
Molecular Analysis of Oryza latifolia Desv. (CCDD Genome)-Derived Introgression Lines and...
14
Journal of Heredity doi:10.1093/jhered/esu032 © The American Genetic Association. 2014. All rights reserved. For permissions, please e-mail: [email protected] 1 Molecular Analysis of Oryza latifolia Desv. (CCDD Genome)-Derived Introgression Lines and Identification of Value-Added Traits for Rice ( O. sativa L.) Improvement ROSALYN B. ANGELES-SHIM, RICKY B. VINARAO, BALRAM MARATHI, AND KSHIROD K. JENA From the Novel Gene Resources Laboratory, Plant Breeding, Genetics, and Biotechnology Division, International Rice Research Institute, DAPO Box 7777, Metro Manila, Philippines (Angeles-Shim, Vinarao, Marathi, and Jena); and the Bioscience and Biotechnology Center, Nagoya University, Chikusa, Nagoya, Aichi 464-8601, Japan (Angeles-Shim). Address correspondence to Kshirod K. Jena at the address above, or e-mail: [email protected]. Abstract Oryza latifolia is a tetraploid wild Oryza species with a CCDD genome that has been reported to harbor resistance to bacterial blight (BB), brown planthopper, and whitebacked planthopper. Aside from these traits, O. latifolia is also being tapped as a new source of resistance to lodging and high biomass production. To explore the genetic potential of O. latifolia as a novel genetic resource for the improvement of existing O. sativa cultivars, 27 disomic derivatives of O. latifolia monosomic alien addition lines (MAAL) were characterized for alien chromosome segment introgressions and evaluated for yield components, BB resistance, and strong stem characteristics. A total of 167 simple sequence repeat, sequence tagged site, and single nucleotide polymor- phism markers, along with newly developed indel markers that were specifically designed to detect O. latifolia chromosome seg- ment introgressions in an O. sativa background, were used to define alien introgressions in 27 disomics derived from O. latifolia MAALs. Genotype data showed that 32 unique introgressions spanning 0.31–22.73 Mb were introgressed in different combina- tions in each of the 27 disomic derivatives. Evaluation of the disomic derivatives for agronomic traits identified lines with puta- tive QTLs for resistance to Philippine races 3A, 4, 9A, and 9D of BB. Putative quantitative trait loci (QTLs) conferring strong stem in 19 out of the 27 disomic derivatives studied were also identified from O. latifolia introgressions on chromosome 6. Subject areas: Genomics and gene mapping; Conservation genetics and biodiversity Key words: disomics, DNA markers, lodging resistance, monosomic alien addition lines, wild introgression, Xanthomonas oryzae pv. oryzae The genus Oryza is relatively small but complex, consisting of 2 cultivated species, Oryza sativa and O. glaberrima, and 22 well-recognized wild relatives (Ge et al. 1999; Zeigler 2013). Based on classical taxonomy and isozyme, restric- tion fragment length polymorphism (RFLP) and sequence analysis, the members of this genus are grouped into 4 species complexes, namely, O. sativa, O. officinalis, O. ridleyi, and O. granulata. Species belonging to the O. sativa complex, including both cultivated species, are diploids (2n = 24) with AA genome. The more distantly related species are either diploid or allotetraploid (2n = 48) with BB, CC, EE, FF, GG, BBCC, CCDD, HHJJ, and KKLL genomes (Khush 1997; Ge et al. 1999; Vaughan et al. 2003; Lu et al. 2009; Sanchez et al. 2013). The wild relatives of rice have long been recognized as a rich source of novel genes underlying traits of agricul- tural importance (Jena and Khush 1990). Earlier attempts to transfer useful genes from wild to cultivated rice have been confined to using only the wild relatives with AA genome via conventional breeding methods. The F 1 hybrids that are derived from such crosses exhibit regular chromosome pair- ing and recombination, thus facilitating an easy transfer of genes into cultivated rice (Khush et al. 1990; Brar and Khush 1997; McCouch et al. 2007). In contrast, F 1 hybrids between cultivated rice and wild species with genomes other than AA are often difficult to produce. Incompatibility barriers and nonhomologous chromosome pairing result in low crossabil- ity and abortion of hybrid embryos. However, male-sterile Journal of Heredity Advance Access published June 17, 2014 at University of Connecticut on June 21, 2014 http://jhered.oxfordjournals.org/ Downloaded from
Transcript of Molecular Analysis of Oryza latifolia Desv. (CCDD Genome)-Derived Introgression Lines and...

Journal of Hereditydoi:10.1093/jhered/esu032
© The American Genetic Association. 2014. All rights reserved. For permissions, please e-mail: [email protected]
1
Molecular Analysis of Oryza latifolia Desv. (CCDD Genome)-Derived Introgression Lines and Identification of Value-Added Traits for Rice (O. sativa L.) ImprovementRosalyn B. angeles-shim, Ricky B. VinaRao, BalRam maRathi, and kshiRod k. Jena
From the Novel Gene Resources Laboratory, Plant Breeding, Genetics, and Biotechnology Division, International Rice Research Institute, DAPO Box 7777, Metro Manila, Philippines (Angeles-Shim, Vinarao, Marathi, and Jena); and the Bioscience and Biotechnology Center, Nagoya University, Chikusa, Nagoya, Aichi 464-8601, Japan (Angeles-Shim).
Address correspondence to Kshirod K. Jena at the address above, or e-mail: [email protected].
AbstractOryza latifolia is a tetraploid wild Oryza species with a CCDD genome that has been reported to harbor resistance to bacterial blight (BB), brown planthopper, and whitebacked planthopper. Aside from these traits, O. latifolia is also being tapped as a new source of resistance to lodging and high biomass production. To explore the genetic potential of O. latifolia as a novel genetic resource for the improvement of existing O. sativa cultivars, 27 disomic derivatives of O. latifolia monosomic alien addition lines (MAAL) were characterized for alien chromosome segment introgressions and evaluated for yield components, BB resistance, and strong stem characteristics. A total of 167 simple sequence repeat, sequence tagged site, and single nucleotide polymor-phism markers, along with newly developed indel markers that were specifically designed to detect O. latifolia chromosome seg-ment introgressions in an O. sativa background, were used to define alien introgressions in 27 disomics derived from O. latifolia MAALs. Genotype data showed that 32 unique introgressions spanning 0.31–22.73 Mb were introgressed in different combina-tions in each of the 27 disomic derivatives. Evaluation of the disomic derivatives for agronomic traits identified lines with puta-tive QTLs for resistance to Philippine races 3A, 4, 9A, and 9D of BB. Putative quantitative trait loci (QTLs) conferring strong stem in 19 out of the 27 disomic derivatives studied were also identified from O. latifolia introgressions on chromosome 6.Subject areas: Genomics and gene mapping; Conservation genetics and biodiversity
Key words: disomics, DNA markers, lodging resistance, monosomic alien addition lines, wild introgression, Xanthomonas oryzae pv. oryzae
The genus Oryza is relatively small but complex, consisting of 2 cultivated species, Oryza sativa and O. glaberrima, and 22 well-recognized wild relatives (Ge et al. 1999; Zeigler 2013). Based on classical taxonomy and isozyme, restric-tion fragment length polymorphism (RFLP) and sequence analysis, the members of this genus are grouped into 4 species complexes, namely, O. sativa, O. officinalis, O. ridleyi, and O. granulata. Species belonging to the O. sativa complex, including both cultivated species, are diploids (2n = 24) with AA genome. The more distantly related species are either diploid or allotetraploid (2n = 48) with BB, CC, EE, FF, GG, BBCC, CCDD, HHJJ, and KKLL genomes (Khush 1997; Ge et al. 1999; Vaughan et al. 2003; Lu et al. 2009; Sanchez et al. 2013).
The wild relatives of rice have long been recognized as a rich source of novel genes underlying traits of agricul-tural importance (Jena and Khush 1990). Earlier attempts to transfer useful genes from wild to cultivated rice have been confined to using only the wild relatives with AA genome via conventional breeding methods. The F1 hybrids that are derived from such crosses exhibit regular chromosome pair-ing and recombination, thus facilitating an easy transfer of genes into cultivated rice (Khush et al. 1990; Brar and Khush 1997; McCouch et al. 2007). In contrast, F1 hybrids between cultivated rice and wild species with genomes other than AA are often difficult to produce. Incompatibility barriers and nonhomologous chromosome pairing result in low crossabil-ity and abortion of hybrid embryos. However, male-sterile
Journal of Heredity Advance Access published June 17, 2014 at U
niversity of Connecticut on June 21, 2014
http://jhered.oxfordjournals.org/D
ownloaded from

Journal of Heredity
2
interspecific F1 hybrids could be produced by embryo rescue (Jena and Khush 1984; Brar and Singh 2011). These hybrids were used as female parents in subsequent backcrossing to produce fertile progenies with normal diploid chromosome complements (2n = 24 or 2n = 24 + 1) (Jena 2010).
Rice plants carrying the normal chromosome comple-ment of O. sativa and an extra chromosome from any wild species (2n + 1) are known as monosomic alien addition lines (MAALs). MAALs are important genetic stocks for broad-ening the existing gene pool of rice. They are useful in the assignment of alien traits to specific chromosomes of the wild relatives and in transferring genes of economic impor-tance from wild species to cultivated rice (Brar and Khush 1997; Jena 2010). MAALs can also provide a convenient way of dissecting wild rice genomes into individual chromosome entities in a functional genomic background. To date, MAALs of O. sativa, trisomic plants carrying single additional chro-mosome, from 1 of 6 different wild rice species have been established. These sets of MAALs include a complete set of MAALs for chromosomes 1–12 of O. officinalis (Jena and Khush 1989) and O. australiensis (Jena et al. 1991); MAALs for 8 chromosomes of O. minuta (Amante-Bordeos et al. 1992); and MAALs for 6, 7, and 11 chromosomes of O. granulata, O. brachyantha, and O. latifolia, respectively (Multani et al. 2003).
