
Modulation of the Growth-inhibitory Effects of Progestins and the...
6
[CANCER RESEARCH 49, 112-116, January 1, 1989] Modulation of the Growth-inhibitory Effects of Progestins and the Antiestrogen Hydroxyclomiphene on Human Breast Cancer Cells by Epidermal Growth Factor and Insulin1 Masafumi Koga,2 Elizabeth A. Musgrove,3 and Robert L. Sutherland4 Garran Institute of Medical Research, St. Vincent's Hospital, Sydney, New South Wales 2010, Australia ABSTRACT The molecular basis of the growth-inhibitory effects of progestins or antiestrogens in human breast cancer has not been fully elucidated. Both direct actions and indirect actions, where the growth inhibition results from modulation of the production of, and/or the response to, growth factors, have been proposed. In this study the ability of some growth factors to modulate progestin-induced inhibition of cell proliferation was investigated in vitro, using 1-471) human breast cancer cells. When I -471) cells grown in insulin-containing medium were treated for 4 to 5 days with the synthetic progestin, ORG 2058, at a concentration of 10 UM, cell numbers were reduced to 10 to 20% of control. Simulta neous treatment with epidermal growth factor (EGF) and ORG 2058 led to a partial reversal of the growth-inhibitory effect of the progestin. The magnitude of the effect of EGF was concentration dependent, being half- maximal at 0.48 ng/ml (0.08 MM)and maximal at concentrations > 5 ng/ ml (> 0.8 n\i ), where cell numbers were increased by 50% compared to those in the presence of ORG 2058 alone. ORG 2058 was no more potent in the absence of insulin, and, after several passages in insulin-free medium, addition of insulin failed to modulate the effect of ORG 2058. However, when maximal concentrations of insulin (5 Mg/ml) and EGF (10 ng/ml) were administered together with ORG 2058, insulin and EGF appeared to act synergistically to reduce the ORG 2058-induced inhibi tion of proliferation. In similar experiments in which cells were treated with hydroxyclom- iphene, a potent antiestrogen, insulin was shown to partially reverse the growth-inhibitory effects of hydroxyclomiphene. Significant increases in cell number above hydroxyclomiphene-treated controls were apparent at insulin concentrations > 50 ng/ml, and at 5 Mg/ml the increase was approximately 2-fold. In contrast to the situation with progestins, simul taneous treatment with EGF and insulin had only an additive effect in reversing the growth-inhibitory effect of the antiestrogen. The results are compatible with the hypothesis that part of the growth- inhibitory effects of progestin and antiestrogen on human breast cancer cell proliferation is mediated by inhibition of autocrine growth factor production. However, they do not exclude more direct mechanisms in volving modulation of progesterone and/or estrogen receptors by EGF and/or insulin. In addition the differential effects of insulin alone, and insulin in combination with EGF, on progestin and antiestrogen-treated cells suggest that, although progestin and antiestrogen-modulated path ways may converge, they are not identical in terms of their interactions with growth factor-mediated pathways. INTRODUCTION For nearly a century it has been known that the ovaries are the major source of the stimulatory signal for the growth of hormone-dependent human breast cancer (1). Estradici was subsequently identified as the active substance (2). The growth Received 6/14/88; revised 8/30/88; accepted 9/26/88. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1Supported by the New South Wales State Cancer Council and the National Health and Medical Research Council of Australia. 2 Visiting Research Fellow from the Third Department of Internal Medicine, Osaka University Hospital, Osaka, Japan, supported by a grant-in-aid from the Mochida Memorial Foundation for Medical and Pharmaceutical Research. 'Government Employees Assistance to Medical Research Fund Doctoral Scholar. 4 To whom requests for reprints should be addressed. of hormone-dependent breast cancer is specifically inhibited by synthetic antiestrogens acting predominantly via the ER,5 and this is thought to involve inhibition of the mitogenic effect of estrogen (3-8). Progestins are also used in the endocrine ther apy of breast cancer where their effect is apparently mediated by the PR (9,10). However, the sequence of postreceptor events leading to the inhibition of the mitogenic response by antiestro gens and progestins in breast cancer cells remains poorly under stood. In addition to estrogen, a variety of mitogens have been identified for human breast cancer cells in vitro. In chemically defined culture conditions, insulin causes a marked increase in the growth rate of most breast cancer cell lines (11-16), some of which also respond to EGF (11-15,17) while estrogen alone is commonly either slightly stimulatory or ineffective (11, 13, 15). It has recently been postulated that estrogens and anties trogens exert their effects on breast cancer cell proliferation indirectly, by regulation of the synthesis and/or secretion of autocrine and paracrine growth factors, including «-TGF and IGF-1 (18-22). In the well-established fibroblast model of mammalian cell cycle regulation, EGF and IGF-1 are "progression" factors, which act at specific points during the d phase of the cell cycle to ensure the cell cycle progression of cells already committed to DNA synthesis (23, 24). If breast cancer cells are regulated in an analogous fashion, growth factors acting through the EGF receptor (a-TGF or EGF) or the IGF-1 receptor (insulin or IGF-1) might be expected to exert their effects at specific times within the GI phase of the cell cycle. It is possible that these sites of action are coincident with the sites of action, early in d phase, of progestins (10) and antiestrogens (6, 25). If this were true, exogenous progression factors would be expected to modify the cellular response to the growth-inhibitory effects of progestins and antiestrogens. Indeed, it has been reported that either EGF or insulin can modulate the growth-inhibitory ef fects of antiestrogens (26-28). In the present study we have further tested this hypothesis by studying the effects of EGF and insulin, alone and in combination, on the response of T- 47D human breast cancer cells to the antiproliferative effects of progestins and antiestrogens. MATERIALS AND METHODS Materials. ORG 2058 and Progesterone were purchased from Amer- sham Australia (Sydney, Australia), and Sigma Chemical Co. (St. Louis, MO), respectively. M PA was obtained from Upjohn Pty, Ltd. (Sydney, Australia), through the courtesy of Dr. Dudley Jacobs. Hydroxyclomi phene was synthesized as previously describe'! (29) and donated by Dr. Peter Ruenitz, College of Pharmacy, University of Georgia, Athens, GA. Murine EGF, tissue culture grade, was supplied by Collaborative 5The abbreviations used are; a-TGF, »-transforming growth factor, /3-TGF, ¿¡-transforming growth factor, IGF, insulin-like growth factor; EGF, epidermal growth factor, MPA, medroxyprogesterone acetate ( 17a-acetoxy-6a-methyl-4- pregnene-3,20-dione); ORG 2058, 16a-ethyl-21-hydroxy-19-norpregn-4-en-3,20- dione; hydroxyclomiphene, [1-(4-0-dimethylaminoethoxyphenyl)-1 -(4-hydroxy- phenyl)-2-chloro-2-phenylethylene); FCS, fetal calf serum; ER, estrogen receptor, PR, progesterone receptor; EGF-R, epidermal growth factor receptor. 112 Research. on February 2, 2019. © 1989 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Modulation of the Growth-inhibitory Effects of Progestins and the...

