
Morphometry of in vivo human white matter association pathways with diffusion-weighted magnetic...
12
Morphometry of In Vivo Human White Matter Association Pathways with Diffusion- Weighted Magnetic Resonance Imaging N. Makris, MD,* A. J. Worth, PhD,* A. G. Sorensen, MD,$ G. M. Papadimitriou, BS,* 0. Wu, MS,$ T. G. Reese, PhD,$ V. J. Wedeen, MD,$ T. L. Davis, MD,S J. W. Stakes, MD,* V. S. Caviness, MD, DPhil,* E. Kaplan, PhD,§ B. R. Rosen, MD, PhD,$ D. N. Pandya, MD,t and D. N. Kennedy, PhD*$ The precise characterization of cortical connectivity is important for the understanding of brain morphological and functional organization. Such connectivity is conveyed by specific pathways or tracts in the white matter. Diffusion- weighted magnetic resonance imaging detects the diffisivity of water molecules in three dimensions. Diffisivity is aniso- tropic in oriented tissues such as fiber tracts. In the present study, we used this method to map (in terms of orientation, location, and size) the “stem” (compact portion) of the principal association, projection, and commissural white matter pathways of the human brain in vivo, in 3 normal subjects. In addition, its use in clinical neurology is illustrated in a patient with left inferior parietal lobule embolic infarction in whom a significant reduction in relative size of the stem of the left superior longitudinal fasciculus was observed. This represents an important method for the characterization of major association pathways in the living human that are not discernible by conventional magnetic resonance imaging. In the clinical domain, this method will have a potential impact on the understanding of the diseases that involve white matter such as stroke, multiple sclerosis, amyotrophic lateral sclerosis, head injury, and spinal cord injury. Makris N, Worth AJ, Sorensen AG, Papadimitriou GM, Wu 0, Reese TG, Wedeen VJ, Davis TL, Stakes JW, Caviness VS, Kaplan E, Rosen BR, Pandya DN, Kennedy DN. Morphometry of in vivo human white matter association pathways with diffusion-weighted magnetic resonance imaging. Ann Neurol 1997;42:95 1-962 The central white matter of the cerebral hemisphere includes axons serving three principal classes of connec- tions, namely, associational, projectional, and commis- sural. It is a general property of these white matter fi- ber pathways that their axons tend to fasciculate into “stems,” converging from origin into compact, more or less homogeneous bundles, from which they ultimately diverge to termination [I], Anatomical connectivity in the cerebral cortex is mediated by long association pathways within the white matter. In experimental an- imals, the complete delineation of the fiber pathways has been performed using anterograde and retrograde tracing techniques [2, 31. In human postmortem ma- terial, traditional techniques such as myelin stain allow the visualization of the stems of these fiber bundles [I]. Using the myelin staining approach, the major white matter tracts have been described precisely in terms of topography, dimension, and orientation of their trajec- tories in the human brain at the level of their stems [ 11. Available histological description of human fiber pathways is incomplete because it does not provide a detailed understanding of their distal or peripheral ter- minations. There is a clear clinical need for methods to characterize human white matter tracts in vivo. Magnetic resonance imaging (MRI) has become the method of choice for the clinical evaluation of the brain in normal and pathological conditions. MRI al- lows anatomical differentiation of gray and white mat- ter tissues and thus permits the topographic and vol- umetric characterization of specific regions of the cerebral cortex and subcortical structures in vivo [4]. Developments in diffusion-weighted imaging (DWI) using magnetic resonance have made it possible to measure the diffusivity of MRI visible water molecules. Water diffusivity in DWI is characterized by the effec- tive diffusion tensor, 0, which describes the magnitude From the *Department of Neurology, Massachusetts General Hos- pital, Harvard Medical School, and Departments of ?Anatomy and Neurobiology, and §Neurology, Psychiatry, and Behavioral Neuro- science, Boston University School of Medicine, Boston; $Depart- ment of Radiology, MGH-NMR Center, Massachusetts General Hospital, Harvard Medical School, Charlestown, MA; and SMallinckrodt Institute of Radiology, Washington University, St Louis, MO. Received Jun 11, 1997, and in revised form Aug 22. Accepted for publication Aug 22, 1997. Address correspondence to Dr Makris, MGH-CMA, 149 13th Street, Charlestown, MA 02129. Copyright 0 1997 by the American Neurological Association 951
-
Upload
dr-n-makris -
Category
Documents
-
view
224 -
download
6
Transcript of Morphometry of in vivo human white matter association pathways with diffusion-weighted magnetic...

Morphometry of In Vivo Human White Matter Association Pathways with Diffusion-
Weighted Magnetic Resonance Imaging N. Makris, MD,* A. J. Worth, PhD,* A. G. Sorensen, MD,$ G. M. Papadimitriou, BS,* 0. Wu, MS,$
T. G. Reese, PhD,$ V. J. Wedeen, MD,$ T. L. Davis, MD,S J. W. Stakes, MD,* V. S. Caviness, MD, DPhil,* E. Kaplan, PhD,§ B. R. Rosen, MD, PhD,$
D. N. Pandya, MD,t and D. N. Kennedy, PhD*$
The precise characterization of cortical connectivity is important for the understanding of brain morphological and functional organization. Such connectivity is conveyed by specific pathways or tracts in the white matter. Diffusion- weighted magnetic resonance imaging detects the diffisivity of water molecules in three dimensions. Diffisivity is aniso- tropic in oriented tissues such as fiber tracts. In the present study, we used this method to map (in terms of orientation, location, and size) the “stem” (compact portion) of the principal association, projection, and commissural white matter pathways of the human brain in vivo, in 3 normal subjects. In addition, its use in clinical neurology is illustrated in a patient with left inferior parietal lobule embolic infarction in whom a significant reduction in relative size of the stem of the left superior longitudinal fasciculus was observed. This represents an important method for the characterization of major association pathways in the living human that are not discernible by conventional magnetic resonance imaging. In the clinical domain, this method will have a potential impact on the understanding of the diseases that involve white matter such as stroke, multiple sclerosis, amyotrophic lateral sclerosis, head injury, and spinal cord injury.
