
Myricetin prevents titanium particle-induced...
13
Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro Chuanlong Wu 1 , Wengang Wang 1 , Bo Tian 1 , Xuqiang Liu, Xinhua Qu, Zanjing Zhai, Haowei Li, Fengxiang Liu, Qiming Fan, Tingting Tang, An Qin *, Zhenan Zhu * Shanghai Key Laboratory of Orthopaedic Implants, Department of Orthopaedics, Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, The People’s Republic of China 1. Introduction Total hip arthroplasty is a highly successful procedure that has become a standard of care to improve the quality of life and to reduce pain in patients with joint disease. However, this surgery is associated with a risk of aseptic loosening, leading to prosthetic failure. Although the pathophysiology of aseptic loosening remains unclear, considerable evidence suggests that periprosthetic osteolysis initiated by inflammatory responses to particulate wear debris is responsible [1,2]. Wear debris is produced by biological responses or mechanical effects after joint implant, including metal debris, ultra-high molecular weight polyethylene (UHMWPE) debris, cement debris, and so on. Among the types of wear debris, titanium (Ti) particles are an important factor contributing to osteolysis in metal-on- metal or metal-on-polyethylene hip arthroplasty. Ti particles can induce the recruitment of macrophages, lymphocytes, and osteoclasts to the bone. Osteoclasts are large, multinucleated cells that arise from hematopoietic monocyte/macrophage lineage precursors [3]. Subsequently, these cells release multiple pro- inflammatory cytokines, such as tumor necrosis factor (TNF)-a, interleukin (IL)-1b, IL-6, IL-11, and IL-17, and prostaglandin E2, into the tissues surrounding the failed implants. These factors induce receptor activator of nuclear factor-kB ligand (RANKL) expression via osteoblasts and bone marrow stromal cells. RANKL induces osteoclast differentiation [4] by binding to receptor activator of nuclear factor-kB (RANK), its cognate receptor. This leads to the activation of downstream signaling molecules, such as mitogen-activated protein kinase (MAPK), extracellular signal- regulated kinase (ERK)1/2, p38, c-Jun N-terminal kinase (JNK)1/2, Biochemical Pharmacology xxx (2014) xxx–xxx A R T I C L E I N F O Article history: Received 17 August 2014 Accepted 29 October 2014 Available online xxx Keywords: Osteoclast formation Osteoclastic bone resorption Periprosthetic osteolysis MAPK signaling NF-kB signaling A B S T R A C T Titanium (Ti) particle-induced periprosthetic osteolysis and subsequent aseptic loosening are a primary reason for total hip arthroplasty failure. The aim of this study was to assess the effect of myricetin on Ti particle-induced osteolysis and osteoclastogenesis. We demonstrated that myricetin, a natural plant extract, exerts potent inhibitory effects on Ti particle-induced osteolysis in a mouse calvarial model. Further histological analysis indicated that the inhibition of osteoclast formation and function, and the secretion of inflammatory factors, are key targets for therapeutic agents in the treatment of wear particle-induced osteolysis. In vitro, we found that myricetin suppressed receptor activator of nuclear factor-kB ligand (RANKL)-mediated osteoclast differentiation, bone resorption, and F-actin ring formation in a dose-dependent manner. Moreover, myricetin significantly reduced the expression of osteoclast-specific markers in mouse bone marrow-derived macrophages, including tartrate-resistant acid phosphatase (TRAP), cathepsin K, the calcitonin receptor, V-ATPase d2, c-fos, and nuclear factor of activated T cells (NFAT) c1. Further investigation revealed that myricetin inhibited osteoclastogenesis through the suppression of the nuclear factor-kB (NF-kB) signaling pathway and mitogen-activated protein kinase (MAPK) pathways involving extracellular signal-regulated kinase 1/2 (ERK1/2), p38, and c-Jun N-terminal kinase 1/2 (JNK1/2). While, the inhibition of TNF-a and IL-1b secretion was another reason for the suppressive effect of myricetin on Ti particle-induced osteolysis. Collectively, these findings suggest that myricetin is a potential natural agent for the treatment of periprosthetic osteolysis and other osteoclast-related osteolytic diseases. ß 2014 Elsevier Inc. All rights reserved. * Corresponding authors at: 639 Zhizaoju Road, Shanghai 200011, PR China. Tel.: +86 2162139920; fax: +86 2162139920. E-mail addresses: [email protected] (A. Qin), [email protected] (Z. Zhu). 1 Contributed equally to the work. G Model BCP-12131; No. of Pages 13 Please cite this article in press as: Wu C, et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL- induced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.10.019 Contents lists available at ScienceDirect Biochemical Pharmacology jo u rn al h om epag e: ww w.els evier.c o m/lo cat e/bio c hem p har m http://dx.doi.org/10.1016/j.bcp.2014.10.019 0006-2952/ß 2014 Elsevier Inc. All rights reserved.
Transcript of Myricetin prevents titanium particle-induced...

Biochemical Pharmacology xxx (2014) xxx–xxx
G Model
BCP-12131; No. of Pages 13
Myricetin prevents titanium particle-induced osteolysis in vivo andinhibits RANKL-induced osteoclastogenesis in vitro
Chuanlong Wu 1, Wengang Wang 1, Bo Tian 1, Xuqiang Liu, Xinhua Qu, Zanjing Zhai,Haowei Li, Fengxiang Liu, Qiming Fan, Tingting Tang, An Qin *, Zhenan Zhu *
Shanghai Key Laboratory of Orthopaedic Implants, Department of Orthopaedics, Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine,
Shanghai, The People’s Republic of China
A R T I C L E I N F O
Article history:
Received 17 August 2014
Accepted 29 October 2014
Available online xxx
Keywords:
Osteoclast formation
Osteoclastic bone resorption
Periprosthetic osteolysis
MAPK signaling
NF-kB signaling
A B S T R A C T
Titanium (Ti) particle-induced periprosthetic osteolysis and subsequent aseptic loosening are a primary
reason for total hip arthroplasty failure. The aim of this study was to assess the effect of myricetin on Ti
particle-induced osteolysis and osteoclastogenesis. We demonstrated that myricetin, a natural plant
extract, exerts potent inhibitory effects on Ti particle-induced osteolysis in a mouse calvarial model.
Further histological analysis indicated that the inhibition of osteoclast formation and function, and the
secretion of inflammatory factors, are key targets for therapeutic agents in the treatment of wear
particle-induced osteolysis. In vitro, we found that myricetin suppressed receptor activator of nuclear
factor-kB ligand (RANKL)-mediated osteoclast differentiation, bone resorption, and F-actin ring
formation in a dose-dependent manner. Moreover, myricetin significantly reduced the expression of
osteoclast-specific markers in mouse bone marrow-derived macrophages, including tartrate-resistant
acid phosphatase (TRAP), cathepsin K, the calcitonin receptor, V-ATPase d2, c-fos, and nuclear factor of
activated T cells (NFAT) c1. Further investigation revealed that myricetin inhibited osteoclastogenesis
through the suppression of the nuclear factor-kB (NF-kB) signaling pathway and mitogen-activated
protein kinase (MAPK) pathways involving extracellular signal-regulated kinase 1/2 (ERK1/2), p38, and
c-Jun N-terminal kinase 1/2 (JNK1/2). While, the inhibition of TNF-a and IL-1b secretion was another
reason for the suppressive effect of myricetin on Ti particle-induced osteolysis. Collectively, these
findings suggest that myricetin is a potential natural agent for the treatment of periprosthetic osteolysis
and other osteoclast-related osteolytic diseases.
� 2014 Elsevier Inc. All rights reserved.
Contents lists available at ScienceDirect
Biochemical Pharmacology
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/b io c hem p har m
1. Introduction
Total hip arthroplasty is a highly successful procedure that hasbecome a standard of care to improve the quality of life and toreduce pain in patients with joint disease. However, this surgery isassociated with a risk of aseptic loosening, leading to prostheticfailure. Although the pathophysiology of aseptic loosening remainsunclear, considerable evidence suggests that periprostheticosteolysis initiated by inflammatory responses to particulate weardebris is responsible [1,2].
Wear debris is produced by biological responses or mechanicaleffects after joint implant, including metal debris, ultra-high
* Corresponding authors at: 639 Zhizaoju Road, Shanghai 200011, PR China.