[CANCER RESEARCH 49, 112-116, January 1, 1989]
Modulation of the Growth-inhibitory Effects of Progestins and the Antiestrogen
Hydroxyclomiphene on Human Breast Cancer Cells by Epidermal GrowthFactor and Insulin1
Masafumi Koga,2 Elizabeth A. Musgrove,3 and Robert L. Sutherland4
Garran Institute of Medical Research, St. Vincent's Hospital, Sydney, New South Wales 2010, Australia
ABSTRACT
The molecular basis of the growth-inhibitory effects of progestins or
antiestrogens in human breast cancer has not been fully elucidated. Bothdirect actions and indirect actions, where the growth inhibition resultsfrom modulation of the production of, and/or the response to, growthfactors, have been proposed. In this study the ability of some growthfactors to modulate progestin-induced inhibition of cell proliferation wasinvestigated in vitro, using 1-471) human breast cancer cells.
When I -471) cells grown in insulin-containing medium were treated
for 4 to 5 days with the synthetic progestin, ORG 2058, at a concentrationof 10 UM, cell numbers were reduced to 10 to 20% of control. Simultaneous treatment with epidermal growth factor (EGF) and ORG 2058 ledto a partial reversal of the growth-inhibitory effect of the progestin. Themagnitude of the effect of EGF was concentration dependent, being half-
maximal at 0.48 ng/ml (0.08 MM)and maximal at concentrations > 5 ng/ml (> 0.8 n\i ), where cell numbers were increased by 50% compared tothose in the presence of ORG 2058 alone. ORG 2058 was no more potentin the absence of insulin, and, after several passages in insulin-free
medium, addition of insulin failed to modulate the effect of ORG 2058.However, when maximal concentrations of insulin (5 Mg/ml) and EGF(10 ng/ml) were administered together with ORG 2058, insulin and EGFappeared to act synergistically to reduce the ORG 2058-induced inhibi
tion of proliferation.In similar experiments in which cells were treated with hydroxyclom-
iphene, a potent antiestrogen, insulin was shown to partially reverse thegrowth-inhibitory effects of hydroxyclomiphene. Significant increases incell number above hydroxyclomiphene-treated controls were apparent at
insulin concentrations > 50 ng/ml, and at 5 Mg/ml the increase wasapproximately 2-fold. In contrast to the situation with progestins, simul
taneous treatment with EGF and insulin had only an additive effect inreversing the growth-inhibitory effect of the antiestrogen.
The results are compatible with the hypothesis that part of the growth-
inhibitory effects of progestin and antiestrogen on human breast cancercell proliferation is mediated by inhibition of autocrine growth factorproduction. However, they do not exclude more direct mechanisms involving modulation of progesterone and/or estrogen receptors by EGFand/or insulin. In addition the differential effects of insulin alone, andinsulin in combination with EGF, on progestin and antiestrogen-treatedcells suggest that, although progestin and antiestrogen-modulated path
ways may converge, they are not identical in terms of their interactionswith growth factor-mediated pathways.
INTRODUCTION
For nearly a century it has been known that the ovaries arethe major source of the stimulatory signal for the growth ofhormone-dependent human breast cancer (1). Estradici wassubsequently identified as the active substance (2). The growth
Received 6/14/88; revised 8/30/88; accepted 9/26/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by the New South Wales State Cancer Council and the National
Health and Medical Research Council of Australia.2Visiting Research Fellow from the Third Department of Internal Medicine,
Osaka University Hospital, Osaka, Japan, supported by a grant-in-aid from theMochida Memorial Foundation for Medical and Pharmaceutical Research.
'Government Employees Assistance to Medical Research Fund Doctoral
Scholar.4To whom requests for reprints should be addressed.
of hormone-dependent breast cancer is specifically inhibited bysynthetic antiestrogens acting predominantly via the ER,5 and
this is thought to involve inhibition of the mitogenic effect ofestrogen (3-8). Progestins are also used in the endocrine therapy of breast cancer where their effect is apparently mediatedby the PR (9,10). However, the sequence of postreceptor eventsleading to the inhibition of the mitogenic response by antiestrogens and progestins in breast cancer cells remains poorly understood. In addition to estrogen, a variety of mitogens have beenidentified for human breast cancer cells in vitro. In chemicallydefined culture conditions, insulin causes a marked increase inthe growth rate of most breast cancer cell lines (11-16), someof which also respond to EGF (11-15,17) while estrogen aloneis commonly either slightly stimulatory or ineffective (11, 13,15). It has recently been postulated that estrogens and antiestrogens exert their effects on breast cancer cell proliferationindirectly, by regulation of the synthesis and/or secretion ofautocrine and paracrine growth factors, including «-TGFandIGF-1 (18-22).