Makris N, Wor th AJ, Sorensen AG, Papadimitriou GM, W u 0, Reese TG, Wedeen VJ, Davis TL, Stakes JW, Caviness VS, Kaplan E, Rosen BR, Pandya D N , Kennedy DN. Morphometry of in vivo human white matter
association pathways with diffusion-weighted magnetic resonance imaging. Ann Neurol 1997;42:95 1-962
The central white matter of the cerebral hemisphere includes axons serving three principal classes of connec- tions, namely, associational, projectional, and commis- sural. It is a general property of these white matter fi- ber pathways that their axons tend to fasciculate into “stems,” converging from origin into compact, more or less homogeneous bundles, from which they ultimately diverge to termination [I] , Anatomical connectivity in the cerebral cortex is mediated by long association pathways within the white matter. In experimental an- imals, the complete delineation of the fiber pathways has been performed using anterograde and retrograde tracing techniques [2, 31. In human postmortem ma- terial, traditional techniques such as myelin stain allow the visualization of the stems of these fiber bundles [I]. Using the myelin staining approach, the major white matter tracts have been described precisely in terms of topography, dimension, and orientation of their trajec-
tories in the human brain at the level of their stems [ 11. Available histological description of human fiber pathways is incomplete because it does not provide a detailed understanding of their distal or peripheral ter- minations. There is a clear clinical need for methods to characterize human white matter tracts in vivo.
Magnetic resonance imaging (MRI) has become the method of choice for the clinical evaluation of the brain in normal and pathological conditions. MRI al- lows anatomical differentiation of gray and white mat- ter tissues and thus permits the topographic and vol- umetric characterization of specific regions of the cerebral cortex and subcortical structures in vivo [4]. Developments in diffusion-weighted imaging (DWI) using magnetic resonance have made it possible to measure the diffusivity of MRI visible water molecules. Water diffusivity in DWI is characterized by the effec- tive diffusion tensor, 0, which describes the magnitude
From the *Department of Neurology, Massachusetts General Hos- pital, Harvard Medical School, and Departments of ?Anatomy and Neurobiology, and §Neurology, Psychiatry, and Behavioral Neuro- science, Boston University School of Medicine, Boston; $Depart- ment of Radiology, MGH-NMR Center, Massachusetts General Hospital, Harvard Medical School, Charlestown, MA; and SMallinckrodt Institute of Radiology, Washington University, St Louis, MO.
Received Jun 11, 1997, and in revised form Aug 22. Accepted for publication Aug 22, 1997.
Address correspondence to Dr Makris, MGH-CMA, 149 13th Street, Charlestown, MA 02129.
Copyright 0 1997 by the American Neurological Association 951

and direction of water mobility in the tissues in three dimensions. Some uses of DWI have focused on the “apparent diffusion coefficient” [ S , 61, a scalar repre- sentation of the total net water diffusion present in each voxel. The use of diffusion anisotropy was intro- duced [7-101 to consider directional specificity. Most recently, methods to characterize the complete diffu- sion tensor have been developed [ 1 1, 121. These meth- ods have allowed the detection of major white matter structures in the central nervous system such as the corpus callosum and the internal capsule in humans in vivo [13, 141 and in experimental animals [15].
Using an orientation color coding system [9] to vi- sualize the primary direction of the diffusion tensor, we tested the hypothesis that DWI can characterize the stems of the principal cerebral white matter pathways in the human brain in terms of their location, size, and trajectory. In the present study, we demonstrated the ability to characterize the stems of more discrete corti- cocortical association pathways than have been previ- ously reported in living humans. This is an advance over the previous literature in that, by using an atlas- guided analytic approach, we have identified the prin- cipal set of association pathways in living human sub- jects. The utility of this type of observation is then demonstrated in a stroke patient. The structural char- acterization of white matter tracts in vivo has impor- tant implications for understanding normal anatomy and function, as well as neurological disorders that present and/or result from damage to such fiber path- ways such as stroke, multiple sclerosis, amyotrophic lat- eral sclerosis, head injury, and spinal cord injury.
Subjects and Methods To test our hypothesis regarding the identification of discrete fiber pathways in the human brain in vivo, we (1) defined our a priori anatomical understanding about a specific subset of fiber pathways of interest, (2) acquired DWI on a set of
subjects, (3) generated maps of various diffusion properties, and (4) evaluated the correspondence of these maps with our anatomical definitions by ascertaining the degree to which the a priori expectations matched the observed data.
Neuroanatomy We have selected a subset of fiber pathways that present a range of orientations and sizes to form the anatomical basis of our hypothesis. These are listed in Table 1, which in- cludes their anatomical location, size, and orientation based on a qualitative interpretation of the Dejerine [l] map of the human brain. This table refers to the compact portion, or stem, of the fiber tracts, an example of which is shown sche- matically in Figure 1. Throughout the remainder of this text when we refer to pathways, we refer to the stem of the path- way. The approximate size of the cross section of different pathways was calculated from the Dejerine anatomical map. From the topographic features of these pathways, we have designated representative anatomical regions of interest (ROIs) on the Dejerine map that indicate their expected blocked-outlined neighborhood location, an example of which is shown in Figure 2a.
Subjects In 3 normal right-handed male subjects (mean age = 35 years) the white matter fiber pathways were analyzed using DWI. In addition, one 69-year-old, right-handed male pa- tient was studied. This patient suffered an embolic infarction in the left inferior parietal lobule at 65 years of age (4.5 years before DWl).
