
Molecular Biology Fourth Edition
60
Molecular Biology Fourth Edition Chapter 17 The Mechanism of Translation I: Initiation Robert F. Weaver Chapter 18 The Mechanism of Translation II: Elongation and Termination Chapter 19 Ribosomes and Transfer RNA
-
Upload
wyoming-bean -
Category
Documents
-
view
43 -
download
1
description
Molecular Biology Fourth Edition. Chapter 18 The Mechanism of Translation II: Elongation and Termination. Chapter 17 The Mechanism of Translation I: Initiation. Chapter 19 Ribosomes and Transfer RNA. Robert F. Weaver. 18.1 Direction of Polypeptide Synthesis and mRNA Translation. - PowerPoint PPT Presentation
Transcript of Molecular Biology Fourth Edition

Molecular BiologyFourth Edition
Chapter 17
The Mechanism of Translation I: Initiation
Robert F. Weaver
Chapter 18The Mechanism of Translation II: Elongation and Termination
Chapter 19
Ribosomes and Transfer RNA

18-2
18.1 Direction of Polypeptide Synthesis and mRNA Translation• Messenger RNAs are read in the 5’3’
direction
• This is the same direction in which they are synthesized
• Proteins are made in the aminocarboxyl direction
• This means that the amino terminal amino acid is added first

18-3
Strategy to Determine Direction of Translation

18-4
18.2 The Genetic Code
• The term genetic code refers to the set of 3-base code words (codons) in mRNA that represent the 20 amino acids in proteins
• Basic questions were answered about translation in the process of “breaking” the genetic code

18-5
Nonoverlapping Codons
• Each base is part of at most one codon in nonoverlapping codons
• In an overlapping code, one base may be part of two or even three codones
AUGUUC
No overlapping AUG UUC
Overlapping AUG UGU GUU UUC
1. If no overlapping, a change of one base in an mRNA would change no more than one a.a. in the resulting protein.
2. If overlapping, up to three adjacent amino acids could be changed.

No-overlapping
One base change in TMV mRNA never caused more than
one a.a. mutation

18-7
No Gaps in the Code
• If the code contained untranslated gaps or “commas”, mutations adding or subtracting a base from the message might change a few codons
• Would still expect ribosome to be back “on track” after the next such comma
• Mutations might frequently be lethal– Many cases of mutations should occur just
before a comma and have little, if any, effect

18-8
Frameshift MutationsFrameshift mutations • Translation starts AUGCAGCCAACG
• Insert an extra base AUXGCAGCCAACG
– Extra base changes not only the codon in which is appears, but every codon from that point on
– The reading frame has shifted one base to the leftCode with commas • Each codon is flanked by one or more
untranslated bases – Commas would serve to set off each codon so that
ribosomes recognize it • Translation starts AUGZCAGZCCAZACGZ
• Insert an extra base AUXGZCAGZCCAZACGZ
– First codon wrong, all others separated by Z, translated normally

18-9
Frameshift Mutation Sequences

18-10
The Triplet Code
• The genetic code is a set of three-base code words, or codons– In mRNA, codons instruct the ribosome to
incorporate specific amino acids into a polypeptide
• Code is nonoverlapping– Each base is part of only one codon
• Devoid of gaps or commas– Each base in the coding region of an mRNA is
part of a codon

18-11
Breaking the Code
• The genetic code was broken– Using:
• Synthetic messengers• Synthetic trinucleotides
– Then observing: • Polypeptides synthesized• Aminoacyl-tRNAs bound to ribosomes
• There are 64 codons– 3 are stop signals– Remainder code for amino acids– The genetic code is highly degenerate

How to test the the triplet code hypothesis
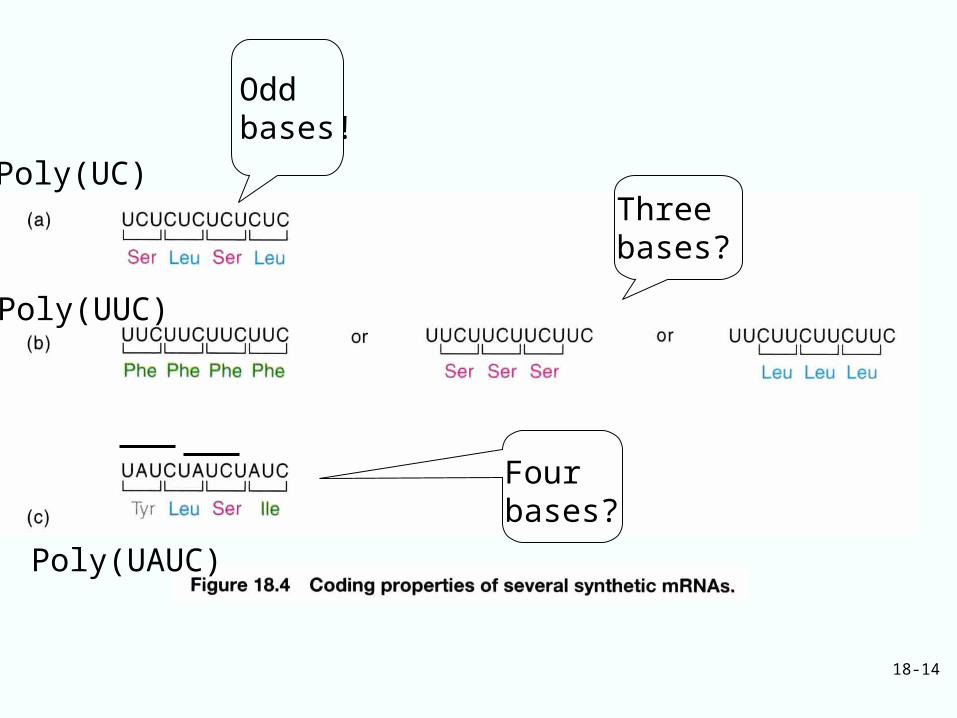
In 1961, Nirenberg and Matthaei,
Poly(U) can be translated into poly-Phe
Synthetic mRNA of defined sequence can shed light on the nature of the code