Tel.: +86 2162139920; fax: +86 2162139920.
E-mail addresses: [email protected] (A. Qin), [email protected]
(Z. Zhu).1 Contributed equally to the work.
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
http://dx.doi.org/10.1016/j.bcp.2014.10.019
0006-2952/� 2014 Elsevier Inc. All rights reserved.
molecular weight polyethylene (UHMWPE) debris, cement debris,and so on. Among the types of wear debris, titanium (Ti) particles
are an important factor contributing to osteolysis in metal-on-
metal or metal-on-polyethylene hip arthroplasty. Ti particles can
induce the recruitment of macrophages, lymphocytes, and
osteoclasts to the bone. Osteoclasts are large, multinucleated cells
that arise from hematopoietic monocyte/macrophage lineage
precursors [3]. Subsequently, these cells release multiple pro-
inflammatory cytokines, such as tumor necrosis factor (TNF)-a,
interleukin (IL)-1b, IL-6, IL-11, and IL-17, and prostaglandin E2,
into the tissues surrounding the failed implants. These factors
induce receptor activator of nuclear factor-kB ligand (RANKL)
expression via osteoblasts and bone marrow stromal cells. RANKL
induces osteoclast differentiation [4] by binding to receptor
activator of nuclear factor-kB (RANK), its cognate receptor. This
leads to the activation of downstream signaling molecules, such as
mitogen-activated protein kinase (MAPK), extracellular signal-
regulated kinase (ERK)1/2, p38, c-Jun N-terminal kinase (JNK)1/2,
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx2
G Model
BCP-12131; No. of Pages 13
and nuclear factor-kB (NF-kB) [5,6]. Excessive formation of RANKLcan stimulate enhanced osteoclast formation and bone resorption,thereby exacerbating the inflammatory response, resulting inperiprosthetic osteolysis finally [7–11]. Macrophage colony-stimulating factor (M-CSF) is also required for osteoclast formation[12]. Targeted inhibition of osteoclast signaling pathways mighttherefore suppress osteoclast formation and/or function, and couldprovide a promising strategy for the treatment of osteoclast-related osteolytic diseases.
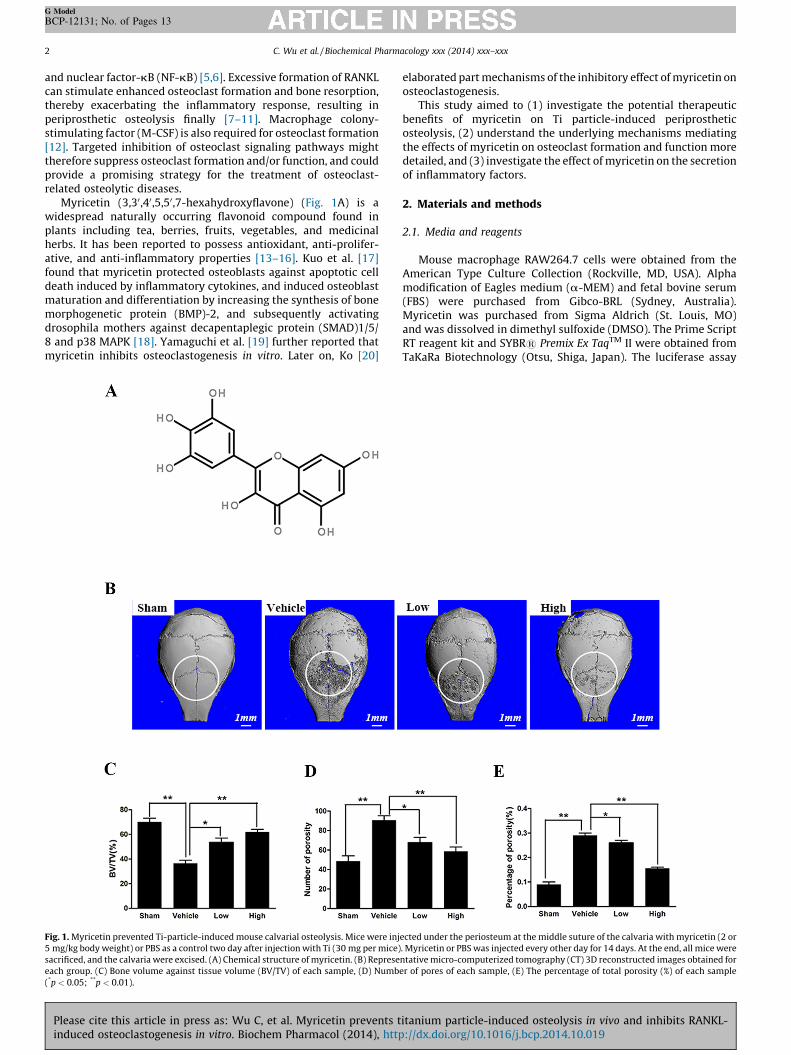
Myricetin (3,30,40,5,50,7-hexahydroxyflavone) (Fig. 1A) is awidespread naturally occurring flavonoid compound found inplants including tea, berries, fruits, vegetables, and medicinalherbs. It has been reported to possess antioxidant, anti-prolifer-ative, and anti-inflammatory properties [13–16]. Kuo et al. [17]found that myricetin protected osteoblasts against apoptotic celldeath induced by inflammatory cytokines, and induced osteoblastmaturation and differentiation by increasing the synthesis of bonemorphogenetic protein (BMP)-2, and subsequently activatingdrosophila mothers against decapentaplegic protein (SMAD)1/5/8 and p38 MAPK [18]. Yamaguchi et al. [19] further reported thatmyricetin inhibits osteoclastogenesis in vitro. Later on, Ko [20]
Fig. 1. Myricetin prevented Ti-particle-induced mouse calvarial osteolysis. Mice were inj
5 mg/kg body weight) or PBS as a control two day after injection with Ti (30 mg per mice)
sacrificed, and the calvaria were excised. (A) Chemical structure of myricetin. (B) Represe
each group. (C) Bone volume against tissue volume (BV/TV) of each sample, (D) Numb
(*p < 0.05; **p < 0.01).
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
elaborated part mechanisms of the inhibitory effect of myricetin onosteoclastogenesis.
This study aimed to (1) investigate the potential therapeuticbenefits of myricetin on Ti particle-induced periprostheticosteolysis, (2) understand the underlying mechanisms mediatingthe effects of myricetin on osteoclast formation and function moredetailed, and (3) investigate the effect of myricetin on the secretionof inflammatory factors.
2. Materials and methods
2.1. Media and reagents
Mouse macrophage RAW264.7 cells were obtained from theAmerican Type Culture Collection (Rockville, MD, USA). Alphamodification of Eagles medium (a-MEM) and fetal bovine serum(FBS) were purchased from Gibco-BRL (Sydney, Australia).Myricetin was purchased from Sigma Aldrich (St. Louis, MO)and was dissolved in dimethyl sulfoxide (DMSO). The Prime ScriptRT reagent kit and SYBR1 Premix Ex TaqTM II were obtained fromTaKaRa Biotechnology (Otsu, Shiga, Japan). The luciferase assay
ected under the periosteum at the middle suture of the calvaria with myricetin (2 or
. Myricetin or PBS was injected every other day for 14 days. At the end, all mice were
ntative micro-computerized tomography (CT) 3D reconstructed images obtained for
er of pores of each sample, (E) The percentage of total porosity (%) of each sample
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 3
G Model
BCP-12131; No. of Pages 13
system was obtained from Promega (Sydney, Australia). Solublehuman recombinant M-CSF and bacteria-derived recombinantmouse RANKL were supplied by R&D (R&D Systems, Minneapolis,MN). Specific antibodies against phospho-p38(#9211),p38(#8690), phospho-JNK1/2(#9251), JNK1/2(#9252), phos-pho-ERK1/2(#9101), ERK1/2(#9102), phospho-IkBa(#9246),IkBa(#9242), MITF (#12590), c-fos(#4384), nuclear factor ofactivated T cells (NFAT) c1(#8032), and b-actin(#4967) werepurchased from Cell Signaling (Danvers, MA). Antibodies againstglyceraldehyde-3-phosphate dehydrogenase (GAPDH) (sc-365062)were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA).The Diagnostic Acid Phosphatase kit for tartrate-resistant acidphosphatase (TRAP) staining, Tris, glycine, NaCl, sodium dodecylsulfate (SDS), and other regents were obtained from Sigma. The CellCounting Kit-8 (CCK8) was obtained from Dojindo MolecularTechnology (Japan).