Neuroimaging MRI scanning was performed using a General Electric 1 .S-T Signa (modified for instant imaging by Advanced NMK, Wilmington, MA). Scans included conventional (multislice T1-weighted sagittal scan for prescription of subsequent im- ages) and echo-planar DWI (Fig 3). The echo-planar-based protocol included automatic magnetic field shimming and coronal diffusion tensor imaging. The coronal imaging plane was set to be perpendicular to the bicommissural (anterior
Table 1. Nomenchture and Features (location, Size, and Orientation) o f the Compact Portions or Sterns o f the Dzferent White Matter Pathways Based on the Dejerine Human Map
White Matter Pathway
Superior longitudinal fasciculus (S1.F) (longitudinal portion) Cingulum bundle (CB) Inferior longitudinal fasciculus and optic radiation complex (ILFIOR) Occipitofrontal fasciculus (OF) Uncinare fasciculus (UF) Extreme capsule (EC) Fornix (Fo) (longitudinal part) Corpus callosurn (CC) Anterior zommissure (AC) Internal capsule (IC) Cerebral peduncle (P)
Extent of Stem: Dejerine May
Approximate Size Range Orientation in Cross Section (mm) (Major Axis)
37-92 30-105 75-137 30-88 40-63
66-92
55-69 43-86
55-83
33-105
75-86
6-1 0 5-8 5-7 4-6 3-5 2-6 2-8 5-15 2-5 5-12 7-12
AP AP AP AP AP AP AP ML ML s1 SI
‘The numbers within the columns showing the extent of the stems in the Dejerine map refer to the range of coronal levels in which thc stems of the specific pathways are present.
AP = anterior-posterior; ML = mediolatcral; SI = superior-inferior.
952 Annals of Neurology Vol 42 No 6 December 1977
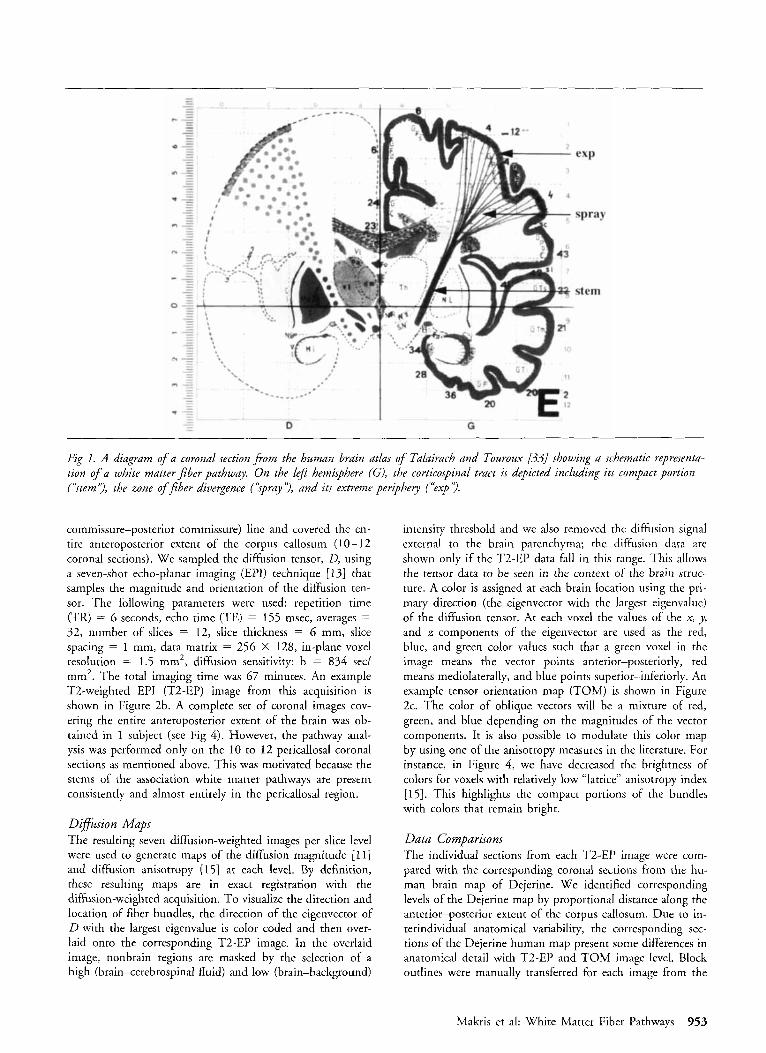
Fig I . A diagram of a coronal section ji-om the human brain adas of Talairach and Touroux [33] showing a schematic representa- tion of a white matter fiber pathway. On the lefi hemisphere (G), the corticospinal tract is depicted including its compact portion (‘item’?, the zone of fiber divergence (“spray’?, and its extreme periphery (“exp 3.
commissure-posterior conimissure) line and covered the en- tire anteroposterior extent of the corpus callosum (10-12 coronal sections). We sampled the diffusion tensor, 0, using a seven-shot echo-planar imaging (EPI) technique [ 131 that samples the magnitude and orientation of the diffusion ten- sor. The following parameters were used: repetition time (TR) = 6 seconds, echo time (TE) = 155 msec, averages = 32, number of slices = 12, slice thickness = 6 mm, slice spacing = 1 mm, data matrix = 256 X 128, in-plane voxel resolution = 1.5 mm2, diffusion sensitivity: b = 834 secl mm2. The total imaging time was 67 minutes. An example T2-weighted EPI (T2-EP) image from this acquisition is shown in Figure 2b. A complete set of coronal images cov- ering the entire anteroposterior extent of the brain was ob- tained in 1 subject (see Fig 4). However, the pathway anal- ysis was performed only on the 10 to 12 pericallosal coronal sections as mentioned above. This was motivated because the stems of the association white matter pathways are present consistently and almost entirely in the pericallosal region.
