
MYCOBACTERIUM LEPRAE FROM LEPROSY SKIN ......Mycobacterium leprae 2 •First bacterial pathogen to...
18
WHOLE GENOME SEQUENCING OF MYCOBACTERIUM LEPRAE FROM LEPROSY SKIN BIOPSIES and skeletons Pushpendra Singh Laboratory of Prof. Stewart Cole Global Health Institute School of Life Sciences Ecole Polytechnique Fédérale de Lausanne, Switzerland.
Transcript of MYCOBACTERIUM LEPRAE FROM LEPROSY SKIN ......Mycobacterium leprae 2 •First bacterial pathogen to...

WHOLE GENOME SEQUENCING OF MYCOBACTERIUM LEPRAE FROM
LEPROSY SKIN BIOPSIES and skeletons
Pushpendra Singh
Laboratory of Prof. Stewart Cole Global Health Institute School of Life Sciences
Ecole Polytechnique Fédérale de Lausanne, Switzerland.

Mycobacterium leprae
2
• First bacterial pathogen to be discovered (Hansen 1873) - still remains uncultured in vitro.
- over 1300 pseudogenes.
• Very long doubling time (2 weeks)
• Very low genetic diversity, very few SNPs: -Challenging for genotyping.
-Optimal for re-sequencing projects and mapping.
= several opportunities

PCR based M. leprae Genotyping
3
• SNPs
- Excellent phylogeographic association
- Very limited resolution (only 16 subtypes known)
- Type 1(A,B,C,D), 2(E,F,G,H), 3(I,J,K,L,M) and 4(N,O,P).
• VNTRs
- High resolution but limited phylogeographic association.
- Intra-patient variability at different sites of infection.

Limitations of PCR-based genotyping
4
• Limited information
- Targeted information and hence not comprehensive.
- Many loci need to be investigated (Not effort-effective).
• Scarce amounts of gDNA from an unculturable pathogen
- Input of whole genome template for a few kb of information.
- not a renewable approach (Empty tubes phenomenon).
Genome-wide analysis (GENOME-Typing) offers great potential, given the
improvements in sequencing technologies and sample prep methods (1-10 ng gDNA).