First, the codons contained odd number of bases.
Poly[UC] or UCUCUCUC…….
Odd number of bases------UCU or CUC will code for di-peptide
Even number of bases-----CUCU or UCUC will code for single repeat
Found…. code for poly(ser-leu)---So, the codons contained an odd number of bases
18-14
Four bases?
Three bases?
Odd bases!
Poly(UUC)
Poly(UC)
Poly(UAUC)

18-15
Four different bases, only twenty amino acids
Triplet 4x4x4=64
Two-base codon 4x4=16 not enough combination
Nine-base codon 49=262,144 too many

18-16
The Genetic Code

18-17
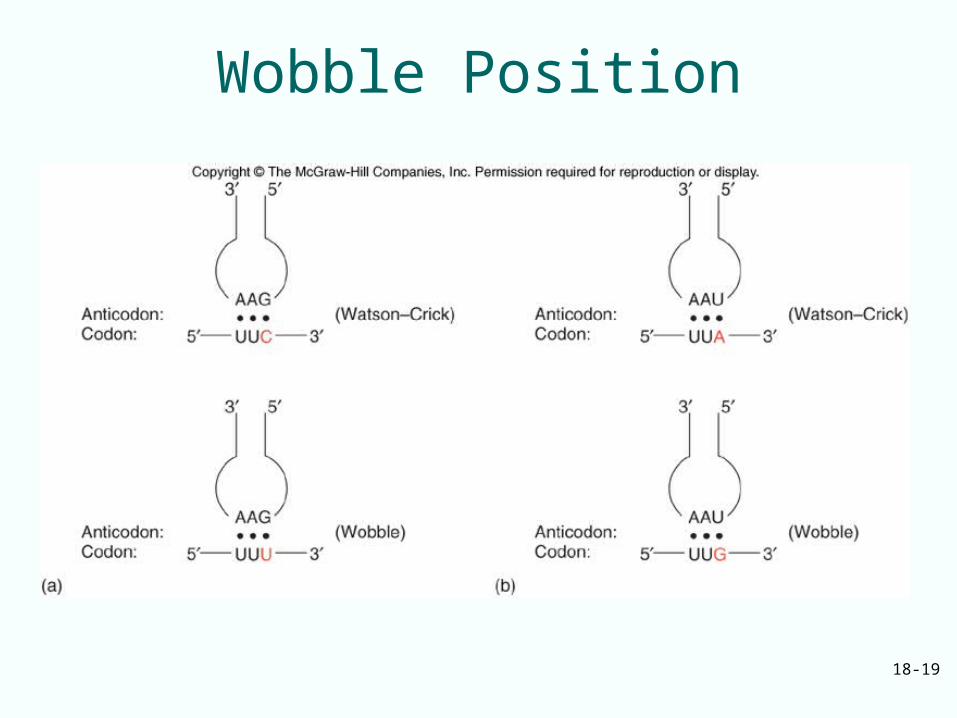
Unusual Base Pairs Between Codon and Anticodon
Degeneracy of genetic code is accommodated by:– Isoaccepting species of tRNA: bind same
amino acid, but recognize different codons– Wobble, the 3rd base of a codon is allowed to
move slightly from its normal position to form a non-Watson-Crick base pair with the anticodon
– Wobble allows same aminoacyl-tRNA to pair with more than one codon (to reduce the number of tRNA to translate genetic code)

18-18
Wobble Base Pairs
• Compare standard Watson-Crick base pairing with wobble base pairs
• Wobble pairs are:– G-U– I-A (or C, or U)
18-19
Wobble Position

18-20
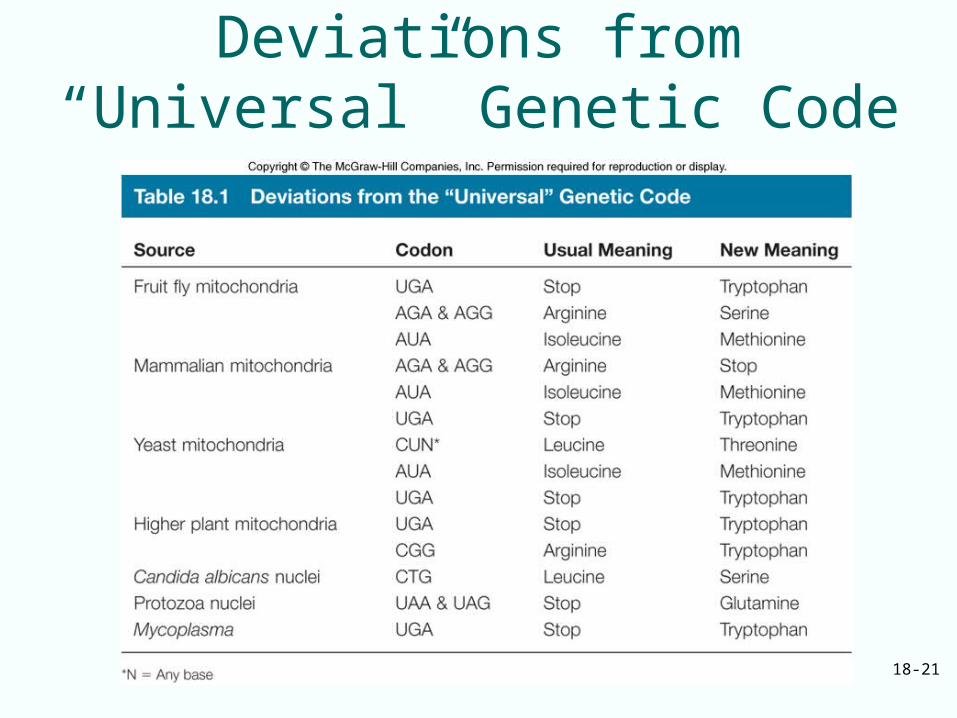
Almost Universal Code• Genetic code is NOT strictly universal• Certain eukaryotic nuclei and mitochondria along
with at least one bacterium– Codons cause termination in standard genetic code
can code for amino acids Trp, Glu– Mitochondrial genomes and nuclei of at least one
yeast have sense of codon changed from one amino acid to another
• Deviant codes are still closely related to standard one from which they evolved
• Genetic code a frozen accident or the product of evolution– Ability to cope with mutations evolution
18-21
Deviations from “Universal” Genetic Code