Di’sion Maps The resulting seven diffusion-weighted images per slice level were used to generate maps of the diffusion magnitude [I I ] and diffusion anisotropy [15] at each level. By definition, these resulting maps are in exact registration with the diffusion-weighted acquisition. To visualize the direction and location of fiber bundles, the direction of the eigenvector of D with the largest eigenvalue is color coded and then over- laid onto the corresponding T2-EP image. In the overlaid image, nonbrain regions are masked by the selection of a high (brain-cerebrospinal fluid) and low (brain-background)
intensity threshold and we also removed the diffusion signal external to the brain parenchyma; the diffusion data are shown only if the T2-EP data fall in this range. This allows the tensor data to be seen in the context of the brain struc- ture. A color is assigned at each brain location using the pri- mary direction (the eigenvector with the largest eigenvalue) of the diffusion tensor. At each voxel the values of the x, y, and z components of the eigenvector are used as the red, blue, and green color values such that a green voxel in the image means the vector points anterior-posteriorly, red means mediolaterally, and blue points superior-inferiorly. An example tensor orientation map (TOM) is shown in Figure 2c. The color of oblique vectors will be a mixture of red, green, and blue depending on the magnitudes of the vector components. It is also possible to modulate this color map by using one of the anisotropy measures in the literature. For instance, in Figure 4, we have decreased the brightness of colors for voxels with relatively low “lattice” anisotropy index [15]. This highlights the compact portions of the bundles with colors that remain bright.
Data Comparisons The individual sections from each T2-EP image were com- pared with the corresponding coronal sections from the hu- man brain map of Dejerine. We identified corresponding levels of the Dejerine map by proportional distance along the anterior-posterior extent of che corpus callosum. Due to in- terindividual anatomical variability, the corresponding sec- tions of the Dejerine human map present some differences in anatomical detail with T2-EP and T O M image level. Block outlines were manually transferred for each image from the
Makris et al: White Matter Fiber Pathways 953

Fig 2
C
Right Left Right Left
Fig 3
954 Annals of Neurology Vol 42 No 6 December 1997

Fig 2. On the 14 (a), diapam shows a coronal section taken fiom the book by Dejerine [I] at the level o f the red nucleus (6 mm anterior to the posterior commissure). The anatomical regions containing major white matter fiber pathways are “blocked ’’ using a geometrical region o f interest outline. SLF = superior longitudinal fasciculus; CB = cingulum bundle; OF = occ+itofiontal fdscicu- lus; EC = extreme capsule; ILF/OR = complex of inferior longitudinalfasciculus and optic radiations; Fo = fornix; CC = corpus callosum; IC = internal capsule; P = cerebral peduncle; BS = brainstem; sf= syluian jssure; lv = lateral ventricle; hm = hemi- spheric margin. On the right, at the top (6) is a T2-weighted echo-planar image of a normal subject at a level anatomically corre- sponding to (a). The predicted regions are scaled to j t the anatomical features that are present at this level. On the right, at the bot- tom (c) is a color-coded tensor orientation map (TOM) that is in exact registration with the image seen in (6). Each pixel in the image is assigned a color based on the direction of the largest eigenualue o f the difision tensor. Blue color coding corresponds to superior-inferior, green to anterior-posterior, and red to mediolateral.
1
Dejerine map to the corresponding T2-EP sections by using visible anatomical landmarks such as the ventricles, sylvian fissure, internal capsule, corpus callosum, and hemispheric margins. This matching procedure was performed manually on each image and required small amounts of correction in terms of scaling, rotation, and translation to maintain the relative anatomical relationship of the regions (see Fig 2b). These matched outlines of T2-EP were then overlaid directly onto its corresponding TOM (see Fig 2c). As the T2-EP and TOM images were in registration, no further spatial correc- tion was required.
Each ROI was evaluated for the presence of a cluster of voxels of the specified orientation and size for the fiber path- way that generated the ROI. In addition, the “size” of white matter tracts could be estimated by counting the number of voxels within the ROIs, which have tensors oriented within 45” of the predicted orientation.
Results The results of this study were threefold: (1) acquisition of “high-resolution’’ diffusion tensor images that were registered to an anatomical map, (2) visualization of tensor orientation by color coding, and (3) comparison of the diffusion tensor orientation information with anatomical predictions of fiber tract location. This analysis was performed on 3 normal young adult sub- jects, and 1 stroke patient.
Difision Imaging For each subject the following data were generated for each coronal level: (1) a set of seven “raw” diffusion- weighted images, (2) a diffusion tensor map, and (3) a color-coded TOM. Each of these data provided unique information about the white matter. We used one of the T2-EP images to identify the anatomical context of the coronal level through the observation of features such as the hemispheric margins, interhemispheric fis- sure, sylvian fissure, internal capsule, corpus callosum,
ventricular system, and brainstem (see Fig 2b). A dis- tinct color-coded pattern of orientation for the com- plete cerebral hemispheres can be seen in the color- coded tensor orientation image (see Fig 2c). The overall appearance of this image is consistent, in its broad context, with the general expectation of fiber pathways in the brain. Specifically, superior-inferiorly oriented tensors (color-coded blue) form a broad, largely homogeneous band that courses parasagittally in each hemisphere and corresponds to the internal cap- sule seen in the T2-EP image. These bands continue inferiorly as the cerebral peduncles and join at the brainstem. Mediolaterally oriented tensors (color-coded red) are apparent as a strip above the roof of the lateral ventricles and correspond to the corpus callosum seen in the T2-EP image. The corpus callosum and the in- ternal capsule form a “frame of reference” for the iden- tification of other fiber pathways. Multiple regions of anteroposteriorly oriented tensors (color-coded green) are observed laterally and adjacent to the internal cap- sules. In addition, this orientation pattern is observed immediately superior to the corpus callosum as well as around the external superior corner of the lateral ven- tricle and at the midline under the floor of the lateral ventricles. As one approaches the lateral hemispheric margins, mediolaterally (color-coded red) oriented ten- sors are predominantly observed. This is in contrast to the neighborhood of the superior hemispheric margin where superior-inferior (color-coded blue) orientation is preferentially observed. These observations are con- sistent with the pattern of the fiber systems of the sub- cortical corona radiata, which would be expected at the lateral cerebral regions to be mainly rendered in red due to their predominant transverse course, as opposed to blue at the superior cerebral regions, which would be consistent with their expected predominantly verti-
Fig 3. Dzfision and TI-weighted images of a patient with a lesion involving the l e j inferior parietal lobule. Difision and TI im- 4 ages are not in exact registration. (a) and (6) are at the equivalent level ofthe normal subject in Figure 26. (c) and (4 are at the level o f the splenium of the corpus callosum. (a and c) TI-weighted images. (b and d) Tensor orientation map images as seen in Figure 2c. (c and d) The location of the lesion is indicated. Note the absence of pixels indicating anterior-posterior orientation (green) in (d) at the site of the lesion. The arrows in (6) show the predicted location of the superior longitudinal fasciculus and the cingulum bundle in both hemispheres. In (b), note the reduction in number of the pixels indicating anterior-posterior orientation (green) for the superior longitudinal fasciculus on the patient? lej. SLF = superior longitudinal fasciculus; CB = cingulum bundle.