2.2. Ti particle-induced calvarial osteolysis model
As described previously, we established a mouse calvarialosteolysis model and used this to measure the osteolysis-suppressing effect of myricetin in vivo [21]. All experimentalprocedures were performed in accordance with the guidelines ofthe Animal Care Committee of Shanghai Jiao Tong University.Commercial pure titanium (Ti) particles were purchased fromJohnson Matthey (Catalog #00681; Ward Hill, MA, USA). Toremove adherent endotoxins from the Ti particles, Ti particles weresterilized by baking at 180 8C for 6 h, followed by treatment with70% (v/v) ethanol for 48 h [22]. Briefly, 24 healthy 8-week-oldC57BL/J6 mice were assigned randomly to 4 groups: PBS control(sham), Ti particles in PBS (vehicle), and Ti particles together withlow (2 mg/kg/day) and high (5 mg/kg/day) doses of myricetin (Lowand High myricetin, respectively). First, 30 mg of Ti particles wereembedded under the periosteum at the middle suture of thecalvaria [23]. Two days after implantation, myricetin or PBS wasinjected into the periosteum, receiving doses every other day for14 d. No adverse effects or mortality occurred during theexperiment. At the end of the experiment, the mice were sacrificed,and the calvaria were excised and fixed in 4% (w/v) paraformalde-hyde for micro-computerized tomography (CT) analysis.
2.3. Micro-CT scanning
The fixed calvaria were analyzed using a high-resolution micro-CT (Skyscan 1072; Skyscan, Aartselaar, Belgium). The scanningprotocol was set at an isometric resolution of 9 mm, and the X-rayenergy settings were 80 kV and 80 mA. After reconstruction, asquare region of interest (ROI) around the midline suture wasselected for further qualitative and quantitative analyses. Bonevolume/tissue volume (BV/TV), the number of pores, and thepercentage of total porosity of each sample were measured asreported previously [24].
2.4. Histological and histomorphometric analysis
After micro-CT scanning, the calvaria samples were decalcifiedin 10% (w/v) ethylenediaminetetraacetic acid (EDTA; pH 7.4) for3 weeks and then embedded in paraffin. Histological sections wereprepared and used for hematoxylin and eosin (H&E) staining andTRAP staining, after which the specimens were examined andphotographed under a high-quality microscope. When analyzingthe middle-suture osteolysis area, the non-osseous tissue adjacentto and continuous with the midline suture was considered as theosteolysis area and this was analyzed using Image Pro-Plus 5.0(Media Cybernetics; USA). The number of TRAP-positive multinu-cleated osteoclasts was determined for each sample.
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
2.5. Bone marrow-derived macrophage isolation and osteoclast
culture
Primary mouse cell culture of bone marrow-derived macro-phages (BMM) were isolated from the long bones of 5-week-oldC57BL/6 mice, as described previously [25]. Briefly, cells wereisolated from the femur and tibiae bone marrow and cultured in aT75 flask ina-MEM supplemented with 10% (v/v) FBS, 1% (w/v)penicillin/streptomycin, and 10 ng/mL M-CSF for 24 h. Non-adherent cells were then removed, and the adherent cells werecultured in an incubator maintained at 37 8C with a 5% CO2 (v/v)atmosphere for a further 3–4 days until cells were fully confluent.Then BMMs were cultured in complete a-MEM medium (a-MEMcontaining 10% (v/v) heat inactivated FBS, 2 mM L-glutamine,100 U/mL penicillin/streptomycin, and 30 ng/mL M-CSF)RAW264.7 cells were cultured in a-MEM containing 10% (v/v)FBS, 2 mM L-glutamine, and 100 U/mL penicillin/streptomycin.These cell cultures were maintained at 37 8C in a humidenvironment with 5% (v/v) CO2.
2.6. Cell viability and cytotoxicty assay
The effect of myricetin on BMM cell viability was determined bya CCK8 assay. The BMM cells were plated in 96-well plates at adensity of 1 � 104 cells/well, and cultured in complete a-MEMmedium containing different concentrations of myricetin (0, 12.5, 25,50, 100, 200, 400, and 800 mM) for 48 h. Next, 10 mL CCK-8 was addedto each well, and the plates were then incubated at 37 8C for anadditional 2 h. The optical density (OD) was then measured with anELX800 absorbance microplate reader (Bio-Tek, Winooski, USA) at awavelength of 450 nm (650 nm reference). The cell viability wascalculated relative to the control by using the following formula:[experimental group OD � zeroing OD]/[control group OD � zeroingOD]. The half-maximal inhibitory concentration (IC50) value wascalculated using GraphPad Prism version 5.0c (San Diego, CA, USA).
2.7. TRAP staining
BMM cells were seeded into a 96-well plate at a density of8 � 103 cells/well with complete a-MEM medium, RANKL (50 ng/mL), and myricetin (0, 10, 25, or 50 mM). The culture medium wasreplaced every 2 days until osteoclasts formed on day 7. The cellswere fixed with 4% (w/v) paraformaldehyde for 20 min and stainedfor TRAP using the Diagnostic Acid Phosphatase kit. TRAP-positivecells with more than 3 nuclei were counted as osteoclast-like (OCL)cells. Three wells were randomly selected for each group for furtheranalysis and the average number of OCL cells was calculated.
2.8. Resorption pit assay
BMM cells (2.4 � 104 cells/cm2) were seeded onto bovine boneslices in complete a-MEM medium. The BMM cells were stimulatedwith RANKL (50 ng/mL), M-CSF (30 ng/mL), and myricetin (0, 10, 25,and 50 mM) until OCL cells were observed on day 7. The OCL cellswere then removed from the bone slices by mechanical agitation andsonication. Resorption pits were visualized using a scanning electronmicroscope (SEM, FEI Quanta 250). Three view fields were randomlyselected for each bone slice for further analysis. Similar independentexperiments were repeated at least three times. The percentage ofresorbed bone surface area was quantified using the Image Jsoftware (National Institutes of Health).
2.9. F-actin ring immunofluorescence
To perform immunofluorescence staining of F-actin rings,myricetin-treated osteoclasts cultured on bovine bone discs were
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx4
G Model
BCP-12131; No. of Pages 13
fixed with 4% (w/v) paraformaldehyde for 15 min and thenpermeabilized for 5 min with 0.1% (v/v) Triton X-100. Cells wereincubated with rhodamine-conjugated phalloidin (1:100; Invitro-gen Life Technologies, Grand Island, NY, USA) diluted in 0.2% (w/v)bovine serum albumin (BSA) in phosphate-buffered saline (PBS) for1 h at room temperature (RT) and then washed extensively with0.2% (w/v) BSA-PBS and PBS. Cells were then incubated withHoechst 3342 dye (1:5000; Invitrogen Life Technologies) to stainnuclei, washed with PBS, and mounted with ProLong Gold anti-fade mounting medium (Invitrogen Life Technologies). Fluorescentstaining was examined using a NIKON A1Si spectral detectorconfocal system equipped with a 20� (dry) lenses. Fluorescenceimages were collected using NIS-C Elements software andanalyzed using Image J software. Three wells were randomlyselected for each group for further analysis and the averagenumber of F-actin ring cells was calculated. Similar independentexperiments were repeated at least three times.
2.10. RNA extraction and quantitative PCR assay
Quantitative PCR was used to measure the expression of specificgenes during osteoclast formation. BMMs were seeded in 6-wellplates at a density of 1 � 105 cells/well and cultured in complete a-MEM supplemented with 30 ng/mL M-CSF and 50 ng/mL RANKL.During RANKL-induced osteoclastogenesis, the BMMs weretreated with either various doses of myricetin (0, 10, or 50 mM)for 5 days or 50 mM myricetin for 0–5 days. Total RNA wasextracted using the Qiagen RNeasy Mini kit (Qiagen, Valencia, CA)in accordance with the manufacturer’s instructions, and cDNA wassynthesized from 1 mg of total RNA using reverse transcriptase(TaKaRa). Real-time PCR was performed using SYBR1 Premix Ex
TaqTM II (TaKaRa) and an ABI 7500 Sequencing Detection System(Applied Biosystems, Foster City, CA). The following cyclingconditions were used: 40 cycles of denaturation at 95 8C for 5 sand amplification at 60 8C for 24 s. GAPDH was amplified as ahousekeeping gene, and all reactions were run in triplicate. Themouse primer sequences for TRAP, cathepsin K (CTSK), calcitoninreceptor (CTR), V-ATPase-d2, c-fos, NFATc1, and GAPDH weredescribed in Table 1.