The disomics (2n) segregate in the progenies of MAALs. The disomics are fertile backcross progenies carrying the normal diploid chromosome complement of O. sativa and limited chromosome introgressions from the wild parent. These progenies mostly resemble the O. sativa parent and are usually recovered in selfed progenies of MAALs. The extremely limited homeologous pairing between the chro-mosomes of cultivated rice and its wild relatives has been proposed to facilitate the rapid recovery of the recurrent phenotype (Stephens 1949; Rick 1971; Jena et al. 1992). Like the MAALs, disomics constitute a valuable genetic resource for the identification of novel genes that control important agronomic traits. Continuous selfing of these disomics pro-duces advanced introgression lines that carry short chromo-somal segments from the wild relative. The uniform genetic background of these introgression lines allows for easy association of a phenotype with the introgressed chromo-some segment from the wild relative. Because of this, QTLs, even those with only minor effects, are easily detected. The uniform genetic background of MAAL-derived disomic introgression lines also provides an advantage in identifying genes/QTLs using simple statistical analysis.
O. latifolia is a tetraploid (CCDD) wild Oryza species endemic to South and Central America. The species is char-acteristically perennial, with tall, erect plants having broad foliage and high biomass, hard stem, and long, lax panicles with a large number of spikelets. It can grow in a wide range of ecosystems, including swamps, savannas, woodlands, hill slopes, and coastal belts (Ying and Song 2003). O. latifolia has been reported to be an important source of resistance to bac-terial leaf blight (BB), brown planthopper, and whitebacked planthopper (Multani et al. 2003) and is being explored for high biomass production and lodging resistance (Jena KK, unpublished data).
MAALs of O. latifolia and their disomic derivatives have been established at the International Rice Research Institute (IRRI) since 2000. However, because of the lack of available DNA markers that are specific for O. latifolia, molecular char-acterization of alien introgressions in disomic lines has not been possible and therefore they cannot be used in breeding programs.
This study aims to define alien introgressions in disomic derivatives of O. latifolia in the genetic background of O. sativa. The simple sequence repeats (SSRs), single nucleotide poly-morphisms (SNPs), and newly developed indel markers that amplify distinctive regions in both O. sativa and O. latifolia genomes were used to identify the wild chromosome segment introgressions in the disomic derivatives. Agronomic evalua-tion of these introgression lines identified value-added traits for yield and yield components and disease resistance for the improvement of existing rice cultivars. Experimental validation of culm strength in the disomic derivatives also identified lines with potential to improve lodging resistance in cultivated rice.
Materials and MethodsPlant Materials
Twenty-seven introgression lines derived from the MAALs 4, 5, 6, 7, 8, 9, 10, 11, and 12 of O. latifolia were selected from a total of 216 plants and used in this study (Table 1). No MAALs for chromosomes 1, 2, and 3 of O. latifolia were available. The MAALs were developed by successive back-crossing of the allotriploid F1 hybrid derived from a cross between an O. sativa elite breeding line (IR31917-45-3-2) and O. latifolia (IRGC 100914) to the O. sativa parent. All intro-gression lines have 2n = 24 and carry a limited number of chromosome segments from O. latifolia (Multani et al. 2003). The breeding scheme used for the development of the intro-gression lines from the O. latifolia MAALs is presented in Figure 1.
Development of O. latifolia-Specific Primers and Detection of Alien Introgression in Disomics
A total of 843 O. sativa-based markers that include 293 SSRs, 166 sequence tagged site (STS), and 384 SNPs were used to determine the extent and location of O. latifolia chromosome segment introgressions in 27 disomic derivatives of O. latifo-lia. The 384-plex SNP set used was a GoldenGate VeraCode oligo pool assay (OPA; VC0013033 OPA) designed for the Illumina BeadXpress Reader (Illumina Inc., San Diego, CA) to differentiate among indica/indica populations (Thomson et al. 2012). For a fuller coverage of the genome, additional indel-based markers were designed based on the existing sequence data for O. sativa and the CCDD genome species O. alta (www.ncbi.nlm.nih.gov; 5 February 2012). Sequence information for O. latifolia is still unavailable. Since O. alta shares the same CCDD genome as O. latifolia, the available bacterial artificial chromosome (BAC) sequences for O. alta were used. Sequence alignment of a total of 127 784 BAC end sequences of O. alta (IRGC 105143) to v7 of the genomic
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
3
sequence of O. sativa subsp. japonica cv. Nipponbare from TIGR-MSU was kindly provided by Professor R. Wing of the Oryza Map Alignment Project (OMAP) of the Arizona Genomics Institute of the University of Arizona, Tuscon, AZ (Ammiraju et al. 2010; Jacquemin et al. 2013). The align-ments were generated using MEGABLAST, an alignment
tool specifically designed to find long alignments between similar sequences, and hence it is the best tool in finding an identical match to any query sequences. Using these align-ments, 35 indel-based markers were designed and validated for their usefulness in differentiating between O. latifolia and O. sativa genomes.
Table 1 Classification of the 27 introgression lines derived from Oryza latifolia based on alien introgression patterns
O. latifolia MAAL source Disomic derivativeIntrogressed chromosome fragment from O. latifolia Introgression type
MAAL 4 WH12-2236 1, 2, 3, 4, 6, 9, 11 MAAL4IWH12-2237 1, 2, 3, 4, 6, 9, 11 MAAL4IWH12-2238 1, 2, 3, 4, 6, 9, 11 MAAL4IWH12-2239 1, 2, 3, 4, 6, 9, 11 MAAL4I
MAAL 5 WH12-2240 4, 5, 6, 8, 12 MAAL5IWH12-2241 4, 5, 6, 8, 12 MAAL5IWH12-2242 4, 5, 6, 8, 12 MAAL5IWH12-2243 4, 5, 6, 8, 12 MAAL5IWH12-2244 4, 5, 6, 8, 12 MAAL5IWH12-2245 4, 5, 6, 8, 12 MAAL5IWH12-2246 4, 5, 6, 8, 12 MAAL5IWH12-2247 4, 5, 6, 8, 12 MAAL5I
MAAL 6 WH12-2248 6, 8 MAAL6IWH12-2249 6, 8 MAAL6IWH12-2250 6, 8, 10 MAAL6IIWH12-2251 6, 8, 10 MAAL6II
MAAL 7 WH12-2252 1, 2, 3, 4, 5, 6, 8, 9 MAAL7IWH12-2253 1, 3, 4, 6, 8, 9, 10, 11 MAAL7IIWH12-2254 1, 3, 4, 6, 8, 9, 10, 11 MAAL7II
MAAL8 WH12-2255 1, 2, 6, 8, 9, 10, 12 MAAL8IWH12-2256 1, 2, 4, 6, 8, 9, 10, 12 MAAL8II
MAAL 9 WH12-2257 4, 6, 8, 10 MAAL9IMAAL 10 WH12-2258 1, 6, 8, 10 MAAL10I
WH12-2259 6, 8, 10 MAAL10IIMAAL 11 WH12-2260 6, 8, 10 MAAL11IMAAL 12 WH12-2261 6, 8, 10 MAAL12I
WH12-2262 6, 8, 10 MAAL12I
Figure 1. Breeding scheme for the production of Oryza latifolia introgression lines.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
4
Agronomic Evaluation of the Disomic Derivatives of O. latifolia for Value-Added Traits
Phenotypic evaluation of the 27 introgression lines was car-ried out during the dry and wet seasons of 2012 at the IRRI in Los Baños, Laguna, Philippines. Twenty-four seedlings of each line, along with the O. sativa elite breeding line IR31917-45-3-2 as a control, were grown to maturity in double-row plots in the field. The seedlings were first germinated in seed-beds and 21-day-old seedlings were transplanted in field plots at 20 × 20-cm spacing.
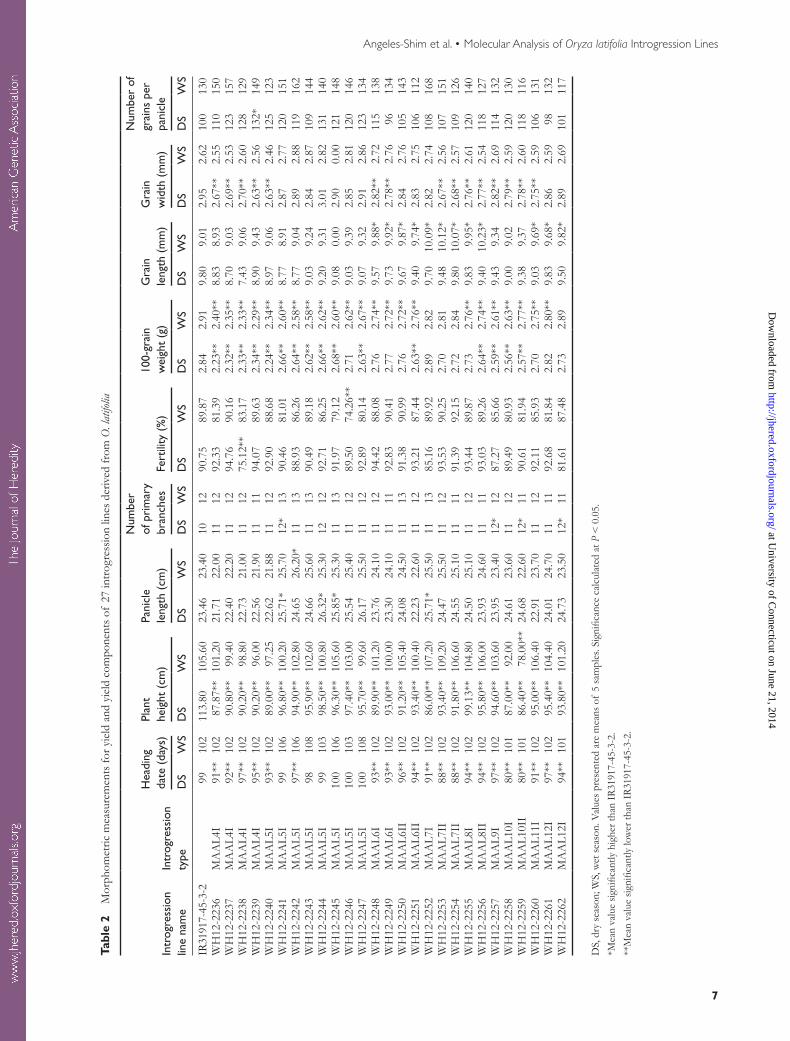
At maturity, 5 healthy plants were selected from each of the introgression lines and phenotyped for plant height, pani-cle length, number of primary branches per panicle, grain number per panicle, grain length and width, 100-grain weight, and fertility. The number of days to heading was determined from the time of sowing up to the actual flowering of 50% of the plants for each line. Data on 100-grain weight were recorded after air drying of seeds in the glasshouse for 7 days to achieve a moisture content of 13–14%.