18-22
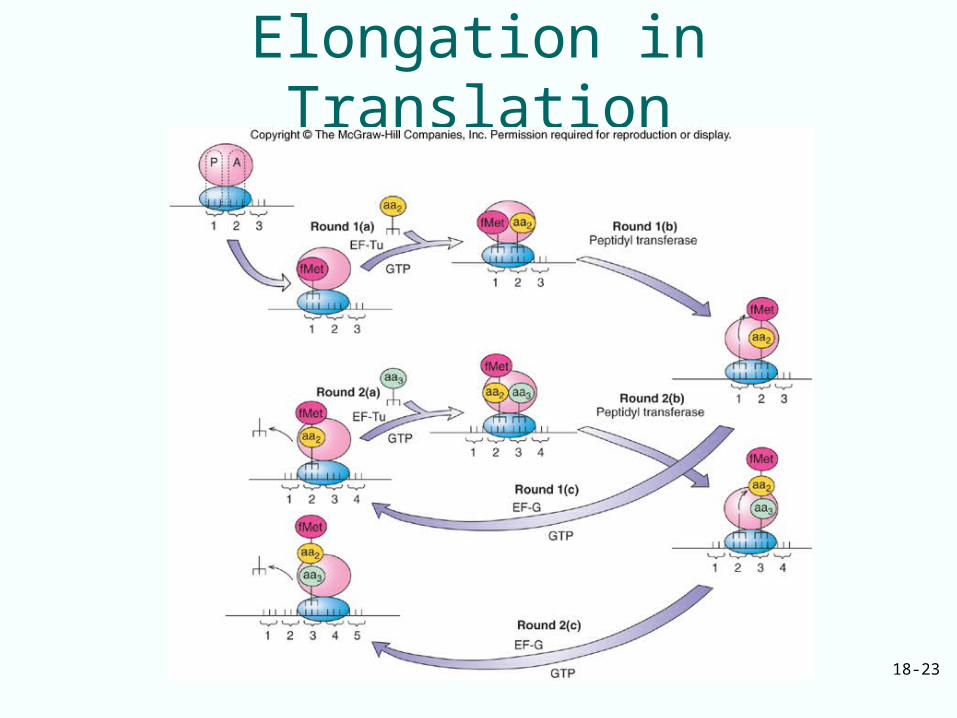
18.3 The Elongation Mechanism
Elongation takes place in three steps:1. EF-Tu with GTP binds aminoacyl-tRNA to
the ribosomal A site2. Peptidyl transferase forms a peptide bond
between peptide in P site and newly arrived aminoacyl-tRNA in the A siteLengthens peptide by one amino acid and shifts it to the A site
3. EF-G with GTP translocates the growing peptidyl-tRNA with its mRNA codon to the P site
18-23
Elongation in Translation

18-24
A Three-Site Model of the Ribosome
• Puromycin – Resembles an aminoacyl-tRNA– Can bind to the A site– Couple with the peptide in the P site– Release it as peptidyl puromycin
• If peptidyl-tRNA is in the A site, puromycin will not bind to ribosome, peptide will not be released
• Two sites are defined on the ribosome:– Puromycin-reactive site (P)– Puromycin unreactive site (A)
• 3rd site (E) for deacylated tRNA bind to E site as exits ribosome
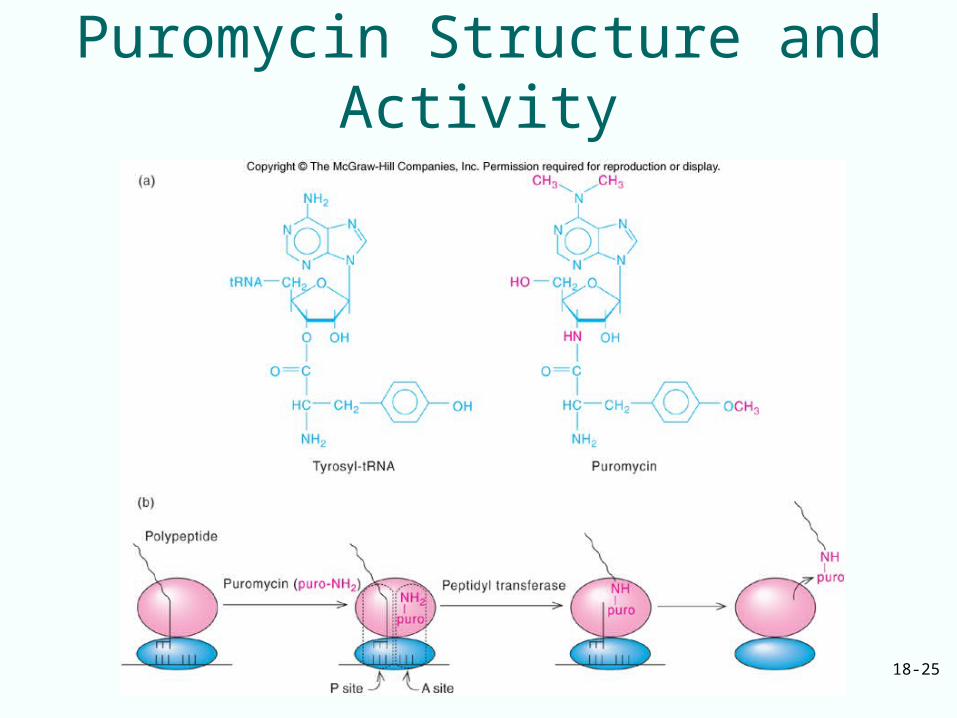
18-25
Puromycin Structure and Activity