2.11. NF-kB luciferase reporter-gene assay
To examine whether myricetin affected NF-kB gene expression,RAW264.7 cells were stably transfected with a p-NF-kB-TA-Lucluciferase reporter construct [26]. Briefly, cells were plated in 24-well plates at a density of 1 � 105 cells/well. After 24 h, the cellswere treated with various concentrations of myricetin (0, 10, 25,and 50 mM) for 1 h, and then incubated with 50 ng/mL RANKL for8 h. Cells were then lysed and the luciferase activity was measured
Table 1Primers used for quantitative PCR.
Gene Forward (F) and reverse (R) primer sequence (50–30)
TRAP F: CTGGAGTGCACGATGCCAGCGACA
R: TCCGTGCTCGGCGATGGACCAGA
CTSK F: CTTCCAATACGTGCAGCAGA
R: TCTTCAGGGCTTTCTCGTTC
CTR F: TGCAGACAACTCTTGGTTGG
R: TCGGTTTCTTCTCCTCTGGA
V-ATPase-d2 F: AAGCCTTTGTTTGACGCTGT
R: TTCGATGCCTCTGTGAGATG
c-fos F: CCAGTCAAGAGCATCAGCAA
R: AAGTAGTGCAGCCCGGAGTA
NFATc1 F: CCGTTGCTTCCAGAAAATAACA
R: TGTGGGATGTGAACTCGGAA
GAPDH F: ACCCAGAAGACTGTGGATGG
R: CACATTGGGGGTAGGAACAC
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
using the Promega Luciferase Assay System, according to themanufacturer’s instructions.
2.12. Western blot analysis
RAW264.7 cells and BMMs were seeded in 6-well plates at adensity of 5 � 105 cells/well. After treatment with vehicle ormyricetin for 2 h, the RAW264.7 cells were stimulated with RANKL(50 ng/mL) for 0, 5, 10, 20, 30, or 60 min. BMMs were cultured with0 or 50 mM myricetin, in the presence of M-CSF (30 ng/mL) andRANKL (50 ng/mL) for 0, 1, 2, 3, 4, and 5 days. The cells were thenwashed twice with PBS and lysed in ice-cold lysis buffer (50 mMTris pH 7.5, 150 mM NaCl, 1% (w/v) Nonidet P-40, 0.1% (w/v) SDS,1% (w/v) sodium deoxycholate) supplemented with PMSF (ShenNeng Bo Cai Corp., China). The lysates were incubated on ice for30 min and then centrifuged at 12,000 rpm for 10 min toprecipitate the debris. The protein concentrations were deter-mined using the BCA protein assay kit (Thermo Scientific, Rockford,IL). Equal amounts of the protein lysates were separated using 10%(w/v) SDS-polyacrylamide gel electrophoresis (PAGE) and electro-blotted onto PVDF membranes (Roche). The membranes were thenblocked with 5% (w/v) skim milk solution for 1 h prior toincubation with primary antibodies at the following dilutions(GAPDH, 1:1000; phospho-p38, 1:1000; p38, 1:1000; phospho-JNK1/2, 1:1000; JNK1/2, 1:1000; phospho-ERK1/2, 1:1000; ERK1/2,1:1000; IkBa, 1:1000; phospho-IkBa, 1:1000; Mitf, 1:1000; c-fos,1:1000; NFATc1, 1:1000; b-actin, 1:1000) at room temperature for4 h. The membranes were then incubated with horseradishperoxidase-conjugated secondary antibodies (1:5000) for 1 h.The antibody reactivity was subsequently visualized using anOdyssey infrared imaging system (LI-COR).
2.13. Measurement of pro-inflammatory cytokines by enzyme-linked
immunosorbent assay (ELISA)
Four groups were assigned for the ELISA: PBS control, Tiparticles in PBS, 50 mM myricetin, and Ti particles together with50 mM myricetin. As described in our previous study[22], the Tisuspensions were sonicated for 30 min using a SB3200 UltrasonicGenerator (Shanghai Branson Ultrasonics Co. Ltd., Shanghai, China)prior to incubation with cells and the concentration of particlesused for incubation was 0.1 mg/mL. BMM were cultured for 6, 24,and 96 h in complete a-MEM medium. Cell culture supernatantswere collected, centrifuged at 1200 � g for 10 min, and exposed toa mixture of protease inhibitors (Kangchen, Shanghai, China).Mouse-specific ELISA kits were used to measure TNF-a and IL-1blevels in accordance with the manufacturer’s instructions andabsorbance was measured at 450 nm. The sensitivities of theassays were: 3 pg/mL for TNF-a, and 7 pg/mL for IL-1b.
2.14. Statistical analyses
Values are presented as the mean � standard deviation (SD). Atleast three independent replicated of each experiment wereconducted separately Statistically significant differences betweenthe groups were determined by Student’s t-test using the SPSS17.0 software (SPSS Inc., USA). Differences were considered signifi-cant at *P < 0.05 or **P < 0.01.
3. Results
3.1. Administration of myricetin prevented wear-particle-induced
bone loss in vivo
To explore the effects of myricetin on pathological osteolysis,we used a mouse calvaria model of Ti particle-induced osteolysis.
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 5
G Model
BCP-12131; No. of Pages 13
Micro-CT scanning and 3D reconstruction revealed extensive in
vivo bone resorption in the Ti group, which was observed asextensive surface erosions on the calvaria (Vehicle; Ti injec-tion), when compared with the negative control (Sham; PBSinjection) (Fig. 1B). However, treatment with myricetinsuppressed Ti-particle-induced osteolysis in a dose-dependentmanner, with bone resorption in mice treated with a highconcentration of myricetin being considerably lower than thatin mice treated with a low concentration of the compound(Fig. 1B). Quantification of bone parameters confirmed thatmyricetin-Low (2 mg/kg/day myricetin) and myricetin-High(5 mg/kg/day myricetin) significantly increased the BV/TV(Fig. 1C) and decreased the number of pores and percentageof porosity (%) (Fig. 1D and E).
Histological assessment and histomorphometric analysisfurther confirmed that myricetin treatment protected againstTi particle-induced bone loss. H&E staining showed fewosteolytic changes in sections from the sham group. Osteolysishad clearly occurred in the vehicle group, whereas themyricetin-treated groups exhibited reduced osteolysis(Fig. 2A). Furthermore, consistent with the micro-CT quantita-tion, histomorphometric analysis demonstrated that bothlow and high doses of myricetin reduced the erosion surface.TRAP staining revealed that the number of multinucleatedosteoclasts at the injection site was increased in the presence ofTi particles, as indicated by the presence of osteoclasts lining theeroded bone surface. However, in both the low- and high-dosemyricetin treatment groups, the number of osteoclasts and thepercentage of osteoclast surface relative to the bone surfacewere decreased (Fig. 2B).
Fig. 2. Myricetin prevented Ti-induced bone loss by inhibiting osteoclast activity. (A) Cal
(5� and 20�) and TRAP staining (20�) were performed using atleast 3 sections per gr
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
3.2. Myricetin inhibited RANKL-induced osteoclast formation without
producing cytotoxicity
To examine the potential cytotoxicity of myricetin, CCK8 assayswere performed. The IC50 value of myricetin was 601.5 mM at 48 h.This indicated that myricetin may partially suppress the prolifera-tion of BMM cells, but only at high concentrations (>601.5 mM).
To investigate the effect of myricetin on osteoclastogenesis,BMMs were stimulated with RANKL (50 ng/mL), M-CSF (30 ng/mL), and myricetin (0, 10, 25, and 50 mM) during osteoclastformation. BMM cells differentiated into characteristic TRAP-positive multinucleated OCL cells (Fig. 3C). However, in cellsexposed to myricetin, the number of TRAP positive OCL cells wassignificantly decreased in a dose dependent manner. Osteoclastformation was suppressed by approximately 30% by 10 mMmyricetin (Fig. 3D), and was almost completely inhibited at higherconcentrations (�50 mM) (Fig. 3E). Myricetin did not affect theproliferation of BMM cells at either 25 mM or 50 mM (Fig. 3A andB). These results suggested that these concentrations of myricetincould impair osteoclast formation without causing cell death.