In the well-established fibroblast model of mammalian cellcycle regulation, EGF and IGF-1 are "progression" factors,
which act at specific points during the d phase of the cell cycleto ensure the cell cycle progression of cells already committedto DNA synthesis (23, 24). If breast cancer cells are regulatedin an analogous fashion, growth factors acting through the EGFreceptor (a-TGF or EGF) or the IGF-1 receptor (insulin orIGF-1) might be expected to exert their effects at specific timeswithin the GI phase of the cell cycle. It is possible that thesesites of action are coincident with the sites of action, early ind phase, of progestins (10) and antiestrogens (6, 25). If thiswere true, exogenous progression factors would be expected tomodify the cellular response to the growth-inhibitory effects ofprogestins and antiestrogens. Indeed, it has been reported thateither EGF or insulin can modulate the growth-inhibitory effects of antiestrogens (26-28). In the present study we havefurther tested this hypothesis by studying the effects of EGFand insulin, alone and in combination, on the response of T-47D human breast cancer cells to the antiproliferative effectsof progestins and antiestrogens.
MATERIALS AND METHODS
Materials. ORG 2058 and Progesterone were purchased from Amer-
sham Australia (Sydney, Australia), and Sigma Chemical Co. (St. Louis,MO), respectively. MPA was obtained from Upjohn Pty, Ltd. (Sydney,Australia), through the courtesy of Dr. Dudley Jacobs. Hydroxyclomiphene was synthesized as previously describe'! (29) and donated by Dr.Peter Ruenitz, College of Pharmacy, University of Georgia, Athens,GA. Murine EGF, tissue culture grade, was supplied by Collaborative
5The abbreviations used are; a-TGF, »-transforming growth factor, /3-TGF,¿¡-transforminggrowth factor, IGF, insulin-like growth factor; EGF, epidermalgrowth factor, MPA, medroxyprogesterone acetate (17a-acetoxy-6a-methyl-4-pregnene-3,20-dione); ORG 2058, 16a-ethyl-21-hydroxy-19-norpregn-4-en-3,20-dione; hydroxyclomiphene, [1-(4-0-dimethylaminoethoxyphenyl)-1 -(4-hydroxy-phenyl)-2-chloro-2-phenylethylene); FCS, fetal calf serum; ER, estrogen receptor,PR, progesterone receptor; EGF-R, epidermal growth factor receptor.
112
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
MODULATION OF PROGESTIN ACTION BY EGF AND INSULIN
Research, Inc. (Lexington, MA). Porcine insulin (Actrapid) was obtained from CSL-Novo Industries (Sydney, Australia) and stored at4°C.Other tissue culture materials were purchased from Flow Labo
ratories (Sydney, Australia).All progestins and hydroxyclomiphene were prepared as 1000-fold
concentrated stock solutions in analytical reagent grade ethanol andstored at -20°C. EGF was stored in aliquots at -20°C, as 1,000-fold
concentrated solutions in tissue culture medium.Cell Culture. T-47D cells in their 74th passage were supplied by E.
G. and G. Mason Research Institute, Worcester, MA, for the NationalCancer Institute Breast Cancer Program Cell Culture Bank, and used,in experiments reported here, between passages 92 and 117. Stockcultures were passaged weekly with an inoculation density of 5 x 10scells/75-cm2 culture flask (Corning Glass, Corning, NY) in 30 ml of
RPMI 1640 medium supplemented with 5 TDMglutamine, 14 mmsodium bicarbonate, 20 mM 4-(2-hydroxyethyl)-l-piperazineethanesul-fonic acid (HEPES) buffer, 20 jig/ml of gentamicin, 10 fig/ml of porcineinsulin, 6 Mg/ml of phenol red, and 10% FCS. Some experiments usedparallel stock cultures maintained in the same medium with insulinomitted.
Cell Growth Experiments. The experimental design consisted ofplating 5 x IO4exponentially growing T-47D cells into 25-cm2 flasks
in 5 ml of RPMI 1640 medium with the concentration of PCS reducedto 5% and containing insulin, except where noted in the text. Whencell numbers had doubled to approximately IO5cells/flask, 5-^1 aliquots
of drug or vehicle were added directly to the culture medium. At varioustimes thereafter, cells were harvested with 0.05% trypsin:0.02% EDTAin Ca2+-Mg2+-freephosphate-buffered saline (1.5 mM KH2PO4:8.1 mM
Na2HPO4:2.7 mM KC1:140 mM NaCl), and viable cell counts weremade on a hemocytometer under phase-contrast microscopy.