18-26
Protein Factors and Peptide Bond Formation
• One factor is T, transfer– It transfers aminoacyl-tRNAs to the ribosome– Actually 2 different proteins
• Tu, u stands for unstable• Ts, s stands for stable
• Second factor is G, GTPase activity
• Factors EF-Tu and EF-Ts are involved in the first elongation step
• Factor EF-g participates in the third step

18-27
Elongation Step 1Binding aminoacyl-tRNA to A site of ribosome
• Ternary complex formed from: – EF-Tu– Aminoacyl-tRNA– GTP
• Delivers aminoacyl-tRNA to ribosome A site without hydrolysis of GTP• Next step:
– EF-Tu hydrolyzes GTP– Ribosome-dependent GTPase activity– EF-Tu-GDP complex dissociates from ribosome
• Addition of aminoacyl-tRNA reconstitutes ternary complex for another round of translation elongation
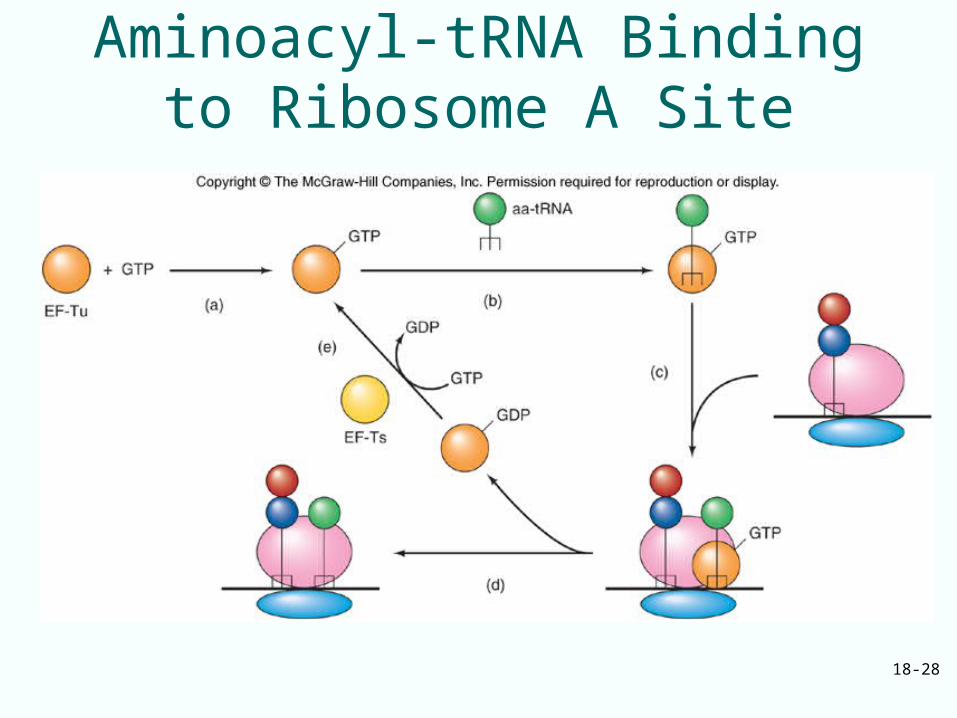
18-28
Aminoacyl-tRNA Binding to Ribosome A Site

18-29
Proofreading
• Protein synthesis accuracy comes from charging tRNAs with correct amino acids
• Proofreading is correcting translation by rejecting an incorrect aminoacyl-tRNA before it can donate its amino acid
• Protein-synthesizing machinery achieves accuracy during elongation in two steps

18-30
Protein-Synthesizing Machinery
• Two steps achieve accuracy:– Gets rid of ternary complexes bearing wrong
aminoacyl-tRNA before GTP hydrolysis– If this screen fails, still eliminate incorrect aminoacyl-
tRNA in the proofreading step before wrong amino acid is incorporated into growing protein chain
• Steps rely on weakness of incorrect codon-anticodon base pairing to ensure dissociation occurs more rapidly than either GTP hydrolysis or peptide bond formation

18-31
Proofreading Balance• Balance between speed and accuracy of
translation is delicate– If peptide bond formation goes too fast
• Incorrect aminoacyl-tRNAs do not have enough time to leave the ribosome
• Incorrect amino acids are incorporated into proteins
– If translation goes too slowly• Proteins are not made fast enough for the
organism to grow successfully
• Actual error rate, ~0.01% per amino acid is a good balance between speed and accuracy

