
Morphological development of testes in ostrich (Struthio camelus) embryo
11
ORIGINAL ARTICLE Morphological development of testes in ostrich (Struthio camelus) embryo Belal Hassanzadeh • Abolghasem Nabipour • Morteza Behnam Rassouli • Hesam Dehghani Received: 27 June 2013 / Accepted: 26 September 2013 / Published online: 15 October 2013 Ó Japanese Association of Anatomists 2013 Abstract Although the histological structure of ostrich testis has been studied, very little information is currently available on the embryonic development of this organ. The aim of this study was to determine the sequence of the histological changes in diverse components of the testis in ostrich embryo from embryonic day (E) 20 to E42. The main findings were categorized into four histological fea- tures, i.e., development of sex cords, interstitial tissue and rete ducts, and the appearance of defective septa. While the lumen of sex cords, tunica albuginea, capsular rete ducts and Leydig cell precursors appeared at E26, the filum- shaped defective septa were visible at E36. The emersion of the lumen in the primary sex cords and formation of cap- sular rete ducts in the ostrich embryo is considerably dif- ferent from that in other birds. However, tunica albuginea and Leydig cell precursors appeared in a similar pattern to those of other birds. An interesting observation was that the primordial germ cell (PGC)-like cells were completely distinct, while the capsular rete ducts were formed by trapping of some Sertoli cell aggregations in the tunica albuginea. This suggests that similar to the primary sex cords, the capsular rete ducts may originate from the Sertoli cell aggregations which had corralled some PGCs. Stereological estimations in the ostrich embryo testis showed the major proportion of testis is occupied by the seminiferous tubules, which is unlike the fowl embryo testis. Keywords Ostrich embryo Primordial germ cells Rete ducts Seminiferous tubules Tunica albuginea Introduction General stages of the testicular development have already been determined in many avian species, and testes in avian embryos are known to arise from the genital ridge at both sides of the dorsal mesentery (Fujimoto et al. 1976; Van Krey 1990). The genital ridges then develop to undiffer- entiated gonads and subsequently to morphologically identical testes (Smith 2007). During this process, Sertoli cell progenitors and primordial germ cells (PGCs) emerge in the primary sex cords where they become wavy and anastomose to each other, forming the reticular features seen in histological sections (Romanoff 1960; Morrish and Sinclair 2002; Gonzalez-Moran and Soria-Castro 2010a). In the fowl embryo, the seminiferous cords lack a lumen by embryonic day (E) 13, but the central axis produces a slit and the lumen appears by E20 (approximate time of hatching) (Gonzalez-Moran and Soria-Castro 2010a). Studies on the morphology and development of testis are mostly performed on mammals and some known birds. To date, the morphological development of testis has been well studied in some avian species, such as domestic fowl and quail (Gonzalez-Moran 1997; Gonzalez-Moran and Soria-Castro 2010a, b; Chang et al. 2012), but pre-hatching studies have been done only on chick and quail testis, with the primary focus on early embryological events in the B. Hassanzadeh A. Nabipour (&) H. Dehghani Department of Basic Science, Faculty of Veterinary Medicine, Ferdowsi University of Mashhad, Mashhad, Iran e-mail: [email protected]; [email protected] M. Behnam Rassouli Department of Biology, Faculty of Science, Ferdowsi University of Mashhad, Mashhad, Iran H. Dehghani Embryonic and Stem Cell Biology and Biotechnology Research Group, Research Institute of Biotechnology, Ferdowsi University of Mashhad, Mashhad, Iran 123 Anat Sci Int (2014) 89:129–139 DOI 10.1007/s12565-013-0207-9
Transcript of Morphological development of testes in ostrich (Struthio camelus) embryo

ORIGINAL ARTICLE
Morphological development of testes in ostrich (Struthio camelus)embryo
Belal Hassanzadeh • Abolghasem Nabipour •
Morteza Behnam Rassouli • Hesam Dehghani
Received: 27 June 2013 / Accepted: 26 September 2013 / Published online: 15 October 2013
� Japanese Association of Anatomists 2013
Abstract Although the histological structure of ostrich
testis has been studied, very little information is currently
available on the embryonic development of this organ. The
aim of this study was to determine the sequence of the
histological changes in diverse components of the testis in
ostrich embryo from embryonic day (E) 20 to E42. The
main findings were categorized into four histological fea-
tures, i.e., development of sex cords, interstitial tissue and
rete ducts, and the appearance of defective septa. While the
lumen of sex cords, tunica albuginea, capsular rete ducts
and Leydig cell precursors appeared at E26, the filum-
shaped defective septa were visible at E36. The emersion of
the lumen in the primary sex cords and formation of cap-
sular rete ducts in the ostrich embryo is considerably dif-
ferent from that in other birds. However, tunica albuginea
and Leydig cell precursors appeared in a similar pattern to
those of other birds. An interesting observation was that the
primordial germ cell (PGC)-like cells were completely
distinct, while the capsular rete ducts were formed by
trapping of some Sertoli cell aggregations in the tunica
albuginea. This suggests that similar to the primary sex
cords, the capsular rete ducts may originate from the Sertoli
cell aggregations which had corralled some PGCs.
Stereological estimations in the ostrich embryo testis
showed the major proportion of testis is occupied by the
seminiferous tubules, which is unlike the fowl embryo
testis.
Keywords Ostrich embryo � Primordial germ cells �Rete ducts � Seminiferous tubules � Tunica albuginea
Introduction
General stages of the testicular development have already
been determined in many avian species, and testes in avian
embryos are known to arise from the genital ridge at both
sides of the dorsal mesentery (Fujimoto et al. 1976; Van
Krey 1990). The genital ridges then develop to undiffer-
entiated gonads and subsequently to morphologically
identical testes (Smith 2007). During this process, Sertoli
cell progenitors and primordial germ cells (PGCs) emerge
in the primary sex cords where they become wavy and
anastomose to each other, forming the reticular features
seen in histological sections (Romanoff 1960; Morrish and
Sinclair 2002; Gonzalez-Moran and Soria-Castro 2010a).
In the fowl embryo, the seminiferous cords lack a lumen by
embryonic day (E) 13, but the central axis produces a slit
and the lumen appears by E20 (approximate time of
hatching) (Gonzalez-Moran and Soria-Castro 2010a).