Outline of the present study 1. DNA isolation directly from leprosy skin
biopsies from diverse geographical areas (n=12) and medieval leprosy skeletons from Europe, 700-1000
yr old).
2. Library preparation and array-based capture of M. leprae DNA.
3. Sequencing and genome-wide comparison (medieval and modern M. leprae).
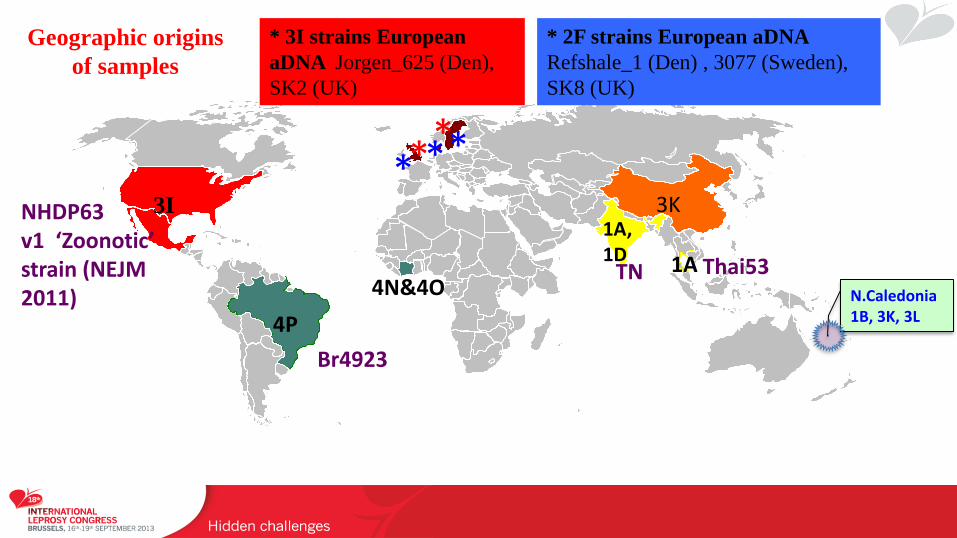
* * * *
* 3I strains European
aDNA Jorgen_625 (Den),
SK2 (UK)
4P
3I
N.Caledonia 1B, 3K, 3L
3K 1A, 1D 1A
* 2F strains European aDNA
Refshale_1 (Den) , 3077 (Sweden),
SK8 (UK)
Br4923
NHDP63 v1 ‘Zoonotic’ strain (NEJM 2011)
TN Thai53 4N&4O
*
Geographic origins
of samples
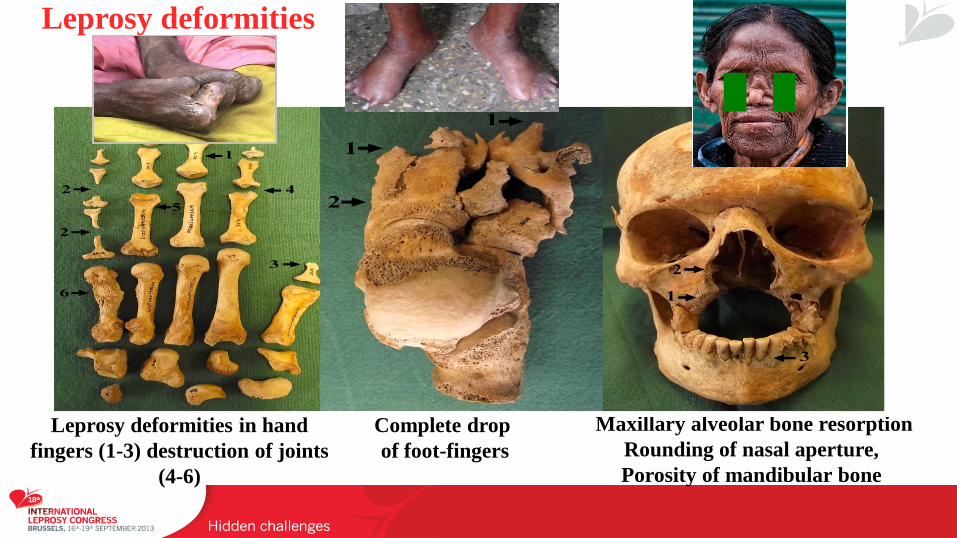
Complete drop
of foot-fingers
Leprosy deformities in hand
fingers (1-3) destruction of joints
(4-6)
Maxillary alveolar bone resorption
Rounding of nasal aperture,
Porosity of mandibular bone
Leprosy deformities
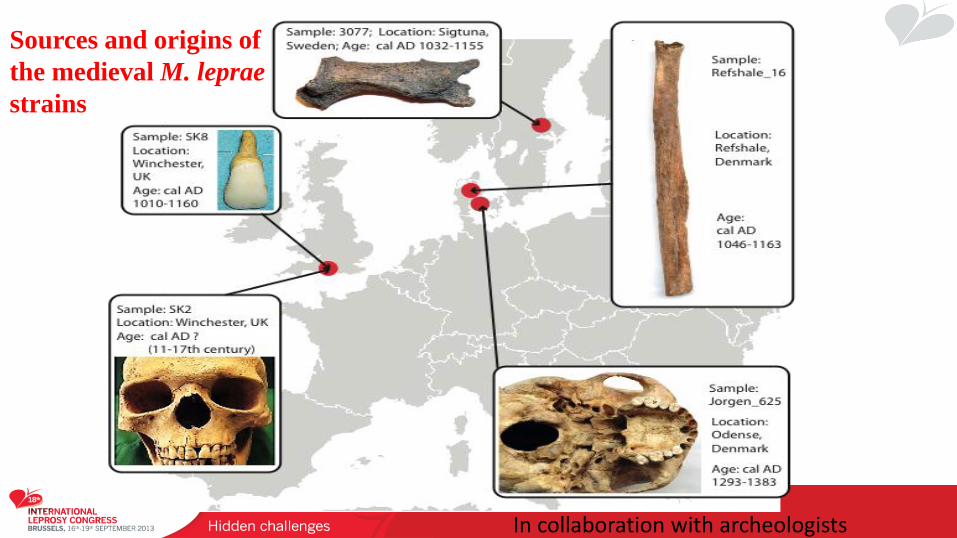
Sources and origins of
the medieval M. leprae
strains
In collaboration with archeologists
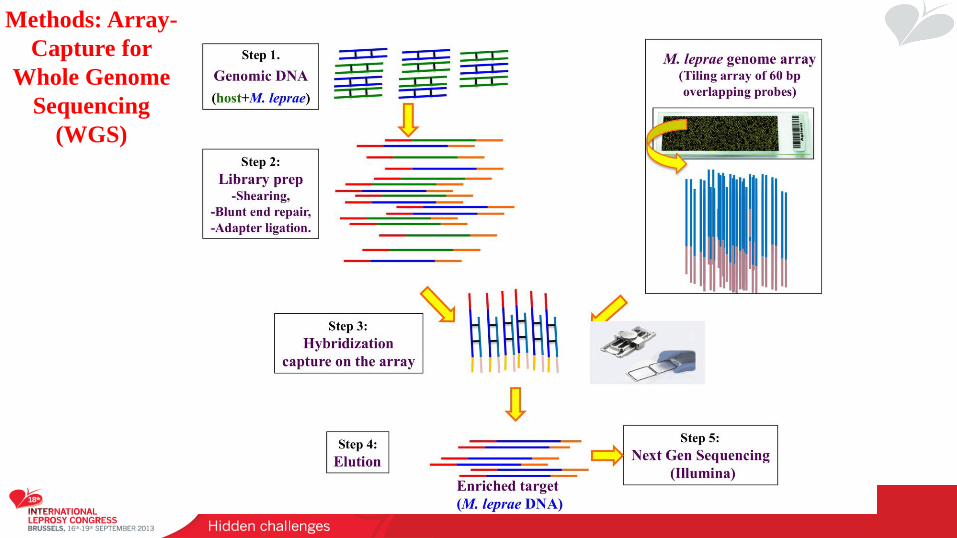
Methods: Array-
Capture for
Whole Genome
Sequencing
(WGS)