RESULTS
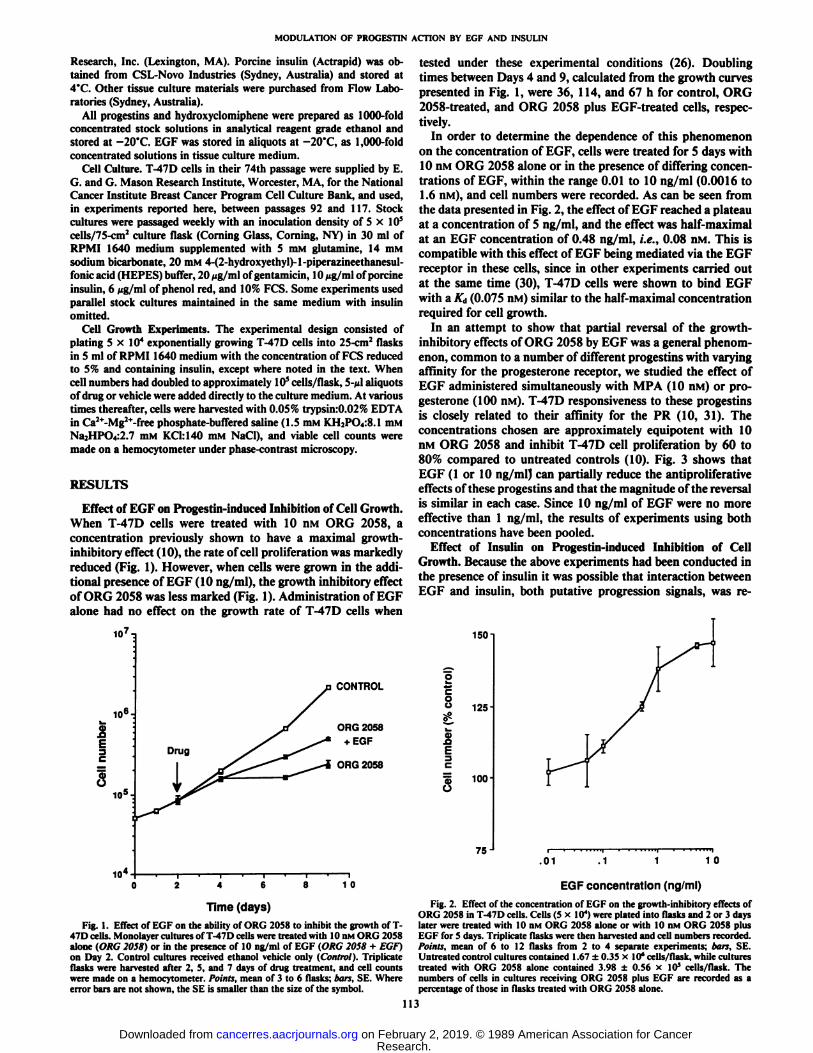
Effect of EGF on Progestin-induced Inhibition of Cell Growth.When T-47D cells were treated with 10 nM ORG 2058, aconcentration previously shown to have a maximal growth-inhibitory effect (10), the rate of cell proliferation was markedlyreduced (Fig. 1). However, when cells were grown in the additional presence of EGF (10 ng/ml), the growth inhibitory effectof ORG 2058 was less marked (Fig. 1). Administration of EGFalone had no effect on the growth rate of T-47D cells when
10'-
CONTROL
ORG 2058
+ EGF
ORG 2058
10
Time (days)Fig. 1. Effect of EGF on the ability of ORG 2058 to inhibit the growth of T-
47D cells. Monolayer cultures of T-47D cells were treated with 10 nM ORG 2058alone (ORG 2058) or in the presence of 10 ng/ml of EGF (ORG 2058 + EGF)on Day 2. Control cultures received ethanol vehicle only (Control)- Triplicateflasks were harvested after 2, 5, and 7 days of drug treatment, and cell countswere made on a hemocytometer. Points, mean of 3 to 6 flasks; bars, SE. Whereerror bars are not shown, the SE is smaller than the size of the symbol.
tested under these experimental conditions (26). Doublingtimes between Days 4 and 9, calculated from the growth curvespresented in Fig. 1, were 36, 114, and 67 h for control, ORG2058-treated, and ORG 2058 plus EGF-treated cells, respec
tively.In order to determine the dependence of this phenomenon
on the concentration of EGF, cells were treated for 5 days with10 nM ORG 2058 alone or in the presence of differing concentrations of EGF, within the range 0.01 to 10 ng/ml (0.0016 to1.6 nM), and cell numbers were recorded. As can be seen fromthe data presented in Fig. 2, the effect of EGF reached a plateauat a concentration of 5 ng/ml, and the effect was half-maximalat an EGF concentration of 0.48 ng/ml, i.e., 0.08 nM. This iscompatible with this effect of EGF being mediated via the EGFreceptor in these cells, since in other experiments carried outat the same time (30), T-47D cells were shown to bind EGFwith a KÕ(0.075 nM) similar to the half-maximal concentrationrequired for cell growth.
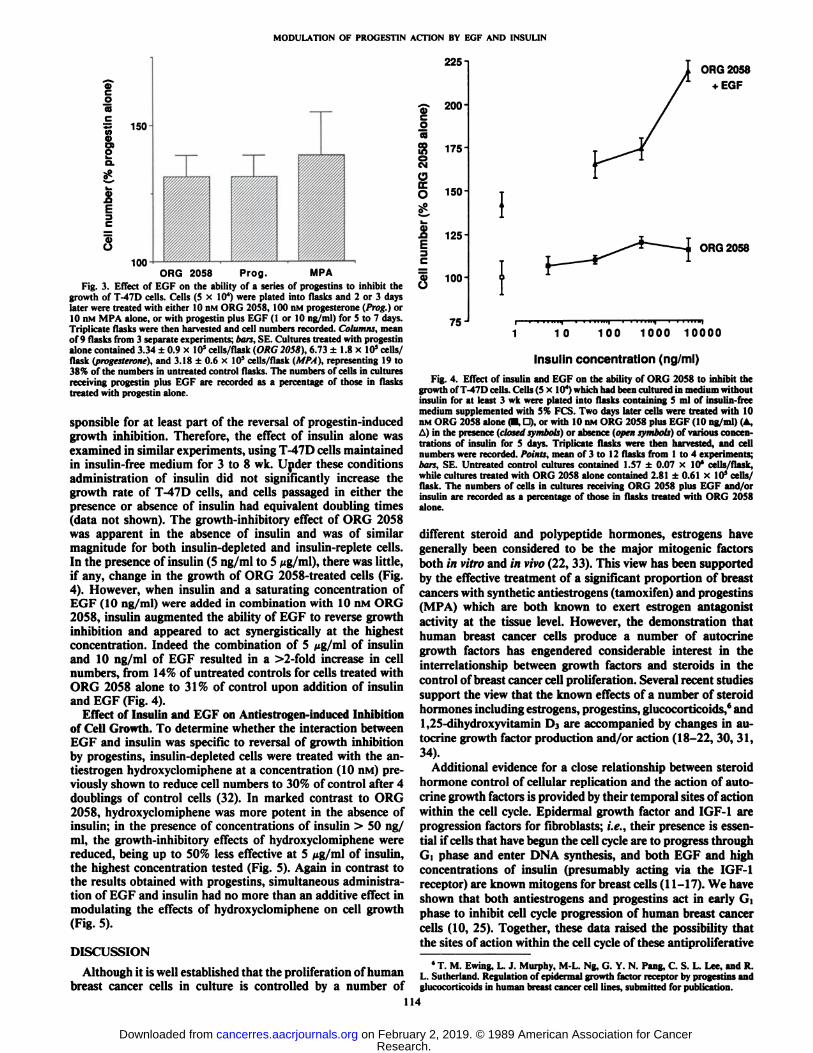
In an attempt to show that partial reversal of the growth-inhibitory effects of ORG 2058 by EGF was a general phenomenon, common to a number of different progestins with varyingaffinity for the progesterone receptor, we studied the effect ofEGF administered simultaneously with MPA (10 nM) or progesterone (100 nM). T-47D responsiveness to these progestinsis closely related to their affinity for the PR (10, 31). Theconcentrations chosen are approximately equipotent with 10HM ORG 2058 and inhibit T-47D cell proliferation by 60 to80% compared to untreated controls (10). Fig. 3 shows thatEGF (1 or 10 ng/ml) can partially reduce the antiproliferativeeffects of these progestins and that the magnitude of the reversalis similar in each case. Since 10 ng/ml of EGF were no moreeffective than 1 ng/ml, the results of experiments using bothconcentrations have been pooled.