The selection of the 27 introgression lines for inclu-sion in this study was based on field evaluation for strong stem. To verify the physical strength of the stems, the sec-tion modulus (SM), a parameter that measures the physical strength of the culm and that is directly influenced by culm morphology (i.e., diameter and wall thickness), was calcu-lated for each of the test materials. Twenty days after head-ing, 5 main culms from each line were sampled and fixed in 1:1:18 FAA (35% formalin:glacial acetic acid:70% alcohol). After a series of washes, tissue samples were sectioned and stained for 4 min with phloroglucinol-hydrochloric acid. Cross-sections of the third internodes of each culm were viewed under an Axioplan stereomicroscope and images were captured using an Olympus DP71 camera. Processing of the captured images for dimensional measurements was carried out using the ImagePro (ver. 7) software. SM was calculated using the formula SM = π/32 × (a1
3b1 − a2
3b2)/a1, where a1 is the outer diameter of the minor axis in an oval cross-section, b1 is the outer diameter of the major axis in an oval cross-section, a2 is the inner diameter of the minor axis in an oval cross-section and b2 is the inner diam-eter of the major axis in an oval cross-section following Ookawa et al. (2010).
Each of the introgression lines, along with IR31917-45-3-2 as a control, was screened for reaction to BB caused by Xanthomonas oryzae pv. oryzae (Xoo) in the screenhouse during the wet season of 2012. The resistance reaction of selected lines during the wet season screening was con-firmed during the dry season of 2013. The inocula of the 10 Philippine races of the pathogen, PXO61 (race 1), PXO86 (race 2), PXO79 (race 3A), PXO340 (race 3B), PXO71 (race 4), PXO112 (race 5), PXO99 (race 6), PXO145 (race 7), PXO280 (race 8), PXO339 (race 9A), PXO349 (race 9B), PXO347 (race 9C), PXO363 (race 9D), and PXO341 (race 10), were prepared by suspending the bacterial mass in sterile water at approximately 109 cells/ml. Inoculation was carried out during the maximum tillering stage of the plants fol-lowing the leaf clipping method of Kauffman et al. (1973).
Scoring for the disease reaction of the test materials 14 days after inoculation was based on lesion length: <3 cm = resist-ant, 3.1–5 cm = moderately resistant, and >5 cm = suscepti-ble (Jeung et al. 2006; Suh et al. 2013).
Statistical Analysis and Detection of Quantitative Trait Loci for Agronomic Traits
A one-way Anova and comparison of means using Dunnett’s multiple comparisons test at 95% confidence interval (P < 0.05) were performed. A probability of 0.005 was used as the threshold for the detection of putative QTLs control-ling any of the traits examined. If the calculated mean value for a trait in any introgression line was significantly different from that of IR31917-45-3-2, the existence of a QTL was considered and assigned in the introgressed segment from the wild donor.
ResultsMolecular Characterization of O. latifolia Introgression in an O. sativa Background
Of the 878 molecular markers used to genotype the 27 intro-gression lines, 169 (38 SSRs, 17 STS, 85 SNPs, and 29 indels) markers detected polymorphism between O. sativa and O. lati-folia. These markers were distributed across the 12 chromo-somes, providing genomic coverage at an average interval of 2.25 Mb. Markers targeting an approximately 12-Mb region spanning the short arm up to the distal part of the long arm of chromosome 10 of O. sativa were not informative when used to amplify targets in both IR31917-45-3-2 and O. latifo-lia, leaving this region on chromosome 10 uncharacterized for O. latifolia introgression (Figure 2).
Genotyping of the disomic derivatives identified a total of 32 unique introgressions of O. latifolia chromosome seg-ments ranging from 0.31 Mb on chromosome 3 to 22.73 Mb on chromosome 6. Based on the different combinations of these wild chromosome segment introgressions, 12 distinct patterns of O. latifolia introgression were observed. Two types of introgression patterns were observed for the lines derived from MAALs 6, 7, 8, and 10, whereas only a single introgres-sion pattern each was observed for the lines derived from MAALs 4, 5, 9, 11, and 12. Introgression lines from MAAL 4 recorded the highest number (11) of introgressions, whereas the derivatives of MAAL 6I recorded the lowest number (2) of wild introgressions (Table 1, Figure 3). Lines belong-ing to MAAL 10II and MAAL 11I had the same introgres-sion patterns. All introgression lines carried different sizes of O. latifolia chromosome segment on chromosome 6, and the introgression lines belonging to MAAL 4I and MAAL 12I harbored O. latifolia segments measuring up to 9.42 Mb. Introgression lines belonging to MAAL 7I carried O. latifolia introgressions covering 22.73 Mb on the same chromosome, whereas the rest of the lines harbored an O. latifolia segment that spanned 22.23 Mb. There was no O. latifolia chromosome segment introgression on the chromosome 7 of the recur-rent parent genome (Figure 3).
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
5
Phenotypic Evaluation of the Introgression Lines
Examination of gross plant morphology showed a close resemblance of the 27 introgression lines to the O. sativa par-ent. None of the undesirable traits of the O. latifolia parent, particularly the tall stature and spreading growth habit, were observed in any of the test materials (Figure 3). Data on the morphometric measurements for different agronomic traits of the 27 introgression lines are presented in Table 2.
Heading date during the dry season was significantly earlier in the introgression lines (80–97 days) than in IR31917-45-3-2 (99 days), except for the 5 lines (WH12-2241, WH12-2243, WH12-2244, WH12-2245, and WH12-2246) that recorded increased number of days to heading (98–100 days) than the control. In the wet season, no significant differences were observed in the heading date of the introgression lines and IR31917-45-3-2.
On average, the introgression lines had a significantly shorter stature (92.9 cm) during the dry season compared with IR31917-45-3-2 with a recorded mean plant height of
99.00 cm. The mean values for panicle length were higher in lines of introgression types MAAL 5I, MAAL 71, MAAL 7II, MAAL 8I, MAAL 8II, MAAL 9I, MAAL 10I, MAAL 10II, and MAAL 12I than in IR31917-45-3-2 during both the dry and wet seasons except for line WH12-2259 of the introgression type MAAL 10II, which recorded shorter pani-cles than IR31917-45-3-2. Significantly longer panicles were observed in lines WH12-2241, WH12-2244, WH12-2245, and WH12-2252 during the dry season (25.71–25.85 cm) and line WH12-2242 during the wet season (26.20 cm).
Alien introgressions from O. latifolia had no significant effects on the number of primary branches per panicle of the test materials during the wet season. However, during the dry season, the lines WH12-2241, WH12-2257, WH12-2259, and WH12-2262 recorded 20% more primary branches than IR31917-45-3-2.
The spikelet fertility of the introgression lines and IR31917-45-3-2 did not show significant differences except the line, WH12-2238 of the introgression type MAAL 4I and the line WH12-2246 of the introgression type MAAL
Figure 2. Molecular markers used for the whole-genome survey of the 27 introgression lines derived from O. latifolia MAAL disomics. Numbers on the left indicate the physical distance (Mb) of each marker (right) along the length of each chromosome. Arrowheads represent the position of the centromere on each chromosome. Marker identification is as follows: rice microsatellite (RM) = SSRs, id and fd = SNPs (blue), Lat = indel-based markers (green) and S = STS.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
6
Figure 3. Morphology and genotype of the plant materials. Gross morphology and graphical genotype of the O. sativa elite breeding line IR31917-43-2, O. latifolia (IRGC 100914), and representative plants from each of the 12 introgression types identified from the 27 introgression lines derived from O. latifolia disomics. Red circles indicate the 32 unique O. latifolia chromosome segments carried by the introgression lines.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from
Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
7
Tabl
e 2
Mor
phom
etric
mea
sure
men
ts fo
r yie
ld a
nd y
ield
com
pone
nts o
f 27
intro
gres
sion
lines
der
ived
from
O. l
atifo
lia
Intr
ogre
ssio
n
line
nam
eIn
trog
ress
ion
ty
pe
Hea
ding
da
te (
days
)Pl
ant
he
ight
(cm
)Pa
nicl
e
leng
th (
cm)
Num
ber
of p
rim
ary
bran
ches
Fert
ility
(%
)10
0-gr
ain
w
eigh
t (g
)G
rain
le
ngth
(m
m)
Gra
in
wid
th (
mm
)
Num
ber
of
grai
ns p
er
pani
cle
DS
WS
DS
WS
DS
WS
DS
WS
DS
WS
DS
WS
DS
WS
DS
WS
DS
WS
IR31
917-
45-3
-299
102
113.
8010
5.60
23.4
623
.40
1012
90.7
589
.87
2.84
2.91
9.80
9.01
2.95
2.62
100
130
WH
12-2
236
MA
AL4
I91
**10
287
.87*
*10
1.20
21.7
122
.00
1112
92.3
381
.39
2.23
**2.
40**
8.83
8.93
2.67
**2.
5511
015
0W
H12
-223
7M
AA
L4I
92**
102
90.8
0**
99.4
022
.40
22.2
011
1294
.76
90.1
62.
32**
2.35
**8.
709.
032.
69**
2.53
123
157
WH
12-2
238
MA
AL4
I97
**10
290
.20*
*98
.80
22.7
321
.00
1112
75.1
2**
83.1
72.
33**
2.33
**7.
439.
062.