WGS results
10
1. Genome wide coverage of 5 ancient and 7 modern strains (coverage of >80% of genome at > 10X).
2. Comparison with 4 reference strains
3. Detailed & Diverse “Phylo-Geo-Chrono”
comparison: 16 genomes (separated by Evolution, Space, and Time):
• Phylogeny: from all four SNP types.
• Geographic: from all 5 continents.
• Chrono (Time): 1000 yr time span (10th century - 2012).
As few as 755 SNPs, and 57 InDels.
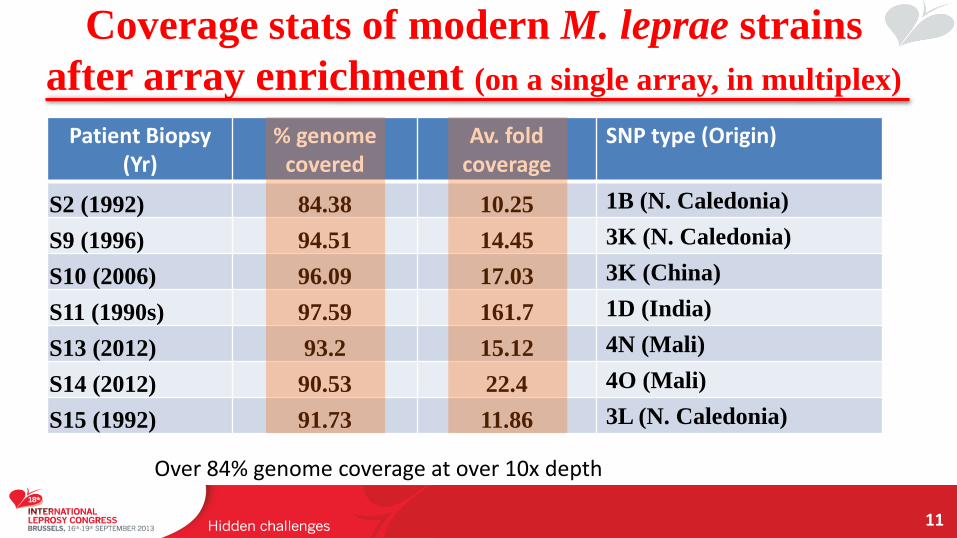
Coverage stats of modern M. leprae strains
after array enrichment (on a single array, in multiplex)
11
Patient Biopsy (Yr)
% genome covered
Av. fold coverage
SNP type (Origin)
S2 (1992) 84.38 10.25 1B (N. Caledonia)
S9 (1996) 94.51 14.45 3K (N. Caledonia)
S10 (2006) 96.09 17.03 3K (China)
S11 (1990s) 97.59 161.7 1D (India)
S13 (2012) 93.2 15.12 4N (Mali)
S14 (2012) 90.53 22.4 4O (Mali)
S15 (1992) 91.73 11.86 3L (N. Caledonia)
Over 84% genome coverage at over 10x depth
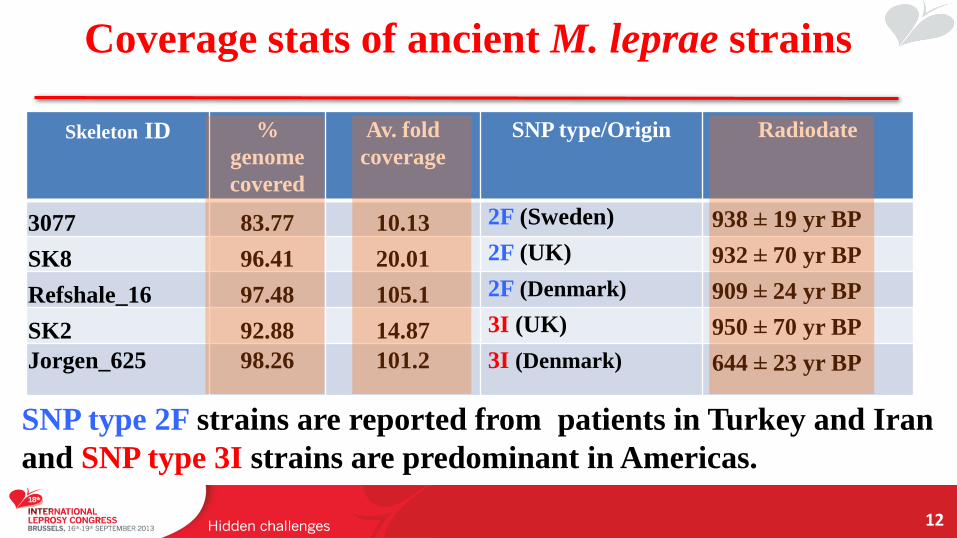
Coverage stats of ancient M. leprae strains
12
Skeleton ID %
genome
covered
Av. fold
coverage
SNP type/Origin Radiodate
3077 83.77 10.13 2F (Sweden) 938 ± 19 yr BP
SK8 96.41 20.01 2F (UK) 932 ± 70 yr BP
Refshale_16 97.48 105.1 2F (Denmark) 909 ± 24 yr BP
SK2 92.88 14.87 3I (UK) 950 ± 70 yr BP
Jorgen_625 98.26 101.2 3I (Denmark) 644 ± 23 yr BP
SNP type 2F strains are reported from patients in Turkey and Iran
and SNP type 3I strains are predominant in Americas.