Studies on the morphology and development of testis are
mostly performed on mammals and some known birds. To
date, the morphological development of testis has been
well studied in some avian species, such as domestic fowl
and quail (Gonzalez-Moran 1997; Gonzalez-Moran and
Soria-Castro 2010a, b; Chang et al. 2012), but pre-hatching
studies have been done only on chick and quail testis, with
the primary focus on early embryological events in the
B. Hassanzadeh � A. Nabipour (&) � H. Dehghani
Department of Basic Science, Faculty of Veterinary Medicine,
Ferdowsi University of Mashhad, Mashhad, Iran
e-mail: [email protected]; [email protected]
M. Behnam Rassouli
Department of Biology, Faculty of Science, Ferdowsi University
of Mashhad, Mashhad, Iran
H. Dehghani
Embryonic and Stem Cell Biology and Biotechnology Research
Group, Research Institute of Biotechnology, Ferdowsi
University of Mashhad, Mashhad, Iran
123
Anat Sci Int (2014) 89:129–139
DOI 10.1007/s12565-013-0207-9

testis (Zaccanti et al. 1990; Sekido and Lovell-Badge 2007;
Tagami et al. 2007) and early or late testicular development
(Csaba et al. 1980; Shahin and Torok 1982; Gonzalez-
Moran and Soria-Castro 2010a, b). There have been few
pre-hatching developmental studies in ostrich and other
ratite birds, and these have focused mainly on embryonic
timing or hatchability of the eggs (Gefen and Ar 2001;
Malecki et al. 2005; Nagai et al. 2011). We have recently
published a morphological study on the juvenile ostrich
testis (Hassanzadeh et al. 2013), and in a few other studies
the authors have explained the morphological features of
adult ostrich testis (Soley 1992; Aire and Soley 2003;
Ozegbe et al. 2006, 2008, 2010; Lan et al. 2007; Zhang
et al. 2011); however, developmental studies of ostrich
testis are still scant and limited to the post-hatching period
(Madekurozwa et al. 2002; Wei et al. 2011). Budras and
Meier (1981) have performed the only developmental
study on the testicular excurrent ducts in ratite birds. This
study covers both the pre- and post-hatching periods, and
the testis in the juvenile and adult ostrich was compared
with that in rhea and emu.
In the study reported here, we studied in detail the mor-
phological development of ostrich testis, including the for-
mation of sex cords, testicular capsule and capsular rete ducts.
We also evaluated the morphometrical changes in the testis
and its tubular compartments quantitatively using stereolo-
gical techniques. Our results provide novel information on the
developmental characteristics of the ostrich testis.
Materials and methods
Animals and tissue samples
A total of 50 fertile ostrich eggs obtained from commercial
farms were incubated at 36–37 �C and 25 ± 2 % relative
humidity with tilting to 90� at 4-h intervals for 20, 26, 36 and
42 days. Due to the coincidence of E42 with hatching day
and in order to minimize the number of animals sacrificed,
hatched chicks were sexed using the method of Malago et al.
(2002). After decapitation of male chicks and embryos and
the drainage of blood, the testicles were anatomically ana-
lyzed, photographed and separated from the body and placed
in Bouin’s solution. Three samples of testes at each of E26,
36 and 42 (size approx. 1 mm3) were taken and placed in 2 %
glutaraldehyde in 1 M cacodylate buffer for embedding in
resin. All procedures were approved by the Ethics Scientific
Committee of the Ferdowsi University of Mashhad.
Morphometry
Stereological techniques were employed to determine the
amount of testicular volume occupied by seminiferous
tubules (volume fraction or volume density of seminiferous
tubules in the testis). After fixation, samples (whole testes)
were dehydrated in increasing degrees of ethanol, cleared in
xylene and embedded in paraffin. Due to the existence of a
directional asymmetry (Moller 1994), the stereological
experiments were done only on the left testicles. Tissue
shrinkage was calculated by measuring testicle diameter just
before fixation and embedding in paraffin. Paraffin-embed-
ded samples were cut into 5-lm-thick serial sections and
sampled (at least 11 sections per sample) according to the
unbiased systematic random sampling (SRS) method. After
staining of the sections with Periodic acid–Schiff (PAS)
stain, several fields from large sections were selected and
imaged according to SRS method. The images were placed
under a stereology grid (with 300 intersection points without
any direction), and when the points were positioned on the
whole section, the reference area and seminiferous tubules
were analyzed and counted. The total volume of the testis
(Tes.V) was estimated according to the Cavalieri’s principle.
The volume fraction of seminiferous tubules in each testis
(Vv) was calculated using Eqs. 1, 2 and 3 based on the De-
lesse principle (Delesse 1848; Howard and Reed 2005):
Vv ¼ �a�A
ð1Þ
�a ¼ a1 þ a2 þ � � � þ ak
kð2Þ
�A ¼ A1 þ A2 þ � � � þ Ak
kð3Þ
Where a, A and k are the area occupied by seminiferous
tubules, the reference area and the number of images,
respectively. The volume of testis was multiplied by Vv to
estimate the total volume of seminiferous tubules (S.T.V).
The results were analyzed by one-way analysis of variance,
and the different groups were determined by Tukey’s
multiple comparison test. Differences among mean Vv of
the seminiferous tubules were considered to be statistically
significant at P \ 0.05.
Histology
The glutaraldehyde-fixed samples were re-fixed in 1 %
osmium tetroxide in 1 M cacodylate buffer. These samples
were dehydrated by the progressive lowering temperature
(PLT) method, embedded in epoxy resin (TAAB Labora-
tories Equipment Ltd, UK) and cut in 1-lm-thick sections.
The sections were stained with toluidine blue in order to
identify the lumen of the sex tubules and the lipid granules
of the Leydig cells (Kuo 2007). Some of the 5-lm-thick
sections from different parts of the right and left testes,
near two poles and the equatorial border, were selected for
staining with PAS, hematoxylin and eosin (H&E), Mas-
son’s trichrome (MT) and Alcian blue (AB) to clarify the
130 B. Hassanzadeh et al.
123

details of seminiferous tubules, interstitial tissue and the
testicular capsule. PGCs were recognized based on their
morphological characteristics, which have been described
previously (Ginsburg and Eyal-Giladi 1986; Ishiguru et al.
2009; Gonzalez-Moran and Soria-Castro 2010a; Wei et al.
2011). Images were collected on a digital camera (model
SX210 IS; Canon Inc., Tokyo, Japan) and using a light
microscope (models BX51 and 60; Olympus, Tokyo,
Japan) equipped with a digital camera (model DP12;
Olympus). The acquired images were prepared by Image J
(National Institutes of Health, Bethesda, MD), Paint
(Microsoft, Redwood, WA), and Snagit Editor (TechSmith
Corp., Okemos, MI) software.