70**
2.60
128
129
WH
12-2
239
MA
AL4
I95
**10
290
.20*
*96
.00
22.5
621
.90
1111
94.0
789
.63
2.34
**2.
29**
8.90
9.43
2.63
**2.
5613
2*14
9W
H12
-224
0M
AA
L5I
93**
102
89.0
0**
97.2
522
.62
21.8
811
1292
.90
88.6
82.
24**
2.34
**8.
979.
062.
63**
2.46
125
123
WH
12-2
241
MA
AL5
I99
106
96.8
0**
100.
2025
.71*
25.7
012
*13
90.4
681
.01
2.66
**2.
60**
8.77
8.91
2.87
2.77
120
151
WH
12-2
242
MA
AL5
I97
**10
694
.90*
*10
2.80
24.6
526
.20*
1113
88.9
386
.26
2.64
**2.
58**
8.77
9.04
2.89
2.88
119
162
WH
12-2
243
MA
AL5
I98
108
95.9
0**
102.
6024
.66
25.6
011
1390
.49
89.1
82.
62**
2.58
**9.
039.
242.
842.
8710
914
4W
H12
-224
4M
AA
L5I
9910
398
.50*
*10
0.80
26.3
2*25
.30
1212
92.7
186
.25
2.66
**2.
62**
9.20
9.31
3.01
2.82
131
140
WH
12-2
245
MA
AL5
I10
010
696
.30*
*10
5.60
25.8
5*25
.30
1113
91.9
779
.12
2.68
**2.
60**
9.08
0.00
2.90
0.00
121
148
WH
12-2
246
MA
AL5
I10
010
397
.40*
*10
3.00
25.5
425
.40
1112
89.5
074
.26*
*2.
712.
62**
9.03
9.39
2.85
2.81
120
146
WH
12-2
247
MA
AL5
I10
010
895
.70*
*99
.60
26.1
725
.50
1112
92.8
980
.14
2.63
**2.
67**
9.07
9.32
2.91
2.86
123
134
WH
12-2
248
MA
AL6
I93
**10
289
.90*
*10
1.20
23.7
624
.10
1112
94.4
288
.08
2.76
2.74
**9.
579.
88*
2.82
**2.
7211
513
8W
H12
-224
9M
AA
L6I
93**
102
93.0
0**
100.
0023
.30
24.1
011
1192
.83
90.4
12.
772.
72**
9.73
9.92
*2.
78**
2.76
9613
4W
H12
-225
0M
AA
L6II
96**
102
91.2
0**
105.
4024
.08
24.5
011
1391
.38
90.9
92.
762.
72**
9.67
9.87
*2.
842.
7610
514
3W
H12
-225
1M
AA
L6II
94**
102
93.4
0**
100.
4022
.23
22.6
011
1293
.21
87.4
42.
63**
2.76
**9.
409.
74*
2.83
2.75
106
112
WH
12-2
252
MA
AL7
I91
**10
286
.00*
*10
7.20
25.7
1*25
.50
1113
85.1
689
.92
2.89
2.82
9.70
10.0
9*2.
822.
7410
816
8W
H12
-225
3M
AA
L7II
88**
102
93.4
0**
109.
2024
.47
25.5
011
1293
.53
90.2
52.
702.
819.
4810
.12*
2.67
**2.
5610
715
1W
H12
-225
4M
AA
L7II
88**
102
91.8
0**
106.
6024
.55
25.1
011
1191
.39
92.1
52.
722.
849.
8010
.07*
2.68
**2.
5710
912
6W
H12
-225
5M
AA
L8I
94**
102
99.1
3**
104.
8024
.50
25.1
011
1293
.44
89.8
72.
732.
76**
9.83
9.95
*2.
76**
2.61
120
140
WH
12-2
256
MA
AL8
II94
**10
295
.80*
*10
6.00
23.9
324
.60
1111
93.0
389
.26
2.64
**2.
74**
9.40
10.2
3*2.
77**
2.54
118
127
WH
12-2
257
MA
AL9
I97
**10
294
.60*
*10
3.60
23.9
523
.40
12*
1287
.27
85.6
62.
59**
2.61
**9.
439.
342.
82**
2.69
114
132
WH
12-2
258
MA
AL1
0I80
**10
187
.00*
*92
.00
24.6
123
.60
1112
89.4
980
.93
2.56
**2.
63**
9.00
9.02
2.79
**2.
5912
013
0W
H12
-225
9M
AA
L10I
I80
**10
186
.40*
*78
.00*
*24
.68
22.6
012
*11
90.6
181
.94
2.57
**2.
77**
9.38
9.37
2.78
**2.
6011
811
6W
H12
-226
0M
AA
L11I
91**
102
95.0
0**
106.
4022
.91
23.7
011
1292
.11
85.9
32.
702.
75**
9.03
9.69
*2.
75**
2.59
106
131
WH
12-2
261
MA
AL1
2I97
**10
295
.40*
*10
4.40
24.0
124
.70
1111
92.6
881
.84
2.82
2.80
**9.
839.
68*
2.86
2.59
9813
2W
H12
-226
2M
AA
L12I
94**
101
93.8
0**
101.
2024
.73
23.5
012
*11
81.6
187
.48
2.73
2.89
9.50
9.82
*2.
892.
6910
111
7
DS,
dry
seas
on; W
S, w
et se
ason
. Val
ues p
rese
nted
are
mea
ns o
f 5
sam
ples
. Sig
nific
ance
cal
cula
ted
at P
< 0
.05.
*Mea
n va
lue
signi
fican
tly h
ighe
r tha
n IR
3191
7-45
-3-2
.**
Mea
n va
lue
signi
fican
tly lo
wer
than
IR31
917-
45-3
-2.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
8
5I which recorded 17.2% and 17.4% lower fertility than the recurrent parent during the dry and wet season, respectively.
One hundred grain weight in all the introgression lines was lower than that of IR31917-45-3-2 during both the sea-sons except for the line WH12-2252 of the introgression type MAAL 7I, which recorded a higher mean 100-grain weight than IR31917-45-3-2 (2.84 g) during the dry season (Table 2).
Differences in the mean values for grain length during the dry season and grain width during the wet season were not significant between the introgression lines and IR31917-45-3-2. However, lines of the introgression types MAAL 6I, MAAL 6II, MAAL 7I, MAAL 7II, MAAL 8I, MAAL 8II, MAAL 11I, and MAAL 12I recorded significantly longer grains (9.74–10.23 mm) than IR31917-45-3-2 (9.01 mm) dur-ing the wet season. The mean values for grain width of the introgression lines during the dry season were lower than that of IR31917-45-3-2 (2.95 mm), except for line WH12-2244 of the introgression type MAAL 5I, which recorded an average grain width of 3.01 mm.
The number of grains per panicle for the introgression lines was not significantly different from that of IR31917-45-3-2 during both the dry and wet seasons except for line WH12-2239, which registered 32% more grains than the recurrent parent during the dry season.
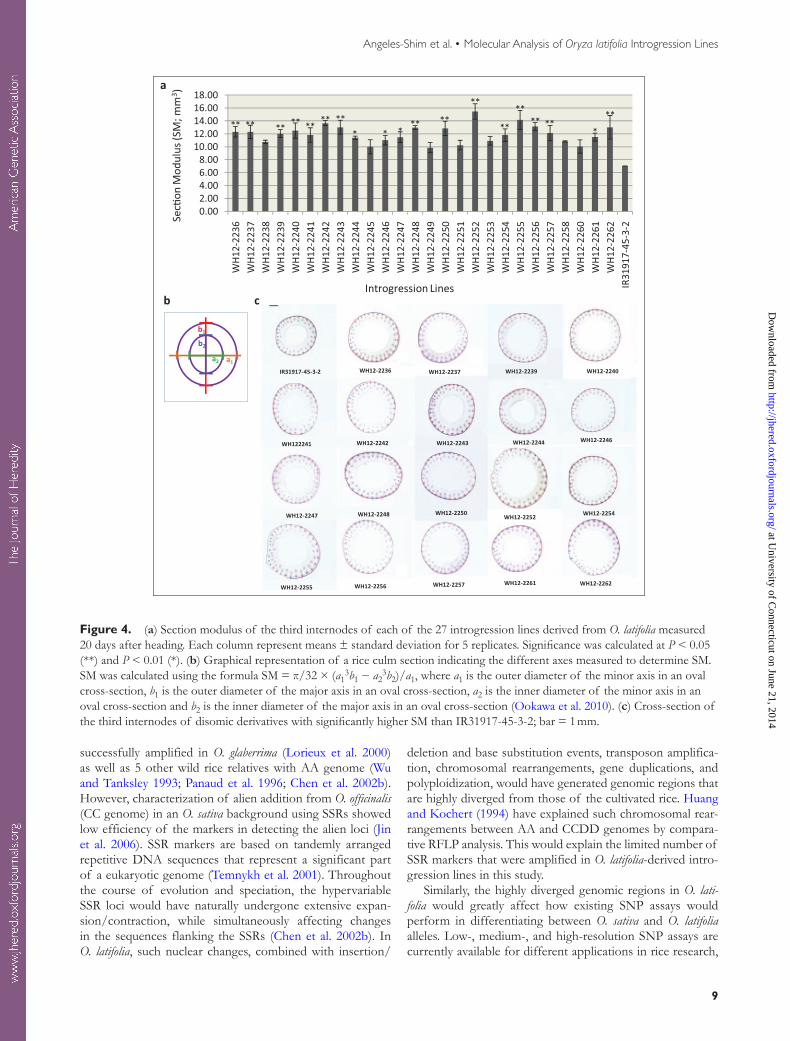
The introgression lines used in this study were selected based on field evaluation for strong stem. SM analysis showed that all introgression lines had a higher SM than IR31917-45-3-2. Measurements were not taken for WH12-2259 due to the poor condition of the fixed samples. Of the remaining 26 introgression lines, 19 exhibited significant increases in SM ranging from 59.5% to 116.5% compared with the mean SM value observed for IR31917-45-3-2 (Figure 4a). Examination of the fourth internode cross-sections showed that all the 19 lines had a wider culm diameter than IR31917-45-3-2 (Figure 4b).