Phylogenetic relationship of M. leprae genomes &
divergence times
Branch 2 strains (2F) are currently reported from Middle East.
Branch 3 strains (3I) are found in Americas, (almost identical to zoonotic v1 strain from southern US
(Truman et al NEJM 2011).
~3400 yrs
300 yrs

Genome-Typing for revealing strain specific
features of the biology of a pathogen
14
• 10 NEW pseudogenes were identified in various strains.
• Premature stop codons by SNPs.
• Frameshift caused by Insertion/Deletion events (InDels).
• Drug resistance mutations
• Thr53Ile in folP1 (Dapsone Resistance mutation in 2 strains).
• High diversity in an immunodominant protein encoded by serine-rich antigen ML0411:
• 10 Non-Synonymous SNPs = immune pressure from the host? (Also reported recently in M. leprae Kyoto-2 genome from Japan (Kai et al, Infect
Genet Evol 2013).

Important findings by genome scale comparisons
15
1. No major variations in M. leprae in past 1000 yrs.
– No evidences of change in the virulence/adaptability etc.
– Sudden decline of leprosy in Europe in 14th century was NOT due
to the medieval European strain of M. leprae losing virulence.
2. Confirmation of European origin of Leprosy in Americas.
3. Link between medieval European strains and those
currently reported from the Middle East. (SNP type 2F strains of
Iran & Turkey were present in medieval skeletons from 3 European countries).

Why genome-typing instead of genotyping?
16
• Comprehensive information
All in one:
- Genotyping.
- Molecular drug susceptibility testing.
- Detailed phylogeny.
- Identification of any additional pseudogenes in a strain.
- Uniform experimental approach for all strains.
- Genome scale investigations into the biology of a strain.
• Why not performed routinely then?
- Costs and technical difficulties.
- streamlining of bioinformatic analysis needed.

Acknowledgements
17
• Stewart Cole
– Andrej Benjak, Philippe Busso, all other members of Prof Cole lab (UPCOL).
• Univ. of Surrey: GM Taylor, GR Stewart, TA Mendum.
• Tuebingen: Johannes Krause Lab (Verena Schuenemann & Kirsten Bos)
• Kay Nieselt Lab (Alexander Herbig, Günter Jäger).
Christos (Swedish aDNA), Ben Kyora (Danish aDNA)
Teams of Dr Samba Sow for samples from Mali, C. Johnson from Benin and collaborators from Niger.
Ongoing studies using array capture approach:
Milton Moraes, Luciana, P Suffys and Dr Samira Buhrer-Sekula and their teams
• All patients, clinicians and researchers for contributing clinical samples & relevant details.
Thank you for your attention
Brazil Swiss Joint Research Project

Thank you for your attention!















![[Micro] mycobacterium leprae](https://static.fdocuments.net/doc/165x107/55d6fd2cbb61eb344d8b45f6/micro-mycobacterium-leprae.jpg)


