybrid screenusing the Met RTK as bait (Holgado-Madruga et al 1996, Weidner et al 1996).Gab1 is a member of a family of adaptor proteins that includes Gab2, IRS-1,IRS-2, and Dos, all of which exhibit sequence homology, and all of which linkplasma membrane RTKs to intracellular signaling cascades (Bausenwein et al2000, Gu et al 1998). Gab1 contains several SH2 and SH3 binding domains thatrecognize PI3 kinase and SHP-2, as well as Grb2, Nck, and Crk (Holgado-Madrugaet al 1996, Weidner et al 1996). Gab1 is tyrosine phosphorylated in response tosignaling downstream from TrkA (Holgado-Madruga et al 1997), and it is alsoinduced to associate with PI3 kinase, recruiting the p85 subunit to the plasmamembrane and eliciting activation. Furthermore, overexpression of Gab1 reducedthe concentration of NGF necessary for mediating cell survival in serum-freeconditions, while expression of a mutant Gab1 lacking the PI3 kinase bindingsites enhanced apoptosis (Holgado-Madruga et al 1997). These data suggest thatanti-apoptotic TrkA signaling to PI3 kinase and the Akt pathway is mediated byGab1. This is supported by the finding that adenovirus-mediated expression ofGab1 in PC12 cells is sufficient to support enhanced survival, even in the absenceof NGF signaling, and that this enhancement is correlated with increased PI3kinase signaling (Korhonen et al 1999). However, Gab1 appears to utilize boththe PI3 kinase pathway and the MAP kinase pathway to mediate its effect on cellsurvival, as pharmacological inhibition of both MEK and PI3 kinase was requiredto fully suppress Gab1-mediated cell survival (Korhonen et al 1999). Finally,adenovirus-expressed Gab1 enhanced neurite outgrowth in response to NGF viaa mechanism that was sensitive to either MEK inhibition or PI3 kinase inhibition(Korhonen et al 1999). These results suggest that Gab1 plays a role as an adaptorprotein for both the PI3 kinase pathway and the MAP kinase pathway downstreamfrom TrkA signaling. However, another member of the Gab family, Gab2, wasrecently identified as a substrate for tyrosine phosphorylation downstream of TrkA,and Gab2 was found in complex with CrkL, C3G, and SHP-2 following NGFtreatment of PC12 cells (CB Wu, CF Lai, WC Mobley, submitted for publication).This finding suggests that Gab2 may adapt TrkA to the Rap1/B-Raf pathway byinducing NGF-dependent activation of C3G, a Rap GTP exchange factor. In that
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1233
activation of the Rap1 pathway leads to MEK activation in parallel with the Raspathway, as described above, it is possible that overexpressed Gab1 subsumes therole of endogenous Gab2 in mediation of neurite outgrowth.
FRS-2 In addition to binding Shc and Gab, tyrosine 490 also appears to mediatethe interaction of TrkA with FRS-2, a novel membrane-anchored adaptor proteinthat is tyrosine phosphorylated in response to NGF (Kouhara et al 1997, Onget al 2000). Phosphorylated FRS-2 binds to the Grb2-Sos signaling unit, forminga multi-protein complex that includes Crk and the protein tyrosine phosphataseSHP-2 (Hadari et al 1998, Kouhara et al 1997, Meakin et al 1999). Formation ofthis complex is necessary for FRS-2 activation of the MAP kinase pathway. FRS-2competes with Shc for binding to tyrosine 490 on TrkA, adding an interestinglayer of complexity to the signaling cascades elicited by NGF treatment (Meakinet al 1999). FRS-2 may or may not be identical to SNT (Friedman & Greene 1999,Kouhara et al 1997), a protein that may be a candidate for the factor that controls thedecision between cell-cycle progression and cell-cycle arrest, a critical componentof differentiative signaling. The ability of SNT to bind the cyclin-dependent kinasesubstrate p13suc1, and the fact that it is rapidly tyrosine phosphorylated in responseto NGF (Rabin et al 1993) suggests that SNT may be the mediator of this keydecision. While the relationship between SNT and FRS-2 is still unresolved, recentevidence indicates that human FRS-2 does bind p13suc1in a constituitive manner(Meakin et al 1999), strengthening the possibility that FRS-2 is an SNT.
It is interesting to note that mutations in tyrosine 490 of TrkA do not abolishNGF induction of the MAP kinase signaling pathway. However, cells expressingTrkA with a double mutation at tyrosine 490 and tyrosine 785 do not exhibitMAP kinase activation or neurite outgrowth in response to NGF (Stephens et al1994). This finding suggests that there is an as yet undiscovered complexity orredundancy to the interaction of adaptor proteins with tyrosines 490 and 785. Onepossible component in this additional complexity is the recent finding that Grb2binds directly to activated TrkA at both tyrosine 785 and the kinase activation looptyrosines (MacDonald et al 2000). This additional route to the Ras pathway maycircumvent loss of either tyrosine 490 or tyrosine 785, but not both.
PLCγ and PKC Pathways Tyrosine 785, near the C terminus of TrkA, is withina consensus site for the binding of the SH2 domain of phospholipase C-γ (PLCγ ).This tyrosine is required for NGF-dependent recruitment of PLCγ to TrkA andfor the phosphorylation and activation of PLCγ (Vetter et al 1991). Followingbinding to tyrosine 785 of TrkA, PLCγ is activated and induced to hydrolyzephosphatidylinositol 4,5-bisphosphate (PI 4,5-P2). PLCγ -mediated hydrolysis ofPI 4,5-P2yields two products that each function as intracellular second messengers:inositol 1,4,5-P3 (IP3), which interacts with its specific receptor on the endoplasmicreticulum to induce the release of intracellular calcium, and diacylglycerol (DAG),which is a potent activator of protein kinase C (PKC) isoforms (Lee & Rhee 1995).IP3-mediated release from intracellular calcium stores leads to the activation of
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1234 SOFRONIEW ¥ HOWE ¥ MOBLEY
calcium-dependent proteins within the cell and to the generation of further IPderivatives such as IP4, IP5, and IP6, which are able to interact with other intra-cellular proteins (Menniti et al 1993). DAG is an activator of several isoforms ofthe serine-threonine calcium-dependent kinase PKC. These include several clas-sical, novel, and atypical PKC isoforms (Bell & Burns 1991; Nishizuka 1988;Liyanage et al 1992; Ono et al 1988; Osada et al 1990, 1992; Marais et al1998). DAG cooperates with calcium, phosphatidylserine, cis-unsaturated fattyacids, and lysophosphatidylcholine to activate the classical PKC isoforms, andit cooperates with phosphatidylserine and cis-unsaturated fatty acids to activatethe δ andε isoforms of novel PKC. PLCγ activation is often accompanied byphospholipase A2–mediated hydrolysis of phosphatidylcholine, directly generat-ing cis-unsaturated fatty acid and lysophosphatidylcholine (Asaoka et al 1992,Nishizuka 1992). These factors, in combination with DAG, serve to tune PKC ac-tivation to signaling downstream from TrkA, leading to phosphorylation of severalproteins critical to survival and differentiation (Coleman & Wooten 1994; Wootenet al 1994, 1997, 1999). One such substrate of PKC is Raf, which is directly acti-vated by PKC-mediated phosphorylation (Carroll & May 1994, Kolch et al 1993,Schonwasser et al 1998, Sozeri et al 1992, van Dijk et al 1997). The association ofPKC with Raf appears to be mediated by binding of the scaffolding protein 14-3-3(Freed et al 1994, Fu et al 1994, Irie et al 1994, van der Hoeven et al 2000). APKC-(14-3-3)-Raf complex may also contribute to PKCθ - and PKCµ-mediatedregulation of the MAP kinase cascade (Hausser et al 1999, Meller et al 1996) andmay account for PKCε-mediated activation of Raf (Cacace et al 1996, Ueffing et al1997). PKC might also mediate activation of the MAP kinase cascade by directlyactivating Ras, leading to the formation of a (Ras-GTP)-Raf complex (Marais et al1998). This finding is consistent with evidence that PKC-mediated activation ofRaf is blocked by mutation in the Ras-binding domain of Raf (Luo et al 1997).Finally, PKC can directly phosphorylate the c-jun protein product, which is alsounder the control of phosphorylation by Erk and which is able to bind to the c-fosprotein product to form the transcriptional regulatory complex AP-1 (Oberwetteret al 1993).
Abl Pathway The juxtamembrane region of TrkA, a unique region in the cyto-plasmic domain of the receptor, has also been implicated in carrying out severalspecific signaling functions downstream from NGF binding. This region appar-ently mediates the association of activated TrkA with Abl, a non–receptor tyrosinekinase that is involved in the regulation of adhesion-dependent signaling and cy-toskeletal remodeling that occurs during neuronal differentiation (Yano et al 2000).The association of Abl with TrkA may lead to its activation, and consequently to thephosphorylation of paxillin (Matsuda et al 1994, Ribon & Saltiel 1996, Teng et al1995, Torres & Bogenmann 1996). It is interesting to note that tyrosine phospho-rylation of paxillin is critical to the increased cell adhesion necessary for neuriteoutgrowth, and that Abl is involved in this pathway in Drosophila (Gertler et al1989, 1993; Wills et al 1999).
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1235
rAPS- and SH2-B-Mediated PathwaysTwo other adaptor proteins that do notappear to interact with either tyrosine 490 or tyrosine 785 are rAPS and SH2-B,which were recently identified as TrkA substrates in developing cortical and sym-pathetic neurons (Qian et al 1998). Both rAPS and SH2-B were found in complexwith Grb2, and either adaptor was able to mediate NGF induction of MAP kinaseactivation. In nnr5 PC12 cells that express extremely low levels of TrkA, cotrans-fection with rAPS and a TrkA mutant lacking all tyrosines except those in thekinase activation loop, or with SH2-B and this TrkA mutant, led to robust neuriteoutgrowth (Qian et al 1998). Moreover, while the interaction between rAPS andGrb2 is at least partially dependent on tyrosine phosphorylation of rAPS, Grb2appears to bind to SH2-B constituitively via an SH3 interaction. Finally, antibodiesto SH2-B inhibited NGF-dependent survival of cultured neonatal sympathetic neu-rons, and transfection with a dominant-interfering mutant of SH2-B completelyblocked the elaboration of axons by cultured sympathetic neurons. This suggeststhat SH2-B and rAPS are critical elements in the TrkA signaling pathway neces-sary for both neurite outgrowth and survival, but that their interaction with TrkAmay utilize a novel association mechanism.
NGF Signaling Through p75NTR
p75NTR was the first identified NGF receptor and for many years was believed tobe the only such receptor. However, following the discovery of a receptor tyrosinekinase for NGF that exhibited readily identifiable signaling properties, p75NTR
was largely relegated to the role of modulating and modifying TrkA signaling.While such a role continues to be an important area of investigation, it has becomeincreasingly clear that p75NTR is a signaling receptor in its own right. In fact, thesignals initiated by p75NTR are likely to be as complex as those for TrkA and tobe critically influenced by the cells in which such signaling arises (Friedman &Greene 1999, Kaplan & Miller 1997). The function of NGF signaling via p75NTR
in the context of cell death and regeneration may be important for understandingNGF actions in controlling the processes of neural repair and neuroprotection.
p75NTR is the first identified member of a superfamily of receptors that includesCD27, CD30, CD40, OX40, Fas (CD95), and the tumor necrosis factor receptors(TNF-R) (Bazan 1990, Cosman et al 1990, Mallett & Barclay 1991, Smith et al1994). These receptors share several common signaling features, including theability to control cell viability via regulation of apoptosis. For example, in theembryonic chick retina, neural precursor cells expressing p75NTR in the absenceof TrkA undergo NGF-dependent apoptosis, suggesting that developmentally pro-grammed death in these cells is mediated by p75NTR (Bredesen & Rabizadeh1997, Carter & Lewin 1997, Frade et al 1996). Furthermore, p75NTR mediatesNGF-induced death of cultured oligodendrocytes (Casaccia-Bonnefil et al 1996,Gu et al 1999, Yoon et al 1998) and cultured hepatic stellate cells (Trim et al2000), and BDNF signaling via p75NTR was shown to induce apoptosis of post-natal sympathetic neurons in culture (Bamji et al 1998). Moreover, an increased
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1236 SOFRONIEW ¥ HOWE ¥ MOBLEY
number of sympathetic neurons are found in BDNF-deficient mice, and there isa delay in sympathetic cell death in p75NTR homozygous knockout mice (Bamjiet al 1998). BDNF-dependent trigeminal neurons are killed via binding of NT-4to p75NTR, even though p75NTR is necessary to the cell survival induced by BDNF(Agerman et al 1999). This indicates that p75NTR signaling is not only dependenton cell context but also on neurotrophin binding specificity.
Ceramide Signaling One signal transduction pathway ascribed to p75NTR thatmay be involved in apoptotic signaling involves generation of the lipid secondmessenger ceramide via activation of sphingomyelinase. In fibroblasts expressingp75NTR but not TrkA, NGF induced the production of ceramide. Furthermore, inT9 glioma cells, NGF induced the activation of sphingomyelinase and the pro-duction of ceramide, and inhibited growth and fiber formation, a process that wasmimicked by incubation with membrane-permeant ceramide analogs (Dobrowskyet al 1994). Other members of the p75NTR superfamily, such as TNF-RI and Fas,also signal via ceramide production (Cifone et al 1994). This signaling functionappears to be mediated at least in part by a region within TNF-RI and Fas termedthe death domain, a C-terminal region in the cytoplasmic domain that is necessaryfor apoptotic signaling downstream from these receptors (Tartaglia et al 1993,Watanabe-Fukunaga et al 1992). Analysis of the p75NTR sequence shows that ahomologous death domain region exists within the intracellular region of this re-ceptor (Liepinsh et al 1997). Recent experiments suggest that the death domainserves to mediate protein:protein interactions. For example, this region mediatesFas and TNF-RI intracellular domain aggregation (Boldin et al 1995a, Song et al1994), and a homologous region has been found within ankyrin, a protein thatanchors transmembrane proteins to the cytoskeleton (Boldin et al 1995b).
Chopper Another death signaling domain was recently discovered within thep75NTR juxtamembrane region. This domain, a 29-residue sequence named chop-per, is necessary and sufficient to induce cell death in several cell types, includingneurons. It is interesting that a peptide corresponding to the chopper domain onlysignaled cell death when associated with the plasma membrane via a lipid anchor.Nonanchored chopper peptide did not mediate cell death and, in fact, acted ina dominant-negative manner to p75NTR-mediated death signaling (Coulson et al2000), suggesting that palmitoylation of p75NTR is a crucial factor in mediatingsignaling from the receptor. This finding also suggests the possibility that prote-olytic cleavage of the intracellular domain may play a role in controlling p75NTR
signaling.
Ligand-Independent p75NTRSignaling Another possible mechanism of p75NTR-mediated cell death was suggested by the observation that overexpression of theintracellular domain of p75NTR induced cell death in several neuronal populationswithin the central and peripheral nervous systems (Majdan et al 1997). This find-ing, plus the observation that immortalized neural cells overexpressing p75NTR
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1237
exhibit enhanced cell death following serum withdrawal (Rabizadeh et al 1993),suggests that p75NTR may signal pro-apoptotically in the absence of ligand bind-ing. In this model, binding of NGF to p75NTR induces a conformational changethat blocks the production of a death signal. Further support for this idea comesfrom work showing that antisense-induced downregulation of p75NTR in neonataldorsal root ganglia sensory neurons enhanced survival (Barrett & Bartlett. 1994).Moreover, identification of an alternatively spliced isoform of p75NTR lacking theneurotrophin-binding domain supports the model of ligand-independent signaling(Dechant & Barde 1997). The receptor produced by this alternative splice eventcontains the transmembrane and intracellular domains, but lacks the ability to bindneurotrophin and may therefore exhibit enhanced cell death signaling consistentwith the function of the death domains described above. Finally, p75NTR appearsto exhibit ligand-independent signaling through the RhoA pathway. In cells trans-fected with p75NTR, RhoA activation was generated in the absence of ligand andwas abolished by addition of ligand, suggesting that p75NTR can signal to reorga-nize the actin cytoskeleton in a manner that is negatively modulated by the presenceof neurotrophin (Yamashita et al 1999b).
NFκκB Pathway Many proteins in the p75NTR superfamily interact with TNFreceptor-associated factors (TRAFs) that modulate signaling through the JNK andNFκB pathways. Six such factors have been identified in signaling evoked by TNF-R, CD30, CD40, and the IL-1 receptor (Arch et al 1998, Rothe et al 1995), andrecently p75NTR was shown to associate with TRAF-2, TRAF-4, and TRAF-6 fol-lowing treatment with NGF (Khursigara et al 1999, Ye et al 1999). Interestingly, theassociation of TRAF-6 with p75NTR is mediated by the receptor’s juxtamembranedomain (Khursigara et al 1999) within a sequence that is absolutely conservedbetween human, rat, and chicken p75NTR (Large et al 1989), suggesting that theinteraction with TRAF-6 is critical to p75NTR function. TRAF-6 is recruited to theIL-1 receptor via binding to IRAK, the IL-1 receptor-associated serine-threoninekinase (Cao et al 1996a,b), and TRAF-6 also signals through NIK, the NFκB induc-ing kinase (Malinin et al 1997), suggesting that one role of the p75NTR-(TRAF-6)interaction may be to couple p75NTR to several different kinase cascades. Theuse of adaptor proteins such as TRAF-6 potentially permits p75NTR, which lacksany intrinsic kinase activity, to recruit and noncatalytically activate several cyto-plasmic non–receptor kinases, thereby linking NGF binding to p75NTR to NFκBactivation.
In addition to apoptosis-related signaling, p75NTR binding of NGF also activatesthe transcription factor NFκB in neuroblastoma cells (Korner et al 1994), culturedsensory and sympathetic neurons (Maggirwar et al 1998, Wood 1995), Schwanncells (Carter et al 1996, Khursigara et al 1999), and oligodendrocytes (Ladiwalaet al 1998, Yoon et al 1998). The activation of NFκB downstream from mostinducer proteins involves the degradation of the IκB protein, an inhibitory factorthat binds heterodimers of the NFκB p50 and p65 subunits and prevents themfrom translocating into the nucleus (Ghosh et al 1998). IκB degradation results
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1238 SOFRONIEW ¥ HOWE ¥ MOBLEY
in NFκB nuclear translocation and in upregulated transcription of several genes,including the IκB gene. In oligodendrocytes, in which p75NTR appears to signalvia both NFκB and the JNK pathway, expression of TrkA abrogates NGF-inducedcell death in a manner that correlates with cessation of JNK signaling, whereasthe NFκB signal downstream from p75NTR is unaffected (Yoon et al 1998). Thissuggests that p75NTR may evoke two separate pathways, one pro-apoptotic, theother anti-apoptotic. The balance of these two pathways, as modulated by TrkAsignaling in some cells, may control the ultimate fate of the cell. However, the exactrole that NFκB plays is unresolved—in some systems it exhibits anti-apoptoticsignaling (Maggirwar et al 1998, Mattson et al 1997), but in others it is associatedwith pro-apoptotic signaling (Schneider et al 1999, Schwaninger et al 1999). TheTNF receptor, generally associated with death signaling, also activates NFκB in apathway that appears to promote survival of lymphoid cells and fibroblasts (Liu et al1996, Van Antwerp et al 1996, Wang et al 1996). Likewise, in hippocampal neuronsthat do not express TrkA, NGF signaling through p75NTR protects these cells fromglucose deprivation-induced apoptosis (Cheng & Mattson 1991). Furthermore,p75NTR appears to play a role in protecting Schwann cells following axotomy. Inthe normal adult animal, Schwann cells do not express p75NTR. However, followingnerve injury, Schwann cells distal to the injury site dramatically upregulate p75NTR
expression (Heumann et al 1987b, Taniuchi et al 1986), and exhibit increased NFκBactivation (Gentry et al 2000). This increase in NFκB activation is correlated withthe absence of apoptosis in Schwann cells distal to the injury (Grinspan et al 1996).It is interesting that during development Schwann cells require axonal contact fortrophic support, and loss of such contact results in cell death. Hence, injury inducedexpression of p75NTR and consequent signaling through NFκB may serve in theadult to maintain Schwann cells in the absence of trophic support from the axon,thereby providing time for the axon to regrow.