BB Resistance of the O. latifolia Introgression Lines
The disease reaction of IR31917-45-3-2 to different races of Xoo used in this study ranged from resistant to moder-ately resistant to susceptible based on the length of lesions in the leaf caused by the pathogen 14 days after inoculation (Table 3). IR31917-45-3-2 showed resistance to races 1, 5, 7, 8, and 10, with lesion lengths ranging from 1.17 to 2.83 cm, and moderate resistance to races 2 and 6, with lesion lengths of 3.67–4.00 cm. IR31917-45-3-2 was susceptible to races 3A, 3B, 4, 9A, 9B, 9C, and 9D, which caused 6.17–13.93-cm lesion length on the leaves. A wide range of reactions to the disease was similarly observed among the introgres-sion lines. Like IR31917-45-3-2, all the test materials were susceptible to races 3B, 9B, and 9C of BB but resistant to moderately resistant to races 1, 5, 7, and 8 of the pathogen. Most of the introgression lines also shared the suscepti-ble reaction of IR31917-43-2 to races 3A, 4, 9A, and 9D. However, some introgression lines showed resistance to these races. In particular, lines WH12-2255 and WH12-2256 showed strong BB resistance, with lesion lengths of only 1.83 and 1.67 cm, respectively, compared with the 12.00-cm
lesions in IR31917-45-3-2 inoculated with race 9A of the pathogen. Both lines have unique introgressions of chromo-some segments from O. latifolia on chromosome 12 span-ning 13.96 Mb. Introgression line WH12-2246 also exhibited resistance to race 9D, recording an average lesion length of 4.00 cm, whereas the rest of the introgression lines remained susceptible like the recurrent parent, O. sativa. Additionally, introgression lines WH12-2236, WH12-2237, WH12-2238, WH12-2240, WH12-2241, WH12-2247, WH12-2254, and WH12-2255 showed moderate resistance to race 3A, whereas lines WH12-2236, WH12-2237, WH12-2239, WH12-2240, WH12-2241, WH12-2244, WH12-2246, WH12-2247, and WH12-2259 showed moderate resistance to race 4 of the pathogen.
DiscussionTargeted Markers for Evaluating Wild Rice Derivatives
MAALs and MAAL-derived disomic introgression lines rep-resent an important genetic resource for both genetic and genomic studies. Molecular characterization of alien intro-gressions in these lines is particularly essential in the study of both single gene and quantitative trait inheritance from wild Oryza species. Isozymes, RFLPs, and genome-specific clones developed via representational difference analysis have been used to characterize alien introgressions in MAALs and their disomic derivatives (Huang and Kochert 1994; Jena et al. 1994; Multani et al 1994; Shim et al. 2010). However, the inherent limitations of each marker had restricted the char-acterization of wild introgressions in these materials, and consequently their application in breeding programs.
A genomic survey using 843 O. sativa-based markers in the form of SSRs, STS, and SNPs was carried out to iden-tify markers that can differentiate between O. latifolia and O. sativa alleles. Of these markers, only 16.5% (139) detected polymorphic targets in O. latifolia and O. sativa, with SNPs being the most informative (21.88% polymorphic), followed by the SSRs (12.97% polymorphic) and the STS (10.24% polymorphic). In comparison, indel-based markers that were designed based on sequence alignments between O. sativa cv. Nipponbare and O. alta exhibited 83% polymorphism.
SSRs have long been used as a marker of choice for appli-cations in plant breeding and genetic studies not only in rice but in other crops, including wheat (Röder et al. 1995), barley (Saghai Maroof et al. 1994), soybean (Maughan et al. 1995), tomato (Smulders et al. 1997), and corn (Senior and Heun 1993). However, a significant number of SSRs have rarely been transferred across species and used in alien introgres-sion studies (Röder et al. 1995). At best, the interspecific transferability of SSRs had been unpredictable. Studies on Hordeum chilense introgression in wheat showed that SSRs were suitable in identifying alien loci (Hernández et al. 2002), whereas reports on the introgression of Lophopyrum elon-gatum in wheat showed the genome specificity of SSRs and, consequently, their low transferability across related species (Mullan et al. 2005). In rice, diversity studies have shown that a high proportion of SSRs designed for O. sativa were
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from
Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
9
successfully amplified in O. glaberrima (Lorieux et al. 2000) as well as 5 other wild rice relatives with AA genome (Wu and Tanksley 1993; Panaud et al. 1996; Chen et al. 2002b). However, characterization of alien addition from O. officinalis (CC genome) in an O. sativa background using SSRs showed low efficiency of the markers in detecting the alien loci (Jin et al. 2006). SSR markers are based on tandemly arranged repetitive DNA sequences that represent a significant part of a eukaryotic genome (Temnykh et al. 2001). Throughout the course of evolution and speciation, the hypervariable SSR loci would have naturally undergone extensive expan-sion/contraction, while simultaneously affecting changes in the sequences flanking the SSRs (Chen et al. 2002b). In O. latifolia, such nuclear changes, combined with insertion/
deletion and base substitution events, transposon amplifica-tion, chromosomal rearrangements, gene duplications, and polyploidization, would have generated genomic regions that are highly diverged from those of the cultivated rice. Huang and Kochert (1994) have explained such chromosomal rear-rangements between AA and CCDD genomes by compara-tive RFLP analysis. This would explain the limited number of SSR markers that were amplified in O. latifolia-derived intro-gression lines in this study.
Similarly, the highly diverged genomic regions in O. lati-folia would greatly affect how existing SNP assays would perform in differentiating between O. sativa and O. latifolia alleles. Low-, medium-, and high-resolution SNP assays are currently available for different applications in rice research,
Figure 4. (a) Section modulus of the third internodes of each of the 27 introgression lines derived from O. latifolia measured 20 days after heading. Each column represent means ± standard deviation for 5 replicates. Significance was calculated at P < 0.05 (**) and P < 0.01 (*). (b) Graphical representation of a rice culm section indicating the different axes measured to determine SM. SM was calculated using the formula SM = π/32 × (a1
3b1 − a23b2)/a1, where a1 is the outer diameter of the minor axis in an oval
cross-section, b1 is the outer diameter of the major axis in an oval cross-section, a2 is the inner diameter of the minor axis in an oval cross-section and b2 is the inner diameter of the major axis in an oval cross-section (Ookawa et al. 2010). (c) Cross-section of the third internodes of disomic derivatives with significantly higher SM than IR31917-45-3-2; bar = 1 mm.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
10
Tabl
e 3
Bact
eria
l lea
f bl
ight
reac
tion
of th
e 27
intro
gres
sion
lines
14
days
afte
r ino
cula
tion
with
Phi
lippi
ne ra
ces o
f X
anth
omon
as or
yzae
pv.
oryz
ae.
Intr
ogre
ssio
n
line
nam
eIn
trog
ress
ion
type
Bact
eria
l blig
ht s
trai
n
Rac
e 1
PXO
61R
ace
2 PX
O86
Rac
e 3A
PX
O79
Rac
e 3B
PX
O34
0R
ace
4 PX
O71
Rac
e 5
PXO
112
Rac
e 6
PXO
99R
ace
7 PX
O14
5R
ace
8 PX
O28
0R
ace
9A
PXO
339
Rac
e 9B
PX
O34
9R
ace
9C
PXO
0347
Rac
e 9D
PX
O36
3R
ace
10
PXO
341
IR31
917-
45-3
-22.
433.
676.
178.
336.
502.
674.
001.
171.
0712
.00
13.9
312
.17
8.67
2.83
WH
12-2
236
MA
AL4
I2.
903.
803.
679.
175.
003.
104.
731.
501.
2712
.67
11.6
713
.33
6.50
2.50
WH
12-2
237
MA
AL4
I2.
673.
834.
509.
504.
002.
004.
331.
101.
5011
.17
15.0
013
.83
11.1
72.
17W
H12
-223
8M
AA
L4I
3.43
4.13
5.00
10.1
75.
602.
674.
001.
931.
0010
.33
16.5
010
.33
11.0
02.
67W
H12
-223
9M
AA
L4I
2.50
4.03
8.43
13.5
05.
003.
675.
430.
501.
109.
5016
.33
12.5
07.
001.
17W
H12
-224
0M
AA
L5I
3.00
3.50
3.33
11.0
04.
172.
276.
670.
671.
3311
.33
13.6
713
.33
8.67
1.83
WH
12-2
241
MA
AL5
I3.
703.
673.
009.
174.
402.
173.
771.
831.
0012
.17
13.6
710
.17
10.3
35.
67W
H12
-224
2M
AA
L5I
2.17
4.73
6.67
8.33
7.77
ND
2.83
1.17
0.83
16.8
39.
5012
.17
9.50
2.17
WH
12-2
243
MA
AL5
I4.
004.
604.
5013
.67
5.67
1.67
9.17
1.17
0.83
13.8
317
.83
12.6
717
.17
1.17
WH
12-2
244
MA
AL5
I1.
175.
835.
5017
.17
4.17
1.33
9.33
1.33
0.50
13.0
0N
D11
.33
13.1
71.
33W
H12
-224
5M
AA
L5I
4.00
3.50
5.83
10.3
36.
171.
675.
170.
831.
1710
.00
12.8
312
.17
12.6
71.
00W
H12
-224
6M
AA
L5I
1.50
3.43
5.50
12.3
33.
671.
337.
171.
170.
6712
.00
18.0
07.
335.
001.
33W
H12
-224
7M
AA
L5I
1.50
2.50
4.33
11.0
03.
832.
174.
671.
330.
379.
5012
.00
8.67
11.3
31.
17W
H12
-224
8M
AA
L6I
3.17
4.83
7.00
12.6
76.
201.
675.
501.
001.
0012
.50
14.0
013
.33
13.0
01.
33W
H12
-224
9M
AA
L6I
2.67
3.33
6.17
13.8
35.
672.
8311
.00
1.17
1.00
14.5
014
.50
11.1
711
.33
2.17
WH
12-2
250
MA
AL6
II1.
673.
508.