Results
Gross anatomical changes
In the 20-day-old embryo, testes having a dirty-white color
were located on the ventral surface of mesonephros kid-
neys. The testes extended from the cranial edge to the mid-
portion of kidneys, where they inclined toward the median
plane of body and were located close to each other. At this
stage, the testes had a curved tortuous or elongated ovoid
shape and were directly attached to the kidneys (Fig. 1a).
In the 26-day-old embryo, the testes had become brighter
and were clearly distinguishable from the kidneys in tex-
ture and color. Their dimensions had increased and the
connection to the kidney had become narrower and distinct.
Their shape changed to elongated ovoid and became reg-
ular relative to the previous stage (Fig. 1b). In the 36-day-
old embryo, the testes had become larger and due to the
regression of mesonephros and growth of metanephros
their location changed caudally toward the metanephros
middle lobe (Fig. 1c). In the 42-day-old embryo or newly
hatched ostrich, the testes extended from the mid-portion of
the cranial lobe of the metanephros to its caudal edge
(Fig. 1d). In all cases the longitudinal axis of testes made
an acute angle with the sagittal axis of the body which was
greater on the right side. The shapes of the testicular
transverse sections changed from elongated in the 20-day-
old embryo to bean-shaped in the 26-day-old embryo, and
then to ovoid in the 36-day old embryo and subsequently to
a circular shape in the 42-day-old embryo.
Morphometrical changes
Stereological techniques were used for evaluating mor-
phological changes in the testis, including the Tes.V, Vv of
the seminiferous tubules in the testis, and S.T.V. in each
testis. Evaluations showed that body weight (B.W.)
increased from E20 to E36 and thereafter slightly
decreased up to the hatching day (Fig. 2a). The Tes.V. and
S.T.V. increased linearly up to the hatching day (Fig. 2a).
In general, the ratio of Tes.V. and S.T.V. to the BW
decreased from E20 to E42 (Fig. 2b). The estimated Vv
decreased from E20 to E26 but increased slowly up to E42
(Fig. 2c).
Histological changes
The 20-day-old embryo
Sertoli cell precursors surrounding the PGCs formed Ser-
toli cell aggregations which extended toward each other
and in some cases came into contact and anastomosed
(Fig. 3). PGCs were distinguishable by their relatively
large, pale, spherically shaped nucleus (Fig. 3a, c). In MT-
and AB-stained sections, the nucleus of mitotic PGCs
appeared as a dark floccus of chromatin (Fig. 3b, d). Sertoli
cell precursors had elongated ovoid or triangular nuclei
which were smaller and more heterochromatic than the
PGC nuclei, whereas when treated with AB stain the nuclei
of the former stained pale relative to those of the PGCs
(Fig. 3). The Sertoli cell precursors had a polar epithelial
Fig. 1 Ventral view of ostrich testes in different embryonic days (E).
a Testes (dashed lines) at E20 are located in the ventral surface of
mesonephros kidneys (Ms). Note that they are inclined toward the
median plane of the body and are located close to each other. b Testes
at E26 are attached to the mesonephros. c Morphology and
topography of the testes at E36 relative to the mesonephros and
metanephros kidneys. d Testes at E42 in the vicinity of metanephros
cranial lobe (Mtc). Note the change in color, shape and location of
testes during in ovo development. Mt Metanephros kidney, R right,
L left. Scale bar 3 mm
Embryonic development of ostrich testis 131
123

cell-like morphology with numerous secretary granules at
the apical pole and the nucleus at the basal pole. The apical
pole was more ramshackle than the basal pole which was
supported by a delicate basement membrane (Fig. 3c).
Interstitial tissue contained only PAS-positive fibers that
appeared pink or purple in color, while Leydig cell pre-
cursors were undistinguishable from mesenchymal cells
(Fig. 3). Testicular capsules consisted of a single layer of
cuboidal cells as a covering epithelium and its underlying
blood vessels (Fig. 3b). Attachment to the kidney was
devoid of covering epithelium, where a few rete ducts
paved by flattened cells were present (Fig. 3a).
The 26-day-old embryo
Elongating and anastomosing cords superseded Sertoli cell
aggregations causing a net-like structure (Fig. 4b, c) to
develop. The number of PGCs was clearly increased rela-
tive to E20, but their morphology was the same except for
their completely eccentric location in the cords (Fig. 4a, b).
Fig. 2 Morphometrical analysis. a Logarithmic plot to show changes
in body weight (B.W.), left testis volume (Tes.V) and total volume of
seminiferous tubules (S.T.V) from E20 to E42. Note that B.W.
increased with age until E36, following which it slightly decreased
due to weight loss during the hatch, while Tes.V. and S.T.V.
constantly increased. b Ratio of S.T.V. and Tes.V. to B.W. from E20
to E42. This logarithmic plot shows that these ratios follow a pattern
opposite to that for B.W. c Estimated volume fraction of seminiferous
tubules in testis (Vv) decreased from E20 to E26, following which it
increased. Data are shown as the mean ± standard deviation (SD)
Fig. 3 Sertoli cell aggregations (some of them defined by broken
lines) in the 20-day-old ostrich embryo. a Illustration of the rete ducts
(R) paved by flattened cells at the attachment site of testis and kidney.
Note the differences between the rete ducts and blood vessels (V).
Hematoxylin and eosin (H&E) staining. b Sertoli cell aggregations
came into contact and anastomosed to each other. Note the
anastomosing site (asterisks), testicular covering epithelium (Ep)
and its underlying blood vessels (V). Masson’s trichrome (MT)