Interactions Between p75NTR and TrkA
Prior to defining p75NTR signaling pathways, a great deal of attention was focusedon a role for p75NTR in regulating NGF signaling through TrkA. A wealth of datashow that p75NTR does impact TrkA signal transduction. It does so by enhancingbinding of NGF to TrkA, by increasing the specificity of TrkA for NGF binding,and through effects on TrkA signal transduction. Moreover, TrkA signaling alsoimpacts signaling through p75NTR. These efffects may be mediated, at least in part,through the direct association of TrkA and p75NTR, as revealed in studies usinga variety of techniques, including photobleaching (Wolf et al 1995), copatching(Ross et al 1996), cross-linking (Ross et al 1998), and coimmunoprecipitation(Bibel et al 1999, Gargano et al 1997, Huber & Chao 1995). Recently, TrkAand p75NTR were colocalized to caveolae-like domains of PC12 cells, and bothTrkA (Huang et al 1999) and p75NTR (Bilderback et al 1997) signal from thesemembranes. These findings highlight an important additional level of complexityfor NGF signaling and point to the need for understanding the cell biology ofreceptor trafficking and signaling.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1239
There are two classes of binding sites for NGF: Low-affinity receptors bindNGF with nanomolar affinity, whereas high-affinity receptors bind NGF with anaffinity that is 100-fold greater (i.e. 10−11 M) (Meakin & Shooter 1992, Sutter et al1979). The two classes are distinguished by the much slower rate of dissociationfrom high-affinity receptors (Landreth & Shooter 1980, Meakin & Shooter 1992,Schechter & Bothwell 1981, Woodruff & Neet 1986). High-affinity receptors arethought to play an important role in mediating NGF actions. Dissociation of NGFfrom TrkA is slow (Meakin et al 1992), which suggests that TrkA contributes to theformation of these receptors. TrkA is often referred to as the high-affinity receptor,a designation that suggests TrkA alone binds NGF with high affinity. However,although there is a small amount of high-affinity binding of NGF in cells expressingonly TrkA, most NGF binding to such cells is of low affinity (Mahadeo et al 1994).In fact, most high-affinity binding appears to reflect the interaction of p75NTR withTrkA. p75NTR has been shown to increase the rate of association of NGF withTrkA, thereby increasing the number of high-affinity receptors (Mahadeo et al1994). Moreover, p75NTR enhances activation of TrkA (Barker & Shooter 1994).Of note, a recent study showed that some receptor complexes from which NGFwas slowly released contained p75NTR (Huang et al 1999).
p75NTR may also interact with TrkA to modify binding specificity. In fibroblaststhat express only TrkA, NT-3 and NT-4/5 are able to activate the receptor, whereasin PC12 cells, which express both p75NTR and TrkA, only NGF is able to activateTrkA (Berkemeier et al 1991, Ip et al 1993). Likewise, mutant PC12 cells thatexpress only very low levels of p75NTR exhibit NT-3-induced TrkA activation(Benedetti et al 1993). Finally, postnatal sympathetic neurons normally exhibit verylimited survival in culture in response to NT-3, but these same neurons isolated fromp75NTR transgenic knockout mice show a much more robust NT-3-induced survivalresponse (Lee et al 1994c). These data suggest that p75NTR may function to tuneindividual neurons to specific neurotrophin responsiveness, thereby controlling theability of such neurons to compete for target-derived neurotrophic support. It isinteresting to note that sympathetic neurons normally undergo a switch in trophicdependence, from an early dependence upon NT-3 to a later dependence on NGF,and that this switch is temporally correlated to the onset of p75NTR expression(Birren et al 1993). Furthermore, as NGF signaling via TrkA appears to controlp75NTR expression in these cells (Miller et al 1991, 1994; Verdi & Anderson1994; Verge et al 1992; Wyatt et al 1990), it is possible that first contact be-tween the innervating sympathetic fibers and NGF available from the target fieldelicits the trophic dependency switch. Also, the expression of p75NTR by cells thathave received an NGF signal from the target may increase the sensitivity of thoseneurons to low levels of target-derived NGF, leading to a situation in which thoseneurons that express p75NTR are better able to compete for synaptic space withinthe target. Hence, the ability of p75NTR to sharpen TrkA binding specificity mayplay a significant role in the maturation of target innervation, and may control thecompetition that defines the adult pattern of innervation. Whether p75NTR playssuch a role in synaptic competition within the central nervous system remains tobe determined.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1240 SOFRONIEW ¥ HOWE ¥ MOBLEY
Perhaps the most interesting facet of the interaction of TrkA and p75NTR isevidence for reciprocal effects on signaling. Barker and colleagues provided ev-idence that signaling through p75NTR inhibits signaling though TrkA. Additionof BDNF to PC12 cells markedly reduced the TrkA activation and downstreamsignaling events that were elicited by treatment with an NGF mutant that onlybinds to TrkA (MacPhee & Barker 1997). BDNF is known to activate sphin-gomyelinase and increase ceramide levels (Dobrowsky et al 1995). Ceramideaddition produced changes similar to those seen with BDNF. It is interestingthat both BDNF and ceramide treatment were shown to increase phosphoserinecontent on the intracellular domain of TrkA (MacPhee & Barker 1997), whichsuggests that BDNF acts through p75NTR and ceramide production to influenceTrkA signaling. Completing the analysis of signaling interactions, there are ex-amples in which TrkA has a negative or restraining effect on p75NTR signaling.Although NGF effectively induced sphingomyelin hydrolysis in cells expressingp75NTR in the absence of TrkA, NGF did not do so in PC12 cells. This effectis apparently mediated by TrkA signaling because inhibition of TrkA signalingby the inhibitor K252a restored the ability of NGF to hydrolyze sphingomyelin(Dobrowsky et al 1995). In another example, when oligodendrocytes were trans-fected with TrkA, treatment with NGF induced activation of the MAPK pathway,suppressed JNK activity, and prevented cell death without influencing the activationof NFκB (Yoon et al 1998). These studies document the existence of robust, recip-rocal interactions between TrkA and p75NTR. An important goal is the elucidationof the molecular basis for these interactions and the definition of their physiologicalsignificance.
Signaling Endosomes
Internalization of the NGF-TrkA complex plays an important role in intracellularsignaling, particularly in neurons, where retrograde transport of the “signal” fromdistant axon terminals is required to trigger signaling in the cell body. Considerableevidence suggests that this internalization involves endocytosis and the formationof “signaling endosomes,” organelles in which NGF continues to be bound to itsactivated receptors (Grimes et al 1996, 1997; Riccio et al 1997; Tsui-Pierchala &Ginty 1999; Ure & Campenot 1997; Watson et al 1999). TrkA and p75NTR acti-vation appears to occur predominantly in caveolae-like membranes that containmany of the intermediates of their signaling pathways (Bilderback et al 1999,Huang et al 1999). It is possible that signaling endosomes are derived from thesemembranes, but it is noteworthy that at least some TrkA appears to be internalizedvia clathrin-coated membranes (CL Howe, JS Valletta, WC Mobley, submittedfor publication). In fact, in recent studies we have shown that NGF increased theassociation of clathrin with membranes and induced the formation of complexescontaining activated TrkA, clathrin heavy chain, and the plasma membrane spe-cific adaptor complex, AP2. Moreover, we discovered that clathrin-coated vesiclesisolated from NGF-treated cells contained NGF bound to activated TrkA, linking
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1241
formation of the TrkA—clathrin complex to endocytosis via clathrin-coated mem-branes. It is exciting to note that Shc was recruited to these membranes, as wasRas and activated Erk1/2. Importantly, we found that NGF-induced clathrin-coatedvesicles were able to signal in an in vitro kinase assay to propagate the NGF sig-nal from Erk to Elk. Our findings indicate that NGF signals from endosomes andthat clathrin-coated vesicles are one source of signaling endosomes produced inresponse to NGF treatment (CL Howe, JS Valletta, WC Mobley, submitted for pub-lication). In other recent work, we have also isolated additional membranes thatcontain the NGF signal, suggesting the existence of a variety of signaling endo-some species (CB Wu, CF Lai, WC Mobley, submitted for publication). Whetherp75NTR signals from endosomes is an interesting possibility that requires furtherstudy, though preliminary evidence suggests that p75NTR does utilize the clathrinpathway for internalization, hinting at the existence of clathrin-coated vesiclesthat contain p75NTR and p75NTR-associated signaling elements (CL Howe, APKruttgen, E Shooter, WC Mobley, unpublished observations). These findings areconsistent with the concept that NGF signaling initiates the endocytosis of special-ized membrane regions to form signaling endosomes that are enriched both in NGFreceptors and their downstream signaling second-messenger target molecules.Signaling endosomes may exist as signaling complexes in the cytoplasm thatcan be transported from the site of their formation along neurites to the cellbody.
Positive Feedback
There are several means by which positive feedback could be exerted in NGFsignaling loops. First, as described above, NGF upregulates the expression of itsown receptors. Second, NGF upregulates expression of such effector moleculesas acetylcholine (Mobley et al 1985) or substance P and related tachykinins(Lindsay & Harmar 1989), which on their release would be expected to upregulatethe expression of NGF by target tissues (Berzaghi et al 1993, French et al 1999,Woolf et al 1994). These changes may lead to reinforcement and strengthening ofNGF signaling in a positive feedback manner (Sofroniew & Mobley 1993). Thefunctional consequences of these effects are poorly understood. In nociceptive neu-rons, NGF upregulates the expression of neuropeptides such as substance P and cal-citonin gene-related peptide, and positive feedback in this system may facilitate theinduction of sensitization and hyperalgesia in response to tissue injury (Malcangioet al 1997; Mendell 1996; Verge et al 1995, 1996). In the forebrain, positive feed-back may represent a means of reinforcing heavily used cholinergic connections,with significant ramifications for learning- and memory-related plasticity (Howe &Mobley 2001).
Interactions Between NGF and Other Molecular Signaling
Glutamate Both NGF and glutamate signaling have well documented effectson the regulation of neuronal survival and neurite outgrowth during development.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1242 SOFRONIEW ¥ HOWE ¥ MOBLEY
Reports also exist of synergistic interactions between glutamate and NGF (Cohen-Cory et al 1991) or other neurotrophins (Morrison & Mason 1998) in sculptingneuronal survival or morphology during development. Interactions between glu-tamate and neurotrophin signaling that modulate neuronal excitablility persist inmature neurons (McAllister 1999). The intracellular signaling mechanisms in-volved in these effects are not yet known, and evidence for direct convergenceof intracellular signaling mechanisms has not yet been reported, but it appearspossible along several pathways, including modulation of cytoplasmic Ca2+ levels(Mattson 1996).
Estrogen Estrogen enhances neuronal growth and differentiation and regulatescytoskeletal and growth-associated gene expression. There is now evidence forboth colocalization of estrogen and NGF receptors in the same cells (Toran-Allerand et al 1992), and for direct convergence of NGF and estrogen signalingthrough the MAPK pathway (Singer et al 1999, Singh et al 1999). Such mechanismsmay contribute to the many modulatory effects of estrogen on neural function. Animportant unresolved issue is how estrogen signals to induce MAPK activation.
Intracellular Signals Much attention has focused on NGF signaling effects onintracellular Ca2+, and there is reason to believe that Ca2+ plays an important rolein many aspects of the biology of NGF and the other neurotrophins. In recent stud-ies using PC12 cells, and fibroblasts transfected with TrkA or p75NTR, NGF wasshown to signal through TrkA and p75NTR to cause acute and transient increasesin intracellular Ca2+ as a result of increased uptake through L-type Ca2+ channels(Jia et al 1999, Jiang et al 1999). TrkA activation also increased intracellular Ca2+
mobilization (Jiang et al 1999). A number of functions can be envisioned for theincreased intracellular Ca2+ that follows NGF treatment. The “Ca2+ set point hy-pothesis” posits that the level of cytoplasmic Ca2+ determines the degree of NGFsignaling required to suppress cell death mechanisms during development (John-son & Deckwerth 1993). Effects of NGF signaling on cytoplasmic Ca2+ levels alsorepresent a means for NGF to influence the plasticity and vulnerability of matureneurons (Mattson et al 1995). It is tempting to suggest that through acute changesin intracellular Ca2+, NGF could influence the behavior of synapses throughincreased release of neurotransmitters or of other neurotrophins (Berninger &Poo 1996, Kr¨uttgen et al 1998).
EFFECTS AND FUNCTIONS OF NGF
Given the many cell types expressing NGF and NGF receptors, and the diverseintracellular signaling cascades triggered by NGF, it is not surprising that NGFsignaling is implicated in many different functions during development and inadults. NGF mediates several types of intercellular communication and has beenshown to act as (a) a retrogradely transported, target-derived factor that influences
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1243
afferent neurons, (b) a locally released paracrine factor that affects both neuronsand nonneuronal cells, (c) an autocrine factor acting on the same cells that produceand release it, and (d) an endocrine factor that acts after transport through the bloodstream (Levi-Montalcini et al 1995, 1996).
NGF Roles in Development
NGF has a number of roles in the development of neuronal and nonneuronal cells.During development NGF promotes the survival and maturation of several pop-ulations of neurons that express TrkA and p75NTR: in the PNS, sympathetic andsensory neurons (Conover & Yancopoulos 1997, Johnson et al 1986, Lehmannet al 1999, Snider 1994) and in the CNS, basal forebrain, and striatal cholinergicneurons (Kew et al 1996, Li et al 1995, Svendsen et al 1994). For each of these pop-ulations, NGF participates in classical target-derived neurotrophic relationships.The importance of these relationships is evident from the results of experimentsin which the genes for NGF or the NGF receptors were disrupted. Knocking outthe NGF gene resulted in the loss of most small nociceptive dorsal root ganglion(DRG) neurons and sympathetic neurons in the PNS (Crowley et al 1994). In theCNS of animals heterozygous for disruption of the NGF gene, there was a clearreduction in the number of basal forebrain cholinergic neurons, atrophy of thesecells, and reduction in the cholinergic innervation of the hippocampus (Chen et al1997). Further study is needed to explore the effect of NGF gene disruption onstriatal cholinergic neurons. However, both basal forebrain and striatal cholinergicneurons are reduced in number and size in animals in which the gene for TrkA wasdisrupted (Fagan et al 1997). Also, there was marked depletion of both small DRGneurons and sympathetic neurons in TrkA knockout animals (Smeyne et al 1994).
Changes are also seen with p75NTR gene disruption in both the PNS and CNS.In the PNS, there are defects in sensory innervation of skin and in sympatheticinnervation of the pineal gland and sweat glands (Lee 1992, 1994b,c). Remarkably,disrupting the gene for p75NTR had little if any effect on the number of sympatheticneurons in animals in which both copies of the NGF gene were present (Brennanet al 1999). However, disrupting p75NTR increased the number of sympathetic neu-rons in animals in which the NGF gene was also disrupted (Brennan et al 1999).The mechanism for this surprising and interesting result is not yet established.However, one possible interpretation is that NGF signaling through p75NTR re-strains the normal survival and development of sympathetic neurons. Another isthat p75NTR gene disruption allows NT-3 to signal through TrkA to enhance neu-ron survival, a suggestion for which there is experimental evidence (Brennan et al1999). Further studies will be needed to clarify the interaction.
Paralleling the results for sympathetic neurons, p75NTR gene disruption appearsto increase the number and size of basal forebrain cholinergic neurons (Greferathet al 2000, Yeo et al 1997, but see Peterson et al 1999). While it is interestingto speculate that p75NTR signaling inhibits the normal development and functionof these neurons, the mechanistic basis for this is yet to be determined. Though
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1244 SOFRONIEW ¥ HOWE ¥ MOBLEY
additional studies are required to detail NGF effects in the developing nervoussystem, gene disruption studies have documented an important role for NGF inthe survival and differentiation of both PNS and CNS neurons.
Interesting additional themes have emerged that have considerably modifiedthe original neurotrophic hypothesis. First, CNS and PNS neurons are not con-tinuously dependent on the constitutive supply of a single target-derived factorthroughout life. Rather, a variety of different molecules from different sourcesinfluence developmental survival and maturation. For example, sensory neuronsare transiently dependent for survival on different neurotrophins at different timepoints as they progress through phases of development (Davies 1994). Second,transiently required growth factors may derive from sources other than the finaltarget region, such as local interactions around the cell bodies (Enokido et al 1999),or intermediate targets that axons encounter and then grow past en route to finaldestinations (Wang & Tessier-Lavigne 1999). Third, NGF signaling can also medi-ate axon sprouting, as well as growth cone turning and local guidance (Campenot1977, Gallo et al 1997, Patel et al 2000, Rice et al 1998, Tuttle & O’Leary 1998).For example, a recent study shows that NGF is critical for the elongation of the pe-ripheral but not the central processes of sensory neurons (Patel et al 2000). Fourth,NGF can also induce the death of certain developing neurons by signaling throughp75NTR in the absence of TrkA, as in the retina (Frade & Barde 1998, Frade et al1996). Regarding glia, NGF may regulate the development of oligodendrocytes,but here too, rather than promoting survival, NGF signaling via p75NTR can undercertain circumstances induce the death of these cells (Casaccia-Bonnefil et al 1996,Chao et al 1998, Gu et al 1999). Taken together, these findings point to the complexneurotrophic environment that guides the development of the nervous system. Arole for TrkA signaling in NGF actions during development is well established andappears to be the dominant theme in signaling events that are required for survivaland differentiation. Nevertheless, it is apparent that p75NTR has important roles toplay in modulation of TrkA signaling and may also independently regulate cellsurvival. The stage is now set for exploring the details of NGF signaling in thedeveloping nervous system.
NGF Roles in Adults
Both NGF and NGF receptors continue to be expressed and dynamically regulatedby many different cells types throughout adult life and aging. Information aboutdifferent NGF functions in adults is emerging for both NGF-responsive neuronsand nonneuronal cell types. Once mature, most neurons lose absolute dependenceon target-derived growth factors for acute survival. In adults, the focus of NGFsignaling shifts away from the regulation of neuronal survival to the regulationof neuronal phenotype and function. In cases well studied thus far, CNS andPNS neurons that are developmentally dependent upon NGF for survival becomeindependent of a constitutive supply of target-derived NGF for acute survival oncethey have established their connections and reached maturity. In adults, sensory
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1245
and CNS cholinergic neurons do not die for many months after NGF withdrawal(Johnson & Deckwerth 1993; Sofroniew et al 1990, 1993; Svendsen et al 1994).Adult sympathetic neurons also do not die acutely after immunologically inducedNGF withdrawal; instead, they undergo a gradual cell death of about 25% afterone month, which increases to about 40% after 3 months (Ruit et al 1990). Theresistance of these neurons to NGF withdrawal may be caused by loss of the c-fos induction that normally follows such withdrawal in young neurons and byconsequent interruption of a cascade that involves both c-fos and Bax (Easton et al1997). Although these mature NGF-responsive neurons become independent ofNGF for acute survival, they all undergo atrophic changes if subjected to NGFwithdrawal. These changes take the form of cell shrinkage (which is often severe)and reduced transmitter-related gene expression.
It is likely that TrkA signaling mechanisms are implicated in these changes,since NGF has been shown to increase cell size in neurons that do not expressp75NTR (Holtzman et al 1995). Because mature NGF receptor expressing neuronsdo not require a constitutive supply of NGF for acute survival, the possibility ex-ists that acute fluctuations in NGF signaling dynamically regulate various typesof activities in mature NGF-responsive cells. As discussed in this section, thesefunctions include modulation of the plasticity of NGF-responsive neurons. A par-ticularly well-documented and striking example of this is the regulation by NGFof mature nociceptive neurons (Woolf et al 1996).
The widespread production of NGF by glial cells and other nonneuronal cellsis leading to new ideas about other types of NGF functions, prominent amongwhich appear to be roles in inflammation and the response to injury in the CNS,PNS, and peripheral tissue. NGF appears likely to have other functions that arecurrently not well understood. For example, NGF infused into the lateral cerebralventricles induces hypophagia and weight loss in rats (Winkler et al 2000), andNGF treatment has been reported to affect appetite in patients in clinical trials(Petty et al 1994).
Plasticity of NGF-Responsive NeuronsThe adult nervous system exhibits aremarkable ability to alter both its structure and function in response to stimuli,a capacity commonly referred to as plasticity. Neurotrophins are implicated asmolecular mediators of specific forms of both structural and functional plasticity.NGF has thus far been associated in particular with effects on structural plastic-ity and has far fewer reported direct effects on neuronal activity in comparisonwith other neurotrophins, such as BDNF (McAllister 1999). Nevertheless, NGFhas reported effects on stimulus-dependent activity in adult somatosensory cortex(Cellerino & Maffei 1996, Gu et al 1994, Prakash et al 1996) that appear to be me-diated by TrkA and facilitated by p75NTR (Pizzorusso et al 1999). The mechanisticbasis for these effects is unclear but may involve NGF-dependent modulation ofcholinergic function and subsequent modification of cortical plasticity (Howe &Mobley 2001). In the PNS, NGF regulates the cell body size, terminal sprout-ing, dendritic arborization, and gene expression of sympathetic neurons and small
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1246 SOFRONIEW ¥ HOWE ¥ MOBLEY
nociceptive sensory neurons (Johnson et al 1986, Ruit et al 1990). In the CNS,NGF regulates the cell body size, dendritic arborization, terminal sprouting, andgene expression of basal forebrain and striatal cholinergic neurons (Cuello 1996,Debeir et al 1999, Howe & Mobley 2001). As discussed above, NGF increases theexpression of its own receptors, as well as of transmitters or transmitter-producingenzymes, and may do so to modulate cell function in a context-specific manner.It is interesting that in both the CNS and PNS, target-derived as well as locallyapplied NGF will exert these effects, and there are both target-derived and lo-cal sources of NGF-producing cells available to the neurons. The significance of,and interactions between, local and target-derived signaling for NGF-responsiveneurons are not yet understood.
Nociception Small nociceptive sensory neurons express both types of NGF re-ceptor throughout life, and NGF has a variety of effects on these cells, includingupregulation of TrkA and p75NTR, CGRP (calcitonin gene related peptide), andtachykinin expression, as well as modulation of cell size, activity, and neuropep-tide release (Malcangio et al 1997; Mendell 1996; Verge et al 1995, 1996). NGFsignaling in these cells leads to hypersensitivity to nociceptive stimuli in the formof allodynia and hyperalgesia in both animals and patients given NGF in clinicaltrials for peripheral neuropathies (Petty et al 1994, Shu & Mendell 1999). NGFis expressed and released in many tissues in response to injury, and blockade ofNGF signaling using function blocking antibodies in experimental animals withskin injury and inflammation prevents the development of hyperalgesia (McMahonet al 1995). NGF elicits both mechanical and thermal hyperalgesia. With respectto the latter, NGF acts as a peripheral sensitizing agent to alter the response ofnociceptors to noxious stimuli. It may accomplish this effect in part by inducingmast cell degranulation and the consequent release of serotonin, histamine, andNGF itself (Shu & Mendell 1999). In terms of mechanical hyperalgesia, NGFmay act centrally by upregulating CGRP, substance P, and BDNF (Shu & Mendell1999). Recent findings suggest that PKC may mediate the actions of NGF on pe-ripheral nociception (Khasar et al 1999). Thus, NGF plays an important role inthe nociceptive response that follows tissue injury by inducing both peripheral andcentral sensitization through signaling that involves several cell types, includingleukocytes and nociceptive sensory and sympathetic neurons (Mendell et al 1999,Shu & Mendell 1999, Woolf et al 1996). NGF may also be an important mediatorin pain due to visceral inflammation (Dmitrieva et al 1997) and in neuropathic painsyndromes induced by peripheral nerve irritation.
Immune and Inflammatory System NGF has numerous effects on immune andinflammatory cells that are generally directed at inducing their state ofactivation and effector functions (Levi-Montalcini et al 1996, Otten et al 1994,Simone et al 1999). NGF increases mast cell number, induces mast cell degran-ulation, and increases mast cell expression of cyclooxygenase and interleukin-6(Marshall et al 1999, Simone et al 1999). NGF activates monocytes, macrophages,
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1247
and CNS microglia by increasing their phagocytic activity and by inducing theirexpression of interleukin-1, Fcγ receptor, and lysosomal proteases of the cathepsinfamily (Liuzzo et al 1999; Susaki et al 1996, 1998). NGF is chemotactic for neu-trophils (Gee et al 1983). NGF influences T and B lymphocyte proliferation, is anautocrine survival factor for B lymphocytes, and stimulates immunoglobulin pro-duction (Levi-Montalcini et al 1996, Otten et al 1994, Torcia et al 1996). Throughits interactions with immune and inflammatory cells, as well as with glia and neu-rons, NGF has been suggested to play a role in various diseases with postulatedautoimmune and inflammatory components, including arthritis and demyelinatingdiseases (Aloe 1998, Bonini et al 1999, Levi-Montalcini et al 1996).
NGF AND NEUROPROTECTION
Substantial evidence suggests that among other functions, NGF acts to protect neu-rons from endogenous toxic events generated during the response to tissue injuryand that NGF signaling facilitates regrowth and repair. The signaling mechanismsengaged in neuroprotection have not been defined. However, it appears that theprotective effects of NGF extend both to neurons known to express NGF receptorsand to those that are not known to express such receptors.