3.3. Myricetin attenuated osteoclastic bone resorption
Because osteoclast differentiation was obviously impaired bymyricetin, we hypothesized that osteoclast bone resorption wouldalso be inhibited. To evaluate this hypothesis, we performed anosteoclastic bone resorption assay. SEM analysis indicated thatosteoclasts actively resorbed the bone surface in vitro. More than50% of this bone resorption activity was effectively inhibited by10 mM myricetin, and almost completely blocked at higher
varias were fixed, decalcified, dehydrated, embedded, and sectioned. H & E staining
oup. (B) The numbers of osteoclasts were determined. (*p < 0.05; **p < 0.01).
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

Fig. 3. Myricetin inhibited RANKL-induced osteoclastogenesis without producing any cytotoxic effects. (A) BMMs were treated with 30 ng/mL M-CSF, 50 ng/mL RANKL, and
the indicated concentrations of myricetin for 48 h prior to measuring cell viability by CCK-8 assay (*p < 0.05; **p < 0.01). (B) The inhibition rate of BMMs was calculated
(GraphPad Prism) and the half-maximal inhibitory concentration (IC50) was determined to be 601.5 mM. (C) BMMs were treated with the indicated concentrations of
myricetin and then with 30 ng/mL M-CSF and 50 ng/mL RANKL for 5 days. Cells were fixed with 4% (w/v) paraformaldehyde and stained for TRAP. (D) The number of
TRAP-positive cells. (E) BMMs were treated with or without 50 mM myricetin and then with 30 ng/mL M-CSF and 50 ng/mL RANKL; cells were then fixed and stained for
TRAP on days 1, 3, and 5. (F) The number of osteoclasts.
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx6
G Model
BCP-12131; No. of Pages 13
Please cite this article in press as: Wu C, et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 7
G Model
BCP-12131; No. of Pages 13
myricetin concentrations (�50 mM) (Fig. 4A and B). This resultclearly demonstrated that the administration of myricetin reducedbone resorption in vitro.
3.4. Myricetin inhibited F-actin ring formation in mature osteoclasts
in vitro
The effects of myricetin on osteoclastogenesis were furtherstudied by examining RANKL-induced osteoclast F-actin ringformation, a readily observable characteristic of mature osteoclastsand a prerequisite for osteoclast bone resorption during osteo-clastogenesis. Confocal microscopy revealed a characteristicF-actin ring formation and morphology in control osteoclastsstained with phalloidin-Alexa Fluor 647 (Fig. 5A and B). However,treatment with myricetin drastically disrupted F-actin ringformation and morphology, with dose-dependent reductionsobserved in both the size and the number of F-actin rings aftermyricetin treatment (Fig. 5A and B). Collectively, these findingssuggested that myricetin treatment impaired osteoclast F-actinring formation in vitro.
3.5. Myricetin suppressed RANKL-induced gene expression
The expression of several genes is upregulated during osteoclastdifferentiation. Therefore, we used quantitative PCR to analyze theinhibitory effect of myricetin on RANKL-induced mRNA expressionof osteoclast-related genes. Quantitative real-time PCR demonstrat-ed that RANKL potently induced the expression of all the evaluatedgenes (TRAP, CTR, CTSK, V-ATPase-d2, c-fos, and NFATc1). However,the RANKL-induced upregulation of these genes was stronglysuppressed in the presence of myricetin in a time- and dose-dependent manner (Fig. 6A and B). Collectively, these results furtherconfirmed that myricetin suppressed osteoclast formation and theexpression of osteoclast-specific genes in vitro.
3.6. Myricetin suppressed RANKL-induced activation of MAPK and
NF-kB signaling pathways
In order to elucidate the mechanism underlying myricetininduced inhibiton of osteoclast formation and function, RANKL-induced signaling pathways were investigated by western blottingand luciferase assays. RAW264.7 cells were stimulated with RANKL(50 ng/mL) for 0, 5, 10, 20, 30, or 60 min to investigate short termsignaling pathways (p-p38, p38, p-ERK1/2, ERK1/2, JNK, p-JNK, p-IkBa and IkBa). The results showed that the phosphorylation ofp38, JNK1/2, and ERK1/2 peaked within 10 min, 20 min, and
Fig. 4. Myricetin attenuated osteoclast-mediated bone resorption. Equal numbers of oste
with 30 ng/mL M-CSF, 50 ng/mL RANKL, and the indicated concentration of myricetin. (A)
are shown. (B) The total area of resorption pits, measured using Image J. Similar result
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
30 min of RANKL stimulation, respectively. However, pretreatmentwith myricetin significantly decreased phosphorylation withinthese signaling pathways (Fig. 7A–D). Collectively, these datasuggested that myricetin inhibited MAPK (p38, ERK1/2, and JNK1/2) signaling pathways during osteoclastogenesis.
As shown in Fig. 7E, NF-kB was activated by RANKL and thisactivation was partially reduced by myricetin in a dose dependentmanner. These results suggested that myricetin might participatein the regulation of RANKL-activated NF-kB signaling, and couldthus contribute to the inhibition of osteoclast formation (Fig. 7Eand F). This finding was further supported by western blot analysisof IkBa phosphorylation and degradation. In the control cells, IkBaphosphorylation and degradation were initiated by RANKLstimulation. When the cells were pretreated with myricetin(50 mM), RANKL-induced IkBa phosphorylation and degradationwere both significantly suppressed (Fig. 7E). These data firmlysuggested that myricetin suppressed the RANKL-induced NF-kBsignaling pathway by inhibiting IkBa phosphorylation anddegradation. The luciferase reporter assay further confirmed thatmyricetin inhibited NF-kB activation in a dose-dependent manner(Fig. 7G).
BMMs were cultured with 0 or 50 mM myricetin, in thepresence of M-CSF (30 ng/mL) and RANKL (50 ng/mL) for 0, 1, 2, 3,4, and 5 days to identify the long-term signaling ways (Mitf, c-fosand NFATc1). The results showed that Mitf, c-fos and NFATc1protein levels were upregulated in response to RANKL on days1 and 3, and that this effect was strongly inhibited by myricetin in atime- and dose-dependent manner (Fig. 7H).
3.7. Myricetin and pro-inflammatory cytokines
To determine whether myricetin can inhibit the production ofpro-inflammatory cytokines that are produced by Ti particles[22],we monitored the concentration of TNF-a, IL-1b in BMM culturesusing ELISA assays. ELISAs conducted in medium collected fromBMM cultures 6 h after seeding showed that the TNF-a and IL-1bconcentration was higher in the presence than in the absence of Tiparticles. The concentration of both cytokines increased further asthe culture progressed (Fig. 8). Exposure of BMM to 50 mMMyricetin in the presence of Ti particles suppressed secretion ofTNF-a and IL-1b. Although myricetin inhibited the release ofcytokines by BMM, the concentration of TNF-a and IL-1b inmedium from BMM was higher in the presence than in the absenceof Ti particles (Fig. 8). This result suggests that the inhibition ofproinflammatory cytokine secretion by myricetin is less pro-nounced than the inhibition of bone resorption.
oclasts were seeded onto bone slices, allowed to adhere to the surface, and treated
Representative scanning electron microscopy (SEM) images of bone-resorption pits
s were obtained from at least 3 independent experiments (*p < 0.05; **p < 0.01).
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

Fig. 5. F-actin ring formation in vitro. BMMs were incubated with 30 ng/mL M-CSF, 50 ng/mL RANKL and then treated with the indicated concentrations of myricetin. Cells
were fixed and stained for F-actin. (A) Fluorescence images are shown. (B) The number and size of F-actin-positive cells. All experiments were carried out at least three times,
and significance was determined by Student–Newman–Keul’s tests (*p < 0.05; **p < 0.01).