staining. c The primordial germ cell (PGCs, P) with a large, pale,
spherical nucleus are located between the Sertoli cells (Sr). The color
of interstitial tissue (I) is because of its Periodic acid–Schiff (PAS)-
positive fibers. Arrows Basal pole of Sertoli cells closed to the
basement membrane, arrowheads apical pole. PAS staining. d Inter-
stitial tissue fibers were distinctly stained by Alcian blue (AB) while
the nuclei of the PGCs stained as dark flocci. Ms mesonephric tubule,
Bg blood globule. Scale bar 30 lm
132 B. Hassanzadeh et al.
123
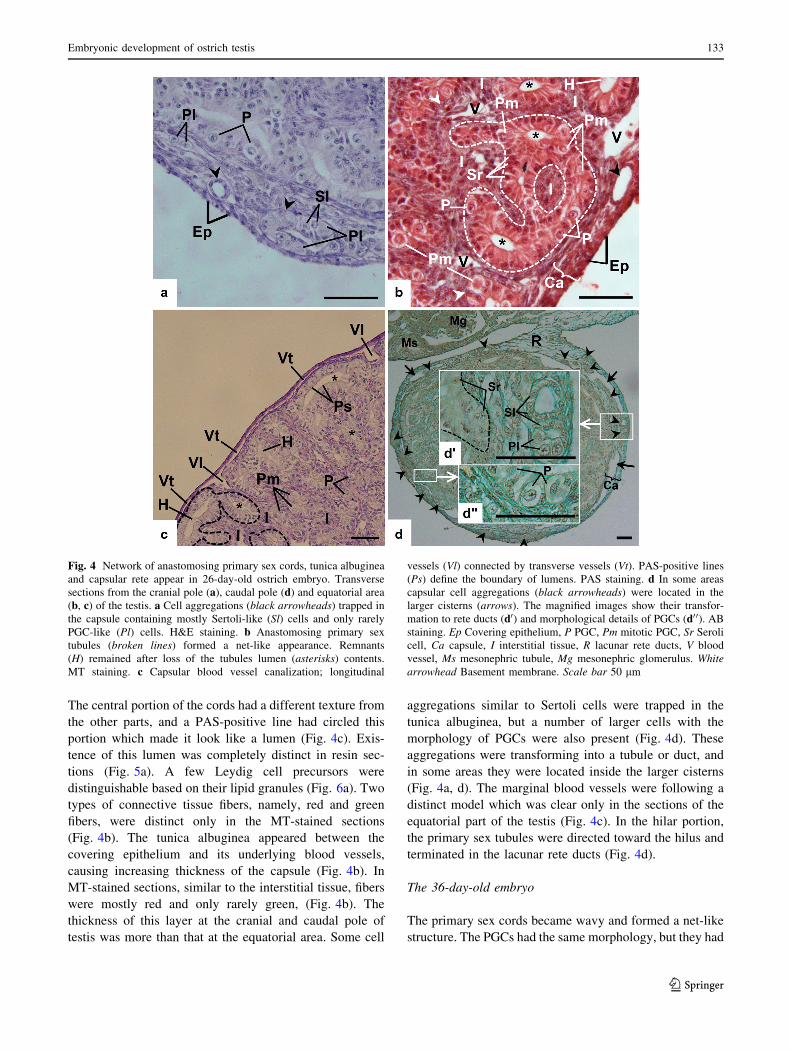
The central portion of the cords had a different texture from
the other parts, and a PAS-positive line had circled this
portion which made it look like a lumen (Fig. 4c). Exis-
tence of this lumen was completely distinct in resin sec-
tions (Fig. 5a). A few Leydig cell precursors were
distinguishable based on their lipid granules (Fig. 6a). Two
types of connective tissue fibers, namely, red and green
fibers, were distinct only in the MT-stained sections
(Fig. 4b). The tunica albuginea appeared between the
covering epithelium and its underlying blood vessels,
causing increasing thickness of the capsule (Fig. 4b). In
MT-stained sections, similar to the interstitial tissue, fibers
were mostly red and only rarely green, (Fig. 4b). The
thickness of this layer at the cranial and caudal pole of
testis was more than that at the equatorial area. Some cell
aggregations similar to Sertoli cells were trapped in the
tunica albuginea, but a number of larger cells with the
morphology of PGCs were also present (Fig. 4d). These
aggregations were transforming into a tubule or duct, and
in some areas they were located inside the larger cisterns
(Fig. 4a, d). The marginal blood vessels were following a
distinct model which was clear only in the sections of the
equatorial part of the testis (Fig. 4c). In the hilar portion,
the primary sex tubules were directed toward the hilus and
terminated in the lacunar rete ducts (Fig. 4d).
The 36-day-old embryo
The primary sex cords became wavy and formed a net-like
structure. The PGCs had the same morphology, but they had
Fig. 4 Network of anastomosing primary sex cords, tunica albuginea
and capsular rete appear in 26-day-old ostrich embryo. Transverse
sections from the cranial pole (a), caudal pole (d) and equatorial area
(b, c) of the testis. a Cell aggregations (black arrowheads) trapped in
the capsule containing mostly Sertoli-like (Sl) cells and only rarely
PGC-like (Pl) cells. H&E staining. b Anastomosing primary sex
tubules (broken lines) formed a net-like appearance. Remnants
(H) remained after loss of the tubules lumen (asterisks) contents.
MT staining. c Capsular blood vessel canalization; longitudinal
vessels (Vl) connected by transverse vessels (Vt). PAS-positive lines
(Ps) define the boundary of lumens. PAS staining. d In some areas
capsular cell aggregations (black arrowheads) were located in the
larger cisterns (arrows). The magnified images show their transfor-
mation to rete ducts (d0) and morphological details of PGCs (d0 0). AB
staining. Ep Covering epithelium, P PGC, Pm mitotic PGC, Sr Seroli
cell, Ca capsule, I interstitial tissue, R lacunar rete ducts, V blood
vessel, Ms mesonephric tubule, Mg mesonephric glomerulus. White
arrowhead Basement membrane. Scale bar 50 lm
Embryonic development of ostrich testis 133
123

increased in number, and most of them were mitotic. Leydig
cell precursors were widespread throughout the interstitial
tissue. The polymorphic nucleus of these cells, as well as
the presence of a few lipid granules, distinguished them
from the fibroblasts and blood cells (Fig. 6b). The testicular
capsule, which completely coated the testis, had acquired a
Fig. 5 Appearance of lumen in the primary sex cords. Transverse
semithin sections of the ostrich testis stained by toluidine blue
indicate the presence of a lumen (arrows) in the primary sex cords on
E26 (a), E36 (b) and E42 (c). Note that the apical pole of Sertoli cells
contains numerous secretory granules and that the basal pole usually
contains the nucleus. d Circling fibroblasts (F), around the seminif-
erous cords, which are presumably the progenitors of the myoid cells.
Ca Testicular capsule, I interstitial tissue. Asterisks PGCs, broken
lines boundary of sex cords. Scale bar 10 lm
Fig. 6 Appearance of Leydig cell lipid granules. Transverse semithin
sections of ostrich testis stained by toluidine blue shows the presence
of lipid granules (arrowheads) on E26 (a), E36 (b) and E42 (c).