NGF Protection of Neurons Known to Express NGF Receptors
Protection from Axotomy Throughout the PNS and CNS, adult neurons varyconsiderably in their vulnerability to axotomy. For unknown reasons, some axo-tomized neurons survive with few obvious changes, others survive but atrophy tomoderate or severe degrees, and others die either rapidly or over a prolonged timecourse (Sofroniew 1999). Among NGF-responsive neurons, sensory neurons andcholinergic neurons in the basal nucleus survive axotomy but exhibit moderate-to-severe atrophy (Sofroniew et al 1983, Verge et al 1996). In contrast, about 50%(but not all) of axotomized sympathetic neurons and septal cholinergic neuronsrapidly die (O’Brien et al 1990, Ruit et al 1990, Tuszynski et al 1990). Neither themechanism of axotomy-induced cell death, nor the reasons that some neurons diewhile others survive but atrophy, are understood. In the adult septum, axotomy-induced cell death is not due to loss of NGF signaling because target lesion andNGF-depletion studies show that these neurons are not acutely dependent on NGFfor survival (Kordower et al 1993; Sofroniew et al 1990, 1993). Nevertheless, thedeath of these neurons can be largely prevented in both rodents and primates byNGF infusions at the time of the axotomy (Hefti 1986, Tuszynski & Gage 1995b,Williams et al 1986). In the septum, this effect appears to be mediated via TrkAsignaling (Lucidi-Phillipi et al 1996), and NGF need only be given transiently fora few weeks after the axotomy and can then be discontinued without subsequentloss of neurons (Tuszynski & Gage 1995a). In addition, NGF is able to preventaxotomy-induced atrophy in cells that are not killed by axotomy, as well as to
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1248 SOFRONIEW ¥ HOWE ¥ MOBLEY
reverse atrophy that has already occurred (Cuello 1994). The intracellular path-ways through which NGF signaling protects TrkA and p75NTR expressing neuronsfrom axotomy-induced death are not defined.
Protection from Glutamate Excitotoxicity Aloe (1987) first reported a protec-tive effect of NGF against glutamate receptor–mediated excitotoxicity, such thatNGF infusions reduced the overall size of excitotoxic lesions in the striatum andprevented the death of cholinergic, NGF receptor–expressing, striatal neurons.These observations have been confirmed by others using local infusions of NGFor grafts of cells genetically modified to secrete NGF (Davies & Beardsall 1992,Frim et al 1993, Martinex-Serrano & Bjorklund 1996, Schumacher et al 1991).NGF has also been reported to protect PC12 cells from anoxia and glucose depriva-tion, or from nitric oxide cytotoxicity (Boniece & Wagner 1993, Wada et al 1996).To examine the potential role of endogenous NGF signaling in the protection ofNGF-responsive neurons from excitotoxicity, we recently studied septal choliner-gic neurons that express NGF receptors and project to hippocampus and are thus,in contrast to striatal cholinergic neurons, well separated from their target cells thatproduce NGF. Both local and target-derived (i.e. retrogradely transported) NGFsignaling significantly attenuated glutamate receptor–mediated excitotoxic deathof these neurons in young adult rats. In addition, aged rats that have a reducedcapacity to retrogradely transport NGF, and young adult rats given target lesionsthat deplete access to retrogradely derived NGF, both exhibited (a) significantlyincreased vulnerability of cholinergic neurons to glutamate receptor–mediatedtoxicity, (b) significantly reduced protective effects of local NGF, and (c) signifi-cantly reduced levels of TrkA. Chronic intracerebroventricular NGF significantlyrestored TrkA and the protective effect of local NGF (Horner et al 1999; HHDLam, CH Horner, J Berke, JD Cooper, RE Brown, SB Dunnett, MV Sofroniew,submitted for publication). These findings suggest that in adult forebrain, signal-ing through TrkA serves ongoing neuroprotective functions. They also suggest thatloss of endogenous NGF signaling leads to increased vulnerability of these cells toexcitotoxicity. However, the intracellular pathways through which NGF signalingprotects TrkA and p75NTR expressing neurons from glutamate receptor–mediateddeath are not yet defined.
NGF Protection of CNS Neurons that donot Appear to Express NGF Receptors
Although readily detectable expression of TrkA and p75NTR is confined to rela-tively few populations of neurons in both the PNS and CNS, NGF is reported toprotect a broad spectrum of neurons from ischemia, glutamate receptor–mediatedexcitotoxicity, and metabolic insults such as glucose deprivation and oxidativestress. Protective effects of NGF on neurons not known to express NGF recep-tors have been described and confirmed by numerous research groups using manydifferent in vivo and in vitro models.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1249
Protection from Glutamate Excitotoxicity and Ischemia In Vivo The initialreport by Aloe (1987) of NGF protection from glutamate receptor–mediated exci-totoxicity in striatum described a reduction in lesion size greater than that whichwould be expected if only TrkA- and p75NTR-expressing neurons had been pro-tected. A generalized and widespread neuroprotective effect of NGF has beenconfirmed in several in vivo experimental models. In response to excitotoxic glu-tamate analogues infused into the striatum, widespread and generalized neuronalprotection is achieved both by simultaneous direct infusions of NGF (Holtzmanet al 1996) and by previously placed grafts of genetically modified cells (us-ing various different cell types) that express and release NGF (Frim et al 1993,Martinex-Serrano & Bjorklund 1996, Schumacher et al 1991). In addition, NGFdelivered by infusion, grafts, or transgenic expression or induced in astrocytes byβ2-adrenoreceptor activation protects diverse populations of retinal, hippocam-pal, cortical, and other forebrain neurons that do not express detectable levels ofTrkA and p75NTR from ischemia or excitotoxic glutamate analogues (Culmsee et al1999, Guegan et al 1997, Shigeno et al 1991, Siliprandi et al 1993). It is interestingthat neutralizing antibodies to NGF block the protective effects of interleukin-1against glutamate excitotoxicity, further indicating that endogenous NGF can exertwidespread protection (Carlson et al 1999, Strijbos & Rothwell 1995).
Protection from Toxins and Metabolic Insults In Vitro NGF neuroprotectiveeffects have been described and characterized extensively in vitro for many typesof neurons not known to express TrkA and p75NTR. NGF protects cortical, hip-pocampal, striatal, retinal, and other types of neurons grown in dispersed celltissue cultures from glutamate excitotoxicity, hypoglycemia, and oxidative stress,as first reported by Mattson and colleagues (Cheng & Mattson 1991, Mattson et al1995) and confirmed, and extended to include ethanol toxicity, in many laborato-ries (Cunha et al 1998, Heaton et al 1993, Luo et al 1997, Mattson & Marck 1996,Singer et al 1999).
Potential Mechanisms of NGF-Mediated Neuroprotection of Non-NGFReceptor-Expressing NeuronsThe signaling mechanisms underlying NGF-mediated protection of neurons that do not express NGF receptors are not un-derstood. In some cases, protection has been shown to involve stabilization ofintracellular Ca2+ levels and prevention of the surge in cytoplasmic Ca2+ associ-ated with cell death, as triggered by excess glutamate signaling or oxidative stress(Mattson 1996). In a recent study, protection of cortical neurons from glutamateexcitotoxicity in vitro by both NGF and estrogen were found to require activationof the MAPK pathway (Singer et al 1999). The means by which NGF signalingleads to widespread neuroprotective effects is not known, and there are severaldifferent options to consider. The first is simply that TrkA and p75NTR expres-sion is more widespread among neurons than is currently appreciated, and verylow levels of TrkA or p75NTR might be able to mediate the protective signaling.The second option is that a novel NGF receptor exists that is widely expressed.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1250 SOFRONIEW ¥ HOWE ¥ MOBLEY
It is difficult simply to dismiss this possibility. Although radiolabeled NGF bind-ing to brain sections defines the known NGF-responsive populations, Altar et al(1991) also found significant binding in the hippocampus, in the subiculum, andin the cingulate, frontal, parietal, and occipital cortex. It is likely that much of thisbinding is attributable to NGF receptor expression on the axons of known NGFresponsive neurons. However, the formal possibility also exists that this bindingrepresents the expression of a novel receptor on cells in these regions. A thirdoption is that widespread neuroprotective effects of NGF on neurons that do notappear to express TrkA and p75NTR are mediated via NGF signaling through suchNGF-responsive nonneuronal cells as glia and inflammatory cells. Several linesof evidence support this possibility. As described above, nonneuronal cells (a)express both NGF and NGF receptors, and expression is generally upregulatedby injury and other CNS insults, (b) respond to NGF signaling, and (c) generatemolecules that are potentially toxic to neurons, such as nitric oxide and reactiveoxygen species, as part of their response to injury and inflammation. Potentiallyneuroprotective effects of NGF include the rapid inhibition of reactive oxygenspecies generation in vitro (Dugan et al 1997) and the rapid inhibition of basal andglutamate receptor–induced nitric oxide synthase in vivo (Lam et al 1998).
NGF AND NEURAL REPAIR
The molecular signals that control cellular interactions during the response toinjury and attempts at repair in the PNS and CNS are beginning to be understood.Available evidence suggests a complex interplay of molecules, including numerousgrowth factors and cytokines. Changes in the expression of NGF and NGF receptorsin both PNS and CNS are compatible with roles in repair, and their involvementwarrants further investigation.
NGF and PNS Repair
Glia and Inflammatory Cells After a peripheral nerve injury that causes axo-tomy, both meylinating and nonmyelinating Schwann cells distal to the injury de-differentiate and reenter the cell cycle. Proliferating and reactive Schwann cells pro-duce growth factors, cytokines, and growth-associated proteins, which are likely toplay key roles in axon regeneration and nerve repair (Frostick et al 1998, Mirsky &Jessen 1999, Verge et al 1996). Changes in gene expression by reactive Schwanncells include a marked upregulation of both NGF and p75NTR (Frostick et al 1998,Lindholm et al 1987, Mirsky & Jessen 1999). Peripheral nerve injury also leadsto substantial infiltration of inflammatory cells, and among these, macrophages,mast cells, and subsets of T cells have the capacity to express NGF. The preciseroles and functions of NGF signaling for different cell types during the response toperipheral nerve injury are not certain. An overall effect of exogenously adminis-tered NGF is to increase both the number and myelination of regenerating axons in
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1251
experimental entubation repair in which transected peripheral nerve stumps regrowthrough bridging tubes containing artificial matrices (Derby et al 1993, Rich et al1989). This may be due to effects of NGF signaling both on regenerating nervefibers, as discussed below, and on Schwann and inflammatory cells. ExogenousNGF promotes Schwann cell migration (Anton et al 1994), and Schwann cell mi-gration is thought to precede and promote axon elongation into entubation repairsites (Madison & Archibald 1994). Denervation is the trigger for Schwann cellproduction of NGF and NGF receptors, and the proximity of regenerating axonssuppresses this expression, probably through diffusible molecules (Taniuchi et al1988). Downregulation of p75NTR on Schwann cells induced by contact with axonsis likely to lead to Schwann cell accumulation around regenerating axons and maypromote eventual remyelination (Madison & Archibald 1994).
Neurons NGF receptor expression in peripheral nerve cells changes after ax-otomy; TrkA and p75NTR expression by sensory neurons decreases (Verge et al1996), whereas motor neurons begin to reexpress detectable levels of p75NTR
(Ernfors et al 1988, Wood et al 1990). Neither the reasons for, nor the conse-quences of, the decline in TrkA expression by axotomized sensory neurons areunderstood. Nevertheless, sensory neurons regenerate robustly and, after regen-eration, reexpress TrkA at preinjury levels. As described above, motor neuronsexpress high levels of p75NTR during the period of axon outgrowth during develop-ment and downregulate expression to undetectable levels after contact with targetstructures. After injury in adult PNS, motor neurons that are allowed to regeneraterecapitulate this developmental event and reexpress p75NTR during the period ofaxon regeneration. Reexpression does not occur in neurons that are not allowed toregenerate and requires retrograde transport from axons growing through injuredperipheral nerve tissue (Bussmann & Sofroniew 1999, Wood et al 1990). It is notyet clear whether p75NTR reexpression plays an essential role in the regenerationof either sensory or motor neurons; however, recent findings show that p75NTR sig-naling through Rho and ceramide pathways may be involved in promoting axonelongation (Brann et al 1999, Lehmann et al 1999, Yamashita et al 1999b).
NGF and CNS Repair
In the CNS, as in the PNS, injury or insults such as trauma, ischemia, or degener-ative disease trigger rapid and substantial upregulation in the expression of NGFand NGF receptors by cell types involved in the repair process, including (a) localastrocytes and microglia, (b) invading inflammatory cells, including macrophages,mast cells, and subsets of T cells, and (c) certain neurons. The purpose underly-ing such changes in expression levels is not yet known. Several lines of evidencesuggest that NGF signaling may in some of these instances facilitate the repair orreorganization of neural connections.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1252 SOFRONIEW ¥ HOWE ¥ MOBLEY
Axon Sprouting and Growth Forebrain cholinergic neurons that express TrkAand p75NTR exhibit neurite outgrowth in response to NGF in adults, particularlyafter injury (Heisenberg et al 1994, Kawaja & Gage 1991, Kordower et al 1994),suggesting that NGF may stimulate regrowth and reorganziation of connectivityof receptor bearing neurons after injury. Several other populations of CNS neuronstransiently express p75NTR during development and, in a manner similar to primarymotor neurons, downregulate this expression after maturity. These populationsinclude cerebellar Purkinje, hippocampal pyramidal neurons, and retinal neurons.Whether these neurons reexpress p75NTR after injury, as do regenerating motorneurons, and whether NGF signaling might increase their axon regeneration, as inthe PNS, is a point for further study. In support of this possibility, NGF inducesneurite outgrowth in developing hippocampal pyramidal neurons in vitro, via ap75NTR-ceramide–mediated signaling pathway (Brann et al 1999), and in retinalneurons, p75NTR signaling activates Rho pathways, which have been associatedwith axon growth (Lehmann et al 1999, Yamashita et al 1999b).
Other intriguing recent observations suggest that stimulation with NGF andother growth factors may regulate the intrinsic capacity of neurons to regeneratetransected axons through the hostile extracellular environment of the injured adultCNS. In adult spinal cord, NGF and other growth factor infusions stimulate theregrowth of fibers of receptor-expressing sensory neurons across the PNS-CNSborder in a manner that does not occur in the absence of growth factor, whichsuggests that the growth factors (including NGF) enabled these axons to over-come environmental cues that inhibit axon elongation in the CNS (Ramer et al2000). The signaling mechanism for this effect is not established, but evidencefrom other studies suggests it may involve cAMP. It has been known for sometime that if the peripheral nerves containing peripheral branches of DRG sensoryneurons receive two lesions within a short period of time, axon regeneration oc-curs more rapidly after the second injury. Moreover, a conditioning lesion madeto the peripheral branches of DRG neurons a number of days prior to a lesionof the central branches results in increased regeneration of these central branchesinto PNS-grafts (Richardson & Issa 1984). Neumann & Woolf (1999) recentlyextended these observations by showing that a peripheral conditioning lesion alsoenables considerable regeneration of the central branches of DRG neurons in theabsence of any other intervention after a second, delayed injury in the adult CNS.The findings of Cai et al (1999) suggest a possible mechanism underlying theseobservations by showing that NGF and other growth factors can mimic the effectsof peripheral conditioning lesions by increasing the capacity of DRG neurons togrow on otherwise inhibitory CNS myelin substrates in vitro. The intracellularsignaling pathway for this effect involves elevation of neuronal cAMP levels andrequires PKA activity. NGF and other growth factors are generated within theinjured peripheral nerve tissue and are transported back to the injured neuronswithin the time frame of the observed enhancements of axon regeneration by theconditioning lesion. Such findings point toward the potential to exploit signalingmechanisms of NGF and other growth factors to facilitate repair in the CNS.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1253
Inappropriate Connectivity It is important to consider that increases in NGF andother growth factors after injury may stimulate formation of inappropriate con-nections and concomitant unwanted side effects. For example, after experimentalspinal cord injury, endogenous NGF-induced sprouting of small-diameter afferentfibers has been implicated as a primary cause of autonomic dysreflexia (Krenz et al1999), an important clinical problem of patients with spinal cord injury. Thus, notall fiber regeneration and connectivity triggered by NGF and other growth factorsafter injury may be beneficial, and aberrant and inappropriate pathways may alsobe formed. Such possibilities must be studied and understood before treatmentstrategies are considered.
FAILURE OF NGF SIGNALING ANDNEURODEGENERATION
Early ideas about the functions of NGF and other members of the neurotrophinfamily naturally focused on their roles as neuronal survival factors (Barde 1989).Strict interpretation of the neurotrophic hypothesis led to the suggestion that neu-rons might be dependent on a continuous, constitutive supply of a single target-derived neurotrophin not only during development but throughout adult life andaging. An obvious extension of this idea is that an interruption in this continuouslyrequired supply of target-derived neurotrophic support might be a direct causeof neuronal death in aging and degenerative disease. Evidence accumulated todate has not substantiated this idea. As discussed above, developmentally NGF-dependent neurons for the most part become independent of a constitutive supplyof target-derived NGF for acute survival once they have established their con-nections and reached maturity. In addition, there is no evidence for a decline inNGF production in aging or degenerative disease; neither NGF mRNA nor proteinare reduced in the cerebral cortex of aged animals (Alberch et al 1991, Crutcher& Weingartner 1991) or patients with Alzheimer’s disease, where NGF proteinlevels may even be increased (Crutcher et al 1993, Goedert et al 1986, Scott et al1995). These observations do not, however, preclude other effects precipitated byan aging- or disease-related failure of NGF signaling, and there is evidence thatfailure of NGF signaling contributes to certain neurodegenerative processes.
NGF and the Age-Related Atrophy and Vulnerabilityof Forebrain Cholinergic Neurons
TrkA and p75NTR expressing basal forebrain cholinergic neurons undergo mode-rate degenerative changes in aging, including cell atrophy, downregulation oftransmitter-synthesizing enzyme, and mild cell loss in both animals and humans,and these degenerative changes are markedly exacerbated in Alzheimer’s disease(de Lacalle et al 1991a,b; Finch 1993; Fischer et al 1987, 1991; Pearson et al 1983;Whitehouse et al 1982). With the exception of cell loss, such atrophic changes
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1254 SOFRONIEW ¥ HOWE ¥ MOBLEY
can also be subacutely induced in young adult animals by disrupting access ofthese neurons to their target cells, which produce NGF and other neurotrophins(Kordower et al 1993; Sofroniew et al 1990, 1993). Furthermore, atrophic changescan be reversed in aged animals by infusions of exogenous NGF (Fischer et al1987). These findings suggest that failure of neurotrophin signaling may underlieor contribute to the atrophic neuronal changes observed in aging or degenerativedisease. However, as described above, most studies agree that NGF levels are notsubstantially reduced in aging or Alzheimer’s disease. Alternatively, available ev-idence does suggest that intrinsic neuronal changes might compromise the abilityof aged basal forebrain cholinergic neurons to derive neurotrophic support. In agedrats, forebrain cholinergic neurons exhibit a reduced capacity for generalized retro-grade transport and a pronounced reduction specifically in the retrograde transportof NGF; cholinergic neurons that do not transport NGF are severely shrunken anddownregulate TrkA expression (Cooper et al 1994, de Lacalle et al 1996). Eventhough NGF protein levels are not reduced in the cortex, they are substantially re-duced in the basal forebrain of aged animals and patients with Alzheimer’s disease(Alberch et al 1991, Scott et al 1995), as well as in mice with Down’s syndrome(JC Cooper, A Salehi, P Belichenko, J-D Delcroix, J Chua-Couzens, J Kilbridge,CL Howe, WC Mobley, submitted for publication). These findings all suggesta disturbance in retrograde transport and NGF signaling mechanisms in these neu-rons in aging and Alzheimer’s disease. Age-related declines of axonal transporthave been reported in other neuronal systems, such as sciatic nerve (McMartin& O’Connor 1979), and disturbances in neurofilaments and axonal transport havebeen proposed as a possible mechanism contributing to neurodegenerative changesassociated with aging and such age-related CNS diseases as Alzheimer’s disease(Gadjusek 1985, Saper et al 1987). Abnormal phosphorylation of the microtubule-associated protein Tau is the basis for the formation of tangles in aged neurons(Goedert 1996), and forebrain cholinergic neurons exhibit tangle formation inaging and Alzheimer’s disease, with likely disruption of microtubule-dependentaxonal transport.
Taken together, these observations suggest that an intrinsic, age-related reduc-tion in the capacity of basal forebrain cholinergic neurons to sustain retrogradeNGF signaling may contribute to the degenerative changes seen in basal forebraincholinergic neurons in aging that is markedly exacerbated in Alzheimer’s disease.Basal forebrain cholinergic neurons may be exquisitely sensitive to interruptionsin NGF signaling as a by-product of the positive feedback in NGF signaling dis-cussed above, such that an initially small failure in NGF signaling may be rapidlyexacerbated by downregulation of NGF receptors and other changes (Sofroniew &Mobley 1993). The consequences of this failure in NGF signaling may be bothdirect, by inducing cellular atrophy and changes in gene expression, and indi-rect, by increasing the vulnerability of the atrophic neurons to other insults, suchas glutamate signaling, which may become excitotoxic in the absence of NGFsignaling. As discussed above, recent studies have shown NGF significantly at-tenuates glutamate receptor–mediated death of forebrain cholinergic neurons, and
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1255
that aged rats with reduced capacity to retrogradely transport NGF exhibit signifi-cantly increased vulnerability of cholinergic neurons to glutamate toxicity. Thesefindings suggest that endogenous NGF serves neuroprotective functions, and thatloss of target-derived NGF signaling, by increasing vulnerability to excitotoxicity,may contribute to the gradual loss of basal forebrain cholinergic neurons in agingand Alzheimer’s disease. The potential for failure of neuroprotective mechanismsprovided by endogenous growth factor signaling to cause or exacerbate neurode-generation is not widely considered in the context of mechanisms that may beoperant in neurodegeneration. A better understanding of such mechanisms mayfacilitate the development of protective interventions.
NGF SIGNALING AND THERAPEUTIC INTERVENTION
The many potentially beneficial effects of NGF signaling in neural protection andrepair raise possibilities of therapeutic intervention using either NGF protein orsmall-molecule analogues (LeSauteur et al 1996, Longo & Mobley 1996, Yuen &Mobley 1996). Nevertheless, the diversity of cells that respond to NGF signalingmay predispose to unwanted side effects when NGF or NGF agonists/antagonistsare delivered systemically. For example, in a randomized and placebo-controlledstudy on the safety of intravenous or subcutaneous administration of NGF, patientsexperienced sustained hyperalgesia, which varied in a dose-dependent manner(Petty et al 1994). Infusions of NGF into the cerebroventricular system also led toa variety of undesirable and unwanted effects in patients and animals (Eriksdotter-Jonhagen et al 1998, Winkler et al 2000). Thus, ways of achieving site-specificdelivery or identifying means of achieving cell-type selective activation of NGFreceptors, or of NGF signalling pathways, may be needed to realize the therapeuticpotential that NGF signaling appears to hold.