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx8
G Model
BCP-12131; No. of Pages 13
4. Discussion
Wear particle-induced periprosthetic osteolysis and subse-quent aseptic loosening are a primary reason for total hiparthroplasty failure. To our knowledge, wear particles increasethe secretion of pro-inflammatory cytokines, thereby inducingosteolysis through the dysregulation of osteoclast and osteoblastactivity. Such osteolytic diseases include aseptic loosening ofprosthesis, osteoporosis, periodontal disease, rheumatoid arthritis,and so on[27,28]. During the last two decades, many advances havebeen made in the treatment of osteolytic diseases, with the use ofbisphosphonates, estrogens, denosumab, and teriparatide to treatthese diseases. However, due to numerous and severe side effects,such as gastrointestinal toxicity, osteonecrosis of the jaw, femurfractures, atrial fibrillation, breast cancer, endometrial cancer,thromboembolism, cellulitis et al., and high cost[29–32], thecurrently available treatments remain far from ideal. In this study,
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
we demonstrated that myricetin was capable of inhibiting Tiparticle-induced osteolysis in vivo. This effect is mediated by thesuppression of the RANKL-induced MAPK (p38, JNK1/2, and ERK1/2) and NF-kB signaling pathways during osteoclast formation andbone resorption, and through the inhibition of TNF-a and IL-1bsecretion. Thus, our findings demonstrated the potential applica-tion of myricetin to treat wear particle-induced periprostheticosteolysis.
We first developed an in vivo Ti particle-induced osteolysismodel. It is well known that adherent endotoxins on Ti particlesare responsible for osteoclast formation, as well as the productionof IL-1b, IL-6, and TNF-a [33]. In view of this, we used a series ofprocedures to remove adherent endotoxins, as previously reported[22]. The present study identified obvious osteolysis in Ti-exposedanimals, whereas the myricetin-treated groups showed significantand dose-dependent suppression of Ti particle-induced osteolysis,without any side-effects. These results were further supported by
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

Fig. 6. Myricetin inhibited RANKL-induced osteoclast-specific gene expression. (A) BMMs were cultured with M-CSF (30 ng/mL), RANKL (50 ng/mL), and the indicated
concentrations of myricetin for 5 days, or (B) with M-CSF (30 ng/mL), RANKL (50 ng/mL), and 50 mM myricetin for 0, 1, 3, and 5 days. Expression of the osteoclast-specific
genes TRAP, CTSK, CTR, V-ATPase-d2, c-fos, and NFATc1 were analyzed using real-time PCR. RNA expression levels were normalized relative to the expression of GAPDH
(*p < 0.05; **p < 0.01).
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 9
G Model
BCP-12131; No. of Pages 13
histological analysis of tissues stained with H&E and for TRAP,because myricetin reduced inflammation, osteolysis, and thenumber of TRAP-positive multinucleated osteoclasts. Therefore,the inhibition of osteoclast formation and function, and thesecretion of inflammatory factors are considered as a key target fortherapeutic agents to treat wear particle-induced osteolysis.
Bone resorption assays revealed that the number and area ofbone resorption pits observed in vitro were dramatically decreasedafter myricetin exposure. These effects were caused by theinhibition of osteoclast formation in a dose-dependent manner.This inhibitory effect was observed at concentrations of 10–50 mM
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
myricetin, and occurred in the absence of cytotoxic effects. A well-polarized F-actin ring is the most discernible characteristic ofmature osteoclasts and it is a prerequisite for osteoclast-mediatedbone resorption [34]. Importantly, myricetin suppressed F-actinring formation, further supporting the notion that myricetininhibits bone resorption.
RANKL-stimulated differentiation of osteoclasts is associatedwith the upregulation of specific genes [5]. We found that theinhibitory effect of myricetin on osteoclastogenesis involvedinhibition of RANKL-induced mRNA-expression of TRAP, CTSK,CTR, V-ATPase-d2, c-fos, and NFATc1.
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

Fig. 7. Myricetin inhibited osteoclast differentiation by specifically impairing RANKL-induced MAPK (p38, JNK, ERK) cascades and the NF-kB pathway. (A, E) RAW264.7 cells
were treated with or without 50 mM myricetin for 4 h and then treated with 50 ng/mL RANKL for the indicated periods. Cell lysates were analyzed using western blotting with
specific antibodies against phospho-p38, p38, phospho-ERK1/2, ERK1/2, phospho-JNK1/2, JNK1/2, b-actin, phospho-IkBa, IkBa, and GAPDH. (B, C, D) The gray levels
corresponding to phosphorylation of the indicated proteins were quantified and normalized relative to b-actin using Image J for p-p38, p-JNK, and p-ERK, (F) The gray levels
corresponding to phosphorylation of the indicated proteins were quantified and normalized relative to GAPDH using Image J for p-IkBa. (G) RAW264.7 cells that had been
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx10
G Model
BCP-12131; No. of Pages 13
Please cite this article in press as: Wu C, et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.10.019

Fig. 8. Production of cytokines in BMM cultures with different treatments. Four
group were assigned: control, Ti particles (0.1 mg/mL), myricetin (50 mM) and Ti
particles (0.1 mg/mL) + myricetin (50 mM). (A) ELISA conducted to evaluate TNF-a
(B) ELISA conducted to evaluate IL-1b. Data were expressed as mean � S.E.
(*p < 0.05; **p < 0.01).
C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 11
G Model
BCP-12131; No. of Pages 13
Previous studies showed that the three major subfamilies of theMAPK signaling pathways (p38, JNK 1/2, ERK1/2) play pivotal rolesin osteoclast development downstream of RANK signaling[35]. The activation of the NF-kB pathway is also crucial forosteoclast formation [36,37]. NF-kB is a nuclear transcriptionfactor that is ubiquitously expressed in all cell types. In restingcells, NF-kB proteins are in the cytoplasm and are associated with anumber of inhibitory IkB proteins including IkBa (the mostabundant form), IkBb, and IkBe [38]. Previous studies found thatNF-kB p50�/� and p52�/� knockout mice showed seriousosteopetrosis because osteclasts failed to form [39,40]. At theprotein level, we revealed that myricetin inhibits osteoclastogen-esis through the suppression of the RANKL-induced MAPK (p38,JNK1/2, ERK1/2) and NF-kB signaling pathways.
NFATc1 is a well-characterized master transcription factor thatis a key target gene of NF-kB during RANKL-induced osteoclas-togenesis. Immediately after NF-kB translocation, NF-kB proteinsare recruited to the NFATc1 promoter, leading to the induction ofNFATc1 auto-amplification during osteoclast formation [41]. Thisidea was supported by Yamashita et al. [42], who found thatNFATc1 induction was impaired in p50/p52-deficient cells. The
stably transfected with an NF-kB luciferase reporter construct were treated with the ind
presence of RANKL for 8 h. Luciferase activity was measured using the Promega Lucifera
cultured with 30 ng/mL M-CSF and 50 ng/mL RANKL with or without 50 mM myricetin fo
The gray levels corresponding to the indicated proteins were quantified and normalized
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
activity of Mitf and c-fos, transcription factors critical for osteoclastdifferentiation, is regulated by MAPK signaling pathways [43–47]. Our findings also demonstrated that inhibition of NF-kBactivation and MAPK phosphorylation by myricetin impaired theinduction of the Mitf/c-fos/NFATc1 cascade, resulting in impairedosteoclast formation.
Ti particles can stimulate macrophages to produce inflamma-tory cytokines such as TNF-a, IL-1a, IL-1b, IL-6, and PGE2 [28,48–52]. The localized release of these molecules can elicit osteoclasticbone resorption [53]. We reported that myricetin inhibits TNF-aand IL-1b release by BMMs, albeit to a lesser extent than theinhibition of bone resorption, suggesting that myricetin preventsTi particle-induced osteolysis partly by inhibiting TNF-a and IL-1bsecretion.
Our study has several limitations. First, although Ko [20]previously reported a partial mechanism of myricetin actionduring osteoclastogenesis, our study is much more thorough andcomprehensive. Ti particle-induced osteolysis in vivo is a complexprocess determined by interactions between bones, cells andinflammatory signals. The possibility that pro-inflammatorycytokines stimulate RANKL production by osteoblasts and bonemarrow stromal cells, thereby eliciting macrophage differentiationinto osteoclasts [9,54,55], was not explored. However, we didinvestigate the inhibition of pro-inflammatory cytokines secretionin vitro. Secondly, UHMWPE is more important to today’s jointreplacement surgeons because significantly more polyethyleneparticles (70–90%) appear in tissues around metal-on-polyethyl-ene prostheses [56,57]. However, though metal particles arerelatively fewer in the tissue, they are also an important factorcontributing to osteolysis [58–60]. Both Ti particle and UHMWPEparticle could ultimately activate osteoclast formation andfunction to induce osteolysis in vivo. Therefore, both animalmodels could be suitable for investigating the effect of myricetinon osteolysis [22,58].