Effect of Insulin on Progestin-induced Inhibition of CellGrowth. Because the above experiments had been conducted inthe presence of insulin it was possible that interaction betweenEGF and insulin, both putative progression signals, was re-
150-
o« 125-1
Q>a
e^ 100o
75 J.01 .1 1 10
EGF concentration (ng/ml)Fig. 2. Effect of the concentration of EGF on the growth-inhibitory effects of
ORG 2058 in T-47D cells. Cells (5 x IO4)were plated into flasks and 2 or 3 days
later were treated with 10 nM ORG 2058 alone or with 10 nM ORG 2058 plusEGF for 5 days. Triplicate flasks were then harvested and cell numbers recorded.Points, mean of 6 to 12 flasks from 2 to 4 separate experiments; han. SE.Untreated control cultures contained 1.67 ±0.35 x 10*cells/flask, while culturestreated with ORG 2058 alone contained 3.98 ±0.56 x 10* cells/flask. The
numbers of cells in cultures receiving ORG 2058 plus EGF are recorded as apercentage of those in flasks treated with ORG 2058 alone.
113
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from
MODULATION OF PROGESTIN ACTION BY EGF AND INSULIN
CO
"Ì 150
100ORG 2058 Prog. MPA
Fig. 3. Effect of EGF on the ability of a series of progestins to inhibit thegrowth of T-47D cells. Cells (5 x IO4) were plated into flasks and 2 or 3 days
later were treated with either 10 nM ORG 2058, 100 nM progesterone (Prog.) or10 nM MPA alone, or with progestin plus EGF (1 or 10 ng/ml) for 5 to 7 days.Triplicate flasks were then harvested and cell numbers recorded. Columns, meanof 9 flasks from 3 separate experiments; bars, SE. Cultures treated with progestinalone contained 3.34 ±0.9 x 10' cells/flask (ORG 2058), 6.73 ±1.8 x 10*cells/flask (progesterone), and 3.18 ±0.6 x 10* cells/flask (MPA), representing 19 to
38% of the numbers in untreated control flasks. The numbers of cells in culturesreceiving progestin plus EGF are recorded as a percentage of those in flaskstreated with progestin alone.
sponsible for at least part of the reversal of progestin-inducedgrowth inhibition. Therefore, the effect of insulin alone wasexamined in similar experiments, using T-47D cells maintainedin insulin-free medium for 3 to 8 wk. Under these conditionsadministration of insulin did not significantly increase thegrowth rate of T-47D cells, and cells passaged in either thepresence or absence of insulin had equivalent doubling times(data not shown). The growth-inhibitory effect of ORG 2058was apparent in the absence of insulin and was of similarmagnitude for both insulin-depleted and insulin-replete cells.In the presence of insulin (5 ng/ml to 5 ¿ig/ml),there was little,if any, change in the growth of ORG 2058-treated cells (Fig.4). However, when insulin and a saturating concentration ofEGF (10 ng/ml) were added in combination with 10 nM ORG2058, insulin augmented the ability of EGF to reverse growthinhibition and appeared to act synergistically at the highestconcentration. Indeed the combination of 5 iig/m\ of insulinand 10 ng/ml of EGF resulted in a >2-fold increase in cellnumbers, from 14% of untreated controls for cells treated withORG 2058 alone to 31 % of control upon addition of insulinand EGF (Fig. 4).
Effect of Insulin and EGF on Antiestrogen-induced Inhibitionof Cell Growth. To determine whether the interaction betweenEGF and insulin was specific to reversal of growth inhibitionby progestins, insulin-depleted cells were treated with the an-tiestrogen hydroxyclomiphene at a concentration (10 nM) previously shown to reduce cell numbers to 30% of control after 4doublings of control cells (32). In marked contrast to ORG2058, hydroxyclomiphene was more potent in the absence ofinsulin; in the presence of concentrations of insulin > 50 ng/ml, the growth-inhibitory effects of hydroxyclomiphene werereduced, being up to 50% less effective at 5 tig/m\ of insulin,the highest concentration tested (Fig. 5). Again in contrast tothe results obtained with progestins, simultaneous administration of EGF and insulin had no more than an additive effect inmodulating the effects of hydroxyclomiphene on cell growth(Fig. 5).