The diversity of NGF-responsive cells also raises the possibility that NGFsignaling may, under certain circumstances, spill over from one cell compartmentto another, causing unwanted and undesirable effects, and may thus represent partof a pathophysiological mechanism. For example, NGF produced and releasedfor signaling intended for regulation of neurons or inflammatory cells may affectglia and present a potential means by which oligodendrocytes could be killed byp75NTR-signaling. Such potential effects will need to be understood before thereare attempts to exploit NGF signaling for therapeutic interventions in the CNS.Therapeutic trials must also be preceded by careful studies in realistic animalmodels of disease or injury. Even when these steps have been completed and havedefined a role for NGF treatment, there is no guarantee of success in humans. Therecent phase III trial of NGF in diabetic neuropathy failed to show an effect of NGF.This may well have resulted from the failure to deliver NGF in adequate amounts tosensory neurons, a consequence of the fact that pain limited the NGF dose that couldbe given. These and other factors will need to be addressed before NGF or NGFanalogues can be successfully used to intervene in neurodegenerative diseases.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1256 SOFRONIEW ¥ HOWE ¥ MOBLEY
CONCLUSIONS
Growth factors play important roles as intercellular signaling molecules throughoutdevelopment, adult life, and aging. Many growth factors influence a wide rangeof cell types and take part in numerous functions. NGF is no exception, and ideasabout its functions have steadily expanded. In addition to its long-recognizeddevelopmental effects on the survival and maturation of a few restricted neuronalpopulations in the PNS and CNS, NGF has effects on numerous types of neuronsand nonneuronal cells throughout adult life and during aging. Among its differentfunctions, NGF signaling appears to play important roles in the response to injuryor disease that subserve neuroprotection and neural repair.
ACKNOWLEDGMENTS
The authors wish to acknowledge the support of the McGowan Charitable Trust,the Adler Foundation, the Christopher Reeve Paralysis Foundation, and NIH GrantNS24054. CL Howe is supported by a Howard Hughes Medical Institute Predoc-toral Fellowship and the Adler Foundation. We also thank numerous colleaguesfor discussions, information, and criticism, including J-D Delcroix, C-B Wu, PBelichenko, A Salehi, J Valletta, S Lai, A Langer-Gould, and A Kr¨uttgen.
Visit the Annual Reviews home page at www.AnnualReviews.org
LITERATURE CITED
Abiru Y, Katoh-Semba R, Nishio C, HatanakaH. 1998. High potassium enhances secretionof neurotrophic factors from cultured astro-cytes.Brain Res.809:115–26
Agerman K, Canlon B, Duan M, Ernfors P.1999. Neurotrophins, NMDA receptors, andnitric oxide in development and protectionof the auditory system.Ann. NY Acad. Sci.884:131–42
Alberch J, Perez-Navarro E, Arenas E, Marsal J.1991. Involvement of nerve growth factor andits receptor in the regulation of the cholin-ergic function in aged rats.J. Neurochem.57:1483–87
Alema S, Casalbore P, Agostini E, Tato F.1985. Differentiation of PC12 phaeochromo-cytoma cells induced by v-src oncogene.Na-ture316:557–9
Alessi DR, Caudwell FB, Andjelkovic M, Hem-mings BA, Cohen P. 1996. Molecular basisfor the substrate specificity of protein kinase
B; comparison with MAPKAP kinase-1 andp70 S6 kinase.FEBS Lett.399:333–38
Alessi DR, Deak M, Casamayor A, Caud-well FB, Morrice N, et al. 1997a. 3-Phos-phoinositide-dependent protein kinase-1(PDK1): structural and functional homologywith the Drosophila DSTPK61 kinase.Curr.Biol. 7:776–89
Alessi DR, James SR, Downes CP, Holmes AB,Gaffney PR, et al. 1997b. Characterizationof a 3-phosphoinositide-dependent proteinkinase which phosphorylates and activatesprotein kinase Balpha.Curr. Biol. 7:261–69
Aloe L. 1987. Intracerebral pretreatment withnerve growth factor prevents irreversiblebrain lesions in neonatal rats injected withibotenic acid.Biotechnology5:1085–86
Aloe L. 1998. Nerve growth factor and autoim-mune diseases: role of tumor necrosis factor-alpha.Adv. Pharmacol.42:591–94
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1257
Altar CA, Dugich-Djordjevic M, ArmaniniM, Bakhit C. 1991. Medial-to-lateral gra-dient of neostriatal NGF receptors: rela-tionship to cholinergic neurons and NGF-like immunoreactivity.J. Neurosci.11:828–36
Andjelkovic M, Jakubowicz T, Cron P, MingXF, Han JW, Hemmings BA. 1996. Acti-vation and phosphorylation of a pleckstrinhomology domain containing protein kinase(RAC-PK/PKB) promoted by serum and pro-tein phosphatase inhibitors.Proc. Natl. Acad.Sci. USA93:5699–704
Angenstein F, Greenough WT, Weiler IJ. 1998.Metabotropic glutamate receptor-initiatedtranslocation of protein kinase p90rsk topolyribosomes: a possible factor regulatingsynaptic protein synthesis.Proc. Natl. Acad.Sci. USA95:15078–83
Anton ES, Weskamp G, Reichardt LF, MatthewWD. 1994. Nerve growth factor and its low-affinity receptor promote Schwann cell mi-gration.Proc. Natl. Acad. Sci. USA91:2795–99
Arch RH, Gedrich RW, Thompson CB. 1998.Tumor necrosis factor receptor-associatedfactors (TRAFs)—a family of adapter pro-teins that regulates life and death.Genes Dev.12:2821–30
Arendt T, Bruckner MK, Krell T, Pagliusi S,Kruska L, Heumann R. 1995. Degenerationof rat cholinergic basal forebrain neurons andreactive changes in nerve growth factor ex-pression after chronic neurotoxic injury. II.Reactive expression of the nerve growth fac-tor gene in astrocytes.Neuroscience65:647–59
Asaoka Y, Oka M, Yoshida K, Sasaki Y,Nishizuka Y. 1992. Role of lysophos-phatidylcholine in T-lymphocyte activation:involvement of phospholipase A2 in signaltransduction through protein kinase C.Proc.Natl. Acad. Sci. USA89:6447–51
Bamji SX, Majdan M, Pozniak CD, BelliveauDJ, Aloyz R, et al. 1998. The p75 neu-rotrophin receptor mediates neuronal apop-tosis and is essential for naturally occur-
ring sympathetic neuron death.J. Cell Biol.140:911–23
Bar-Sagi D, Feramisco JR. 1985. Microinjec-tion of the ras oncogene protein into PC12cells induces morphological differentiation.Cell 42:841–48
Barde YA. 1989. Trophic factors and neuronalsurvival.Neuron2:1525–34
Barker PA, Shooter EM. 1994. Disruption ofNGF binding to the low affinity neurotrophinreceptor p75LNTR reduces NGF binding toTrkA on PC12 cells.Neuron13:203–15
Barouch R, Appel E, Kazimirsky G, Braun A,Renz H, Brodie C. 2000. Differential regu-lation of neurotrophin expression by mito-gens and neurotransmitters in mouse lym-phocytes.J. Neuroimmunol.103:112–21
Barrett GL, Bartlett PF. 1994. The p75 nervegrowth factor receptor mediates survival ordeath depending on the stage of sensory neu-ron development.Proc. Natl. Acad. Sci. USA91:6501–5
BartLett SE, Reynolds AJ, Weible M, HeydonK, Hendry IA. 1997. In sympathetic but notsensory neurones, phosphoinositide-3 kinaseis important for NGF-dependent survival andthe retrograde transport of 125I-βNGF.BrainRes.761:257–62
Basu T, Warne PH, Downward J. 1994. Roleof Shc in the activation of Ras in responseto epidermal growth factor and nerve growthfactor.Oncogene9:3483–91
Bausenwein BS, Schmidt M, Mielke B, RaabeT. 2000. In vivo functional analysis of thedaughter of sevenless protein in receptor ty-rosine kinase signaling.Mech. Dev.90:205–15
Bazan JF. 1990. Structural design and molec-ular evolution of a cytokine receptor super-family.Proc. Natl. Acad. Sci. USA87:6934–8
Beattie EC, Howe CL, Wilder A, Brodsky FM,Mobley WC. 2001. NGF signals throughTrkA to increase clathrin at the plasma mem-brane and enhance clathrin-mediated mem-brane trafficking.J. Neurosci.In press
Bell RM, Burns DJ. 1991. Lipid activation ofprotein kinase C.J. Biol. Chem.266:4661–64
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1258 SOFRONIEW ¥ HOWE ¥ MOBLEY
Benedetti M, Levi A, Chao MV. 1993. Differen-tial expression of nerve growth factor recep-tors leads to altered binding affinity and neu-rotrophin responsiveness.Proc. Natl. Acad.Sci. USA90:7859–63
Berg-von der Emde K, Dees WL, Hiney JK,Hill D, Dissen GA, et al. 1995. Neu-rotrophins and the neuroendocrine brain:different neurotrophins sustain anatomicallyand functionally segregated subsets of hy-pothalamic dopaminergic neurons.J. Neu-rosci.15:4223–37
Berkemeier LR, Winslow JW, Kaplan DR,Nikolics K, Goeddel DV, Rosenthal A. 1991.Neurotrophin-5: a novel neurotrophic factorthat activates trk and trkB.Neuron7:857–66
Berkowitz LA, Riabowol KT, Gilman MZ.1989. Multiple sequence elements of a singlefunctional class are required for cyclic AMPresponsiveness of the mouse c-fos promoter.Mol. Cell. Biol.9:4272–81
Berninger B, Poo M. 1996. Fast actions ofneurotrophic factors.Curr. Opin. Neurobiol.6:324–30
Berra E, Diaz-Meco MT, Dominguez I, Muni-cio MM, Sanz L, et al. 1993. Protein kinaseC zeta isoform is critical for mitogenic signaltransduction.Cell 74:555–63
Berra E, Diaz-Meco MT, Lozano J, Frutos S,Municio MM, et al. 1995. Evidence for arole of MEK and MAPK during signal trans-duction by protein kinase C zeta.EMBO J.14:6157–63
Berzaghi MP, Cooper JD, Castren E, Zafra F,Sofroniew MV, et al. 1993. Cholinergic reg-ulation of brain-derived neurotrophic factor(BDNF) and nerve growth factor (NGF) butnot neurotrophin-3 (NT-3) mRNA levels inthe developing rat hippocampus.J. Neurosci.13:3818–26
Bibel M, Hoppe E, Barde YA. 1999. Bio-chemical and functional interactions betweenthe neurotrophin receptors trk and p75NTR.EMBO J.18:616–22
Bilderback TR, Gazula VR, Lisanti MP, Do-browsky RT. 1999. Caveolin interacts withTrk A and p75(NTR) and regulates neu-
rotrophin signaling pathways.J. Biol. Chem.274:257–63
Bilderback TR, Grigsby RJ, DobrowskyRT. 1997. Association of p75(NTR) withcaveolin and localization of neurotrophin-induced sphingomyelin hydrolysis to cave-olae.J. Biol. Chem.272:10922–27
Birren SJ, Lo L, Anderson DJ. 1993. Sympa-thetic neuroblasts undergo a developmentalswitch in trophic dependence.Development119:597–610
Bizon JL, Lauterborn JC, Gall CM. 1999. Sub-populations of striatal interneurons can bedistinguished on the basis of neurotrophicfactor expression.J. Comp. Neurol.408:283–98
Boise LH, Gottschalk AR, Quintans J, Thomp-son CB. 1995. Bcl-2 and Bcl-2-related pro-teins in apoptosis regulation.Curr. Top. Mi-crobiol. Immunol.200:107–21
Boldin MP, Mett IL, Varfolomeev EE, Chu-makov I, Shemer-Avni Y, et al. 1995a. Self-association of the “death domains” of thep55 tumor necrosis factor (TNF) receptor andFas/APO1 prompts signaling for TNF andFas/APO1 effects.J. Biol. Chem.270:387–91
Boldin MP, Varfolomeev EE, Pancer Z, MettIL, Camonis JH, Wallach D. 1995b. A novelprotein that interacts with the death domainof Fas/APO1 contains a sequence motif re-lated to the death domain.J. Biol. Chem.270:7795–98
Boniece I, Wagner J. 1993. Growth factorsprotect PC12 cells against ischemia by amechanism that is independent of PKA,PKC, and protein synthesis.J. Neurosci.13:4220
Bonini S, Lambiase A, Bonini S, Levi-SchafferF, Aloe L. 1999. Nerve growth factor: animportant molecule in allergic inflammationand tissue remodelling.Int. Arch. Allergy Im-munol.118:159–62
Bonni A, Ginty DD, Dudek H, Greenberg ME.1995. Serine 133-phosphorylated CREBinduces transcription via a cooperativemechanism that may confer specificity to
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1259
neurotrophin signals.Mol. Cell. Neurosci.6:168–83
Bothwell M. 1995. Functional interactions ofneurotrophins and neurotrophin receptors.Annu. Rev. Neurosci.18:223–53
Brann AB, Scott R, Neuberger Y, Abulafia D,Boldin S, et al. 1999. Ceramide signalingdownstream of the p75 neurotrophin receptormediates the effects of nerve growth factor onoutgrowth of cultured hippocampal neurons.J. Neurosci.19:8199–206
Bredesen DE, Rabizadeh S. 1997. p75NTRand apoptosis: Trk-dependent and Trk-independent effects.Trends Neurosci.20:287–90
Brennan C, Rivas-Plata K, Landis SC. 1999.The p75 neurotrophin receptor influencesNT-3 responsiveness of sympathetic neuronsin vivo. Nat. Neurosci.2:699–705
Brown MT, Cooper JA. 1996. Regulation, sub-strates and functions of src.Biochim. Bio-phys. Acta1287:121–49
Burgering BM, Coffer PJ. 1995. Protein kinaseB (c-Akt) in phosphatidylinositol-3-OH ki-nase signal transduction.Nature 376:599–602
Bush TG, Puvanachandra N, Horner CH, PolitoA, Ostenfeld T, et al. 1999. Leukocyte in-filtration, neuronal degeneration and neuriteoutgrowth after ablation of scar-forming, re-active astrocytes in adult transgenic mice.Neuron23:297–308
Butte MJ, Hwang PK, Mobley WC, FletterickRJ. 1998. Crystal structure of neurotrophin-3 homodimer shows distinct regions are usedto bind its receptors.Biochemistry37:16846–52
Cacace AM, Ueffing M, Philipp A, Han EK,Kolch W, Weinstein IB. 1996. PKC epsilonfunctions as an oncogene by enhancing acti-vation of the Raf kinase.Oncogene13:2517–26
Cai D, Shen Y, De Bellard ME, Tang S, FilbinMT. 1999. Prior exposure to neurotrophinsblocks the inhibition of axonal regenerationby MAG and myelin via a cAMP-dependentmechanism.Neuron22:89–101
Campenot RB. 1977. Local control of neuritedevelopment by nerve growth factor.Proc.Natl. Acad. Sci. USA74:4516–19
Cance WG, Craven RJ, Bergman M, Xu L, Al-italo K, Liu ET. 1994. Rak, a novel nucleartyrosine kinase expressed in epithelial cells.Cell Growth Differ.5:1347–55
Cao Z, Henzel WJ, Gao X. 1996a. IRAK: akinase associated with the interleukin-1 re-ceptor.Science271:1128–31
Cao Z, Xiong J, Takeuchi M, Kurama T, Goed-del DV. 1996b. TRAF6 is a signal transducerfor interleukin-1.Nature383:443–46
Carlson N, Wieggel W, Chen J, Bacchi A,Rogers S, Gahring L. 1999. Inflammatory cy-tokines IL-1 alpha, IL-1 beta, IL-6, and TNF-alpha impart neuroprotection to an excito-toxin through distinct pathways.J. Immunol.163:3963–68
Carroll MP, May WS. 1994. Protein kinase C-mediated serine phosphorylation directly ac-tivates Raf-1 in murine hematopoietic cells.J. Biol. Chem.269:1249–56
Carter BD, Kaltschmidt C, Kaltschmidt B, Of-fenhauser N, Bohm-Matthaei R, et al. 1996.Selective activation of NF-kappa B by nervegrowth factor through the neurotrophin re-ceptor p75.Science272:542–45
Carter BD, Lewin GR. 1997. Neurotrophinslive or let die: does p75NTR decide?Neu-ron 18:187–90
Casaccia-Bonnefil P, Carter BD, DobrowskyRT, Chao MV. 1996. Death of oligodendro-cytes mediated by the interaction of nervegrowth factor with its receptor p75.Nature383:716–19
Casademunt E, Carter BD, Benzel I, Frade JM,Dechant G, Barde YA. 1999. The zinc fingerprotein NRIF interacts with the neurotrophinreceptor p75(NTR) and participates inprogrammed cell death.EMBO J.18:6050–61
Casey PJ. 1995. Mechanisms of protein preny-lation and role in G protein function.Biochem. Soc. Trans.23:161–66
Cellerino A. 1995. Expression of messengerRNA coding for the nerve growth factor
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1260 SOFRONIEW ¥ HOWE ¥ MOBLEY
receptor trkA in the hippocampus of the adultrat.Neuroscience70:613–16
Cellerino A, Maffei L. 1996. The action of neu-rotrophins in the development and plasticityof the visual cortex.Prog. Neurobiol.49:53–71
Chao M, Casaccia-Bonnefil P, Carter B, Chitt-ka A, Kong H, Yoon SO. 1998. Neurotrophinreceptors: mediators of life and death.BrainRes. Rev.26:295–301
Chen KS, Nishimura MC, Armanini MP, Crow-ley C, Spencer SD, Phillips HS. 1997.Disruption of a single allele of the nervegrowth factor gene results in atrophy of basalforebrain cholinergic neurons and memorydeficits.J. Neurosci.17:7288–96
Chen RH, Sarnecki C, Blenis J. 1992. Nu-clear localization and regulation of erk- andrsk-encoded protein kinases.Mol. Cell. Biol.12:915–27
Cheng B, Mattson MP. 1991. NGF and bFGFprotect rat hippocampal and human corti-cal neurons against hypoglycemic damageby stabilizing calcium homeostasis.Neuron7:1031–41
Chou MM, Hou W, Johnson J, Graham LK, LeeMH, et al. 1998. Regulation of protein kinaseC zeta by PI 3-kinase and PDK-1.Curr. Biol.8:1069–77
Chrivia JC, Kwok RP, Lamb N, Hagiwara M,Montminy MR, Goodman RH. 1993. Phos-phorylated CREB binds specifically to thenuclear protein CBP.Nature365:855–59
Cifone MG, De Maria R, Roncaioli P, RippoMR, Azuma M, et al. 1994. Apoptotic sig-naling through CD95 (Fas/Apo-1) activatesan acidic sphingomyelinase.J. Exp. Med.180:1547–52
Clary DO, Reichardt LF. 1994. An alternativelyspliced form of the nerve growth factor re-ceptor TrkA confers an enhanced responseto neurotrophin 3.Proc. Natl. Acad. Sci. USA91:11133–37
Cohen P, Alessi DR, Cross DA. 1997. PDK1,one of the missing links in insulin signaltransduction?FEBS Lett.410:3–10
Cohen-Cory S, Dreyfus CF, Black IB. 1991.