Makris et al: White Matter Fiber Pathways 955
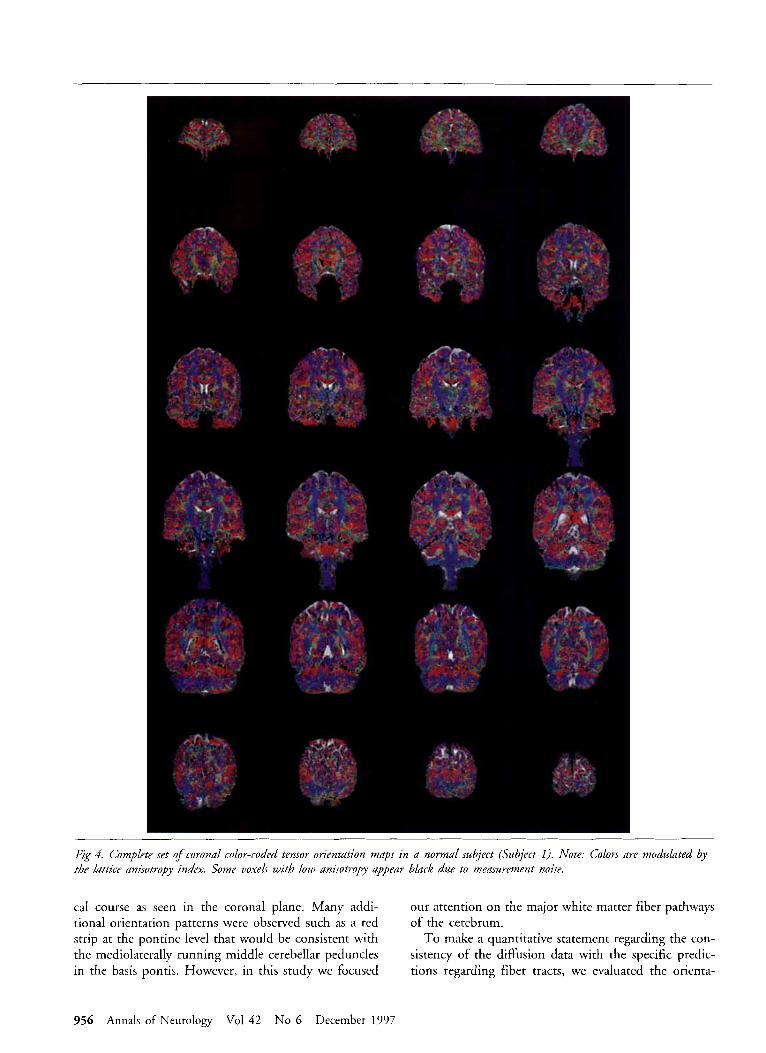
Fig 4. Complete set of covonal color-coded tensor orientation maps in a normal subject (Subject 1). Note: Colors are modulated by tbe Lattice anisotropy index. Some voxels with low anisotropy appear black due to measurement noise.
cal course as seen in the coronal plane. Many addi- tional orientation patterns were observed such as a red strip at the pontine level that would be consiscent with the mediolaterally running middle cerebellar peduncles in the basis pontis. However, in this study we focused
our attention on the major white matter fiber pathways of the cerebrum.
To make a quantitative statement regarding the con- sistency of the diffusion data with the specific predic- tions regarding fiber tracts, we evaluated the orienta-
956 Annals of Neurology Vol 42 No 6 December 1397

Midsagittal of Dejerine
SLF
CB
Fig 5. Schematic representation of the observations of a full set o f coronal sections taken through the anteroposterior extent of the corpus callosum, corresponding to coronal levels 33 through 105 of the Dejerine human map o f 3 normal subjects (Subjects I , 2, and 3) and I patient (Subject P). Each coronal section was analyzed for the presence or absence of a j b e r tract for all regions o f interest in the right and left hemispheres. In our patient (Subject P), a lesion was present in the left inferior parietal lobule involv- ing also the posterior one-third of the left SLF. SLF = superior longitudinal fasciculus; CB = cingulum bundle; ILF/OR = com- plex o f inferior longitudinal fmciculus and optic radiation; OF = JFonto-occipital fasciculus; UF = uncinate fasciculus. The regions where the j b e r pathways are predicted to be present are indicated, as well uj the observed locations of tensor orientation and size, consistent with prediction in the right and left (black and white, respectively) hemispheres.
Makris et al: White Matter Fiber Pathways 957

tion information contained within each ROI in each slice. The results of this analysis are shown in Figure 5, where the correspondence between predictions and ob- served orientation is tabulated graphically for a set of representative association pathways. Specific hypotheses for tensors oriented in the anterior-posterior, mediolat- eral, and superior-inferior directions for fiber systems as identified in Table 1 were tested. In summary, the normal subjects demonstrated excellent correspondence between the direction information of the tensors and that provided by the Dejerine map in terms of loca- tion, size, and orientation of fiber tracts. Specifically, 96% of these hypotheses for localized orientation were consistent with the diffusion data. For the most part, the fiber pathways identified in the stroke patient were similar (see Fig 5 ) . In this patient, the lesion was lo- cated in the left hemisphere extending from the pari- etal cortex to the underlying white matter in the four posteriormost coronal sections that corresponded to the posterior one-third of the total anteroposterior length of the superior longitudinal fasciculus (SLF). In these sections the region that was anatomically defined based on the SLF did not show significant anterior-posterior directionality of the diffusion tensors. As shown in Ta- ble 2, in the coronal sections anterior to the lesion (an- terior two-thirds of the total anteroposterior length) the number of anterior-posteriorly oriented tensors in the left SLF region was reduced by a factor of 2 com- pared with the homologous region on the right (no. of
Table 2. Ratios of the Number of Voxeh, Between the Two Sides o f the Brain for the Cingulum Bundle and the Anterior Two- Thirds o f the Superior Longitudinal Fasciculus in Three Normal Subjects ( I , 2, and 3) and One Patient
Average Number of Pixels per
Right/ Coronal Section Left
White Matter Pathway Ratio Right Left
Superior longitudinal fasciculus (anterior two-thirds)
Subject 1 Subject 2 Subject 3
Mean of Subjects 1, 2, and 3 Patient 1
Cingulum bundle (entire anteroposterior
length) Subject 1 Subject 2 Subject 3
Mean of Subjects 1, 2, and 3 Patient 1
0.89 23 1.42 17 1.01 21 1.03 21 2.09 14
0.97 14 1.146 15 1.129 16 1.076 15 1.112 12
26 12 21 20 7
14 13 14 14 10
For thr above-mentioned fiber tracts, thew is also reported the av- erage number of pixels per coronal section for both sides of the brain.