18-32
Elongation Step 2
• One the initiation factors and EF-Tu have done their jobs, the ribosome has fMet-tRNA in the P site and aminoacyl-tRNA in the A site
• Now form the first peptide bond• No new elongation factors participate in
this event• Ribosome contains the enzymatic activity,
peptidyl transferase, that forms peptide bond
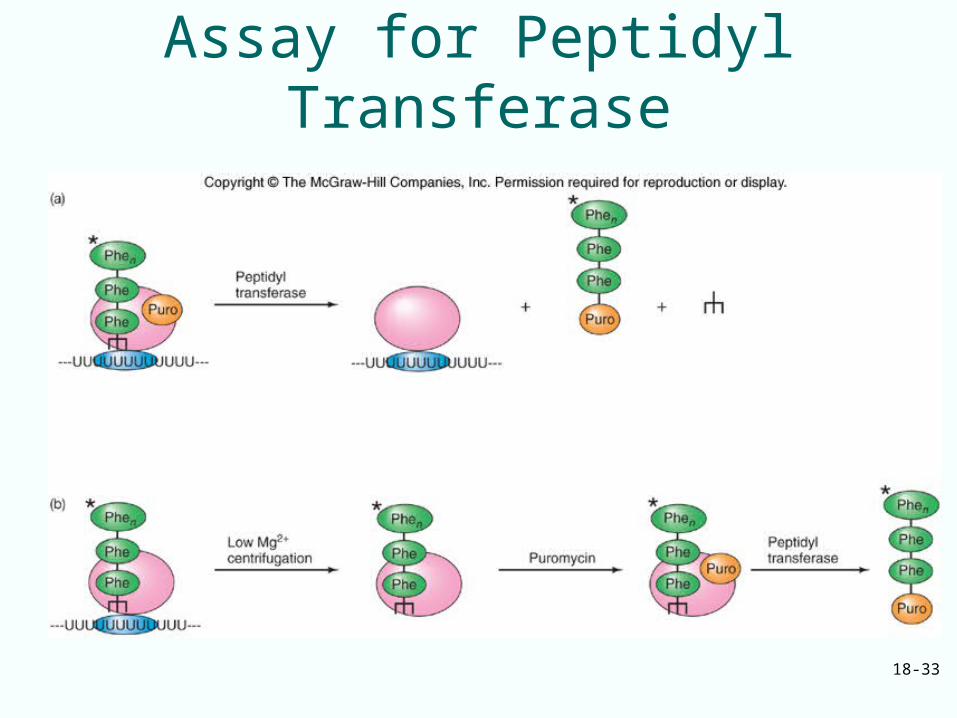
18-33
Assay for Peptidyl Transferase

18-34
Peptide Bond Formation
• The peptidyl transferase resides on the 50S ribosomal particle
• Minimum components necessary for activity are 23S rRNA and proteins L2 and L3
• 23S rRNA is at the catalytic center of peptidyl transferase

18-35
Elongation Step 3
• When peptidyl transferase has worked:– Ribosome has peptidyl-tRNA in the A site– Deacylated tRNA in the P site
• Translocation, next step, moves mRNA and peptidyl-tRNA one codon’s length through the ribosome– Places peptidyl-tRNA in the P site– Ejects the deacylated tRNA– Process requires elongation factor EF-G
which hydrolyzes GTP after translocation is complete
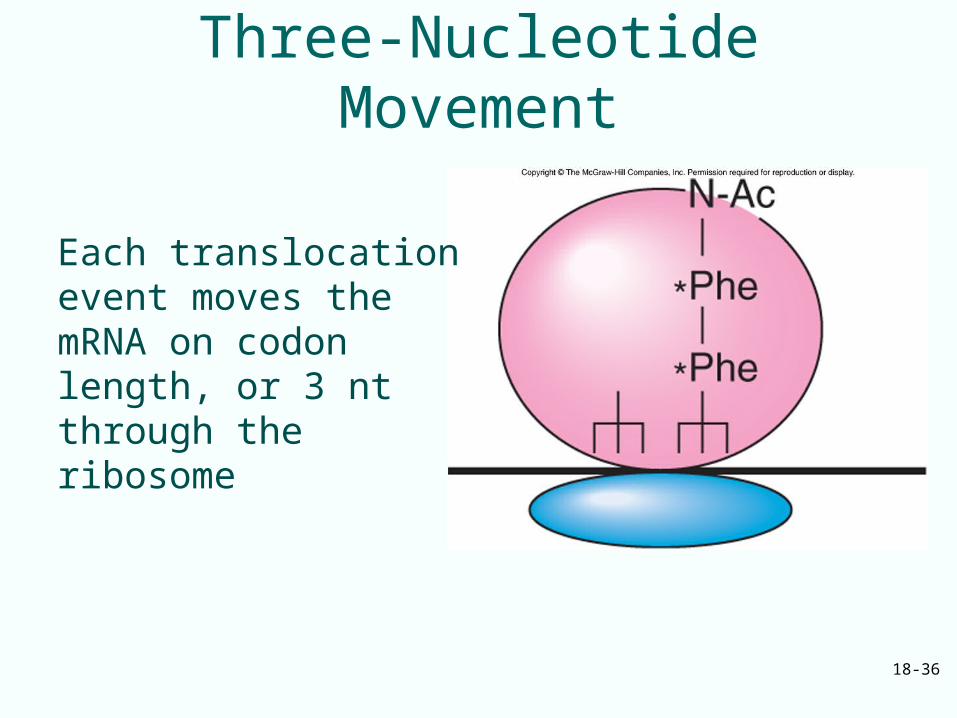
18-36
Three-Nucleotide Movement
Each translocation event moves the mRNA on codon length, or 3 nt through the ribosome

18-37
Role of GTP and EF-G• GTP and EF-G are necessary for
translocation– Translocation activity appears to be inherent
in the ribosome– This activity can be expressed without EF-G
and GTP
• GTP hydrolysis – Precedes translocation– Significantly accelerate translocation
• New round of elongation occurs if:– EF-G must be released from the ribosome– Release depends on GTP hydrolysis

18-38
GTPases and Translation• Some translation factors harness GTP
energy to catalyze molecular motions• These factors belong to a large class of G
proteins– Activated by GTP– Have intrinsic GTPase activity activated by an
external factor (GAP)– Inactivated when they cleave their own GTP
to GDP– Reactivated by another external factor
(guanine nucleotide exchange protein) that replaces GDP with GTP

18-39
Structures of EF-Tu and EF-G
• Three-dimensional shapes determined by x-ray crystallography: – EF-Tu-tRNA-GDPNP
ternary complex– EF-G-GDP binary
complex
• As predicted, the shapes are very similar

18-40
18.4 Termination
• Elongation cycle repeats over and over– Adds amino acids one at a time– Grows the polypeptide product
• Finally ribosome encounters a stop codon– Stop codon signals time for last step– Translation last step is termination