There are many strength to this study. We provide the firstcharacterization of the inhibitory effects of myricetin in Ti particle-induced osteolysis. Compared with synthetic drugs, Chinese herbshave fewer side effects and are less prone to causing drugresistance. Therefore, to investigating the effects of myricetin is apromising approach to develop noval treatments for asepticloosening.
Collectively, our study demonstrated that myricetin hadinhibitory effects on Ti particle-induced osteolysis in vivo. Wealso clarified that myricetin functioned through the suppression ofthe MAPK (p38, JNK1/2, ERK1/2) and NF-kB signaling pathwaysand inhibition of TNF-a and IL-1b secretion. Taken together, ourdata strongly suggest that myricetin may represent a potentialagent for the treatment of periprosthetic osteolysis or otherosteoclast-related osteolytic diseases.
Conflicts of interest
None of the authors has any conflicts of interest to declare.
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (No. 81171729), and the Fund for KeyDisciplines of Shanghai Municipal Education Commission (J50206).
icated concentrations of myricetin for 1 h, followed by incubation in the absence or
se Assay System. All experiments were performed at least 3 times (H) BMMs were
r 0–5 days. Cell lysates were prepared and analyzed using western blotting. (I, J, K)
relative to b-actin using Image J for Mitf, c-fos and NFATc1 (*p < 0.05; **p < 0.01).
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx12
G Model
BCP-12131; No. of Pages 13
References
[1] Anderson JM, Rodriguez A, Chang DT. Foreign body reaction to biomaterials.Semin Immunol 2008;20:86–100.
[2] Long M, Rack HJ. Titanium alloys in total joint replacement–a materials scienceperspective. Biomaterials 1998;19:1621–39.
[3] Udagawa N, Takahashi N, Akatsu T, Tanaka H, Sasaki T, Nishihara T, et al. Originof osteoclasts: mature monocytes and macrophages are capable of differenti-ating into osteoclasts under a suitable microenvironment prepared by bonemarrow-derived stromal cells. Proc Natl Acad Sci USA 1990;87:7260–4.
[4] Takahashi N, Udagawa N, Tanaka S, Suda T. Generating murine osteoclastsfrom bone marrow. Methods Mol Med 2003;80:129–44.
[5] Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation.Nature 2003;423:337–42.
[6] Feng X. RANKing intracellular signaling in osteoclasts. IUBMB Life 2005;57:389–95.
[7] Clohisy JC, Frazier E, Hirayama T, Abu-Amer Y. RANKL is an essential cytokinemediator of polymethylmethacrylate particle-induced osteoclastogenesis. JOrthop Res Off Publ Orthop Res Soc 2003;21:202–12.
[8] Gowen M, Wood DD, Ihrie EJ, McGuire MK, Russell RG. An interleukin 1 likefactor stimulates bone resorption in vitro. Nature 1983;306:378–80.
[9] Pioletti DP, Kottelat A. The influence of wear particles in the expression ofosteoclastogenesis factors by osteoblasts. Biomaterials 2004;25:5803–8.
[10] Holding CA, Findlay DM, Stamenkov R, Neale SD, Lucas H, Dharmapatni AS,et al. The correlation of RANK, RANKL and TNFalpha expression with bone lossvolume and polyethylene wear debris around hip implants. Biomaterials2006;27:5212–9.
[11] Jiranek WA, Machado M, Jasty M, Jevsevar D, Wolfe HJ, Goldring SR, et al.Production of cytokines around loosened cemented acetabular components.Analysis with immunohistochemical techniques and in situ hybridization. JBone Joint Surg Am Vol 1993;75:863–79.
[12] Novack DV. Role of NF-kappaB in the skeleton. Cell Res 2011;21:169–82.[13] Hertog MG, Feskens EJ, Hollman PC, Katan MB, Kromhout D. Dietary antioxi-
dant flavonoids and risk of coronary heart disease: the Zutphen Elderly Study.Lancet 1993;342:1007–11.
[14] Yanez J, Vicente V, Alcaraz M, Castillo J, Benavente-Garcia O, Canteras M, et al.Cytotoxicity and antiproliferative activities of several phenolic compoundsagainst three melanocytes cell lines: relationship between structure andactivity. Nutr Cancer 2004;49:191–9.
[15] Blonska M, Czuba ZP, Krol W. Effect of flavone derivatives on interleukin-1beta(IL-1beta) mRNA expression and IL-1beta protein synthesis in stimulated RAW264.7 macrophages. Scand J Immunol 2003;57:162–6.
[16] Wang J, Mazza G. Inhibitory effects of anthocyanins and other phenoliccompounds on nitric oxide production in LPS/IFN-gamma-activated RAW264.7 macrophages. J Agric Food Chem 2002;50:850–7.
[17] Kuo PL. Myricetin inhibits the induction of anti-Fas IgM-, tumor necrosisfactor-alpha- and interleukin-1beta-mediated apoptosis by Fas pathway in-hibition in human osteoblastic cell line MG-63. Life Sci 2005;77:2964–76.
[18] Hsu YL, Chang JK, Tsai CH, Chien TT, Kuo PL. Myricetin induces humanosteoblast differentiation through bone morphogenetic protein-2/p38mitogen-activated protein kinase pathway. Biochem Pharmacol 2007;73:504–14.
[19] Yamaguchi M, Hamamoto R, Uchiyama S, Ishiyama K. Effects of flavonoid oncalcium content in femoral tissue culture and parathyroid hormone-stimulat-ed osteoclastogenesis in bone marrow culture in vitro. Mol Cell Biochem2007;303:83–8.
[20] Ko SY. Myricetin suppresses LPS-induced MMP expression in human gingivalfibroblasts and inhibits osteoclastogenesis by downregulating NFATc1 inRANKL-induced RAW 264.7 cells. Arch Oral Biol 2012;57:1623–32.
[21] Jin S, Park JY, Hong JM, Kim TH, Shin HI, Park EK, et al. Inhibitory effect of (-)-epigallocatechin gallate on titanium particle-induced TNF-alpha release andin vivo osteolysis. Exp Mol Med 2011;43:411–8.
[22] Liu F, Zhu Z, Mao Y, Liu M, Tang T, Qiu S. Inhibition of titanium particle-inducedosteoclastogenesis through inactivation of NFATc1 by VIVIT peptide. Bioma-terials 2009;30:1756–62.
[23] Qin A, Cheng TS, Lin Z, Cao L, Chim SM, Pavlos NJ, et al. Prevention of wearparticle-induced osteolysis by a novel V-ATPase inhibitor saliphenylha-lamide through inhibition of osteoclast bone resorption. PloS One2012;7:e34132.
[24] Wedemeyer C, Xu J, Neuerburg C, Landgraeber S, Malyar NM, von Knoch F,et al. Particle-induced osteolysis in three-dimensional micro-computed to-mography. Calcified Tissue Int 2007;81:394–402.
[25] Toro EJ, Zuo J, Ostrov DA, Catalfamo D, Bradaschia-Correa V, Arana-Chavez V,et al. Enoxacin directly inhibits osteoclastogenesis without inducing apopto-sis. J Biol Chem 2012;287:17894–904.
[26] Wang C, Steer JH, Joyce DA, Yip KH, Zheng MH, Xu J. 12-O-tetradecanoyl-phorbol-13-acetate (TPA) inhibits osteoclastogenesis by suppressing RANKL-induced NF-B activation.pdf>. J Bone Miner Res 2003;18.
[27] Helfrich MH. Osteoclast diseases and dental abnormalities. Arch Oral Biol2005;50:115–22.
[28] Abu-Amer Y, Darwech I, Clohisy JC. Aseptic loosening of total joint replace-ments: mechanisms underlying osteolysis and potential therapies. ArthritisRes Therapy 2007;9(Suppl. 1):S6.
[29] Marx RE. The deception and fallacies of sponsored randomized prospectivedouble-blinded clinical trials: the bisphosphonate research example. Int J OralMaxillofacial Implants 2014;29:e37–44.