Periphery of sex cords (S) are marked by broken lines. Bl Blood cells,
I interstitial tissue. Asterisks PGCs. Scale bar 10 lm
134 B. Hassanzadeh et al.
123

tree-partite structure that included the tunica serosa, tunica
albuginea and tunica vasculosa (Fig. 7a, b). Extending from
the tunica albuginea, some meandering rays penetrated the
interstitial tissue, while embedded testicular internal arter-
ies had the typical structure of a muscular artery (Fig. 7b0,d00). Lacunar rete ducts in the hilar portion were connected
to the capsular rete system which extended up to the free
surface of the testis (Fig. 7a–d). The capsular rete system
consisted of large cisterns, duct-like structures and trapped
cell aggregations which were obviously transforming into
the duct-like structures (Fig. 7a, c).
The 42-day-old embryo
In the hatching chick, the primary sex cords became wavier
and formed a reticular structure while the confines of their
lumen became more distinct (Fig. 5c). It appeared that the
number of PGCs had not increased relative to E36. The
tree-partite structure of the capsule had become more dis-
tinct and completely coated the testis (Fig. 8a). The cap-
sular rete ducts had expanded toward the free surface of the
testis (Fig. 8a, b) and the thickness of the capsule was
increased due to intensification of fibers in the tunica
Fig. 7 Histology of testis in the 36-day-old ostrich embryo. Trans-
verse sections from the cranial pole (a, c) and equatorial area (b, d) of
ostrich testis on E42 stained by H&E (a), MT (b), PAS (c) and AB
(d). a Lacunar rete ducts (R) were present in the testicular hilus, while
capsular rete ducts (arrowheads) inside the large cisterns (arrows)
extended up to the testicular free surface. Magnified parts (a0 and a0 0)illustrate details of the capsular rete cuboidal epithelium which is
different from a blood vessel endothelium. b Meandering rays from
the tunica albuginea (Ta), defective septa (Ds), penetrate the
interstitial tissue (I). In the central part of the testis some arteries
can be seen to be embedded inside the defective septa (b0).c Distribution of capsular rete (arrowheads) and large cisterns
(arrows) in the testicular free surface and their histological details (c0
and c0 0). Note the presence of PGC-like (Pl) and Sertoli-like (Sl) cells
in the structure of capsular rete ducts, and the cuboidal epithelium
(Co) of the large cisterns. d Histological details of capsular cell
aggregation (arrowheads) which still had not acquired a duct-like
shape (d0). Note the typical structure of a muscular artery (d00)including the tunica intima (In), tunica media (Md) and a fibrous
tunica adventitia (Ad) which merged with the fibrous texture of their
container defective septum. Bg Blood globule, Ca testicular capsule,
P PGC, Pm mitotic PGC, S seminiferous tubule, Sr Sertoli cell, Ts
tunica serosa, Tv tunica vasculosa, V blood vessel in tunica vasculosa.
Asterisk Lumen of seminiferous tubules, arrows large cisterns,
arrowheads capsular rete ducts. Scale bar 50 lm
Embryonic development of ostrich testis 135
123
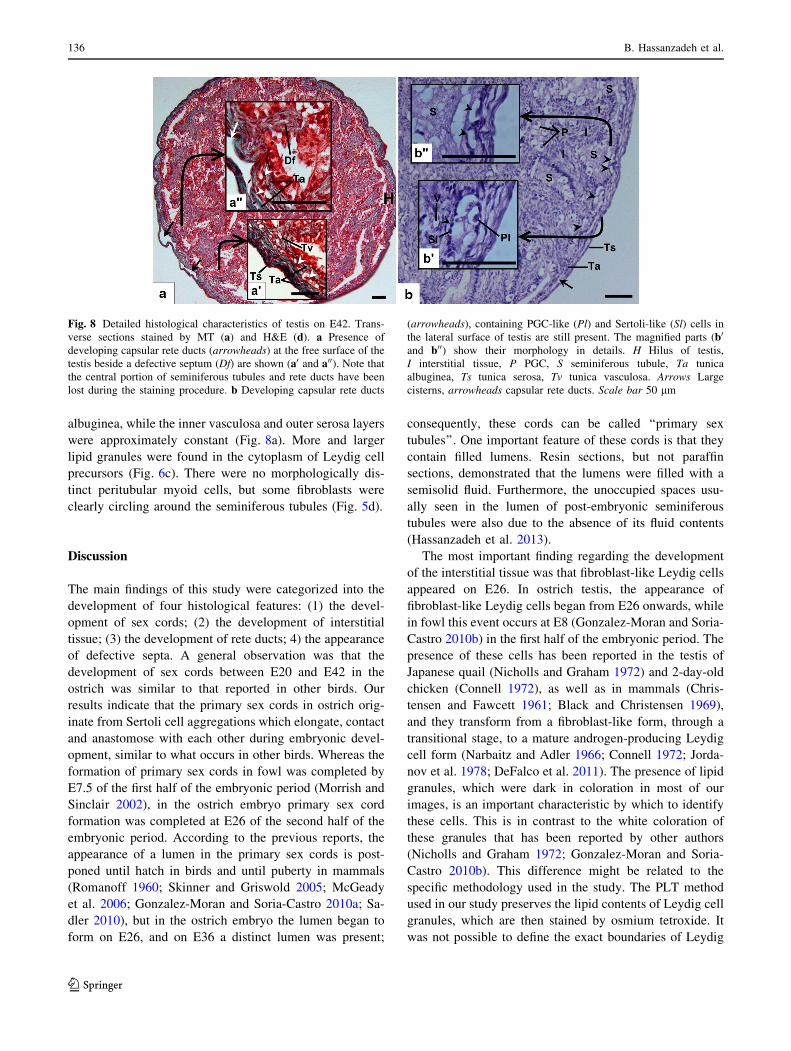
albuginea, while the inner vasculosa and outer serosa layers
were approximately constant (Fig. 8a). More and larger
lipid granules were found in the cytoplasm of Leydig cell
precursors (Fig. 6c). There were no morphologically dis-
tinct peritubular myoid cells, but some fibroblasts were
clearly circling around the seminiferous tubules (Fig. 5d).