voxels, right SLFIno. of voxels, left SLF = 2.09). This is contrasted to the 3 normal subjects in which the number of anterior-posteriorly oriented tensors in the SLF regions of the two sides were nearly symmetric (no. of voxels, right SLFIno. of voxels, left SLF = 1.03 for the anterior two-thirds of the SLF). We also mea- sured the number of anterior-posteriorly oriented ten- sors in the ROI defined by the cingulum bundle (CB) as a control pathway, and observed that they were nearly symmetric in the 3 normal subjects as well as in the patient (no. of voxels, right CB/no. of voxels, left CB = 1.08 and 1.11 for the normal subjects and pa- tient, respectively). It was assumed that this bundle would have remained intact in the patient, because the cortical areas related to the CB were not affected pri- marily by the lesion.
Discussion Information regarding the white matter pathways in the human brain has been based on methods that rely on postmortem material. The description of human anatomical connectivity at a level of peripheral or distal terminations is not possible at this time, because there is no technique capable of addressing this issue satis- factorily. The closest inferences at this level of descrip- tion are obtained from white matter degeneration stud- ies of brains with specific neurological damage. These studies lack specificity due to the relatively large size of the lesions and most of them deal with corticosubcor- tical connections [ 16, 171. In experimental animals, one can delineate the peripheral terminations of the cortical pathways with certainty. The knowledge of cortical association pathways derived from nonhuman primates during the past several years has provided sig- nificant understanding of the basic principles that un- derlie the organization of the brain (eg, see References 2, 18, and 19).
The study of anatomical connectivity of the human brain aims at clarifying the pathways by which differ- ent cortical and/or subcortical structures are intercon- nected. Among the different white matter pathway sys- tems, namely, the commissural, the corticosubcortical (projection), and the corticocortical (association), dam- age to the last of these is mainly responsible for many disturbances of higher brain functions such as aphasias, apraxias, and agnosias [20]. The main pathways of the corticocortical system are the SLF, the CB, the inferior longitudinal fasciculus, the occipitofrontal fasciculus, the uncinate fasciculus, and the extreme capsule. Dis- connections of these pathways as well as of the com- missural system, in particular the corpus callosum, have provided insight into lateralization of function and ce- rebral dominance in the human brain (eg, see Refer- ences 20-25). It should be pointed out that in a num- ber of neurological disorders the pathways are involved at the level of their stems. which makes their in vivo
958 Annals of Neurology Vol 42 No 6 December 1997

a
b
Right Left Right Left
Fig 6 Summary of radiological findings for patient. (a) Schematic representation of the location o f the lesion related to the topo- graphically based cortical parcellation system o f Rademacher and colledgztes [34/. The lesion intersects the supramarginal (SGa and SGp) and angular (AG) gyri. (6) A n axial T,-weighted magnetic resonance imaging scan acquired on day 12 aJter stroke. A zone o f increased signal intensig in the inferior parietal lobule is noted. (c and d) Sagittal and axial 7’1-weighted images acquired at the time of the difision imaging, 4.5 years after the onset of stroke. The horimntal line in (c) indicates the level of the axial im- age shown in (d); in a similar manner, the vertical line in (d) indicates the level o f the sagittal image in (c).
morphometry important for the understanding of the physiological, behavioral, and clinical correlates of their anatomy.
It was hypothesized that by using DWI we would be able to identify the stems of the major association pathways of the human brain in vivo. The high degree of correspondence between the tensor orientation and that predicted by orientations in the Dejerine human map supports the assertion that this orientation infor- mation corresponds to the actual fiber pathways. These observations, taken together, also validate the DWI
technique in terms of its utility for characterizing dis- crete fiber pathways. This method presupposes that most of a pathway runs in the compact portions of the bundle. The exact relationship between the compact portion of a fiber pathway and the trajectories of indi- vidual fibers themselves is not known in detail for hu- mans. However, primate data (eg, see References 26 and 27), and the limited amount of histological and tracer data in humans, strongly support the assertion that fibers of neighboring origin and termination fol- low a common course. The degree to which diffusion
Makris et al: White Matter Fiber Pathways 959

imaging itself will permit the clarification of specific targets in the peripheral terminations of a pathway re- mains unclear. Increased spatial resolution is expected to enhance the observations of pathways toward their target zones. Even if specific targets are not identified by diffusion imaging, as anatomical and tracer studies in nonhuman primates accumulate knowledge regard- ing these derails for each specific fiber pathway, addi- tional inferences can be drawn from alterations in the quantitative topography of the compact portion ob- served in humans.