NGF and excitatory neurotransmitters reg-ulate survival and morphogenesis of cul-tured cerebellar Purkinje cells.J. Neurosci.11:462–71
Coleman ES, Wooten MW. 1994. Nerve growthfactor-induced differentiation of PC12 cellsemploys the PMA-insensitive protein kinaseC-zeta isoform.J. Mol. Neurosci.5:39–57
Conover JC, Yancopoulos GD. 1997. Neu-rotrophin regulation of the developing ner-vous system: analyses of knockout mice.Rev.Neurosci.8:13–27
Cooper JD, Lindholm D, Sofroniew MV. 1994.Reduced transport of 125I-NGF by cholin-ergic neurons and downregulated TrkA ex-pression in the medial septum of aged rats.Neuroscience62:625–29
Cosman D, Lyman SD, Idzerda RL, BeckmannMP, Park LS, et al. 1990. A new cytokinereceptor superfamily.Trends Biochem. Sci.15:265–70
Coulson EJ, Reid K, Baca M, Shipham KA,Hulett SM, et al. 2001. Chopper, a new deathdomain of the p75 neurotrophin receptor thatmediates rapid neuronal cell death.J. Biol.Chem.In press
Cowley S, Paterson H, Kemp P, Marshall CJ.1994. Activation of MAP kinase kinase isnecessary and sufficient for PC12 differen-tiation and for transformation of NIH 3T3cells.Cell 77:841–52
Crews CM, Alessandrini A, Erikson RL. 1992.Erks: their fifteen minutes has arrived.CellGrowth Differ.3:135–42
Crews CM, Erikson RL. 1992. Purificationof a murine protein-tyrosine/threonine ki-nase that phosphorylates and activates theErk-1 gene product: relationship to the fissionyeast byr1 gene product.Proc. Natl. Acad.Sci. USA89:8205–9
Crowder RJ, Freeman RS. 1998. Phosphatidyli-nositol 3-kinase and Akt protein kinase arenecessary and sufficient for the survival ofnerve growth factor-dependent sympatheticneurons.J. Neurosci.18:2933–43
Crowley C, Spencer SD, Nishimura MC, ChenKS, Pitts-Meek S, et al. 1994. Mice lacking
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1261
nerve growth factor display perinatal lossof sensory and sympathetic neurons yet de-velop basal forebrain cholinergic neurons.Cell 76:1001–11
Crutcher KA, Scott SA, Liang S, Everson WV,Weingartner J. 1993. Detection of NGF-likeactivity in human brain tissue: increasedlevels in Alzheimer’s disease.J. Neurosci.13:2540–50
Crutcher KA, Weingartner J. 1991. Hippocam-pal NGF levels are not reduced in the agedfischer 344 rat.Neurobiol. Aging12:449–54
Cuello AC. 1994. Trophic factor therapy in theadult CNS: remodelling of injured basalo-cortical neurons.Prog. Brain Res.100:213–21
Cuello AC. 1996. Effects of trophic factors onthe CNS cholinergic phenotype.Prog. BrainRes.109:347–58
Culmsee C, Semkova I, Krieglstein J. 1999.NGF mediates the neuroprotective effect ofthe β2-adrenoceptor agonist clenbuterol invitro and in vivo: evidence from an NGF-antisense study.Neurochem. Int.35:47–57
Cunha GMA, Moraes RA, Moraes GA, FrancaMC Jr, Moraes MO, Viana GSB. 1998. Nervegrowth factor, ganglioside and vitamin E re-verse glutamate cytotoxicity in hippocampalcells.Eur. J. Pharmacol.367:107–12
Cunningham ME, Stephens RM, Kaplan DR,Greene LA. 1997. Autophosphorylation ofactivation loop tyrosines regulates signalingby the TRK nerve growth factor receptor.J.Biol. Chem.272:10957–67
Datta K, Bellacosa A, Chan TO, TsichlisPN. 1996. Akt is a direct target of thephosphatidylinositol 3-kinase. Activation bygrowth factors, v-src and v-Ha-ras, in Sf9 andmammalian cells.J. Biol. Chem.271:30835–39
Datta SR, Brunet A, Greenberg ME. 1999. Cel-lular survival: a play in three Akts.GenesDev.13:2905–27
Datta SR, Dudek H, Tao X, Masters S, Fu H,et al. 1997. Akt phosphorylation of BAD cou-ples survival signals to the cell-intrinsic deathmachinery.Cell 91:231–41
Davies AM. 1994. The role of neurotrophins inthe developing nervous system.J. Neurobiol.25:1334–48
Davies SW, Beardsall K. 1992. Nerve growthfactor selectively prevents excitotoxin in-duced degeneration of striatal cholinergicneurons.Neurosci. Lett.140:161–64
Debeir T, Saragovi HU, Cuello AC. 1999. Anerve growth factor mimetic TrkA antago-nist causes withdrawal of cortical choliner-gic boutons in the adult rat.Proc. Natl. Acad.Sci. USA96:4067–72
Dechant G, Barde YA. 1997. Signalling throughthe neurotrophin receptor p75NTR.Curr.Opin. Neurobiol.7:413–18
de Lacalle S, Cooper JD, Svendsen CN, Dun-nett SB, Sofroniew MV. 1991a. Degenera-tive changes in basal forebrain cholinergicneurons in aged rats. A quantitative analysisof cells stained for ChAT or NGFr, or retro-gradely labelled with fluorescent tracer.Soc.Neurosci. Abstr.17:1508
de Lacalle S, Cooper JD, Svendsen CN, Dun-nett SB, Sofroniew MV. 1996. Reducedretrograde labeling with fluorescent tracer ac-companies neuronal atrophy of basal fore-brain cholinergic neurons in aged rats.Neu-roscience75:19–27
de Lacalle S, Iraizoz I, Gonzalo LM. 1991b. Dif-ferential changes in cell size and number intopographic subdivisions of human basal nu-cleus in normal aging.Neuroscience43:445–56
del Peso L, Gonzalez-Garcia M, Page C,Herrera R, Nunez G. 1997. Interleukin-3-induced phosphorylation of BAD through theprotein kinase Akt.Science278:687–89
Derby A, Engleman VW, Frierdich GE, NeisesG, Rapp SR, Roufa DG. 1993. Nerve growthfactor facilitates regeneration across nervegaps: morphological and behavioral studiesin rat sciatic nerve.Exp. Neurol.119:176–91
Dmitrieva N, Shelton D, Rice A, McMahonS. 1997. The role of nerve growth factor ina model of visceral inflammation.Neurosci.78:449–59
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1262 SOFRONIEW ¥ HOWE ¥ MOBLEY
Dobrowsky RT, Jenkins GM, Hannun YA.1995. Neurotrophins induce sphingomyelinhydrolysis. Modulation by co-expression ofp75NTR with Trk receptors.J. Biol. Chem.270:22135–42
Dobrowsky RT, Werner MH, Castellino AM,Chao MV, Hannun YA. 1994. Activationof the sphingomyelin cycle through thelow-affinity neurotrophin receptor.Science265:1596–99
Dougherty K, Milner T. 1999. p75NTR im-munoreactivity in the rat dentate gyrus ismostly within presynaptic profiles but is alsofound in some astrocytic and postsynapticprofiles.J. Comp. Neurol.407:77–91
Douville E, Downward J. 1997. EGF inducedSOS phosphorylation in PC12 cells involvesP90 RSK-2.Oncogene15:373–83
Dudek H, Datta SR, Franke TF, Birnbaum MJ,Yao R, et al. 1997. Regulation of neuronalsurvival by the serine-threonine protein ki-nase Akt.Science275:661–65
Dugan LL, Creedon DJ, Johnson EM, HoltzmanDM. 1997. Rapid suppression of free radicalformation by nerve growth factor involvesthe mitogen-activated protein kinase path-way. Proc. Natl. Acad. Sci. USA94:4086–91
Easton RM, Deckwerth TL, Parsadanian AS,Johnson EM Jr. 1997. Analysis of the mech-anism of loss of trophic factor dependenceassociated with neuronal maturation: a phe-notype indistinguishable from bax deletion.J. Neurosci.17:9656–66
Eckenstein F. 1988. Transient expressionof NGF-receptor-like immunoreactivity inpostnatal rat brain and spinal cord.Brain Res.446:149–54
Edwards RH, Selby MJ, Mobley WC, WeinrichSL, Hruby DE, Rutter WJ. 1988. Processingand secretion of nerve growth factor: expres-sion in mammalian cells with a vaccinia virusvector.Mol. Cell. Biol.8:2456–64
Edwards RH, Selby MJ, Rutter WJ. 1986.Differential RNA splicing predicts two dis-tinct nerve growth factor precursors.Nature319:784–87
Elkabes S, DiCicco-Bloom E, Black I. 1996.Brain microglia/macrophages express neu-rotrophins that selectively regulate mi-croglial proliferation and function.J. Neu-rosci.16:2508–21
Elkabes S, Peng L, Black IB. 1998.Lipopolysaccharide differentially regulatesmicroglial trk receptor and neurotrophinexpression.J. Neurosci. Res.54:117–22
Ellinger-Ziegelbauer H, Brown K, Kelly K,Siebenlist U. 1997. Direct activation of thestress-activated protein kinase (SAPK) andextracellular signal-regulated protein kinase(ERK) pathways by an inducible mitogen-activated protein kinase/ERK kinase ki-nase 3 (MEKK) derivative.J. Biol. Chem.272:2668–74
Enokido Y, Wyatt S, Davies AM. 1999. Devel-opmental changes in the response of trigem-inal neurons to neurotrophins: influence ofbirthdate and the ganglion environment.De-velopment126:4365–73
Eriksdotter-J¨onhagen M, Nordberg A, AmberlaK, Backman L, Ebendal T, et al. 1998. Intra-cerebroventricular infusion of nerve growthfactor in three patients with Alzheimer’s dis-ease.Dement. Geriatr. Cogn. Disord.9:246–57
Erikson E, Maller JL. 1991. Purification andcharacterization of ribosomal protein S6 ki-nase I from Xenopus eggs.J. Biol. Chem.266:5249–55
Erikson E, Maller JL, Erikson RL. 1991. Xeno-pus ribosomal protein S6 kinase II.MethodsEnzymol.200:252–68
Ernfors P, Hallbook F, Ebendal T, Shooter EM,Radeke MJ, et al. 1988. Developmental andregional expression of beta-nerve growth fac-tor receptor mRNA in the chick and rat.Neu-ron 1:983–96
Ernfors P, Henschen A, Olson L, Persson H.1989. Expression of nerve growth factorreceptor mRNA is developmentally regulatedand increased after axotomy in rat spinal cordmotoneurons.Neuron2:1605–13
Erpel T, Courtneidge SA. 1995. Src familyprotein tyrosine kinases and cellular signal
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1263
transduction pathways.Curr. Opin. Cell Biol.7:176–82
Facchinetti F, Dawson VL, Dawson TM. 1998.Free radicals as mediators of neuronal injury.Cell. Mol. Neurobiol.18:609–20
Fagan AM, Garber M, Barbacid M, Silos-Santiago I, Holtzman DM. 1997. A role forTrkA during maturation of striatal and basalforebrain cholinergic neurons in vivo.J. Neu-rosci.17:7644–54
Fahnestock M, Woo JE, Lopez GA, SnowJ, Walz DA, et al. 1991. beta-NGF-endopeptidase: structure and activity of akallikrein encoded by the gene mGK-22.Bio-chemistry30:3443–50
Finch CE. 1993. Neuron atrophy during aging:programmed or sporadic.Trends Neurosci.16:104–10
Fischer W, Chen KS, Gage FH, Bj¨orklund A.1991. Progressive decline in spatial learningand integrity of forebrain cholinergic neuronsin rats during aging.Neurobiol. Aging13:9–23
Fischer W, Wictorin K, Bjorklund A, WilliamsLR, Varon S, Gage FH. 1987. Ameliorationof cholinergic neuronal atrophy and spatialmemory impairment in aged rats by nervegrowth factor.Nature329:65–68
Frade J, Barde Y. 1998. Microglia-derived nervegrowth factor causes cell death in the devel-oping retina.Neuron20:35–41
Frade JM, Rodriguez-Tebar A, Barde Y. 1996.Induction of cell death by endogenous nervegrowth factor through its p75 receptor.Na-ture383:166–68
Franke TF, Kaplan DR, Cantley LC. 1997.PI3K: downstream AKTion blocks apopto-sis.Cell 88:435–37
Franke TF, Yang SI, Chan TO, Datta K, Kaz-lauskas A, et al. 1995. The protein kinase en-coded by the Akt proto-oncogene is a targetof the PDGF-activated phosphatidylinositol3-kinase.Cell 81:727–36
Freed E, Symons M, Macdonald SG, Mc-Cormick F, Ruggieri R. 1994. Binding of 14-3-3 proteins to the protein kinase Raf and ef-fects on its activation.Science265:1713–16
French SJ, Humby T, Horner CH, SofroniewMV, Rattray M. 1999. Hippocampal neu-rotrophin and trk receptor mRNA levels arealtered by local administration of nicotine,carbachol and pilocarpine.Mol. Brain Res.67:124–36
Friedman WJ, Greene LA. 1999. Neurotrophinsignaling via Trks and p75.Exp. Cell Res.253:131–42
Friedman WJ, Thakur S, Seidman L, RabsonAB. 1996. Regulation of nerve growth fac-tor mRNA by interleukin-1 in rat hippocam-pal astrocytes is mediated by NFκB. J. Biol.Chem.271:31115–20
Frim DM, Uhler TA, Short MP, Ezzedine ZD,Klagsbrun M, et al. 1993. Effects of bio-logically delivered NGF, BDNF and bFGFon striatal excitotoxic lesions.NeuroReport4:367–70
Frostick SP, Yin Q, Kemp GJ. 1998. Schwanncells, neurotrophic factors, and peripheralnerve regeneration.Microsurgery 18:397–405
Fu H, Xia K, Pallas DC, Cui C, Conroy K, et al.1994. Interaction of the protein kinase Raf-1 with 14-3-3 proteins.Science266:126–29
Gadjusek DC. 1985. Hypothesis: interferencewith axonal transport of neurofilament as acommon pathogenic mechanism in certaindiseases of the central nervous system.N.Engl. J. Med.312:714–19
Gage FH, Batchelor P, Chen KS, Chin D, Hig-gins GA, et al. 1989. NGF receptor reexpres-sion and NGF-mediated cholinergic neuronalhypertrophy in the damaged adult neostria-tum.Neuron2:1177–84
Gall CM, Isackson PJ. 1989. Limbic seizuresincrease neuronal production of messen-ger RNA for nerve growth factor.Science245:758–61
Gallo G, Lefcort FB, Letourneau PC. 1997. ThetrkA receptor mediates growth cone turningtoward a localized source of nerve growthfactor.J. Neurosci.17:5445–54
Gargano N, Levi A, Alema S. 1997. Modula-tion of nerve growth factor internalization by
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1264 SOFRONIEW ¥ HOWE ¥ MOBLEY
direct interaction between p75 and TrkA re-ceptors.J. Neurosci. Res.50:1–12
Gee AP, Boyle MD, Munger KL, LawmanMJ, Young M. 1983. Nerve growth factor:stimulation of polymorphonuclear leukocytechemotaxis in vitro.Proc. Natl. Acad. Sci.USA80:7215–18
Gentry JJ, Casaccia-Bonnefil P, Carter BD.2000. Nerve growth factor activation of nu-clear factorκB through its p75 receptor is ananti-apoptotic signal in RN22 Schwannomacells.J. Biol. Chem.275:7558–65
Gertler FB, Bennett RL, Clark MJ, HoffmannFM. 1989. Drosophila abl tyrosine kinasein embryonic CNS axons: a role in axono-genesis is revealed through dosage-sensitiveinteractions with disabled.Cell 58:103–13
Gertler FB, Hill KK, Clark MJ, HoffmannFM. 1993. Dosage-sensitive modifiers ofDrosophila abl tyrosine kinase function:prospero, a regulator of axonal outgrowth,and disabled, a novel tyrosine kinase sub-strate.Genes Dev.7:441–53
Ghoda L, Lin X, Greene WC. 1997. The 90-kDa ribosomal S6 kinase (pp90rsk) phospho-rylates the N-terminal regulatory domain ofIkappaBalpha and stimulates its degradationin vitro. J. Biol. Chem.272:21281–88
Ghosh S, May MJ, Kopp EB. 1998. NF-kappaB and Rel proteins: evolutionarily conservedmediators of immune responses.Annu. Rev.Immunol.16:225–60
Gille H, Kortenjann M, Thomae O, Moomaw C,Slaughter C, et al. 1995. ERK phosphoryla-tion potentiates Elk-1-mediated ternary com-plex formation and transactivation.EMBO J.14:951–62
Ginty DD, Bonni A, Greenberg ME. 1994.Nerve growth factor activates a Ras-dependent protein kinase that stimulatesc-fos transcription via phosphorylation ofCREB.Cell 77:713–25
Goedert M, Fine A, Hunt SP, Ullrich A. 1986.Nerve growth factor mRNA in peripheral andcentral rat tissues and in the human centralnervous system: lesion effects in the rat brain
and levels in Alzheimer’s disease.Mol. BrainRes.1:85–92
Goedert M. 1996. Tau protein and the neu-rofibrillary pathology of Alzheimer’s dis-ease.Ann. NY Acad. Sci.777:121–31
Goss JR, O’Malley ME, Zou L, Styren SD,Kochanek PM, DeKosky ST. 1998. Astro-cytes are the major souce of nerve growthfactor upregulation following traumatic braininjury in the rat.Exp. Neurol.149:301–9
Gottlieb M, Matute C. 1999. Expression ofnerve growth factor in astrocytes of the hip-pocampal CA1 area following transient fore-brain ischemia.Neuroscience91:1027–34
Gotz R, Koster R, Winkler C, Raulf F, Lott-speich F, et al. 1994. Neurotrophin-6 is a newmember of the nerve growth factor family.Nature372:266–69
Greene LA, Kaplan DR. 1995. Early events inneurotrophin signalling via Trk and p75 re-ceptors.Curr. Opin. Neurobiol.5:579–87
Greenberg ME, Hermanowski AL, Ziff EB.1986. Effect of protein synthesis inhibitorson growth factor activation of c-fos, c-myc,and actin gene transcription.Mol. Cell. Biol.6:1050–57
Greferath U, Bennie A, Kourakis A, BartLettPF, Murphy M, Barrett GL. 2000. Enlargedcholinergic forebrain neurons and improvedspatial learning in p75 knockout mice.Eur J.Neurosci.12:885–93
Grimes ML, Beattie E, Mobley WC. 1997.A signaling organelle containing the nervegrowth factor-activated receptor tyrosine ki-nase, TrkA. Proc. Natl. Acad. Sci. USA94:9909–14
Grimes ML, Zhou J, Beattie EC, Yuen EC,Hall DE, et al. 1996. Endocytosis of activatedTrkA: evidence that nerve growth factor in-duces formation of signaling endosomes.J.Neurosci.16:7950–64
Grinspan JB, Marchionni MA, Reeves M,Coulaloglou M, Scherer SS. 1996. Axonalinteractions regulate Schwann cell apopto-sis in developing peripheral nerve: neureg-ulin receptors and the role of neuregulins.J.Neurosci.16:6107–18
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1265
Gross A, Jockel J, Wei MC, Korsmeyer SJ.1998. Enforced dimerization of BAX resultsin its translocation, mitochondrial dysfunc-tion and apoptosis.EMBO J.17:3878–85
Gu C, Casaccia-Bonnefil P, Srinivasan A, ChaoMV. 1999. Oligodendrocyte apoptosis me-diated by caspase activation.J. Neurosci.19:3043–49
Gu H, Pratt JC, Burakoff SJ, Neel BG. 1998.Cloning of p97/Gab2, the major SHP2-binding protein in hematopoietic cells, re-veals a novel pathway for cytokine-inducedgene activation.Mol. Cell 2:729–40
Gu Q, Liu Y, Cynader MS. 1994. Nerve growthfactor-induced ocular dominance plasticity inadult cat visual cortex.Proc. Natl. Acad. Sci.USA91:8408–12
Guegan C, Onteniente B, Makiura Y, Merad-Boudia M, Ceballos-Picot I, Sola B. 1997.Reduction of cortical infarction and impair-ment of apoptosis in NGF-transgenic micesubjected to permanent focal ischemia.Mol.Brain Res.55:133–40
Hadari YR, Kouhara H, Lax I, SchlessingerJ. 1998. Binding of Shp2 tyrosine phos-phatase to FRS2 is essential for fibroblastgrowth factor-induced PC12 cell differenti-ation.Mol. Cell. Biol.18:3966–73
Hagag N, Halegoua S, Viola M. 1986. Inhibi-tion of growth factor-induced differentiationof PC12 cells by microinjection of antibodyto ras p21.Nature319:680–82
Hakak Y, Hsu YS, Martin GS. 2000. Shp-2 me-diates v-Src-induced morphological changesand activation of the anti-apoptotic proteinkinase Akt.Oncogene19:3164–71
Hasen¨ohrl RU, Soderstrom S, Mohammed AH,Ebendal T, Huston JP. 1997. Reciprocalchanges in expression of mRNA for nervegrowth factor and its receptors TrkA andLNGFR in brain of aged rats in relationto maze learning deficits.Exp. Brain Res.114:205–13
Hausser A, Storz P, Link G, Stoll H, Liu YC,et al. 1999. Protein kinase C mu is negativelyregulated by 14-3-3 signal transduction pro-teins.J. Biol. Chem.274:9258–64
Hawley RJ, Scheibe RJ, Wagner JA. 1992.NGF induces the expression of the VGF genethrough a cAMP response element.J. Neu-rosci.12:2573–81
Heaton MB, Paiva M, Swanson DJ, Walker DW.1993. Modulation of ethanol neurotoxicityby nerve growth factor.Brain Res.620:78–85
Heese K, Fiebich BL, Bauer J, Otten U.1998. NF-κB modulates lipopolysaccharide-induced microglial nerve growth factor ex-pression.Glia 22:401–7
Hefti F. 1986. Nerve growth factor promotessurvival of septal cholinergic neurons afterfimbrial transections.J. Neurosci.6:2155–62
Heisenberg C-P, Cooper JD, Berke J, SofroniewMV. 1994. NMDA potentiates NGF-inducedsprouting of septal cholinergic neurons.Neu-roReport5:413–16
Heumann R, Korsching S, Bandtlow C, Thoe-nen H. 1987a. Changes of nerve growth fac-tor synthesis in nonneuronal cells in responseto sciatic nerve transection.J. Cell Biol.104:1623–31
Heumann R, Lindholm D, Bandtlow C, MeyerM, Radeke MJ, et al. 1987b. Differentialregulation of mRNA encoding nerve growthfactor and its receptor in rat sciatic nerve dur-ing development, degeneration, and regener-ation: role of macrophages.Proc. Natl. Acad.Sci. USA84:8735–39
Hill CS, Marais R, John S, Wynne J, Dalton S,Treisman R. 1993. Functional analysis of agrowth factor-responsive transcription factorcomplex.Cell 73:395–406
Holgado-Madruga M, Emlet DR, MoscatelloDK, Godwin AK, Wong AJ. 1996. A Grb2-associated docking protein in EGF- andinsulin-receptor signalling.Nature379:560–64
Holgado-Madruga M, Moscatello DK, Em-let DR, Dieterich R, Wong AJ. 1997.Grb2-associated binder-1 mediates phos-phatidylinositol 3-kinase activation and thepromotion of cell survival by nerve growthfactor.Proc. Natl. Acad. Sci. USA94:12419–24
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1266 SOFRONIEW ¥ HOWE ¥ MOBLEY
Holtzman DM, Kilbridge J, Li Y, CunninghamET, Lenn NJ, et al. 1995. TrkA expression inthe CNS: evidence for the existence of sev-eral novel NGF-responsive CNS neurons.J.Neurosci.15:1567–76
Holtzman DM, Li Y, Parada LF, Kinsman S,Chen CK, et al. 1992. p140trk mRNA marksNGF-responsive forebrain neurons: evidencethat trk gene expression is induced by NGF.Neuron9:465–78