18-41
Termination Codons
• Three codons are the natural stop signals at the ends of coding regions in mRNA– UAG– UAA– UGA
• Mutations can create termination codons within an mRNA causing premature termination of translation– Amber mutation creates UAG– Ochre mutation creates UAA– Opal mutation creates UGA
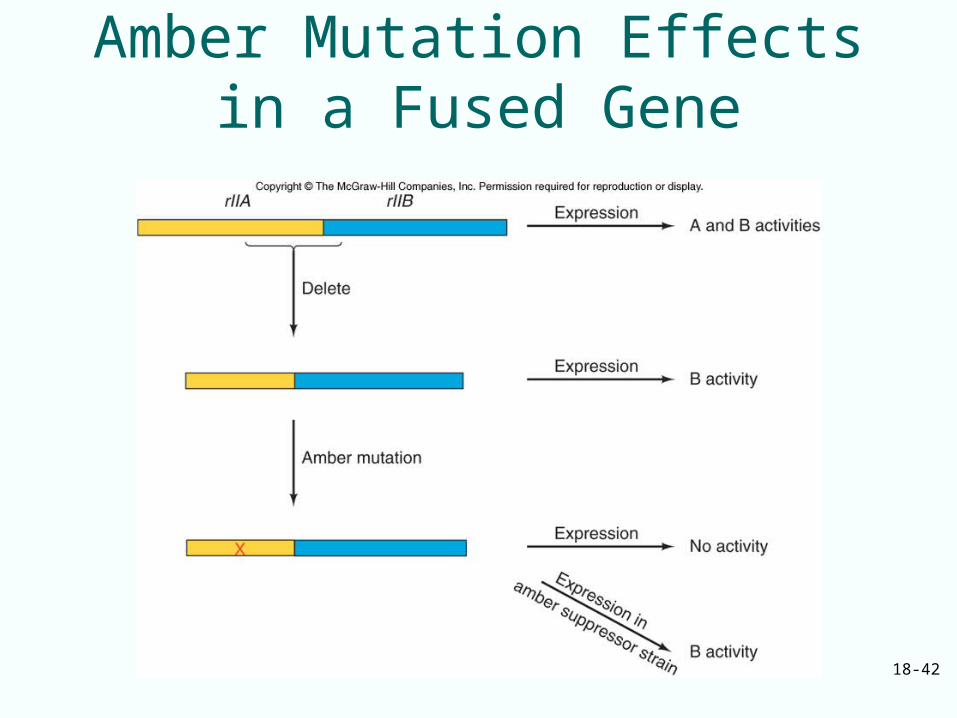
18-42
Amber Mutation Effects in a Fused Gene

18-43
Termination Mutations
• Amber mutations are caused by mutagens that give rise to missense mutations
• Ochre and opal mutations do not respond to the same suppressors as do the amber mutations– Ochre mutations have their own suppressors– Opal mutations also have unique suppressors
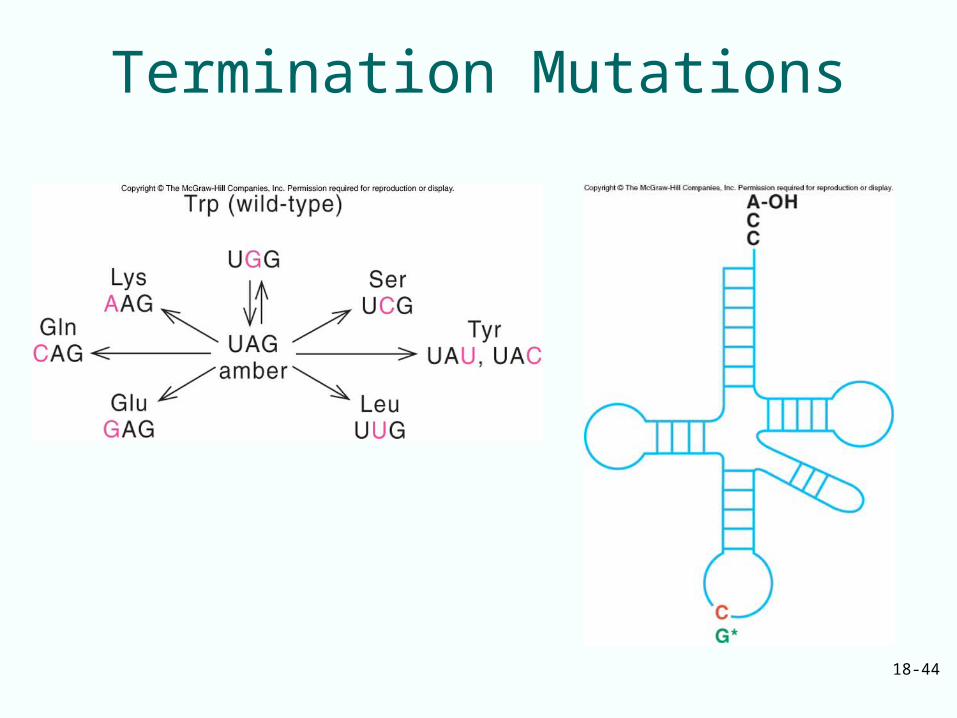
18-44
Termination Mutations

18-45
Stop Codon Suppression
• Most suppressor tRNAs have altered anticodons: – Recognize stop
codons– Prevent termination by
inserting an amino acid
– Allow ribosome to move on to the next codon