The SLF, the CB, and the inferior longitudinal fas- ciculus in combination with the optic radiation are readily observed in this study primarily due to their size (5-10 mm in diameter), which is substantially larger than our sampling volume. In addition, these tracts present a homogeneity of orientation that is nec- essary for their detection. Of particular interest is their relative topography in relation to neighboring struc- tures. The SLF remains orthogonal to the rest of the white matter (ie, corona radiata and U fibers) for al- most its entire course above the insula and the sylvian fissure. Also, the CB courses orthogonally to the un- derlying corpus callosum and to adjacent U fibers and fibers of the corona radiata. The inferior longitudinal fasciculus in combination with the optic radiation re- main orthogonal to neighboring white matter fiber sys- tems as well. In addition, other smaller bundles, namely, the occipitofrontal fasciculus and the fornix, present optimal features of anatomical topography and orientation for visualization on the diffusion orienta- tion map. The uncinate fasciculus is less prominent, as it does not present an optimal orientation, because it arches over the limen of insula for a considerable part of its course. The relative topography of the different pathways with adjacent fiber systems, such as corona radiata and U fibers, could well explain the presence of different tensor orientations, and therefore colors, within a ROI that surround the anatomically predicted location. Anatomical variability of brains and size of pathways relative to the diffusion sampling volume are likely the primary sources of mismatch between the predicted and observed pathway features.
Use of D WI in Clinicounutornical Correlation We used this technique to study secondary degenera- tion of the SLF in 1 patient with conduction aphasia. This 65-year-old (at the onset of stroke), right-handed male patient suffered an embolic infarction of the left inferior parietal lobule, involving the parietal opercu- lum as well as the supramarginal and angular gyri. MRI comparison of lesion location and extent between the time of lesion onset and the time of diffusion im- aging revealed no significant differences (Fig 6) . The DWI pattern observed in this patient showed remark- able reduction in preferred diffusion orientation for the
left SLF, compared with the patient’s right hemisphere, as well as with the normal subjects (see Fig 3 and Ta- ble 2). This finding suggests damage to the left SLF of this patient.
The details of anatomical connectivity are critical to a number of newly emerging methods for the analysis of functional neuroimaging data. Fiber pathway dis- connections are implicated in altered cerebral metabo- lism pattern observed in stroke patients [28]. Func- tional activation studies using positron emission tomography and functional MRI contribute to the knowledge of the spatial distribution of cortical and subcortical processing elements. The details of func- tional connectivity (eg, see Reference 29), however, are influenced directly by the a priori model of anatomical connectivity being used in structural equation model- ing [30-321. Use of actual individual information re- garding white matter fiber pathway size and trajectory in vivo may permit the creation of more advanced models.
It should be pointed out that there are technical and practical limitations inherent in this technique. In this study, our sampling volume is anisotropic. Thus, the relative sensitivity is different within the coronal plane of acquisition (mediolateral, superior-inferior) com- pared with the anterior-posterior direction. A fiber tract must have a homogeneous orientation that com- prises most of the volume of the voxel for that tract and orientation to be accurately reflected in the result- ing map. When multiple tracts and orientations are present within the voxel, the net result is an average orientation. Therefore, for definitive identification, the fiber tract should be at least the size of the voxel. Dif- ferent orientation and voxel size parameters could be prescribed and optimized for studies of specific white matter ROIs. The absence of identifiable SLF in some locations in our patient can be due to either complete destruction of the tract or reduction of the tract below the size of detectability. General application of this technique will undoubtedly benefit from a more nearly isotropic acquisition.
The present results are limited to the compact parts or stems of the fiber tracts, because these portions of the bundles are larger than the diffusion sampling vol- ume. In this investigation we used a large number of averages at the cost of a relatively long imaging session to achieve a relatively high signal-to-noise ratio. Addi- tional work is required to achieve the optimum trade- off between sensitivity, resolution, and patient compli- ance relative to total imaging time.
Additional studies related to the fundamental neuro- anatomical underpinnings of this method are currently under way. First, direct correlation of the actual fiber pathways to their imaging results can be attempted in postmortem material. This type of endeavor is not without significant technical difficulties pertaining to
960 Annals of Neurology Vol 42 No 6 December 1997

altered MRI parameters of postmortem tissue prepara- tion relative to in vivo values, precise registration of imaging and histological results, and precise pathway delimitation in the histological preparation. Second, the generation of quantitative population statistics is required to determine the specific sensitivity of this method for identification of subtle abnormalities of fi- ber pathway topology. Principally, abnormalities will appear either as absence of a fiber pathway (as demon- strated in the stroke case presented here) or as a “sig- nificant” alteration of the location, orientation, or size of a particular pathway (inferred in the stroke case pre- sented here). For the former, the population data we compiled on the three normal subjects make us confi- dent in the identification of the missing portion of the SLF in the region of the lesion. In the latter type of observation, the significance of an alteration in size (as of the SLF remote from the lesion, in this case) or orientation or location i s more dependent on a quan- titative assessment of the normative population data and requires more extended subject sampling.
Conclusion The data presented demonstrate that the stems of ma- jor association pathways can be visualized and quanti- fied in vivo in the human brain. In addition, this is the first time that a color-coded tensor orientation map has been correlated with a human brain anatomical map to validate the DWI technique in terms of known fiber pathway orientation in living humans. This methodol- ogy opens up various possibilities. Anatomical connec- tivity can be studied in detail in vivo, within and across subjects, in both a qualitative and a quantitative man- ner. In addition, this method allows the characteriza- tion of damage to or disconnection of white matter pathways and could provide further diagnostic and functional insight in different neurological disorders in- volving white matter such as stroke, multiple sclerosis, amyotrophic lateral sclerosis, head injury, and spinal cord injury. Moreover, knowledge of white matter pathways derived by DWI could well contribute to the understanding of language processing and in other cog- nitive domains as well as shed light on the mechanisms of recovery of function or its failure. Future technolog- ical development will decrease imaging time as well as improve the spatial resolution and availability of this imaging capability. This may allow, more ,readily, ac- curate clarification of the white matter pathways and provide additional understanding of the organizational principles underlying the structure and function of the human brain in health and disease.
This study was supported in part by National Public Health Service grants Pol-DA09467, RO1-HL39810, RO1-NS34189, and P o l - N S2795O.