Holtzman DM, Sheldon RA, Jaffe W, ChengY, Ferriero DM. 1996. Nerve growth factorprotects the neonatal brain against hypoxic-ischemic injury. Ann. Neurol. 39:114–22
Horner CH, Lam HHD, Berke J, Cooper JD,Sofroniew MV. 1999. Local and target-derived nerve growth factor protect basalforebrain cholinergic neurons from gluta-mate receptor-mediated toxicity.Soc. Neu-rosci. Abstr.25:770
Horvath CM, Wolven A, Machadeo D, Huber J,Boter L, et al. 1993. Analysis of the trk NGFreceptor tyrosine kinase using recombinantfusion proteins.J. Cell Sci.17:223–28
Howe CL, Mobley WC. 2001. Nerve growthfactor effects on cholinergic modulation ofhippocampal and cortical plasticity. InNeu-robiology of the Neurotrophins, ed. I Moc-chetti, FP Graham, pp. 255–307
Huang CS, Zhou J, Feng AK, Lynch CC,Klumperman J, et al. 1999. Nerve growthfactor signaling in caveolae-like domainsat the plasma membrane.J. Biol. Chem.274:36707–14
Huber LJ, Chao MV. 1995. A potential interac-tion of p75 and trkA NGF receptors revealedby affinity crosslinking and immunoprecipi-tation.J. Neurosci. Res.40:557–63
Hutton L, deVellis J, Perez-Polo J. 1992.Expression of p75NGFR TrkA, and TrkBmRNA in Rat C6 glioma and type U astrocytecultures.J. Neurosci. Res.32:375–83
Hutton L, Perez-Polo J. 1995. In vitro glial re-sponses to nerve growth factor.J. Neurosci.Res.41:185–96
Ibanez CF. 1994. Structure function relation-
ships in the neurotrophin family.J. Neuro-biol. 25:1349–61
Ibanez CF, Ebendal T, Barbany G, Murray-RustJ, Blundell TL, Persson H. 1992. Disruptionof the low affinity receptor-binding site inNGF allows neuronal survival and differen-tiation by binding to the trk gene product.Cell 69:329–41
Ip NY, Stitt TN, Tapley P, Klein R, Glass DJ,et al. 1993. Similarities and differences in theway neurotrophins interact with the Trk re-ceptors in neuronal and nonneuronal cells.Neuron10:137–49
Irie K, Gotoh Y, Yashar BM, Errede B, NishidaE, Matsumoto K. 1994. Stimulatory effectsof yeast and mammalian 14-3-3 proteins onthe Raf protein kinase.Science265:1716–19
Jaiswal RK, Moodie SA, Wolfman A, LandrethGE. 1994. The mitogen-activated proteinkinase cascade is activated by B-Raf in re-sponse to nerve growth factor through inter-action with p21ras.Mol. Cell. Biol.14:6944–53
Janknecht R, Ernst WH, Pingoud V, NordheimA. 1993. Activation of ternary complex factorElk-1 by MAP kinases.EMBO J.12:5097–104
Jia M, Li M, Liu XW, Jiang H, NelsonPG, Guroff G. 1999. Voltage-sensitive cal-cium currents are acutely increased by nervegrowth factor in PC12 cells.J. Neurophysiol.82:2847–52
Jiang H, Takeda K, Lazarovici P, Katagiri Y, YuZX, et al. 1999. Nerve growth factor (NGF)-induced calcium influx and intracellular cal-cium mobilization in 3T3 cells expressingNGF receptors.J. Biol. Chem.274:26209–16
Joel PB, Smith J, Sturgill TW, Fisher TL,Blenis J, Lannigan DA. 1998. pp90rsk1 reg-ulates estrogen receptor-mediated transcrip-tion through phosphorylation of Ser-167.Mol. Cell. Biol.18:1978–84
Johnson EM, Deckwerth TL Jr. 1993. Molec-ular mechanisms of developmental neuronaldeath.Annu. Rev. Neurosci.16:31–46
Johnson EM, Rich KM, Yip HK. 1986. The role
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1267
of NGF in sensory neurons in vivo.TrendsNeurosci.9:33–37
Kaplan DR, Hempstead BL, Martin-ZancaD, Chao MV, Parada LF. 1991a. The trkproto-oncogene product: a signal transduc-ing receptor for nerve growth factor.Science252:554–58
Kaplan DR, Martin-Zanca D, Parada LF. 1991b.Tyrosine phosphorylation and tyrosine ki-nase activity of the trk proto-oncogene prod-uct induced by NGF.Nature350:158–60
Kaplan DR, Miller FD. 1997. Signal trans-duction by the neurotrophin receptors.Curr.Opin. Cell. Biol.9:213–21
Karlsson M, Clary DO, Lefcort FB, ReichardtLF, Karten HJ, Hallbook F. 1998. Nervegrowth factor receptor TrkA is expressedby horizontal and amacrine cells duringchicken retinal development.J. Comp. Neu-rol. 400:408–16
Kawaja MD, Gage FH. 1991. Reactive astro-cytes are substrates for the growth of adultCNS axons in the presence of elevated levelsof nerve growth factor.Neuron7:1019–30
Kew JNC, Smith DW, Sofroniew MV. 1996.Nerve growth factor withdrawal inducesthe apoptotic death of developing septalcholinergic neuronsin vitro: protection bycAMP analogue and high K+. Neuroscience70:329–39
Khasar SG, Lin YH, Martin A, Dadgar J,McMahon T, et al. 1999. A novel nociceptorsignaling pathway revealed in protein kinaseC epsilon mutant mice.Neuron24:253–60
Khursigara G, Orlinick JR, Chao MV. 1999. As-sociation of the p75 neurotrophin receptorwith TRAF6.J. Biol. Chem.274:2597–600
Klein R, Jing SQ, Nanduri V, O’Rourke E, Bar-bacid M. 1991. The trk proto-oncogene en-codes a receptor for nerve growth factor.Cell65:189–97
Klesse LJ, Parada LF. 1998. p21 ras andphosphatidylinositol-3 kinase are requiredfor survival of wild-type and NF1 mutant sen-sory neurons.J. Neurosci.8:10420–28
Klippel A, Kavanaugh WM, Pot D, WilliamsLT. 1997. A specific product of phos-
phatidylinositol 3-kinase directly activatesthe protein kinase Akt through its pleckstrinhomology domain.Mol. Cell. Biol. 17:338–44
Knipper M, Berzaghi MP, Blochl A, BreerH, Thoenen H, Lindholm D. 1994. Positivefeedback between acetylcholine and the neu-rotrophins nerve growth factor and brain-derived growth factor in the rat hippocampus.Eur. J. Neurosci.6:668–71
Kobayashi M, Nagata S, Kita Y, Nakatsu N,Ihara S, et al. 1997. Expression of a consti-tutively active phosphatidylinositol 3-kinaseinduces process formation in rat PC12 cells.Use of Cre/loxP recombination system.J.Biol. Chem.272:16089–92
Koh S, Higgins GA. 1991. Differential regula-tion of the low-affinity nerve growth factorreceptor during postnatal development of therat brain.J. Comp. Neurol.313:494–508
Kolch W, Heidecker G, Kochs G, Hummel R,Vahidi H, et al. 1993. Protein kinase C alphaactivates RAF-1 by direct phosphorylation.Nature364:249–52
Kordower JH, Burke-Watson M, Roback JD,Wainer BH. 1993. Stability of septohip-pocampal neurons following excitotoxic le-sions of the rat hippocampus.Exp. Neurol.117:1–16
Kordower JH, Winn SR, Liu Y, Mufson EJ,Sladek JR, et al. 1994. The aged monkeybasal forebrain: rescue and sprouting of axo-tomized basal forebrain neurons after graftsof encapsulated cells secreting human nervegrowth factor.Proc. Natl. Acad. Sci. USA91:10898–902
Korhonen JM, Said FA, Wong AJ, KaplanDR. 1999. Gab1 mediates neurite outgrowth,DNA synthesis, and survival in PC12 cells.J. Biol. Chem.274:37307–14
Korner M, Tarantino N, Pleskoff O, Lee LM,Debre P. 1994. Activation of nuclear factorkappa B in human neuroblastoma cell lines.J. Neurochem.62:1716–26
Kouhara H, Hadari YR, Spivak-Kroizman T,Schilling J, Bar-Sagi D, et al. 1997. A lipid-anchored Grb2-binding protein that links
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1268 SOFRONIEW ¥ HOWE ¥ MOBLEY
FGF-receptor activation to the Ras/MAPKsignaling pathway.Cell 89:693–702
Kremer NE, D’Arcangelo G, Thomas SM, De-Marco M, Brugge JS, Halegoua S. 1991. Sig-nal transduction by nerve growth factor andfibroblast growth factor in PC12 cells re-quires a sequence of src and ras actions.J.Cell Biol. 115:809–19
Krenz NR, Meakin SO, Krassioukov AV,Weaver LC. 1999. Neutralizing intraspinalnerve growth factor blocks autonomic dys-reflexia caused by spinal cord injury.J. Neu-rosci.19:7405
Kroemer G. 1997. The proto-oncogene Bcl-2and its role in regulating apoptosis.Nat. Med.3:614–20
Kruttgen A, Moller J, Heymach JV, ShooterEM. 1998. Neurotrophins induce releaseof neurotrophins by the regulated secre-tory pathway.Proc. Natl. Acad. Sci. USA95:9614–19
Kumar S, Pena L, de Vellis J. 1993. CNS glialcells express neurotrophin receptors whoselevels are regulated by NGF.Brain Res. Mol.Brain Res.17:163–68
Kuo WL, Chung KC, Rosner MR. 1997. Differ-entiation of central nervous system neuronalcells by fibroblast-derived growth factor re-quires at least two signaling pathways: rolesfor Ras and Src.Mol. Cell. Biol.17:4633–43
Labouyrie E, Parrens M, de Mascarel A, BlochB, Merlio JP. 1997. Distribution of NGF re-ceptors in normal and pathologic human lym-phoid tissues.J. Neuroimmunol.77:161–73
Ladiwala U, Lachance C, Simoneau SJ, BhakarA, Barker PA, Antel JP. 1998. p75 neu-rotrophin receptor expression on adult hu-man oligodendrocytes: signaling without celldeath in response to NGF.J. Neurosci.18:1297–304
Lai KO, Fu WY, Ip FCF, Ip NY. 1998. Cloningand expression of a novel neurotrophin,NT-7, from carp.Mol. Cell. Neurosci.11:64–76
Lam HHD, Bhardwaj A, O’Connell MT, Han-ley DF, Traystman RJ, Sofroniew MV. 1998.Nerve growth factor rapidly suppresses basal,
NMDA-evoked, and AMPA-evoked nitricoxide synthase activity in rat hippocampusinvivo. Proc. Natl. Acad. Sci. USA95:10926–31
Lambiase A, Bracci-Laudiero L, Bonini S,Bonini S, Starace G, et al. 1997. HumanCD4+ T cell clones produce and releasenerve growth factor and express high-affinitynerve growth factor receptors.J. Allergy Clin.Immunol.100:408–14
Landreth GE, Shooter EM. 1980. Nerve growthfactor receptors on PC12 cells: ligand-induced conversion from low- to high-affinity states.Proc. Natl. Acad. Sci. USA77:4751–55
Lange-Carter CA, Johnson GL. 1994. Ras-dependent growth factor regulation of MEKkinase in PC12 cells.Science265:1458–61
Large TH, Weskamp G, Helder JC, Radeke MJ,Misko TP, et al. 1989. Structure and develop-mental expression of the nerve growth factorreceptor in the chicken central nervous sys-tem.Neuron2:1123–34
Le Good JA, Ziegler WH, Parekh DB, AlessiDR, Cohen P, Parker PJ. 1998. Protein kinaseC isotypes controlled by phosphoinositide3-kinase through the protein kinase PDK1.Science281:2042–45
Lee J, Wang Z, Luoh SM, Wood WI, Scad-den DT. 1994a. Cloning of FRK, a novel hu-man intracellular SRC-like tyrosine kinase-encoding gene.Gene138:247–51
Lee KF, Bachman K, Landis S, Jaenisch R.1994b. Dependence on p75 for innerva-tion of some sympathetic targets.Science263:1447–49
Lee KF, Davies AM, Jaenisch R. 1994c. p75-deficient embryonic dorsal root sensory andneonatal sympathetic neurons display a de-creased sensitivity to NGF.Development120:1027–33
Lee KF, Li E, Huber LJ, Landis SC, Sharpe AH,et al. 1992. Targeted mutation of the geneencoding the low affinity NGF receptor p75leads to deficits in the peripheral sensory ner-vous system.Cell 69:737–49
Lee SB, Rhee SG. 1995. Significance of PIP2
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1269
hydrolysis and regulation of phospholipase Cisozymes.Curr. Opin. Cell Biol.7:183–89
Lehmann M, Fourneir A, Selles-Navarro I,Dergham P, Sebok A, et al. 1999. Inactiva-tion of rho signaling pathway promotes CNSaxon regeneration.J. Neurosci.19:7537–47
Lemke G, Chao M. 1988. Axons regu-late Schwann cell expression of the majormyelin and NGF receptor genes.Develop-ment102:499–504
Leon A, Buriani A, Dal Toso R, Fabris M, Ro-manello S, et al. 1994. Mast cells synthesize,store, and release nerve growth factor.Proc.Natl. Acad. Sci. USA91:3739–43
LeSauteur L, Cheung NV, Lisbona R, SaragoviHU. 1996. Small molecule nerve growth fac-tor analogues image receptors in vivo.Nat.Biotechnol.14:1120–22
Levi-Montalcini R, Hamburger V. 1951. Selec-tive growth stimulating effects of mouse sar-coma on the sensory and sympathetic ner-vous system of chick embryo.J. Exp. Zool.116:321–61
Levi-Montalcini R, Hamburger V. 1953. A dif-fusible agent of mouse sarcoma, producinghyperplasia of sympathetic ganglia and hy-perneurotization of viscera in the chick em-bryo.J. Exp. Zool.123:233–87
Levi-Montalcini R, Skaper SD, Toso RD, Pe-trelli L, Leon A. 1996. Nerve growth factor:from neurotrophin to neurokine.Trends Neu-rosci.19:514–20
Levi-Montalcini R, Toso RD, Valle FD, SkaperSD, Leon A. 1995. Update of the NGF saga.J. Neurol. Sci.130:119–27
Lewis TS, Shapiro PS, Ahn NG. 1998. Signaltransduction through MAP kinase cascades.Adv. Cancer Res.74:49–139
Li Y, Holtzman DM, Kromer LF, Kaplan DR,Chua-Couzens J, et al. 1995. Regulation ofTrkA and ChAT expression in developing ratbasal forebrain: evidence that both exoge-nous and endogenous NGF regulate differ-entiation of cholinergic neurons.J. Neurosci.15:2888–905
Liepinsh E, Ilag LL, Otting G, Ibanez CF. 1997.NMR structure of the death domain of the p75
neurotrophin receptor.EMBO J. 16:4999–5005
Lindholm D, Heumann R, Meyer M, ThoenenH. 1987. Interleukin-1 regulates synthesis ofnerve growth factor in non-neuronal cells ofrat sciatic nerve.Nature330:658–59
Lindsay RM, Harmar AJ. 1989. Nerve growthfactor regulates expression of neuropep-tide genes in adult sensory neurons.Nature337:362–64
Lindvall O, Kokaia Z, Bengzon J, Elmer E,Kokaia M. 1994. Neurotrophins and brain in-sults.Trend. Neurosci.17:490–96
Liu YZ, Chrivia JC, Latchman DS. 1998. Nervegrowth factor upregulates the transcriptionalactivity of CBP through activation of thep42/p44(MAPK) cascade.J. Biol. Chem.273:32400–407
Liu ZG, Hsu H, Goeddel DV, Karin M. 1996.Dissection of TNF receptor 1 effector func-tions: JNK activation is not linked to apopto-sis while NF-kappaB activation prevents celldeath.Cell 87:565–76
Liuzzo J, Petanceska S, Devi L. 1999. Neuro-trophic factors regulate cathepsin S in ma-crophages and microglia: a role in the degra-dation of myelin basic protein and amyloidbeta peptide.Mol. Med.5:334–43
Liyanage M, Frith D, Livneh E, Stabel S. 1992.Protein kinase C group B members PKC-delta, -epsilon, -zeta and PKC-L(eta). Com-parison of properties of recombinant proteinsin vitro and in vivo.Biochem. J.283:781–87
Loeb DM, Maragos J, Martin-Zanca D, ChaoMV, Parada LF, Greene LA. 1991. The trkproto-oncogene rescues NGF responsivenessin mutant NGF-nonresponsive PC12 celllines.Cell 66:961–66
Loeb DM, Stephens RM, Copeland T, KaplanDR, Greene LA. 1994. A Trk nerve growthfactor (NGF) receptor point mutation affect-ing interaction with phospholipase C-gamma1 abolishes NGF-promoted peripherin induc-tion but not neurite outgrowth.J. Biol. Chem.269:8901–10
Longo FM, Holtzman DM, Grimes ML, Mob-ley WC. 1993. Nerve growth factor: actions
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1270 SOFRONIEW ¥ HOWE ¥ MOBLEY
in the peripheral and central nervous systems.In Neurotrophic Factors, ed. SE Loughlin,JH Fallon. San Deigo, CA: Academic. 209–56
Longo FM, Mobley WC. 1996. Minimizedhormones grow in stature.Nat. Biotechnol.14:1092
Lucidi-Phillipi CA, Clary DO, Reichardt LF,Gage FH. 1996. TrkA activation is sufficientto rescue axotomized cholinergic neurons.Neuron16:653–63
Luo J, West JR, Pantazis NJ. 1997. Nervegrowth factor and basic fibroblast growth fac-tor protect rat cerebellar granule cells in cul-ture against ethanol-induced cell death.Al-cohol Clin. Exp. Res.21:1108–20
Luo Z, Diaz B, Marshall MS, Avruch J. 1997.An intact Raf zinc finger is required for op-timal binding to processed Ras and for ras-dependent Raf activation in situ.Mol. Cell.Biol. 17:46–53
MacDonald JI, Gryz EA, Kubu CJ, Verdi JM,Meakin SO. 2000. Direct binding of the sig-naling adapter protein Grb2 to the activationloop tyrosines on the nerve growth factor re-ceptor tyrosine kinase, TrkA.J. Biol. Chem.275:18225–33
MacPhee IJ, Barker PA. 1997. Brain-derivedneurotrophic factor binding to the p75 neu-rotrophin receptor reduces TrkA signalingwhile increasing serine phosphorylation inthe TrkA intracellular domain.J. Biol. Chem.272:23547–51
Madison RD, Archibald SJ. 1994. Point sourcesof Schwann cells result in growth into a nerveentubulation repair site in the absence of ax-ons: effects of freeze-thawing.Exp. Neurol.128:266–75
Mahadeo D, Kaplan L, Chao MV, HempsteadBL. 1994. High affinity nerve growth factorbinding displays a faster rate of associationthan p140trk binding. Implications for multi-subunit polypeptide receptors.J. Biol. Chem.269:6884–91
Maggirwar SB, Sarmiere PD, Dewhurst S,Freeman RS. 1998. Nerve growth factor-dependent activation of NF-kappaB con-
tributes to survival of sympathetic neurons.J. Neurosci.18:10356–65
Majdan M, Lachance C, Gloster A, Aloyz R,Zeindler C, et al. 1997. Transgenic miceexpressing the intracellular domain of thep75 neurotrophin receptor undergo neuronalapoptosis.J. Neurosci.17:6988–98
Malcangio M, Garrett NE, Tomlinson DR.1997. Nerve growth factor treatment in-creases stimulus-evoked release of sensoryneuropeptides in the rat spinal cord.Eur. J.Neurosci.9:1101–4
Malinin NL, Boldin MP, Kovalenko AV, Wal-lach D. 1997. MAP3K-related kinase in-volved in NF-kappaB induction by TNF,CD95 and IL-1.Nature385:540–44
Mallett S, Barclay AN. 1991. A new super-family of cell surface proteins related to thenerve growth factor receptor.Immunol. To-day12:220–23
Marais R, Light Y, Mason C, Paterson H, OlsonMF, Marshall CJ. 1998. Requirement of Ras-GTP-Raf complexes for activation of Raf-1by protein kinase C.Science280:109–12
Marshall CJ. 1994. MAP kinase kinase kinase,MAP kinase kinase and MAP kinase.Curr.Opin. Genet. Dev.4:82–89
Marshall JS, Gomi K, Blennerhassett MG, Bie-nenstock J. 1999. Nerve growth factor modi-fies the expression of inflammatory cytokinesby mast cells via a prostanoid-dependentmechanism.J. Immunol.162:4271–76
Martin-Zanca D, Hughes SH, Barbacid M.1986a. A human oncogene formed by thefusion of truncated tropomyosin and proteintyrosine kinase sequences.Nature319:743–48
Martin-Zanca D, Mitra G, Long LK, BarbacidM. 1986b. Molecular characterization of thehuman trk oncogene.Cold Spring Harb.Symp. Quant. Biol.51:983–92
Martin-Zanca D, Oskam R, Mitra G, CopelandT, Barbacid M. 1989. Molecular andbiochemical characterization of the humantrk proto-oncogene.Mol. Cell. Biol.9:24–33
Martinex-Serrano A, Bjorklund A. 1996. Pro-tection of the neostriatum against excitotoxic
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1271
damage by neurotrophin-producing, geneti-cally modified neural stem cells.J. Neurosci.16:4604
Martınez-Murillo R, Fern´andez AP, BenturaML, Rodrigo J. 1998. Subcellular local-ization of low-affinity nerve growth factorreceptor-immunoreactive protein in adult ratpurkinje cells following traumatic injury.Exp. Brain Res.119:47–57
Matsuda M, Hashimoto Y, Muroya K,Hasegawa H, Kurata T, et al. 1994. CRKprotein binds to two guanine nucleotide-releasing proteins for the Ras family andmodulates nerve growth factor-induced ac-tivation of Ras in PC12 cells.Mol. Cell. Biol.14:5495–500
Mattson M. 1996. Calcium and free radicals:mediators of neurotrophic factor and ex-citatory transmitter-regulated developmentalplasticity and cell death.Perspect. Dev. Neu-robiol. 3:79–91
Mattson M, Marck R. 1996. Excitotoxicity andexcitoprotection in vitro.Adv. Neurol.71:1–30
Mattson MP. 1997. Neuroprotective signaltransduction: relevance to stroke.Neurosci.Biobehav. Rev.21:193–206
Mattson MP, Goodman Y, Luo H, Fu W, Fu-rukawa K. 1997. Activation of NF-kappaBprotects hippocampal neurons against oxida-tive stress-induced apoptosis: evidence forinduction of manganese superoxide dismu-tase and suppression of peroxynitrite produc-tion and protein tyrosine nitration.J. Neu-rosci. Res.49:681–97
Mattson MP, Lovell MA, Furukawa K, Markes-bery WR. 1995. Neurotrophic factors at-tenuate glutamate-induced accumulation ofperoxides, elevation of intracellular Ca2+concentration, and neurotoxicity and in-crease antioxidant enzyme activities in hip-pocampal neurons.J. Neurochem.65:1740–51
McAllister AK. 1999. Neurotrophins andsynaptic plasticity.Annu. Rev. Neurosci.22:295–318
McCormick F. 1994. Activators and effectors
of ras p21 proteins.Curr. Opin. Genet. Dev.4:71–76
McDonald NQ, Lapatto R, Murray-Rust J, Gun-ning J, Wlodawer A, Blundell TL. 1991. Newprotein fold revealed by a 2.3-A resolutioncrystal structure of nerve growth factor.Na-ture354:411–14
McMahon SB, Bennete DLH, Priestley JV,Shelton DL. 1995. The biological effects ofendogenous nerve growth factor on adult sen-sory neurons revealed by a trkA-IgG fusionmolecule.Nat. Med.1:774
McMartin DN, O’Connor JA. 1979. Ef-fect of age on axoplasmic transport ofcholinesterase in rat sciatic nerves.Mech.Ageing Dev.10:241–48
Meakin SO, MacDonald JI, Gryz EA, Kubu CJ,Verdi JM. 1999. The signaling adapter FRS-2 competes with Shc for binding to the nervegrowth factor receptor TrkA. A model for dis-criminating proliferation and differentiation.J. Biol. Chem.274:9861–70