Discussion
The main findings of this study were categorized into the
development of four histological features: (1) the devel-
opment of sex cords; (2) the development of interstitial
tissue; (3) the development of rete ducts; 4) the appearance
of defective septa. A general observation was that the
development of sex cords between E20 and E42 in the
ostrich was similar to that reported in other birds. Our
results indicate that the primary sex cords in ostrich orig-
inate from Sertoli cell aggregations which elongate, contact
and anastomose with each other during embryonic devel-
opment, similar to what occurs in other birds. Whereas the
formation of primary sex cords in fowl was completed by
E7.5 of the first half of the embryonic period (Morrish and
Sinclair 2002), in the ostrich embryo primary sex cord
formation was completed at E26 of the second half of the
embryonic period. According to the previous reports, the
appearance of a lumen in the primary sex cords is post-
poned until hatch in birds and until puberty in mammals
(Romanoff 1960; Skinner and Griswold 2005; McGeady
et al. 2006; Gonzalez-Moran and Soria-Castro 2010a; Sa-
dler 2010), but in the ostrich embryo the lumen began to
form on E26, and on E36 a distinct lumen was present;
consequently, these cords can be called ‘‘primary sex
tubules’’. One important feature of these cords is that they
contain filled lumens. Resin sections, but not paraffin
sections, demonstrated that the lumens were filled with a
semisolid fluid. Furthermore, the unoccupied spaces usu-
ally seen in the lumen of post-embryonic seminiferous
tubules were also due to the absence of its fluid contents
(Hassanzadeh et al. 2013).
The most important finding regarding the development
of the interstitial tissue was that fibroblast-like Leydig cells
appeared on E26. In ostrich testis, the appearance of
fibroblast-like Leydig cells began from E26 onwards, while
in fowl this event occurs at E8 (Gonzalez-Moran and Soria-
Castro 2010b) in the first half of the embryonic period. The
presence of these cells has been reported in the testis of
Japanese quail (Nicholls and Graham 1972) and 2-day-old
chicken (Connell 1972), as well as in mammals (Chris-
tensen and Fawcett 1961; Black and Christensen 1969),
and they transform from a fibroblast-like form, through a
transitional stage, to a mature androgen-producing Leydig
cell form (Narbaitz and Adler 1966; Connell 1972; Jorda-
nov et al. 1978; DeFalco et al. 2011). The presence of lipid
granules, which were dark in coloration in most of our
images, is an important characteristic by which to identify
these cells. This is in contrast to the white coloration of
these granules that has been reported by other authors
(Nicholls and Graham 1972; Gonzalez-Moran and Soria-
Castro 2010b). This difference might be related to the
specific methodology used in the study. The PLT method
used in our study preserves the lipid contents of Leydig cell
granules, which are then stained by osmium tetroxide. It
was not possible to define the exact boundaries of Leydig
Fig. 8 Detailed histological characteristics of testis on E42. Trans-
verse sections stained by MT (a) and H&E (d). a Presence of
developing capsular rete ducts (arrowheads) at the free surface of the
testis beside a defective septum (Df) are shown (a0 and a00). Note that
the central portion of seminiferous tubules and rete ducts have been
lost during the staining procedure. b Developing capsular rete ducts
(arrowheads), containing PGC-like (Pl) and Sertoli-like (Sl) cells in
the lateral surface of testis are still present. The magnified parts (b0
and b00) show their morphology in details. H Hilus of testis,
I interstitial tissue, P PGC, S seminiferous tubule, Ta tunica
albuginea, Ts tunica serosa, Tv tunica vasculosa. Arrows Large
cisterns, arrowheads capsular rete ducts. Scale bar 50 lm
136 B. Hassanzadeh et al.
123

cell precursors due to the compression of cells and the
randomness of the fibers in the interstitial tissue. At first,
the fibers of the interstitial tissue were not distinct collagen
fibers (Eurell and Van Sickle 2006), but during develop-
ment collagen fibers appeared, increased in number, and
finally caused the change in interstitial tissue color from
red to a mixed color of green and red. At the territorial
portions of the interstitial tissue, a group of interstitial
tissue fibroblasts similar to peritubular myoid cells were
observed encircling the seminiferous tubules. These cells,
however, did not have the distinct morphological charac-
teristics of the myoid cells which are seen in the juvenile
ostrich testis (Hassanzadeh et al. 2013). It might be possi-
ble that these encircling fibroblasts acquire the morphol-
ogy of myoid cells during post-hatching developmental
processes.
Rete ducts are other noticeable structures of ostrich
testis that consist of three parts of intratesticular, ex-
tratesticular or lacunar, and intracapsular rete ducts or true
rete with large cisterns (Budras and Meier 1981; Aire and
Soley 2003). We recently investigated the capsular rete, the
important part of this system, and reported its extension up
to the free surface of the testis in the juvenile ostrich
(Hassanzadeh et al. 2013). In accordance with that report,
our results also show that the true rete is present in the
medial, lateral and free surfaces of the testis from the pre-
hatching period. Since rete ducts were frequently seen in
the capsule of the polar parts, we can conclude that these
ducts might first appear in the polar parts of the testis and
subsequently extend toward the equatorial parts. Budras
and Meier (1981) proposed that the buds of glomerular
capsules of mesonephros are the origin of rete testis, but
our results indicate that the capsular rete develops from the
cell aggregations trapped inside the tunica albuginea which
are comprised of two cell types: Sertoli-like and PGC-like
cells. Thus, it is a plausible speculation that the capsular
rete ducts may originate from the primary sex cords.
Despite the presence of primary sex cords inside the testis,
the thickness of the epithelium in these capsular aggrega-
tions decreases during embryo development and ultimately
forms a squamous or flattened cuboidal epithelium (Has-
sanzadeh et al. 2013). However, regarding the structural
differences between different parts of the rete system
(Budras and Meier 1981; Aire 1982; Barker and Kendall
1984; Aire and Soley 2003), it is possible that its different
parts develop from diverse origins.
As in other birds, in the ostrich the covering epithelium
and its underlying blood vessels are present from the early
stages of testis formation, while its third layer, the tunica
albuginea, appears later (Gonzalez-Moran and Soria-Castro
2010a). It has been shown that tunica albuginea, the main
part of the testicular capsule, is considerably different in
the ostrich (Ozegbe et al. 2008; Hassanzadeh et al. 2013).
In the ostrich embryo, this layer first consists of a loose
connective tissue, but due to intensification of collagen
fibers (which stain green by MT), changes to a dense
connective tissue in the newly hatched ostrich (Nicholls
and Graham 1972; Eurell and Van Sickle 2006). Initially
this layer had a loose structure, and its internal compo-
nents, such as the capsular rete ducts and blood vessels,
appeared in their real shapes in the sections. After densi-
fication, which forces these structures to collapse, it was
difficult to find these internal components in the sections.