DISCUSSION
Although it is well established that the proliferation of humanbreast cancer cells in culture is controlled by a number of
2ocoSoCMOO.O
¿I
225 n
200
175-
150
125
100-
75 J
ORG 2058•fEGF
ORG 2058
10 100 1000 10000
Insulin concentration (ng/ml)
Fig. 4. Effect of insulin and EGF on the ability of ORG 2058 to inhibit thegrowth of T-47D cells. Cells (5x10*) which had been cultured in medium withoutinsulin for at least 3 wk were plated into flasks containing 5 ml of insulin-freemedium supplemented with 5% FCS. Two days later cells were treated with 10nM ORG 2058 alone (•.D), or with 10 nM ORG 2058 plus EGF (10 ng/ml) (A,A) in the presence (closed symbols) or absence (open symbols) of various concentrations of insulin for 5 days. Triplicate flasks were then harvested, and cellnumbers were recorded. Points, mean of 3 to 12 flasks from 1 to 4 experiments;bars, SE. Untreated control cultures contained 1.57 ±0.07 x 10' cells/flask,while cultures treated with ORG 2058 alone contained 2.81 ±0.61 x 10s cells/
flask. The numbers of cells in cultures receiving ORG 2058 plus EGF and/orinsulin are recorded as a percentage of those in flasks treated with ORG 2058alone.
different steroid and polypeptide hormones, estrogens havegenerally been considered to be the major mitogenic factorsboth in vitro and in vivo (22, 33). This view has been supportedby the effective treatment of a significant proportion of breastcancers with synthetic antiestrogens (tamoxifen) and progestins(MPA) which are both known to exert estrogen antagonistactivity at the tissue level. However, the demonstration thathuman breast cancer cells produce a number of autocrinegrowth factors has engendered considerable interest in theinterrelationship between growth factors and steroids in thecontrol of breast cancer cell proliferation. Several recent studiessupport the view that the known effects of a number of steroidhormones including estrogens, progestins, glucocorticoids,6 and1,25-dihydroxyvitamin D3 are accompanied by changes in autocrine growth factor production and/or action (18-22, 30, 31,34).
Additional evidence for a close relationship between steroidhormone control of cellular replication and the action of autocrine growth factors is provided by their temporal sites of actionwithin the cell cycle. Epidermal growth factor and IGF-1 areprogression factors for fibroblasts; i.e., their presence is essential if cells that have begun the cell cycle are to progress throughG i phase and enter DNA synthesis, and both EGF and highconcentrations of insulin (presumably acting via the IGF-1receptor) are known mitogens for breast cells (11-17). We haveshown that both antiestrogens and progestins act in early Gìphase to inhibit cell cycle progression of human breast cancercells (10, 25). Together, these data raised the possibility thatthe sites of action within the cell cycle of these antiproliferative
»T.M. Ewing, L. J. Murphy, M-L. Ng, G. Y. N. Pang, C. S. L. Lee, and R.
L. Sutherland. Regulation of epidermal growth factor receptor by progestins andglucocorticoids in human breast cancer cell lines, submitted for publication.
114
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

MODULATION OF PROGESTIN ACTION BY EGF AND INSULIN
300
ocuE 200
JO
>100-
0J
OH-CLOM
+ EGF
OH-CLOM
1 0 100 1000 10000
Insulin concentration (ng/ml)Fig. 5. Effect of insulin and EGF on the ability of hydroxyclomiphene to
inhibit the growth of T-47D cells. Cells (5 x IO4) which had been cultured in
medium without insulin for at least 3 wk were plated into flasks containing 5 mlof insulin-free medium supplemented with 5% FCS. Two days later cells weretreated with 10 nin hydroxyclomiphene (OH-CLOM) alone {•,D), or with 10 nMhydroxyclomiphene plus EGF (10 ng/ml) (A, A) in the presence (closed symbols)or absence (open symbols) of various concentrations of insulin for 5 days. Triplicate flasks were then harvested and cell numbers recorded. Points, mean of 3 to12 flasks from 1 to 4 experiments; bars, SE. Untreated control cultures contained1.73 ±0.24 x 10' cells/flask, while cultures treated with hydroxyclomiphenealone contained 7.7 ±1.7 x 10s cells/flask. The number of cells in cultures
receiving hydroxyclomiphene plus EGF and/or insulin are recorded as a percentage of those in flasks treated with hydroxyclomiphene alone.
agents are the same as those at which progression factorsstimulate proliferation.
If antiestrogens and progestins inhibit cell proliferation, inwhole or in part, by inhibiting the production of autocrinegrowth factors, then their growth-inhibitory activity might beexpected to be modulated by addition of exogenous progressionfactors in the form of EGF or insulin. The data presented heresupport this hypothesis, clearly demonstrating that growth canbe partly restored to progestin-inhibited T-47D cells by EGF,and to antiestrogen-inhibited cells by either EGF or insulin. Innone of these cases, however, are control growth rates achieved,in contrast to the complete reversal of antiestrogen effects uponaddition of estrogen (32, 35). The failure to achieve completereversal does not necessarily reflect an irreversible lesion in theproliferative pathway, and other growth factors may furtheraugment the partial restoration of growth by EGF or insulin.
Although insulin and EGF are both capable of diminishingthe growth-inhibitory effects of antiestrogens or progestins,they apparently act through different mechanisms, such thatcells are capable of responding to insulin with a further reversalof growth inhibition, even in the presence of a maximallyeffective concentration of EGF. Our data show apparent synergy between insulin and EGF in the reversal of ORG 2058-
mediated growth inhibition. Insulin alone is essentially ineffective in this case, although it augments the decrease in growthinhibition of progestin-treated cells by EGF > 2-fold (Fig. 4).However, T-47D cells are more resistant to the antiestrogen,hydroxyclomiphene, in the presence of insulin than in thepresence of EGF, and in combination the effects of the twostimulatory agents are additive. These data, illustrating differences in growth factor effects, suggest that the growth-arrested
states induced via the ER (by antiestrogens) or via the PR (byprogestins) are qualitatively different and thus that, althoughin both cases growth arrest occurs in the d phase of the cellcycle, different mechanisms must be opérant.