1713
.00
5.50
2.33
9.17
1.17
1.33
15.6
712
.50
13.8
314
.33
2.17
WH
12-2
251
MA
AL6
II1.
832.
837.
3315
.33
5.17
2.17
10.3
30.
831.
3314
.67
21.3
314
.67
11.1
72.
50W
H12
-225
2M
AA
L7I
1.83
5.67
10.3
320
.33
8.67
3.17
10.8
30.
671.
3317
.67
22.0
015
.33
19.3
30.
67W
H12
-225
3M
AA
L7II
2.33
2.07
5.83
13.5
06.
171.
176.
670.
830.
5010
.83
19.3
312
.17
16.3
34.
33W
H12
-225
4M
AA
L7II
1.50
6.33
4.43
16.1
79.
000.
679.
170.
830.
2711
.17
12.8
39.
839.
502.
20W
H12
-225
5M
AA
L8I
2.73
5.17
3.50
12.8
35.
832.
5710
.67
1.33
1.17
1.83
15.6
712
.50
10.8
31.
67W
H12
-225
6M
AA
L8II
3.50
3.67
5.10
14.5
05.
431.
176.
831.
332.
331.
6714
.67
13.3
314
.00
ND
WH
12-2
257
MA
AL9
I3.
677.
879.
1717
.83
9.67
2.30
7.67
1.17
1.50
14.8
317
.50
16.5
017
.50
3.00
WH
12-2
258
MA
AL1
0I3.
008.
337.
8315
.67
7.10
1.67
7.83
1.33
0.67
15.1
716
.83
19.1
714
.33
1.67
WH
12-2
259
MA
AL1
0II
1.67
4.60
8.83
14.8
34.
671.
6715
.00
0.83
1.67
15.8
316
.00
16.0
013
.25
2.43
WH
12-2
260
MA
AL1
1I2.
177.
178.
6712
.33
6.50
1.17
2.83
1.17
1.67
12.0
013
.83
15.8
312
.63
3.17
WH
12-2
261
MA
AL1
2I2.
337.
007.
5016
.00
5.33
2.33
11.1
71.
171.
0014
.17
22.0
015
.83
14.1
72.
33W
H12
-226
2 M
AA
L12I
4.50
4.50
5.50
16.0
010
.00
2.33
11.1
71.
170.
8313
.67
18.8
317
.00
14.0
02.
17
ND
, no
data
. Val
ues p
rese
nted
are
mea
ns o
f 3
sam
ples
; <3
cm =
resis
tant
, 3.1
–5 c
m =
mod
erat
ely
resis
tant
, >5
cm =
susc
eptib
le.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
11
although the SNP discovery pools used for these assays were all based on sequences of O. sativa varieties and of a few wild relatives with AA genome (O. rufipogon, O. nivara, and O. barthii) (McNally et al. 2009; Huang et al. 2010; McCouch et al. 2010; Chen et al. 2011; Thomson et al. 2012). For this study, a low-resolution 384-plex SNP assay designed to define variation within the O. sativa subspecies indica varietal group was used. The target SNPs for this assay were particu-larly selected following a SNP discovery effort that provides information about the frequency of polymorphism within an indica germplasm. Of the 384 SNPs in the assay, only 84 suc-cessfully differentiated between O. sativa and O. latifolia. SNPs are biallelic and the individual base change that is detected as a SNP is expected to have occurred only once in evolu-tionary time. Because of this, SNPs are generally informative only for a particular set of genetic materials (McCouch et al. 2010). In the current study, the SNP assay used was specifi-cally optimized for indica populations, which explains the low polymorphism rate observed between O. sativa and O. latifolia. Our study suggests that, for a SNP assay to work for a spe-cific population, the SNP set has to be optimized first for the population to be studied.
In order to fully harness the potential of genetic resources that are derived from wild species, molecular markers that are tailored to detect alien chromosome introgressions in any given background must be developed. This will not only accelerate selection in breeding programs that involve wild rice derivatives but will also advance the identification and transfer of novel genes from wild species into culti-vated rice. To this end, the progress in the end sequencing of the BAC and construction of physical maps of 12 rep-resentative genomes of rice by the OMAP project offers an unprecedented opportunity to develop unique, genome-spe-cific molecular markers that can accurately evaluate genetic resources derived from highly diverged wild rice relatives (Ammiraju et al. 2010). The highly polymorphic indel mark-ers that were developed for this study using the sequence alignment between cv. Nipponbare and the O. alta BAC ends that were generated by the OMAP project highlight the sig-nificance of wild reference genomes toward the develop-ment of reliable markers to define wild introgressions in any given genetic background.
Characterization of the MAAL Disomic-Derived Introgression Lines
Molecular characterization of the disomic derivatives of the O. latifolia MAALs demonstrated a conserved pattern of wild genome introgression into an O. sativa background, with more than one line carrying the same set of intro-gressed chromosome segments from O. latifolia. Across introgression types, transmission of common chromosome segments was also observed. In particular, varying lengths of O. latifolia chromosome segments were systematically transferred into chromosome 6 in all the disomic deriva-tives, whereas none were transmitted into chromosome 7. In cotton, systematic chromosome or chromosome seg-ment transmission has also been reported in several studies.
Genotyping of trispecies hybrids and backcross progenies used in breeding for low gossypol content in seeds and high gossypol in plants showed that 4 AFLP markers specific to Gossypium sturtianum (C genome) were systematically present in all the backcross progenies of 2 trispecies hybrids ([G. hir-sutum × G. raimondii] × G. sturtianum and [G. raimondii × G. sturtianum] × G. hirsutum) (Vroh Bi et al. 1999). Similarly, Benbouza et al. (2007) reported the conserved transmission of 3 SSR markers specific to G. sturtianum in all selected progenies of the trispecies hybrid (G. hirsutum × G. raimon-dii) × G. sturtianum. In both cases, the higher pairing affinity of the chromosome segment of G. sturtianum harboring the specific AFLP or SSR loci with the chromosomes of the other parents resulted in the preferential transmission of those particular chromosome segments. In this study, the disomic derivatives showed a similar pattern of preferential introgression of small chromosome segments, marked by the conserved transmission patterns of the wild SSR, STS, SNP, and indel loci on specific chromosomes of O. sativa. The systemic replacement of these loci with chromosome segments from the wild species demonstrates crossing over that would have been facilitated only by the limited home-ologous chromosome pairing between the 2 excessively divergent species.
The variable length of O. latifolia chromosome segment introgressions on chromosome 6 of the O. sativa parent may be associated with chromosome breakage and repairs dur-ing homeologous recombination and crossing over, similar to what has been reported in rice and cotton (Jena et al. 1994; Benbouza et al. 2007).
Single Gene Transfer and Quantitative Trait Loci Inheritance in Disomic Derivatives of O. latifolia MAALs
Yield Components
Significant divergence of the mean values for different agro-nomic traits of the disomic introgression lines from those of the control, IR31917-45-32, indicates the presence of putative QTLs in the O. latifolia introgressions. These puta-tive QTLs, alone or in combination, conferred either positive or negative effects on the traits we examined. The variable expression in heading date, plant height, number of primary branches per panicle, number of grains per panicle, and grain width and length during the wet and dry seasons indicates that these traits are highly affected by the environment (Yano et al. 2000; Cao et al. 2001; Luan et al. 2009). It may also be possible that the wide range of phenotypic variations observed are due to genetic or epigenetic variations induced by genomic shock imposed due to wild species chromosome segments in the recipient genome (Shan et al. 2005; Wang et al. 2005). In our earlier collaboration with University of Georgia, Athens, GA, USA to prove the possibility of the role of transposable elements for creating genetic variations by CC genome wild species introgression in the recipient genome due to genomic shock, it was concluded that there was no role of transposable element in creating novel genetic variation of the introgression lines (Wessler S, unpublished data).
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
12
BB Resistance
BB is one of the most serious diseases of rice in Asia, caus-ing yield losses of up to 50% during disease severity (Huang et al. 1997; Chen et al. 2002a). The identification of single or multiple genes of a dominant nature (Xa1, Xa2, Xa3/Xa26, Xa4, Xa7, Xa10, Xa11, Xa12, Xa14, Xa16, Xa17, Xa18, Xa21, Xa22, Xa25(t), and Xa27) and/or recessive nature (xa5, xa8, xa9, xa13, xa15, xa19, xa20, xa24, xa25/Xa25(t), xa26(t), xa28(t), xa31(t), xa33(t), and xa34(t)) for BB resistance (Khush and Kinoshita 1991; Kinoshita 1995; Lin et al. 1996; Chen et al. 2002a; Li and Wang 2013) in existing rice varieties has been the most effective and economical control of the disease. However, as in many host plant–pathogen systems, resistance of the hosts to the pathogens eventually breaks down after many years of cultivation due to shifts in patho-gen race frequency (Rao et al. 2002), hence the need for the identification of new sources of resistance to the disease.
The O. sativa elite breeding line IR31917-45-3-2 has been reported to have resistance to BB races 1 and 5 of the Philippines, whereas O. latifolia has known resistance to races 1, 2, and 3 (Multani et al. 2003). In the current study, the recurrent parent also exhibited moderate to high resistance to races 2, 6, 7, 8, and 10 of the disease, whereas it dem-onstrated susceptibility to the rest of the races tested. The O. latifolia introgressions in the background of IR31917-45-3-2 demonstrated either a positive or negative influence on the inherent reaction of the recurrent parent to the different races of BB. For example, some introgression lines exhib-ited weaker resistance to races 1, 2, 6, and 10 compared with the recurrent parent, suggesting the presence of putative QTLs on the introgressed segments coming from O. latifo-lia that have negative effects on BB resistance. Conversely, a few lines demonstrated resistance to race 9A and 9D of BB despite the susceptibility of the recurrent parent to the pathogen. In particular, lines WH12-2255 and WH12-2256 demonstrated a high resistance to race 9A of the dis-ease (PXO339), recording lesion lengths of only 1.83 and 1.67 cm, respectively, compared with the 12.48 cm average lesion length of the rest of the lines, including the recur-rent parent. Both lines share a common introgression on chromosome 12 from O. latifolia, suggesting the presence of a putative QTL for BB resistance in the chromosome seg-ment introgressed from wild rice. Previous studies identified the BB resistance gene, Xa25(t) from cv Minghui 63 in the centromeric region of chromosome 12. This gene confers resistance to Philippine race 9 of Xoo during both the seed-ling and adult stages of the rice plant. The linkage map for Xa25(t) shows that the genomic location of this gene coin-cides with the map position of the O. latifolia introgression identified in this study (Chen et al. 2002a), indicating the presence of an allelic variant of Xa25(t) in the O. latifolia genome. A survey of gene annotations showed the presence of several NBS-LRR genes containing a nucleotide binding site and a leucine-rich repeat within the map position of the introgressed O. latifolia chromosome segment. Further stud-ies are in progress to fine map the resistance gene(s) inher-ited from O. latifolia.