Hardening and thickening of the tunica albuginea has
another outcome, i.e., the appearance of defective septa. In
mammals, testicular septa branch from the inner part of the
capsule and conduct blood vessels and nerves into and out
of the testicular substance (Davis et al. 1970). In earlier
studies, it was believed that these septa were absent in birds
(Lake 1971; Hodges 1974), but their presence were sub-
sequently reported in ostrich (Soley and Groenewald 1999;
Hassanzadeh et al. 2013). Our observations confirm the
presence of defective septa and suggest that the mamma-
lian lamellar septa are the evolutionary remnants of the
defective septa in ostrich which have a filum-like shape.
The defective septa penetrated the interstitial tissue in a
meandering way while conducting blood vessels and
nerves into and out of the testis.
While classical measurements are routinely used in
anatomical and embryological studies, new methods such
as stereology have been applied recently in studies on avian
developmental biology (Gonzalez-Moran 1997, 2011;
Gonzalez-Moran and Soria-Castro 2010a, b). In addition to
measurements of B.W. and testis diameter, we used ster-
eological techniques to estimate the Tes.V and Vv of
seminiferous tubules. The weight of embryos increased in a
linear manner during the period leading up to the hatching
day, but it decreased during the day of hatching (Fig. 2a).
This is a common finding in newborns in both mammals
and birds (Mortola 2009; Eidelman et al. 2012). Our results
show that testis and seminiferous tubule volumes increased
during the embryonic period and that this pattern of
increase was similar to that of the respectivegrowth curves.
Thus, the ratios of Tes.V/B.W. and S.T.V/B.W. fell on E26
(Fig. 2b). Considering that the Vv of seminiferous tubules
generally decreases from E20 to E42 and reaches about
0.56 in the juvenile ostrich (Hassanzadeh et al. 2013), it is
possible to conclude that there is no direct relationship
between the B.W. increase and changes in the Vv of
tubular compartments. However, a decrease in Vv between
E20 and E26 (Fig. 2c) could be related to histological
changes in the components of the testis other than semi-
niferous tubules, including changes in the testis capsule
(appearance of tunica albuginea) and the development of
new blood vessels. In comparison with other animals,
increased Vv of seminiferous tubules from prenatal to
Embryonic development of ostrich testis 137
123

pre-pubertal period is very different from the increasing
pattern in domestic fowls and bonnet monkeys (Macaca
radiata) reported previously (Gonzalez-Moran and Soria-
Castro 2010a; Prakash et al. 2008).
In summary, we report here developmental changes in
the testis of the ostrich embryo based on histological and
morphometrical results and highlight differences in the
development of capsular rete ducts, defective septa and
changes in the Vv of tubular compartments in testis of
ostrich embryo in comparison with other birds. Our results
provide new information on the appearance of the lumen in
the primary sex cords, which is unique to the ostrich among
birds and mammals.
Acknowledgments This research has been supported by a grant
(No. 881) from the Research Council of the Ferdowsi University of
Mashhad. We especially thank Mrs. Naseri (Central lab of FUM), Dr.
J. Sadeghinezhad and Dr. J. Ashrafi Helan for their invaluable help.
Conflict of interest None.
References
Aire TA (1982) The rete testis of birds. J Anat 135(1):97–110
Aire TA, Soley JT (2003) The morphological features of the rete testis
of the ostrich (Struthio camelus). Anat Embryol 207:355–361
Barker SG, Kendall MD (1984) A study of the rete testis epithelium in
several wild birds. J Anat 138(1):139–152
Black VH, Christensen AK (1969) Differentiation of interstitial cells and
Sertoli cells in fetal guinea pig testes. Am J Anat 124(2):211–237
Budras KD, Meier U (1981) The epididymis and its development in
ratite birds (ostrich, emu, rhea). Anat Embryol (Berl)
162(3):281–299
Chang G, Chen R, Qin Y, Zhang Y, Dai A, Chen G (2012) The
development of primordial germ cells (PGCs) and Testis in the
Quail Embryo. Pak Vet J 32(1):88–92
Christensen AK, Fawcett DW (1961) The normal fine structure of
opossum testicular interstitial cells. J Biophys Biochem Cytol
9:653–670
Connell CJ (1972) The effect of luteinizing hormone on the
ultrastructure of the Leydig cell of the chick. Z Zellforsch
128:139–151
Csaba G, Shahin MA, Dobozy O (1980) The overlapping effect of
gonadotropins and TSH on embryonic chicken gonads. Arch
Anat Histol Embryol 63:31–38
Davis JR, Langford GA, Kirby PJ (1970) The testicular capsule. In:
Johnson AD, Gomes WR, Vandemark NL (eds) The testis
development, anatomy and physiology. Academic Press, Lon-
don, pp 281–337
DeFalco T, Takahashi S, Capel B (2011) Two distinct origins for
Leydig cell progenitors in the fetal testis. Dev Biol 352:14–26
Delesse A (1848) Procede mecanique pour determiner la composition
des roches. Ann Min 13:379–388
Eidelman AI, Schanler RJ, Johnston M, Landers S, Noble L, Szucs K,
Viehmann L (2012) Breastfeeding and the use of human milk.
Pediatrics 129:e827–e841
Eurell JA, Van Sickle DC (2006) Connective and supportive tissues.
In: Eurell JA, Frappier BL (eds) Dellmann’s textbook of
veterinary histology. Wiley-Blackwell, London, p 37
Fujimoto T, Ukeshima A, Kiyofuji R (1976) The origin, migration
and morphology of the primordial germ cells in the chick
embryo. Anat Rec 185(2):139–145
Gefen E, Ar A (2001) Morphological description of the developing
ostrich embryo: a tool for embryonic age estimation. Isr J Zool
47:87–97
Ginsburg M, Eyal-Giladi H (1986) Temporal and spatial aspects of
the gradual migration of primordial germ cells from the epiblast
into the germinal crescent in the avian embryo. J Embryol Exp
Morphol 95:53–71
Gonzalez-moran G (1997) A stereological study of the different cell
populations in chicken testes treated with follicle-stimulating
hormone during embryonic development. Anat Histol Embryol
26:311–317
Gonzalez-Moran MG (2011) Histological and stereological changes
in growing and regressing chicken ovaries during development.