Meakin SO, Shooter EM. 1992. The nervegrowth factor family of receptors.TrendsNeurosci.15:323–31
Meakin SO, Suter U, Drinkwater CC, WelcherAA, Shooter EM. 1992. The rat trk pro-tooncogene product exhibits properties char-acteristic of the slow nerve growth factor re-ceptor.Proc. Natl. Acad. Sci. USA89:2374–78
Meller N, Liu YC, Collins TL, Bonnefoy-Berard N, Baier G, et al. 1996. Direct inter-action between protein kinase C theta (PKCtheta) and 14-3-3 tau in T cells: 14-3-3 over-expression results in inhibition of PKC thetatranslocation and function.Mol. Cell. Biol.16:5782–91
Mendell LM. 1996. Neurotrophins and sen-sory neurons: role in development, mainte-nance and injury. A thematic summary.Phi-los. Trans. R. Soc. London Ser. B351:463–67
Mendell LM, Albers KM, Davis BM. 1999.Neurotrophins, nociceptors, and pain.Mi-crosc. Res. Technol.45:252–61
Menniti FS, Oliver KG, Putney JW Jr, ShearsSB. 1993. Inositol phosphates and cell
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1272 SOFRONIEW ¥ HOWE ¥ MOBLEY
signaling: new views of InsP5 and InsP6.Trends Biochem. Sci.18:53–56
Merry DE, Korsmeyer SJ. 1997. cl-2 gene fam-ily in the nervous system.Annu. Rev. Neu-rosci.20:245–67
Micera A, Vigneti E, Aloe L. 1998. Changesof NGF presence in nonneuronal cellsin response to experimental allergic en-cephalomyelitis in lewis rats.Exp. Neurol.154:41–46
Michael GJ, Kaya E, Averill S, Rattry M, ClayDO, Priestley JV. 1997. TrkA immunoreac-tive neurones in the rat spinal cord.J. Comp.Neurol.385:441–55
Middlemas DS, Meisenhelder J, Hunter T.1994. Identification of TrkB autophosphory-lation sites and evidence that phospholipaseC-gamma 1 is a substrate of the TrkB recep-tor. J. Biol. Chem.269:5458–66
Miller FD, Mathew TC, Toma JG. 1991. Reg-ulation of nerve growth factor receptor geneexpression by nerve growth factor in the de-veloping peripheral nervous system.J. CellBiol. 112:303–12
Miller FD, Speelman A, Mathew TC, Fabian J,Chang E, et al. 1994. Nerve growth factor de-rived from terminals selectively increases theratio of p75 to trkA NGF receptors on maturesympathetic neurons.Dev. Biol. 161:206–17
Miranti CK, Ginty DD, Huang G, ChatilaT, Greenberg ME. 1995. Calcium activatesserum response factor-dependent transcrip-tion by a Ras- and Elk-1-independent mech-anism that involves a Ca2+/calmodulin-dependent kinase.Mol. Cell. Biol.15:3672–84
Mirsky R, Jessen KR. 1999. The neurobiologyof Schwann cells.Brain Pathol.9:293–311
Mizuma H, Takagi K, Miyake K, Takagi N,Ishida K, et al. 1999. Microsphere embolism-induced elevation of nerve growth factor leveland appearance of nerve growth factor im-munoreativity in activated T-lymphocytes inthe rat brain.J. Neurosci. Res.55:749–61
Mobley WC, Rutkowski JL, Tennekoon GI,Buchanan K, Johnston M. 1985. Choline
acetyltransferase activity in striatum ofneonatal rats increased by nerve growth fac-tor. Science229:284–87
Moller DE, Xia CH, Tang W, Zhu AX,Jakubowski M. 1994. Human rsk iso-forms: cloning and characterization of tissue-specific expression.Am. J. Physiol.266:C351–59
Morrison ME, Mason CA. 1998. Granule neu-ron regulation of purkinje cell development:striking a balance between neurotrophin andglutamate signaling.J. Neurosci.18:3563–73
Mueller CG, Nordheim A. 1991. A protein do-main conserved between yeast MCM1 andhuman SRF directs ternary complex forma-tion. EMBO J.10:4219–29
Mufson EJ, Lavine N, Jaffar S, Kordower JH,Quirion R, Saragovi HU. 1997. Reduction inp140-TrkA receptor protein within the nu-cleus basalis and cortex in Alzheimer’s dis-ease.Exp. Neurol.146:91–103
Muragaki Y, Chou TT, Kaplan DR, TrojanowskiJQ, Lee VM. 1997. Nerve growth factor in-duces apoptosis in human medulloblastomacell lines that express TrkA receptors.J. Neu-rosci.17:530–42
Muroya K, Hattori S, Nakamura S. 1992. Nervegrowth factor induces rapid accumulationof the GTP-bound form of p21ras in ratpheochromocytoma PC12 cells.Oncogene7:277–81
Nakajima T, Fukamizu A, Takahashi J, GageFH, Fisher T, et al. 1996. The signal-dependent coactivator CBP is a nuclear targetfor pp90RSK.Cell 86:465–74
Neumann S, Woolf CJ. 1999. Regeneration ofdorsal column fibers into and beyond the le-sion site following adult spinal cord injury.Neuron23:83–91
Nishizuka Y. 1988. The molecular heterogene-ity of protein kinase C and its implicationsfor cellular regulation.Nature334:661–65
Nishizuka Y. 1992. Intracellular signaling byhydrolysis of phospholipids and activation ofprotein kinase C.Science258:607–14
Noda M, Ko M, Ogura A, Liu DG, Amano
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1273
T, et al. 1985. Sarcoma viruses carrying rasoncogenes induce differentiation- associatedproperties in a neuronal cell line.Nature318:73–75
Obermeier A, Bradshaw RA, Seedorf K,Choidas A, Schlessinger J, Ullrich A. 1994.Neuronal differentiation signals are con-trolled by nerve growth factor receptor/Trkbinding sites for SHC and PLC gamma.EMBO J.13:1585–90
Oberwetter JM, Conrad KE, Gutierrez-Hartmann A. 1993. The Ras and proteinkinase C signaling pathways are functionallyantagonistic in GH4 neuroendocrine cells.Mol. Endocrinol.7:915–23
O’Brien TS, Svendsen CN, Isacson O,Sofroniew MV. 1990. Loss of True blue la-belling from the medial septum followingtransection of the fimbria-fornix: evidencefor the death of cholinergic and noncholin-ergic neurons.Brain Res.508:249–56
Ong SH, Guy GR, Hadari YR, Laks S, Go-toh N, et al. 2000. FRS2 proteins recruitintracellular signaling pathways by bindingto diverse targets on fibroblast growth factorand nerve growth factor receptors.Mol. Cell.Biol. 20:979–89
Ono Y, Fujii T, Ogita K, Kikkawa U, IgarashiK, Nishizuka Y. 1988. The structure, expres-sion, and properties of additional membersof the protein kinase C family.J. Biol. Chem.263:6927–32
Osada S, Mizuno K, Saido TC, Akita Y,Suzuki K, et al. 1990. A phorbol ester recep-tor/protein kinase, nPKC eta, a new memberof the protein kinase C family predominantlyexpressed in lung and skin.J. Biol. Chem.265:22434–40
Osada S, Mizuno K, Saido TC, Suzuki K,Kuroki T, Ohno S. 1992. A new memberof the protein kinase C family, nPKC theta,predominantly expressed in skeletal muscle.Mol. Cell. Biol.12:3930–38
Oshima M, Sithanandam G, Rapp UR, GuroffG. 1991. The phosphorylation and activationof B-raf in PC12 cells stimulated by nervegrowth factor.J. Biol. Chem.266:23753–60
Otten U, Scully JL, Ehrhard PB, Gadient RA.1994. Neurotrophins: signals between thenervous and immune systems.Prog. BrainRes.103:293–305
Pang L, Sawada T, Decker SJ, Saltiel AR.1995. Inhibition of MAP kinase kinaseblocks the differentiation of PC-12 cells in-duced by nerve growth factor.J. Biol. Chem.270:13585–88
Pascual M, Rocamora N, Acs´ady L, FreundTF, Soriano E. 1998. Expression of nervegrowth factor and neurotrophin-3 mRNAsin hippocampal interneurons: morphologi-cal characterization, levels of expression, andcolocalization of nerve growth factor andneurotrophin-3.J. Comp. Neurol.395:73–90
Patel TD, Jackman A, Rice FL, Kucera J, SniderWD. 2000. Development of sensory neuronsin the absence of NGF/TrkA signaling in vivo[see comments].Neuron25:345–57
Payne DM, Rossomando AJ, Martino P, Er-ickson AK, Her JH, et al. 1991. Identi-fication of the regulatory phosphorylationsites in pp42/mitogen-activated protein ki-nase (MAP kinase).EMBO J.10:885–92
Pearson RCA, Sofroniew MV, Cuello AC, Pow-ell TPS, Eckenstein F, et al. 1983. Persis-tance of cholinergic neurons in the basal nu-cleus in a brain with senile dementia of theAlzheimer’s type demonstrated by immuno-histochemical staining for choline acetyl-transferase.Brain Res.289:375–79
Pechan PA, Chowdhury K, Gerdes W, SeifertW. 1993. Glutamate induces the growth fac-tors NGF and bFGF, the receptors FGF-R1and c-fos mRNA expression in rat astrocyteculture.Neurosci. Lett.16:111–14
Pechan PA, Chowdhury K, Seifert W. 1992.Free radicals induce gene expression of NGFand bFGF in rat astrocyte culture.NeuroRe-port 3:469–72
Peterson DA, Dickinson-Anson HA, LeppertJT, Lee KF, Gage FH. 1999. Central neuronalloss and behavioral impairment in mice lack-ing neurotrophin receptor p75.J. Comp. Neu-rol. 404:1–20
Petty B, Cornblath DR, Adornato BT, Chaudry
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1274 SOFRONIEW ¥ HOWE ¥ MOBLEY
V, Flexner C, et al. 1994. The effect of sytemi-cally administered recombinant human nervegrowth factor in healthy human subjects.Ann. Neurol.36:244–46
Pitts AF, Miller MW. 2000. Expression of nervegrowth factor, brain-derived neurotrophicfactor, and neurotrophin-3 in the somatosen-sory cortex of the mature rat: coexpressionwith high-affinity neurotrophin receptors.J.Comp. Neurol.418:241–54
Pizzorusso T, Berardi N, Rossi FM, Viegi A,Venstrom K, et al. 1999. TrkA activation inthe rat visual cortex by antirat trkA IgG pre-vents the effect of monocular deprivation.Eur J. Neurosci.11:204–12
Prakash N, Cohen-Cory S, Frostig RD. 1996.Rapid and opposite effects of BDNF andNGF on the functional organization of theadult cortex in vivo.Nature381:702–6
Qian X, Riccio A, Zhang Y, Ginty DD. 1998.Identification and characterization of novelsubstrates of Trk receptors in developing neu-rons.Neuron21:1017–29
Qui MS, Green SH. 1992. PC12 cell neuronaldifferentiation is associated with prolongedp21ras activity and consequent prolongedERK activity.Neuron9:705–17
Rabin SJ, Cleghon V, Kaplan DR. 1993.SNT, a differentiation-specific target of neu-rotrophic factor-induced tyrosine kinase ac-tivity in neurons and PC12 cells.Mol. Cell.Biol. 13:2203–13
Rabizadeh S, Oh J, Zhong LT, Yang J, BitlerCM, et al. 1993. Induction of apoptosis by thelow-affinity NGF receptor.Science261:345–48
Ramer MS, Priestley JV, McMahon SB. 2000.Functional regeneration of sensory axonsinto the adult spinal cord.Nature403:312–16
Ramirez S, Ait-Si-Ali S, Robin P, Trouche D,Harel-Bellan A. 1997. The CREB-bindingprotein (CBP) cooperates with the serumresponse factor for transactivation of thec-fos serum response element.J. Biol. Chem.272:31016–21
Ribon V, Saltiel AR. 1996. Nerve growth fac-tor stimulates the tyrosine phosphorylation
of endogenous Crk-II and augments its asso-ciation with p130Cas in PC-12 cells.J. Biol.Chem.271:7375–80
Riccio A, Ahn S, Davenport CM, Blendy JA,Ginty DD. 1999. Mediation by a CREB fam-ily transcription factor of NGF-dependentsurvival of sympathetic neurons.Science286:2358–61
Riccio A, Pierchala BA, Ciarello CL, GintyDD. 1997. An NGF-TrkA-mediated retro-grade signal to transcription factor CREBin sympathetic neurons.Science277:1097–100
Rice FL, Albers KM, Davis BM, Silos-SantiagoI, Wilkinson GA, et al. 1998. Differential de-pendency of unmyelinated and A delta epi-dermal and upper dermal innervation on neu-rotrophins, trk receptors, and p75LNGFR.Dev. Biol.198:57–81
Rich KM, Alexander TD, Pryor JC, HollowellJP. 1989. Nerve growth factor enhances re-generation through silicone chambers.Exp.Neurol.105:162–70
Richardson PM, Issa VM. 1984. Peripheral in-jury enhances entral regeneration of primarysensory neurones.Nature309:791–93
Robinson RC, Radziejewski C, Spraggon G,Greenwald J, Kostura MR, et al. 1999.The structures of the neurotrophin 4 homo-dimer and the brain-drived neurotrophic fac-tor/neurotrophin 4 heterodimer reveal a com-mon Trk-binding site.Protein Sci.8:2589–97
Robinson RC, Radziejewski C, Stuart DI, JonesEY. 1995. Structure of the brain-derived neu-rotrophic factor/neurotrophin 3 heterodimer.Biochemistry34:4139–46
Ross AH, Daou MC, McKinnon CA, CondonPJ, Lachyankar MB, et al. 1996. The neu-rotrophin receptor, gp75, forms a complexwith the receptor tyrosine kinase TrkA.J.Cell Biol. 132:945–53
Ross GM, Shamovsky IL, Lawrance G, Solc M,Dostaler SM, et al. 1998. Reciprocal modu-lation of TrkA and p75NTR affinity states ismediated by direct receptor interactions.Eur.J. Neurosci.10:890–98
Rothe M, Sarma V, Dixit VM, Goeddel DV.
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1275
1995. TRAF2-mediated activation of NF-kappa B by TNF receptor 2 and CD40.Sci-ence269:1424–27
Rothstein JD, Dykes-Hoberg M, Pardo CA,Bristol LA, Jin L, et al. 1996. Knockout ofglutamate transporters reveals a major rolefor astroglial transport in excitotoxicity andclearance of glutamate.Neuron16:675–86
Rozakis-Adcock M, McGlade J, Mbamalu G,Pelicci G, Daly R, et al. 1992. Associationof the Shc and Grb2/Sem5 SH2-containingproteins is implicated in activation of theRas pathway by tyrosine kinases.Nature360:689–92
Ruit KG, Osborne PA, Schmidt RE, JohnsonEM Jr, Snider WD. 1990. Nerve growth fac-tor regulates sympathetic ganglion cell mor-phology and survival in the adult mouse.Neu-roscience10:2412–19
Ryden M, Ibanez CF. 1997. A second deter-minant of binding to the p75 neurotrophinreceptor revealed by alanine-scanning muta-genesis of a conserved loop in nerve growthfactor.J. Biol. Chem.272:33085–91
Saper CB, Wainer BH, German DC. 1987. Ax-onal and transneuronal transport in the trans-mission of neurological disease: potentialrole in systems degeneration.Neuroscience23:389–97
Schechter AL, Bothwell MA. 1981. Nervegrowth factor receptors on PC12 cells: evi-dence for two receptor classes with differingcytoskeletal association.Cell 24:867–74
Schneider A, Martin-Villalba A, Weih F, Vo-gel J, Wirth T, Schwaninger M. 1999. NF-kappaB is activated and promotes cell deathin focal cerebral ischemia.Nat. Med.5:554–59
Schonwasser DC, Marais RM, Marshall CJ,Parker PJ. 1998. Activation of the mitogen-activated protein kinase/extracellular signal-regulated kinase pathway by conventional,novel, and atypical protein kinase C isotypes.Mol. Cell. Biol.18:790–98
Schouten GJ, Vertegaal AC, Whiteside ST, Is-rael A, Toebes M, et al. 1997. IkappaB al-pha is a target for the mitogen-activated 90
kDa ribosomal S6 kinase.EMBO J.16:3133–44
Schumacher JM, Short MP, Hyman BT, Break-field XO, Isacson O. 1991. Intracerebral im-plantation of nerve growth factor-producingfibroblasts protects striatum against neuro-toxic levels of excitatory amino acids.Neu-roscience43:567–70
Schwaninger M, Sallmann S, Petersen N,Schneider A, Prinz S, et al. 1999. Bradykinininduces interleukin-6 expression in astro-cytes through activation of nuclear factor-kappaB.J. Neurochem.73:1461–66
Scott SA, Mufson EJ, Weingartner JA, SkauKA, Crutcher KA. 1995. Nerve growth fac-tor in Alzheimer’s disease: increased levelsthroughout the brain coupled with declinesin nucleus basalis.J. Neurosci.15:6213–21
Seibenhener ML, Roehm J, White WO, NeidighKB, Vandenplas ML, Wooten MW. 1999.Identification of Src as a novel atypical pro-tein kinase C-interacting protein.Mol. Cell.Biol. Res. Commun.2:28–31
Semkova I, Krieglstein J. 1999. Cilliary neu-rotrophic factor enhances the expression ofNGF and p75 low-affinity NGF receptor inastrocytes.Brain Res.838:184–92
Sheng M, Greenberg ME. 1990. The regulationand function of c-fos and other immediateearly genes in the nervous system.Neuron4:477–85
Shigeno T, Mima T, Takakura K, Graham DI,Kato G, et al. 1991. Amelioration of de-layed neuronal death in the hippocampus bynerve growth factor.J. Neurosci.11:2914–19
Shu XQ, Mendell LM. 1999. Neurotrophinsand hyperalgesia.Proc. Natl. Acad. Sci. USA96:7693–96
Siliprandi R, Canella R, Carmignoto G.1993. Nerve growth factor promotes func-tional recovery of retinal ganglion cells af-ter ischemia.Invest. Ophthalmol. Vis. Sci.34:3232–45
Simone M, De Santis S, Vigneti E, Papa G,Amadori S, Aloe L. 1999. Nerve growth
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1276 SOFRONIEW ¥ HOWE ¥ MOBLEY
factor: a survey of activity on immune andhematopoietic cells.Hematol. Oncol.17:1–10
Singer CA, Figueroa-Masot XA, Batchelor RH,Dorsa DM. 1999. The mitogen-activated pro-tein kinase pathway mediates estrogen neuro-protection after glutamate toxicity in primarycortical neurons.J. Neurosci.19:2455
Singh M, Setalo GJ, Guan X, Warren M, Toran-Allerand CD. 1999. Estrogen-induced acti-vation of mitogen-activated protein kinase incerebral cortical explants: convergence of es-trogen and neurotrophin signaling pathways.J. Neurosci.19:1179–88
Smeyne RJ, Klein R, Schnapp A, Long LK,Bryant S, et al. 1994. Severe sensory andsympathetic neuropathies in mice carrying adisrupted Trk/NGF receptor gene [see com-ments].Nature368:246–49
Smith CA, Farrah T, Goodwin RG. 1994. TheTNF receptor superfamily of cellular and vi-ral proteins: activation, costimulation, anddeath.Cell 76:959–62
Snider WD. 1994. Functions of the neu-rotrophins during nervous system develop-ment: what the knockouts are teaching us.Cell 77:627–38
Sofroniew MV. 1999. Neuronal responses to ax-otomy. InCNS Regeneration: Basic Scienceand Clinical Advances, ed. M Tuszynski, JKordower, pp. 3–26. San Diego, CA: Aca-demic
Sofroniew MV, Cooper JD, Svendsen CN,Crossman P, Ip NY, et al. 1993. Atrophy butnot death of adult septal cholinergic neuronsafter ablation of target capacity to producemRNAs for NGF, BDNF and NT-3.J. Neu-rosci.13:5263–76
Sofroniew MV, Galletly NP, Isacson O, Svend-sen CN. 1990. Survival of adult basal fore-brain cholinergic neurons after loss of targetneurons.Science247:338–42
Sofroniew MV, Mobley WC. 1993. On the pos-sibility of positive feedback in trophic inter-actions between afferent and target neurons.Semin. Neurosci.5:309–12
Sofroniew MV, Pearson RCA, Eckenstein F,
Cuello AC, Powell TPS. 1983. Retrogradechanges in the basal forebrain of the rat fol-lowing cortical damage.Brain Res.289:370–74
Song HY, Dunbar JD, Donner DB. 1994. Ag-gregation of the intracellular domain of thetype 1 tumor necrosis factor receptor definedby the two-hybrid system.J. Biol. Chem.269:22492–95
Sozeri O, Vollmer K, Liyanage M, Frith D, KourG, et al. 1992. Activation of the c-Raf proteinkinase by protein kinase C phosphorylation.Oncogene7:2259–62
Steller H. 1995. Mechanisms and genes of cel-lular suicide.Science267:1445–49
Stephens L, Anderson K, Stokoe D, Erdjument-Bromage H, Painter GF, et al. 1998. Proteinkinase B kinases that mediate phosphatidyli-nositol 3,4,5-trisphosphate-dependent acti-vation of protein kinase B.Science279:710–14
Stephens RM, Loeb DM, Copeland TD, Paw-son T, Greene LA, Kaplan DR. 1994. Trkreceptors use redundant signal transductionpathways involving SHC and PLC-gamma 1to mediate NGF responses.Neuron12:691–705
Stokoe D, Stephens LR, Copeland T, GaffneyPR, Reese CB, et al. 1997. Dual roleof phosphatidylinositol-3,4,5-trisphosphatein the activation of protein kinase B.Science277:567–70
Strauss S, Otten U, Joggerst B, Pluss K, Volk B.1968. Increased levels of nerve growth factor(NGF) protein and mRNA and reactive glio-sis following kainic acid injection into the ratstriatum.Neurosci. Lett.168:193–96
Strijbos P, Rothwell N. 1995. Interleukin-1 betaattenuates excitatory amino acid-inducedneurodegeneration in vitro: involvement ofnerve growth factor.J. Neurosci.15:3468
Sturgill TW, Ray LB, Erikson E, MallerJL. 1988. Insulin-stimulated MAP-2 kinasephosphorylates and activates ribosomal pro-tein S6 kinase II.Nature334:715–18
Susaki Y, Shimizu S, Katakura K, WatanabeN, Kawamoto K, et al. 1996. Functional
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1277
properties of murine macrophages promotedby nerve growth factor.Blood88:4630–37
Susaki Y, Tanaka A, Honda E, Matsuda H. 1998.Nerve growth factor modulates Fc gammareceptor expression on murine macrophageJ774A.1 cells.J. Vet. Med. Sci.60:87–91
Sutter A, Riopelle RJ, Harris-Warrick RM,Shooter EM. 1979. Nerve growth factorreceptors. Characterization of two distinctclasses of binding sites on chick embryo sen-sory ganglia cells.J. Biol. Chem.254:5972–82
Svendsen CN, Kew JNC, Staley K, SofroniewMV. 1994. Death of developing septal cholin-ergic neurons following NGF withdrawal invitro: protection by protein synthesis inhibi-tion. J. Neurosci.14:75–87
Taiwo YO, Levine JD, Burch R, Woo JE, Mob-ley WC. 1991. Hyperalgesia induced in therat by the amino-terminal octapeptide ofnerve growth factor.Proc. Natl. Acad. Sci.USA88:5144–48
Taniuchi M, Clark HB, Johnson EM Jr. 1986.Induction of nerve growth factor receptorin Schwann cells after axotomy.Proc. Natl.Acad. Sci. USA83:4094–98