Strong Stem
The success of the green revolution in rice and wheat was attributed to the identification and incorporation of the semi-dwarf1 (sd1) and reduced height1 (Rht1) genes, respectively (Sakamoto and Matsuoka 2008). A reduction in plant stat-ure has been the predominant strategy in breeding for high-yielding and lodging-resistant crop varieties (Hargrove and Cabanilla 1979). However, despite the short plant stature conferred by sd1 or Rht1 genes in improved rice and wheat varieties, respectively, lodging remains a big problem. To realize higher crop production, it is imperative to identify genes that will improve resistance to lodging (Terashima et al. 1992; Berry et al. 2004). The morphology of the rice culm is one of the parameters that define its strength, which in turn is closely associated with lodging resistance (Ookawa et al. 2010).
The wild rice species O. latifolia has a plant height of 100–200 cm. At maturity, the leaf blades of the plant start to droop, while the stems remain erect up to 1 m from the plant base. Qualitative evaluation for stem strength showed that all the disomic derivatives from O. latifolia MAALs exhibit a strong stem. Verification of the physical strength of the stems showed that all introgression lines had higher SM than the recurrent parent, indicating the presence of putative QTLs for stem strength in the introgression lines we examined. Genotypic examination of the disomic derivatives showed that all lines share a common chromo-some segment of O. latifolia on chromosome 6 spanning 9.42–22.72 Mb. A 9.42 overlapping segment flanked by the SNP markers id6004481 and id6009055 is common in all the lines, suggesting that a putative QTL controlling culm diameter may lie in this introgressed region. Differences in the absolute value of SM in the disomic derivatives may be attributed to the presence of other QTLs with minor effects on the trait. Mapping of this QTL using a popula-tion generated from any of the disomic derivatives might reveal an allelic variant of SCM2 from O. latifolia (Ookawa et al. 2010). Further studies are underway to precisely iden-tify the gene controlling lodging resistance derived from O. latifolia.
Conclusions
The molecular characterization of MAALs and their dis-omic derivatives is fundamental for the development of appropriate strategies for the purposeful and efficient use of exotic genetic resources in rice breeding programs. The identification of wild species introgressions in the 27 O. lat-ifolia-derived disomic derivatives, as well as their agronomic evaluation in the field, is only the first step in tapping into the large amount of genetic diversity present in O. latifolia. The localization of O. latifolia introgressed segments in the uniform genetic background of O. sativa allowed the easy identification by association of putative QTLs controlling valuable traits from the wild species. In particular, putative QTLs exhibiting strong control of resistance to Philippine races 3A, 4, 9A, and 9D of BB as well as strong stem were
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Angeles-Shim et al. • Molecular Analysis of Oryza latifolia Introgression Lines
13
identified. Cloning and functional validation of these QTLs in different genetic backgrounds would contribute in wid-ening the resource base for BB resistance as well as for lodging resistance in rice. The molecular markers that were identified to differentiate between O. sativa and O. latifolia genomes would not only aid in the rapid mapping and clon-ing of the genes identified from these lines but would also facilitate both genomic and genetic studies on breeding resources derived from wild Oryza species that share the same genome as O. latifolia. The evaluation of the disomic derivatives for other biotic and abiotic stresses could facili-tate the identification of several novel genes from O. latifolia that can be used to improve existing rice cultivars.
FundingGlobal Rice Science Partnership (GRiSP) program of the International Rice Research Institute (A-2012-180), Manila, Philippines.
AcknowledgementsWe would like to thank N. T. Quilloy and R. M. Malabanan for providing excellent technical help during the experiment. We are grateful to 2 anony-mous internal reviewers for their critical review of the manuscript. We are also grateful to Dr J. Jacquemin of Arizona Genomics Institute, USA for providing us the sequence alignments between O. alta and O. sativa cv Nipponbare. We also thank B. Hardy, Science Editor, IRRI for editing the manuscript.
ReferencesAmante-Bordeos A, Sitch LA, Nelson R, Dalmacio RD, Oliva NP, Aswidinnoor H, Leung H. 1992. Transfer of bacterial blight and blast resist-ance from the tetraploid wild rice Oryza minuta to cultivated rice, Oryza sativa. Theor Appl Genet. 84:345–354.
Ammiraju JSS, Song X, Luo M, Sisneros N, Angelova A, Kudrna D, Kim HR, Yu Y, Goicoechea JL, Loriuex M, et al. 2010. The Oryza BAC resource: a genus-wide and genome scale tool for exploring rice genome evolution and leveraging useful genetic diversity from wild relatives. Breed Sci. 60:536–543.
Benbouza H, Diouf FBH, Baudoin JP, Mergeai G. 2007. Preferential trans-mission of Gossypium sturtianum chromosome fragments in the prog-eny of [(G. hirsutum x G. raimondii)2 x G. sturtianum] trispecific hybrid. In: Proceedings of the World Cotton Research Conference-4. Lubbock, Texas, USA. 10–14 September 2007.
Berry PM, Spink JH, Gay AP, Craigon JA. 2004. A comparison of root and stem lodging risks among winter wheat cultivars. J Agric Sci. 141:191–202.
Brar DS, Khush GS. 1997. Alien introgression in rice. Plant Mol Biol. 35:35–47.
Brar DS, Singh K. 2011. Oryza. In: Kole C, editor. Wild crop relatives: genomic and breeding resources: cereals. Dordrecht London, New York: Springer Heidelberg. p. 321–326.
Cao G, Zhu J, He C, Gao Y, Yan J, Wu P. 2001. Impact of epistasis and QTL x environment interaction on the developmental behavior of plant height in rice (Oryza sativa L.). Theor Appl Genet. 103:153–160.
Chen H, He H, Zou Y, Chen W, Yu R, Liu X, Yang Y, Gao YM, Xu JL, Fan LM, et al. 2011. Development and application of a set of breeder-friendly SNP markers for genetic analyses and molecular breeding of rice (Oryza sativa L.). Theor Appl Genet. 123:869–879.
Chen H, Wang S, Zhang Q. 2002a. New gene for bacterial blight resistance in rice located on chromosome 12 identified from minghui 63, an elite restorer line. Phytopathology. 92:750–754.
Chen X, Cho YG, McCouch SR. 2002b. Sequence divergence of rice microsatellites in Oryza and other plant species. Mol Genet Genomics. 268:331–343.
Ge S, Sang T, Lu BR, Hong DY. 1999. Phylogeny of rice genomes with emphasis on origins of allotetraploid species. Proc Natl Acad Sci USA. 96:14400–14405.
Hargrove TR, Cabanilla VL. 1979. The impact of semi-dwarf varieties on Asian rice-breeding programs. Bioscience. 29:731–735.
Hernández P, Laurie DA, Martín A, Snape JW. 2002. Utility of barley and wheat simple sequence repeat (SSR) markers for genetic analysis of Hordeum chilense and tritordeum. Theor Appl Genet. 104:735–739.
Huang H, Kochert G. 1994. Comparative RFLP mapping of an allotetra-ploid wild rice species (Oryza latifolia) and cultivated rice (O. sativa). Plant Mol Biol. 25:633–648.
Huang N, Angeles ER, Domingo J, Magpantay G, Singh S, Zhang G, Kumaravadivel N, Bennet J, Khush GS. 1997. Pyramiding of bacterial blight resistance genes in rice: marker-assisted selection using RFLP and PCR. Theor Appl Genet. 95:313–320.
Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu T, Zhang Z, et al. 2010. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat Genet. 42:961–967.
Jacquemin J, Bhatia D, Singh K, Wing RA. 2013. The International Oryza Map Alignment Project: development of a genus-wide comparative genom-ics platform to help solve the 9 billion-people question. Curr Opin Plant Biol. 16:147–156.
Jena KK. 2010. The species of the genus Oryza and transfer of useful genes from wild species into cultivated rice, O. sativa. Breed Sci. 60:518–523.
Jena KK, Khush GS. 1984. Embryo-rescue of interspecific hybrids and its scope in rice improvement. Rice Genet Newl. 1:133–134.
Jena KK, Khush GS. 1989. Monosomic alien addition lines of rice: produc-tion, morphology, cytology, and breeding behavior. Genome. 37:217–221.
Jena KK, Khush GS. 1990. Introgression of genes from Oryza officinalis Well ex Watt to cultivated rice, O. sativa L. Theor Appl Genet. 80:737–745.
Jena KK, Kochert G, Khush GS. 1992. RFLP analysis of rice (Oryza sativa L.) introgression lines. Theor Appl Genet. 84:608–616.
Jena KK, Khush GS, Kochert G. 1994. Comparative RFLP mapping of a wild rice, Oryza officinalis, and cultivated rice, O. sativa. Genome. 37:382–389.
Jena KK, Multani DS, Khush GS. 1991. Monosomic alien addition lines of Oryza australiensis and alien gene transfer. In: Rice genetics II. Philippines: International Rice Research Institute. p. 728.
Jeung JU, Heu SG, Shin MS, Vera Cruz CM, Jena KK. 2006. Dynamics of Xanthomonas oryzae pv. oryzae populations in Korea and their relationship to known bacterial blight resistance genes. Phytopathology. 96:867–875.