Anat Rec 294:893–904
Gonzalez-Moran MG, Soria-Castro E (2010a) Changes in the tubular
compartment of the testis of Gallus domesticus during develop-
ment. Br Poult Sci 51:296–307
Gonzalez-Moran MG, Soria-Castro E (2010b) Histological and
stereological studies on Leydig cells in the testes of Gallus
domesticus from pre-hatching to sexual maturity. Anim Reprod
Sci 120(1–4):129–135
Hassanzadeh B, Nabipour A, Rassouli M, Dehghani H (2013)
Microanatomical study of testis in juvenile ostrich (Struthio
camelus). Anat Sci Int 88(3):134–140
Hodges RD (1974) The histology of the fowl. Academic Press,
London
Howard V, Reed MG (2005) Unbiased stereology: three-dimensional
measurement in microscopy. Garland Science/BIOS Scientific
Publishers, New York
Ishiguru S, Minematsu T, Naito M, Kanai Y, Tajima A (2009)
Migratory ability of chick primordial germ cells transferred into
quail embryos. J Reprod Dev 55(2):183–186
Jordanov J, Angelova P, Boyadjieva-Michailova A, Bakalska M
(1978) Ultrastructure of developing interstitial cells in chick
embryonic gonad in relation to their genesis and steroidogenic
function. Z Mikrosk Anat Forsch 92(3):449–464
Kuo J (2007) Electron microscopy methods and protocols. Humana
Press, Totowa
Lake PE (1971) The male in reproduction. In: Bell DJ, Freeman BM
(eds) Physiology and biochemistry of the domestic fowl.
Academic Press, London, pp 1411–1447
Lan W, Ke-mei P, Lai-qiang L, Hui S, Yan W, Sheng-he L, Li T, An-
na D, Er-hui J, Jia-xiang W (2007) Morphological study on the
reproductive organs of the male ostrich chicks. Acta Vet Zootech
Sinica 38(8):851–855
Madekurozwa M-C, Chabvepi TS, Matema S, Teerds KJ (2002)
Relationship between seasonal changes in spermatogenesis in the
juvenile ostrich (Stuthio camelus) and the presence of the LH
receptor and 3b-hydroxysteroid dehydrogenase. Reproduction
123(5):735–742
Malago W Jr, Franco HM, Matheucci E Jr, Medaglia A, Henrique-
Silva F (2002) Large scale sex typing of ostriches using DNA
extracted from feathers. BMC Biotechnol 2:19
Malecki IA, Horbanczuk JO, Reed CE, Martin GB (2005) The ostrich
(Struthio camelus) blastoderm and embryo development follow-
ing storage of eggs at various temperatures. Br Poult Sci
46(6):652–660
McGeady TA, Quinn PJ, FitzPatrick ES, Ryan MT, Cahalan S (2006)
Veterinary embryology. Blackwell, London
Moller AP (1994) Directional selection on directional asymmetry:
testes size and secondary sexual characters in birds. Proc R Soc
Lond B 258:147–151
138 B. Hassanzadeh et al.
123

Morrish BC, Sinclair AH (2002) Vertebrate sex determination: many
means to an end. Reproduction 124(4):447–457
Mortola JP (2009) Gas exchange in avian embryos and hatchlings.
Comp Biochem Phys A 153(4):359–377
Nagai H, Mak S–S, Weng W, Nakaya Y, Ladher R, Sheng G (2011)
Embryonic development of the emu, Dromaius novaehollandiae.
Dev Dyn 240(1):162–175
Narbaitz R, Adler R (1966) Submicroscopical observations on the
differentiation of chick gonads. J Embryol Exp Morph
16(1):41–47
Nicholls TJ, Graham GP (1972) Observations on the ultrastructure
and differentiation of Leydig cells in the testis of the Japanese
quail (Coturnix coturnix japonica). Biol Reprod 6(2):179–192
Ozegbe PC, Aire TA, Soley JT (2006) The morphology of the efferent
ducts of the testis of the ostrich, a primitive bird. Anat Embryol
211(5):559–565
Ozegbe PC, Aire TA, Madekurozwa MC, Soley JT (2008) Morpho-
logical and immunohistochemical study of testicular capsule and
peritubular tissue of emu (Dromaius novaehollandiae) and
ostrich (Struthio camelus). Cell Tissue Res 332(1):151–158
Ozegbe PC, Kimaro W, Madekurozwa MC, Soley JT, Aire TA (2010)
The excurrent ducts of the testis of the emu (Dromaius
novaehollandiae) and ostrich (Struthio camelus): microstereol-
ogy of the epididymis and immunohistochemistry of its
cytoskeletal systems. Anat Histol Embryol 39(1):7–16
Prakash S, Prithiviraj E, Suresh S (2008) Developmental changes of
seminiferous tubule in prenatal, postnatal and adult testis of
bonnet monkey (Macaca radiata). Anat Histol Embryol
37(1):19–23
Romanoff AL (1960) The avian embryo. Structural and functional
development. Macmillan, New York
Sadler TW (2010) Langman’s medical embryology. Lippincott
Williams & Wilkins, Philadelphia
Sekido R, Lovell-Badge R (2007) Mechanisms of gonadal morpho-
genesis are not conserved between chick and mouse. Dev Biol
302:132–142
Shahin MA, Torok O (1982) Differentiation of male chick embryo
gonads during the last week of incubation in organ and tissue
culture. Z Mikrosk Anat Forsch 96(1):157–170
Skinner MK, Griswold MD (2005) Sertoli cell biology. Academic
Press, London
Smith CA (2007) Avian, sex determination and gonadal sex
differentiation. In: Jamieson BGM (ed) Reproductive biology
and phylogeny of birds. Science Publishers, Enfield, pp 479–506
Soley JT (1992) A histological study of spermatogenesis in the ostrich
(Struthio camelus). PhD thesis. University of Pretoria, Pretoria
Soley JT, Groenewald HB (1999) Reproduction. In: Deeming DC (ed)
The ostrich: biology production and health. CABI Publishing,
New York, pp 129–159
Tagami T, Kagami H, Matsubara Y, Harumi T, Naito M, Takeda K,
Hanada H, Nirasawa K (2007) Differentiation of female
primordial germ cells in the male testes of chicken (Gallus
gallus domesticus). Mol Reprod Dev 74(1):68–75
Van Krey H (1990) Reproductive biology in relation to breeding and
genetics. In: Crawford RD (ed) Poultry breeding and genetics.
Elsevier, Amsterdam, pp 61–90
Wei L, Peng KM, Liu H, Song H, Wang Y, Tang L (2011)
Histological examination of testicular cell development and
apoptosis in the ostrich chick. Turk J Vet Anim Sci 35(1):7–14
Zaccanti F, Vallisneri M, Quaglia A (1990) Early aspects of sex
differentiation in the gonads of chick embryos. Differentiation
43(2):71–80
Zhang Y, Ren Z, Tang L (2011) Anatomic study on the main male
reproductive organs of ostrich. Global J Health Sci 3(1):181–184
Embryonic development of ostrich testis 139
123


