Taniuchi M, Clark HB, Schweitzer JB, John-son EM. 1988. Expression of nerve growthfactor receptors by Schwann cells of axo-tomized peripheral nerves: ultrastructural lo-cation, suppression by axonal contact, andbinding properties.J. Neurosci.8:664–81
Tartaglia LA, Ayres TM, Wong GH, GoeddelDV. 1993. A novel domain within the 55 kdTNF receptor signals cell death.Cell74:845–53
Teng KK, Lander H, Fajardo JE, Hanafusa H,Hempstead BL, Birge RB. 1995. v-Crk mod-ulation of growth factor-induced PC12 celldifferentiation involves the Src homology 2domain of v-Crk and sustained activationof the Ras/mitogen-activated protein kinasepathway.J. Biol. Chem.270:20677–85
Thomas SM, Brugge JS. 1997. Cellular func-tions regulated by Src family kinases.Annu.Rev. Cell Dev. Biol.13:513–609
Thomas SM, Hayes M, D’Arcangelo G, Arm-
strong RC, Meyer BE, et al. 1991. Inductionof neurite outgrowth by v-src mimics crit-ical aspects of nerve growth factor-induceddifferentiation. Mol. Cell. Biol. 11:4739–50
Thuveson M, Albrecht D, Zurcher G, AndresAC, Ziemiecki A. 1995. iyk, a novel intra-cellular protein tyrosine kinase differentiallyexpressed in the mouse mammary gland andintestine.Biochem. Biophys. Res. Commun.209:582–89
Toran-Allerand CD, Miranda RC, BenthamWD, Sohrabji F, Brown TJ, et al. 1992. Es-trogen receptors colocalize with low-affinitynerve growth factor receptors in cholinergicneurons of the basal forebrain.Proc. Natl.Acad. Sci. USA89:4668–72
Torcia M, Bracci-Laudiero L, Lucibello M,Nencioni L, Labardi D, et al. 1996. Nervegrowth factor is an autocrine survival factorfor memory B lymphocytes.Cell 85:345–56
Torres M, Bogenmann E. 1996. Nerve growthfactor induces a multimeric TrkA recep-tor complex in neuronal cells that includesCrk, SHC and PLC-gamma 1 but excludesP130CAS.Oncogene12:77–86
Traverse S, Cohen P. 1994. Identification of a la-tent MAP kinase kinase kinase in PC12 cellsas B-raf.FEBS Lett.350:13–18
Traverse S, Gomez N, Paterson H, MarshallC, Cohen P. 1992. Sustained activation ofthe mitogen-activated protein (MAP) kinasecascade may be required for differentiationof PC12 cells. Comparison of the effects ofnerve growth factor and epidermal growthfactor.Biochem. J.288:351–55
Treisman R. 1992. The serum response element.Trends Biochem. Sci.17:423–26
Trim N, Morgan S, Evans M, Issa R, Fine D,et al. 2000. Hepatic stellate cells expressthe low affinity nerve growth factor recep-tor p75 and undergo apoptosis in responseto nerve growth factor stimulation.Am. J.Pathol.156:1235–43
Tsui-Pierchala BA, Ginty DD. 1999. Charac-terization of an NGF-P-TrkA retrograde-signaling complex and age-dependent
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1278 SOFRONIEW ¥ HOWE ¥ MOBLEY
regulation of TrkA phosphorylation in sym-pathetic neurons.Neuroscience19:8207–18
Tuszynski MH, Armstrong DM, Gage FH.1990. Basal forebrain cell loss following fim-bria/fornix transection.Brain Res.508:241–48
Tuszynski MH, Gage FH. 1995a. Bridginggrafts and transient nerve growth factor infu-sions promote long-term central nervous sys-tem neuronal rescue and partial functional re-covery.Proc. Natl. Acad. Sci. USA92:4621–25
Tuszynski MH, Gage FH. 1995b. Maintain-ing the neuronal phenotype after injury inthe adult CNS. Neurotrophic factors, axonalgrowth substrates, and gene therapy.Mol.Neurobiol.10:151–67
Tuttle R, O’Leary DDM. 1998. Neurotrophinsrapidly modulate growth cone response to theaxon guidance molecule, collapsin-1.Mol.Cell. Neurosci.11:1–8
Twiss JL, Shooter EM. 1995. Nerve growth fac-tor promotes neurite regeneration in PC12cells by translational control.J. Neurochem.64:550–57
Ueffing M, Lovric J, Philipp A, Mischak H,Kolch W. 1997. Protein kinase C-epsilon as-sociates with the Raf-1 kinase and inducesthe production of growth factors that stimu-late Raf-1 activity.Oncogene15:2921–27
Ure DR, Campenot RB. 1997. Retrograde trans-port and steady-state distribution of125I-nerve growth factor in rat sympathetic neu-rons in compartmented cultures.J. Neurosci.17:1282–90
Urfer R, Tsoulfas P, Soppet D, Escandon E,Parada LF, Presta LG. 1994. The binding epi-topes of neurotrophin-3 to its receptors trkCand gp75 and the design of a multifunctionalhuman neurotrophin.EMBO J. 13:5896–909
Vaillancourt RR, Gardner AM, Johnson GL.1994. B-Raf-dependent regulation of theMEK-1/mitogen-activated protein kinasepathway in PC12 cells and regulation bycyclic AMP. Mol. Cell. Biol.14:6522–30
Vaillant AR, Mazzoni I, Tudan C, Boudreau
M, Kaplan DR, Miller FD. 1999. Depolar-ization and neurotrophins converge on thephosphatidylinositol 3-kinase-Akt pathwayto synergistically regulate neuronal survival.J. Cell Biol.146:955–66
Van Aelst L, Barr M, Marcus S, PolverinoA, Wigler M. 1993. Complex formation be-tween RAS and RAF and other protein kina-ses.Proc. Natl. Acad. Sci. USA90:6213–17
Van Antwerp DJ, Martin SJ, Kafri T, Green DR,Verma IM. 1996. Suppression of TNF-alpha-induced apoptosis by NF-kappaB.Science274:787–89
van der Hoeven PCJ, van der Wal JCM, Ru-urs P, van Dijk MCM, van Blitterswijk WJ.2000. 14-3-3 isotypes facilitate coupling ofprotein kinase C-zeta to Raf-1: negative reg-ulation by 14-3-3 phosphorylation.Biochem.J. 345:297–306
van Dijk MC, Hilkmann H, van BlitterswijkWJ. 1997. Platelet-derived growth factoractivation of mitogen-activated protein ki-nase depends on the sequential activation ofphosphatidylcholine-specific phospholipaseC, protein kinase C-zeta and Raf-1.Biochem.J. 325:303–7
Venero JL, Knusel B, Beck KD, Hefti F. 1994.Expression of neurotrophin and trk receptorgenes in adult rats with fimbria transections:effect of intraventricular nerve growth factorand brain-derived neurotrophic factor admin-istration.Neuroscience59:797–815
Verdi JM, Anderson DJ. 1994. Neurotrophinsregulate sequential changes in neurotrophinreceptor expression by sympathetic neuro-blasts.Neuron13:1359–72
Verge VM, Gratto KA, Karchewski LA,Richardson PM. 1996. Neurotrophins andnerve injury in the adult.Philos. Trans. R.Soc. London Ser. B351:423–30
Verge VM, Merlio JP, Grondin J, Ernfors P,Persson H, et al. 1992. Colocalization of NGFbinding sites, trk mRNA, and low-affinityNGF receptor mRNA in primary sensoryneurons: responses to injury and infusion ofNGF.J. Neurosci.12:4011–22
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1279
Verge VMK, Richardson PM, Benoit R, Ri-opelle RJ. 1989. Histochemical characteri-zation of sensory neurons with high-affinityreceptors for nerve growth factor.J. Neuro-cytol.18:583–91
Verge VM, Richardson PM, Wiesenfeld-HallinZ, Hokfelt T. 1995. Differential influence ofnerve growth factor on neuropeptide expres-sion in vivo: a novel role in peptide suppres-sion in adult sensory neurons.J. Neurosci.15:2081–96
Vetter ML, Martin-Zanca D, Parada LF, BishopJM, Kaplan DR. 1991. Nerve growth factorrapidly stimulates tyrosine phosphorylationof phospholipase C-gamma 1 by a kinase ac-tivity associated with the product of the trkprotooncogene.Proc. Natl. Acad. Sci. USA88:5650–54
von Bussmann KA, Sofroniew MV. 1999. Re-expression of p75NTR by adult motor neu-rons after axotomy is triggered by retrogradetransport of a positive signal from regenerat-ing peripheral nerve.Neuroscience91:273–81
Wada K, Okada N, Yamamura Y, Koizumi S.1996. Nerve growth factor induces resistanceof PC12 cells to nitric oxide cytotoxicity.Neurochem. Int.29:461–67
Wang CY, Mayo MW, Baldwin AS Jr. 1996.TNF- and cancer therapy-induced apoptosis:potentiation by inhibition of NF-kappaB.Sci-ence274:784–87
Wang H, Tessier-Lavigne M. 1999. En pas-sant neurotrophic action of an intermediateaxonal target in the developing mammalianCNS.Nature401:765–69
Watanabe-Fukunaga R, Brannan CI, CopelandNG, Jenkins NA, Nagata S. 1992. Lympho-proliferation disorder in mice explained bydefects in Fas antigen that mediates apopto-sis.Nature356:314–17
Watson FL, Heerssen HM, Moheban DB,Lin MZ, Sauvageot CM, et al. 1999.Rapid nuclear responses to target-derivedneurotrophins require retrograde transportof ligand-receptor complex.J. Neurosci.9:7889–900
Weidner KM, Di Cesare S, Sachs M, Brink-mann V, Behrens J, Birchmeier W. 1996.Interaction between Gab1 and the c-Met re-ceptor tyrosine kinase is responsible for ep-ithelial morphogenesis.Nature384:173–76
Welch H, Maridonneau-Parini I. 1997. Lyn andFgr are activated in distinct membrane frac-tions of human granulocytic cells.Oncogene15:2021–29
Whitehouse PJ, Price DL, Struble RG,Clark AW, Coyle JT, DeLong MR. 1982.Alzheimer’s disease and senile dementia:loss of neurons in the basal forebrain.Sci-ence215:1237–39
Wiesmann C, Ultsch MH, Bass SH, de Vos AM.1999. Crystal structure of nerve growth factorin complex with the ligand-binding domainof the TrkA receptor.Nature401:184–88
Williams LR, Varon S, Peterson GM, Wic-torin K, Fischer W, et al. 1986. Continu-ous infusion of nerve growth factor preventsbasal forebrain neuronal death after fimbriafornix transection.Proc. Natl. Acad. Sci. USA83:9231–35
Wills Z, Marr L, Zinn K, Goodman CS, VanVactor D. 1999. Profilin and the Abl tyrosinekinase are required for motor axon outgrowthin the Drosophila embryo.Neuron22:291–99
Wilson JX. 1997. Antioxidant defense of thebrain: a role for astrocytes.Can. J. Physiol.Pharmacol.75:1149–63
Winkler J, Ramirez GA, Thal LJ, Waite JJ.2000. Nerve growth factor (NGF) aug-ments cortical and hippocampal cholinergicfunctioning afer p75NGF receptor-mediateddeafferentation but impairs inhibitory avoid-ance and induces fear-related behaviors.J.Neurosci.20:834–44
Wolf DE, McKinnon CA, Daou MC, StephensRM, Kaplan DR, Ross AH. 1995. Interac-tion with TrkA immobilizes gp75 in the highaffinity nerve growth factor receptor com-plex.J. Biol. Chem.270:2133–38
Wood JN. 1995. Regulation of NF-kappa B ac-tivity in rat dorsal root ganglia and PC12 cellsby tumour necrosis factor and nerve growthfactor.Neurosci Lett.192:41–44
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
1280 SOFRONIEW ¥ HOWE ¥ MOBLEY
Wood KW, Sarnecki C, Roberts TM, Blenis J.1992. ras mediates nerve growth factor recep-tor modulation of three signal-transducingprotein kinases: MAP kinase, Raf-1, andRSK.Cell 68:1041–50
Wood SJ, Pritchard J, Sofroniew MV. 1990. Re-expression of nerve growth factor receptorafter axonal injury recapitulates a develop-mental event in motor neurons: differentialregulation when regeneration is allowed orprevented.Eur. J. Neurosci.2:650–57
Woodruff NR, Neet KE. 1986. Beta nervegrowth factor binding to PC12 cells. Asso-ciation kinetics and cooperative interactions.Biochemistry25:7956–66
Woolf C, Ma Q, Allchorne A, Poole S. 1996.Peripheral cell types contributing to the hy-peralgesic action of nerve growth factor ininflammation.J. Neurosci.16:2716–23
Woolf CJ, Safieh-Garabedian B, Ma QP, CrillyP, Winter J. 1994. Nerve growth factorcontributes to the generation of inflamma-tory sensory hypersensitivity.Neuroscience62:327–31
Wooten MW, Seibenhener ML, Neidigh KB,Vandenplas ML. 2000. Mapping of atypicalprotein kinase C within the nerve growth fac-tor signaling cascade: relationship to differ-entiation and survival of PC12 cells.Mol.Cell. Biol.20:4494–504
Wooten MW, Seibenhener ML, Zhou G, Van-denplas ML, Tan TH. 1999. Overexpres-sion of atypical PKC in PC12 cells enhancesNGF- responsiveness and survival through anNF-kappaB dependent pathway.Cell DeathDiffer. 6:753–64
Wooten MW, Zhou G, Seibenhener ML, Cole-man ES. 1994. A role for zeta protein kinaseC in nerve growth factor-induced differentia-tion of PC12 cells.Cell Growth Differ.5:395–403
Wooten MW, Zhou G, Wooten MC, Seiben-hener ML. 1997. Transport of protein kinaseC isoforms to the nucleus of PC12 cells bynerve growth factor: association of atypicalzeta-PKC with the nuclear matrix.J. Neu-rosci. Res.49:393–403
Wyatt S, Shooter EM, Davies AM. 1990. Ex-pression of the NGF receptor gene in sensoryneurons and their cutaneous targets prior toand during innervation.Neuron4:421–27
Xiang J, Chao DT, Korsmeyer SJ. 1996. BAX-induced cell death may not require inter-leukin 1 beta-converting enzyme-like protea-ses.Proc. Natl. Acad. Sci. USA93:14559–63
Xing J, Ginty DD, Greenberg ME. 1996. Cou-pling of the RAS-MAPK pathway to gene ac-tivation by RSK2, a growth factor-regulatedCREB kinase.Science273:959–63
Xing J, Kornhauser JM, Xia Z, Thiele EA,Greenberg ME. 1998. Nerve growth fac-tor activates extracellular signal-regulated ki-nase and p38 mitogen-activated protein ki-nase pathways to stimulate CREB serine 133phosphorylation.Mol. Cell. Biol. 18:1946–55
Xue L, Murray JH, Tolkovsky AM. 2000.The Ras/phosphatidylinositol 3-kinase andRas/ERK pathways function as independentsurvival modules each of which inhibits a dis-tinct apoptotic signaling pathway in sympa-thetic neurons.J. Biol. Chem.275:8817–24
Yamashita H, Avraham S, Jiang S, Dikie I,Avraham H. 1999a. The Csk homologous ki-nase associates with TrkA receptors and isinvolved in neurite outgrowth of PC12 cells.J. Biol. Chem.274:15059–65
Yamashita T, Tucker KL, Barde Y. 1999b. Neu-rotrophin binding to the p75 receptor modu-lates rho activity and axonal outgrowth.Neu-ron 24:585–93
Yano H, Cong F, Birge RB, Goff SP, Chao MV.2000. Association of the Abl tyrosine kinasewith the Trk nerve growth factor receptor.J.Neurosci. Res.59:356–64
Yao R, Cooper GM. 1995. Requirement forphosphatidylinositol-3 kinase in the preven-tion of apoptosis by nerve growth factor.Sci-ence267:2003–6
Ye X, Mehlen P, Rabizadeh S, VanArsdale T,Zhang H, et al. 1999. TRAF family proteinsinteract with the common neurotrophin re-ceptor and modulate apoptosis induction.J.Biol. Chem.274:30202–8
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

P1: GDL
April 13, 2001 18:17 Annual Reviews AR121-40
NGF, NEUROPROTECTION, AND REPAIR 1281
Yeo TT, Chua-Couzens J, Butcher LL, Bre-desen DE, Cooper JD, et al. 1997. Absenceof p75NTR causes increased basal forebraincholinergic neuron size, choline acetyltrans-ferase activity, and target innervation.J. Neu-rosci.17:7594–605
Yoon SO, Casaccia-Bonnefil P, Carter B, ChaoMV. 1998. Competitive signaling betweenTrkA and p75 nerve growth factor recep-tors determines cell survival.J. Neurosci.18:3273–81
York RD, Yao H, Dillon T, Ellig CL, Eckert SP,et al. 1998. Rap1 mediates sustained MAPkinase activation induced by nerve growthfactor.Nature392:622–26
Yoshida K, Gage FH. 1991. Fibroblast growthfactors stimulate nerve growth factor synthe-sis and secretion by astrocytes.Brain Res.538:118–26
Yoshida K, Gage FH. 1992. Cooperative regu-lation of nerve growth factor synthesis andsecretion in fibroblasts and astrocytes by fi-broblast growth factor and other cytokines.Brain Res.569:14–25
Yuen EC, Mobley WC. 1996. Therapeutic
potential of neurotrophic factors for neu-rological disorders.Ann. Neurol.40:346–54
Zafra F, Castren E, Thoenen H, LindholmD. 1991. Interplay between glutamate andgamma-aminobutyric acid transmitter sys-tems in the physiological regulation of brain-derived neurotrophic factor and nerve growthfactor in hippocampal neurons.Proc. Natl.Acad. Sci. USA88:10037–41
Zha J, Harada H, Yang E, Jockel J, KorsmeyerSJ. 1996. Serine phosphorylation of death ag-onist BAD in response to survival factor re-sults in binding to 14-3-3 not BCL-X(L).Cell87:619–28
Zhao Y, Bjorbaek C, Moller DE. 1996. Regu-lation and interaction of pp90(rsk) isoformswith mitogen-activated protein kinases.J.Biol. Chem.271:29773–79
Zhao Y, Bjorbaek C, Weremowicz S, MortonCC, Moller DE. 1995. RSK3 encodes a novelpp90rsk isoform with a unique N-terminalsequence: growth factor-stimulated kinasefunction and nuclear translocation.Mol. Cell.Biol. 15:4353–63
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

Annual Review of Neuroscience Volume 24, 2001
CONTENTSPDZ DOMAINS AND THE ORGANIZATION OF SUPRAMOLECULAR COMPLEXES, Morgan Sheng, Carlo Sala 1
THE ROLE AND REGULATION OF ADENOSINE IN THE CENTRAL NERVOUS SYSTEM, Thomas V. Dunwiddie, Susan A. Masino
31
LOCALIZATION AND GLOBALIZATION IN CONSCIOUS VISION, S. Zeki 57
GLIAL CONTROL OF NEURONAL DEVELOPMENT, Greg Lemke 87TOUCH AND GO: Decision-Making Mechanisms in Somatosensation, Ranulfo Romo, Emilio Salinas 107
SYNAPTIC MODIFICATION BY CORRELATED ACTIVITY: Hebb''s Postulate Revisited, Guo-qiang Bi, Mu-ming Poo 139
AN INTEGRATIVE THEORY OF PREFRONTAL CORTEX FUNCTION, Earl K. Miller, Jonathan D. Cohen 167
THE PHYSIOLOGY OF STEREOPSIS, B. G. Cumming, G. C. DeAngelis 203
PARANEOPLASTIC NEUROLOGIC DISEASE ANTIGENS: RNA-Binding Proteins and Signaling Proteins in Neuronal Degeneration, Kiran Musunuru, Robert B. Darnell
239
ODOR ENCODING AS AN ACTIVE, DYNAMICAL PROCESS: Experiments, Computation, and Theory, Gilles Laurent, Mark Stopfer, Rainer W Friedrich, Misha I Rabinovich, Alexander Volkovskii, Henry DI Abarbanel
263
PROTEIN SYNTHESIS AT SYNAPTIC SITES ON DENDRITES, Oswald Steward, Erin M. Schuman 299
SIGNALING AND TRANSCRIPTIONAL MECHANISMS IN PITUITARY DEVELOPMENT, Jeremy S. Dasen, Michael G. Rosenfeld
327
NEUROPEPTIDES AND THE INTEGRATION OF MOTOR RESPONSES TO DEHYDRATION , Alan G. Watts 357
THE DEVELOPMENTAL BIOLOGY OF BRAIN TUMORS, Robert Wechsler-Reya, Matthew P. Scott 385
TO EAT OR TO SLEEP? OREXIN IN THE REGULATION OF FEEDING AND WAKEFULNESS, Jon T. Willie, Richard M. Chemelli, Christopher M. Sinton, Masashi Yanagisawa
429
SPATIAL PROCESSING IN THE BRAIN: The Activity of Hippocampal Place Cells, Phillip J. Best, Aaron M. White, Ali Minai 459
THE VANILLOID RECEPTOR: A Molecular Gateway to the Pain Pathway, Michael J Caterina, David Julius 487
PRION DISEASES OF HUMANS AND ANIMALS: Their Causes and Molecular Basis, John Collinge 519
VIKTOR HAMBURGER AND RITA LEVI-MONTALCINI: The Path to the Discovery of Nerve Growth Factor, W. Maxwell Cowan 551
EARLY DAYS OF THE NERVE GROWTH FACTOR PROTEINS, Eric M. Shooter 601
SEQUENTIAL ORGANIZATION OF MULTIPLE MOVEMENTS: Involvement of Cortical Motor Areas, Jun Tanji 631
INFLUENCE OF DENDRITIC CONDUCTANCES ON THE INPUT-OUTPUT PROPERTIES OF NEURONS, Alex Reyes 653
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.

NEUROTROPHINS: Roles in Neuronal Development and Function, Eric J Huang, Louis F Reichardt 677
CONTRIBUTIONS OF THE MEDULLARY RAPHE AND VENTROMEDIAL RETICULAR REGION TO PAIN MODULATION AND OTHER HOMEOSTATIC FUNCTIONS, Peggy Mason
737
ACTIVATION, DEACTIVATION, AND ADAPTATION IN VERTEBRATE PHOTORECEPTOR CELLS, Marie E Burns, Denis A Baylor 779ACTIVITY-DEPENDENT SPINAL CORD PLASTICITY IN HEALTH AND DISEASE, Jonathan R Wolpaw, Ann M Tennissen 807QUANTITATIVE GENETICS AND MOUSE BEHAVIOR, Jeanne M Wehner, Richard A Radcliffe, Barbara J Bowers 845
EARLY ANTERIOR/POSTERIOR PATTERNING OF THE MIDBRAIN AND CEREBELLUM, Aimin Liu, Alexandra L Joyner 869
NEUROBIOLOGY OF PAVLOVIAN FEAR CONDITIONING, Stephen Maren 897
{{alpha}}-LATROTOXIN AND ITS RECEPTORS: Neurexins and CIRL/Latrophilins, Thomas C Südhof 933IMAGING AND CODING IN THE OLFACTORY SYSTEM, John S Kauer, Joel White 963
THE ROLE OF THE CEREBELLUM IN VOLUNTARY EYE MOVEMENTS, Farrel R Robinson, Albert F Fuchs 981
ROLE OF THE REELIN SIGNALING PATHWAY IN CENTRAL NERVOUS SYSTEM DEVELOPMENT, Dennis S Rice, Tom Curran 1005
HUMAN BRAIN MALFORMATIONS AND THEIR LESSONS FOR NEURONAL MIGRATION, M Elizabeth Ross, Christopher A Walsh 1041
MORPHOLOGICAL CHANGES IN DENDRITIC SPINES ASSOCIATED WITH LONG-TERM SYNAPTIC PLASTICITY, Rafael Yuste, Tobias Bonhoeffer
1071
STOPPING TIME: The Genetics of Fly and Mouse Circadian Clocks, Ravi Allada, Patrick Emery, Joseph S. Takahashi, Michael Rosbash
1091
NEURODEGENERATIVE TAUOPATHIES, Virginia M-Y Lee, Michel Goedert, John Q Trojanowski 1121
MATERNAL CARE, GENE EXPRESSION, AND THE TRANSMISSION OF INDIVIDUAL DIFFERENCES IN STRESS REACTIVITY ACROSS GENERATIONS, Michael J Meaney
1161
NATURAL IMAGE STATISTICS AND NEURAL REPRESENTATION, Eero P Simoncelli, Bruno A Olshausen 1193
Nerve Growth Factor Signaling, Neuroprotection, and Neural Repair, Michael V Sofroniew, Charles L Howe, William C Mobley 1217
FLIES, GENES, AND LEARNING, Scott Waddell, William G Quinn 1283
Ann
u. R
ev. N
euro
sci.
2001
.24:
1217
-128
1. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
08/2
3/10
. For
per
sona
l use
onl
y.


