18-46
Release Factors
• Prokaryotic translation termination is mediated by 3 factors:– RF1 recognizes UAA and UAG– RF2 recognizes UAA and UGA– RF3 is a GTP-binding protein facilitating
binding of RF1 and RF2 to the ribosome
• Eukaryotes has 2 release factors:– eRF1 recognizes all 3 termination codons– eRF3 is a ribosome-dependent GTPase
helping eRF1 release the finished polypeptide
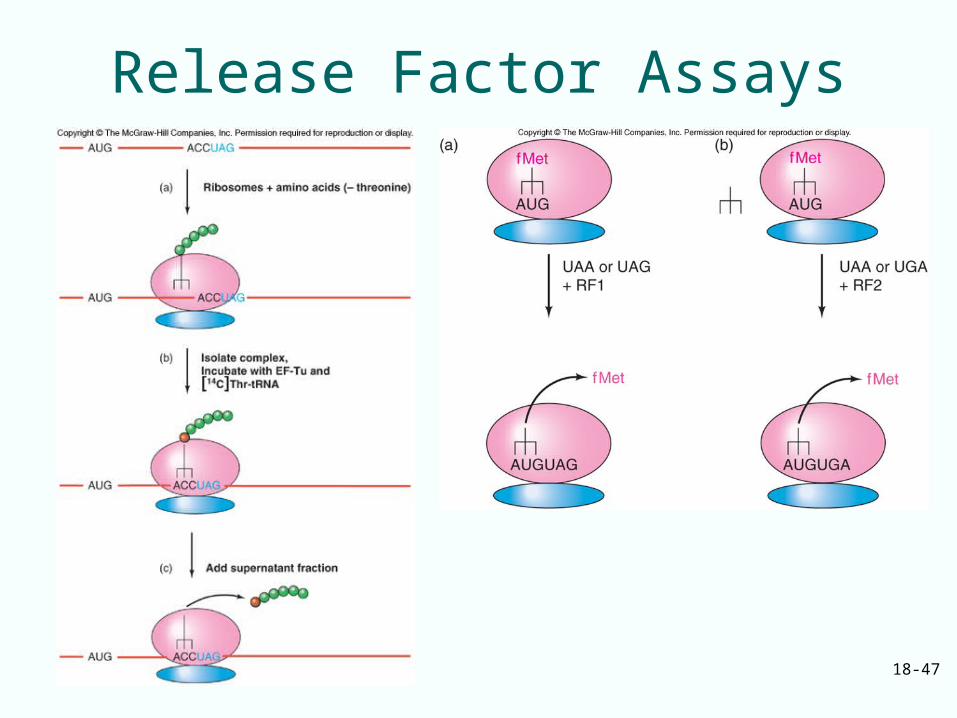
18-47
Release Factor Assays

18-48
Dealing with Aberrant Termination
• Two kinds of aberrant mRNAs can lead to aberrant termination– Nonsense mutations can occur that cause premature
termination– Some mRNAs (non-stop mRNAs) lack termination
codons• Synthesis of mRNA was aborted upstream of termination
codon• Ribosomes translate through non-stop mRNAs and then stall
• Both events cause problems in the cell yielding incomplete proteins with adverse effects on the cell– Stalled ribosomes out of action– Unable to participate in further protein synthesis

18-49
Non-Stop mRNAs• Prokaryotes deal with non-stop mRNAs by tmRNA-
mediated ribosome rescue– Alanyl-tmRNA resembles alanyl-tRNA– Binds to vacant A site of a ribosome stalled on a non-
stop mRNA– Donates its alanine to the stalled polypeptide
• Ribosome shifts to translating an ORF on the tmRNA (transfer-messenger RNA)– Adds another 9 amino acids to the polypeptide before
terminating– Extra amino acids target the polypeptide for destruction– Nuclease destroys non-stop mRNA
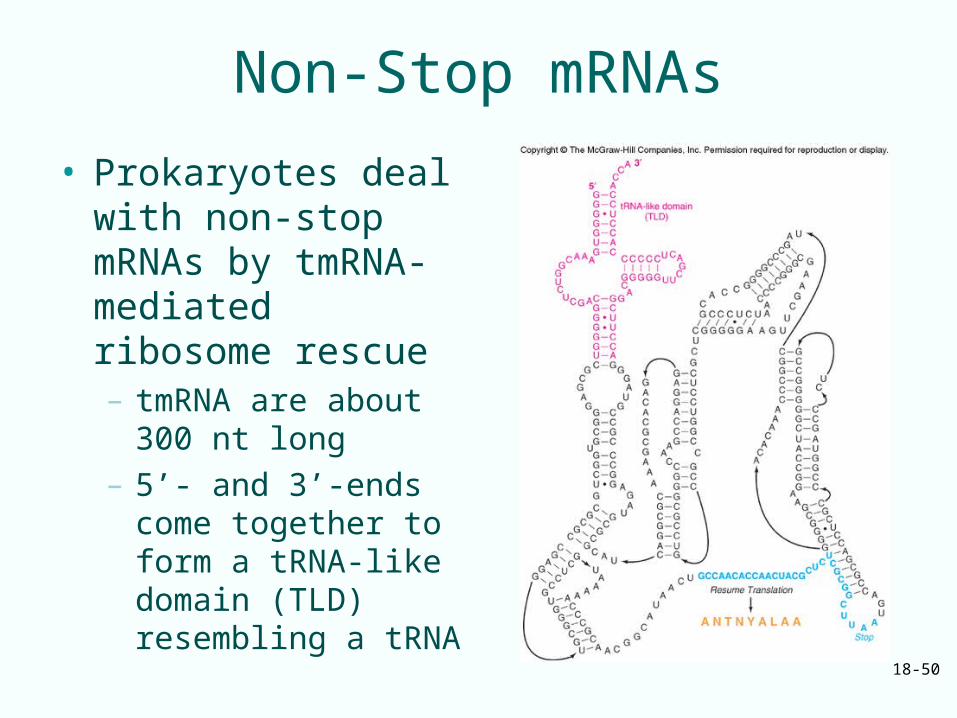
18-50
Non-Stop mRNAs
• Prokaryotes deal with non-stop mRNAs by tmRNA-mediated ribosome rescue– tmRNA are about 300
nt long– 5’- and 3’-ends come
together to form a tRNA-like domain (TLD) resembling a tRNA