Modulation of growth factor sensitivity by steroids, or viceversa, may provide a basis for apparent differences in action.The increased resistance of T-47D cells to antiestrogens in thepresence of insulin correlates with published data using MCF-7 cells (27, 28). The earlier of these studies (27) noted aninsulin-induced 3-fold decrease in ER sites/cell, which couldaccount for reduced sensitivity to antiestrogens. The minimaleffect of insulin in reversing progestin actions may then be theconsequence of an inability to alter PR expression (28). Sinceneither progestins (31) nor antiestrogens (36) induce insulinreceptors, differential sensitivity to insulin treatment is unlikelyto contribute to the differential response. Although the half-maximal response to EGF occurs at a concentration comparablewith the affinity of EGF for its receptor, a 3-fold induction ofthe EGF-R by the progestin MPA (31) is not accompanied byan enhanced response to EGF in progestin-treated cells compared to antiestrogen-treated cells, in which EGF-R levels donot change (26). Furthermore, the at least additive effect ofinsulin and EGF in combination might not have been expected,given that insulin has been shown to decrease EGF-R levels(37). Reciprocal regulation of steroid and growth factor sensitivity thus provides an attractive explanation for the responseto insulin described here, although not for that described forEGF. Therefore, these data do not allow distinction betweendirect (i.e., via receptor modulation) and indirect (i.e., viaregulation of autocrine growth factor production) models ofsteroid hormone action in breast cancer cells.
Interpretation of the results is complicated by the presenceof FCS in the culture medium used for these experiments. Anumber of growth factors and steroids, including estrogen, arepresent in FCS and, as outlined above, might be expected tomodulate the response of cells to the addition of further inhibitory or stimulatory agents. Although the present data clearlydemonstrate modulation of steroid effects on proliferation byEGF and insulin, they emphasize that the response of any celltype to a given growth factor or steroid will depend on theprecise combination of other agents present. Further dissectionof the complex, interrelated mechanisms governing hormonaland growth factor regulation of breast cancer cell cycle progression will require more detailed studies in hormonally defined and/or growth factor-depleted conditions; such studiesare currently in progress in this laboratory.
ACKNOWLEDGMENTS
The authors wish to thank Grace Pang for her expert technicalassistance and Dr. Christine Clarke for her valuable comments anddiscussion during preparation of the manuscript.
REFERENCES
1. Beatson, G. T. On the treatment of inoperable cases of carcinoma of themamma: suggestions for a new method of treatment, with illustrative cases.Lancet, 2: 104-107, 1896.
2. Pearson, O. H., West, D. C, Hollander, V. P., and Trêves,N. E. Evaluationof endocrine therapy for advanced breast cancer. J. Am. Med. Assoc., 154:234-239, 1954.
3. Lippman, M., Bolán,G., and Huff, K. The effects of estrogens and antiestrogens on hormone-responsive human breast cancer in long term tissue culture.Cancer Res., 36: 4595-4601, 1976.
4. Coezy, E., Borgna, J. L., and Rochefort, H. Tamoxifen and metabolites inMCF-7 cells: correlation between binding to estrogen receptor and inhibitionof cell growth. Cancer Res., 42: 317-323, 1982.
5. Sutherland, R. L., Hall, R. E., and Taylor, I. W. Cell proliferation kinetics
115
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

MODULATION OF PROGESTIN ACTION BY EGF AND INSULIN
of MCF-7 human mammary carcinoma cells in culture and effects of tamox-ifen on exponentially growing and plateau phase cells. Cancer Res., 43:3998-4006, 1983.
6. Sutherland, R. L., Murphy, L. C, Hall, R. E., Reddel, R. R., Watts, C. K.W., and Taylor, I. W. Effects of antiestrogens on human breast cancer cellsin vitro. Interaction with high affinity intracellular binding sites and effectson cell proliferation kinetics. Prog. Cancer Res. Ther., 31: 193-212, 1984.
7. Jordan, V. C. Biochemical pharmacology of antiestrogen action. Pharmacol.Rev., 36: 245-275, 1984.
8. Jordan, V. C. (ed.). Estrogen/Antiestrogen Action and Breast Cancer Therapy. Madison: University of Wisconsin Press, 1986.
9. Sedlacek, S. M., and Horwitz, K. B. The role of progestins and progesteronereceptors in the treatment of breast cancer. Steroids, 44:467-484, 1984.
10. Sutherland, R. L., Hall, R. E., Pang, G. Y. N., Musgrove, E. A., and Clarke,C. L. Effect of medroxyprogesterone acetate on proliferation and cell cyclekinetics of human mammary carcinoma cells. Cancer Res., 48: 5084-5091,1988.
11. Barnes, 1)., and Sato, G. Growth of a human mammary tumour cell line ina serum-free medium. Nature (Lond.), 281: 388-389, 1979.
12. Osborne, C. K., Hamilton, B., Titus, G., and Livingston, R. B. Epidermalgrowth factor stimulation of human breast cancer cells in culture. CancerRes., 40: 2361-2366, 1980.
13. Briand, P., and Lykkesfeldt, A. E. Long-term cultivation of a human breastcancer cell line, MCF-7, in a chemically defined medium. Effect of estradici.Anticancer Res., 6: 85-90, 1986.
14. Medrano, E. E., Cafferata, E. G. A., and Larcher, F. Role of thrombin in theproliferative response of T-47D mammary tumor cells. Exp. Cell Res., / 72:354-364, 1987.
15. van der Burg, B., Ruttemen, G. R., Blankenstein, M. A., de Laat, S. W., andvan Zoelen, E. J. J. Mitogenic stimulation of human breast cancer cells in agrowth factor-defined medium: synergistic action of insulin and estrogen. J.Cell. Physiol., 134:101-108, 1988.
16. Osborne, C. K., Bolán,G., Monaco, M. E., and Lippman, M. E. Hormoneresponsive human breast cancer in long-term tissue culture: effect of insulin.Proc. Nati. Acad. Sci. USA, 75:4326-4540, 1976.
17. Fitzpatrick, S. L., LaChance, M. P., and Schultz, G. S. Characterization ofepidermal growth factor receptor and action on human breast cancer cells inculture. Cancer Res., 44: 3442-3447,1984.