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
[30] Maeda SS, Lazaretti-Castro M. An overview on the treatment of postmeno-pausal osteoporosis. Arquivos Brasileiros de Endocrinologia e Metabologia2014;58:162–71.
[31] Nakamura T, Sugimoto T, Nakano T, Kishimoto H, Ito M, Fukunaga M, et al.Randomized Teriparatide [human parathyroid hormone (PTH) 1-34] Once-Weekly Efficacy Research (TOWER) trial for examining the reduction in newvertebral fractures in subjects with primary osteoporosis and high fracturerisk. J Clin Endocrinol Metab 2012;97:3097–106.
[32] Hiligsmann M, Boonen A, Dirksen CD, Ben Sedrine W, Reginster JY. Cost-effectiveness of denosumab in the treatment of postmenopausal osteoporoticwomen. Expert Rev Pharmacoecon Outcomes Res 2013;13:19–28.
[33] Bi Y, Seabold JM, Kaar SG, Ragab AA, Goldberg VM, Anderson JM, et al. Adherentendotoxin on orthopedic wear particles stimulates cytokine production andosteoclast differentiation. J Bone Miner Res Off J Am Soc Bone Miner Res2001;16:2082–91.
[34] Wilson SR, Peters C, Saftig P, Bromme D, Cathepsin. K activity-dependentregulation of osteoclast actin ring formation and bone resorption. J BiologChem 2009;284:2584–92.
[35] Stevenson DA, Schwarz EL, Carey JC, Viskochil DH, Hanson H, Bauer S, et al.Bone resorption in syndromes of the Ras/MAPK pathway. Clin Genet 2011;80:566–73.
[36] Keating SE, Maloney GM, Moran EM, Bowie AG. IRAK-2 participates in multipletoll-like receptor signaling pathways to NFkappaB via activation of TRAF6ubiquitination. J Biolog Chem 2007;282:33435–43.
[37] Takaesu G, Ninomiya-Tsuji J, Kishida S, Li X, Stark GR, Matsumoto K. Interleu-kin-1 (IL-1) receptor-associated kinase leads to activation of TAK1 by induc-ing TAB2 translocation in the IL-1 signaling pathway. Mol Cell Biol 2001;21:2475–84.
[38] Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell2002;109(Suppl):S81–96.
[39] Franzoso G, Carlson L, Xing L, Poljak L, Shores EW, Brown KD, et al. Require-ment for NF-kappaB in osteoclast and B-cell development. Genes Develop1997;11:3482–96.
[40] Iotsova V, Caamano J, Loy J, Yang Y, Lewin A, Bravo R. Osteopetrosis in micelacking NF-kappaB1 and NF-kappaB2. Nature Med 1997;3:1285–9.
[41] Asagiri M, Sato K, Usami T, Ochi S, Nishina H, Yoshida H, et al. Autoamplifica-tion of NFATc1 expression determines its essential role in bone homeostasis. JExp Med 2005;202:1261–9.
[42] Yamashita T, Yao Z, Li F, Zhang Q, Badell IR, Schwarz EM, et al. NF-kappaB p50and p52 regulate receptor activator of NF-kappaB ligand (RANKL) and tumornecrosis factor-induced osteoclast precursor differentiation by activating c-Fos and NFATc1. J Biol Chem 2007;282:18245–53.
[43] Sharma SM, Bronisz A, Hu R, Patel K, Mansky KC, Sif S, et al. MITF and PU.1recruit p38 MAPK and NFATc1 to target genes during osteoclast differentia-tion. J Biol Chem 2007;282:15921–29.
[44] Matsumoto M, Kogawa M, Wada S, Takayanagi H, Tsujimoto M, Katayama S,et al. Essential role of p38 mitogen-activated protein kinase in cathepsin Kgene expression during osteoclastogenesis through association of NFATc1 andPU.1. J Biol Chem 2004;279:45969–79.
[45] Huang H, Chang EJ, Ryu J, Lee ZH, Lee Y, Kim HH. Induction of c-Fos and NFATc1during RANKL-stimulated osteoclast differentiation is mediated by the p38signaling pathway. Biochem Biophys Res Commun 2006;351:99–105.
[46] Lucas JJ, Yamamoto A, Scearce-Levie K, Saudou F, Hen R. Absence of fenflur-amine-induced anorexia and reduced c-Fos induction in the hypothalamusand central amygdaloid complex of serotonin 1B receptor knock-out mice. JNeurosci Off J Soc Neurosci 1998;18:5537–44.
[47] Takayanagi H, Kim S, Koga T, Nishina H, Isshiki M, Yoshida H, et al. Induction andactivation of the transcription factor NFATc1 (NFAT2) integrate RANKL signalingin terminal differentiation of osteoclasts. Develop Cell 2002;3:889–901.
[48] Greenfield EM, Bi Y, Ragab AA, Goldberg VM, Van De Motter RR. The role ofosteoclast differentiation in aseptic loosening. J Orthop Res Off Publ OrthopRes Soc 2002;20:1–8.
[49] Holt G, Murnaghan C, Reilly J, Meek RM. The biology of aseptic osteolysis. ClinOrthop Relat Res 2007;460:240–52.
[50] Merkel KD, Erdmann JM, McHugh KP, Abu-Amer Y, Ross FP, Teitelbaum SL.Tumor necrosis factor-alpha mediates orthopedic implant osteolysis. Am JPathol 1999;154:203–10.
[51] Rakshit DS, Ly K, Sengupta TK, Nestor BJ, Sculco TP, Ivashkiv LB, et al. Weardebris inhibition of anti-osteoclastogenic signaling by interleukin-6 and in-terferon-gamma. Mechanistic insights and implications for periprostheticosteolysis. J Bone Joint Surg Am Vol 2006;88:788–99.
[52] Nakashima Y, Sun DH, Trindade MC, Maloney WJ, Goodman SB, Schurman DJ,et al. Signaling pathways for tumor necrosis factor-alpha and interleukin-6expression in human macrophages exposed to titanium-alloy particulatedebris in vitro. J Bone Joint Surg Am Vol 1999;81:603–15.
[53] Wang W, Ferguson DJ, Quinn JM, Simpson AH, Athanasou NA. Biomaterialparticle phagocytosis by bone-resorbing osteoclasts. J Bone Joint Surg Brit Vol1997;79:849–56.
[54] Purdue PE, Koulouvaris P, Potter HG, Nestor BJ, Sculco TP. The cellular andmolecular biology of periprosthetic osteolysis. Clin Orthop Relat Res2007;454:251–61.
[55] Datta HK, Ng WF, Walker JA, Tuck SP, Varanasi SS. The cell biology of bonemetabolism. J Clin Pathol 2008;61:577–87.
[56] Hirakawa K, Bauer TW, Stulberg BN, Wilde AH. Comparison and quantitationof wear debris of failed total hip and total knee arthroplasty. J Biomed MaterRes 1996;31:257–63.
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019

C. Wu et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 13
G Model
BCP-12131; No. of Pages 13
[57] Baumann B, Seufert J, Jakob F, Noth U, Rolf O, Eulert J, et al. Activation of NF-kappaB signalling and TNFalpha-expression in THP-1 macrophages by TiAlV-and polyethylene-wear particles. J Orthop Res Off Publ Orthop Res Soc2005;23:1241–8.
[58] von Knoch M, Jewison DE, Sibonga JD, Sprecher C, Morrey BF, Loer F, et al. Theeffectiveness of polyethylene versus titanium particles in inducing osteolysisin vivo. J Orthop Res Off Publ Orthop Res Soc 2004;22:237–43.
Please cite this article in press as: Wu C, et al. Myricetin prevents tinduced osteoclastogenesis in vitro. Biochem Pharmacol (2014), http
[59] Masui T, Sakano S, Hasegawa Y, Warashina H, Ishiguro N. Expressionof inflammatory cytokines, RANKL and OPG induced by titanium,cobalt-chromium and polyethylene particles. Biomaterials 2005;26:1695–702.
[60] Baumann B, Rader CP, Seufert J, Noth U, Rolf O, Eulert J, et al. Effects ofpolyethylene and TiAlV wear particles on expression of RANK, RANKL and OPGmRNA. Acta Orthop Scand 2004;75:295–302.
itanium particle-induced osteolysis in vivo and inhibits RANKL-://dx.doi.org/10.1016/j.bcp.2014.10.019


