18-51
Eukaryotic Aberrant Termination• Eukaryotes do not have tmRNA• Eukaryotic ribosomes stalled at the end of the
poly(A) tail contain 0 – 3 nt of poly(A) tail– This stalled ribosome state is recognized by carboxyl-
terminal domain of a protein called Ski7p– Ski7p also associates tightly with cytoplasmic
exosome, cousin of nuclear exosome– Non-stop mRNA recruit Ski7p-exosome complex to
the vacant A site– Ski complex is recruited to the A site
• Exosome, positioned just at the end of non-stop mRNA, degrades that RNA
• Aberrant polypeptide is presumably destroyed

18-52
Exosome-Mediated Degradation
• This stalled ribosome state is recognized by carboxyl-terminal domain of a protein called Ski7p
• Ski7p also associates tightly with cytoplasmic exosome, cousin of nuclear exosome
• Non-stop mRNA recruit Ski7p-exosome complex to the vacant A site
• Ski complex is recruited to the A site

18-53
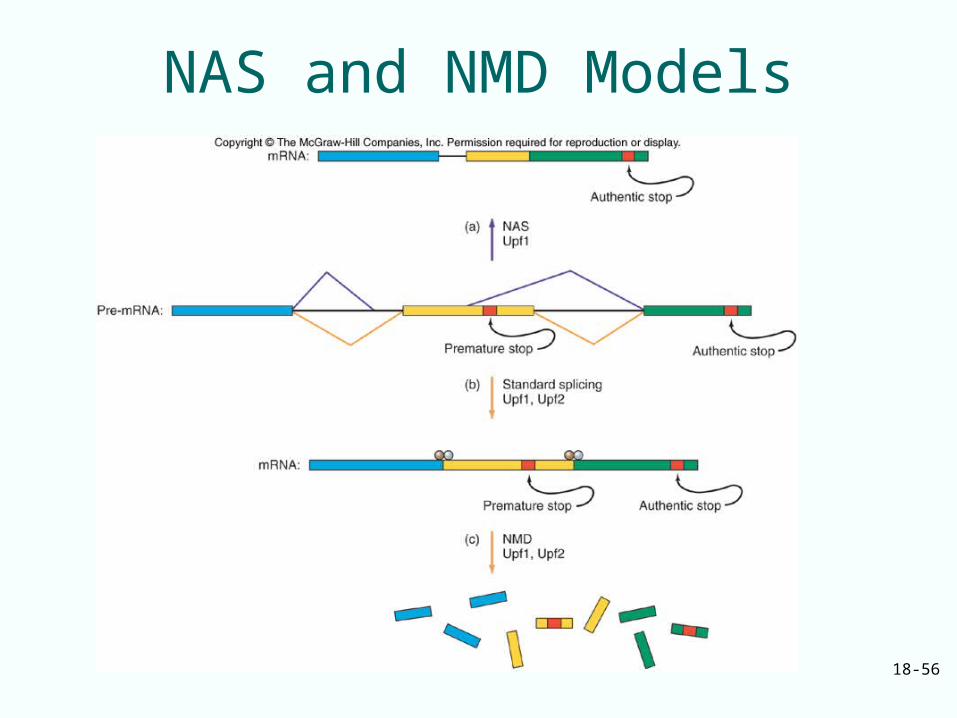
Premature Termination• Eukaryotes deal with premature
termination codons by 2 mechanisms:– NMD (nonsense-mediated mRNA decay)
• Mammalian cells use a downstream destabilizing element
• Yeast cells appear to recognize a premature stop codon
– NAS (nonsense-associated altered splicing)• Senses a stop codon in the middle of a reading
frame• Changes the splicing pattern so premature stop
codon is spliced out of mature mRNA
– Both mechanisms require Upf1

18-54
Mammalian NMD
NMD in mammalian cells involves a downstream destabilizing element
• Upf1• Upf2
– Bind to mRNA at exon-exon junction that measures distance to a stop codon
– Codon far enough upstream• Looks like a stop codon• Activates downstream destabilizing element to
degrade mRNA

18-55
Yeast NMD
• Yeast cells appear to recognize a premature stop codon by the absence of a normal 3’-UTR or poly (A) nearby
• Ribosome stopping at premature stop codon moves to an upstream AUG
• This may mark the mRNA for destruction
18-56
NAS and NMD Models

18-57
Use of Stop Codons to Insert Unusual Amino Acids
Unusual amino acids are incorporated into growing polypeptides in response to termination codons
– Selenocysteine uses a special tRNA • Anticodon for UGA codon• Charged with serine then converted to
selenocysteine• Selenocysteyl-tRNA escorted to ribosome by
special EF-Tu
– Pyrrolysine uses a special tRNA synthetase that joins preformed pyrrolysine with a special tRNA having an anticodon recognizing UAG

18-58
18.5 Posttranslation
• Translation events do not end with termination– Proteins must fold properly– Ribosomes need to be released from mRNA
and engage in further translation rounds
• Folding is actually a cotranslational event occurring as nascent polypeptide is being made

18-59
Folding Nascent Proteins
• Most newly-made polypeptides do not fold properly alone– Polypeptides require folding help from
molecular chaperones– E. coli cells use a trigger factor
• Associates with the large ribosomal subunit• Catches the nascent polypeptide emerging from
ribosomal exit tunnel in a hydrophobic basket to protect from water
– Archaea and eukaryotes lack trigger factor, use freestanding chaperones

18-60
Release of Ribosomes from mRNA
• Ribosomes do not release from mRNA spontaneously after termination
• Help is required from ribosome recycling factor (RRF) and EF-G– RRF resembles a tRNA
• Binds to ribosome A site• Uses a position not normally taken by a tRNA
– Collaborates with EF-G in releasing either 50S ribosome subunit or whole ribosome


