18. Dickson, R. B., Huff, K. K., Spencer, E. M., and Lippman, M. E. Inductionof epidermal growth factor-related polypeptides by 17/3-estradiol in MCF-7human breast cancer cells. Endocrinology, 118:138-142, 1985.
19. Dickson, R., Bates, S. E., McManaway, M. E., and Lippman, M. E. Characterization of estrogen responsive transforming activity in human breastcancer cell lines. Cancer Res., 46:1707-1713, 1986.
20. Huff, K. K., Kaufman, D., Gabbay, K. H., Spencer, E. M., Lippman, M. E.,and Dickson, R. B. Secretion of an insulin-like growth fuclor-I-related proteinby human breast cancer cells. Cancer Res., 46:4613-4619,1986.
21. Dickson, R. B., and Lippman, M. E. Estrogenic regulation of growth andpolypeptide growth factor secretion in human breast carcinoma. EndocrineRev., «.-29-43,1987.
22. Bates, S. E., Davidson, N. E., Valverius, E. M. Fréter,C. E., Dickson, R. B.,Tarn, J. P., Kudlow, J. E., Lippman, M. E., and Salomon, D. S. Expression
of transforming growth factor a and its messenger ribonucleic acid in humanbreast cancer: its regulation by estrogen and its possible functional significance. Mol. Endocrino!., 2: 543-555, 1988.
23. Leof, E. B., Wharton, W., van Wyk, J. J., and Pledger, W. J. Epidermalgrowth factor (EGF) and somatomedin C regulate Gl progression in competent BALB/C-3T3 cells. Exp. Cell Res., 141: 107-115, 1982.
24. O'Keefe, E. J., and Pledger, W. J. A model of cell cycle control: sequentialevents regulated by growth factors. Mol. Cell. Endocrinol., 31: 167-186,1983.
25. Taylor, I. W., Hodson, P. J., Green, M. D., and Sutherland, R. L. Effects oftamoxifen on cell cycle progression of synchronous MCF-7 human mammarycarcinoma cells. Cancer Res., 43: 4007-4011, 1983.
26. Koga, M., and Sutherland, R. L. Epidermal growth factor partially reversesthe inhibitory effects of antiestrogens on T-47D human breast cancer cellgrowth. Biochem. Biophys. Res. Commun., 146: 739-745, 1987.
27. Butler, W. B., Kelsey, W. H., and Goran, N. Effects of serum and insulin onthe sensitivity of the human breast cancer cell line MCF-7 to estrogen andantiestrogens. Cancer Res., 41: 82-88, 1981.
28. Vignon, F., Bouton, M-M., and Rochefort, H. Antiestrogens inhibit themitogenic effect of growth factors on breast cancer cells in the total absenceof estrogens. Biochem. Biophys. Res. Commun., 146:1502-1508, 1987.
29. Ruenitz, P. C., Bagley, J. R., and Mokier, C. M. Metabolism of clomiphenein the rat. Estrogen receptor affinity and antiestrogenic activity of clomiphenemetabolites. Biochem. Pharmacol., 32: 2941-2947, 1983.
30. Koga, M., Eisman, J. A., and Sutherland, R. L. Regulation of epidermalgrowth factor receptor levels by 1,25-dihydroxyvitamin D3 in human breastcancer cells. Cancer Res., 48: 2734-2739, 1988.
31. Murphy, L. J., Sutherland, R. L., Stead, B., Murphy, L. C., and Lazarus, L.Progestin regulation of epidermal growth factor receptor in human mammarycarcinoma cells. Cancer Res., 46: 728-734, 1986.
32. Ruenitz, P. C., Bagley, J. R., Watts, C. K. W., Hall, R. E., and Sutherland,R. L. Vinyl substituted hydroxytriarylethylenes, l-|4-[2-(diethylamino)]-phenyl|-l-(4-hydroxyphenyl)-2-phenylethylene: synthesis and effects onMCF-7 breast cancer cell proliferation. J. Med. Chem., 29: 2511-2519,1986.
33. Sutherland, R. L., Reddel, R. R., and Green, M. D. Effects of oestrogens oncell proliferation and cell cycle kinetics. A hypothesis on the cell cycle effectsof antioestrogens. Eur. J. Cancer Clin. Oncol., 19: 307-318, 1983.
34. Liu, S. C., Sanfilippo, B., Perroteau, I., Derynck, R., Salomon, D. S., andKidwell, W. R. Expression of transforming growth factor a (TGF-a) indifferentiated rat mammary tumors: estrogen induction of TGF-a production.Mol. Endocrinol., I: 683-692, 1987.
35. Reddel, R. R., Murphy, L. C., and Sutherland, R. L. Factors affecting thesensitivity of T-47D human breast cancer cells to tamoxifen. Cancer Res.,44: 2398-2405, 1984.
36. Horwitz, K. B., and Freidenberg, G. R. Growth inhibition and increase ofinsulin receptors in antiestrogen-resistant T47D,,, human cancer cells byprogestins: implications for endocrine therapies. Cancer Res., 45: 167-173,1985.
37. Ryan, P. D., and Hilf, R. Characterization and regulation of epidermalgrowth factor (EGF) receptors in R3230AC rat mammary tumors. Proc. Am.Assoc. Cancer Res., 29: 247, 1988.
116
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from

1989;49:112-116. Cancer Res Masafumi Koga, Elizabeth A. Musgrove and Robert L. Sutherland Cells by Epidermal Growth Factor and Insulinthe Antiestrogen Hydroxyclomiphene on Human Breast Cancer Modulation of the Growth-inhibitory Effects of Progestins and
Updated version
http://cancerres.aacrjournals.org/content/49/1/112
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/49/1/112To request permission to re-use all or part of this article, use this link
Research. on February 2, 2019. © 1989 American Association for Cancercancerres.aacrjournals.org Downloaded from


