We thank Drs David Caplan, Jeremy Schmahmann, Jean Paul von Sattel, Edward Yeterian, and the anonymous reviewers for providing useful comments on the manuscript, and Terry Campbell and Mary Foley for subject scanning.
References 1.
2.
3.
4.
5.
6.
7.
8.
9
10
11
12
13.
14.
15.
16.
17.
18.
19.
20.
Dejerine J. Anatomie des centres nerveux, Vol 1. Paris: Rueff et CIE, 1895 Jones EG, Powell TPS. An anatomical study of converging sen- sory pathways within the cerebral cortex of the monkey. Brain
Pandya DN, Yeterian E. In: Peters A, Jones E, eds. Cerebral cortex. New York: Plenum Publishing, 1985 Caviness VS, Filipek PA, Kennedy DN. Magnetic resonance technology in human brain science: blueprint for a program based upon morphometry. Brain Dev 1989;ll:l-13 Le Bihan D, Breton E, Lallemand D, et a!. MR imaging of intravoxel incoherent motions: applications to diffusion and perfusion in neurologic disorders. Radioloby 1986; 161:401- 407 Turner R, Le Bihan D, Delannoy J, Pekar J . Echo-planar dif- fusion and perfusion imaging at 2.0 Tesla. In: Abstracrs of the Society of Magnetic Resonance in Medicine, 7th annual meet- ing. Berkeley, CA, 1989 Moseley ME, Cohen Y, Kucharczyk J, et al. Diffusion-weighted MR imaging of anisotropic water diffusion in cat central ner- vous system. Radiology 1990;176:439-446 Moseley ME, Kucharczyk J, Mintorovitch J, et al. Diffusion- weighted MR imaging of acute stroke: correlation with T2- weighted and magnetic susceptibiliry-enhanced MR imaging in cats. AJNR 1990;11:423-429 Douek P, Turner R, Pekar J, et al. MR color mapping of my- elin fiber orientation. J Comput Assist Tomogr 1991;15:923- 929 Chien D, Buxron RB, Kwong KK, et al. MR diffusion imaging of the human brain. J Comput Assist Tomogr 1990;14:514- 520 Basser PJ, Mattiello J, Le Bihan D. Estimation of the effective self-diffusion tensor from the NMR spin echo. J Magn Reson [B] 1994; 103:247-254 Garrido L, Wedeen VJ, Kwong KK, er al. Anisotropy of water diffusion in the myocardium of the rat. Circ Res 1994;74:789- 793 Davis TL, Wedeen VJ, Weisskof RM, et al. White matter tract visualization by echo-planar MRI. In: Abstracts of the Society of Magnetic Resonance in Medicine, 12th annual meeting. New York, 1993 Pierpaoli CJ, Basser PJ, Barnett A, Di Chiro G. Diffusion ten- sor MR imaging of the human brain. Radiology 1996;201:637- 648 Pierpaoli C, Basser PJ. Toward a quantitative assessment of dif- fusion anisotropy. Magn Reson Med 1996;36:893-906 Dejerine J. Anatomie des centres nerveux, Vol 2. Paris: Rueff et CTE, 1895 Clarke S, Miklossy J. Occipital cortex in man: organization of callosal connections, related myelo- and cytoarchitecture, and putative boundaries of functional visual areas. J Comp Neurol 1990;298: 188-214 Pandya DN, Kuypers HGJM. Cortico-cortical connections in the rhesus monkey. Brain Res 1969;13:13-36 Pandya DN, Seltzer B, Barbas H. Input-output organization of the primate cerebral cottex. Comp Primate Biol 1988;4:39-80 Geschwind N. Disconnexion syndromes in animals and man. In: MWW, Cohen RS, eds. Selected papers on language and the brain. Dordrecht, The Netherlands: D Reidel Publishing, 1974: 105-236
1970;93:793-820
Makris et al: White Matter Fiber Pathways 961

21. Akelaitis AJ, Wilford AR, Yorke Herren K, Van Wagenen WP. Studies on the corpus callosum: 111. A contribution to the study of dyspraxia and apraxia following partial and complete section of the corpus callosum. Arch Neurol Psychiatry 1942;47:971- 1007
22. Sperry RW. Cerebral organization and behavior. Science 1961;
23. Geschwind N, Kaplan E. A human cerebral disconnection syn- drome. Neurology 1962;12:675-685
24. Gazzaniga MS. The bisected brain. New York: Appleton- Centuty-Crofts, 1970
25. Geschwind N, Galaburda A. Cerebral lateralization. Cam- bridge, MA: MIT Press, 1987
26. Petrides M, Pandya DN. Association fiber pathways to the frontal cortex from the superior temporal region in the rhesus monkey. J Comp Neurol 1988;273:52-66
27. Petrides M, I'andya DN. Projections to the frontal cortex from the posterior parietal region in the rhesus monkey. J Comp Neurol 1984;228:105-116
133:1749-1757
28. Kosslyn SM, Daly PF, McPeek RM, et al. Using locations to store shape: an indirect effect of a lesion. Ceteb Cortex 1993;
29. Friston K. Functional and effective connectivity in neuroimaging: a synthesis. Hum Brain Map 1994;2:56-78
30. McIntosh AR, Gonzalez-Lima F. Structural equation modelling and its application to network analysis in functional brain im- aging. Hum Brain Map 1994;2:2-24
31. Gonzalez-Lima F, McIntosh AR. Neural network interactions related to auditory learning analyzed with structural equation modelling. Hum Brain Map 1994;2:23-44
32. Honvitz B. Data analysis paradigms for metabolic-flow data: combining neural modeling and functional neuroimaging. Hum Brain Map 1994;2:112-122
33. Talairach J, 'Tournoux P. Co-planar stereotaxic atlas of the human brain. New York: Thieme Medical Publishers, 1988
34. Rademacher J, Galaburda A, Kennedy D, et al. Human cerebral cortex: localization, parcellation and morphometry with mag- netic resonance imaging. J Cogn Neurosci 1992;4:352-374
3:567-582
962 Annals of Neurology Vol 42 No 6 December 1997


