Jin H, Tan G, Brar DS, Tang M, Li G, Zhu L, He G. 2006. Molecular and cytogenetic characterization of an Oryza officinalis-O. sativa chromosome 4 addition line and its progenies. Plant Mol Biol. 62:769–777.
Kauffman HE, Reddy APK, Hsieh SPY, Merca SD. 1973. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae pv. oryzae. Plant Dis Rep. 57:537–541.
Khush GS. 1997. Origin, dispersal, cultivation and variation of rice. Plant Mol Biol. 35:25–34.
Khush GS, Bacalangco E, Ogawa T. 1990. A new gene for resistance to bac-terial blight from O. longistaminata. Rice Genet Newsl. 8:106–109.
Khush GS, Kinoshita T. 1991. Rice karyotype, marker gene and link-age groups. In: Khush GS, Toenniessen GH, editors. Rice biotechnology. Wallingford, UK: CABI Int. p. 83–108.
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

Journal of Heredity
14
Kinoshita T. 1995. Report of committee on gene symbolization, nomencla-ture and linkage groups. Rice Genet Newsl. 12:9–153.
Li H, Wang S. 2013. Disease resistance. In: Zhang Q, Wing RA, editors. Genetics and genomics of rice, plant genetics and genomics: crops and models 5. Springer. p. 161–175.
Lin XH, Zhang DP, Xie YF, Gao HP, Zhang Q. 1996. Identifying and map-ping a new gene for bacterial blight resistance in rice based on RFLP mark-ers. Phytopathology. 86:1156–1159.
Lorieux M, Ndjiondjop MN, Ghesquiere A. 2000. A first interspeci Oryza sativa x Oryza glaberrima microsatellite-based genetic map. Theor Appl Genet. 100:591–601.
Lu F, Ammiraju JS, Sanyal A, Zhang S, Song R, Chen J, Li G, Sui Y, Song X, Cheng Z, et al. 2009. Comparative sequence analysis of MONOCULM1-orthologous regions in 14 Oryza genomes. Proc Natl Acad Sci USA. 106:2071–2076.
Luan W, Chen H, Fu Y, Si H, Peng W, Song S, Liu W, Hu G, Sun Z, Xie D, et al. 2009. The effect of the crosstalk between photoperiod and tempera-ture on the heading-date in rice. PLoS One. 4:e5891.
Maughan PJ, Saghi Maroof MA, Buss GR. 1995. Microsatellite and amplified sequence length polymorphisms in cultivated and wild soybean. Genome. 38:715–723.
McCouch SR, Sweeney M, Li J, Jiang H, Thomson M, Septiningsih E, Edwards J, Moncada P, Xiao J, Garris A, et al. 2007. Through the genetic bottleneck; O. rufipogon as a source of trait-enhancing alleles for O. sativa. Euphytica. 154:317–339.
McCouch SR, Zhao K, Wright M, Tung CW, Ebana K, Thomson M, Reynolds A, Wang D, DeClerck G, Ali Md L, et al. 2010. Development of genome-wide SNP assays for rice. Breed Sci. 60:524–535.
McNally KL, Childs KL, Bohnert R, Davidson RM, Zhao K, Ulat VJ, Zeller G, Clark RM, Hoen DR, Bureau TE, et al. 2009. Genomewide SNP variation reveals relationships among landraces and modern varieties of rice. Proc Natl Acad Sci USA. 106:12273–12278.
Mullan DJ, Platteter A, Teakle NL, Appels R, Colmer TD, Anderson JM, Francki MG. 2005. EST-derived SSR markers from defined regions of the wheat genome to identify Lophopyrum elongatum specific loci. Genome. 48:811–822.
Multani DS, Jena KK, Brar DS, de Los Reyes BG, Angeles ER, Khush GS. 1994. Development of monosomic alien addition lines and introgression of genes from Oryza australiensis Domin. to cultivated rice O. sativa L. Theor Appl Genet. 88:102–109.
Multani DS, Khush GS, delos Reyes BG, Brar DS. 2003. Alien genes intro-gression and development of monosomic alien addition lines from Oryza latifolia Desv. to rice, Oryza sativa L. Theor Appl Genet. 107:395–405.
Ookawa T, Hobo T, Yano M, Murata K, Ando T, Miura H, Asano K, Ochiai Y, Ikeda M, Nishitani R, et al. 2010. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat Commun. 1:132.
Panaud O, Chen X, McCouch SR. 1996. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol Gen Genet. 252:597–607.
Rao KK, Lakshminarasu M, Jena KK. 2002. DNA markers and marker-assisted breeding for durable resistance to bacterial blight disease in rice. Biotechnol Adv. 20:33–47.
Röder MS, Plaschke J, König SU, Börner A, Sorrells ME, Tanksley SD, Ganal MW. 1995. Abundance, variability and chromosomal location of microsatel-lites in wheat. Mol Gen Genet. 246:327–333.
Rick CM. 1971. Further studies on segregation and recombination in back-cross derivatives of a tomato species hybrid. Biol Zentrabl. 91:209–220.
Saghai Maroof MA, Biyashev RM, Yang GP, Zhang Q, Allard RW. 1994. Extraordinarily polymorphic microsatellite DNA in barley: species diversity, chromosomal locations, and population dynamics. Proc Natl Acad Sci USA. 91:5466–5470.Sakamoto T, Matsuoka M. 2008. Identifying and exploiting grain yield genes in rice. Curr Opin Plant Biol. 11:209–214.
Sanchez PL, Wing RA, Brar DS. 2013. The wild relative of rice: genomes and genomics. In: Zhang Q, Wing RA, editors. Genetics and genomics of rice, plant genetics and genomics; crops and models 5. Springer. p. 10–25.Senior ML, Heun M. 1993. Mapping maize microsatellites and polymer-ase chain reaction confirmation of the targeted repeats using a CT primer. Genome. 36:884–889.Shan X, Liu Z, Dong Z, Wang Y, Chen Y, Lin X, Long L, Han F, Dong Y, Liu B. 2005. Mobilization of the active MITE transposons mPing and Pong in rice by introgression from wild rice (Zizania latifolia Griseb.). Mol Biol Evol. 22:976–990.Shim J, Panaud O, Vitte C, Mendioro MS, Brar DS. 2010. RDA derived Oryza minuta specific clones to probe genomic conservation across Oryza and introgression into rice (O. sativa L). Euphytica. 176:269–279.Smulders MJM, Bredemeijer G, Rus-Kortekaas W, Arenas P, Vosman B. 1997. Use of short microsatellites from database sequences to generate pol-ymorphisms among Lycopersicon esculentum cultivars and accessions of other Lycopersicon species. Theor Appl Genet. 94:264–272.Stephens SG. 1949. The cytogenetics of speciation in Gossypium; selec-tive elimination of the donor parent genotype in interspecific backcrosses. Genetics. 34:627–637.Suh JP, Jeung JU, Noh TH, Cho YC, Park SH, Park HS, Shin MS, Kim CK, Jena KK. 2013. Development of breeding lines with three pyramided resist-ance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice (N Y). 6:5.Temnykh S, DeClerck G, Lukashova A, Lipovich L, Cartinhour S, McCouch S. 2001. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 11:1441–1452.Terashima K, Akita S, Sakai N. 1992. Ecophysiological characteristics related with lodging tolerance of rice in direct sowing cultivation. I. Comparison of the root lodging tolerance among cultivars by measurement of pushing resistance. Jpn J Crop Sci. 61:380–387.Thomson MJ, Zhao K, Wright M, McNally KL, Rey J, Tung CW, AReynolds, Scheffler B, Eizenga, GMcClung A, et al. 2012. High-throughput single nucleotide polymorphism genotyping for breeding applications in rice using BeadXpress platform. Mol Breed. 29:875–886.Vaughan DA, Morishima H, Kadowaki K. 2003. Diversity in the Oryza genus. Curr Opin Plant Biol. 6:139–146.Vroh Bi I, Maquet A, Baudoin JP, Du Jardin P, Jacquemin JM, Mergeai G. 1999. Breeding for “low-gossypol seed and high-gossypol plants” in upland cotton: analysis of tri-species hybrids and backcross progenies using AFLPs and mapped RFLPs. Theor Appl Genet. 99:1233–1244.Wang YM, Dong ZY, Zhang ZJ, Lin XY, Shen Y, Zhou D, Liu B. 2005. Extensive de Novo genomic variation in rice induced by introgression from wild rice (Zizania latifolia Griseb.). Genetics. 170:1945–1956.Wu KS, Tanksley SD. 1993. Abundance, polymorphism and genetic mapping of microsatellites in rice. Mol Gen Genet. 241:225–235.Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, Baba T, Yamamoto K, Umehara Y, Nagamura Y, et al. 2000. Hd1, a major pho-toperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell. 12:2473–2484.Ying B, Song GE. 2003. Identification of Oryza species with the CD genome using RFLP analysis on nuclear ribosomal ITS sequences. Acta Bot Sin. 45:762–765.
Zeigler RS. 2013. Food security, climate change and genetic resources. In: Jackson M, Ford-Lloyd B, Parry M, editors. Plant genetic resources and cli-mate change. UK: CAB International. p. 1–15.
Received September 22, 2013; First decision January 25, 2014; Accepted April 21, 2014
Corresponding Editor: Kenneth Olsen
at University of C
onnecticut on June 21, 2014http://jhered.oxfordjournals.org/
Dow
nloaded from

![1. iv congreso internacional [desv] cip](https://static.fdocuments.net/doc/165x107/58d0273d1a28ab04288b47ab/1-iv-congreso-internacional-desv-cip.jpg)


![Ergebnisliste U14 - kreis100-bayerwald.de · Manuela Hallhuber Kohlschmid Franz, Christian Rimsl DESV Jugendwartin Feichtgruber Simon,Meyer Stefan DESV Sportwart [Wettbewerbsleiter]](https://static.fdocuments.net/doc/165x107/5d50509288c993e32d8bbd2a/ergebnisliste-u14-kreis100-manuela-hallhuber-kohlschmid-franz-christian.jpg)













